4
TINJAUAN PUSTAKA
A. Kajian Teori
1. Penyembuhan luka
1.1. Healing Cascade
Healing cascade dimulai segera setelah terjadinya perlukaan, dimana
terjadi kontak antara Trombosit dengan kolagen dari jaringan yang terpapar terhadap darah, yang mana akan menyebabkan pelepasan faktor pembekuan dan deposisi fibrin kedalam lokasi luka, bentukan ini bukan hanya berfungsi untuk menghentikan perdarahan, namun juga akan menjadi matrik dan mendasari tahap selanjutnya dari pemyembuhan luka. Platelet melepaskan faktor pembekuan dan berbagai mediator kimia yang dikenal sebagai Sitokin dan growth factor, dua yang terutama adalah PDGF dan TGF-β(Rajan dan Murray, 2008).
PDGF akan memicu proses kemotaksis dari Netrofil, Makrofag, otot polos dan Fibroblas, dan juga memulai proses mitosis dari sel Fibroblas dan otot polos. TGF-β berperan dalam menarik Makrofag dan menstimulasi pelepasan Sitokin -sitokin lain seperti FGF, TNF-α, dan IL-1. TGF-β juga diketahui memperkuat
kemotaksis dari Fibroblas dan otot polos, dan memodulasi pembentukan kolagen dan kolagenase. Proses ini secara keseluruhan akan menyebabkan deposisi jaringan ikat baru kedalam lokasi luka yang dikenal sebagai fase proliferasi, dan setelah semua proses epithelialisasi, granulasi, dan neovaskularisasi selesai, akan diikuti oleh suatu proses remodelling untuk mengembalikan struktur yang baru terbentuk mendekat kondisi awalnya (Eming et. al. 2007).
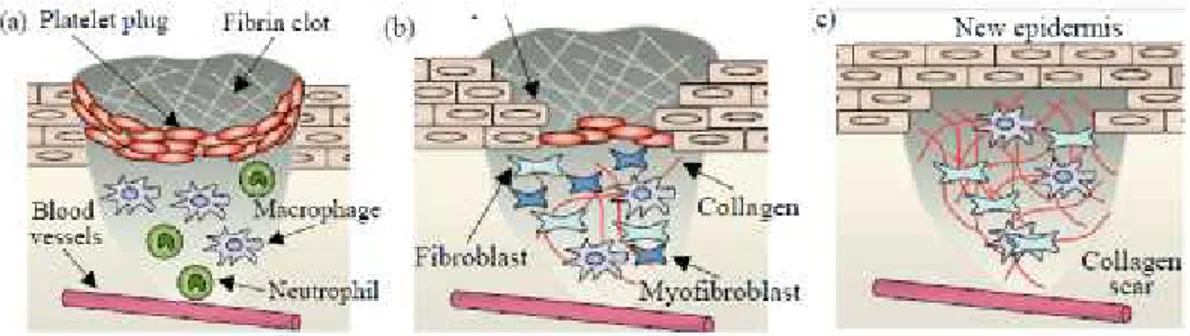
Gambar 1 : Tahapan Penyembuhan luka (Rajan dan Murray, 2008). a. Fase Inflamasi b. Fase Proliferasi c. Fase Remodelling
1.1.1 Fase inflamasi
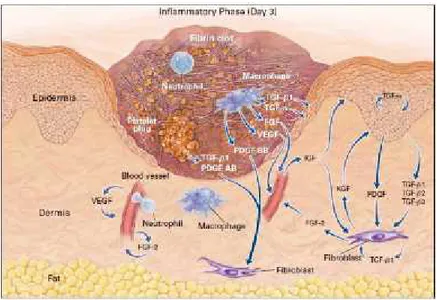
Netrofil merupakan sel radang pertama yang dijumpai pada daerah luka, biasanya mulai muncul dalam 24 jam pertama setelah kerusakan, fungsi utamanya untuk mengeliminasi benda asing, bakteri, sel dan matrik jaringan yang rusak. Sel Mast merupakan sel yang kaya dengan granula berisi berbagai macam enzim, Histamin dan berbagai jenis mediator kimia lain yang bertanggung jawab terhadap terjadinya inflamasi pada daerah sekitar luka. Bahan aktif yang dilepaskannya akan memicu serangkaian proses yang menyebabkan peningkatan permeabilitas pembuluh darah sehingga sel monosit bisa dengan mudah bermigrasi kedalam jaringan yang luka (Eming et. al. 2007).
Sel Monosit dalam darah akan menjadi teraktivasi dan menjadi Makrofag setelah 48 jam, yang berperan besar dalam tahap inflamasi penyembuhan luka dan gangguan terhadap fungsi Makrofag akan mengganggu penyembuhan luka. Setelah teraktivasi, sel Makrofag sendiri juga akan menghasilkan PDGF dan TGF-β. Sifat fagositik dari Makrofag
bertujuan untuk mengeliminasi sel dan matrik yang rusak, Netrofil yang penuh dengan patogen, benda asing dan sisa bakteri yang masih tersisa. Adanya Wound Macrophage menandakan akhir proses inflamasi dan segera dimulainya proses proliferasi. Limfosit juga dijumpai pada lokasi terjadinya luka, namun sel ini dinyatakan tidak terlalu memiliki peran yang
menonjol dalam proses peyembuhan luka dan peran pastinya masih perlu ditelaah lebih lanjut (Rajan dan Murray, 2008).
Gambar 2 : Fase Inflamasi, sel-sel dan mediator yang berperan didalamnya (Epstein et. al, 1999)
1.1.2 Fase proliferasi
Fase proliferasi terdiri atas proses reepitelialisasi, neovaskularisasi, dan pembentukan jaringan granulasi, dalam fase ini peran TGF-β yang
dilepaskan oleh Trombosit, Makrofag memegang peranan penting sebagai pengatur fungsi Fibroblas. TGF-β memiliki beberapa peran penting dalam
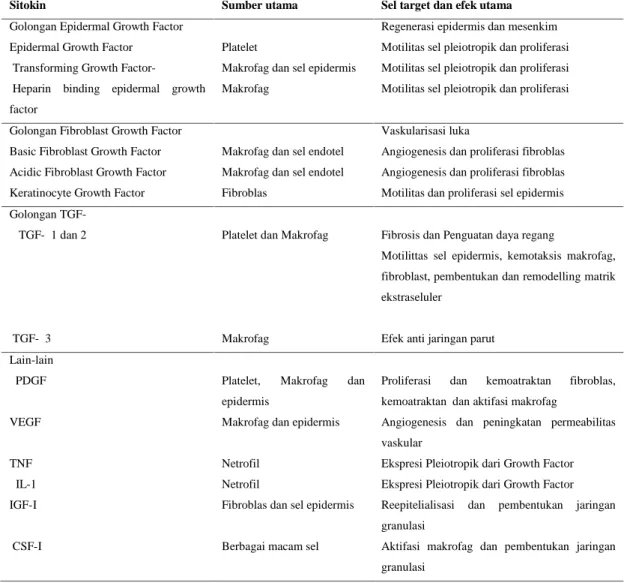
pembentukan matrik ekstraselular, yaitu meningkatkan pergerakan sel epidermis, pembentukan kolagen, proteoglikan, dan fibronektin, serta mengurangi produksi dari enzim protease yang merusak matrik. Sitokin-sitokin lain yang berperan dalam proses penyembuhan luka dirangkum pada tabel 1 (Diegelmann, 2004 ; Epstein et. al, 1999).
