KARAKTER BIOLOGI JARAK PAGAR (Jatropha curcas L.)
TRIMONOECIOUS
Kurniati
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI
Dengan ini saya menyatakan bahwa tesis yang berjudul Karakter Biologi Jarak Pagar (Jatropha curcas L.) Trimonoecious merupakan gagasan dan karya saya bersama komisi pembimbing dan belum diajukan dalam bentuk apapun kepada perguruan tinggi manapun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain disebutkan dalam teks dan dicantumkan dalam daftar pustaka di bagian akhir tesis ini.
Bogor, April 2014
Kurniati
RINGKASAN
KURNIATI. Karakter Biologi Jarak Pagar (Jatropha curcas L.) Trimonoecious. Dibimbing oleh TRIADIATI dan UTUT WIDYASTUTI.
Jarak pagar (Jatropha curcas L.) merupakan tanaman tahunan berumah satu atau biasa disebut tanaman tipe monoecious. Saat ini diketahui terdapat tanaman jarak pagar yang mempunyai bunga hermaprodit asal tanaman monoecious namun jumlahnya terbatas. Tanaman jarak pagar berbunga hermaprodit termasuk tipe tanaman andromonoecious (hanya mempunyai bunga hermaprodit dan jantan) dan tipe tanaman trimonoecious (memiliki bunga jantan, betina dan hermaprodit). Tanaman jarak pagar trimonoecious menghasilkan bunga hermaprodit dan betina yang keduanya memiliki peluang untuk menghasilkan buah, sehingga peluang untuk menghasilkan jumlah buah lebih banyak dibandingkan tanaman jarak pagar monoecious yang hanya menghasilkan bunga betina. Permasalahan utama pada tanaman jarak pagar trimonoecious yaitu adanya kesulitan untuk membedakan antara tanaman monoecious dan trimonoecious pada fase bibit dan ini merupakan faktor pembatas dalam pengembangan jarak pagar trimonoecious. Tanaman trimonoecious ini baru dapat dikenali saat tanaman berumur 6 bulan setelah penanaman yaitu pada saat muncul bunga pertama. Penelitian ini bertujuan untuk melakukan identifikasi karakter biologi pada tanaman jarak pagar trimonoecious yang dapat digunakan sebagai penciri sifat trimonoecious. Karakter yang dievaluasi berupa karakter agronomi, analisis ukuran kromosom, analisis isozim, dan analisis viabilitas polen.
Penelitian dilaksanakan bulan Desember 2011 – Februari 2013. Percobaan dilakukan dengan menggunakan Rancangan Acak Kelompok (RAK) satu faktor yaitu genotipe jarak pagar. Analisis viabilitas polen dan karakter agronomi menggunakan analisis sidik ragam dan dilanjutkan dengan uji lanjut Duncan Multiple Range Test (DMRT) taraf 5%. Pengelompokan nilai heritabilitas arti luas menurut Stansfield (1983). Analisis pengelompokan jarak genetik dilakukan dengan metode pengelompokan program minitab 15.
Dendogram hasil analisis kekerabatan dapat membedakan antara tanaman trimonoecious dan monoecious pada koefisien kesamaan 27.25.
SUMMARY
KURNIATI. Biological Characteristics of Jatrophacurcas Trimonoecious. Under supervision of TRIADIATI and UTUT WIDYASTUTI.
Physic nut (Jatropha curcas L.) is an original plant from tropical America belonging to the family of Euphorbiaceae and an annual plant of monoecious.
Commonly J.curcas have male and female flowers on the same plant. There are currently some J. curcas which have hermaphrodite flowers from monoecious plants, but very rarely. J. curcas with hermaphrodite flowers are classified as andromonoecious type (having hermaphrodite and male in the same plant) and trimonoecious type (having male, female, and hermaphrodite flowers in the plant). Trimonoecious plants with hermaphrodite flowers were tend to perform self-pollination (geitonogamy) and produce a homogenous and high-productivity in next generation. J. curcas plants is difficult in differentiating between monoecious and trimonoecious plants when it seedling, and this is a limiting factor to know development of trimonoecious J. curcas. Trimonoecious J.curcas
can be identified after six months of planting, and when it have started flowering. Therefore, the trimonoecious J. curcas biological characteristics needs to be an observation. The trimonoecious trait in J. curcas plants can be identified through several analyses, i.e. the analysis of agronomical characteristics, the chromosomes analysis, the isozyme analysis, and the analysis of pollen viability.
The study was conducted in December 2011 - February 2013. The experiments were conducted using the Randomized Complete Block Design for one factor, the J. curcas genotype with 5 replicates. Each unit of the experiment consisted of 5 plants. Aagronomics characteristic analyses were conducted using analysis of variance (ANOVA) followed by the Duncan Multiple Range Test
(DMRT) at 5% level. The classification of heritability and Genetic Variation Coefficient (GVC) criteria and relative Phenotypic Variation Coefficient (PVC). The analysis of genetic distance clustering was done using the Minitab 1.5 program clustering method.
J. curcas stem trimonoecious characters is the primary branches angle, stem diameter, number of primary and secondary branches have a higher size than monoecious plants. Diameter and thickness fruit flesh origins from monoecious plants larger than fruit from plants trimonoecious. Characteristics of the trimonoecious plants in this study were almost the same with their mother plant. Number chromosomes number of J. curcas plant genotypes have 2n = 22 chromosomes The average length of the trimonoecious plant’s chromosome was longer than the average for monoecious plants. The length of chromosomes can differ among one family, even though the numbers of the chromosomes are similar. Isozymes peroxidase (Rf 0.83) and esterase (Rf 0.20) could be used as markers to differentiate between trimonoecious and monoecious plants. Male flower pollen viability of plant origin trimonoecious have a slower rate of decrease than male flowers monoecious plants. The dendrogram from the cluster analysis results could differentiate between trimonoecious and monoecious plants with a variation coefficient of 27.25.
©Hak Cipta milik IPB, tahun 2014
Hak Cipta dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan yang wajar IPB
KARAKTER BIOLOGI JARAK PAGAR (Jatropha curcas L.)
TRIMONOECIOUS
Kurniati
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains pada
Program Studi Biologi Tumbuhan
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Judul Tesis : Karakter Biologi Jarak Pagar (Jatropha curcas L.) Trimonoecious
Nama : Kurniati NIM : G353100061
Disetujui
Komisi Pembimbing
Dr. Triadiati, M.Si Dr. Ir. Utut Widyastuti, M.Si
Ketua Anggota
Diketahui
Ketua Program Studi Biologi Tumbuhan Dekan Sekolah Pascasarjana
Dr. Ir. Miftahudin, M.Si Dr. Ir. Dahrul Syah, M.Sc.Agr
PRAKATA
Puji syukur penulis panjatkan kehadirat Allah SWT atas segala karuniaNya sehingga penelitian yang berjudul “Karakter Biologi Jarak Pagar (Jatropha curcas
L.) Trimonoecious” telah diselesaikan.
Terima kasih penulis ucapkan kepada Dr. Triadiati, M.Si dan Dr. Ir. Utut Widyastuti, M.Si selaku pembimbing atas saran, bimbingan serta dukungannya dalam pelaksanaan penelitian dan penyusunan karya ilmiah ini. Kepada Dr. Ir. Miftahudin, M.Si, atas saran dan bimbingannya. Kepada Dr. Ir. Darda Efendi, MSi terima kasih atas saran dan informasinya.
Akhirnya ucapan terima kasih juga disampaikan kepada Bapak, Ibu dan adik saya serta seluruh keluarga atas doa dan kasih sayangnya. Seseorang yang terkasih terimakasih atas pengorbanan, dan ketulusannya dalam memberi motivasi dan semangat.
Terima kasih juga kepada Bapak Adi dan pak Pras, juga kepada teman-teman di Program Biologi Tumbuhan yang kesemuanya tidak dapat saya sebutkan satu per satu, penulis mengucapkan banyak terima kasih atas bantuan dan kebersamaannya
Semoga hasil penelitian ini bermanfaat bagi mereka yang memerlukan.
