Fakultas Matematika dan Ilmu Pengetahuan Alam
Institut Pertanian Bogor
Bogor
2009
ISBN: 978-979-95093-5-2
Bogor, 14 November 2009
Prosiding
Seminar Nasional Sains II
ISBN: 978-979-95093-5-2
Prosiding
Seminar Nasional Sains II
Peningkatan Peran Sains dalam
Pertanian dan Industri
Bogor, 14 November 2009
Fakultas Matematika dan Ilmu Pengetahuan Alam
Institut Pertanian Bogor
Bogor
__________________________________________________________________
Copyright© 2009
Fakultas Matematika dan Ilmu Pengetahuan Alam (FMIPA), Institut Pertanian Bogor (IPB) Prosiding Seminar Nasional Sains: “Peningkatan Peran Sains dalam Pertanian dan Industri” Bogor, 14 November 2009
FMIPA-IPB, Jalan Meranti Kampus IPB Dramaga, Bogor 16680 Telp/Fax: 0251-8625481/8625708
http://fmipa.ipb.ac.id
ix + 553 halaman
KATA PENGANTAR
Sektor pertanian dan sektor industri, khususnya industri yang menopang pertanian,
merupakan tumpuan perekonomian Bangsa Indonesia. Efisiensi dan efektivitas
merupakan dua hal yang harus diperhatikan dalam upaya meningkatkan produktivitas
baik di sektor pertanian maupun industri. Kedua hal ini hanya mungkin dicapai secara
signifikan bila berlandaskan sains dan teknologi yang tepat melalui pemahaman,
pengembangan dan penerapannya yang disesuaikan dengan tuntutan dan tantangan
zaman.
Banyak perguruan tinggi dan lembaga litbang departemen bahkan divisi litbang di
perusahaan terus berupaya untuk meningkatkan produktivitas melalui penelitian dan
pengembangan yang didasarkan pada pemanfaatan dan pengembangan sains dan
teknologi. Seminar Nasional Sains II (2009) ini diharapkan menjadi sarana dan upaya
untuk menjalin komunikasi antar pelaku dan institusi yang terlibat untuk mengoptimumkan
pemanfaatan peran sains dalam pertanian maupun industri.
Seminar ini merupakan rangkaian dari kegiatan Pesta Sains 2009 yang
diselenggarakan oleh FMIPA-IPB pada tanggal 13-15 November 2009. Selain acara
seminar juga diselenggarakan kegiatan Workshop Penulisan Buku Ajar yang diikuti oleh
guru-guru SMA dan dosen.
Sebanyak 60 makalah hasil penelitian dipresentasikan pada empat kelas paralel
yaitu Biosains (1 & 2), Nanosains & Material, serta Penginderaan Jauh, Sensor &
Pemodelan. Selain itu beberapa makalah juga ditampilkan pada sesi Poster.
Makalah-makalah tersebut sebagian besar merupakan isi dari prosiding ini. Seminar dihadiri oleh
peneliti dari balitbang-balitbang terkait dan dosen-dosen perguruan tinggi, mahasiswa
pascasarjana serta guru-guru SMA.
Ucapan terima kasih disampaikan kepada FMIPA-IPB yang telah mendukung penuh
kegiatan Seminar Nasional Sains II ini. Juga kepada Panitia Seminar dan mahasiswa dari
tim Pesta Sains 2009, dan semua pihak yang telah mensukseskan acara seminar ini.
Kami juga sangat berterima kasih kepada semua pemakalah atas kerjasamanya,
sehingga memungkinkan prosiding ini terbit. Semoga prosiding ini bermanfaat bagi semua
pihak.
Bogor, November 2009
Panitia Seminar Nasional Sains II
PANITIA SEMINAR NASIONAL SAINS II
Penanggung Jawab
: Dr. drh Hasim, DEA
(Dekan FMIPA-IPB)
Ketua Pelaksana
: Dr. Kiagus Dahlan
Wakil Ketua Pelaksana
: Dr. Ir Ence Darmo J Supena
Sekretaris
: Dr. Ir Suryani
Bendahara
: Dr. Dyah Iswantini
Pubdok & Promosi
: Dr. Akhiruddin M (Koord.)
Dr. Sri Nurdiati
Faozan, M.Si
Dr. Muhammad Nur Aidi
Acara & Persidangan
: Dr. Miftahuddin (Koordinator)
Mersi Kurniati, M.Si
Makalah & Prosiding
: Ir. Indahwati, M.Si (Koordinator)
Ir. AE Zainal Hasan, M.Si
Perlengkapan & Konsumsi
: Dr. Aris Tjahjoleksono (Koord.)
Mansur
Fitri
Samsudin
DAFTAR ISI
No. PENULIS JUDUL Hal
BIOSAINS 1
1 Mardi Santoso, M. Holil, S. Alfarisi
Pembuatan 4-Formil-2-Metoksifenil Isobutirat dari Daun Cengkeh
2
2 Christiani Tumilisar Effect of Rodent Tuber Extract (Typhonium Flagelliforme (Lodd)BL.) on Cancer Cell Line Proliferation Inhibition
8
3 Samanhudi, Ahmad Yunus, Wangi Satutik
Pengaruh Macam Nutrisi dan Pemberian Ekstrak Buah Pisang terhadap Pertumbuhan Plantlet Anggrek Dendrobium Secara In Vitro
15
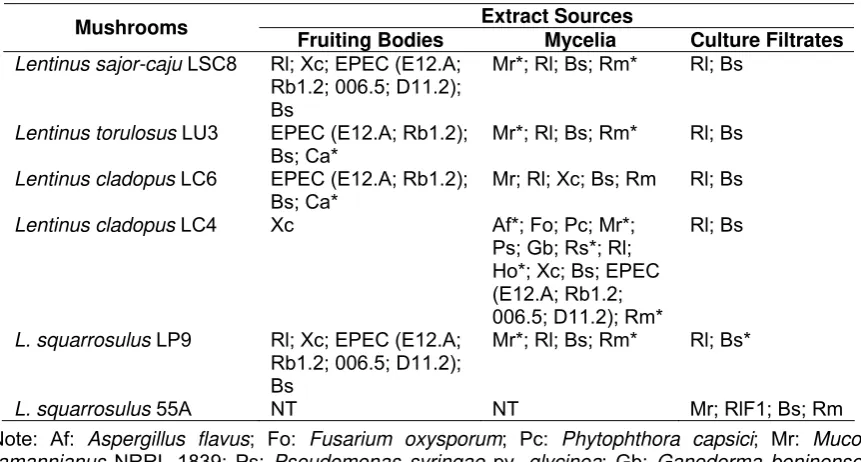
4 Lisdar I. Sudirman Potensi Jamur Pelapuk Kayu Tropis dalam Menghasilkan Senyawa Antimikroba
26
5 It Jamilah, Anja Meryandini, Iman Rusmana, Antonius Suwanto, Nisa R Mubarik
Karakterisasi Protease dan Amilase Bacillus sp. DA 5.2.3 yang Diisolasi dari Tambak Udang
37
6 Dyah Iswantini, Gustini Syabirin dan Yusuf Affandi S
Daya Hambat Ekstrak Air dan Etanol Keladi Tikus (Typhonium flagelliforme) terhadap Enzim Tirosin Kinase Secara In Vitro
47
7 Dyah Iswantini, Gustini Syabirin dan Maya Puspitasari S
Inhibisi Ekstrak Air dan Etanol Sambiloto (Andrographis paniculata [Burm.f.] Nees) terhadap Aktivitas Enzim Tirosin Kinase secara In Vitro
59
8 Dyah Iswantini, Latifah K Darusman dan Dede Yulianto
Inhibisi Xantin Oksidase secara In Vitro oleh Ekstrak Rosela (Hibiscus sabdariffa L.) dan Herba Ciplukan (Physalis angulata)
73
9 Dyah Iswantini, Latifah K Darusman dan Chintya Galuh TW
Potensi Ekstrak Tempuyung (Sonchus arvensis) dan Meniran (Phyllanthus niruri) Sebagai Anti Asam Urat: Aktivitas Inhibisinya terhadap Xantin Oksidase
89
10 Anak Agung Istri
Ratnadewi, Muh.Naqib, I Nyoman Adi Winata, Laode Muh. Dzuhri Abdullah
Hidrolisis Oat-Spelt Xylan oleh Enzim Xilanase serta Deteksi Xilooligosakarida Secara Kromatografi
103
11 Anak Agung Istri Ratnadewi, Muhammad Naqib, Nuri, Zora Olivia
Populasi Bifidobacterium spp. Akibat Suplementasi Roti Tawar Berprebiotik Xilooligosakarida pada Diet Tikus Rattus norvegicus Berkenhout strain WISTAR
113
12 Charlena, Abdu Haris, Karwati
Degradasi Hidrokarbon pada Tanah Tercemar Minyak Bumi dengan Isolat A10 dan D8
124
13 Lucy Arianie, Ahmad Mulyadi, Afghani Jayuska
Pengaruh Pemupukan Urea Termodifikasi Lignin Terhadap Pertumbuhan Sawi
137
14 Gunawan, Tatik Chikmawati, Miftahudin, Dwi Susilaningsih
Mikroalga dari Sumber Air Panas Ciater yang Berpotensi Sebagai Sumber Biodisel
No. PENULIS JUDUL Hal
15 Arinana, Yudi Rismayadi, Noor Farikhah Haneda
Karakterisasi Serangan Kumbang Bubuk Kayu Kering pada Kayu Konstruksi Rumah Tinggal
155
16 Abdul Rauf Pengujian Rumput Tapak Liman (Elephantopus scaber L.) Sebagai Tanaman Penutup Tanah terhadap
Beberapa Sifat Tanah Inceptisol dan Bibit Kelapa Sawit
161
17 Boedi Rachmandan Sata Yoshida Srie Rahayu
Pertumbuhan Kerang Mutiara Air Tawar (Anodonta woodiana, Lea) dengan Tipe Pemeliharaan yang Berbeda
167
NANOSAINS DAN MATERIAL 177
1 Muhammad Ali Zulfikar, Efni Novita
Penurunan Intensitas Warna Air Gambut Menggunakan Cangkang Telur
178
2 S.T. Wahyudi, J.Juansah, E.Mahrani
Karakterisasi Kekuatan Mekanik Membran Telur Ayam Kampung
185
3 Purwantiningsih Sugita, Suminar S. Achmadi, Yuyu Yundhana
Perilaku Disolusi Ketoprofen Tersalut Gel Kitosan-Karboksimetilselulosa (CMC)
192
4 Gerald E Timuda, Akhiruddin M, Irmansyah
Pengaruh Waktu Pemaparan Gelombang Ultrasonik terhadap Komposisi Fase, Ukuran dan Parameter Kisi Kristal dari Nanopartikel TiO2 yang Disintesis
Menggunakan Metode Sonokimia
202
5 Taofik Jasa Lesmana, Akhiruddin M, Irmansyah
Pengaruh Konsentrasi Donor H+ pada Polianilin Terhadap Sel Surya Hibrid ITO/CdS/Klorofil/PANI/ITO
210
6 H. Syafutra, Irzaman, H. Darmasetiawan, H. Hardhienata, F. Huriawati, M. Hikam, P. Arifin
Penumbuhan Film Tipis BST di atas Substrat Si (100) Tipe-p untuk Aplikasi Sensor Cahaya
216
7 Betty Marita Soebrata, Moh. Khotib, Maipa Diapati
Ampas Tebu Sebagai Adsorben Zat Warna Reaktif CIBACRON RED
225
8 Tetty Kemala, Emil Budianto, Bambang Soegiyono
Pembuatan dan Pencirian Polipaduan Poliasamlaktat dan Polikaprolakton Sebagai Bahan Dasar Mikrosfer
237
9 H. A.E. Zainal Hasan, I Made Artika, Vita Rosaline Fahri, Nurmala Sari
Penerapan Teknologi Nanopartikel untuk Sediaan Obat (Antibiotik Berbasis Bahan Alam, Propolis Trigona spp.)
