PRODUKTIVITAS SERASAH DAN DEKOMPOSISI SEMI
AEROBIK DAUN MAHONI (Swietenia macrophylla King)
SETYO ANDI NUGROHO
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA*
Dengan ini saya menyatakan bahwa tesis berjudul produktivitas serasah dan dekomposisi semi aerobik daun mahoni (Swietenia macrophylla King) adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, Mei 2014
Setyo Andi Nugroho
RINGKASAN
SETYO ANDI NUGROHO. Produktivitas Serasah dan Dekomposisi Semi Aerobik Daun Mahoni (Swietenia macrophylla King). Dibimbing oleh MUHADIONO dan IMAN RUSMANA.
Mahoni (Swietenia macrophylla King) merupakan salah satu jenis pohon tropis yang paling berharga di dunia. Mahoni dijadikan prioritas utama dalam rangka pembangunan hutan buatan seperti tanaman hutan kota. Pohon mahoni yang ditanam menghasilkan produktivitas cukup banyak. Akan tetapi serasah yang dihasilkan mahoni tidak dimanfaatkan secara maksimal. Serasah bisa dimanfaatkan sebagai kompos dengan cara mendekomposisi. Penelitian ini bertujuan untuk mengkaji produktivitas serasah mahoni, mengkaji pengaruh ukuran potongan daun mahoni, dan mengkaji pengaruh konsentrasi EM4.
Produktivitas serasah pada tegakan mahoni diukur menggunakan litter-trap method. Serasah yang jatuh ditampung menggunakan litter trap berukuran 1m x 1m sebanyak 24 buah yang diletakkan pada lokasi di bawah tegakan mahoni, dengan tinggi trap 50 cm dari permukaan tanah. Dekomposisi semi aerobik dilakukan dengan menyediakan ember dilengkapi jaring. Tutup ember diberi lubang (ventilasi udara) diameter 2 mm jumlah 3 buah. Lubang bekerja untuk memasukkan udara dan mempertahankan kelembaban. Setiap minggu dilakukan pengamatan dan pengukuran suhu serta pH. Pengaruh kompos diuji menggunakan rancangan acak lengkap faktorial dengan dua faktor. Faktor pertama Perlakuan potongan serasah daun mahoni P1 dengan luas daun 25-30 cm2, P2 luas 7-10 cm2, dan P3 luas 0,7-1 cm2. Faktor kedua konsentrasi EM4, K1 adalah rasio larutan gula (100 g gula dilarutkan 1 liter air) yaitu K1 sebanyak 5 ml, K2 dengan 10 ml dan K3 dengan 20 ml. Data dianalisis menggunakan uji F pada tingkat kepercayaan 95%. Apabila ada perlakuan nyata selanjutnya dilakukan uji Duncan (DMRT).
Isolasi bakteri menggunakan 4 media selektif, yaitu MRSA untuk
Lactobacillus sp; SCA untuk Actinomycetes sp; medium sistrom’s untuk bakteri fotosintetik, dan PDA untuk yeast. Analisis produktivitas serasah menggunakan rerata serasah yang dikumpulkan selama periode percobaan. Analisis dekomposisi semi aerobik menggunakan perbedaan penurunan bobot serasah. Pada akhir percobaan dilakukan uji kadar C organik, N, P, dan K.
Hasil penelitian menunjukkan produktivitas serasah tertinggi terjadi pada minggu ke-16 sebesar 89,24 g/m2/minggu pada bulan mei-juni yaitu bertepatan dengan musim kemarau. Total produktivitas serasah terbesar terdapat pada pohon 2 yaitu pohon berdiameter 33 cm, tinggi 20 m, dan tebal tajuk 5,5 m dengan total produktivitas 1415 g/m2/minggu. Hasil penelitian menunjukkan perlakuan P3K2 lebih efisien pada minggu ke-7 karena sudah berwarna hitam, dengan konsentrasi EM4 lebih rendah dibanding perlakuan P3K3. Hasil terbaik dekomposisi daun mahoni adalah potongan P3 dengan luas daun 0,7-1 cm2, sedang proses dekomposisi paling efisien pada konsentrasi K2 yaitu larutan gula (100 gr gula dilarutkan 1 liter air) dengan 10 ml EM4.
SUMMARY
SETYO ANDI NUGROHO. Litterfall Production and Semi Aerobic Decomposition of Leaf Mahogany (Swietenia macrophylla King). Supervised by MUHADIONO and IMAN RUSMANA.
Mahogany (Swietenia macrophylla King) is one of the most valuable tropical tree in the world, due to their potential use in the development of artificial forest such as a forest city. Mahogany trees produced enough litter, however the litter was not used maximally. These litter can be used as a compost fertilizer after composting decomposition. This research aims were to assess the productivity of litter mahogany, to evaluate the effect of leaf size mahogany and addition of EM4. Mahogany litter productivity was measured using litter-trap method. Litter leaf fall was trapped by 1m x 1m sample plot. There were 24 sample plots placed under mahagony stands. The plot height was 50 cm above the soil surface. Semi aerobic decomposition was set up in bucket with nets. The cover of the bucket had 3 holes with a diameter of 2 mm for air ventilation and maintaining moisture. Temperature and pH were monitored weekly. The experiment was set up with two factors completely randomized design. The first factor was the size of litter with 3 different sizes i.e. 25-30 cm2 (P1), 7-10 cm2 (P2) and 0,7-1 cm2 (P3). The second factor was microbes (EM4) addition with 3 different volume i.e. 5 ml (K1), 10 ml (K2) and 20 ml (K3). The microbes (EM4) was prepared with 100 g of sugar in 1 liter water. Data were analyzed using the F test at the 95% confidence level. If there was significancy, then continued with DMRT.
Enumeration of bacterial groups was conducted using 4 selective media i.e. MRSA media for Lactobacillus sp, SCA for Actinomycetes sp, sistrom’s media for photosynthetic bacteria, and PDA for yeast. Productivity of litterfall was determined from the average of collected litter. The reduction rate of semi aerobic litter decomposition were used to show the decomposition rate. Total concentration organic carbon, nitrogen, phosphate and potash were analised at the end of experiment.
The results showed that the highest litter productivity of mahogany at sixteen weeks of composting periode, it was 89.24 g/m2/week in May-June (the dry season). The highest litter productivity was found in tree no 2 (with 33 cm in diameter, 20 m in height, and 5.5 m crown thickness). Total productivity was 1415 g/m2/week. The results showed that P3K2 treatment was the most efficient decomposition process, it was shown at seven week of composting periode. This treatment was combination of 0.7-1 cm2 litter size and addition of 10 ml EM4.
© Hak Cipta Milik IPB, Tahun 2014
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains
pada
Program Studi Biologi Tumbuhan
PRODUKTIVITAS SERASAH DAN DEKOMPOSISI SEMI
AEROBIK DAUN MAHONI (Swietenia macrophylla King)
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
BOGOR 2014
Judul Tesis : Produktivitas Serasah dan Dekomposisi Semi Aerobik Daun Mahoni (Swietenia macrophylla King)
Nama : Setyo Andi Nugroho
NIM : G353110131
Disetujui oleh
Komisi Pembimbing
Dr Ir Muhadiono, MSc Ketua
Dr Ir Iman Rusmana, MSi Anggota
Diketahui oleh
Ketua Program Studi Biologi Tumbuhan
Dr Ir Miftahudin, MSi
Dekan Sekolah Pascasarjana
Dr Ir Dahrul Syah, MScAgr
PRAKATA
Puji dan syukur penulis panjatkan kepada Allah Subhanahu Wa Ta’ala atas segala karunia-Nya sehingga karya ilmiah ini berhasil diselesaikan. Judul yang dipilih dalam penelitian ini adalah Produktivitas Serasah dan Dekomposisi Semi Aerobik Daun Mahoni (Swietenia macrophylla King). Penelitian ini berlangsung dari bulan Februari 2013 sampai Desember 2013.
Penulis mengucapkan terima kasih kepada Bapak Dr Ir Muhadiono, MSc dan Bapak Dr Ir Iman Rusmana, MSi selaku pembimbing yang telah banyak memberikan nasihat, saran serta bimbingan. Ucapan terimakasih penulis sampaikan kepada Bapak Zainal atas segala ilmu dan bantuannya dalam proses mencari data dihutan penelitian Dramaga Bogor. Terimakasih juga penulis ucapakan kepada Bapak Jaka dan Ibu Heni selaku teknisi laboratorium mikrobiologi IPB yang telah membantu penulis dalam menyelesaikan isolasi bakteri.
Penulis juga mengucapakan terima kasih kepada bapak dan ibu pengajar Biologi Tumbuhan (BOT) atas semua ilmu, pengalaman, bimbingan, dan nasihat selam ini. Ucapan terimakasih untuk teman-teman Biologi Tumbuhan angkatan 2011 atas kebersamaan, kecerian, kehangatan dan semangat yang telah diberikan. Serta untuk seluruh teman-teman perkumpulan beasiswa unggulan (BU) atas dukungan dan persahabatan selama ini. Ucapan terimakasih penulis berikan kepada orang tua, adik-adik, dan seluruh keluarga atas segala doa, kasih sayang, semangat, dan dukungannya selama ini.
Semoga karya ilmiah ini dapat bermanfaat bagi pengembangan ilmu pengetahuan.