Bogor, April 2014
RIWAYAT HIDUP
DAFTAR ISI
Karakterisasi Agronomi ... 3
Analisis Ukuran Kromosom ... 3
Isozim Esterase dan Peroksidase ... 4
Viabilitas Polen ... 5
3 BAHAN DAN METODE Waktu dan Tempat Penelitian ... 6
Bahan Penelitian ... 6
Tahap Penelitian ... 6
Penanaman dan Pemeliharaan ... 7
Analisis Karakter Agronomi ... 7
Analisis Kualitatif ... 7 Keadaan Umum Lokasi Penelitian ... 11
Analisis Karakter Agronomi ... 11
Karakter Batang ... 11
Karakter Daun... 14
Karakter Bunga ... 15
Karakter Buah ... 18
Karakter Biji ... 20
Analisis Ukuran Kromosom ... 21
Analisis Isozim ... 23
Analisis Viabilitas Polen... 24
Pendugaan Parameter Genetik ... 27
5 SIMPULAN DAN SARAN
Simpulan ... 29
Saran ... 30
DAFTAR PUSTAKA ... 30
DAFTAR TABEL
Halaman
1 Karakter batang dan daun umur 4 - 6 BST jarak pagar trimonoecious dan
monoecious ... 13
2 Karakter daun jarak pagar trimonoecious dan monoecious ... 14
3 Karakter bunga jarak pagar trimonoceious dan monoecious per tanaman 7 BST ... 15
4 Karakter buah 5-12 BST jarak pagar trimonoecious dan monoecious ... 19
5 Karakter biji 5-12 BST jarak pagar trimonoecious dan monoecious ... 21
6 Panjang kromosom jarak pagar trimonoecious dan monoecious ... 22
7 Viabilitas polen bunga jantan pada jarak pagar monoecious dan trimonoecious ... 25
8 Parameter genetik beberapa karakter kuantitatif jarak pagar trimonoecious ... 27
DAFTAR GAMBAR
Halaman
1 Bagan alur penelitian ... 6
2 Bentuk batang dan warna batang 4 BST ... 12
3 Cabang yang muncul dari batang utama pada genotipe jarak pagar 6 BST 12 4 Pertumbuhan tanaman jarak pagar umur 6 BST ... 13
5 Daun jarak pagar ... 14
6 Bunga jarak pagar ... 15
7 Fluktuasi tanaman trimonoecious asal bunga hermaprodit dan betina ... 17
8 Bentuk buah jarak pagar ... 18
9 Bentuk biji jarak pagar trimonoecious dan monoceious ... 20
10Kromosom masing-masing genotipe jarak pagar trimonoecious dan monoecious ... 22
11Pola pita isozim esterase dan peroksidase ... 23
12Bentuk, warna dan ukuran polen jarak pagar ... 25
13Karakter bunga/warna kepala sari bunga jantan jarak pagar ... 26
DAFTAR LAMPIRAN
Halaman
1 Komposisi larutan pewarna isozim EST dan PER
(Wendel & Weeden1989) ... 38 2 Data iklim bulanan Dramaga Bogor ... 39 3 Perkembangan jumlah bunga rata-rata pada tanaman trimonoecious
1
1 PENDAHULUAN
Latar belakang
Jarak pagar (Jatropha curcas L.) merupakan tanaman tahunan berumah satu dengan bunga jantan dan betina terdapat pada satu tanaman yang sama, atau biasa disebut tanaman tipe monoecious (Hartati et al. 2009). Saat ini diketahui terdapat tanaman jarak pagar yang mempunyai bunga hermaprodit asal tanaman monoecious namun jumlahnya terbatas. Tanaman jarak pagar berbunga hermaprodit termasuk tipe tanaman andromonoecious (hanya mempunyai bunga hermaprodit dan jantan) dan tipe tanaman trimonoecious (memiliki bunga jantan, betina dan hermaprodit) (Makkar et al. 2008).
Tanaman jarak pagar trimonoecious dengan rangkaian bunga hermaprodit dapat melakukan penyerbukan sendiri (geitonogami) dan menghasilkan generasi berikutnya yang lebih homogen dibandingkan dengan tanaman yang tidak memiliki bunga hermaprodit (Raju et al. 2002). Tanaman jarak pagar trimonoecious menghasilkan bunga hermaprodit dan betina yang keduanya memiliki peluang untuk menghasilkan buah, sehingga peluang untuk menghasilkan jumlah buah lebih banyak dibandingkan tanaman jarak pagar monoecious yang hanya menghasilkan bunga betina. Keberhasilan penyerbukan sangat tergantung pada polen bunga jantan. Permasalahan utama pada tanaman jarak pagar trimonoecious yaitu adanya kesulitan untuk membedakan antara tanaman monoecious dan trimonoecious pada fase bibit dan ini merupakan faktor pembatas dalam pengembangan jarak pagar trimonoecious. Tanaman trimonoecious ini baru dapat dikenali saat tanaman berumur 6 bulan setelah penanaman saat muncul bunga pertama, sehingga perlu dilakukan pengamatan lebih awal karakter biologi tanaman jarak pagar trimonoecious.
2
Tujuan penelitian
Penelitian ini bertujuan untuk melakukan identifikasi karakter biologi pada tanaman jarak pagar trimonoecious yang dapat digunakan sebagai penciri sifat trimonoecious.
2 TINJAUAN PUSTAKA
Jarak pagar (Jatropha curcas L.)
Jarak pagar (Jatropha curcas L.) termasuk dalam famili Euphorbiaceae, merupakan tanaman tahunan berumah satu yang memiliki bunga uniseksual (monoecious). Tanaman ini merupakan tanaman semak berkayu yang cukup berkembang di daerah tropis (Jun et al. 2010). Walaupun telah lama dikenal sebagai bahan pengobatan dan racun (Wei et al. 2005), saat ini jarak pagar makin mendapat perhatian sebagai tanaman penghasil minyak non-edible yang mayoritas digunakan sebagai bahan baku penghasil biodiesel karena kandungan minyak pada bijinya (Raju et al. 2002). Jarak pagar merupakan tanaman yang dapat tumbuh di berbagai kondisi lahan dan lingkungan dan pada tanah yang ketersediaan air dan unsur-unsur haranya terbatas atau lahan-lahan marginal (Parthiban et al. 2010).
Bunga tanaman jarak pagar adalah bunga majemuk berbentuk malai, berwarna kuning kehijauan, berkelamin tunggal dan berumah satu (putik dan benang sari pada bunga yang berbeda dalam satu tanaman). Bunga jantan berjumlah 4-5 kali lebih banyak dari pada bunga betina. Bunga jantan maupun bunga betina tersusun dalam rangkaian berbentuk cawan yang tumbuh di ujung batang atau ketiak daun (Santoso et al. 2011). Umur berbunga jarak pagar umumnya setelah umur 3 – 4 bulan sejak perkecambahan, sedangkan pembentukan buah dimulai 1 bulan kemudian. Pemanenan dilakukan jika buah telah masak, dicirikan kulit buah berwarna kuning dan kemudian mulai mengering (Camelia et al. 2012).
3
trimonoecious. Karakterisasi pertama yang dapat dilakukan yaitu karakterisasi agronomi yang kemudian dapat diperkuat dengan penanda karakter tanaman trimonoecious melalui analisis ukuran kromosom, analisis isozim dan analisis viabilitas polen.
Karakter agronomi
Karakter agronomi banyak dimanfaatkan pada penelitian karakteristik, karena identifikasi dapat dilakukan dengan cepat pada tingkat lapang. Variabilitas genetik organ tanaman seperti daun, bunga, batang merupakan organ tanaman yang paling sering digunakan sebagai karakter. Penanda agronomi telah terbukti dapat diandalkan sebagai alat yang dapat menggambarkan variabilitas intraspesifik yang berasal dari provenan yang berbeda pada beberapa spesies (Jubera 2008).
Karakterisasi agronomi biasanya diikuti dengan karakterisasi lainnya. Keanekaragaman genetik pada tunisian tall fescue (Festuca arundinaceae Schre.) dapat dijelaskan dengan menggunakan karakterisasi agronomi dan genetik (Ghorbel et al. 2011). Karakterisasi morfologi agronomi dikolerelasikan dengan karakterisasi berdasarkan pola pita isozim pada tanaman talas (Colocasia esculenta) (Trimanto et al. 2011). Selain itu identifikasi berbagai variasi Adenium obesum dari enam varietas dilakukan berdasarkan sifat agronomi, kariotipe, serta pola pita protein (Hastuti et al. 2009).
Tahap awal identifikasi tanaman, biasanya dilakukan secara morfologi. Sifat morfologi ini dapat berupa sifat kualitatif maupun kuantitatif, yang memiliki tipe dan aksi gen yang berbeda (Fitmawati 2008). Penanda morfologi masih banyak digunakan oleh para peneliti untuk mendapatkan karakteristik pohon dan daun. Kelemahan penanda morfologi dipengaruhi oleh tahap perkembangan tanaman dan lingkungan. Kadang-kadang sulit membedakan genotipe yang diamati, karena secara morfologi tampak sama, walaupun sebenarnya genotipe tersebut berbeda. Hal ini terjadi akibat sifat resesif tertutup oleh sifat dominan (Bakhtiar 2002). Sehingga juga diperlukan marka lainnya untuk mendukung identifikasi karakterisasi tanaman.
Analisis ukuran kromosom
Tujuan utama analisis kromosom adalah mengetahui karateristik dan morfologi dari kromosom, seperti jumlah kromosom, struktur dan aktivitas kromosom selama pembelahan sel berlangsung. Metode analisis kromosom berbeda pada masing-masing spesies (Gianfranco et al. 2008). Jumlah kromosom bervariasi dari satu spesies dengan spesies lain dan pada spesies yang sama jumlah kromosom adalah konstan. Spesies-spesies yang mempunyai hubungan kekerabatan dekat mempunyai jumlah kromosom sama, sedangkan spesies yang tidak mempunyai hubungan, memiliki jumlah kromosom berbeda (Schubert 2007).
4
2003b; Carvalho et al. 2008; Dahmer et al. 2009; Kikuchi et al. 2010; Reddy et al. 2013). Analisis kromosom juga telah banyak digunakan untuk menganalisis keanekaragaman tanaman lainnya, seperti yang dilakukan oleh Escudero et al. (2010) yang mengevaluasi variasi jumlah kromosom pada Cyperaceae. Talukdar (2010) menggunakan analisis kromosom untuk mengkarakterisasi dan membandingkan tiga varietas unggul tanaman grass pea. Mekanisme hereditas keanekaragaman Mallotus juga diamati melalui analisis kromosom (Soontornchainaksaeng et al. 2003a). Mao et al. (2010) mengevaluasi pola distribusi aksesi Elytrigia pada tingkat ploid di 25 negara. Identifikasi variasi
Adenium obesum dari enam varietas (obesum, cery, red lucas, red fanta, white bigben dan Harrypotter) diamati berdasarkan analisis kromosom (Hastuti et al.
2009).
Isozim peroksidase dan esterase
Keragaman genetik tidak cukup hanya diamati secara sitologis melalui pengamatan kromosom selama pembelahan sel. Hal ini terjadi karena seringkali keragaman yang timbul pada tanaman berbeda satu sama lainnya dan hanya disebabkan oleh sejumlah kecil gen saja (Aisyah 2006).
Untuk mengukur keragaman spesies tanaman, telah banyak dilakukan melalui teknik isozim. Isozim adalah enzim yang merupakan produk langsung dari gen yang memiliki molekul aktif dan struktur kimia yang berbeda tetapi mengkatalis reaksi kimia yang sama. Enzim merupakan protein biokatalisator untuk proses-proses fisiologis yang perannya dikendalikan secara genetik (Asante & Laing 2001).