Tabel 1. Berbagai jenis Sitokin yang berperan pada penyembuhan luka (Epstein et. al, 1999)
Sitokin Sumber utama Sel target dan efek utama Golongan Epidermal Growth Factor
Epidermal Growth Factor Transforming Growth Factor-α
Heparin binding epidermal growth factor
Platelet
Makrofag dan sel epidermis Makrofag
Regenerasi epidermis dan mesenkim Motilitas sel pleiotropik dan proliferasi Motilitas sel pleiotropik dan proliferasi Motilitas sel pleiotropik dan proliferasi
Golongan Fibroblast Growth Factor Basic Fibroblast Growth Factor Acidic Fibroblast Growth Factor Keratinocyte Growth Factor
Makrofag dan sel endotel Makrofag dan sel endotel Fibroblas
Vaskularisasi luka
Angiogenesis dan proliferasi fibroblas Angiogenesis dan proliferasi fibroblas Motilitas dan proliferasi sel epidermis Golongan TGF-β
TGF-β1 dan 2
TGF-β3
Platelet dan Makrofag
Makrofag
Fibrosis dan Penguatan daya regang
Motilittas sel epidermis, kemotaksis makrofag, fibroblast, pembentukan dan remodelling matrik ekstraseluler
Efek anti jaringan parut Lain-lain PDGF VEGF TNFα IL-1 IGF-I CSF-I
Platelet, Makrofag dan epidermis
Makrofag dan epidermis
Netrofil Netrofil
Fibroblas dan sel epidermis
Berbagai macam sel
Proliferasi dan kemoatraktan fibroblas, kemoatraktan dan aktifasi makrofag
Angiogenesis dan peningkatan permeabilitas vaskular
Ekspresi Pleiotropik dari Growth Factor Ekspresi Pleiotropik dari Growth Factor Reepitelialisasi dan pembentukan jaringan granulasi
Aktifasi makrofag dan pembentukan jaringan granulasi
Fibroblas akan berikatan dengan serabut dari matrik fibrin dan mulai memproduksi kolagen, sampai saat ini telah diketahui ada 23 jenis kolagen, yang dominan ditemukan pada kulit adalah kolagen tipe 1. Pembentukan kolagen dimulai dari pembentukan prokolagen dengan karakter khas triple helix, setelah di sekresikan ke dalam ruang ekstraselular, kemudian akan mengalami hidroksilasi dan kemudian mengalami pembelahan pada gugus terminal peptida prokolagen N dan C oleh enzim Lysyl Oxydase yang memungkinkan terjadinya crosslink yang
lebih stabil. Kolagen normal pada kulit tersusun teratur dan memiliki kekuatan regangan yang setara dengan baja, namun pada jaringan parut, ukurannya lebih kecil dan tidak beraturan, sehingga lebih lemah dan mudah sekali rusak dibandingkan jaringan sekitarnya (Diegelman, 2014; Rajan dan Murray, 2008).
Reepitelialisasi terjadi dalam beberapa jam setelah terjadi luka, dan
Sitokin yang berperan adalah EGF dan TGFα yang dihasilkan oleh
Platelet, Makrofag, dan keratinosit. Karena proses ini memiliki aktivitas metabolik yang tinggi, maka akan timbul peningkatan kebutuhan oksigen dan nutrisi. Penurunan pH, oxygen tension, dan peningkatan laktat dilokasi sekitar luka akan memicu serangkaian proses yang mendorong terbentuknya pembuluh darah baru atau yang lazim dikenal sebagai angiogenesis atau neovaskularisasi, yang terutama dipengaruhi oleh VEGF, bFGF dan TGF-β. Proses ini vital dalam kelangsungan proses
selanjutnya yaitu pembentuk jaringan granulasi pada hari ke 4-7 (Diegelman, 2004; Rajan dan Murray, 2008).
Proses angiogenesis bisa dijelaskan secara singkat sebagai berikut: trauma akan menimbulkan kerusakan jaringan, dan bFGF akan segera dilepaskan oleh Makrofag dan VEGF oleh sel epidermis yang mengalami hipoksia. Enzim proteolitik yang dilepaskan akan merusak protein matrik ekstraselular, dan fragmen protein yang dihasilkan akan berfungsi untuk merekrut sel monosit menuju lokasi kerusakan jaringan, yang nantinya Monosit akan teraktivasi dan berubah menjadi Makrofag. Beberapa Sitokin yang dihasilkan Makrofag, seperti bFGF akan menstimulasi endotel untuk melepas tPA yang akan mengubah Plasminogen menjadi Plasmin dan Prokolagenase yang mengaktifkan Polagenase, kedua enzim proteolitik ini akan merusak membran basalis, sehingga memungkinkan sel Endotel yang terstimulasi untuk bergerak dan membentuk pembuluh darah baru dilokasi cedera. Proses angiogenesis akan terhenti setelah terbentuk granulasi dan pembuluh darah baru yang banyak tersebut akan mengalami disintegrasi
akibat apoptosis, dengan berakhirnya tahap ini, proses penyembuhan dilanjutkan oleh fase remodelling (Epstein et. al, 1999; Wulff, 2012). 1.1.3 Fase Remodelling
Sebagian molekul kolagen terdegradasi oleh enzim kolagenase yang didapatkan pada Fibroblas, Makrofag, dan Netrofil pada fase
remodelling, disamping itu juga terjadi kontraksi luka (wound contraction)
yang merupakan suatu proses kompleks dimana melibatkan berbagai jenis sel, matrik, dan Sitokin. Pada periode ini, Fibroblas memiliki suatu gambaran fenotipe yang disebut myofibroblas, yang mampu melakukan kontraksi, adanya fenomena ini menunjukan adanya pemadatan dari
jaringan ikat dan kontraksi dari luka. Proses ini diduga dipicu oleh TGF β1 atau β2 dan PDFG (Rajan dan Murray, 2008).
Remodelling dari kolagen dipengaruhi oleh keseimbangan antara
sintesis dan katabolisme kolagen. Degradasi kolagen pada luka juga dipengaruhi oleh beberapa enzim proteolitik yang disebut MMP yang dihasilkan oleh sel Makrofag, epidermis, endothel dan Fibroblas. Keseimbangan antara MMP dan inhibitor dari MMP akan menentukan perkembangan penyembuhan luka. Proses remodelling memungkinan kekuatan jaringan baru yang terbentuk bisa mendekati aslinya, pada 3 minggu pertama setelah cedera, kekuatan ini hanya berkisar 20% dari semula, dalam proses remodelling akan terjadi penggantian serabut kolagen dengan serabut yang lebih besar disertai oleh penguatan
crosslinking dari masing masing serabut yang membentuk jaringan yang
lebih kuat. Kekuatan maksimal yang bisa dicapai oleh jaringan parut baru hanyalah 70% dari kulit yang normal (Demidova-Rice, et al. 2012; Epstein et. al, 1999).