Bogor, Mei 2014
DAFTAR ISI
DAFTAR TABEL vii
DAFTAR GAMBAR vii
DAFTAR LAMPIRAN vii
1 PENDAHULUAN 1
Latar Belakang 2
Tujuan Penelitian 2
Manfaat Peneliitian 2
Hipotesis 2
2 METODE 3
Waktu dan Tempat Penelitian 3
Alat dan bahan 3
Metode Pengumpulan Data Vegetasi 3
Metode Pengumpulan Data produktivitas Serasah 4
Metode Pengomposan Semi Aerobik 4
Metode Isolasi Bakteri Serasah Mahoni 5
Analisis Data Produktivitas 5
Analisis Data Laju Dekomposisi Semi Aerobik 5
Analisis Data Pengaruh Pengomposan Semi Aerobik 6
3 HASIL DAN PEMBAHASAN 6
HASIL 6
Produktivitas Serasah 6
Dekomposisi Serasah 8
PEMBAHASAN 11
Produktivitas Serasah 11
Dekomposisi Serasah 14
4 SIMPULAN DAN SARAN 17
DAFTAR PUSTAKA 18
LAMPIRAN 23
DAFTAR TABEL
1 Tinggi pohon, diameter pohon, dan laju produktivitas serasah daun mahoni 8 2 Pengaruh potongan daun dan konsentrasi EM4 terhadap laju dekomposisi
semi aerobik 8
3 Warna kematangan hasil dekomposisi semi aerobik 9 4 Konsentrasi unsur hara daun mahoni minggu ke-10 10
DAFTAR GAMBAR
1 Produktivitas serasah dan parameter lingkungan tegakan mahoni (Swietenia
macrophylla King) di Kebun Percobaan Dramaga Bogor 7
2 Temperatur dan pH dekomposisi semiaerobik daun mahoni 10 3 Perhitungan koloni bakteri EM4 minggu ke-10 dekomposisi semiaerobik
daun Mahoni 11
DAFTAR LAMPIRAN
1. Analisis ragam laju dekomposisi serasah mahoni (Swietenia macrophylla
King) 23
2. Hasil dekomposisi semi aerobik daun mahoni 24 3. Koloni bakteri minggu ke-10 dengan medium selektif 26 4. Penelitian lain mengenai scanning electron microscopy (SEM) bakteri yang
terlibat dalam EM4 27
1
PENDAHULUAN
Latar Belakang
Mahoni (Swietenia macrophylla King) merupakan salah satu jenis pohon tropis yang paling berharga di dunia (Rodan et al. 1992; Veríssimo et al. 1995). Mahoni dijadikan prioritas utama dalam rangka pembangunan hutan buatan seperti tanaman hutan kota. Pohon mahoni yang ditanam pada hutan kota menghasilkan produktivitas serasah cukup banyak. Akan tetapi serasah yang dihasilkan mahoni tidak dimanfaatkan dengan maksimal. Serasah bisa dimanfaatkan sebagai kompos dengan cara dekomposisi serasah. Dalam pembuatan kompos tidak terlepas dari produksi serasah dan dekomposisi serasah. Produksi serasah dan dekomposisi serasah adalah dua sarana penting dalam sebuah ekosistem (Karma 1970).
Produktivitas serasah dan dekomposisi serasah merupakan rantai dasar ekosistem secara fungsional dalam sebuah vegetasi. Produksi serasah dan dekomposisi adalah proses yang saling berkaitan dan memberikan umpan balik positif (Kitayama et al. 2004). Dekomposisi memberikan nutrisi yang diperlukan untuk produktivitas serasah dengan daur ulang bahan organik, sedang peningkatan biomassa tanaman berhubungan positif untuk menyediakan substrat dalam proses dekomposisi serasah. Oleh karena itu, laju dekomposisi dapat mengatur siklus nutrisi di komunitas tumbuhan dan dapat memperkirakan produktivitas bersih serasah secara tidak langsung (Clark et al. 2001).
Serasah merupakan salah satu jalur utama dari siklus unsur hara (Vitousek 1984). Ketersediaan unsur hara penting bagi pertumbuhan tanaman secara normal. Hilangnya beberapa unsur hara dari daerah perakaran akan menyebabkan kesuburan tanah menurun sehingga tanah tidak mampu mendukung pertumbuhan tanaman secara normal. Ketersediaan unsur hara yang cukup dan seimbang diindikasikan oleh perbedaan produksi serasah dan dekomposisi serasah. Serasah memindahkan unsur hara, nutrisi, dan energi dari vegetasi ke dalam tanah dan ini merupakan paling dominan dalam siklus biogeokimia (Facelli dan Pickett 1991; Liu et al. 2004). Pengetahuan ini bersifat komprehensif dari dekomposisi bahan organik dan pola pelepasan hara dari serasah daun memaksimalkan keberlanjutan produktivitas tanah dan tanaman.
Produktivitas dan dekomposisi serasah merupakan proses sangat komplek melibatkan beberapa faktor (Dezzeo et al. 1998). Salah satunya ditentukan oleh kualitas dan lingkungan fisik (Villela dan Proctor 2002). Faktor lingkungan produktivitas serasah meliputi kecepatan angin, suhu, dan kelembaban, sedang dekomposisi serasah meliputi fakor pH, temperatur, dan komposisi kimia serasah dan mikroorganisme tanah (Van Breemen 1995; Aerts dan Caluwe 1997; Saetre 1998; Guo dan Sims 1999). Selain lingkungan faktor lain yang mempengaruhi laju dekomposisi serasah dan produksi serasah adalah iklim, musim, kualitas serasah, substrat dan jenis vegetasi (Melillo et al. 1982; Upadhyay et al. 1989; Hobbie 1992; Vitousek et al. 1994).
2
aerobik memberi keuntungan yaitu mempercepat biodegradasi senyawa organik dan stabilisasi (Robison dan Peter 1983). Sistem semi aerobik menguntungkan dengan nitrifikasi dan denitrifikasi terjadi secara bersamaan dan mempercepat proses stabilisasi (Theng et al. 2005).
Spesies yang tumbuh pada lingkungan miskin hara lebih sulit terdekomposisi dan menyebabkan lambatnya proses siklus hara pada lingkungan tersebut dibanding serasah berasal dari tanaman hidup pada lingkungan kaya hara (Van Breemen 1995; Aerts dan Caluwe 1997). Dekomposisi serasah lebih cepat pada kondisi aerobik dibanding kondisi anaerobik (Johnson dan Damman 1991). Serasah yang berada pada daerah mempunyai jumlah mikro organisme banyak cenderung lebih cepat terdekomposisi dibanding pada daerah mempunyai jumlah mikro organisme sedikit (Saetre 1998).
Proses dekomposisi dalam mempercepat kematangannya ditambahkan dengan effective innoculant atau aktivator (Saptoadi 2001). Salah satu bahan aktivator yang digunakan adalah Effective Microorganism 4 (EM4). Penggunaan mikrobia terpilih EM4 mempercepat dekomposisi bahan organik dari 3 bulan menjadi 7-14 hari. EM4 merupakan kultur campuran mikrobia terpilih seperti
Lactobacillus sp, bakteri fotosintetik, Actinomycetes sp dan yeast yang bekerja secara sinergik dalam proses dekomposisi (Higa dan Wididana 1994).
Tujuan Penelitian
Penelitian dilakukan dengan tujuan sebagai berikut :
1. Mengkaji produktivitas serasah mahoni (Swietenia macrophylla King). 2. Mengkaji pengaruh ukuran potongan daun mahoni (Swietenia macrophylla
King) terhadap dekomposisi semi aerobik.
3. Mengkaji pengaruh konsentrasi EM4 terhadap dekomposisi semi aerobik
Manfaat Penelitian
Hasil penelitian diharapkan memberi informasi tentang produktivitas serasah dan dekomposisi serasah semi aerobik.
Hipotesis
Hipotesis penelitian sebagai berikut:
1. Produktivitas serasah tinggi terjadi pada saat musim kemarau, kecepatan angin tinggi, temperatur udara tinggi, dan kelembaban udara rendah. 2. Terdapat pengaruh luas potongan daun terhadap dekomposisi serasah daun
mahoni (Swietenia macrophylla King).
3
2
METODE
Waktu dan Tempat Penelitian
Penelitian dilaksanakan bulan Februari sampai Desember 2013. Penelitian dilakukan di beberapa tempat yaitu produktivitas serasah dilakukan di Kebun Percobaan Dramaga Bogor, kemudian dekomposisi serasah dilakukan di laboratorium Ekologi Departemen Biologi FMIPA IPB, dan isolasi bakteri dilakukan di laboratorium Mikrobiologi FMIPA IPB serta analisis uji kandungan C organik, N, P, dan K di laboratorium Ilmu Tanah BIOTROP.