247
10 Nur Aisyah Nuzulia, Akhiruddin Maddu, Kiagus Dahlan
Synthesizing and Characterization of Biphasic Calcium Phosphate Ceramic
257
11 Jajang Juansah, Mersi Kurniati, Kiagus Dahlan dan F. Jannah
Studi Membran Telur Ayam Melalui Pengukuran Listrik 265
12 Akhiruddin Maddu, Nendar Herdianto dan Irmansyah
Studi Fotoelektrokimia Elektroda Nanokristal TiO2 untuk Aplikasi Fotovoltaik
No. PENULIS JUDUL Hal
13 Fifia Zulti, Kiagus Dahlan, Purwantiningsih Sugita
Sintesis dan Karakterisasi Membran untuk Filtrasi Limbah
286
14 M. Kurniati, A.L Kencana, J. Juansah, A Maddu
Perlakuan Sonikasi Terhadap Kitosan: Viskositas dan Bobot Molekul Kitosan
293
PENGINDERAAN JAUH DAN SENSOR 302
1 Suyadi Tropical Deforestation in Bukit Barisan Selatan National Park, Sumatra, Indonesia
303
2 M. Rahmat, Teguh P.N, H. Alatas, Irmansyah
Desain dan Fabrikasi Sensor Real Time berbasis Kristal Fotonik Satu Dimensi untuk Deteksi Konsentrasi
Larutan Gula
318
3 Kris Sunarto Kontribusi Survei dan Pemetaan terhadap Pembangunan Bidang Pertanian
328
4 Ucuk Darusalam, Retno W.P.
Piranti Optik Pengukur Kelimpahan Fitoplankton dengan Metoda Fluoresensi
337
5 Gunady Haryanto, Retno Wigajatri P.
Perancangan Probe Optik Berbasis Fluoresensi untuk Mengukur Konsentrasi Fitoplankton
350
6 Liliana Adia K, Akhirudin Maddu, Irmansyah
Pembuatan Sensor Serat Optik dengan Cladding Dye Methyl Violet untuk Mendeteksi Gas H2S
356
7 Teguh P Negara, H Alatas Sensor Optik Berbasis Kristal Fotonik Satu Dimensi dengan Sensitivitas Terkontrol
364
8 Jessi L Tambunan, Akhiruddin Maddu, Iriani Setyaningsih
Karakteristik Optik dan Elektronik Ekstrak Klorofil Spirulina fusiformis
375
9 Novita G. Pamungkas, Irzaman
Kajian Efisiensi Termal Heating Mantel untuk
Penerapan Penyulingan Minyak Atsiri dari Bahan Serai Dapur
384
PEMODELAN 389
1 Rietje J.M Bokau, Wamiliana
Desain Model Matematika untuk Sistem Produksi Pakan Udang
390
2 Mohammad Masjkur Perbandingan Metode Kuadrat Terkecil Linear dan Nonlinear Marquardt-Levenberg Pendugaan Model Jerapan Fosfor
400
3 Mohammad Masjkur Perbandingan Metode Kuadrat Terkecil Marquardt-Levenberg dan Kemungkinan Maksimum EM
Pendugaan Parameter Model Nonlinear Jerapan Fosfor
414
4 Muhammad Nur Aidi Deteksi Pola Sebaran Titik Spasial Secara Reguler Melalui Penelusuran Fungsi Massa Peluang, Metode Kuadran dan Tetangga Terdekat
425
5 Aji Hamim Wigena Penggunaan Regresi Kuadrat Terkecil Parsial dalam Statistical Downscaling
No. PENULIS JUDUL Hal
6 Endar H. Nugrahani Model Dinamika Sistem Ekonomi Berdasarkan Akumulasi Modal
440
POSTER 450
1 Ninik Setyowati dan Nurul Sumiasri
Variasi Jenis Tanaman dalam Upaya Peningkatan Produktivitas Lahan Pekarangan di Cibinong
451
2 Mukhtar Effendi, Sehah Peningkatan Kepekaan Sistem Deteksi Spektrometer Fotoakustik Gas Lacakan dengan Cara Optimasi Daya Laser CO2 yang Digunakan
460
3 Destario Metusala Studi Waktu Aplikasi dan Dosis Herbisida Campuran Atrazine dan Mesotrione Terhadap Pertumbuhan Gulma pada Pertanaman Jagung
470
4 Agung Sri Darmayanti, Destario Metusala
Pengaruh Media Tanam Terhadap Pertumbuhan Vegetatif Cempaka (Michelia Champaca)
478
5 Samanhudi, Praswanto, dan Edi Dwiyono
Induksi Kalus Tanaman Mahkota Dewa (Phaleria macrocarpa) dengan Perlakuan Kondisi Gelap dan 2,4-D
485
6 Sarjiya Antonius, Dwi Agustyani, Nurlaili, Ronald B. P. Simbolon
Sifat Biologi dan Kimia Tanah pada Beberapa Komoditas Pertanian di Malinau-Kaltim
495
7 Rini Riffiani Pengujian Enzim Peroksidase pada Kultur Suspensi Sel Raphanus sativus yang Diperkaya dengan Hormon Pertumbuhan
501
8 Nurul Sumiasri, Yani Cahyani, Dody Priadi
Pengaruh berbagai Media dan Pematahan Dormansi Biji terhadap Pertumbuhan Biji Jarak Pagar (Jatropha curcas L)
510
9 Sudarmono Pendekatan Konservasi Berdasarkan Variasi Genetika Populasi pada Tumbuhan: Suatu Kasus pada Salvia Sp. (LAMIACEAE)
521
10 Sudarmono, Sumanto Variasi Genetika pada Populasi Scutellaria Sp. (Lamiaceae) di Gunung Slamet, Jawa Tengah
529
11 Sri Hartin Rahayu Pengaruh Rhizobium dan Puminal Terhadap Pertumbuhan dan Hasil Kacang Hijau dalam
Pengembangan Usahatani di Lahan Bekas Galian Emas Jampang-Sukabumi
537
12 Hartutiningsih-M. Siregar, R. S. Purwantoro,
Sudarmono, A. Agusta
Pengungkapan Potensi Obat pada Tiga Jenis Begonia Terpilih (Begonia muricata Blume, B. multangula Blume, B. ”Bacem Kebo”.) Melalui Uji Anti Bakteri Escherichia coli dan Staphylococcus aureus Secara In Vitro
543
13 Eko Murniyanto Keragaan Daun Kimpul (xanthosoma sagittifolium l.schoot) yang Terpapar pada Penyinaran Matahari
B
PEMBUATAN 4-FORMIL-2-METOKSIFENIL ISOBUTIRAT Biosains
PEMBUATAN 4-FORMIL-2-METOKSIFENIL ISOBUTIRAT
DARI DAUN CENGKEH
Mardi Santoso*, Mohammad Holil, Salman Alfarisi
Jurusan Kimia, Fakultas MIPA, Institut Teknologi Sepuluh Nopember Kampus ITS Sukolilo Surabaya 60111
Telp. (031) 5924930, Fax. (031) 5945813 [email protected]
Abstrak
Distilasi daun cengkeh (Syzigium aromaticum L. Mp) segar dari Pasuruan diperoleh minyak daun cengkeh dengan rendemen 1,5%. Eugenol sebagai komponen terbesar (80%) dalam minyak daun cengkeh dipisahkan dengan menerapkan prinsip reaksi asam basa dan perbedaan kelarutan. Oksidasi eugenol dalam campuran nitrobenzena, kalium hidroksida, dimetil sulfoksida, dan akuades diperoleh vanilin dengan rendemen 19%. Adisi eliminasi vanilin dalam piridina dengan butiril klorida diperoleh 4-formil-2-metoksifenil isobutirat dengan rendemen 91%. Analisis sensori oleh panelis terlatih menunjukkan bahwa transformasi vanilin menjadi 4-formil-2-metoksifenil isobutirat menyebabkan terjadinya perubahan karakteristik sensori yang signifikan.