1.2 Sel-sel inflamasi yang berperan dalam penyembuhan luka 1.2.1 Netrofil
Segera setelah terjadi cedera, maka akan segera dibentuk bekuan darah untuk menyumbat pembuluh darah yang terbuka. Sel Trombosit dan
Netrofil yang terperangkap didalam bekuan darah akan mengeluarkan berbagai macam faktor yang memperkuat proses agregasi Trombosit, koagulasi dan menarik sel-sel yang berperan dalam proses inflamasi. Dalam beberapa jam, akan terjadi pergerakan sel Netrofil menembus Endotel, yang mana dipicu oleh Sitokin pro inflamasi seperti IL-1β, TNF
-α, dan IFN-γ yang akan memicu dihasilkannya molekul adhesi yang
berperan dalam proses adhesi dan diapedesis leukosit, seperti Endothelial
P and E-selectins, ICAM 1 dan 2. Mediator lain yang ikut berperan adalah
IL-8, MCP-1 dan growth related oncogene-α. Produk dari bakteri seperti LPS, juga dapat memperkuat proses pergerakan Leukosit (Eming et. al. 2007; Sabol ,2012).
Netrofil yang terekrut akan segera memulai proses debridement dari jaringan yang mati dan memfagositir sel-sel patogen, proses ini membutuhkan berbagai macam senyawa antimikroba seperi ROS, peptida kation, dan enzim Protease. Jenis mediator yang dihasilkan atau berperan pada proses rekrutmen Netrofil telah banyak diketahui, namun sampai saat ini belum diketahui pasti apakah Netrofil bersifat merugikan atau menguntungkan pada penyembuhan luka, beberapa penelitian oleh Simpson dan Ross di tahun 1972 dan Dovi et al. di tahun 2003 menunjukkan bahwa parameter penyembuhan tidak terpengaruh oleh netropenia, dan bahkan didapatkan reepitelialisasi lebih cepat pada kondisi tersebut. Sehingga memunculkan dugaan bahwa mungkin ada sel lain yang lebih berperan dalam proses pemyembuhan luka, seperti halnya sel Makrofag (Eming et al. 2007; Sabol ,2012).
1.2.2 Monosit / Makrofag
Pada keadaan normal beberapa hari setelah perlukaan, tanpa stimulus yang berlanjut tidak akan terjadi lagi rekrutmen Netrofil kedaerah cedera. Proses ini akan diikuti oleh masuknya Makrofag, yang akan memfagosit sel Netrofil yang telah penuh terisi debris dan patogen. Umumnya Makrofag yang didapatkan didaerah luka mulai ditemukan pada hari ke -2 setelah terjadinya cedera dan berasal dari monosit darah, disamping sel Makrofag resident didalam kulit sendiri. Bila pada sel PMN proses ekstravasasi diatur oleh komplek CD11/18 dan ICAMs, maka pergerakan monosit dari pembuluh darah menuju luka dipengaruhi oleh interaksi dari very late antigen-4 (α4β1 integrin) dan endothelial vascular cell adhesion molecule-1. Sedangkan infiltrasi Makrofag pada luka
dipengaruhi oleh berbagai macam faktor kemotaktik, seperti faktor pertumbuhan, Sitokin pro inflamasi dan chemokines macrophage inflammatory protein 1α, MCP-1. Trombosit merpakan sumber utama zat-zat tersebut yang terperangkap didalam bekuan darah, keratinosit ditepi luka yang berada dalam tahap hiperproliferasi, Fibroblas dan Leukosit lain (Eming et al., 2007; Wulff, 2012).
Monosit akan mengalami aktivasi dan berdiferensiasi menjadi Makrofag jaringan yang mature setelah meninggalkan pembuluh darah, diikuti oleh perubahan ekspresi gen, yang dipengaruhi oleh berbagai mediator yang ditemukan pada lingkungan mikro disekitar luka yang menyebabkan sel Makrofag mengalami perubahan sifat sesuai dengan kebutuhan di lokasi luka. Makrofag berperan sebagai sel penyaji antigen dan Fagosit selama proses penyembuhan luka, juga diduga memiliki peran dalam proses penyembuhan melalui sintesis berbagai macam faktor pertumbuhan yang penting seperti TGF-α, TGF-β, bFGF, PDGF dan
VEGF yang akan meningkatkan proliferasi sel dan sintesis matrik ekstraseluler oleh sel kulit (Rajan dan Murray, 2008).
Sel Mast umumnya dikenal sebagai sel-sel "alergi" yang menyebabkan gejala dini dari segala jenis alergi dan biasanya didapatkan pada lokasi di mana jaringan inang berkontak dengan antigen eksternal, alergen, racun dan mikroba, misalnya lapisan atas dermis, saluran pernapasan dan mukosa usus. Peran fisiologis sel Mast masih belum sepenuhnya bisa dijelaskan, bukti yang diperoleh dari model tikus menunjukkan bahwa sel-sel Mast pada dasarnya terlibat dalam melindungi kulit dari infeksi bakteri dan parasit yang berat, selain itu, sel-sel Mast mengatur penyembuhan luka kulit setelah trauma (Harvima dan Nilsson, 2011).
Sitokin yang dihasilkan sel Mast akan meningkatkan produksi Sitokin proinflamasi oleh sel resident . Pelepasan vasoactive amine merangsang permeabilitas pembuluh darah, memacu masuknya Netrofil, Makrofag, dan sel Mast tambahan dalam jumlah besar ke dalam jaringan, konsekuensinya aktivasi dan degranulasi sel Mast resident meningkat dan memperpanjang respon inflamasi. Sel Mast sangat menonjol di jaringan parut, dan sel Mast yang teraktivasi akan berdiam di jaringan parut hingga setahun setelah terjadi perlukaan. Aktivasi sel Mast dapat mempengaruhi
remodelling akibat inflamasi berlebih dan produksi Sitokin memacu formasi
jaringan parut. Penyembuhan tanpa disertai jaringan parut mukosa mulut pada babi Duroc merah, sebagai model utama jaringan parut hipertropi, memiliki jumlah sel Mast yang lebih rendah dibandingkan luka pada kulit (Chen Lin et al., 2014)
Sel Mast dalam jaringan manusia dapat diklasifikasikan ke dalam MCTC, MCT, dan MCc berdasarkan kandungan proteinasenya. Sel MCTC mengandung Triptase, Chymase, karboksipeptidase dan katepsin G-seperti proteinase, sel MCT hanya berisi Triptase dan sel MCc menunjukkan Chymase dan karboksipeptidase, tapi tidak mengandung Triptase. Semua jenis sel Mast mengandung Histamin. Sebagian besar sel Mast pada kulit manusia bertipe MCTC, sedangkan sel-sel MCT mendominasi di paru-paru dan mukosa usus. Hal ini menunjukkan bahwa semua jenis enzim tipe "C",
seperti Chymase, Karboksipeptidase dan cathepsin G-like proteinase, memiliki fungsi sepsifik di kulit setelah dilepaskan dari sel Mast melalui proses degradasi protein dan peptida yang bervariasi (Eming, et al. 2007; Wulff dan Wilgus, 2013).
Dalam model tradisional, secara sederhana sel Mast diaktifkan untuk mengalami degranulasi dan melepaskan mediator melalui proses cross
linking antara alergen dengan IgE dan reseptor FcεRI-nya pada membran sel. Namun, kini diketahui bahwa Sel Mast kulit juga dapat
mengekspresikan reseptor FcγRI- dan FcγRIIa dan dengan demikian dapat
diaktifkan oleh mekanisme yang dependen terhadap IgG. Peptida Endogen dan protein tertentu dapat mengaktifkan sel-sel Mast untuk melepas mediator, antara lain C3a dan C5a, neuropeptida termasuk substansi P dan peptida vasoaktif intestinal, faktor sel induk, TNF, Triptase, cathelicidin LL-37, α-melanosit stimulating hormone dan corticotrophin-releasing hormone
(Harvima dan Nilsson, 2011; Rajan dan Murray, 2008).