Alat dan bahan
Peralatan dan bahan penunjang yang digunakan dalam penelitian produktivitas serasah yaitu pita ukur 1,5 meter dan 25 meter, litter-trap (alat penampung serasah) terbuat dari kain kasa/nylon berukuran 1 m x 1 m x 0,5 m, tali plastik, patok bambu, timbangan, cangkul, golok, spiegel relaskop, 4 in 1. Sedang alat untuk dekomposisi yaitu ember dengan ukuran volume 21 liter, kantong plastik dan oven, termometer, 4 in 1, pH meter, gelas ukur 1000 ml, spray dengan ukuran 1000 ml, dan timbangan analitik. Bahan utama yang digunakan pada penelitian adalah serasah dari tegakan Mahoni (Swietenia macrophylla King) dan media isolasi mikro organisme meliputi 4 media isolasi yaitu media MRSA
terdiri protein dari kasein, ekstrak daging, ekstrak ragi, glukosa, magnesium sulfat, agar bacto, dipotassium hidrogen, phosphate, tween 80, diamonium hidrogen sitrat, natrium asetat, mangan sulfat, kemudian medium soluble-casein Agar 100 ml dengan komposisi 1 g soluble starch, 0,03 g casein, 0,2 g KNO3, 0,2 karbon, dan medium PDA 100 ml dengan komposisi kentang 25 g, agar bacto 2 g, dan dextrose 2 g.
Metode Pengumpulan Data Vegetasi
Serasah pada tegakan Mahoni dikumpulkan menggunakan litter-trap
method (Metode alat penampung serasah). Langkah pelaksanaan yaitu
4
Metode Pengumpulan Data produktivitas Serasah
Pengambilan serasah daun dilakukan dengan menempatkan ke dalam plastik untuk masing-masing trap. Pengambilan serasah setiap hari minggu sekitar pukul 08.00-10.00 WIB. Serasah yang diamati terdiri helaian daun (lamina) dan tangkai daun (petiol). Setiap pengambilan daun, dilakukan perhitungan jumlah daun dan penimbangan berat kering daun. Pengambilan daun hanya dilakukan pada daun mahoni (Swietenia macrophylla King), untuk daun komunitas lain yang tertampung dalam trap tidak dilakukan perhitungan dan penimbangan. Perlakuan daun yang didapatkan dalam kondisi basah, dilakukan pengeringan udara ( air-dry), terlebih dahulu selama 1 hingga 2 minggu. Kemudian, perlakuan daun dibungkus kertas untuk dimasukkan ke dalam oven bersuhu 105oF (60oC) selama 24 jam sehingga didapatkan berat kering daun konstan. Perhitungan berat kering daun dihitung per satuan luas trap per waktu pengambilan (g/m2/minggu).
Data iklim selama penampungan serasah daun dilakukan pengamatan dan pengumpulan yaitu meliput temperatur udara, kelembaban udara, dan kecepatan angin. Pengukuran tinggi dan diameter pohon sebagai data penunjang dilakukan dengan mengukur tinggi masing-masing pohon menggunakan spiegel relaskop
dan mengukur diameter pohon menggunakan meteran. Pengamatan tinggi pohon diukur tinggi tajuk sampai ujung dan tinggi tajuk bebas cabang untuk mengetahui tebal tajuk pohon.
Metode Pengomposan Semi Aerobik
Pengomposan semi aerobik yaitu menyediakan ember dilengkapi jaring (rajut). Fungsi jaring (rajut) untuk memisahkan antara serasah padat dengan bagian cair serta mempermudah mengambil serasah di ember. Penutup ember diberi lubang (ventilasi udara) dengan diameter 2 mm sebanyak 3 buah. Lubang berfungsi memasukkan udara dalam rangka menjaga kelembaban. Setiap seminggu sekali dilakukan pengamatan dan pengukuran suhu dan pH. Proses dekomposisi serasah dilakukan selama 10 minggu.
5
Metode Isolasi Bakteri Serasah Mahoni
Isolasi bakteri diambil dari hasil deomposisi serasah pada bagian padat dengan perlakuan potongan daun Mahoni (P1,P2,P3) dan 3 konsentrasi EM4 (K1,K2,K3). Isolasi menggunakan 4 medium selektif mengindikasikan bakteri tertentu yang tumbuh dari aktivator EM4. Isolasi bakteri Lactobacillus sp menggunakan medium deMann Rogosa Sharpe Agar (MRSA), kemudian bakteri
Actinomycetes sp menggunakan medium soluble-casein, selanjutnya bakteri fotosintetik menggunakan medium Sistrom's, dan jamur fermentasi menggunakan medium PDA yaitu media berguna untuk menumbuhkan yeast pada perlakuan.
Analisis Data Produktivitas
Nilai tengah (rata-rata) produktivitas serasah per plot diperoleh dari setiap pengamatan dengan rumus :
gr/m2/minggu
Dimana:
Xj : rata-rata produksi serasah per plot setiap periode (minggu)
Xi : produksi serasah per plot setiap periode n : (trap)
Analisis Data Laju Dekomposisi Semi Aerobik
Analisis laju dekomposisi semi aerobik menggunakan rumus adalah sebagai berikut :
Penurunan bobot didapat dengan rumus:
W = W0 – Wt x 100% W0
Dimana :
W0 = berat kering awal serasah
Wt = berat kering akhir serasah (gram) per periode waktu t W = Penurunan bobot
Laju dekomposisi diduga dengan rumus:
D = Penurunan bobot Minggu
6
Analisis Data Pengaruh Pengomposan Semi Aerobik
Pengaruh potongan luas daun dan konsentrasi EM4 dalam sistem kompos diukur melalui rancangan Percobaan Dua Faktor dalam Rancangan Acak Lengkap (RAL) atau Faktorial RAL. Penelitian mengkaji perbedaan luas potongan serasah daun Mahoni (P1,P2,P3) dengan pemberian konsentrasi EM4 (K1,K2,K3). Dengan demikian ada sebanyak 3x3= 9 kombinasi perlakuan. Setiap kombinasi perlakuan dilakukan pengulangan 3 kali. Banyak unit percobaan adalah 9x3 = 27 unit. Seluruh ember yang digunakan dianggap seragam. Data diperoleh dari hasil pengamatan dianalisis menggunakan analisis ragam (Uji F) pada tingkat kepercayaan 95%. Apabila ada perlakuan nyata dilakukan uji Duncan (DMRT).
3
HASIL DAN PEMBAHASAN
HASIL
Produktivitas Serasah
Uji produktivitas serasah dilakukan di Kebun Percobaan Dramaga Bogor. Kebun percobaan menurut administrasi pemerintahan termasuk ke dalam wilayah Situ Gede dan Bubulak, Kecamatan Bogor Barat, Kotamadya Bogor. Batasan secara geografis lokasi penelitian terletak pada 6 033’8” sampai dengan 6 033’38” LS dan 106 0 44’50” sampai dengan 106 0105’19” BT. Jarak lokasi dari Bogor sekitar 9 km ke arah Barat.
Luas kebun percobaan 60 ha, status kebun penelitian milik Departemen Kehutanan RI Badan Penelitian dan Pengembangan Kehutanan. Iklim rata-rata curah hujan setahun 3.552 mm, hari hujan 187. Temperatur maximum rata-rata: 30,10oC minimum rata-rata 20,10oC dengan rata-rata kelembaban 88,33%. Menurut klasifikasi iklim Schmidt dan Ferguson (1951) termasuk type A yaitu iklim tropika basah.
7
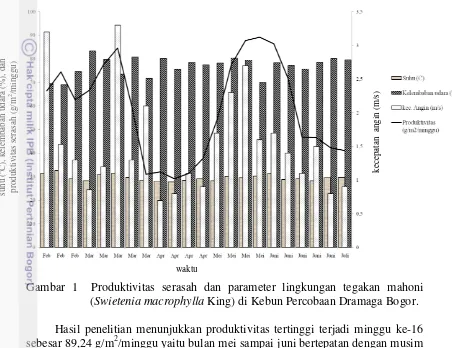
Pengamatan produktivitas serasah di Kebun Percobaan Dramaga Bogor dilakukan selama 22 minggu. jumlah pohon yang diamati sebanyak 6 pohon, setiap pohon ada 4 trap (alat penampung serasah), sehingga trap yang digunakan penelitian sebanyak 24 trap. Jarak tanam pohon satu dengan lain yaitu berjarak 5 meter. Hasil produktivitas serasah ditunjukkan Gambar 1 sebagai berikut:
Gambar 1 Produktivitas serasah dan parameter lingkungan tegakan mahoni (Swietenia macrophylla King) di Kebun Percobaan Dramaga Bogor.
Hasil penelitian menunjukkan produktivitas tertinggi terjadi minggu ke-16 sebesar 89,24 g/m2/minggu yaitu bulan mei sampai juni bertepatan dengan musim kemarau. Produktivitas terendah terjadi minggu ke-10 sebesar 29,12 g/m2/minggu bertepatan musim hujan. Produktivitas serasah tinggi dijumpai pada musim hujan yaitu bulan maret atau minggu ke-6 sebesar 84,61 g/m2/minggu.