Kata kunci: daun cengkeh, eugenol, sensori, 4-formil-2-metoksifenil isobutirat
1. PENDAHULUAN
Cengkeh (Syzigium aromaticum L. mp) merupakan tanaman asli Indonesia yang
kemungkinan berasal dari Maluku. Penanaman cengkeh dilakukan di Maluku, Jawa,
Sulawesi, Irian Jaya (Sastromahmidjoyo, 2004). Minyak cengkeh diperoleh dari bunga
maupun daun cengkeh melaui proses distilasi. Minyak cengkeh dengan rasa dan aroma
khas dimanfaatkan sebagai stimulan, anestetik, karminatif, antiseptik, dan antipasmodik
(Nurdjannah, 2004). Minyak daun cengkeh mengandung eugenol sebagai komponen
utama, disertai senyawa-senyawa non fenolat seperti β-kariofilena, α-kubebena, α
-kopaena, humelena, δ-kadinena (Wahidi, 2001; Sastrohamidjojo, 2004). Kadar eugenol
dari minyak daun cengkeh yang tumbuh di India adalah sebesar 64-81% (Lawrence,
1994; Lawrence, 1997; Verheij dan Coronel, 1997), minyak dari Lumajang mengandung
92% (Wahidi, 2001), sedangkan yang di pasaran berkisar 72% (Sastrohamidjojo, 2004).
Eugenol telah ditransformasi menjadi isoeugenol, vanilin, metil eugenol, metil
eugenol bromida, metil eugenol format, 1-(3,4-metoksifenil)-2-propanol, 1-(3,4-metoksi
fenil)-2-propanon, amfetamin (Lampman et al, 1976; Pybus dan Sell, 1999; dan
Sastrohamidjojo, 2004). 4-Formil-2-metoksifenil isobutirat merupakan senyawa komersial
PEMBUATAN 4-FORMIL-2-METOKSIFENIL ISOBUTIRAT Biosains
flavourchocominty (Kraft et al, 2000). Pembuatan 4-formil-2-metoksifenil isobutirat sejauh
pengetahuan penulis belum pernah diungkap. Paper ini bermaksud melaporkan
pemanfaatan daun cengkeh sebagai bahan dasar pembuatan 4-formil-2-metoksifenil
isobutirat, berikut uji sensori 4-formil-2-metoksifenil isobutirat dibandingkan terhadap
vanillin.
2. BAHAN DAN METODA
Bahan-bahan yang digunakan adalah daun cengkeh (Syzigium aromaticum L. mp)
segar dari Pasuruan, dietil eter, natrium hidroksida, kalium hidroksida, asam klorida,
nitrobenzena, dimetil sulfoksida, isobutiril klorida, sikloheksana, natrium bisulfit,
magnesium sulfat anhidrat, piridina, akuades.
2.1. Distilasi Daun Cengkeh (Syzigium aromaticum L. mp)
Daun cengkeh (Syzigium aromaticum L. mp) segar sebanyak 100 gram didistilasi
menggunakan peralatan distilasi Clevenger yang dimodifikasi selama 6 jam. Minyak daun
cengkeh dalam dietil eter yang diperoleh selanjutnya dipisahkan, dikeringkan, dan
diuapkan sehingga diperoleh minyak daun cengkeh (1,5 gram; 1,5%). Minyak daun
cengkeh selanjutnya dianalisis dengan kromatografi gas spektrometer massa.
2.3. Pemisahan dan Pemurnian Eugenol
Pemisahan dan pemurnian eugenol dilakukan dengan mengadaptasi prosedur yang
dilaporkan Sastrohamidjojo (2004). Campuran minyak daun cengkeh dan larutan natrium
hidroksida 0,17 M pada pH 11 diaduk pada suhu kamar selama 30 menit. Campuran
kemudian diekstrak dengan dietil eter. Fasa air selanjutnya dipisahkan, diasamkan, dan
diekstrak reaksikan dengan dietil eter. Ekstrak selanjutnya dikeringkan, diuapkan pada
tekanan rendah sehingga diperoleh eugenol yang selanjutnya dianalisis dengan
kromatografi gas spektrometer massa. Spektrum massa (EI): (m/z) 166 (M+2, 5%), 165
(M+1, 10), 164 (M, 100), 149 (30), 137 (20), 121 (10), 103 (20), 91 (20), 77 (20).
2.4. Oksidasi Eugenol
Oksidasi eugenol dilaksanakan dengan mengadaptasi prosedur yang dilaporkan
Lampman et al. (1976), menggunakan campuran eugenol (2,14 gram; 0,013 mol), kalium
hidroksida (4,75 gram; 0,084 mol), dimetil sulfoksida (20 ml; 0,282 mol), akuades (10 ml),
nitrobenzena (10 ml; 0,097 mol). Campuran hasil reaksi selanjutnya diasamkan, dan
diekstrak dengan dietil eter. Ekstrak kemudian dikocok dengan larutan natrium bisulfit.
Fasa air selanjutnya dipisahkan, diasamkan, dan diekstrak dengan dietil eter. Ekstrak
kemudian dikeringkan, diuapkan pada tekanan rendah, direkristalisasi menggunakan
PEMBUATAN 4-FORMIL-2-METOKSIFENIL ISOBUTIRAT Biosains
titik leleh 79-80 oC. 1H NMR (CDCl
3): δ 3,94 (s, 3H), 6,51 (sl, 1H), 6,98-7,46 (m, 3H), 9,81
(s, 1H). Spektrum massa (EI): (m/z) 153 (M+1, 10%), 152 (M, 95), 151 (100), 137 (5), 123
(15), 109 (20), 93 (5), 81 (20), 77 (5).
2.5. Sintesis 4-Formil-2-metoksifenil Isobutirat
Sintesis 4-formil-2-metoksifenil isobutirat dilakukan dengan mengadaptasi prosedur
sintesis 2-benzoiloksiasetofenon dari reaksi 2-hidroksiasetofenon dengan benzoil klorida
yang dilaporkan Harwood dan Moody (1990) menggunakan vanilin (1,51 gram; 0,010
mol), piridin (2,00 ml; 0,025 mol), isobutiril klorida (1,50 ml; 0,015 mol). Hasil sintesis
diekstrak dengan dietil eter. Ekstrak yang diperoleh selanjutnya dicuci dengan akuades,
dikeringkan dengan magnesium sulfat anhidrat, diuapkan pada tekanan rendah sehingga
diperoleh 4-formil-2-metoksifenil isobutirat sebagai cairan jernih kekuningan (2,14 gram;
91%). 1H NMR (CDCl
3): δ 1,28 (d, 6H), 2,78-2,94 (m, 1H), 3,89 (s, 3H), 7,14-7,53 (m, 3H),
9,94 (s, 1H). 13C NMR (CDCl3): δ 18,89; 33,96; 55,98; 110,78; 123,18; 124,44; 134,92;
145,10; 151,84; 174,28; 190,77. Spektrum massa (EI): (m/z) 223 (M+1, 5%), 222 (M, 15),
153 (20), 152 (100), 151 (65), 137 (5), 123 (10), 109 (10), 71 (45).
3. HASIL DAN PEMBAHASAN
3.1. Distilasi Daun Cengkeh (Syzigium aromaticum L. mp)
Distilasi daun cengkeh (Syzigium aromaticum L. mp) segar sebanyak 100 gram
didistilasi diperoleh minyak daun cengkeh sebanyak 1,5 gram atau dengan rendemen
1,5%. Minyak daun cengkeh yang diperoleh jernih kekuningan dengan bau seperti sampel
daun. Analisis dengan kromatografi gas spektrometer massa menunjukkan adanya 13
komponen, dengan eugenol sebagai komponen terbesar (80%).
3.2. Pemisahan dan Pemurnian Eugenol
Pemisahan dan pemurnian eugenol dilakukan dengan menerapkan prinsip reaksi
asam basa dan perbedaan kelarutan. Pemisahan diawali dengan penambahan larutan
natrium hidoksida ke dalam minyak daun cengkeh untuk mengubah eugenol menjadi
natrium eugenolat yang larut dalam air, sehingga terpisah komponen-komponen lain yang
larut dalam dietil eter. Fasa air selanjutnya diasamkan untuk mendapatkan eugenol
kembali yang kemudian dilarutkan dalam dietil eter. Larutan dietil eter selanjutnya
dikeringkan, diuapkan pada tekanan rendah sehingga diperoleh eugenol sebagai cairan
jernih. Analisis dengan kromatografi gas spektrometer massa memberikan kromatogram
yang menunjukkan adanya pucak tunggal dengan spektrum massa yang menunjukkan
puncak ion molekul yang sekaligus merupakan puncak dasar pada m/z 164 yang sesuai
PEMBUATAN 4-FORMIL-2-METOKSIFENIL ISOBUTIRAT Biosains
hasil pelepasan radikal metil dari ion molekul, dan puncak dengan m/z 137 merupakan
puncak kation hasil pelepasan radikal vinil.