Beberapa isozim diketahui berasosiasi atau terkait dengan karakter agronomi (Stalker & Mozingo 2001). Dibandingkan dengan ciri-ciri morfologi, isozim dapat lebih menunjukkan genotipe suatu organisme. Pita-pita isozim dapat dievaluasi dari segi genetik (yaitu frekuensi alel pada setiap lokus) dan dari segi fenotipe (yaitu dengan menganggap setiap pita tunggal sebagai suatu karakter kodominan, yang keberadaannya dinyatakan dengan skor). Fungsi utama isozim adalah mengendalikan aktivitas metabolik suatu organisme. (Rahayu 2012). Perbedaan ukuran, konfigurasi dan muatan ion di antara isozim membuatnya dapat dideteksi dan ditampilkan dengan berbagai cara pemisahan melalui elektroforesis (Zeidler 2000).
Enzim esterase adalah enzim hidrolase yang memisahkan gugus-gugus ester menjadi sebuah asam dan alkohol di dalam suatu reaksi kimia dengan air yang disebut hidrolisis (Ying et al. 2006). Selain itu esterase juga berperan dalam modifikasi, aktivasi dan inaktivasi berbagai jenis enzim (Wu & Tan 2003). Esterase dapat digunakan untuk identifikasi taksonomi dan membedakan analisis taksa pada beberapa spesies Curcuma L (Apavatjrut et al. 1999). Hubungan genetik antara varietas mentimun dapat dijelaskan dengan menggunakan enzim esterase (Juliansah et al. 2008). Selain itu esterase juga digunakan untuk mengevaluasi 36 aksesi spesies Trifolium, yaitu T. incarnatum, T. polymorphum, T. pratense. T. resupinatum, T. riograndense, T. subterraneum dan T. vesiculosum
5
Klaten (Jawa Tengah) serta Bantul (Yogyakarta) diuji dengan menggunakan enzim esterase (Rahmawati dan Mahajoeno 2010).
Enzim Peroksidase tergolong dalam kelompok oksidoreduktase (Ying et al.
2006). Analisis variabilitas pola pita pada beberapa aksesi nenas berdasarkan analisis isozim dengan menggunakan enzim peroksidase (Hadiati & Sukmadjaja 2002). Keragaman Puccinia arachidis Speg dapat diidentifikasi melalui variasi pola pita isozim dengan menggunakan enzim peroksidase (Tashildar et al. 2012). Isozim esterase dan peroksidase digunakan untuk membantu penentuan jenis kelamin pada Hem (Cannabis sativa L.) yang pada dasarnya cukup sulit dilakukan (Tru et al. 2002). Begitupula Sharma et al. (2010) menggunakan enzim peroksidase dan esterase untuk mengidentifikasi penanda jenis kelamin pada awal pertumbuhan tanaman dioecious Hippophae rhamniodes L.
Viabilitas polen
Serbuk sari (polen) adalah alat reproduksi jantan yang terdapat pada tumbuhan yang membawa material genetik jantan (Jiong et al. 2008). Polen berada dalam kepala sari (anther) tepatnya dalam kantung yang disebut ruang serbuk sari (theca) (Wang et al. 2003).
Penentuan viabilitas polen berperan besar dalam proses reproduksi tanaman, karena dapat menunjukkan kemampuan butir polen untuk menghantarkan sel sperma ke kantong embrio pada proses penyerbukan (Rahayu 2012). Polen dikatakan viabel apabila mampu menunjukkan fungsinya menghantarkan sperma kepada ovul segera setelah penyerbukan (Brownfield et al. 2009). Keefektifan penyerbukan dan pembuahan akan mempengaruhi pembentukan buah dan biji. Viabilitas polen yang rendah menyebabkan polen tidak dapat berkecambah yang pada akhirnya akan mempengaruhi efisiensi pembuahan (Widiastuti dan Palupi 2008). Kegagalan dalam penyerbukan dapat disebabkan karena tidak adanya polen yang sesuai. Inkompatibilitas dapat berupa tidak melekatnya polen pada stigma sehingga polen tidak berkecambah (Wilcock & Neiland 2002).
Pengujian viabilitas polen dengan menggunakan pewarna I2KI telah banyak digunakan, diantaranya pengujian viabilitas polen Musa (Ssali et al. 2012), polen pada bunga Carica papaya L. (Junior et al. 2009), polen pada bunga Oriza sativa
(Jong et al. 2010), polen pada Manihot esculenta Crantz (Parera et al. 2012), polen bunga tropical japonica rice (Hairmansis et al. 2005) dan polen pada bunga
Elaeagnus umbellata (Ma & Han 2010).
6
3 BAHAN DAN METODE
Waktu dan tempat penelitianPenelitian dilaksanakan bulan Desember 2011 – Februari 2013. Analisis karakter agronomi dilakukan di kebun percobaan IPB Cikabayan. Analisis isozim, analisis kromosom dan viabilitas polen Jarak pagar dilaksanakan di Laboratorium Hayati, Pusat Studi Bioteknologi dan Sumberdaya Hayati IPB.
Bahan penelitian
Bahan tanaman yang digunakan adalah lima genotipe biji jarak pagar yaitu biji asal bunga betina (BTB) dan hermaprodit (BTH) tanaman trimonoecious asal daerah Banten dan biji asal bunga betina (LTB) dan hermaprodit (LTH) tanaman trimonoecious asal daerah Lampung, serta biji dari bunga betina monoceius asal daerah Banten (BMB). Tanaman trimonoecious ini merupakan bibit yang berasal dari tanaman jarak pagar andromonoecious.
Tahapan penelitian
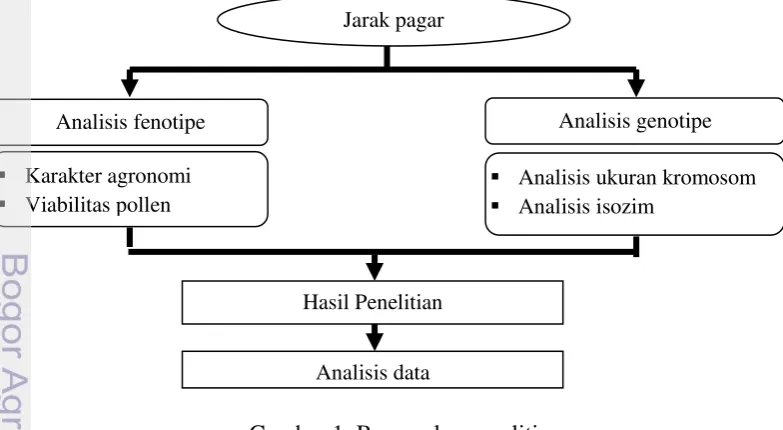
Penelitian yang dilakukan dengan menggunakan tanaman Jatropha curcas ini terdiri dari beberapa bagian analisis yang terdiri dari analisis uuran kromosom, isozim dengan pewarnaan esterase dan peroksidase, analisis viabilitas pollen dan analisis karakter agronomi. Tahapan penelitian ini dapat dilihat pada bagan alir di bawah ini (Gambar 1):
Gambar 1 Bagan alur penelitian
Jarak pagar
trimonoecious
Analisis fenotipe
Karakter agronomi
Viabilitas pollen
Analisis genotipe
Analisis ukuran kromosom
Analisis isozim
Hasil Penelitian
7
Penanaman dan pemeliharaan
Masing-masing genotipe terpilih dibibitkan dalam polybag dengan daya tampung 6 kg tanah. Tanaman berumur 1 BST (bulan setelah tanam) siap unuk dipindahkan ke lapang. Penanaman bibit dilakukan pada lubang tanam berukuran 40 x 40 x 40 cm dengan jarak tanam 2 x 2 m. Pemupukan dilakukan pada saat penanaman bibit di lapangan dengan dosis 2.5 kg pupuk kandang + 20 g urea + 20 g SP 36 + 10 g KCl (Mahmud et al. 2008). Penyiraman dilakukan tiga kali seminggu.
Analisis karakter agronomi
Pengamatan karakter agronomi terdiri atas karakter kualitatif dan karakter kuantitatif mengikuti Panduan Pengujian Individual ubi kayu (Manihot esculenta
Crantz.) (PVT 2007).
A. Pengamatan karakter kualitatif meliputi
Pengamatan karakter agronomi terdiri atas karakter kualitatif dan karakter kuantitatif. Pengamatan dilakukan pada seluruh tanaman dari setiap genotipe pada masing-masing unit percobaan. Pelaksanaan pengamatan karakter agronomi mengikuti Panduan Pengujian Individual ubi kayu (Manihot esculenta Crantz.) (Arisanti 2010). Pengamatan karakter kualitatif
Pengamatan batang yang dilakukan saat tanaman berumur 6 BST (bulan setelah tanam) adalah: (1) Tipe tanaman (tidak bercabang atau bercabang); (2) Warna batang muda diamati menggunakan Munsel Color Chart yaitu dengan mengamati batang bagian atas yang dekat dengan pucuk; (3) Warna batang tua diamati menggunakan Munsel Color Chart yaitu dengan mengamati batang bagian bawah; (4) Bentuk batang.
Pengamatan daun yang dilakukan pada saat tanaman berumur 4 BST adalah: (1) Bentuk daun; (2) Bentuk ujung daun (runcing atau tumpul); (3) Tipe tulang daun; (4) Bulu pada daun muda, ada atau tidak ada bulu pada daun muda diamati pada daun yang berada di bagian pucuk yang belum terbuka sempurna; (5) Warna daun muda diamati pada daun ke-2 dibagian pucuk yang belum membuka sempurna; (6) Warna daun tua diamati pada daun ke-15 yang berada pada batang bagian bawah menggunakan Munsel Color Chart; (7) Tekstur daun muda diamati pada daun ke-2 pada bagian pucuk yang belum membuka sempurna; (8) Tekstur daun tua diamati pada daun ke-15 yang berada pada batang bagian bawah.