8
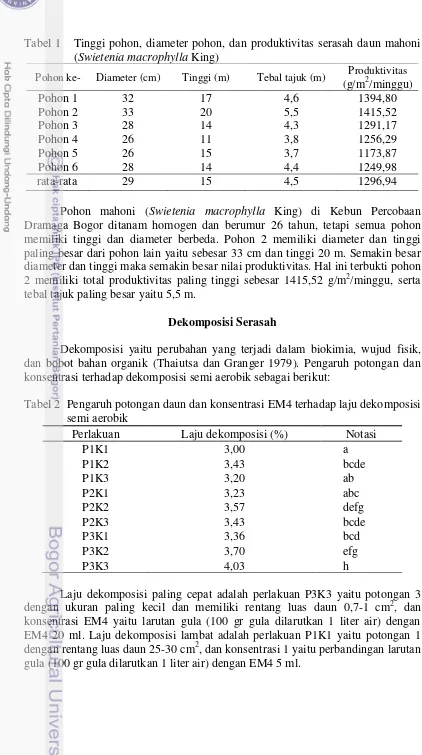
Tabel 1 Tinggi pohon, diameter pohon, dan produktivitas serasah daun mahoni (Swietenia macrophylla King)
Pohon ke- Diameter (cm) Tinggi (m) Tebal tajuk (m) Produktivitas (g/m2/minggu)
Pohon mahoni (Swietenia macrophylla King) di Kebun Percobaan Dramaga Bogor ditanam homogen dan berumur 26 tahun, tetapi semua pohon memiliki tinggi dan diameter berbeda. Pohon 2 memiliki diameter dan tinggi paling besar dari pohon lain yaitu sebesar 33 cm dan tinggi 20 m. Semakin besar diameter dan tinggi maka semakin besar nilai produktivitas. Hal ini terbukti pohon 2 memiliki total produktivitas paling tinggi sebesar 1415,52 g/m2/minggu, serta tebal tajuk paling besar yaitu 5,5 m.
Dekomposisi Serasah
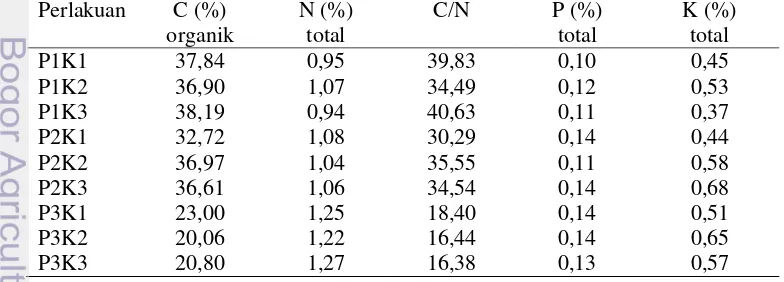
Dekomposisi yaitu perubahan yang terjadi dalam biokimia, wujud fisik, dan bobot bahan organik (Thaiutsa dan Granger 1979). Pengaruh potongan dan konsentrasi terhadap dekomposisi semi aerobik sebagai berikut:
Tabel 2 Pengaruh potongan daun dan konsentrasi EM4 terhadap laju dekomposisi semi aerobik
Perlakuan Laju dekomposisi (%) Notasi
9
Tabel 3 Warna kematangan hasil dekomposisi semi aerobik daun mahoni
Minggu ke-
Ket: 1. Warna asli (tingkat kematangan 0-25 %) 2. Cokelat (tingkat kematangan 26%-50%) 3. Cokelat Pekat (tingkat kematangan 51%-75%) 4. Hitam (Tingkat kematangan 76%-100%)
Karakteristik yang digunakan untuk menilai kematangan dan kualitas kompos salah satunya meliputi warna (Lekasi et al. 2003). Berdasarkan hasil penelitian menunjukkan bahwa perlakuan P3K2 lebih efisien karena pada minggu ke-7 sudah berwarna hitam dengan kebutuhan konsentrasi EM4 rendah dibandingkan dengan perlakuan P3K3. Dekomposisi semi aerobik matang ditunjukkan minggu terakhir perlakuan yaitu perlakuan P3K1, P3K2 dan P3K3.
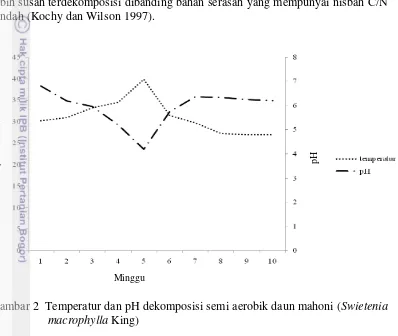
Dekomposisi serasah dipengaruhi oleh beberapa kondisi lingkungan, kualitas kimia serasah, dan adanya kelimpahan organisme pengurai dari sebuah vegetasi (Polyakova dan Billor 2007). Faktor dominan yang mempengaruhi aktivitas mikroorganisme dalam perombakan dan penguraian serasah adalah jenis tanaman dan iklim. Efek terhadap jenis tanaman terhadap mikroflora ditentukan oleh sifat fisik dan kimia daun, keduanya tercermin dalam C/N rasio (Thaiutsa dan Granger 1979). Berikut konsentrasi unsur hara daun mahoni pada proses dekomposisi semi aerobik minggu ke-10 sebagai berikut:
10
Nilai rasio C/N bahan organik daun serasah mahoni paling efisien yaitu perlakuan potongan 3 dengan rentang luas 0,7-1 cm2, memiliki nilai rasio C/N kecil dibandingkan dengan perlakuan lain yaitu 16,38 sampai 18,40. Standar Nasional Indonesia (SNI) kompos matang berkisar antara 10-20%. Prinsip pengomposan yaitu nilai rasio C/N bahan organik menjadi sama dengan rasio C/N tanah. Rasio C/N adalah hasil perbandingan antara karbohidrat dan nitrogen yang terkandung di dalam bahan serasah. Bahan serasah mempunyai nisbah C/N tinggi lebih susah terdekomposisi dibanding bahan serasah yang mempunyai nisbah C/N rendah (Kochy dan Wilson 1997).
Gambar 2 Temperatur dan pH dekomposisi semi aerobik daun mahoni (Swietenia macrophylla King)
Pengukuran temperatur dan pH dilakukan selama 10 minggu. Hasil penelitian menunjukkan pengukuran temperatur pada minggu pertama sebesar 30oC, kemudian naik dengan cepat dan mencapai temperatur puncak pada minggu kelima sebesar 39oC, tumpukan serasah kemudian mengalami fase pendinginan dan pematangan yang ditandai dengan penurunan temperatur dari temperatur puncak menuju kestabilan. Pengukuran pH minggu pertama 6,8 kemudian minggu kelima mengalami penurunan sejalan dengan kenaikan temperatur yaitu sebesar 4,2 dan naik kembali menuju kestabilan saat kompos mendekati matang atau matang.
pH
Minggu
tem
p
er
atu
r
(
11
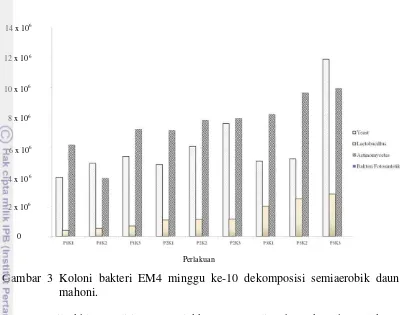
Gambar 3 Koloni bakteri EM4 minggu ke-10 dekomposisi semiaerobik daun mahoni.
Hasil akhir penelitian menunjukkan yeast paling banyak pada perlakuan P3K3 dengan total koloni 11,9 x 106, sedangkan Lactobacillus paling banyak pada perlakuan P3K3 dengan total koloni 2,84 x 106, selanjutnya bakteri Actinomycetes paling banyak pada perlakuan P3K3 dengan total koloni 9,94 x 106, dan tidak terdapat bakteri fotosintetik pada pengamatan akhir minggu ke-10.
PEMBAHASAN Produktivitas Serasah
Produktivitas serasah dipengaruhi oleh faktor fisik seperti proses mekanik dari hujan, angin dan fisiologis respon tanaman terhadap perubahan lingkungan (Santiago dan Mulkey 2005). Berdasarkan hasil penelitian menunjukkan bahwa produktivitas tertinggi terjadi pada minggu ke-16 sebesar 89,24 g/m2/minggu yaitu bulan mei sampai juni bertepatan dengan musim kemarau. Produktivitas terendah pada minggu ke-10 sebesar 29,12 g/m2/minggu bertepatan dengan musim hujan. Produktivitas serasah tinggi dijumpai pada musim hujan yaitu bulan maret atau minggu ke-6 sebesar 84,61 g/m2/minggu. Musim memiliki pengaruh kuat dalam penentuan jumlah serasah, musim kemarau jumlah produktivitas serasah tinggi, sedangkan musim hujan produktivitas serasah rendah, namun akan berbeda dengan tipe hutan lain (Hopkins 1966; John 1973; Songwe et al. 1988; Muoghalu et al. 1993). Produktivitas serasah meningkat dan mencapai maksimum pada musim kemarau dan menurun pada musim hujan, hal ini terjadi karena pada musim kemarau terjadi persaingan antar tanaman dan antar organ dalam suatu tanaman untuk mendapatkan cahaya matahari, sehingga menyebabkan terjadinya
Perlakuan 2 x 106
4 x 106 6 x 106 8 x 106 10 x 106 12 x 106 14 x 106
12
efisiensi dalam proses fotosintesis dan tanaman cepat melakukan regenerasi (Sallata et al. 1990). Tingkat serasah di areal kebun Percobaan Dramaga musim hujan puncak serasah minggu ke-6 sebesar 84,61 g/m2/minggu, berbeda pada musim kemarau tingkat serasah merata. Penelitian ini sejalan yang dilakukan oleh Cuevas dan Medina (1986), serta Barbosa dan Fearnside (1996) menemukan tingkat serasah merata selama musim kemarau. Beberapa penelitian lain mengungkapkan bahwa musim kemarau adalah puncak serasah, hal ini karena tanaman mengalami stres air dan respon vegetatif musiman (Arato et al. 2003). Pohon mahoni saat musim kemarau menggugurkan daun dalam jumlah banyak, tetapi hasil penelitian juga menunjukkanjumlah serasah banyak saat musim hujan (Gambar 1) karekteristik ini menjadi catatan tersendiri oleh Arato et al. (2003) dan Ara´ujo et al. (2002) di hutan dengan iklim musim antara intensitas curah hujan sedang dan tinggi. Menurut Luiz˜ao (1989) ada hubungan positif selama hujan yang intens ditambah dengan angin kencang yang diikuti kekeringan singkat menghasilkan tingkat serasah yang tinggi.