3.3. Oksidasi Eugenol
Oksidasi eugenol dilaksanakan dengan merefluks dalam campuran kalium
hidroksida, nitrobenzena, dimetil sulfoksida dan akuades. Monitoring reaksi menggunakan
kromatografi gas spektrometer massa menunjukkan bahwa pada vanilin terbentuk saat
reaksi berjalan selama 3 jam, dan reaksi berjalan tuntas saat reaksi berlangsung selama
48 jam. Hasil oksidasi selanjutnya didinginkan sehingga mencapai suhu kamar,
diasamkan, dan diekstrak dengan dietil eter. Ekstrak selanjutnya direaksikan dengan
larutan natrium bisulfit. Fasa air selanjutnya dipisahkan, diasamkan, dan diekstrak dengan
dietil eter. Ekstrak selanjutnya dikeringkan, diuapkan pada tekanan rendah sehingga
diperoleh vanilin kotor yang selanjutnya dimurnikan dengan rekristalisasi menggunakan
sikloheksana. Vanilin murni yang diperoleh berbentuk jarum bening dengan titik leleh
79-80 oC dengan rendemen sebesar 19%. Analisis vanillin hasil sintesis dengan kromatografi
gas spektrometer massa memberikan spektrum massa yang menunjukkan ion molekul
pada m/z 152 yang sesuai dengan massa relatif vanilin. Spektrum juga menunjukkan
pucan dasar pada m/z 151 yang merupakan puncak dari fragmen hasil pelepasan atom
hidrogen dari ion molekul khas gugus aldehid, kation ini selanjutnya melepaskan CO
menghasilkan kation dengan m/z 123. Puncak pada m/z 137 merupakan puncak fragmen
hasil pelepasan radikal metil dari ion molekul yang selanjutnya melepaskan CO
menghasilkan puncak dengan m/z 109. Analisis vanilin hasil sintesis dengan spektrometer
NMR (dalam CDCl3) memberikan spektrum 1H NMR yang menunjukkan adanya empat
signal. Signal singlet dengan pergeseran kimia pada 3,94 ppm merupakan signal dari
ketiga proton gugus metoksi; signal singlet lebar pada pergeseran kimia 6,51 ppm
merupakan signal dari proton hidroksi; signal multiplet pada pergeseran kimia 6,98-7,46
ppm merupakan signal dari ketiga proton aromatik; dan signal singlet pada pergeseran
kimia 9,81 ppm merupakan signal dari proton gugus aldehid.
3.4. Sintesis 4-Formil-2-metoksifenil Isobutirat
4-Formil-2-metoksifenil isobutirat disintesis dari reaksi vanillin dalam piridin dengan
isobutiril klorida pada suhu kamar. Reaksi yang berjalan dipantau dengan kromatografi
gas spektrometer massa, dan reaksi berjalan tuntas setelah berlangsung selama 5 jam.
Hasil reaksi kemudian diasamkan, dan diekstrak dengan dietil eter. Ekstrak selanjutnya
dikeringkan dan diuapkan pada tekanan rendah sehingga diperoleh
4-formil-2-metoksifenil isobutirat berupa cairan jernih kekuningan dengan rendemen sebesar 91%.
PEMBUATAN 4-FORMIL-2-METOKSIFENIL ISOBUTIRAT Biosains
spektrum massa yang menunjukkan ion molekul pada m/z 222, sesuai dengan massa
relatif 4-formil-2-metoksifenil isobutirat. Puncak dasar pada m/z 152 merupakan puncak
dari radikal kation vanilin, yang selanjutnya melepaskan atom hidrogen khas aldehid
menghasilkan kation dengan m/z 151. Fragmen dengan m/z 152 juga melepaskan radikal
metil sehingga terbentuk kation dengan m/z 137, kation ini selanjutnya melepaskan CO
menghasilkan fragmen dengan m/z 109 yang kemudian melepaskan CO lebih lanjut
menghasilkan fragmen dengan m/z 81. Radikal kation vanilin dengan m/z 152 juga
melepaskan gugus aldehid sehingga terbentuk kation dengan m/z 123. Puncak pada m/z
71 merupakan puncak dari kation hasil pelepasan radikal vanilil.
Analisis lebih lanjut dengan spektrometer NMR memperkuat kesimpulan hasil
sintesis sebagai 4-formil-2-metoksifenil isobutirat. Spektrum 1H NMR (dalam CDCl3) yang
menunjukkan adanya lima signal. Signal doublet pada pergeseran kimia 1,28 ppm
merupakan signal dari keenam proton dari dua gugus metil; signal multiplet pada
pergeseran kimia 2,78-2,94 ppm merupakan signal dari proton gugus metin yang
dikopling oleh proton-proton kedua gugus metil tetangga; signal singlet pergeseran kimia
pada 3,89 ppm merupakan signal dari proton-proton gugus metoksi. Ketiga proton
aromatik memberikan signal multiplet pada pergeseran kimia 7,14-7,53 ppm; dan proton
gugus aldehid memberikan signal singlet pada pergeseran kimia 9,94 ppm. Spektrum 13C
NMR menunjukkan adanya sebelas jenis karbon sebagaimana dijumpai pada struktur
4-formil-2-metoksifenil isobutirat. Kedua karbon metil memberikan signal pada pergeseran
kimia pada 18,89 ppm, sedangkan karbon metin menunjukkan signal pada pergeseran
kimia 33,96 ppm. Karbon metoksi memberikan pada pergeseran kimia 55,98 ppm; kedua
karbon karbonil menunjukkan pergeseran kimia pada 174,28 ppm untuk karbon karbonil
ester dan pada 190,77 ppm untuk karbon gugus aldehid. Keenam karbon aromatik
selanjutnya memberikan signal pada pergeseran kimia 110,78; 123,18; 124,44; 134,92;
145,10; dan 151,84 ppm.
Uji sensori vanilin dan 4-formil-2-metoksifenil isobutirat oleh lima panelis terlatih
menunjukkan bahwa open/stick aroma 4-formil-2-metoksifenil isobutirat hampir sama
dengan vanillin, sedangkan aroma vanilli 4-formil-2-metoksi isobutirat sedikit lebih rendah
dari pada vanilli. Atribut-atribut lain 4-formil-2-metoksi isobutirat secara umum mengalami
peningkatan, sebagai contoh fruity aroma 4-formil-2-metoksi isobutirat lebih tiga kali lipat
dibandingkan vanillin. Aroma coklat 4-formil-2-metoksi isobutirat lima kali lebih lipat
dibandingkan vanillin, fresh aroma hampir dua kali lipat, dan penerimaan lebih dua kali
PEMBUATAN 4-FORMIL-2-METOKSIFENIL ISOBUTIRAT Biosains
4. KESIMPULAN DAN PROSPEK
Daun cengkeh telah dimanfaatkan sebagai bahan dasar pembuatan
4-formil-2-metoksifenil isobutirat. Reaksi vanillin dengan berbagai asil halida dan anhidrida asam
tengah dilaksanakan untuk mendapatkan 4-formil-2-metoksifenil isobutirat sehingga
karakter sensorinya dapat dioptimalkan.
UCAPAN TERIMA KASIH
Penulis menyampaikan terima kasih kepada Ir. Trisnawati Trisnajuwana atas
bantuan analisis menggunakan kromatografi gas spektrometer massa dan uji sensori, Dr.
Marcellino Rudyanto, MS. atas bantuannya dalam analisis NMR, Kepala Kebun Raya
Purwodadi atas bantuan sampel berikut determinasinya. DP2M DIKTI atas hibah
penelitian yang diberikan.
DAFTAR PUSTAKA
Harwood, M.L., dan Moody, J.C., (1990), Experimental Organic Chemistry, 1st edition, Blackweel Scientific Publications, London.
Kraft, P., Bajgrowicz, J.A., Denis, C., dan Frater, G., (2000), Angew, Chem. Int. Ed., Vol. 39, 2980-3010.
Lampman, G. M., Andrews, J., Bratz, W., Hanssen, O., Kelley, K., dan Ridgeway, A., (1976), J. Chem. Educ., Vol. 54, No. 12, 776-778.
Lawrence, B.M., (1994), Perfurmer & Flavorist, Vol. 19, 92-94.
Lawrence, B.M., (19974), Perfurmer & Flavorist, Vol. 22, 83-84.
Nurdjannah, N., (2004), Perspektif, Vol. 3, No. 2, 61-70.
Pybus, D., dan Sell, C., (1999), The Chemistry of Fragrances, RSC Paperbacks, Quest International, Ashford, Kent, UK.
Sastrohamidjojo, H., (2004), Kimia Minyak Atsiri, Edisi pertama, Gadjah Mada University Press, Yogyakarta.
Pengaruh Ekstrak Umbi Keladi Tikus Biosains
Pengaruh Ekstrak Umbi Keladi Tikus (
Typhonium flagelliforme
(Lodd.)BL.)
Terhadap Penghambatan Proliferasi
Cell Line
Kanker
ChristianiTumilisar
Jurusan Biologi FMIPA Universitas Negeri Jakarta Jl. Pemuda 10, Jakarta Timur 13220, fax: 021.4894909
email: christiani_tumilisar @yahoo.com
Abstract
The research’s purpose is to know the effect of rodent tuber extract (Typhonium flagelliforme (Lodd.)Bl.) on K-562, A-132 and HeLa cancer cell line proliferation inhibition. The research used experiment method with block random design. There were 6 treatments with 3 replications each. The independent variable was rodent tuber extract concentration, whereas the
dependent variable was Inhibition Index (IP/Indeks Penghambatan) of cancer
cell line. Data were collected using MTT method. Statistical analysis used was one-way anova F test and followed by LSD test. The result showed that the
extract of rodent tuber affected the K-562 and HeLa cell line proliferation
inhibition, whereas on the separate research, the tuber extract didn’t
significantly affect the A-132 and K-562 cell line proliferation inhibition at the
same concentration.
Key words: Thyphonium flagelliforme (Lodd.)Bl., cell line, proliferation.
1. PENDAHULUAN
Keladi tikus (Typhonium flagelliforme (Lodd)BL.) termasuk familia Aracea.
Tanaman ini banyak dijumpai tumbuh liar di bawah pohon rindang atau di pinggir sawah,
bahkan ada yang menyebutnya sebagai gulma. Ciri tanaman ini mempunyai pangkal
daun lebar dan bertoreh, sedangkan ujung daun meruncing. Bunganya berwarna merah
gelap dan mempunyai bagian yang memanjang menyerupai ekor tikus. Umbinya
berbentuk bulat lonjong berwarna putih. Tanaman ini diindikasikan sebagai salah satu
tanaman obat yang dapat mengobati penyakit kanker.
Kanker merupakan salah satu penyakit degeneratif yang mendapat perhatian
besar dalam ilmu kedokteran. Sebagian besar penderitanya berakhir dengan kematian.