Telah diketahui bahwa sel-sel Mast tidak hanya bisa diaktifkan untuk rilis mediator dengan mekanisme sederhana yaitu dalam keadaan istirahat atau degranulasi anafilaksis yang cepat dan luas, namun juga melalui proses degranulasi lambat dan parsial sedikit demi sedikit, selain itu mekanisme sekresi lainnya juga telah dijelaskan, seperti sekresi exosome, dan sekresi mediator selektif yang independen terhadap. Sebagai contoh, peningkatan konsentrasi Histamin interstitial dalam plak psoriasis menunjukkan aktivitas sel Mast dan degranulasi yang tinggi dan dalam lesi peradangan kronis tanpa disertai proses anafilaksis dan gambaran urtikaria (Eming et. al. 2007; Harvima dan Nilsson, 2011).
Meskipun sel Mast penting dalam timbulnya alergi tipe langsung dan terlibat dalam reaksi fisiologis kulit terhadap trauma dan infeksi, mereka juga dapat mempengaruhi sistem kekebalan tubuh, meningkatkan peradangan atau bahkan menekannya. Mediator pre-formed disimpan dalam granul sekretori termasuk berbagai jenis protease, Histamin, proteoglikan
heparin, kondroitin sulfat E, acidic hydrolase, berbagai Sitokin dan faktor pertumbuhan (Wulff dan Wilgus, 2013).
Sel Mast resident umumnya mengalami degranulasi setelah terjadinya luka, sehingga jarang ditemukan pada awal cedera, jumlahnya mulai kembali normal dan meningkat setelah 48 jam setelah cedera. Beberapa penelitian menunjukkan bahwa sel Mast berperan dalam proses migrasi Netrofil, namun tidak berpengaruh pada sel Makrofag dan limfosit-T. Penelitian lain menunjukkan bahwa defisiensi sel Mast tidak berpengaruh pada epitelialisasi, sintesa kolagen dan angiogenesis, walaupun masih ada kontroversi terkait temuan ini (Eming et. al. 2007; Sabol ,2012).
Selain Histamin, protein utama dalam butiran sel Mast, adalah β -tryptase, yang merupakan suatu serin proteinase yang menyerupai tripsin,
yang memiliki struktur cincin tetramerik. Berdasarkan struktur ini, β -tryptase tahan terhadap sebagian besar inhibitor protease endogen, dan untuk menstabilkan enzim ini diperlukan heparin. Dalam kulit yang meradang, Triptase secara histokimia berperan sebagai katalis aktif. Peran
β-tryptase dalam patofisiologi penyembuhan luka masih belum sepenuhnya jelas. Namun, ada beberapa temuan eksperimental yang menunjukkan perannya dalam aktivasi dan perekrutan jenis sel yang berbeda, termasuk sel-sel endotel, sel-sel mononuklear, sel T dan Netrofil di darah perifer. Pada hewan coba, suntikan Triptase menginduksi akumulasi Netrofil, Eosinofil dan sel-sel lain dari sistem kekebalan tubuh pada kulit marmut. Triptase mungkin meningkatkan peradangan neurogenik dengan mengaktifkan PAR-2 pada saraf yang menyebabkan pelepasan substansi P, neuropeptida dan peptida yang terkait gen-kalsitonin (Harvima dan Nilsson, 2011).
Triptase dapat mengaktifkan keratinosit secara langsung melalui aktivasi PAR-2 pada permukaan mereka. Triptase juga mampu mengaktifkan enzim metaloprotenase dan pro-urokinase, atau dapat berfungsi sebagai gelatinase. Dengan demikian, Triptase dapat membuat ruang untuk sel T dan Netrofil dalam matrik ekstraseluler dan membran
basalis yang memungkinkan migrasi ke dalam epidermis, namun Triptase mungkin juga memiliki fungsi penghambatan, karena dapat menurunkan kemokin, neuropeptida dan cathelicidin LL-37 (Eming et. al. 2007; Wulff dan Wilgus, 2013).
Tabel 2 Fungsi stimulasi dan inhibisi dari enzim Triptase (Harvima dan Nilsson, 2011)
Stimulasi Inhibisi
Angiogenesis, MCP-1 dan IL-8 pada sel endotel
Pembelahan eotaxin dan RANTES
Aktivasi sel MN dan PMN di darah tepi untuk produksi Sitokin
Pembelahan dari neuropeptida VIP dan CGRP
Aktivasi saraf dan keratinosit melalui reseptor PAR-2
Pembelahan cathelicidin LL-37
Aktivasi MMP-3 & 9, dan pro urokinase
α-chymase adalah suatu chymotryptic serin proteinase yang disimpan dalam jumlah tinggi dalam butiran sekret sel Mast. Seperti Triptase, Chymase juga berikatan kuat dengan heparin, tapi kompleks proteoglikan chymase-heparin berukuran lebih besar dan terletak di sub regional yang berbeda dari granul dari kompleks proteoglikan Triptase-heparin. Sehingga, Triptase dapat menyebar melalui matrik ekstraselular, sedangkan Chymase cenderung tetap di lokasi aktivasi. Berbeda dengan Triptase, Chymase bisa teraktivasi walaupun tidak ada heparin. Aktivitas Chymasediatur oleh inhibitor protease plasma α1-PI dan α1-AC, yang juga mengalami peningkatan kadar di dalam sel Mast pada kulit yang meradang (Nauta et al., 2013).
Chymase diduga berperan dalam perekrutan sel inflamasi, dari beberapa eksperimen ditunjukkan injeksi Chymase manusia ke dalam kulit marmut akan merangsang akumulasi Netrofil dan Eosinofil, dan merangsang Monosit, Netrofil, Limfosit dan sel-sel Eosinofilik in vitro.
Chymase dapat mempromosikan peradangan secara tidak langsung dengan mengaktifkan pro-IL-1β menjadi IL-1β, dan pro-IL-18 menjadi IL-18, dan menghasilkan 31-amino acid endothelin-1, suatu kemoatraktan kuat untuk Netrofil dan Monosit. Namun, Chymase juga dapat mengatur inflamasi dengan mendegradasi IL-6 dan IL-13, dan sampai batas tertentu IL-5 dan TNF-α. Chymase dapat mempengaruhi epidermis dan menginduksi
pembentukan bullae dalam beberapa kondisi, karena melepaskan Keratinosit dari lapisan substratum dan mendegradasi Fibronektin (Wulff dan Wilgus, 2013).