8
B. Pengamatan karakter kuantitatif meliputi
Pengamatan batang yang terdiri dari karakter sudut cabang primer, diameter pangkal batang, dan panjang ruas dilakukan pada saat tanaman berumur 6 BST yang terdiri dari: (1) Sudut cabang primer; (2) Diameter pangkal batang, diamati dengan mengukur bagian pangkal batang 5 cm di atas permukaan tanah; (3) Panjang ruas; (4) Tinggi tanaman pada saat muncul bunga pertama, diukur pada saat muncul bunga pertama; (5) Jumlah cabang primer; (6) Jumlah cabang sekunder; (7) Jumlah cabang produktif.
Pada pengamatan daun yang diamati adalah karakter panjang tangkai daun, panjang daun dan lebar daun dilakukan pada saat tanaman berumur 5 BST sebanyak 5 daun pada setiap tanaman.
Pengamatan bunga dilakukan pada semua bunga yang muncul di setiap malai pada tanaman saat tanaman berumur 6 BST. Karakter yang diamati terdiri dari: (1) Umur berbunga diamati pada saat muncul bunga pertama; (2) Jumlah bunga jantan per malai; (3) Jumlah bunga betina per malai; (4) Jumlah bunga hermaprodit per malai; (5) Jumlah malai per tanaman. Pengamatan karakter bunga dilakukan selama 7 bulan setelah inisiasi bunga, Juni – Desember 2012.
Pada pengamatan buah matang fisiologis dilakukan pada semua buah yang terbentuk pada setiap malai. Karakter yang diamati adalah: (1) Tebal daging buah; (2) Panjang buah masak; (3) Diameter buah masak; (4) Jumlah buah per tanaman, diamati dengan menghitung buah yang terbentuk selama periode satu kali panen; (5) Jumlah buah per malai; (6) Bobot buah rata-rata.
Pada pengamatan karakter biji yang diamati adalah: (1) Panjang biji; (2) Tebal biji; (3) Jumlah biji rata-rata per buah; (4) Jumlah biji per tanaman, dihitung dengan mengalikan jumlah buah per tanaman dengan jumlah biji rata-rata per buah; (5) Bobot basah biji, diukur dengan menimbang setiap biji dari buah matang fisiologis yang dipanen dan dirata-ratakan; (6) Bobot kering biji, diukur dengan menimbang sejumlah biji yang telah dikeringkan dan dirata-ratakan.
Analisis ukuran kromosom
Ujung akar dipanen dari tunas yang berakar pada tanaman jarak pagar berumur 1 minggu, dipotong ± 1 cm dari ujung akar, dan dimasukkan ke dalam botol yang berisi hidroksiquinolin 0.8%, dan dibiarkan selama 3-5 jam pada suhu 20°C. Selanjutnya dibilas dengan aquades hingga bersih. Akar kemudian direndam dalam larutan asam asetat 45% selama 10 menit, dipindahkan dalam larutan 1 N HCl : asam asetat 45% (3:1, v/v) dan dibiarkan selama 3 menit dalam
water bath bersuhu 60°C.
9
Preparat selanjutnya diamati di bawah mikroskop dengan perbesaran 400x dan 1.000x. Panjang kromosom diukur menggunakan aplikasi ImageJ 1.42q (http://rsbweb.nih.gov/ij/).
Analisis isozim
Metode analisis isozim yang digunakan merupakan modifikasi dari teknik Soltis & Soltis (1989). Analisis isozim meliputi penyiapan bahan tanaman, pembuatan buffer, pembuatan gel pati, ekstraksi enzim, elektroforesis, pewarnaan dan pencucian serta pengumpulan data.
Persiapan bahan tanaman yang digunakan untuk analisis isozim adalah daun yang masih segar (urutan ketiga dari pucuk). Daun tanaman jarak pagar dipetik dari pohon di lokasi dan disusun pada kertas koran basah. Setelah itu masing-masing daun yang telah disusun dengan koran basah dimasukkan ke dalam kantong plastik dan diberi label. Selanjutnya di laboratorium, contoh daun tersebut dipindahkan ke dalam refrigerator untuk digunakan sebagai bahan ekstraksi enzim. Daun dianalisis berupa ekstrak bahan kasar.
Selanjutnya dilakuan pembuatan buffer pengekstrak, fungsi buffer pengekstrak ini adalah dapat membantu menghancurkan sel dalam suatu jumlah minimum tanpa menimbulkan panas terhadap ekstrak dan perubahan warna terhadap daun yang diekstrak.
Gel dibuat dari pati kentang dengan kadar 13% (Sigma). Pati kentang dan bufer gel dimasukkan ke dalam labu didih, dikocok merata, dan dimasak suhu 100°C, sambil diputar agar larutan pati masak merata. Gelembung udara yang ada dalam larutan dibuang dengan menggunakan pompa vakum. Larutan selanjutnya dituangkan ke cetakan gel yang sudah dilapisi parafin. Gel kemudian didinginkan, ditutup plastik wrap, dan dibiarkan sampai mengeras.
Proses ekstraksi enzim dilakukan dengan menggerus sampel daun sebanyak 2 gram yang telah di beri buffer ekstrak sebanyak 0,5 ml dan pasir kuarsa dengan menggunakan mortar yang telah didiamkan di dalam lemari pendingin selama 5 menit. Untuk pemindahan sampel ke dalam cetakan gel dilakukan dengan cara menyerap ekstrak sampel dengan kertas saring, kemudian dimasukkan ke dalam gel. Pada salah satu lubang yang paling pinggir disisipkan kertas saring yang telah diberi cairan bromophenol blue sebagai indikator mobilitas elektroforesis.
Proses elektroforesis dilakukan pada gel yang telah siap dibuat lubang/sumur sebagai tempat sampel. Kertas saring yang mengandung sampel disisipkan ke dalam sumur. Cetakan gel kemudian dimasukkan ke dalam sistem reservoar elektroforesis yang berisi bufer elektroda. Elektroforesis dilakukan dalam ruang pendingin pada suhu 10°C. Waktu yang dibutuhkan untuk elektroforesis adalah 30 menit dengan tegangan awal 100 volt, kemudian pada 200 volt selama 3-4 jam.
pita-10
pita pada gel yang cukup jelas. Perendaman dalam larutan pewarna memerlukan waktu antara 3 sampai 4 jam atau lebih. Setelah pewarnaan selesai selanjutnya gel dicuci dengan air bersih untuk menghilangkan sisa pewarna.
Interpretasi pola pita isozim dilakuan dengan meletakkan gel di atas plastik bening dan diletakkan di atas lampu pengamatan untuk diambil data dan difoto. Hasil pengukuran jarak pergerakan ditentukan dengan pola pada Rf (relative electrophoresis mobility) (Sastrosumarjo et al. 2006):
Rf =
Uji viabilitas pollen
Sampel polen untuk uji viabilitas polen bunga jantan diambil dari 17 bunga jantan dari 4 tanaman jarak pagar trimonoecious dan 17 bunga jantan dari 1 tanaman jarak pagar monoecious. Pengambilan bunga jantan dilakukan pada pukul 09.00-10.00 WIB. Pada uji pewarnaan, polen diambil dari bunga jantan yang telah antesis. Polen diambil dengan bantuan tusuk gigi atau pinset.
Polen ditetesi dengan pewarna I2KI 0,2% selama beberapa menit dan diamati di bawah mikroskop. Pada setiap genotipe dilakukan 3 kali ulangan dengan jumlah minimum 50 butir polen untuk masing-masing ulangan. Untuk mempermudah perhitungan, pengamatan dilakukan hanya terhadap butir-butir polen yang tidak bergerombol. Butir polen yang terwarnai hitam dianggap viabel dan dinyatakan dalam hitungan persen (Scott 2001).
Analisis data
Percobaan dilakukan dengan menggunakan Rancangan Acak Kelompok (RAK) satu faktor yaitu genotipe jarak pagar. Analisis viabilitas polen dan karakter agronomi menggunakan analisis sidik ragam dan dilanjutkan dengan uji lanjut Duncan Multiple Range Test (DMRT) taraf 5%.
Rumus parameter genetik yang digunakan adalah sebagai berikut (Singh dan Chaudhary 1979):
r = ulangan KKG = koefisien keragaman genetik
11
Nilai ragam lingkungan diperoleh dari tabel ANOVA, selanjutnya nilai heritabilitas diperoleh dari hasil pembagian antara ragam genotipe dibagi dengan ragam genotipe dibagi ragam lingkungan (Mohammadi & Amri 2011). Pengelompokan nilai heritabilitas arti luas menurut Stansfield (1983): rendah (h2 < 20%), sedang (20% < h2 < 50%) dan tinggi (50% < h2 < 100%). Kriteria KKG relatif adalah rendah (0 < x < 25%), agak rendah (25% < x < 50%), cukup tinggi (50% < x < 75%), dan tinggi (75% < x ≤ 100%) (Moedjiono dan Mejaya 1994). Kemiripan antar genotipe dicari berdasarkan pada koefisien kesamaan (similarity coefficients). Analisis pengelompokan jarak genetik dilakukan dengan metode pengelompokan program minitab 15.
4 HASIL DAN PEMBAHASAN
Kondisi umum lokasi penelitian
Percobaan dilaksanakan di Kebun Percobaan Cikabayan, Institut Pertanian Bogor yang berada pada ketinggian 216 m dpl, 06°31’ LS dan 106°44’ BT pada bulan Desember 2011 – Februari 2013 Berdasarkan data Badan Meteorologi Klimatologi dan Geofisika Darmaga Bogor, Kota Bogor memiliki rata-rata curah hujan tinggi (323.63 mm/bln), jumlah hari hujan 297 hari/tahun, suhu harian antara 21.2°C – 32.4°C, jumlah rata-rata bulan basah 12 bulan/tahun. Data klimatologi wilayah Darmaga Bogor dari Desember 2011 – Februari 2013 disajikan pada Lampiran 2.