Penyakit ini dapat menyerang semua bagian organ tubuh, seperti paru-paru, kulit, prostat,
serviks, darah, payudara dan lainnya. Kanker menjadi momok bagi sebagian besar
masyarakat, karena pengobatan terhadap penyakit ini memerlukan biaya medis yang
cukup besar. Oleh karena itu perlu dicarikan alternatif pengobatan yang dapat
meringankan beban penderita dan keluarganya.
Berdasarkan alasan di atas, tanaman keladi tikus yang diindikasikan berkhasiat
sebagai tanaman obat dan dapat mengobati penyakit kanker dapat dimanfaatkan dan
Pengaruh Ekstrak Umbi Keladi Tikus Biosains
aktivitas penghambatan proliferasi sel kanker dari tanaman keladi tikus. Pada penelitian
ini umbi tanaman menjadi bahan utama yang diujikan pada 3 macam cell line kanker
yaitu: K-562 (leukemia), A-132 (kulit) dan Hela (serviks). Adapun tujuan dari penelitian ini
adalah mengetahui efektivitas umbi keladi tikus terhadap 3 macam cell line kanker
tersebut.
2. BAHAN DAN METODE
Umbi keladi tikus umur 4 bulan diekstraksi dengan menggunakan teknik maserasi.
Umbi dicuci bersih dan dikeringanginkan selama 1 minggu, kemudian umbi diblender
sampai berbentuk serbuk. Sebanyak 25 gram serbuk umbi ditambahkan dengan 200 ml
etanol 96%, lalu ditutup dan dibiarkan selama 3 hari. Selanjutnya ekstrak disaring dengan
kertas whatman no. 42. Ampas umbi diberi etanal 50 ml, diaduk dan didiamkan kembali
selama 3 hari, diulang sebanyak 3 kali. Evaporasi dilakukan pada suhu 600 C.
Uji fitokimia ekstrak keladi tikus menggunakan metode Liberman, meliputi uji
triterpenoid dimana ekstrak ditambah 2-3 tetes anhidrat asam asetat dan 1-2 tetes H2SO4
pekat. Penanda berupa perubahan warna merah menjadi ungu, sedangkan adanya
steroid ditandai dengan terbentuknya warna hijau sampai biru. Untuk uji soponin, ekstrak
dalam tabung reaksi dikocok sampai terbentuk busa. Sedangkan untuk uji kuinon, ekstrak
ditambah 10 ml dietil eter lalu dikocok sampai terbentuk warna kuning atau merah.
Kemudian tambahkan 10 tetes NaOH 5% sampai warna ekstrak hilang, lalu tambahkan
HCl encer. Jika warna ekstrak muncul kembali menunjukkan adanya kuinon.
Pembuatan larutan ekstrak keladi tikus, disesuaikan dengan konsumsi keladi tikus
oleh manusia per hari yang masuk ke dalam 6000 ml darah dimana konsentrasi tersebut
setara dengan jumlah ekstrak yang dimasukkan ke dalam 150 µl larutan kultur sel (C2 =
7,08x10-5 g/ml). Sedangkan konsentrasi lain C1 setara dengan setengah kali konsumsi
keladi tikus, C3 setara dengan dua kali konsumsi keladi tikus, C4 setara dengan empat kali
konsumsi keladi tikus dan C5 setara dengan delapan kali konsumsi keladi tikus.
Penentuan konsentrasi dilakukan dengan melarutkan ekstrak ke dalam RPMI 20%.
Larutan ekstrak yang sudah terbentuk disterilisasi dengan filter 0,22 µm.
Untuk medium stok, bubuk RPMI-1640 sebanyak 8,1 gram dilarutkan dalam
akuabides, sehingga diperoleh 500 ml larutan RPMI-1640. Kemudian ditambahkan 1
gram NaHCO3, dan 1% Penicilin Streptomicin. Medium dihomogenkan dan pH diatur
antara 7,2 -7,4. Kemudian medium disterilisasi dengan menggunakan filter 0,22 µm.
Pemeliharaan dan kultur sel. Cell line dalam keadaan beku diinkubasi pada suhu
370 C, dan kemudian dipindahkan ke dalam tabung sentrifugasi yang berisi 3 ml media
Pengaruh Ekstrak Umbi Keladi Tikus Biosains
dan pellet ditambahkan media stok kemudian disentrifus kembali selama 5 menit dengan
kecepatan 1500 rpm. Setelah itu pellet ditambah 5 ml media dan dihomogenkan.
Suspensi sel dipeindahkan kedalam flask dan diinkubasi pada inkubator dengan 5% CO2
pada suhu 370 C.
Observasi dilakukan setiap hari di bawah mikroskop untuk melihat bagaimana
pertumbuhan dan keadaan sel (ada tidaknya kontaminan). Penggantian media dilakukan
setiap 3 hari sekali, atau bila terjadi kontaminasi. Suspensi sel diambil dari flask dengan
menggunakan pipet dan dimasukkan ke dalam tabung sentrifus kemudian disentrifugasi
selama 5 menit dengan kecepatan 1500 rpm. Supernatan dibuang dan pellet
ditambahkan 5 ml medium. Kemudian suspensi dipindahkan dalam flask dan diinkubasi
kembali. Jumlah sel yang digunakan sebanyak Y x 104 sel/well dalam medium RPMI-1640
steril. Pengujian dilakukan dengan menambahkan 100 µl ekstrak dalam 50 µl suspensi
sel.
Penghitungan IP proliferasi sel dengan metode MTT. Metode ini merupakan suatu
metode kuantitatif yang dilakukan dengan menggunakan prinsip colorimetric. Bahan
utama yang dipakai adalah 3-(4,5-Dimethylthiazol-2-yl)-2,5-Diphenyl Tetrazolium Bromida
(MTT). Prinsip kerja dari metode ini adalah mengukur panjang gelombang warna
formazan yang terbentuk dengan menggunakan Spektrofotometric Elisa Plate Reader
sebanding dengan jumlah sel yang hidup (Freshney, 1992). Untuk pengukuran dengan
metode MTT, 6 jam sebelum masa inkubasi berakhir, kultur ditambahkan dengan 10 µl
larutan MTT, 0,5% dan diinkubasi pada suhu 370 C dengan CO2. Setelah masa inkubasi
berakhir, ditambahkan 100 µl HCl-Isopropanol 0,04 N pada setiap sumur. Kemudian nilai
absorbansi masing-masing sumur dibaca dengan Spektrofotometric Elisa Plate Reader
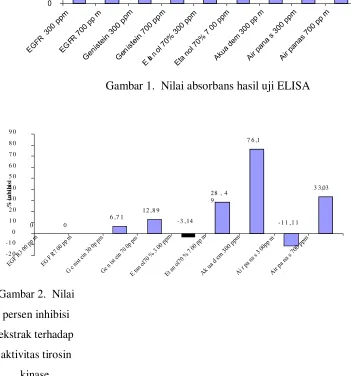
pada panjang gelombang 540 nm.
Hasil absorbansi sampel yang terbaca dibandingkan dengan absorbansi kontrol
sehingga didapatkan nilai indeks penghambatan. Nilai Indeks Penghambatan (IP) dan
Persentase Indeks Penghambatan, dihitung dengan rumus sebagai berikut:
OD (perlakuan)
IP = 1 - ; % IP = IP x 100% OD (kontrol)
3. HASIL DAN PEMBAHASAN
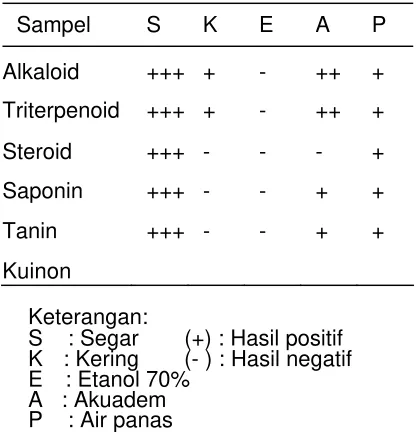
Hasil uji fitokimia ekstrak umbi keladi tikus dapat dilihat pada Tabel 1. Golongan
senyawa yang terkandung pada ekstrak umbi keladi tikus adalah saponin, steroid dan
triterpenoid. Menurut Indiranti (2002), ekstrak umbi keladi tikus mengandung senyawa
golongan triterpenoid dan steroid. Penelitian lain menyatakan bahwa serbuk daun keladi
Pengaruh Ekstrak Umbi Keladi Tikus Biosains
mengandung saponin, steroid dan triterpenoid (Handoko, 2005). Hasil ini sesuai dengan
uji fitokimia ekstrak umbi keladi tikus pada penelitian ini.
Tabel 1. Hasil Uji fitokimia ekstrak umbi keladi tikus.
Golongan Senyawa Hasil Uji Fitokimia
Saponin + Kuinon - Steroid + Triterpenoid +
Rerata Persentase Indeks Penghambatan (IP) proliferasi pada 3 macam cell line
dapat dilihat pada Tabel 2.
Tabel 2. Rerata persentase ip proliferasi pada 3 macam cell line.
IP (%) Konsentrasi Ekstrak
(g/ml) K-562 A-132 HeLa
Kontrol 0 0 0
3,54 x 10-5 0,03 0,21 0,28
7,08 x 10-5 0,04 0,23 0,29
14,16 x 10-5 0,25 0,06 0,37
28,32 x 10-5 0,16 0.01 0,35
56,64 x 10-5 0,32 0.02 0,39
Tabel 2 menunjukkan bahwa ekstrak umbi keladi tikus pada konsentrasi 56,64 x 10-5 g/ml
memiliki rerata persentase IP tertinggi terhadap cell line K-562. Namun pada konsentrasi
28,32 x 10-5 g/ml, rerata persentase IP justru menunjukkan nilai lebih kecil dibandingkan
pada konsentrasi 14,16 x 10-5. Mungkin hal ini disebabkan kurang homogennya jumlah
cell line K-562 pada saat dimasukkan dalam sumur cawan Elisa.