1.2.4 Sel - Limfosit T
Pada fase remodelling, saat penutupan luka telah terjadi, dan infeksi telah teratasi, sel T merupakan salah satu jenis leukosit yang paling banyak ditemukan pada kulit. Kemokin yang berperan terhadap kemotaksis dan fungsi dari sel ini antara lain MCP-1 yang ditemukan pada hari ke 4 setelah cedera, dipicu oleh IFN-γ-inducible protein-10 dan monokin yang
diinduksi-IFN-γ. Mediator-mediator ini terutama diproduksi oleh Makrofag. Keberadaan Sitokin ini menunjukkan pergeseran jenis Sitokin dari pro-inflamasi ke anti pro-inflamasi. Pada tikus coba dengan defisiensi IFN-γ, jumlah
Netrofil, Makrofag dan sel-T berkurang secara signifikan pada lokasi luka, hal ini mungkin berkaitan dengan berkurangnya aktivitas endothel, hal ini mendukung dugaan bahwa sel –T berperan pada proses remodelling. Ada dua subset dari Sel-T didaerah luka, yaitu Th1 dan Th2,Sel Th1 menghasilkan IFN-γ, IL-2 dan TNFα, Th2 menghasilkan sel IL4,5, dan -10. Sel T juga berinteraksi dengan keratinosit, Fibroblas, Platelet dan Makrofag yang mengekspresikan CD-40, yang akan mengubah ekspresi dari mediator pro inflamasi yang dihasilkan oleh sel-sel tersebut dan efeknya terhadap penyembuhan luka. Temuan terakhir adalah jenis baru dari limfosit T yang disebut Limfosit T-γδ yang merupakan suatu DETC, terletak
spesifik di lapisan epidermis saja, sel ini berfungsi untuk mendeteksi antigen yang dihasilkan oleh keratinosit yang mengalami kerusakan pada epidermis. Sel DETC menghasilkan berbagai macam faktor pertumbuhan seperti
FGF-7 dan 10, dan juga IGF-1, sehingga mempengaruhi proliferasi dan diferensiasi dari keratinosit. Disamping itu sel ini juga mempengaruhi deposit Hyaluronan pada matrik ekstraseluler yang diperantarai oleh keratinosit, dan infiltrasi macrofag kedalam luka. (Eming et. al. 2007; Harvima dan Nilsson, 2011; Sabol ,2004).
2. Pengaruh inflamasi terhadap penyembuhan luka
2.1. Mekanisme resolusi peradangan pada luka
Inflamasi merupakan bagian dari proses penyembuhan luka, namun untuk agar bisa berlangsung sempurna, perlu adanya resolusi peradangan pada saat yang tepat, sampai saat ini masih banyak hal yang perlu dipelajari mengenai hal ini. Mekanisme resolusi inflamasi, bisa terjadi karena downregulation dari berbagai kemokin pro inflamasi oleh Sitokin anti inflamasi seperti IL-10, atau TGF-β1,
atau upregulation dari molekul anti inflamasi seperti antagonis reseptor IL-1 atau reseptor TNF. MMP juga dapat mengurangi inflamasi melalui pembelahan kemokin, yang mana akan bersifat sebagai antagonis. Beberapa mediator apoptosis seperti CD44 dan kaspase, faktor transkripsi seperti Nrf-2, yang merupakan target dari keratinocyte growth factor-1 juga berperan terhadap proses resolusi inflamasi (Eming et. al. 2007).
2.2. Inflamasi dan kualitas penyembuhan luka
Fibrosis adalah suatu proses fisiologis dan tidak dapat dihindari pada suatu proses penyembuhan luka pada mammalia, dan bukti-bukti yang ada menyokong dugaan bahwa inflamasi berperan dalam menentukan derajat pembentukan jaringan parut. Evolusi pada mammalia menyebabkan pada kondisi luka akan terjadi proses keradangan yang terjadi secara cepat, hal ini dimaksudkan agar tetap bisa terjadi penyembuhan luka tanpa infeksi, walaupun pada kondisi luka yang kotor. Sehingga bisa dikatakan bahwa jaringan parut adalah suatu harga yang harus dibayar, untuk mendapatkan penyembuhan luka yang cepat tersebut (Harvima dan Nilsson,2011; Wulff dan Wilgus, 2013).
Ada perbedaan dalam pembentukan jaringan parut, pada individu dewasa cenderung mudah terbentuk jaringan parut, sedangkan pada embrio umumnya penyembuhan luka bisa terjadi tanpa pembentukan jaringan parut, dan meskipun
ada banyak faktor yang diduga menyebabkan perbedaan tersebut, namun bukti terakhir menunjukkan bahwa banyak yang tidak berhubungan dengan hal tersebut, seperti adanya kondisi lingkungan yang steril dalam uterus, adanya cairan amnion, dan hal-hal lain. Satu hal utama yang membedakan proses pembentukan jaringan parut diantara keduanya adalah derajat inflamasi yang ditemukan didalam luka, dimana pada fetus yang sistem imunitasnya belum berkembang, sel-sel radang yang ditemukan dalam luka, jauh lebih rendah baik dalam jumlah, maupun tingkat aktivitasnya. Bahkan Makrofag, umumnya baru mengalami rekrutmen, setelah terjadinya penutupan luka, hal ini berbeda dengan proses normal yang ditemukan pada individu dewasa(Diegelman, 2004; Eming et. al. 2007).
Kondisi yang mirip ditemukan pada beberapa bagian tubuh individu dewasa, seperti mukosa mulut, yang pada umumnya tidak bisa memicu reaksi inflamasi seperti pada kulit, sehingga memiliki kadar Netrofil dan Makrofag yang lebih rendah, sehingga bisa sembuh dengan derajat scarring yang lebih minimal. (Rajan dan Murray, 2008).
Gambar 3
Mediator dan mekanisme pada tahap inflamasi dan resolusi inflamasi penyembuhan luka
3. Peran sel Mast dalam penyembuhan luka
Peran sel Mast dalam proses penyembuhan luka masih kontradiktif sampai saat ini, beberapa penilitian, seperti yang dilakukan oleh Nauta dan kawan-kawan di tahun 2013, menunjukkan bahwa sel Mast tidak dibutuhkan pada penyembuhan luka eksisi pada tikus coba. Hal ini bertentangan dengan penilitian lain, seperti yang dilakukan oleh Weller dan kawan-kawan di tahun 2006, yang menemukan bahwa sel Mast diperlukan untuk penyembuhan luka pada tikus. Ada beberapa hipotesis mengenai peran sel Mast dalam penyembuhan luka, yaitu: sel Mast berperan dalam meningkatkan respon inflamasi akut, sel Mast menstimulasi proses proliferasi, dan sel Mast meningkatkan pembentukan jaringan parut (Nauta et al., 2013; Weller et al., 2006).
3.1 Sel Mast berperan dalam meningkatkan respon inflamasi akut
Sebagai sel yang banyak didapatkan pada dermis, sel Mast sangat mungkin menjadi sel pengendali inflamasi akut. Walaupun proses pasti mengenai cara aktifasi sel Mast pada kulit yang terluka belum seluruhnya diketahui, beberapa hal yang diduga bisa mencetuskannya antara lain, patogen, produk dari patogen, dan berbagai Sitokin, disamping stimulasi mekanik dan perubah kondisi mekanik.Seperti telah diketahui, bahwa aktivasi dari sel Mast akan menyebabkan dilepaskannya berbagai macam mediator pro inflamasi yang menimbulkan tanda dari proses inflamasi, yaitu vasodilatasi, peningkatan permeabilitas, dan aktifasi/rekrutmen dari sel imunitas dalam darah tepi (Wulff dan Wilgus, 2013). 3.2 Sel Mast menstimulasi proses proliferasi
Sel Mast yang teraktivasi juga diketahui memproduksi Sitokin dan faktor-faktor pertumbuhan yang bisa mendorong terjadinya proliferasi dan migrasi dari beberapa jenis sel kulit, seperti stimulasi keratinosit, membantu restorasi barrier epidermis saat reepiteliasasi, aktivasi Fibroblas dan neovaskularisasi. Yang pada akhirnya akan meningkatkan fase proliferasi pada penyembuhan. Dalam hal ini, penelitian yang dilaporkan masih banyak yang bertentangan satu dengan yang lain, beberapa penelitian menunjukkan bahwa pencegahan degranulasi dari sel Mast tidak mempengaruhi penutupan luka, namun penelitian lain menunjukkan bahwa terjadi penundaan penutupan luka. Hal yang sama juga ditemukan dalam
hubungan antara degranulasi sel Mast dan proses angiogenesis (Harvima dan Nilsson, 2011, Wulff dan Wilgus, 2013).