Analisis karakter agronomi
Karakter batang
12
a b c
Gambar 2 Bentuk batang dan warna batang; (a) batang muda tanaman monoecious umur 2 BST; (b) batang muda tanaman trimonoecious umur 2 BST; (c) batang tua umur 4 BST
a b c d e
Gambar 3 Cabang yang muncul dari batang utama pada genotipe jarak pagar umur 6 BST; (a) BTB: Banten trimonoecious betina; (b) BTH: Banten trimonoecious hermaprodit; (c) LTH: Lampung trimonoecious hermaprodit; (d) LTB: Lampung trimonoecious betina; (e) BMB: Banten monoecious betina
Semua karakter batang pada genotipe BMB mempunyai ukuran lebih kecil dibandingkan genotipe yang berasal dari tanaman trimonoecious (Tabel 1), sehingga pertumbuhannya lebih tegak. Jumlah cabang primer dan jumlah cabang sekunder mempengaruhi tipe pertumbuhan tanaman jarak pagar (Santoso 2009). Sudut cabang tanaman trimonoecious lebih besar dibandingkan tanaman monoecious, sehingga semakin besar sudut cabang primer maka arstitektur kanopi lebih rimbun (Raden et al. 2009) (Gambar 4). Cabang produktif adalah cabang yang menghasilkan buah. Jumlah cabang akan menentukan jumlah bunga, buah dan biji karena bunga jarak pagar muncul disetiap ujung batang atau ketiak daun. Sehingga semakin banyak cabang diharapkan akan meningkatkan jumlah bunga dan buah yang dihasilkan (Xiu dan Gui 2012).
0.5 cm 1 cm
0.5 cm
13
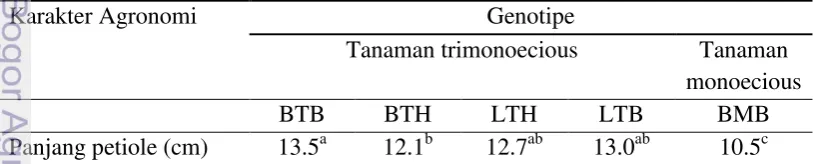
Tabel 1 Karakter batang dan daun umur 4 - 6 BST jarak pagar trimonoecious dan monoecious
Huruf yang sama pada lajur yang sama menunjukkan tidak berbeda nyata (uji DMRT taraf 5%). BTB: Banten trimonoecious betina; BTH: Banten trimonoecious hermaprodit; LTH: Lampung trimonoecious hermaprodit; LTB: Lampung trimonoecious betina; BMB: Banten monoecious betina
a b c d e
Gambar 4 Pertumbuhan tanaman jarak pagar umur 6 BST; (a) BTB: Banten trimonoecious betina; (b) BTH: Banten trimonoecious hermaprodit; (c) LTH: Lampung trimonoecious hermaprodit; (d) LTB: Lampung trimonoecious betina; (e) BMB: Banten monoecious betina
Diameter pangkal batang tanaman monoecious lebih kecil dibandingkan tanaman trimonoecious, sehingga pertumbuhan tanaman trimonoecious lebih kokoh karena diameter batang yang besar mempunyai kemampuan pertumbuhan yang lebih besar dan tidak mudah roboh (Sudjijo 2009). Ukuran diameter pangkal batang suatu genotipe akan bertambah seiring dengan semakin bertambahnya jumlah cabang primer, karena percabangan (cabang primer) banyak terbentuk di pangkal batang dekat permukaan tanah (Santoso 2009).
Karakter panjang buku dipengaruhi oleh jenis tanaman monoecious dan trimonoecious, panjang buku pada tanaman monoecious memiliki panjang buku yang lebih rendah dibandingkan tanaman trimonoecious.
10 cm 10 cm
14
Karakter daun
Daun jarak pagar bertipe daun tunggal yang terletak pada buku batang yang didukung oleh tangkai daun, dengan tangkai daun berbentuk silinder dan tak berongga. Susunan atau tata letak daun (filotaksis) jarak pagar disebut tersebar (folia sparsa). Susunan daun tersebut mengikuti rumus daun (divergensi) 5/13 serah putaran jarum jam. Orientasi daun terhadap batang tempat daun duduk bervariasi dari tegak hingga horisontal. Orientasi tegak jika daun masih muda dan kemudian menjadi horisontal setelah dewasa. Daun jarak pagar berbentuk jantung dengan bentuk ujung daun yang runcing dan pada pangkal daun berlekuk dalam, memiliki tipe tulang daun menjari dengan lima tulang daun utama (Gambar 5).
a b c d e
Gambar 5 Daun jarak pagar 4 BST; (a) BTB: Banten trimonoecious betina; (b) BTH: Banten trimonoecious hermaprodit; (c) LTH: Lampung trimonoecious hermaprodit; (d) LTB: Lampung trimonoecious betina; (e) BMB: Banten monoecious betina
Daun muda tidak memiliki bulu. Tekstur permukaan atas dan bawah daun muda dan daun tua pada jarak pagar licin. Tangkai daun yang menghubungkan helaian daun dengan batang umumnya berwarna ungu khususnya pada pangkal (dekat buku) dan ujung tangkai daun. Seiring pertumbuhan dan perkembangan daun, warna ungu berkurang bahkan menghilang dan menjadi warna kuning kehijauan. Daun muda pada tanaman jarak pagar umumnya berwarna coklat (coklat kemerahan, hijau kecoklatan, colat kehijauan). Daun tua berwarna hijau pada keseluruhan genotipe.
Tabel 2 Karakter daun jarak pagar trimonoecious dan monoecious 4 BST
Karakter Agronomi Genotipe BTB: Banten trimonoecious betina; BTH: Banten trimonoecious hermaprodit; LTH: Lampung trimonoecious hermaprodit; LTB: Lampung trimonoecious betina; BMB: Banten monoecious betina
Panjang tangkai daun dipengaruhi oleh jenis tanaman jarak pagar pada masing-masing genotipe. Panjang tangkai daun antara genotipe terdapat perbedaan. Masing-masing genotipe memiliki panjang tangkai daun yang berbeda.
15
Ukuran panjang tangkai daun yang paling panjang terdapat pada genotipe Banten trimonoecious sedangkan genotipe Banten monoecious memiliki panjang tangkai daun terpendek. Hal ini dapat dijadikan salah satu penanda antara tanaman trimonoecious dan monoceious. Tanaman trimonoecious memiliki ukurang tangkai daun yang lebih panjang dibandingkan tanaman monoecious. Pada daun-daun muda, tangkai daun-daun membentuk sudut kecil kemudian melebar hingga mencapai maksimum umur 14 minggu terhadap posisi cabang (Raden et al 2008).
Karakter Bunga bunga hermaprodit, (c) bunga jantan, (d) bunga betina
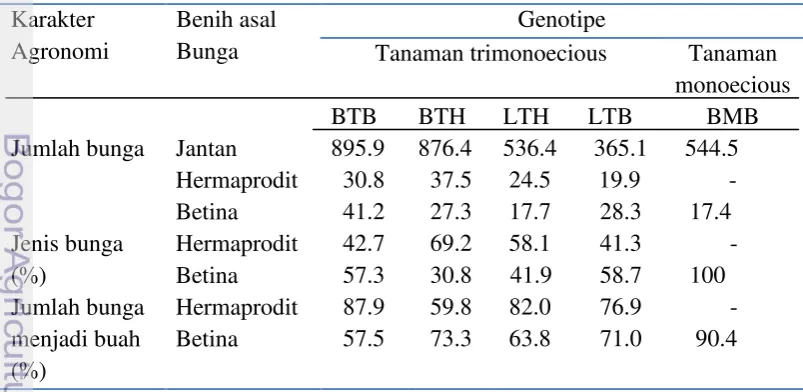
Tabel 3 Karakter bunga bunga jarak pagar trimonoceious dan monoecious per BTB: Banten trimonoecious betina; BMB: Banten monoecious betina; BTH: Banten trimonoecious hermaprodit; LTH: Lampung trimonoecious hermaprodite; LTB: Lampung trimonoecious betina
0.1 cm 0.1 cm
16
Jumlah bunga betina dan hermaprodit lebih sedikit dibandingkan dengan bunga jantan (Tabel 3). Hal ini dapat disebabkan potensi pembentukan bunga betina dan hermaprodit memang rendah, tetapi dapat juga dipengaruhi oleh faktor lingkungan atau terlalu banyak hujan dan lain sebagainya (Xiu & Gui 2012). Selain itu menurut Jun et al. (2010) bahwa jarak pagar memiliki bunga betina yang tidak muncul di situs bunga jantan, tetapi bunga jantan dapat berkembang di situs bunga betina sehingga jumlah bunga jantan lebih banyak.
17
(a)
(b)
Gambar 7 (a) Fluktuasi pembungaan tanaman trimonoecious asal bunga hermaprodit; (b) Fluktuasi pembungaan tanaman trimonoecious asal bunga betina
Perkembangan bunga tanaman trimonoecious asal bunga hermaprodit pada empat bulan pertama (Juni – September) menghasilkan bunga betina dan hermaprodit, bulan selanjutnya hanya menghasilkan bunga hermaprodit. Perkembangan tanaman trimonoecious asal bunga betina pada bulan Juni hanya menghasilkan bunga betina, bulan selanjutnya menghasikan bunga hermaprodit dan betina (Gambar 7) (Lampiran 3). Fluktuasi perkembangan jenis bunga yang muncul setiap bulan dapat dipengaruhi oleh ekspresi gen-gen yang mengendalikan jenis gamet pada tanaman yang sangat dipengaruhi oleh lingkungan seperti fotoperiode, suhu dan lain sebagainya (Roy 2000). Berdasarkan data dari BMKG Darmaga Bogor, rata-rata curah hujan 117 – 217 mm/bulan pada bulan Juli hingga September 2012. Pada rentang waktu tersebut muncul bunga hermaprodit. Hal ini sesuai dengan penelitian Jun et al. (2010) bahwa bunga hermaprodit pada jarak pagar paling banyak dihasilkan pada saat empat bulan pembungaan pertama (Juni – September) dengan curah hujan yang tidak terlalu tinggi hanya berkisar < 500 mm/tahun. Curah hujan yang optimal akan meningkatkan perkembangan bunga betina dan hermaprodit.