Hal serupa juga terjadi pada rerata persentase IP pada cell line A-132, dimana
pada konsentrasi 3,54 x 10-5 g/ml dan 7,08 x 10-5g/ml menunjukkan rerata persentase IP
yang lebih tinggi, mencapai nilai 0,21 dan 0,23, dibandingkan dengan pada konsentrasi ≥
14,16 x 10-5 g/ml. Hasil analisis varian satu arah IP pada cell line K-562 dapat dilihat pada
Tabel 3.
Tabel 3. Anava Satu Arah Indeks Penghambatan (IP) Proliferasi Cell Line K-562.
Ftabel
Sumber Variasi dK JK RJK Fhitung
5% 1%
Perlakuan 5 0,26 0,05 1,25 ts 3,11 5,06
Galat 12 0,49 0,04 --- --- ---
Total 17 0,75 --- --- --- ---
Dari hasil anava satu arah di atas, diperoleh Fhitung (= 1,25) < Ftabel untuk α = 0,05 (=3,11)
Pengaruh Ekstrak Umbi Keladi Tikus Biosains
secara signifikan terhadap penghambatan proliferasi cell line K-562. Sedangkan hasil
Anava satu arah IP cellline A-132 dapat dilihat pada Tabel 4.
Tabel 4. Anava satu arah indeks penghambatan (ip) proliferasi cell line a-132.
Ftabel
Sumber Variasi dK JK RJK Fhitung
5% 1%
Perlakuan 5 0,18 0,04 2,00 ts 3,11 5,06
Galat 12 0,24 0,02
Total 17 0,42
Dari hasil anava satu arah di atas, diperoleh Fhitung (= 2,00) < Ftabel untuk α = 0,05 (=3,11)
dan untuk α = 0,01 (= 5,06). Ini berarti bahwa ekstrak umbi keladi tikus tidak berpengaruh
secara signifikan terhadap penghambatan proliferasi cell line A-132. Namun demikian
baik cell line K-562 dan cell line A-132, menghasilkan rerata persentase IP yang lebih
besar dari kontrol, artinya terjadi penghambatan proliferasi terhadap cell line K-562 dan
cell line A-132, walaupun penghambatan tersebut tidak signifikan (lihat Tabel 2).
Hal yang berbeda tampak pada rerata persentase IP pada cell line HeLa, yang
menunjukkan kecenderungan semakin meningkat konsentrasi ekstrak umbi keladi tikus
maka semakin tinggi nilai rerata persentase IP (lihat Tabel 2). Hasil analisis varian satu
arah IP cell line HeLa dapat dilihat pada Tabel 5.
Tabel 5. Anava Satu Arah Indeks Penghambatan (IP) Proliferasi Cell Line HeLa.
Ftabel
Sumber Variasi dK JK RJK Fhitung
5% 1%
Perlakuan 5 0,3101 0,0620 9,9 3,11 5,06
Galat 12 0,0751 0,063 --- --- ---
Total 17 0,3852 --- --- --- ---
Berdasarkan hasil anava di atas, diperoleh Fhitung (= 9,9) > Ftabel untuk α = 0,05 (=3,11) dan
untuk α = 0,01 (= 5,06). Ini berarti bahwa ekstrak umbi keladi tikus berpengaruh sangat
signifikan terhadap penghambatan proliferasi cell line HeLa.
Rupanya pengaruh ekstrak umbi keladi tikus ini spesifik terhadap ketiga macam
cell line tersebut. Ekstrak umbi keladi tikus memiliki senyawa aktif berupa triterpenoid dan
steroid. Kedua senyawa ini berperan penting sebagai anti inflamasi, analgesik dan
sitotoksik. Senyawa ini juga menunjukkan aktivitas antibakteri dan antivirus
(Robinson,1990). Triterpenoid menghambat proliferasi sel kanker dengan menghambat
kerja enzim DNA Topoisomerase. Enzim tersebut adalah enzim yang berperan dalam
proses replikasi, transkripsi dan rekombinasi DNA dan juga proliferasi dan diferensiasi sel
kanker; enzim ini merupakan target bahan bioaktif tanaman yang memiliki aktivitas anti
kanker, karena dengan dihambatnya enzim DNA Topoisomerase, maka proses dalam sel
Pengaruh Ekstrak Umbi Keladi Tikus Biosains
penghambatan DNA Topoisomerase, ada beberapa mekanisme lain yang dapat
menghambat proliferasi sel kanker diantaranya, penghambatan kerja kinase oleh
senyawa flavonoid dan peningkatan supresor gen p53 yang menyebabkan terjadinya
kematian sel (Sismindari, 2003).
Kecenderungan terjadinya penurunan rerata persentase IP apabila konsentrasi
ekstrak ditingkatkan seperti yang terlihat pada cell line A-132, diduga karena
kemungkinan sel resisten terhadap senyawa aktif yang terkandung dalam ekstrak. Ada
beberapa mekanisme yang menyebabkan sel kanker menjadi resisten terhadap senyawa
aktif yang diberikan, antara lain meningkatnya kemampuan ekstruksi sel terhadap
senyawa aktif yang diberikan termasuk komponen-komponen yang terkandung di
dalamnya. Hal tersebut menyebabkan senyawa aktif tidak dapat mencapai target karena
dikeluarkan dari sel sebelum bekerja. Resistensi sel kanker juga dapat terjadi karena
detoksifikasi dari senyawa aktif yang diberikan. Mekanisme lainnya adalah manipulasi dari
protein inti sel, sehingga kadar enzim Topoisomerase menjadi rendah, padahal salah satu
mekanisme kerja dari senyawa aktif kanker adalah dengan menghambat kerja dari enzim
tersebut (Davies dan Hickson, 1996). Penelitian ini menunjukkan bahwa ada perbedaan
karakteristik pada ketiga cell line dalam merespons ekstrak umbi keladi tikus.
4. KESIMPULAN DAN PROSPEK
Ekstrak umbi keladi tikus berpengaruh terhadap penghambatan proliferasi ketiga
macam cell line kanker. Cell line HeLa menunjukkan kecenderungan adanya korelasi
positif antara konsentrasi ekstrak dan persentase IP. Sedangkan ekstrak umbi tidak
berpengaruh secara signifikan terhadap penghambatan proliferasi pada cell line K-562
dan A-132.
Mengingat prospek tanaman ini di masa mendatang, penelitian ini perlu
dilanjutkan dengan memanfaatkan teknik kultur jaringan untuk mendapatkan klon yang
stabil terhadap kandungan senyawa aktif yang ada pada umbi keladi tikus, sehingga kelak
dapat diperoleh obat herbal yang bermanfaat untuk menyembuhkan penyakit kanker.
UCAPAN TERIMA KASIH
Kami mengucapkan terima kasih kepada Pimpinan Lambaga Penelitian UNJ yang
telah mendanai penelitian ini melalui Dana DIPA RUTIN UNJ, No. 54/SPK/LP-UNJ/DIPA
Pengaruh Ekstrak Umbi Keladi Tikus Biosains
DAFTAR PUSTAKA
Asri, Christiani, Supriyatin. 2007. Pengaruh ekstrak umbi keladi tikus (Typhonium
flagelliforme (Lodd)BL.) terhadap penghambatan proliferasi cell-line A-132 dan K-562. Bioma V: 1-4.
Brown, R. 1996. Chemotheraputic Druginduced DNA Damage and Repair. Di dalam Molecular Biology for Oncologist. Chapman & Hall: London.
Christiani, dkk. 2006. Pemanfaatan umbi keladi tikus (Typhonium flagelliforme (Lodd)BL.)
sebagai upaya pengobatan penyakit kanker (belum dipublikasikan).
Davies, S.L. dan Hickson. I.D. 1996. Molecular Basic of Drug Resistance. Di dalam Molecular Biology for Oncologist. Chapman & Hall: London.
Feshney, R.I. 1992. Introduction to Basic Priciples. Di dalam Freshney Animal Cell Culture
A Practical Approach, Second Edition. Oxford University Press, New York, Hlm: 1-13.
Handoko, B.J.F. 2005. Karakteristik dan uji hayati pendahuluan metabolit sekunder aktif
dari fraksi etil asetat umbi keladi tikus (Typhonium divaricatum(L.) Decne). (Skripsi).
Jakarta: Universitas Pancasila.
Indriati. 2002. Isolasi senyawa bioaktif antikanker dari tanaman keladi tikus (Typhonium
divaricatum (L.) Decne). (Skripsi). Jakarta: Universitas Negeri Jakarta.
Maruer. 1992. Toward serum-free, chemically defined media for mamalia cell culture. Di
dalam Freshney Animal Cell Culture A Practical Approach, Second Edition. Edited.
Oxford University Press, New York, Hlm: 15-46.
Ratna, Christiani, Arleni. 2006. Uji aktivitas penghambatan proliferasi cell line K-562 oleh
ekstrak keladi tikus (Typhonium flagelliforme (Lodd)BL.) secara in vitro. Bioma V:
14-18.
Robinson, T. 1995. Kandungan Organik Tumbuhan Tingkat Tinggi. Terjemahan Kosasih Padmawinata. Penerbit ITB Bandung.
Sismindari. 2003. Cytotoxic effects of methanol ectract isolated from Eryrhrina fusca
Lourvleaves on cancer cell-lines. Berkala Ilmu Kedokteran Vol. 35 No. 2. Hlm.