3.3 Sel Mast meningkatkan pembentukan jaringan parut.
Eksperimen telah menunjukkan bahwa sel Mast mempengaruhi aktivitas dari Fibroblas yang berperan dalam deposisi kolagen dan remodelling saat fase proliferasi dan remodelling penyembuhan luka. Beberapa penelitian menunjukkan bahwa jumlah sel Mast atau sel Mast teraktivasi yang lebih rendah pada jaringan yang luka berkaitan erat dengan derajat jaringan parut yang lebih minimal. Contoh lain adalah pada mukosa mulut, bisa sembuh tanpa jaringan parut, dan dari penelitian histopatologi didapatkan jumlah sel Mast yang jauh lebih sedikit dibandingkan pada kulit. Demikian juga dengan sel kulit fetus, yang memiliki jumlah sel Mast lebih sedikit dan tidak mengalami degranulasi setelah cedera, umumnya akan sembuh tanpa jaringan parut. Beberapa penelitian juga menunjukkan adanya pengaruh dari sel Mast terhadap maturasi dan remodelling kolagen yang berperan terhadap timbulnya jaringan parut(Nauta et al., 2013; Wulff et al., 2012).
Ada beberapa mekanisme yang diduga mendasari pengaruh sel Mast terhadap timbulnya jaringan parut/fibrosis. Sel Mast yang teraktivasi bisa menghasilkan beberapa mediator profibrosis seperti TGF-β dan PDGF, dan beberapa mediator
lain. Histamin juga dapat memicu terjadinya migrasi, proliferasi Fibroblas, dan diferensiasinya menjadi Myofibroblast yang kontraktil. Triptase juga ditemukan dalam jumlah tinggi didalam jaringan parut manusia, dan enzim ini diketahui meningkatkan proliferasi dan kemotaksis Fibroblas, serta menstimulasi pembentukan kolagen, differensiasi Fibroblas menjadi myofibroblast dan
kontraksinya. Chymase diketahui bisa meningkatkan pembelahan prokolagen type I dan pembentukan fibril kolagen (Harvima dan Nilsson, 2011; Weller et al., 2006).
Penelitian terakhir menunjukkan bahwa sel Mast dan fibroblas dapat membentuk gap junction yang heteroseluler, dan memungkinkan komunikasi pintas sel, yang berefek terhadap stimulasi proliferasi, differensiasi menjadi
Myofibroblas dan kontraksi, walaupun pengaruh temuan ini pada kondisi klinis masih belum diketahui pasti (Wulff et al., 2012).
4. Ketotifen
Ketotifen merupakan derivat dari benzocycloheptathiophene yang mempunyai efek anti histamin dan anti anafilaktik. Hal itu ditunjukkan dengan kemampuan Ketotifen dalam mengeblok pelepasan mediator dari sel Mast peritonium tikus secara in vitro. Fungsi Ketotifen antara lain mencegah terjadinya pelepasan Histamin dan Leukotrien dari Basofil dan jaringan paru, merupakan antagonis Histamin pada reseptor H1, mampu menghambat ambilan kalsium, dapat memblokade reaksi anafilaktik kulit pasif, dan untuk mencegah asma baik yang disebabkan oleh obat atau yang disebabkan oleh alergen. Beberapa penelitian tentang Ketotifen menunjukkan efek yang yang bermanfaat dalam terapi asma (Sayeed, 2011).
Ketotifen fumarat merupakan obat antihistamin dan antialergi yang telah diketahui menghambat degranulasi sel Mast melalui mekanisme calcium-dependent, dan memblok Histamin secara non kompetitif pada reseptor H1. Ketotifen telah disetujui oleh FDA sebagai terapi tambahan pada dewasa dan anak diatas 15 tahun dengan asma, dan baru-baru ini FDA mengijinkan Ketotifen dipakai sebagai terapi alergi pada mata (Monument et al., 2012).
4.1 Struktur kimia
Ketotifen memiliki nama bangun yaitu 4-(1-metilpiperidin-ylidene)- 4,9-dihidro-10H-benzo [4,5] siklopenta [1,2-b] tiofen-10-satu hydrogen (E)-butadiana. Formula molekul dari Ketotifen adalah C23H23NO5S, dan memiliki berat molekul 425,49742 (Serna, 2006).
Myofibroblas dan kontraksi, walaupun pengaruh temuan ini pada kondisi klinis masih belum diketahui pasti (Wulff et al., 2012).
4. Ketotifen
Ketotifen merupakan derivat dari benzocycloheptathiophene yang mempunyai efek anti histamin dan anti anafilaktik. Hal itu ditunjukkan dengan kemampuan Ketotifen dalam mengeblok pelepasan mediator dari sel Mast peritonium tikus secara in vitro. Fungsi Ketotifen antara lain mencegah terjadinya pelepasan Histamin dan Leukotrien dari Basofil dan jaringan paru, merupakan antagonis Histamin pada reseptor H1, mampu menghambat ambilan kalsium, dapat memblokade reaksi anafilaktik kulit pasif, dan untuk mencegah asma baik yang disebabkan oleh obat atau yang disebabkan oleh alergen. Beberapa penelitian tentang Ketotifen menunjukkan efek yang yang bermanfaat dalam terapi asma (Sayeed, 2011).
Ketotifen fumarat merupakan obat antihistamin dan antialergi yang telah diketahui menghambat degranulasi sel Mast melalui mekanisme calcium-dependent, dan memblok Histamin secara non kompetitif pada reseptor H1. Ketotifen telah disetujui oleh FDA sebagai terapi tambahan pada dewasa dan anak diatas 15 tahun dengan asma, dan baru-baru ini FDA mengijinkan Ketotifen dipakai sebagai terapi alergi pada mata (Monument et al., 2012).
4.1 Struktur kimia
Ketotifen memiliki nama bangun yaitu 4-(1-metilpiperidin-ylidene)- 4,9-dihidro-10H-benzo [4,5] siklopenta [1,2-b] tiofen-10-satu hydrogen (E)-butadiana. Formula molekul dari Ketotifen adalah C23H23NO5S, dan memiliki berat molekul 425,49742 (Serna, 2006).
Myofibroblas dan kontraksi, walaupun pengaruh temuan ini pada kondisi klinis masih belum diketahui pasti (Wulff et al., 2012).