Ekspresi seksual pada tanaman berbunga berada di bawah kendali genetik, seperti faktor epigenetik (kondisi lingkungan, fitohormon, fotoperiode dll). Faktor fotoperiodisme pada tanaman Hemp dapat menimbulkan pengaruh maskulinisasi maupun feminisasi. Hari panjang dan meningkatnya suhu lingkungan dapat menginisiasi pembentukan kelamin jantan, sedangkan hari pendek dan menurunnya suhu dapat menginisiasi pembentukan kelamin betina (Manoj et al.
-5 0 5 10 15
June July Aug Sept Oct Nov Dec
J
June July Aug Sept Oct Nov Dec
18
2005). Oleh karena itu, fluktuasi yang terjadi pada kedua jenis tanaman jarak pagar tersebut biasa saja terjadi, dikarenakan pengaruh lingkungan.
Perkembangan bunga betina mengalami penurunan jumlah, sedangkan jumlah bunga hermaprodit cenderung konstan dan bahkan meningkat (Gambar 8a). Hal ini dapat terjadi karena antara bunga betina dan hermaprodit berbeda dalam pola alokasi fotosintat. Pada kondisi lingkungan tercekam bunga betina dapat bertahan. Pola ini menunjukkan bahwa bunga betina dapat mengalokasikan fotosintat sehingga lebih meningkatkan kemampuan bertahan pada kondisi lingkungan yang tidak menguntungkan dibandingkan bunga hermaprodit, sehingga berdampak pada berkurangnya jumlah bunga betina seiring perkembangan tanaman (Delph 2003). Hal ini berkaitan dengan hasil penelitian Miyake et al. (2009) bahwa total berat buah, jumlah buah, jumlah biji dan jumlah bunga dipengaruhi oleh biomassa. Ketika biomassa digunakan dalam proses pertahanan maka pembentukan bunga tersebut akan menyebabkan perbedaan tingkat fitness pada kedua jenis bunga betina dan hermaprodit. Hal yang sama dikemukakan oleh Ramsai & Vaughton (2001), bahwa bunga betina lebih banyak mengalokasikan simpanan cadangan ataupun unsur hara ke dalam biji dibandingkan bunga hermaprodit. Bunga betina dan hermaprodit memiliki strategi investasi hara yang berbeda.
Karakter buah
19
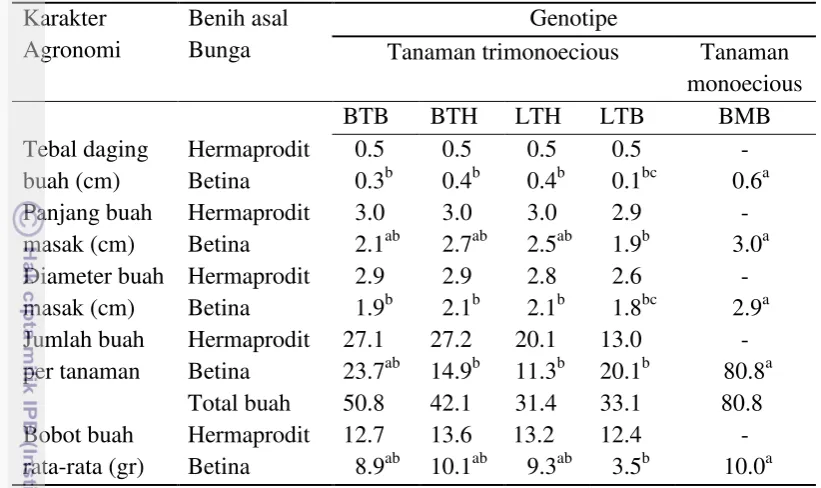
Tabel 4 Karakter buah 5-12 BST jarak pagar trimonoecious dan monoecious Karakter
Huruf yang sama pada lajur yang sama menunjukkan tidak berbeda nyata (uji DMRT taraf 5%). BTB: Banten trimonoecious betina; BTH: Banten trimonoecious hermaprodit; LTH: Lampung trimonoecious hermaprodit; LTB: Lampung trimonoecious betina; BMB: Banten monoecious betina
Jumlah buah pertanaman hanya dipengaruhi oleh genotipe tanaman jarak pagar trimonoecious asal buah dari bunga betina. Tingginya jumlah buah betina yang dihasilkan pada genotipe BMB dikarenakan, pada genotipe ini hanya menghasilkan bunga betina saja, sedangkan genotipe lain selain menghasilkan bunga betina juga menghasilkan bunga hermaprodit (Tabel 4). Semakin banyak jumlah bunga betina dan hermaprodit maka akan semakin banyak pula jumlah buah yang dihasilkan, karena bunga betina dan hermaprodit yang akan menjadi buah setelah terjadi antesis. Meskipun demikian tidak semua bunga menghasilkan buah karena pengaruh beberapa faktor, antara lain adalah bunga betina yang telah mengalami penyerbukan gagal berkembang menjadi buah, bunga betina terkena penyakit seperti tepung putih, buah yang masih inisiasi menguning / diserang kepik penghisap (Ahkamulloh et al. 2013).
Ukuran buah dari hermaprodit memiliki nilai yang lebih besar dibandingkan dengan buah asal bunga betina, dilihat dari tebal daging buah, panjang dan diameter buah masak. Adanya kemampuan menghasilkan buah yang lebih baik dari bunga hermaprodit dibandingkan bunga betina dilaporkan oleh Reale et al
20
dibandingkan dengan bunga betina yang penyerbukannya tergantung dari bunga jantan yang mekar tidak bersamaan dengan bunga betina.
Karakter biji
Biji jarak pagar sesaat setelah dipanen tampak berwarna hitam kecoklatan. Bentuk biji oval sampai melonjong, tergantung pada bentuk buah, dan jumlah biji yang terdapat pada satu buahnya (Gambar 9). Semakin banyak biji yang terdapat dalam satu buah maka ukurannya relatif lebih kecil, sedangkan jika biji yang terdapat pada satu buah hanya sekitar 1 - 2 biji, maka ukurannya dapat lebih besar.
a b c
d e
21
Huruf yang sama pada lajur yang sama menunjukkan tidak berbeda nyata (uji DMRT taraf 5%). BTB: Banten trimonoecious betina; BTH: Banten trimonoecious hermaprodit; LTH: Lampung trimonoecious hermaprodit; LTB: Lampung trimonoecious betina; BMB: Banten monoecious betina
Panjang biji buah betina dari genotipe LTB memiliki perbedaan dibandingkan dengan biji genotipe lainnya yang memiliki panjang biji yang paling kecil. Bobot basah dan bobot kering biji asal bunga betina dipengaruhi oleh gentipe jarak pagar trimonoecious dan monoceious. Bobot basah biji, bobot kering biji dari buah asal bunga hermaprodit dan bobot total kering biji dari buah asal betina dipengaruhi oleh genotipe tanaman jarak pagar. Bentuk dan ukuran biji tanaman trimonoecious dan monoecious relatif sama (Tabel 5). Periode perkembangan buah yang panjang terutama bila terjadi pada kondisi lingkungan yang baik akan menambah bobot biji (Foidl et al. 1996).
Analisis ukuran kromosom
Berdasarkan panjang kromosom, tanaman jarak pagar memiliki kromosom berukuran kecil. Oleh karena itu, disarankan untuk mengidentifikasi kromosom jarak pagar dilakukan pada saat sel dalam keadaan prometaphase, karena ukuran kromosom lebih panjang dan struktur kromosom lebih jelas.
22
Tanaman jarak pagar kelima genotipe memiliki jumlah kromosom 2n = 22, sesuai dengan penelitian yang dilakukan oleh Carvalho et al. (2008). Berdasarkan panjang kromosom, jarak pagar memiliki kromosom berukuran kecil, panjangnya berkisar antara 1.19 –2.69 μm (Tabel 6). Data ini sesuai dengan penelitian Reddy
et al. (2013) dengan ukuran kromosom jarak pagar berkisar 1–3.67 μm. Tabel 6 Panjang kromosom jarak pagar trimonoecious dan monoecious
Pasangan
BTB: Banten trimonoecious betina; BTH: Banten trimonoecious hermaprodit; LTH: Lampung trimonoecious hermaprodit; LTB: Lampung trimonoecious betina; BMB: Banten monoecious betina
23
a b c d e
Gambar 10 Kromosom masing-masing genotipe jarak pagar trimonoecious dan monoecious (a) BTB: Banten trimonoecious betina; (b) BTH: Banten trimonoecious hermaprodit; (c) LTH: Lampung trimonoecious hermaprodit; (d) LTB: Lampung trimonoecious betina; (e) BMB: Banten monoecious betina
Analisis isozim
Pada zimogram dapat dilihat bahwa isozim EST dan PER masing-masing menunjukkan adanya perbedaan jumlah pita. Kedua isozim peroksidase dan esterase yang diujikan memberikan polimorfisme pada pola pitanya. Perbedaan jarak pagar monoecious dan trimonoecious dapat terlihat dengan menggunakan kedua enzim tersebut.