Produksi serasah berhubungan dengan faktor lingkungan (Finer 1996; Hart
et al. 1992), faktor lingkungan yang diukur dalam penelitian yaitu kecepatan angin, kelembaban udara, dan suhu. Berdasarkan hasil penelitian kecepatan angin memiliki pengaruh terhadap besar kecilnya produktivitas serasah, semakin besar kecepatan angin sebesar 2,7 m/s semakin besar pula produktivitas serasah sebesar 87,77 g/m2/minggu. Angin menyebabkan daun berdekatan bergesekan satu sama lain, menciptakan berbagai macam kerusakan. Thompson (1974) melaporkan penelitian mengenai tumbuhan Festuca arundinacea Schreb dengan perlakuan lubang angin sebesar 3,5 m/s dan dilihat secara mikroskopis terjadi kerusakan pada bagian epidermis. Kerusakan menyebabkan pecahnya epidermal sel, keretakan pada kutikula, dan semakin menipisnya lapisan lilin. Dalam percobaan menunjukkan bahwa daun rusak akan kehilangan banyak kapasitas untuk kontrol kehilangan air (Grace 1974). Angin menyebabkan kerusakan daun dan telah dilaporkan dapat menyebabkan daun mengering (Boodle 1920; Tsuboi 1961). Angin menyebabkan daun bergesekan sehingga menimbulkan goresan, luka, dan pecah (Aston dan Bradshaw 1966; Taylor dan Sexton 1972; Wilson 1980), dilihat secara makroskopik kerusakan bahkan lebih jelas ketika angin membawa partikel pasir, tanah atau es (Fryrear 1971; Armbrust et al. 1974). Pengaruh lain yang sering disebut dalam literatur adalah stomata yang segera menutup dalam menanggapi perlakuan angin (Martin dan Clements 1935; Tranquillini 1969; Caldwell 1970).
13
tinggi dapat meningkatkan stres pada tanaman (Vose dan Allen 1991). Kondisi kekeringan merupakan salah satu faktor yang dapat menekan pertumbuhan (Schwanz dan Polle 2001). Gejala pertama yang disebabkan oleh kekeringan ialah penurunan potensial air kemudian diikuti oleh penutupan stomata (Chaves 1991; Brodribb dan Holbrook 2003) sehingga menyebabkan pengambilan CO2 proses fotosintesis terhambat yang akhirnya menurunkan laju fotosintesis (Lawlor 2002; Neumann 2008). Apabila kekeringan berlanjut menyebabkan pertumbuhan fase generatif terganggu sehingga melakukan mekanisme menggugurkan daun (Neumann 2008). Hamim (2004) menyatakan bahwa pada tahap awal, kekeringan menyebabkan berkurangnya pembukaan stomata untuk meminimalisir kehilangan air di bawah kondisi cahaya berlebihan. Peristiwa ini mengakibatkan terjadinya penurunan konsentrasi CO2 intrasel, sehingga tanaman mengalami overreduksi pada transfer elektron fotosintesis. Overreduksi terjadi karena pembentukan NADPH reaksi terang tidak diimbangi oleh pemakaian NADPH pada reaksi gelap karena penurunan konsentrasi CO2intrasel. Hal ini mengakibatkan terbentuknya
reactive oxygen species (ROS) diawali dengan pengikatan elektron pada transpor elektron fotosintesis oleh oksigen. Proses selanjutnya terbentuk berbagai bentuk senyawa ROS seperti; superoksida (O2-), singlet oksigen (.O2), radikal hidroksil (OH) dan hidrogen peroksida (H2O2) (Mckersie dan Leshem 1994). Senyawa
reactive oxygen species (ROS) dapat menimbulkan kerusakan pada tanaman (Aroca et al. 2001). Jika hal ini dibiarkan, maka daun akan gugur (Apel dan Hirt 2004).
Suhu dan kelembaban udara memiliki keterkaitan dengan produktivitas serasah. Naiknya suhu udara menyebabkan menurunnya kelembaban udara sehingga transpirasi meningkat, dan untuk menguranginya daun harus segera digugurkan (Yuliadi dan Suci 2008). Hal ini sesuai dengan hasil penelitian terlihat minggu ke-1 dan minggu ke-2 memiliki suhu tertinggi sebesar 31,40C dan 32,60C, sebaliknya minggu ke-1 dan minggu ke-2 untuk pengukuran kelembaban udara memiliki nilai paling kecil yaitu sebesar 69,5% dan 69,1%.
14
Dekomposisi Serasah
Dekomposisi serasah merupakan proses yang sangat penting dalam dinamika hara pada suatu ekosistem (Regina dan Tarazona 2001). Kecepatan kematangan dalam proses dekomposisi bervariasi (Kochy dan Wilson 1997). Secara umum kematangan dalam proses dekomposisi mengacu pada sedikit banyaknya bakteri, jamur, air, dan kelembaban serasah (Lekasi et al. 2003). Beberapa karakteristik yang digunakan untuk menilai kematangan dan kualitas kompos meliputi tekstur, warna, bau, dan aktivitas biologis (Lekasi et al. 2003). Hasil penelitian menunjukkan bahwa perlakuan P3K2 lebih efisien karena pada minggu ke-7 sudah berwarna hitam dengan kebutuhan konsentrasi EM4 rendah dibandingkan dengan perlakuan P3K3. Kompos matang dengan karakteristik tekstur serasah kasar menjadi lebih halus, kompos matang diperkirakan tinggal 30% dari biomassa awal, selanjutnya warna kompos cokelat seragam gelap atau hitam, dan tidak berbau busuk (Kochy dan Wilson 1997).
Hasil penelitian menunjukkan F hitung 24,77 > F tabel 3,555 atau P value 0,000 < α 0,05 sehingga terdapat pengaruh perbedaan potongan serasah daun mahoni terhadap proses dekomposisi (Lampiran 1). Perlakuan P3K2 dengan luas daun 0,7-1 cm2 lebih cepat matang dan efisien pada perlakuan lain (Tabel 2). Ukuran partikel serasah kecil mempengaruhi pergerakan oksigen ke dalam tumpukan yaitu berkaitan dengan kerja mikroba dalam mendekomposisi serasah yang melibatkan fungsi enzim dan substrat. Partikel kecil dari bahan organik meningkatkan permukaan area, sehingga mikroba dalam dekomposisi serasah bekerja secara maksimal. Ukuran serasah kecil membuat pergerakan karbon dioksida dalam tumpukan serasah sehingga proses dekomposisi lebih cepat atau matang (Zia et al. 2003). Ukuran potongan daun yang lebih kecil menjadikan mikroba bekerja secara maksimal dalam mendekomposisi serasah, sedangkan ukuran potongan yang lebih besar memiliki luas permukaan area yang lebar dan terdapat rongga udara yang menyebabkan mikroba bekerja kurang maksimal (Marie et al. 2002). Proses semi aerobik terdapat lubang (ventilasi udara) pada penutup ember percobaan. Lubang berfungsi meminimalkan kerugian menguapkan selama resirkulasi (Pohland 1980). Dekomposisi secara semi aerobik menyebabkan kerja bakteri lebih cepat mendekomposisi seperti bakteri aerobik (Hanashima et al. 1981). Selain itu dekomposisi secara semi aerobik memberikan keuntungan yaitu mempercepat biodegradasi senyawa organik dan stabilisasi (Robison dan Peter 1983). Sistem semi aerobik menguntungkan dengan nitrifikasi dan denitrifikasi terjadi secara bersamaan dan mempercepat proses stabilisasi (Theng et al. 2005).
Hasil penelitian menunjukkan F hitung 17,95 > F tabel 3,555 atau P value
0,000 < α 0,05 sehingga terdapat pengaruh perbedaan konsentrasi Effective Microorganism 4 (EM4) terhadap proses dekomposisi (Lampiran 1). Konsentrasi paling efisien terdapat pada perlakuan P3K2 yaitu larutan gula (100 gr gula dilarutkan 1 liter air) dengan EM4 10 ml (Tabel 1). Effective Microorganism 4 (EM4) berfungsi mempercepat penguraian bahan organik, menghilangkan bau yang timbul selama proses penguraian dan meningkatkan aktivitas mikro organisme (Syafrudin dan Badrus 2007). Hasil penelitian lainnya menunjukkan F
hitung 3,77 > F tabel 2,928 atau P value 0,021 < α 0,05 sehingga terdapat
15
dekomposisi serasah (Lampiran 1). Dekomposisi serasah adalah proses penghancuran bahan organik seperti daun mahoni menjadi menjadi unsur hara terlarut dan partikel yang lebih kecil yaitu berkurangnya bobot serasah. Kehilangan bobot yang cepat dalam tahap awal dekomposisi serasah terkait dengan karbohidrat mudah terurai, Sedang pada tahap selanjutnya relatif lambat karena akumulasi senyawa yang lebih rekalsitran seperti lignin dan selulosa (Songwe et al. 1988; Berg 2000).