4. Ketotifen
Ketotifen merupakan derivat dari benzocycloheptathiophene yang mempunyai efek anti histamin dan anti anafilaktik. Hal itu ditunjukkan dengan kemampuan Ketotifen dalam mengeblok pelepasan mediator dari sel Mast peritonium tikus secara in vitro. Fungsi Ketotifen antara lain mencegah terjadinya pelepasan Histamin dan Leukotrien dari Basofil dan jaringan paru, merupakan antagonis Histamin pada reseptor H1, mampu menghambat ambilan kalsium, dapat memblokade reaksi anafilaktik kulit pasif, dan untuk mencegah asma baik yang disebabkan oleh obat atau yang disebabkan oleh alergen. Beberapa penelitian tentang Ketotifen menunjukkan efek yang yang bermanfaat dalam terapi asma (Sayeed, 2011).
Ketotifen fumarat merupakan obat antihistamin dan antialergi yang telah diketahui menghambat degranulasi sel Mast melalui mekanisme calcium-dependent, dan memblok Histamin secara non kompetitif pada reseptor H1. Ketotifen telah disetujui oleh FDA sebagai terapi tambahan pada dewasa dan anak diatas 15 tahun dengan asma, dan baru-baru ini FDA mengijinkan Ketotifen dipakai sebagai terapi alergi pada mata (Monument et al., 2012).
4.1 Struktur kimia
Ketotifen memiliki nama bangun yaitu 4-(1-metilpiperidin-ylidene)- 4,9-dihidro-10H-benzo [4,5] siklopenta [1,2-b] tiofen-10-satu hydrogen (E)-butadiana. Formula molekul dari Ketotifen adalah C23H23NO5S, dan memiliki berat molekul 425,49742 (Serna, 2006).
Gambar 4. Rumus molekul Ketotifen (Sumber : Dewoto, 2009)
4.2 Mekanisme kerja
Infiltrasi dan degranulasi sel Mast memiliki peran dalam proses inflamasi. Degranulasi sel Mast akan melepaskan berbagai macam mediator inflamasi seperti Sitokin, endothelin, growth factor dan enzim proteolitik. Sehubungan dengan antagonis resptor Histamin, beberapa efeknya kemungkinan berhubungan dengan inhibisi terhadap pelepasan sel Mast dan derivat Netrofil mediator inflamasi. Pada banyak penelitian dan kondisi klinis, Ketotifen tercatat mampu mengurangi dreganulasi sel Mast dan mengurangi pelepasan Histamin, protease sel Mast, myeloperoxidase, leukotriens, PAF dan bermacam-macam Prostaglandin. Ketotifen juga menghambat agregasi polimorfonuklear dan migrasi serta mengurangi respon inflamasi. Hal ini secara langsung akan mengurangi fungsi Eosinofil dan viabilitasnya (Khurana et al., 2011).
Ketotifen melakukan blokade secara non kompetitif terhadap ikatan Histamin 1 dengan reseptornya dan menghambat degranulasi sel Mast yang diperantarai oleh kalsium. Ketotifen merupakan agen stabilisator sel Mast yang mencegah degranulasi sel Mast dengan cara mencegah influk transmembran dari ion kalsium. Ketotifen dapat memblokade pelepasan mediator oleh sel Mast tikus secara in vitro. (Khurana et al., 2011). Ketotifen juga memblokade penurunan konsentrasi cyclyc-AMP (c-AMP) yang diperlukan pada saat akhir degranulasi vesikel (Monument et al., 2010).
Ketotifen menghambat produksi Sitokin dari sel TH2. NO adalah modulator sel Mast yang menginduksi aksi pro-inflamasi. Sel Mast juga berperan pada kerusakan ginjal melalui aktivasi lokal sistem renin-angitensin dalam nefrophati IgA. Sekresi Sitokin dari sel Mast dan sel Th2 seperti TGF-B yang memfasilitasi produksi IgA. Disini ada peningkatan IL-4,5,6 yang merupakan Sitokin dari sel TH2 dan sel Mast. Produksi IgA intestinal yang berlebihan diketahui sebagai salah satu penyebab nefrophati IgA. Ketotifen mengaktivasi distribusi NOS di lapisan luar korteks dan glomerulus dan menyebabkan penurunan resistensi pembukuh darah renal (Young-Sun
et al., 2009).
Ketotifen diabsorbsi dengan baik setelah pemberian secara oral, mencapai kadar puncak dalam plasma dalam 2-4 jam setelah pemberian. Namun demikian, belum ada informasi yang cukup mengenai absorbsi dari Ketotifen sediaan tablet lepas lambat. Karena adanya efek first-pass metabolism, bioavailabilitas obat ini hanya 50%. Kadar puncak dalam plasma setelah dosis oral multipel sebesar 1mg, 2 kali sehari adalah 1.92 mg/L pada dewasa dan 3.25 mg/L pada anak-anak, dengan daerah dibawah kurva konsentrasi-waktu 16.98 mg/L dan 20,72 mg/L. Obat ini dilaporkan 75% terikat protein (Grant et al., 1990).
Eliminasi waktu paruh dari Ketotifen yaitu 18,3 ± 6 ,7 jam dan memiliki durasi sekitar 12 jam. Ketotifen memiliki volume distribusi sebesar 56 L/Kg dan 75% berikatan dengan protein plasma. Ketotifen dimetabolisme di hepar dan memiliki rute eliminasi utama di renal, sehingga butuh penyesuaian dosis pada kasus insufisiensi renal dan hepar. Dosis normal untuk orang dewasa yaitu 1-2 mg/12 jam. Dosis anak-anak 0,05 mg/kg/hari (Cuvillo et al., 2006; Criado et al., 2010).
Ketotifen dimetabolisme menjadi Ketotifen-N-glukoronide yang inaktif dan nor-Ketotifen yang aktif secara farmakologi, dan jumlah metabolit-metabolit ini dalam urine adalah sebanyak 50% dan 10% dari dosis, hanya sebanyak 1% yang dikeluarkan sebagai senyawa asal. Pembersihan obat ini dari plasma bersifat bifasik, dengan waktu paruh distribusi 3 jam dan waktu paruh eliminasi 22 jam pada orang dewasa. Pola eliminasi pada anak-anak sama dengan pada dewasa. Tidak tersedia data mengenai efek farmakokinetik obat ini pada usia lanjut atau pada penyakit tertentu (Grant et al., 1990). 4.4 Efek samping
Efek samping Ketotifen sama seperti efek samping AH1. Ketotifen pernah dinyatakan mampu meningkatkan nafsu makan dan menambah berat badan. Kombinasi Ketotifen dengan antidiabetik oral telah dilaporkan dapat menurunkan jumlah Trombosit secara reversibel, karena itu kombinasi kedua obat tersebut harus dihindarkan. Ketotifen harus diberikan secara hati-hati pada pasien yang alergi terhadap obat ini (Dewoto, 2009).