Enzim esterase (EST) dari lima genotipe jarak pagar yang diamati hanya dibedakan menjadi dua pola pita dengan tiga sampai empat pita. Pola pita pada genotipe BTB, BTH, LTH dan LTB mempunyai kesamaan yaitu memiliki empat pita. Hanya genotipe BMB yang memiliki tiga pita (Gambar 11), sehingga dapat dikatakan terdapat keragaman pola pita isozim yang kemungkinan menyandi enzim tersebut.
a b
Gambar 11 Pola pita (a) isozim esterase dan (b) Peroksidase. BTB: Banten trimonoecious betina; BTH: Banten trimonoecious hermaprodit; LTH: Lampung trimonoecious hermaprodit; LTB: Lampung trimonoecious betina; BMB: Banten monoecious betina
2 μm 2 μm
2 μm 2 μm
24
Enzim peroksidase (PER) membedakan genotipe jarak pagar yang diamati ke dalam dua variasi pola pita dengan memiliki tiga sampai empat alel. Pola pita pada genotipe BTB, BTH, LTH dan LTB mempunyai kesamaan yaitu memiliki empat pita. Hanya genotipe BMB yang memiliki tiga alel (Gambar 12). Adanya enzim peroksidase mudah dideteksi karena aktivitas dan stabilitasnya yang tinggi dan dapat menggunakan sejumlah substrat sebagai donor hidrogen (Cahyarini 2004). Hasil analisis kekerabatan menggunakan isozim esterase dan peroksidase menunjukkan variasi genetik yang cukup rendah pada kelima genotipe Jarak pagar.
Isozim dalam penelitian ini berperan sebagai penanda genetik untuk mengidentifikasi genotipe jarak pagar trimonoecious. Sifat morfologi yang membedakan antara jarak pagar monoecious dan trimonoecious seperti bunga dan buah memerlukan waktu cukup lama sampai bisa diamati serta sulit digunakan untuk pemeriksaan pada tingkat benih atau bibit. Pada Gambar 11 dapat terlihat bahwa isozim EST dan PER dapat digunakan sebagai penanda tanaman J. curcas
monoecious dan trimonoecious ketika tanaman masih dalam keadaan bibit dengan menggunakan sampel daun. Seperti Sharma et al. (2010) yang menggunakan enzim peroksidase sebagai penanda alat reproduksi betina pada Hippophae rhamniodes.
Berdasarkan hasil elektroforesis baik dengan pewarnaan peroksidase maupun esterase, keragaman pola pita isozim lebih cenderung tergolong ke dalam keragaman secara kualitatif, yaitu ada atau tidaknya pita pada gel, sedangkan ketebalan pita yang merupakan sifat kuantitatif sebagian besar adalah sama. Pola pita tanaman trimonoecious dan monoecious menghasilkan variasi pola pita yang berbeda. Hal ini menunjukkan adanya perbedaan genetik pada kedua tanaman tersebut. Perbedaan ini terjadi karena adanya lebih dari satu gen dalam tiap tanaman tersebut yang mengkode setiap isozim (Dewatisari et al. 2008).
Pada umunya tanaman memiliki satu daerah aktif pada sitoplasma dan satu daerah di plastid yang masing-masing dikendalikan 1 lokus. Isozim merupakan produk gen, sehingga analisisnya dapat mencerminkan secara langsung aktivitas gen, termasuk bila terjadi perubahan dalam sekuen DNA yang mengakibatkan perubahan asam aminonya (Hartana 2003). Enzim sebagai golongan protein dalam jumlah paling banyak di dalam sel, mempunyai fungsi sebagai katalisator reaksi biokimia. Protein itu sendiri merupakan ekspresi dari DNA dimana proses sintesisnya melalui penerjemahan kodon-kodon pada mRNA menjadi polipeptida, dan setiap kodon tersusun atas basa-basa nukleotida (Wirahadikusuma 2001).
Analisis viabilitas polen
25
dikandungnya, sehingga diasumsikan semakin tinggi kandungan pati dalam polen, semakin tinggi viabilitas polen tersebut (Junior et al. 2009).
Pada pengamatan terdapat dua perbedaan intensitas warna yaitu hitam dan tidak berwarna/bening (Gambar 12). Butir polen yang berwarna hitam adalah butir polen yang viabel, dan yang tidak berwarna adalah butir polen yang tidak viabel. Perhitungan terhadap polen yang viabel hanya dilakukan pada butir polen yang berwarna hitam. Uji yang sama, yaitu uji pewarnaan I2KI juga pernah dilakukan oleh Abdelgadir et al. (2011) untuk mengetahui tingkat viabilitas polen pada tanaman J. curcas.
a b
Gambar 12 Bentuk, Warna dan ukuran polen jarak pagar (a) polen viabel (b) polen aviabel
Tabel 7 Viabilitas polen bunga jantan pada jarak pagar monoecious dan trimonoecious
Huruf yang sama pada lajur yang sama menunjukkan tidak berbeda nyata (uji DMRT taraf 5%); umur berbunga terhitung sejak inisiasi bunga; BTB: Banten trimonoecious betina; BTH: Banten trimonoecious hermaprodit; LTH: Lampung trimonoecious hermaprodit; LTB: Lampung trimonoecious betina; BMB: Banten monoecious betina
Iodine Potassium Iodide (I2KI) merupakan senyawa yang sering dipakai juga untuk mendeteksi kandungan gula/pati. Pati berperan dalam menunjang polen, sehingga diasumsikan semakin tinggi kandungan pati dalam polen, semakin tinggi viabilitas polen tersebut (Junior et al. 2009). Pati pada setiap butir polen tersebut digunakan sebagai sumber energi sehingga mampu berkecambah. Oleh karena itu banyaknya pati yang dikandung dapat digunakan sebagai
26
indikator tingkat viabilitas polen. Semakin banyak kandungan pati maka viabilitas polen semakin tinggi. Hal ini ditunjukkan dengan semakin hitam (pekat) warna yang dihasilkan pada uji I2KI (Ssali et al. 2012).
Viabilitas polen bunga jantan jarak pagar tertinggi saat bunga dalam keadaan kuncup dan mekar hari ke-20 dan ke-22 sejak inisiasi bunga (Tabel 7), dan ini merupakan keadaan yang tepat untuk melakukan penyerbukan karena pada keadaan tersebut memiliki kandungan pati dan jumlah polen viabel yang tinggi sehingga mempunyai peluang berkecambah yang besar (Abdelgadir et al. 2011). Hal ini didukung oleh penelitian pada perkecambahan polen bunga labu yaitu polen yang diambil pada awal periode reseptif memiliki kemampuan berkecambah yang besar (Nepi dan Pacini 2001). Viabilitas polen juga dapat mempengaruhi viabilitas benih yang dihasilkan, polen dengan viabilitas tinggi akan lebih dahulu membuahi sel telur serta menghasilkan buah bermutu baik dan benih berviabilitas tinggi (Widiastuti dan Palupi 2008).
a b c d e Gambar 13 Karakter bunga/warna kepala sari bunga jantan jarak pagar. (a) hari
ke-21, (b) hari ke-22, (c) hari ke-23, (d) hari ke-24, (e) hari ke-25
Tanaman trimonoecious mengalami penurunan viabilitas polen yang lebih lambat dibandingkan tanaman monoecious. Hal ini disebabkan karena tanaman dengan rangkaian bunga hermaprodit memiliki kandungan pati yang tinggi pada pistil dan stamen dibandingkan tanaman tanpa bunga hermaprodit (Reale et al.
2009), sehingga mempengaruhi kecepatan penurunan viabilitas polen pada tanaman jarak pagar monoecious. Viabilitas polen yang menurun menunjukkan bahwa jumlah polen yang viabel juga menurun (Gambar 13). Hal ini dapat disebabkan oleh umur bunga yang semakin tua dan adanya degenerasi kandungan pati pada polen (Doyle et al. 2002). Selain itu, faktor lingkungan juga turut berperan terhadap durasi viabilitas polen, dengan dehidrasi atau cahaya UV merupakan salah satu faktor yang menyebabkan rusaknya atau menurunnya viabilitas polen (Dafni & Dange 2000)
Uji I2KI pada penelitian ini dapat membedakan beberapa tingkat viabilitas polen pada umur bunga yang berbeda. Hal ini sama pada penelitian yang dilakukan oleh Sasikala et al. (2009) yang meneliti viabilitas polen dari sepuluh jenis Jatropha spp menggunakan I2KI, dari kesembilan dari sepuluh spesies Jatropha memiliki tingkat kesuburan polen lebih dari 84%.
27
Pendugaan parameter genetik
Pendugaan parameter genetik bertujuan untuk mengetahui potensi genetik lima genotipe jarak pagar yang diuji. Analisis dilakukan terhadap ragam genotipe, ragam fenotipe, heritabilitas, koefisien keragaman genetik, dan koefisien keragaman fenotipe. Hasil analisis nilai duga parameter genetik disajikan pada Tabel 8. Untuk karakterisasi suatu tanaman hal yang perlu diperhatikan adalah karakter-karakter yang dapat diturunkan (Rahayu 2012), sehingga data yang disajikan hanya terbatas pada beberapa karakter yang memiliki nilai heritabilitas yang tinggi. Berdasarkan kriteria Stanfield (1983) Pengelompokan nilai heritabilitas tergolong tinggi jika (50% < h2 < 100%) dan KKG tergolong rendah jika (0 < x < 25%).