Rasio karbon dengan hara lain sangat penting dalam proses dekomposisi. Proses dekomposisi semi aerobik melibatkan mikroorganisme yang bekerja secara sinergik. Mikroorganisme membutuhkan sumber karbon untuk menyediakan energi dan pasokan nitrogen untuk protein sel. Berdasarkan hasil penelitian perlakuan dengan potongan 3 dengan luas 0,7-1 cm2 memiliki nilai rasio C/N rasio lebih kecil dibanding dengan potongan lain dengan rentang 16,38-18,40. Penurunan rasio C/N disebabkan karena kenaikan kadar N dan penurunan kadar C. Peningkatan kadar N akibat terjadinya penguraian protein menjadi asam amino selama pengomposan dengan bantuan kegiatan mikroorganisme heterotropik, seperti bakteri, fungi dan Actinomycetes. Asam amino kemudian mengalami amonifikasi menghasilkan ammonium yang selanjutnya dioksidasi menjadi nitrat (Oades 1994). Penurunan unsur karbon (C) disebabkan karena senyawa karbon organik digunakan sebagai sumber energi bagi organisme dan selanjutnya karbon tersebut hilang sebagai CO2. EM4 merupakan kultur campuran mikroorganisme aerob dan anerob seperti Lactobacillus, bakteri fotosintetik, Actinomycetes dan sedikit jamur. Mikroorganisme tersebut bekerja secara sinergik untuk menguraikan bahan organik secara terus menerus, dan suksesif terbukti adanya koloni bakteri tertangkap pada minggu ke-10 (Lampiran 3).
16
Faktor penting dalam proses dekomposisi selain suhu yaitu pH. Dalam proses dekomposisi pH berkisar 4-12 (Rizwan et al. 2007). Tidak ada pH spesifik yang diperlukan untuk proses dekomposisi (Gaur 1997). Proses dekomposisi serasah melibatkan kegiatan metabolik yang berpengaruh terhadap pH kompos. Meningkatnya pH disebabkan oleh deaminasi yaitu reaksi kimiawi metabolisme melepaskan gugus amina dari molekul senyawa asam amino. Gugus amina terlepas akan terkonversi menjadi amonia sebaliknya produksi asam organik selama dekomposisi karbohidrat dan lipid mengurangi pH. Peningkatan pH erat kaitannya dengan proses dekomposisi. Bahan organik yang telah terdekompisisi dapat meningkatkan aktivitas ion OH- yang bersumber dari gugus karboksil (-COOH) dan gugus hidroksil (OH-). Ion OH- akan menetralisir ion H+ yang berada dalam larutan. Brady dan Weil (2002), menyatakan bahwa naik turunnya pH merupakan fungsi ion H+ dan OH-, jika konsentrasi ion H+ dalam larutan naik, maka pH akan turun dan jika konsentrasi ion OH- naik maka pH akan naik, lebih lanjut dijelaskan pula bahwa bahan organik yang telah terdekomposisi akan dapat menghasilkan ion OH- yang dapat menetralisir aktivitas ion H+. Peningkatan pH tersebut erat kaitannya dengan hasil dekomposisi bahan organik. Hasil penelitian menunjukkan pH dalam kondisi asam hingga minggu 5 yaitu sebesar 4 sampai 4,3 kemudian pH netral hingga proses dekomposisi selesai, hal ini sesuai dengan Hernando et al. (1989) melaporkan bahwa produk kompos pengukuran pH pada tahap awal asam dan kompos matang dengan pH netral.
Proses dekomposisi merupakan proses dinamis yang melibatkan banyak mikroba seperti bakteri fotosintetik, Lactobacillus, Actinomycetes, dan yeast (Crawford 1983). Hasil penelitian menunjukkan bahwa aktivator EM4 diisolasi dengan menggunakan medium selektif, yeast menggunakan medium PDA dimana yeast paling banyak terdapat pada perlakuan P3K3 dengan total koloni 11,9 x 106, sedangkan Lactobacillus menggunakan medium MRSA bakteri paling banyak pada perlakuan P3K3 dengan total koloni 2,84 x 106, selanjutnya bakteri
Actinomycetes menggunakan medium SCA bakteri paling banyak terdapat pada perlakuan P3K3 dengan total koloni 9,94 x 106, dan medium sistrom tidak terdapat bakteri fotosintetik. Bakteri fotosintetik membentuk zat-zat bermanfaat yang menghasilkan asam amino, asam nukleat dan zat-zat bioaktif yang berasal dari gas berbahaya dan berfungsi mengikat nitrogen dari udara, bakteri tidak terlihat saat kompos mendekati matang atau matang (Atkinson et al. 1996). Bakteri Lactobacillus pada proses dekomposisi berfungsi untuk fermentasi bahan organik jadi asam laktat, percepat perombakan bahan organik, lignin dan cellulose, dan menekan pathogen dengan asam laktat yang dihasilkan.
Actinomycetes menghasilkan zat anti mikroba dari asam amino yang dihasilkan bakteri fotosintetik. Ragi menghasilkan zat anti biotik, menghasilkan enzim dan hormon, sekresi ragi menjadi substrat untuk Actinomycetes. Jamur fermentasi mampu mengurai bahan organik secara cepat yang menghasilkan alkohol ester anti mikroba, menghilangkan bau busuk (Higa dan Wididana 1994).
17
bakteri mesofilik terhenti pada temperatur >40oC. Asam organik yang dihasilkan pada tahap ini menurunkan pH. Termofilik, yaitu suhu yang meningkatkan aktivitas mikroba menyebabkan kenaikan 40-60oC. Meningkatnya aktivitas mikroba termofilik (>40oC) menghasilkan amoniak sehingga terjadi kenaikan pH (Strom 1985). Temperatur >60oC aktivitas jamur termofilik terhenti, selanjutnya digantikan oleh kelompok bakteri dan Actinomycetes hingga temperatur 70-80oC. Kenaikan suhu tersebut secara langsung mematikan berbagi jenis pathogen. Apabila suhu melampaui 65-70oC, aktivitas mikroba dan proses dekomposisi senyawa organik terhambat karena suhu tinggi tersebut mematikan mikroba. Tahap selanjutnya pendinginan yaitu berkurangnya substrat dan tingginya temperatur menimbulkan kematian pada mikroba dan aktivitas metabolisme menurun sehingga temperatur turun kembali ke fase awal mesofilik (Tuomela et al. 2000).
4 SIMPULAN DAN SARAN
Simpulan
Produktivitas tertinggi terjadi minggu ke-16 sebesar 89,24 g/m2/minggu yaitu bulan mei-juni bertepatan dengan musim kemarau. Total produktivitas serasah terbesar terdapat pada pohon 2 yaitu diameter 33 cm, tinggi 20 m, dan tebal tajuk 5,5 m dengan total produktivitas 1415 g/m2/minggu. Hasil penelitian menunjukkan bahwa perlakuan P3K2 lebih efisien karena pada minggu ke-7 sudah berwarna hitam dengan konsentrasi EM4 lebih rendah dibanding perlakuan P3K3. Terdapat pengaruh ukuran potongan daun mahoni pada dekomposisi semi aerobik. Hasil penelitian menunjukkan paling cepat matang adalah potongan 3 dengan luas 0,7-1 cm2. Terdapat pengaruh konsentrasi EM4 pada dekomposisi semi aerobik, proses dekomposisi paling efisien adalah konsentrasi K2 yaitu larutan gula (100 gr gula dilarutkan 1 liter air) dengan EM4 10 ml.
Saran
18
DAFTAR PUSTAKA
Aerts R, Caluwe HD. 1997. Nutritional and plant-mediated controls on leaf litter decomposition of Carex species. Ecol. 78: 244-260.
Albrektson A. 1988. Needle litterfall in stands of Pinus sylvestris L. in Sweden, in relation to site quality, stand age and latitude. Scand. J For Res. 3:333-342. Anni R, deni R, Karunia HP. 2003. Pengaruh agitasi terhadap proses
pengomposan sampah organik. Infomatek. 5:4-7.
Apel K, Hirt H. 2004. Reactive oxygen species: metabolism, oxydative stress, and signal transduction. Plant Biol. 55:373-399.
Arato HD, Martins SV, Ferrari SHDS. 2003. Produc’aoedecomposic¸ aodeserapilheira emumsistemaagroflorestal implantado para recuperac¸ao de´area degradada em Vic¸osa-MG. Revista A´ rvore. 27:715–721.
Ara´ujo AC, Nobre AD, Kruijt B, Elbers JA, Dallarosa R, Stefani P, Randow CV, Manzi AO, Culf AD, Gash JHC, Valentini R, Kabat P. 2002. Comparative measurements of carbon dioxide fluxes from two nearby towers in a central Amazonian rainforest: The Manaus LBA site. JGeo Res. 107:1–20.
Armbrust DV, Paulson GM, Ellis R. 1974. Physiological responses to wind and sandblast damaged winter wheat plants. Agron J. 66:421-423.
Aroca R, Juan JI, Manuel SD. 2001. Photosynthetic characteristics and protective mechanisms against oxidative stress during chilling and subsequent recovery in two maize varieties differing in chilling sensitivity. Plant Sci
161:719–726.