PENGARUH MACAM NUTRISI DAN PEMBERIAN EKSTRAK Biosains
PENGARUH MACAM NUTRISI DAN PEMBERIAN EKSTRAK BUAH PISANG TERHADAP PERTUMBUHAN PLANTLET ANGGREK DENDROBIUM
SECARA IN VITRO
(The Influence of Nutrition Kinds and Banana Extract Toward the In Vitro Growth of Dendrobium Plantlet)
Samanhudi1)*, Ahmad Yunus1), dan Wangi Satutik2)
1)
Jurusan Agronomi Fakultas Pertanian Universitas Sebelas Maret Surakarta Jl. Ir. Sutami 36 A, Surakarta 57126 Telp/Fax. (0271) 632451
*
Penulis untuk Korespondensi : HP. 0813 290 60000 Email : [email protected]
2) Mahasiswa Jurusan Agronomi, Fakultas Pertanian UNS
Abstrak
Penelitian ini bertujuan untuk mencari nutrisi media alternatif dan konsentrasi ekstrak pisang yang tepat untuk pembiakan anggrek Dendrobium secara in vitro. Penelitian dilaksanakan di Laboratorium Fisiologi Tumbuhan dan Bioteknologi Fakultas Pertanian Universitas Sebelas Maret Surakarta pada bulan Januari sampai Mei 2009. Penelitian ini menggunakan Rancangan Acak Lengkap yang disusun secara faktorial. Faktor pertama adalah macam nutrisi yaitu VW, Growmore, Formula AB-Mix. Faktor kedua adalah konsentrasi ekstrak pisang yaitu 0, 50l, 100, dan 150 g/l sehingga terdapat 12 kombinasi perlakuan masing-masing diulang sebanyak tiga kali. Hasil penelitian menunjukkan bahwa semua macam nutrisi yang diujikan dapat digunakan sebagai media alternatif untuk pembiakan anggrek Dendrobium secara in vitro. Media tanpa penambahan ekstrak pisang menunjukkan hasil terbaik pada variabel tinggi plantlet, jumlah daun, saat muncul tunas dan jumlah tunas, sedangkan media dengan penambahan ekstrak pisang pada konsentrasi 50 g/l menghasilkan panjang akar yang terpanjang. Kombinasi perlakuan nutrisi AB-Mix dengan penambahan ekstrak pisang 50 g/l menghasilkan saat muncul akar tercepat (14 HST). Kombinasi perlakuan AB-Mix tanpa penambahan ekstrak pisang menghasilkan jumlah akar terbanyak (10 akar). Perlakuan yang menghasilkan kalus antara lain pada kombinasi perlakuan nutrisi VW, Growmore dan AB-Mix dengan penambahan ekstrak pisang 0 dan 50 g/l. Tekstur kalus yang dihasilkan kompak dan warna kalus hijau..
Kata kunci: Nutrisi, ekstrak buah pisang, anggrek Dendrobium, in vitro.
1. PENDAHULUAN
Anggrek merupakan salah satu tanaman hias yang sangat prospektif sebagai
komoditas non migas. Anggrek termasuk ke dalam famili Orchidaceae, suatu famili yang
sangat besar dan sangat bervariasi. Anggrek Dendrobium menarik minat para penggemar
tanaman hias karena mempunyai warna, bentuk dan ukuran yang sangat beragam, serta
mempunyai daya tahan kesegaran bunga lebih lama sebagai bunga potong.
Teknik perbanyakan tanaman anggrek yang telah lazim dilakukan adalah dengan
PENGARUH MACAM NUTRISI DAN PEMBERIAN EKSTRAK Biosains
dalam bentuk bibit dalam botol atau kultur biji (Sriyanti, 2007). Media kultur yang biasa
digunakan dalam kultur jaringan anggrek merupakan media yang teramu dalam media
MS (Murashige and Skoog), VW (Vacin and Went) dan Knudson. Sayangnya bahan baku
media kultur tersebut, selain masih sulit diperoleh juga relatif mahal untuk skala produksi
bagi para petani atau para pemula anggrek (Sandra, 2004).
Oleh karena itu, diperlukan media kultur alternatif dengan berbagai macam nutrisi
yang dapat terjangkau dan mudah didapat. Nutrisi yang digunakan untuk kultur anggrek
selain nutrisi VW dapat menggunakan salah satu pupuk lengkap anorganik misal
Growmore dengan kandungan NPK (32-10-10) dan nutrisi hidroponik misal resep formula
AB-Mix EC 2 mS/cm (Electro Conductivity atau daya hantar listrik dengan satuan milli
siemens persenti meter), dengan penambahan suplemen dan ZPT alami. Penggunaan
pisang merupakan penggunaan salah satu bahan organik yang sangat umum diberikan
dalam media kultur anggrek. Buah pisang sering digunakan sebagai sumber karbohidrat,
suplemen dan ZPT (auksin) dalam penanaman anggrek secara in vitro, dapat
meningkatkan pertumbuhan dan deferensiasi sel pada tanaman. Pemberian buah pisang
ambon pada subkultur plantlet anggrek dendrobium dapat memacu pertumbuhan
(Widiastoety dan Bahar, 1995). Penelitian ini bertujuan untuk mencari media alternatif
serta konsentrasi pisang yang tepat untuk pembiakan Dendrobiumsp secara in vitro.
2. METODE PENELITIAN
Penelitian ini dilaksanakan pada bulan Januari sampai Mei 2009 bertempat di
Laboratorium Fisiologi Tumbuhan dan Bioteknologi Fakultas Pertanian Universitas
Sebelas Maret Surakarta. Bahan dan alat yang digunakan dalam penelitian ini meliputi :
bahan tanam yang dipergunakan dalam penelitian ini meliputi plantlet anggrek silangan
Dendrobium Imelda romualdez / Alice noda >< Dendrobium Tomie / Waipahu pink,
berumur kurang lebih 4 bulan setelah semai dengan pertumbuhan calon daun tinggi 1-2
cm, belum membuka sempurna dan belum ada akar. Bahan kimia yang dipergunakan
meliputi media vacin and Went (VW). Nutrisi pupuk lengkap anorganik (Growmore), nutrisi
hidroponik formula AB Mix, alkohol 96%, spiritus, arang aktif, larutan deterjen, NaOH 1N,
HCl 1 N. Bahan organik yang digunakan meliputi air kelapa hijau muda, pisang raja bulu,
agar-agar powder, gula pasir dan akuades. Alat-alat yang digunakan dalam penelitian
adalah botol kultur, LAF (Laminar Air Flow Cabinet), refrigerator, autoklaf, petridish,
erlenmeyer, pH meter, timbangan digital, pipet ukur, gelas ukur, hot plate magnetic stirrer,
pisau scalpel, pinset, lampu Bunsen, alumunium foil, kertas label, plastik tahan panas, EC
PENGARUH MACAM NUTRISI DAN PEMBERIAN EKSTRAK Biosains
Penelitian ini menggunakan Rancangan Acak Lengkap (RAL) yang disusun secara
faktorial dengan dua faktor perlakuan yaitu macam nutrisi VW (N1), Growmore (N2),
Formula AB-Mix (N3) dan konsentrasi ekstrak pisang 0 g/l (P0), 50 g/l (P1), 100 g/l (P2),
150 g/l (P3). Setiap kombinasi perlakuan dilakukan ulangan sebanyak tiga kali. Penelitian
ini dilaksanakan melalui tahap-tahap sebagai berikut: sterilisasi botol dan alat, pembuatan
larutan stok, penyiapan media, penanaman plantlet dan pemeliharaan. Peubah yang
diamati meliputi tinggi plantlet, jumlah daun, saat muncul akar, jumlah akar, panjang akar,
saat muncul tunas, jumlah tunas dan saat muncul kalus.
Data hasil penelitian dianalisis dengan menggunakan analisis ragam dengan uji F
pada taraf 5% dan apabila signifikan dilanjutkan pada DMRT taraf 5%. Variabel saat
muncul kalus dianalisis secara deskriptif.
3. HASIL DAN PEMBAHASAN
Keberhasilan penggunaan metode kultur jaringan sangat tergantung pada jenis
media atau nutrisi yang digunakan. Media kultur tidak hanya mengandung unsur makro
dan mikro, tetapi juga karbohidrat sebagai sumber karbon atau bahan organik lainnya
(Widiastoety dan Purbadi, 2003). Untuk mendapatkan media yang lebih ekonomis adalah
dengan mengembangkan metode pengkombinasian berbagai komposisi nutrisi dengan
ZPT organik. Penggunaan komposisi nutrisi dan ekstrak buah pisang dengan komposisi
yang berbeda diduga dapat mempengaruhi pertumbuhan plantlet anggrek Dendrobium
secara in vitro.
3.1. Tinggi Plantlet
Pertumbuhan adalah proses kehidupan tanaman yang mengakibatkan perubahan
ukuran tanaman semakin besar. Tinggi tanaman merupakan ukuran yang paling sering
diamati baik sebagai indikator pertumbuhan maupun sebagai parameter yang digunakan
untuk mengukur pengaruh perlakuan yang diterapkan.
1.4467a
Gambar 1. Pengaruh konsentrasi pisang terhadap tinggi plantlet anggrek
PENGARUH MACAM NUTRISI DAN PEMBERIAN EKSTRAK Biosains
Hasil analisis ragam menunjukkan bahwa penambahan konsentrasi pisang
memberikan pengaruh berbeda nyata terhadap variabel tinggi plantlet, sedangkan
komposisi nutrisi dan interaksi keduanya memberikan pengaruh tidak nyata terhadap
tinggi plantlet. Setelah dilakukan uji regresi, terdapat kecenderungan bahwa peningkatan
konsentrasi buah pisang hingga 150 g/l menghambat pertumbuhan tinggi plantlet
Dendrobium. Pisang mengandung auksin dan giberelin yang berperan dalam pembesaran
dan pemanjangan sel, sehingga sangat mungkin penggunaannya dalam jumlah tertentu
dapat membantu meningkatkan tinggi tanaman.
Komposisi nutrisi dan interaksi antara kedua perlakuan memberikan respon yang
tidak berbeda nyata terhadap tinggi plantlet. Dengan demikian dapat dikatakan bahwa,
peningkatan maupun penurunan tinggi plantlet lebih dipengaruhi oleh penambahan
konsentrasi buah pisang.
3.2. Jumlah Daun
Jumlah daun merupakan salah satu indikator pertumbuhan tanaman dan dapat
digunakan sebagai data penunjang untuk menjelaskan proses pertumbuhan yang terjadi
(Sitompul dan Guritno, 1995). Penghitungan jumlah daun dilakukan pada akhir
pengamatan dengan menghitung tiap helai daun pada tiap plantlet dan tunas.