4.5 Ketotifen sebagai stabilisator sel Mast
Ketotifen juga diketahui memiliki efek sebagai Mast cell stabilizer. Zat yang mencegah keluarnya mediator dari sel-sel Mast disebut Mast cell stabilizer. Target
utama di dalam sel yang dihambat saat aktivasi sel Mast adalah PLCγ1 yang berfungsi
sebagai katalisator pada pemecahan membran fosfolipid PIP2 untuk menghasilkan second messanger inositol-1,4,5-triphosphate (IP3) dan DAG. Molekul-molekul ini bertanggung jawab sebagai signal yang mengaktivasi isoform PKC dan pelepasan Kalsium (Ca2 +) yang berasal dari simpanan di intraseluler, hal ini menyebabkan kenaikan sementara Ca2 + bebas di intraseluler yang kemudian memicu masuknya kalsium dari ekstraselular dan diikuti oleh degranulasi dan pelepasan mediator kimia dari sel Mast. Ketotifen menghambat pelepasan mediator sel Mast dengan kombinasi 2 mekanisme, yaitu dengan menghalangi masuknya kalsium ke intraseluler dari ekstra-seluler dan dengan menghalangi penurunan konsentrasi siklik AMP-(cAMP) yang dibutuhkan untuk proses akhir degranulasi vesikel. Kompleksitas dari proses signaling sel Mast hanya sebagian dipahami dan membutuhkan penelitian lebih lanjut (Finn dan Walsh, 2013; Monument et al., 2010).
B. Penelitian yang relevan
Hiperplasia sel Mast sering dijumpai pada jaringan fibrosis. Sel Mast juga diketahui sebagai penghasil beberapa mediator profibrosis, berdasarkan hal tersebut, dipikirkan bahwa dengan menghambat hiperplasi sel Mast dan pelepasan mediator sel Mast pasca cedera otot, akan mengurangi derajat kontraktur dan akan mengurangi hiperplasia myofibroblast pada selaput sendi. Penggunaan Ketotifen sebagai stabilisator sel Mast dalam penelitian ini terbukti efektif dalam mengurangi biomekanika dan manifestasi dari fibrosis selaput sendi pada binatang percobaan kelinci yang mengalami kontraktur pada sendi pasca trauma (Monument et al., 2010).
Weller dan kawan-kawan (2006), meneliti peran sel Mast dalam penyembuhan luka tikus coba, pada percobaan ini mereka menggunakan tikus dengan MC-deficient KitW / Kit WƲ
, tikus normal dengan Kit +/+, dan tikus dengan MC-reconstituted Kit W / Kit WƲ. Hasilnya mereka menemukan bahwa sel Mast dan pelepasan Histamin dibutuhkan untuk untuk penyembuhan luka kulit tikus secara normal, hal ini kontradiktif dengan penelitian oleh Nauta dan kawan-kawan (2013), menggunakan tikus C57BL/6-Kit(W-sh/W-sh), WBB6F1-Kit(W/W-v), and Cpa3-Cre; Mcl-1(fl/fl) dan tikus wild type sebagai kontrol, pada penelitian ini dilakukan eksisi dan kemudian dilakukan penjahitan. Dari percobaan ini,
peneliti berkesimpulan bahwa sel Mast tidak mempengaruhi re-epitelialisasi, ukuran jaringan parut yang terbentuk, ataupun kandungan kolagen dalam jaringan parut, dibandingkan dengan kontrol (Weller et al., 2009).
Penelitian oleh Wulff dan kawan-kawan (2012), menggunakan fetus tikus berusia 15 hari (E15) dibandingkan dengan fetus usia 18 hari (E18), dan fetus E18 dengan defisiensi sel Mast (Kitw/w-v) dibandingkan dengan fetus normal Kit+/+. Mereka menemukan bahwa pada E15, dimana jumlah sel Mast kulit lebih sedikit dan tidak mengalami degranulasi saat terjadi perlukaan, derajat jaringan parut yang terbentuk lebih ringan dibandingkan pada fetus E18, temuan yang sama didapatkan pada fetus dengan defisiensi sel Mast, didapatkan bahwa derajat jaringan parut yang terbentuk lebih ringan dibandingkan dengan fetus normal, dari kedua temuan ini, mereka berkesimpulan bahwa sel Mast berperan dalam pembentukan jaringan parut pada penyembuhan luka fetus (Wulff et al., 2012).
C. Kerangka Pikir
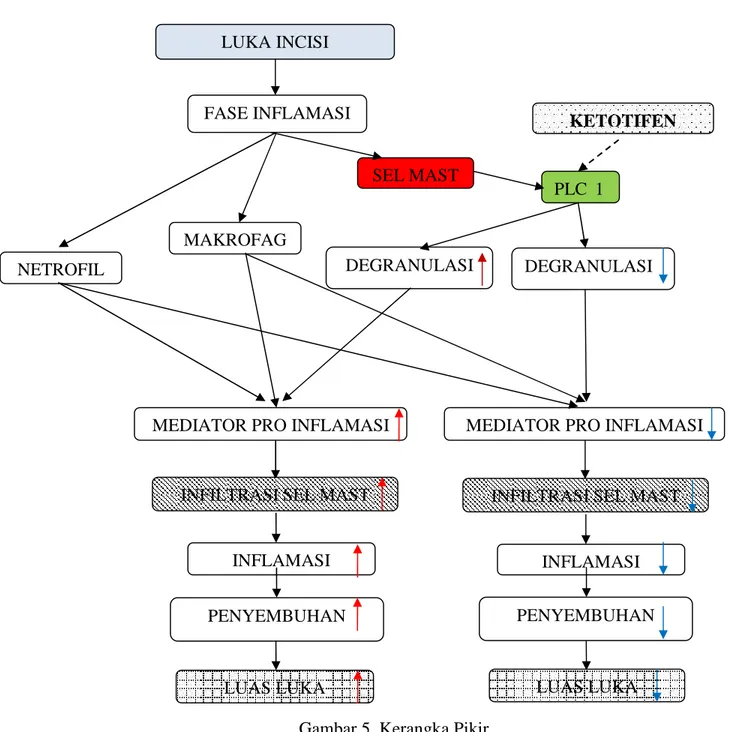
Gambar 5. Kerangka Pikir
INFLAMASI INFLAMASI
LUAS LUKA INFILTRASI SEL MAST LUKA INCISI FASE INFLAMASI SEL MAST MAKROFAG NETROFIL KETOTIFEN DEGRANULASI
MEDIATOR PRO INFLAMASI
LUAS LUKA
DEGRANULASI
MEDIATOR PRO INFLAMASI
INFILTRASI SEL MAST
PLCγ1
PENYEMBUHAN PENYEMBUHAN
Keterangan gambar 5 :
: Ketotifen oral dosis 0.3mg/kg per 12 jam sebagai stabilisator membran sel Mast mempengaruhiPLCγ1 mencegah Ca++keluar dari sel
: Infiltrasi sel Mast di sekitar luka incisi dihitung dengan cara histopatologi : Luas luka diukur setelah hari ke-6 pasca incisi
/ : Perubahan yang terjadi (naik/ turun)
Sesaat setelah dilakukan incisi, fase inflamasi yang melibatkan Netrofil, Makrofag dan sel Mast akan dimulai. Pada proses normal sel Mast akan ter-degranulasi mengeluarkan mediator-mediator pro inflamasi, sehingga waktu terjadinya resolusi fase inflamasi akan memanjang, penyembuhan luka lama, dan luas luka juga menjadi lebih luas. Pada pemberian Ketotifen, degranulasi sel Mast akan menurun sehingga mediator pro inflamasi juga akan menurun, infiltrasi sel Mast berkurang, inflamasi berkurang,, luas luka juga berkurang.
C. Hipotesis
Terdapat pengaruh Ketotifen terhadap infiltrasi sel Mast dan luas luka incisi tikus Wistar yaitu infiltrasi sel Mast berkurang dan luas luka menjadi lebih sempit.