Aston JL, Bradshaw AD. 1966. Evolution in closely adjacent plant populations. II.
Agrostis tenuis in maritime habitats. Heredity. 21:649-664.
Atkinson CF, Jones DD, Gauthier JJ. 1996. Biodegradabilities and microbial activities during composting of oxidation ditch sludge. Compost Sci Utiliz.
4:84-96.
Barbosa R, Fearnside P. 1996. Carbon and nutrient flows in an Amazonian forest: fine litter production and composition at Apia´u, Roraima, Brazil. Trop Ecol. 37:115–125.
Berg B. 2000. Litter decomposition and organic matter turnover in northern forest soils. For Ecol Manage. 133:13–22.
Brady NC, Weil RR. 2002. The Nature and Properties of Soils. 31th ed. Prentice-Hall, Upper Saddle River, New York. 511 p.
Brodribb TJ, Holbrok NM. 2003. Stomatal closure during leaf dehydration, correlation with other leaf physiological traits. Plant Physiol. 132:2166-2173.
Boodle LA. 1920. The scorching of foliage by sea-winds. J Minist Agric. 27:479-486.
Caldwell MM. 1970. Plant gas exchange at high wind speeds. Plant Physiol.
46:535-357.
Clark DA, Brown S, Kichlighter DK, Charmbers JQ, Thomlinson JR, Ni J, Holland EA. 2001. Net primary production in tropical forests, an evaluation and synthesis of existing field data. Ecol. 11:371-384.
Chaves M. 1991. Effect water deficit on carbon assimilation. J Exp Bot. 42:1-6. Crawford JH. 1983. Composting of agricultural wastes: a review. Proc Biochem.
19 Venezuelan Guayana. Biogeochem. 43: 197-210.
Dimock EJ. 1958. Litterfall in a young stand of Douglas fir. Northw Sci. 32:19-29. Epstein E, Willson GB, Burg WD, Mullen DC, Enkiri NK. 1976. A forced aeration system for composting waste water sludge. J Water Pollut. 48: 688-691.
Facelli JM, Pickett STA. 1991. Plant litter its dynamics and effects on plant community structure. J Bot. 57:1-32.
Fryrear DW. 1971. Survival and growth of cotton plants damaged by wind blown sand. Agron J. 63:638-642.
Finer L. 1996. Variation in the amount and quality of litterfall in a Pinus sylvestris
L. stand growing on a bog. Forest Ecol Manag. 80:1-11
Gaur AC. 1997. Bulky Organic Manures and Crop Residues. in: tandon hls, ed., Fertilizers, Organic Manures, Recyclable Wastes and Biofertilizers, FDCO, New Delhi, India, pp. 37-51.
Grace J. 1974. The effect of wind on grasses cuticular and stomatal transpiration.
J Exp Bot. 25:542-551.
Gosz JR, Likens GE, Bormann FH. 1972. Nutrient content of litter fall on the hubbard brook experimental forest, New Hampshire. Ecol. 53: 769-784 Guo LB, Sim REH. 1999. Litter decomposition and nutrient release via litter
decomposition in New Zealand eucalypt short rotation forests. Agricul Eco and Environ. 75: 133-140.
Hanashima M, Yamasaki K, Matsufuji Y. 1981 Experimental study of the landfill structure for solid waste Disposal. J Hydraulic. 310:69-76.
Hamim. 2004. Underlying drought stress effect on plant: inhibition of photosynthesis. J Hayati Biosci. 11:164-169.
Hart SC, Firestone MK, Paul EA. 1992. Decomposition and nutrient dynamics of ponderosa pine needles in a Mediterranean-type climate. Can J For Res.
22:306-314.
Hennessey TC, Dougherty PM, Cregg BM, Wittwer RF. 1992. Annual variation in needle fall of a loblolly pine stand in relation to climate and stand density.
For Ecol Manag. 51:329-338.
Hernando S, Lobo MC, Polo A. 1989. Effect of the application of municipal refuse compost on the physical and chemical properties of a soil. Sci Tot Environ. 81:589-596.
Higa T, Wididana GN. 1994. The Concept and Theories of Effective Microorganisms. Second International Conference at Khon Kaen University. Thailand:hal 118-124.
Hobbie SE. 1992. Effect of plant species on nutrient cycling. Trends in Ecol and Evo. 7:336-339.
20
Hopkins B. 1966. Vegetation of the Olokemeji Forest Reserve, Nigeria IV. The litter and soil with special reference to their seasonal changes. J Ecol. 54: 687-703.
John DM. 1973. Accumulation and decay of litter and net production of forest in tropical west Africa. Oikos. 24:430-435.
Johnson LC, Damman AWH. 1991. Species-controlled sphagnum decay on a south Swedish raised bog. Oikos. 61:234-242.
Karlsson K, Malmberg P. 1989. Characterization of exposure to molds and actinomycetes in agricultural dusts by scanning electron microscopy, fluorescence microscopy and the culture method. Scand J Work Environ Health. 15:353-359
Karma MA. 1970. Litter production. In: Philipson J (ed) method of study in soil ecology. Unesco IBP. 6:139-143.
Kitayama K, Suzuki S, Hori M, Takyu M, Aiba S, Majalap-lee N, Kikuzawa K. 2004. On the relationships between leaf-litter lignin and net primary productivity in tropical rain forests. Oecol. 2:335-339.
Kochy K, Wilson SD. 1997. Litter decomposition and nitrogen dynamic in aspen forest and mixed-grass prairie. Ecol. 78:732-739.
Lawlor DW. 2002. Limitation to photosynthesis in water-stressed leaves: stomata vs metabolism and the role of ATP. Ann Bot. 89:871-885.
Lekasi JK, Tanner JC, Kimani SK, Harris PJC. 2003. Cattle manure quality in Maragua District, Central Kenya: Effect of management practices and development of simple methods of assessment. Agr Ecosyst Environ.
94:289-298.
Liu C, Westman CJ, Berg B, Kutsch W, Wang GZ, Man R, Ilvesniemi H. 2004. Variation in litterfall-climate relationships between coniferous and broadleaf forests in Eurasia. Global Ecol and Biogeogr. 13:105-114.
Luiza˜OF. 1989. Litter production and mineral element input to the forest in a
Central Amazonian forest. Geojournal. 19:407–417.
Marie CD, Louise B, Michele H. A study of the impact of particle size and adsorption phenomena in a compost-based biological filter. Chem Engin Scie. 57:4999 – 5010.
Martin EV, Clements FE. 1935. Studies of the effect of artificial wind on growth and transpiration in Helianthus annuus. Plant Physiol. 10: 613-636.
McKersie BD, Leshem YY. 1994. Stress and Stress Coping in Cultivated Plants. Netherlands: Kluwer Academic Publishers.
Mellilo JM, Aber JD, Muratore JF. 1982. Nitrogen and lignin control of hardwood leaf litter decomposition dynamics. Ecol. 63:621-626.
Michael S, Anna S, Tatu H, Paivi M, Helja SM. 2005. Models of litterfall production for Scots pine (Pinus sylvestris L.) in Finland using stand, site and climate factors. For Ecol Manag. 205:215–225.
Muoghalu JI, Akanni SO, Eretan OO. 1993. Litter fall and nutrient dynamics in a Nigeria rainforest seven years after a ground fire. J VegScie. 4: 323-328. Neumann PM. 2008. Coping mechanisms for crop plants in drought-prone
environments. Ann Bot. 101:901-907.
21
Pohland FG. 1980. Leachate recycle as landfill management option. J Environ Engin. 106:1057–1069.
Polyakova O, Billor N. 2007. Impact of deciduous tree species on litterfall quality, decomposition rates and nutrient circulation in pine stands. Forest Ecol. 253:11–18.
Regina IS, Tarazona T. 2001. Nutrient pools to the soil through organic matter and throughfall under a Scot pine plantation in the Sierra de la Demanda, Spain. European Journal of Soil Biology. 37: 125-133.
Rizwan A, Ghulam J, Muhammad A, Zahir AZ, Azeem K. 2007. Bio-conversion of organic wastes for their recycling in agriculture: an overview of perspectives and prospects. Annals of Microb. 57:471-479.
Robioson HD, Peter JM. 1983. The treatment of leachates from domestic wastes in landfills-I: Aerobic biological treatment of a medium-strength leachate.
Water Research. 17:1537–1548.
Rodan B, Newton A, Veríssimo A. 1992. Mahogany conservation: status and policy initiatives. Environ Conser. 19:331-342.
Saetre P. 1998. Decomposition, microbial community strusture, and earthworm effects along a birch-spure soil gradient. Ecol. 79:834-846.
Sallata M, Kudeng, Halidah. 1990. Produksi dan penghancuran serasah di bawah hutan alam sekunder di Tabo-tabo Sulawesi Selatan. J Penelitian Kehutanan. 4:19-25.
Santiago LS, Mulkey SS. 2005. Leaf productivity along a precipitation gradient in lowland Panama: patterns from leaf to ecosystem. Structure and Function.
19:349-356.
Saptoadi. 2001. Utilization of organic matter from municipal solid waste in compost industries. Jurnal Manusia Lingkungan. 8:119-129.