Tabel 8 Parameter genetik beberapa karakter kuantitatif jarak pagar trimonoecious
Karakter panjang ruas; JUR: jumlah ruas; JAR: jarak antar ruas; PTD: panjang petiole: HPM: jumlah bunga hermaprodit per malai: BPM: jumlah betina per malai; JMPT: Jumlah malai per tanaman; TDBB: Tebal daging buah tanaman asal bunga betina; PBMB: panjang buah masak asal bunga betina; DBMB: diameter buah masak asal bunga betina; JBPTB: jumlah buah per tanaman asal bunga betina; BBRH: Bobot buah rata-rata tanaman asal bunga hermaprodit; BBRB: bobot buah rata-rata tanaman asal bunga betina; PBB: panjang biji tanaman asal bunga betina; TBB: tebal biji tanaman asal bunga betina; BBBH: bobot biji buah hermaprodit; BKBH: bobot kering biji asal bunga
28
Berdasarkan kriteria Stanfield (1983), nilai heritabilitas beberapa karakter kuantitatif jarak pagar di atas tergolong tinggi. Karakter yang memiliki heritabilitas luas berarti seleksi terhadap karakter tersebut berlangsung efektif dan mampu meningkatkan potensi genetik karakter pada generasi selanjutnya (Zen dan Bahar 2001). Heritabilitas luas berarti faktor genetik lebih dominan terhadap karakter yang ditampilkan dibandingkan dengan faktor lingkungan dan akan diwariskan secara kuat terhadap keturunannya (Wicaksana 2001). Rendahnya nilai h2 berarti bahwa fenotipe tidak berkorelasi positif dengan genotipe, dengan kata lain kondisi lingkungan lebih mempengaruhi tanaman (Singh 2005). Nilai KKG pada karakter di atas menunjukkan nilai yang tergolong rendah, yang diasumsikan sebagai karakter yang memiliki keragaman genetik sempit.
Analisis gabungan data kualitatif, kuantitatif, ukuran kromosom, isozim dan pollen
Matrik kemiripan hasil dari analisis gabungan data kualitatif, kuantitatif, viabilitas polen, ukuran kromosom dan analisis isozim disajikan pada Tabel 9. Matrik kemiripan menunjukkan rentang kemiripan 0.080 – 0.965. nilai koefisien kemiripan tertinggi 0.965 diperoleh genotipe BMB dan BTB dan koefisien kemiripan terendah 0.080 pada genotipe BTH dan LTH. Berdasarkan hasil beberapa analisis tersebut maka diperoleh dua kelompok genotipe. Kelompok I terdiri dari BTB, BTH, LTH, dan LTB sedangkan kelompok II terdiri dari genotipe BMB dengan koefisien similaritas 9.42 (Gambar 14).
Tabel 9 Matrik kemiripan 5 genotipe jarak pagar trimonoecious dan monoecious berdasarkan gabungan data kualitatif, kuantitatif, viabilitas polen, ukuran kromosom dan analisis isozim
BTB: Banten trimonoecious betina; BTH: Banten trimonoecious hermaprodit; LTH: Lampung trimonoecious hermaprodit; LTB: Lampung trimonoecious betina; BMB: Banten monoecious betina
29
Gambar 14 Dendrogram analisis gabungan data kualitatif, kuantitatif, analisis isozim analisis ukuran kromosom, dan viabilitas polen. BTB: Banten trimonoecious betina; BTH: Banten trimonoecious hermaprodit; LTH: Lampung trimonoecious hermaprodit; LTB: Lampung trimonoecious betina; BMB: Banten monoecious betina
Hasil di atas dapat digunakan sebagai acuan dalam penentuan induk untuk pembuatan bibit. Semakin jauh hubungan kekerabatan antar sampel, maka semakin kecil keberhasilan persilangan, tetapi kemungkinan untuk memperoleh genotip unggul lebih besar jika persilangan berhasil. Semakin beragam genetik, maka semakin besar kemungkinan diperoleh genotipe unggul. Perkawinan antara individu berjarak genetik dekat atau hubungan kekerabatannya sama mempunyai efek peningkatan homozigositas, sebaliknya perkawinan antara individu berjarak genetik besar atau kekerabatannya jauh mempunyai efek peningkatan heterozigositas. Informasi ini berdampak baik bagi proses pembuatan bibit unggul. Perkawinan tetua dengan variasi genetik yang relatif tinggi akan menghasilkan individu dengan heterozigositas lebih tinggi (Juliansaniah et al
2008).
5 SIMPULAN DAN SARAN
SIMPULAN
30
peroksidase (Rf 0.83) dan esterase (Rf 0.20) dapat dijadikan penanda untuk membedakan tanaman trimonoecious dan monoecious. Viabilitas polen bunga jantan asal tanaman trimonoecious memiliki tingkat penurunan yang lebih lambat dibandingkan bunga jantan tanaman monoecious. Karakter tanaman trimonoecious pada penelitian ini memiliki karakter yang hampir sama dengan tetuanya. Dendogram hasil analisis kekerabatan dapat membedakan antara tanaman trimonoecious dan monoecious pada koefisien similarity 27.05.
SARAN
Studi jarak pagar perlu dilanjutkan untuk menggali berbagai informasi lainnya baik yang terkait dengan hasil tanaman maupun berbagai karakter-karakter penting lainnya seperti karakter-karakter buah masak serempak, karakter-karakter ketahanan terhadap cekaman biotik maupun abiotik.
DAFTAR PUSTAKA
Abdelgadir HA, Johnson SD, Staden JV. 2012. Pollen viability, pollen germination ad pollen tube growth in the biofuel seed crop Jatropha curcas
(Euphorbiaceae). SouthAfrican Journal of Botany. 79: 132-139.
Ahkamulloh A, Murniati E, Surahman M. 2013. Keragaman pertumbuhan dan produksi beberapa aksesi jarak pagar (Jatropha curcas L.) di daerah pesisir pantai. Buletin Agrohorti. 1(1):34-44.
Aisyah SI. 2006. Induksi mutagen fisik pada anyelir (Dianthus caryophyllus
Linn.) dan pengujian stabilitas mutannya yang diperbanyak secara vegetatif [Tesis]. Bogor, Pascasarjana Institut Pertanian Bogor.
Apavatjrut P, Anuntalabhochai S, Sirirugsa P, Alisi C. 1999. Molecular markers in the identification of some early flowering Curcuma L. (Zingiberaceae) species. Annals of Botany. 84: 529-534.
Arisanti Y. 2010. Analisis karakter agronomi dan pola pita isozim jarak pagar (Jatopha curcas L.) di daerah beriklim basah [Tesis]. Bogor, Pascasarjana Institut Pertanian Bogor.
Asante IK, Laing EI. 2001. Isozyme variation and genetic diversity at 3 phosphoglucose-isomerase (PGI) [glucose-1-phospate] gen loci in nine Cowpea accessions (Vigna unguiculata L. Walp) from three agroecological zones. West African Journal of Applied Ecology. 2:1-8.
Bakhtiar. 2002. Analisis keragaman genetik gladiol dengan penanda RAPD dan evaluasi ketahanan turunan dari beberapa kombinasi silangan terhadap Fusarium [Tesis]. Bogor, Pascasarjana, Institut Pertanian Bogor.
31
Cahyarini RD. 2004. Identifikasi keragaman genetik beberapa varietas lokal kedelai di Jawa berdasarkan analisis isozim. [Tesis]. Surakarta, Pascasarjana Universitas Sebelas Maret.
Camellia N, Thohirah NA, Abdullah NAP. 2012. Floral biology, flowering behaviour and fruit set development of Jatropha curcas L. in Malaysia.
Journal Tropical Agriculture Science. 35(4): 725-736.
Campos ET, Espinosa MAG, Warbuton ML, Varela AS, Monter AV. 2005. Characterization of mandarin (Citrus sp) using morphological and AFLP markers. Intersciencia. 11: 687-692.
Carvalho CR, Clarindo WR, Praca MM, Araujo FS, Carels N. 2008. Genome size, base composition and karyotipe of Jatropha curcas L., an important biofuel plant. Plant Science. 174:613-617.
Dafni A, Firmage D. 2000. Pollen viability and longevity: practical, ecological and evolutionary implications. Plant Systematics and Evolution. 222: 113-132.
Dahmer N, Wittmanni TS, Dias LADS. 2009. Chromosomes numbers of Jatropha curcas L.: an important agrofuel plant. Crop Breeding and Applied Biotechnology. 9: 386-289.
Delph LF. 2003. Sexual dimorphism in gender plasticity and its consequences for breeding system evolution. Evolution Development. 5:34-39.
Dewatisari WF, Suranto, Setyomo P. 2008. Kenakeragaman beberapa varietas
Sansevieria trifasciata berdasarkan karakter anatomi, isozim dan kandungan saponin. Bioteknologi. 5(2): 56-62.
Doyle JH, Verhoeven RL, Bester C, Wingfield BD, Botha AM. 2002. Germ-furrow morphology and storage conditions determine the degree of viability of Pinus caribea pollen. South African Journal of Botany. 68:457-463. Escudero M, Hipp AL, Luceno M. 2010. Karyotype stability and predictors of
chromosome number variation in sedges: A study carex section spirostachyae (Cyperaceae). Molecular Phylogenetics and Evolution. 57: 353-363.
Fitmawati. 2008. Biosistematika mangga Indonesia. [Disertasi]. Bogor, Pascasarjana Institut Pertanian Bogor.
Foidl N, Foidl G, Sanchez M, Mittelbach M, Hackel S. 1996. Jartopha curcas as a source for production of biofuel in Nicaragua. Bioresource Technology.
58:77-82.
Ghorbel NC, Chakroun M, Elazreg H, Farah NT. 2011. Agronomic evaluation and genetic variation of tunisian tall fescue (Festuca arundinacea Schreb.).
International Journal of Agronomy. 2011: 1-6.
Gianfranco V, Ravalli C, Cremonini R. 2008. The karyotipe as a tool to identify species: Vicia species belonging to Vicia subgenus. Caryologia. 61(3): 300-319.
Hadiati B, Sukmadjaja D. 2002. Keragaman pola pita beberapa aksesi nenas berdasarkan analisis isozim. Jurnal Bioteknologi Pertanian. 7(2): 62-70. Hairmansis A, Aswidinnoor H, Trikoesoemaningtyas, Suwarno. 2005.
Identification of wide compatibility varieties in some tropical japonica rice.
Journal of Agricultural Science. 6(1): 28-31.