Schwanz P, Polle A. 2001. Differential stress responses of antioxidative systems to drought in penduculate oak (Quercus robur) and maritime pine (Pine pinaster) grown under high CO2 concentration. J Exp Bot. 52:133-143. Songwe NC, Fasehun FE, Okali DUU. 1988. Litter fall and productivity in a
tropical rainforest, Southern Bakundu Forest, Cameroon. JTrop Ecol. 4: 25-37.
Strom PF. 1985. Effect of temperature on bacterial species diversity in thermophilic solid-waste composting. Environ Microbiol. 50:899-905. Syafrudin, Badrus Z. 2007. Pengomposan limbah teh hitam dengan penambahan
kotoran kambing pada variasi yang berbeda dengan menggunakan starter EM4 (efective microorganism-4). Teknik.28:852-859
Tadaki Y, Kagawa T. 1968. Studies on the production structure of forest: 13. Seasonal change of litterfall in some evergreen stands. J Japan For Soc.
50:7-13
Taylor SE, Sexton OJ. 1972. Some implications of leaf tearing in Musaceae. Ecol.
53:143-149.
Thaiutsa B, Granger O. 1979. Climate and decomposition rate of tropical forest litter. Unasylva. 31:28-35.
22
Thompson JR. 1974. The effect of wind on grasses. II. Mechanical damage in
Festuca arundinacea Schreb. J Exp Bot. 25:965-972.
Tuomela M, Vikman M, Hatakka A, Itavaara M. 2000. Biodegradation of lignin in a compost environment: a review. Biores Tech. 72:169-183.
Tranquillini W. 1969. Photosynthese und transpiration einiger holzarten bei verschieden starkern wind. Cbl. Ges. Forstwesen. 86: 35-48.
Tsuboi Y. 1961. Ecological studies on rice plants with regard to damages caused by wind. Bull. Nat. Inst. Agric Sci. 8:1-156.
Upadhyay VP, Singh J, Meentemeyer V. 1989. Dynamics and weight loss of leaf litter in central Himalayan forests: abiotic versus leaf litter quality influences. Ecol. 77:147–161.
Van Breemen N. 1995. Nutrient cycling strategies. Plant and Soil. 169:321-326. Veríssimo A, Barreto P, Tarifa R, Uhl C. 1995. Extraction of a high-value natural
resource in Amazonia: the case of mahogany. For Ecol Manag. 72:39-60. Vitousek PM. 1984. Litterfall, nutrient cycling, and nutrient limitation in tropical
forests. J Ecol. 65:285–298.
Vitousek PM, Turner DR, Parton WJ, Sanford RL. 1994. Litter decomposition on the Mauna Loa environmental matrix, Hawaii: Patterns, mechanisms and Models. Ecol. 75:418-429.
Villela DM, Proctor J. 2002. Leaf litter decomposition and monodominance in the Peltogyne Forest of Maraca Island, Brazil. Biotropica. 34:334–347.
Vose JM, Allen HL. 1991. Quantity and timing of needlefall in N and P fertilized loblolly pine stands. For Ecol Manag. 41:205-219
Wilson J. 1980. Macroscopic features of wind damage to leaves of Acer pseudoplatanus L and its relationship with season, leaf age and windspeed.
Ann Bot. 46: 303-311.
Yongzhen T, Yanling H, Yongqiang W, Fanghua L, Xinfeng L, Wenming Z, Zhihua Z. 2008. Characteristics of a new photosynthetic bacterial strain for hydrogen production and its application in wastewater treatment. Int J Hydrogen Energy. 33:963-973
Yu H, Zhi LX, Chun SR, Jia LZ, De ZW, Yu NW, Teng CM. 2005. Inactivation of yeast by dielectric barrier discharge (DBD) plasma in helium at atmospheric pressure. IEEE. 33:1405-1410
Yuliadi Z, Suci R. 2008. Produksi serasah hutan mangrove di perairan pantai teluk sepi, Lombok Barat. Biodiversitas. 9:284-287.
23
Lampiran 1 Analisis ragam laju dekomposisi serasah mahoni (Swietenia macrophylla King)
Analisis varian dekomposisi serasah mahoni (Swietenia macrophylla King) Sumber
keragaman
db JK KT F hitung F tabel P value
potongan Konsentrasi Interaksi Galat
2 2 4 18
1,09 0,79 0,33 0,4
0,545 0,395 0,083 0,022
24,77 17,95 3,77
F 0,05;2;18 = 3,555 F 0,05;2;18 = 3,555 F 0,05;4;18 = 2,928
0,000 0,000 0,021
Total 26 2,61
24
Lampiran 2 Hasil dekomposisi semi aerobik daun mahoni
25
Perlaku an
Dokementasi
Minggu 1 Minggu 6 Minggu 10
P3K1
P3K2
P3K3
Ket: 1. Warna asli (tingkat kematangan 0-25 %) 2. Cokelat (tingkat kematangan 26%-50%) 3. Cokelat Pekat (tingkat kematangan 51%-75%) 4. Hitam (Tingkat kematangan 76%-100%)
P3K1 (1)
P3K1 (3)
P3K2 (1)
P3K2 (4) P3K2
(3)
P3K1 (4)
P3K3 (1)
P3K3 (3)
26
Lampiran 3 Koloni bakteri minggu ke-10 dengan medium selektif
Dokumentasi Keterangan
Medium Potato Dextrose Agar (PDA)
Medium ini berfungsi untuk menangkap yeast pada EM4
Pada pengenceran 10-5
Fungsi menghasilkan zat anti biotik, menghasilkan enzim, dan hormon, sekresi ragi menjadi substrat untuk actinomycetes. Jamur fermentasi menguraikan bahan organik secara cepat menghasilkan alkohol ester anti mikroba, menghilangkan bau busuk (Higa dan Wididana 1994)
Medium deMann Rogosa Sharpe Agar
(MRSA)
Medium ini berfungsi untuk menangkap
Lactobacillus pada EM4
Pada pengenceran 10-5
Fungsi untuk fermentasi bahan organik jadi asam laktat, percepat perombakan bahan organik, lignin dan cellulose (Higa dan Wididana 1994)
Medium Soluble Casein Agar (SCA)
Medium ini berfungsi untuk menangkap
Actinomycetes pada EM4
Pada pengenceran 10-3
Fungsi menghasilkan zat anti mikroba dari asam amino yang dihasilkan bakteri fotosintetik (Higa dan Wididana 1994
Medium Sistrom
Medium ini berfungsi untuk menangkap Bakteri fotosintetik pada EM4
Pada pengenceran 10-3
27
Lampiran 4 Penelitian lain mengenai scanning electron microscopy (SEM) bakteri yang terlibat dalam EM4
Dokumentasi Keterangan
(Yu et al. 2005)
Yeast
Medium PDA
Warna putih bening
Menghasilkan senyawa-senyawa bermanfaat bagi pertumbuhan tanaman dari asam amino dan gula yang dike luarkan oleh bakteri fotosintetik
(Hood and Zottola 1987)
Lactobacillus
Medium MRSA
Warna merah bening
Termasuk bakteri gram positif
menghancurkan bahan organik seperti lignin dan selulosa.
(Karlsson dan Malmber 1989)
Actinomycetes
Medium SCA
Warna putih keruh
Termasuk bakteri gram positif
Actinomycetes menghasilkan zat-zat anti mikroba dari asam amino yang dihasilkan bakteri fotosintetik.
(Yongzhen et al. 2008)
Bakteri fotosintetik
Medium Sistrom
Warna putih bening
Termasuk bakteri gram negatif
28
Lampiran 5 Bahan media selektif sebagai penangkap bakteri EM4
Jenis bahan
Jenis media
Jumlah PDA
(100 ml)
MRSA (100 ml)
SCA (100 ml)
Sistrom (100 ml)
1. PDA (Kemasan jadi) 5,3 g 5,3 g
2. MRSA (Kemasan jadi) 3,6 g 3,6 g
3. Soluble starch 1 g 1 g
4. Casein 0,03 g 0,03 g
5. KNO3 0,2 g 0,2 g
6. K2HPO4 0,2 g 3,48 g 3,68 g
7. MgSO4.7H2O 0,005 g 0,3 g 0,305 g
8. CaCO3 0,002 g 0,002 g
9. FeSO4,7H2O. 0,001 g 0,002 g 0,003 g
10. NH4CI 0,5 g 0,5 g
11. NaCI 0,5 g 0,5 g
12. CaCI2.2H20 0,04 g 0,04 g
13. EDTA 0,17 mg 0,17mg
14. MnSO4.2H20 0,15 mg 0,15mg
15. ZnSO4.7H20 0,5 mg 0,5 mg
16. CuSO4.5H20 0,03 mg 0,03mg
17. Co(N03)2.6H20 0,02 mg 0,02mg
18. H3B03 0,02 mg 0,02mg
Keterangan:
1. Medium Potato dextrose Agar (PDA) untuk mengisolasi yeast pada perlakuan.
2. Medium deMann Rogosa Sharpe Agar (MRSA) mengisolasi bakteri Lactobacillus sp.
3. Medium Soluble Casein Agar (SCA) untuk mengisolasi bakteri Actinomycetes sp.
29
RIWAYAT HIDUP
Penulis dilahirkan di Jember, Jawa Timur pada tanggal 24 Juli 1988 sebagai putra dari pasangan Bapak Miftahul Ulum dan Ibu Sumarlik. Penulis merupakan anak pertama dari dua bersaudara.