Hasil analisis sidik ragam menunjukkan bahwa penambahan konsentrasi buah
pisang menunjukkan pengaruh yang sangat nyata terhadap jumlah daun, sedangkan
komposisi nutrisi dan interaksi antara keduanya menunjukkan pengaruh yang tidak nyata
terhadap jumlah daun.
Gambar 2. Pengaruh konsentrasi buah pisang terhadap jumlah daun plantlet anggrek Dendrobium secara in vitro.
Hasil uji regresi menunjukkan terdapat kecenderungan bahwa penambahan buah
pisang justru menyebabkan terhambat jumlah daun dengan pola linear. Pertmbuhan
terhambat jumlah daun ini lebih disebabkan terhambatnya jumlah tunas, sehingga
semakin sedikit tunas yang terbentuk, semakin sedikit pula terbentuknya daun pada
PENGARUH MACAM NUTRISI DAN PEMBERIAN EKSTRAK Biosains
penggunaan sumber karbohidrat dengan konsentrasi tinggi. Secara visual, tanaman yang
mengalami tekanan karena pengaruh osmotik berupa penghambatan pertumbuhan
ukuran dan jumlah daun. Widiastoety dan Purbadi (2003) menjelaskan bahwa tekanan
yang disebabkan oleh perubahan osmotik akan merangsang akumulasi asam absisat
(ABA) di dalam jaringan tanaman yang dapat menghambat pertumbuhan tanaman dalam
media. Selain akumulasi ABA, terjadi pula penghambatan sintesis sitokinin yang efeknya
memperkuat penghambatan pertumbuhan yang diakibatkan oleh pengaruh ABA.
Komposisi nutrisi dan interaksi antara kedua perlakuan menunjukkan respon tidak
berbeda nyata. Hal ini diduga karena kondisi fisiologis masing-masing eksplan akan
memberikan respon yang berbeda terhadap perlakuan yang diberikan, mengingat
eksplant berasal dari pembiakan secara generatif yaitu melalui biji, sehingga terdapat
variasi genetik yang mengakibatkan perbedaan kondisi fisiologis masing-masing individu.
3.3. Saat Muncul Akar
Akar merupakan organ vegetatif utama yang memasok air, mineral dan
bahan-bahan yang penting untuk pertumbuhan dan perkembangan tanaman (Gardner et al.,
1991). Saat munculnya akar menjadi faktor yang penting dalam pertumbuhan tanaman
karena tanaman akan lebih mudah menyerap unsur-unsur yang terdapat dalam media
kultur.
Hasil analisis ragam menunjukkan bahwa perlakuan komposisi nutrisi dan
penambahan buah pisang memberikan pengaruh tidak berbeda nyata terhadap saat
muncul akar plantlet Dendrobium. Namun, interaksi keduanya memberikan pengaruh
nyata terhadap saat muncul akar plantlet Dendrobium. Penambahan buah pisang tidak
mampu mempercepat kemunculan akar, dan jika dikombinasikan dengan komposisi
nutrisi, perlakuan N3P1 (Nutrisi AB-Mix dan penambahan ekstrak pisang 50 g/l)
menunjukkan rata-rata waktu pembentukan akar paling lambat, yaitu 45 HST (HST).
Keadaan ini mengindikasikan bahwa kemunculan akar tidak bergantung pada komposisi
nutrisi yang digunakan untuk media kultur.
Penambahan buah pisang juga tidak mampu mempercepat kemunculan akar. Hal ini
dimungkinkan kandungan unsur-unsur yang terdapat pada pisang mampu melengkapi
maupun menambah kuantitas unsur-unsur pada masing-masing nutrisi, sehingga interaksi
antara keduanya dapat memberikan pengaruh yang nyata terhadap saat muncul akar.
Pengaruh interaksi antara kedua perlakuan yang terbaik terdapat pada kombinasi
PENGARUH MACAM NUTRISI DAN PEMBERIAN EKSTRAK Biosains
Gambar 3. Pengaruh macam nutrisi dan konsentrasi buah pisang pada media kultur terhadap saat muncul akar plantlet Dendrobium.
3.4. Jumlah Akar
Jumlah unsur hara yang diserap tanaman tergantung pada kesempatan untuk
mendapatkan nutrisi dalam media. Variabel pengamatan jumlah akar merupakan indikasi
pertumbuhan dan pelebaran akar dalam usahanya mempeluas permukaan bidang serap.
Berdasarkan hasil penelitian, semua kombinasi perlakuan mampu memunculkan
akar. Hasil analisis ragam menunjukkan bahwa kompisisi nutrisi dan penambahan buah
pisang serta interaksi antara keduanya memberikan pengaruh tidak berbeda nyata
terhadap jumlah akar Dendrobium.
0
Gambar 4. Pengaruh komposisi nutrisi dan konsentrasi ekstrak pisang pada media kultur terhadap jumlah akar plantlet Dendrobium.
Gambar 4 menunjukkan bahwa jumlah akar terbanyak terdapat pada kombinasi
perlakuan N3P0 (nutrisi AB-Mix tanpa penambahan buah pisang), sedangkan jumlah akar
terkecil terdapat pada kombinasi perlakuan (N1P3). Hal ini dapat diinterpretasikan bahwa
PENGARUH MACAM NUTRISI DAN PEMBERIAN EKSTRAK Biosains
Perbedaan komposisi masing-masing jenis nutrisi baik dari segi kandungan maupun
jumlah unsur-unsurnya diduga mengakibatkan respon yang berbeda terhadap jumlah akar
masing-masing perlakuan. Penambahan auksin dari buah pisang diperlukan jaringan
tanaman untuk membentuk akar. Meskipun demikian penambahan auksin tidak
selamanya meningkatkan jumlah akar sebab penambahan auksin jenis tertentu dengan
konsentrasi tertentu dapat pula menghambat jumlah akar (Fuchs, 1986).
Kombinasi perlakuan memberikan respon tidak berbeda nyata terhadap jumlah akar.
Hal ini diduga disebabkan komposisi nutrisi dan buah pisang yang ditambahkan ke dalam
media kultur belum mencukupi kebutuhan nutrisi eksplan anggrek, sehingga
perkembangan akar sedikit terhambat. Pertumbuhan akar juga tergantung pada peran
unsur fosfor, kalsium, mangan, besi, dan boron. Sebagian besar fosfor didalam tanaman
adalah sebagai zat pembangun dan terikat dalam senyawa-senyawa organik dan hanya
sebagian kecil terdapat dalam bentuk anorganik sebagai ion-ion fosfat (Anonim, 2007).
3.5. Panjang Akar
Panjang akar merupakan hasil perpanjangan sel-sel di belakang meristem
ujung (Dewi, 2007). Pengukuran panjang akar dilakukan dengan mengukur akar
terpanjang pada plantlet. Selain untuk menyerap unsur hara, akar berfungsi sebagai
penguat berdirinya tanaman.
Berdasarkan hasil analisis ragam diketahui bahwa komposisi nutrisi tidak
berpengaruh terhadap variabel panjang akar, tetapi penambahan buah pisang
berpengaruh nyata terhadap variabel panjang akar, sedangkan interaksi antara kedua
perlakuan memberikan pengaruh tidak berbeda nyata terhadap panjang akar. Pemberian
auksin pada plantlet anggrek Dendrobium dapat merangsang pertumbuhan akar
(Widiastoety et al., 2004). Hasil penelitian Arditti dan Ernts (1992) menunjukkan bahwa
buah pisang mengandung hormon tumbuh seperti auksin dan giberelin.
1.5767a
Gambar 5. Pengaruh konsentrasi ekstrak pisang terhadap panjang akar plantlet anggrek
PENGARUH MACAM NUTRISI DAN PEMBERIAN EKSTRAK Biosains
Berdasarkan uji regresi, terdapat kecenderungan bahwa dengan peningkatan
konsentrasi pisang sampai 150 g/l akan menghambat pertumbuhan panjang akar. Hal ini
sesuai dengan hasil penelitian pada kultur Piretrum (chrysanthemum cinerariifolium)
bahwa penghambatan panjang akar sejalan dengan konsentrasi auksin yang semakin
meningkat (Rostiana dan Seswita 2007) pemberian dapat membentuk akar lebih banyak,
tetapi akan menghambat proses pemanjangan akar lateral. Hal ini sesuai dengan
pendapat Salisbury dan Ross (1995) bahwa konsentrasi zat pengatur tumbuh yang terlalu
tinggi untuk suatu jenis tanaman tertentu akan mendorong sintesis etilen yang kemudian
menghambat pemanjangan akar.
Selain akibat peningkatan auksin yang menyebabkan sintesis etilen yang dapat
menghambat pemanjangan akar, penghambatan pemanjangan akar ini juga disebabkan
oleh kelebihan karbohidrat atau zat gula yang diakibatkan oleh penambahan buah pisang.
Peningkatan konsentrasi gula dalam media kultur akan megakibatkan kondisi di luar sel
tanaman menjadi hipertonis, sehingga kondisi ini menyebabkan tekanan osmotik sel
tanaman meningkat, akibatnya terjadi plasmolisis yang menyebabkan kematian sel-sel
somatik pada akar. Macam nutrisi yang digunakan, baik VW, Growmore maupun AB-Mix,
tidak mampu meningkatkan pemanjangan akar karena pemanjangan akar dipengaruhi
oleh konsentrasi auksin dan karbohidrat yang terkandung dalam buah pisang. Analisis
ragam juga menunjukkan bahwa tidak terdapat interaksi antara kedua perlakuan.
3.6. Saat Muncul Tunas
Tunas merupakan ranting muda yang baru tumbuh atau calon tanaman baru yang
tumbuh dari bagian tanaman (Rahardja dan Wiryanta, 2003). Pengamatan saat muncul
tunas dilakukan untuk mengetahui tingkat kefektifan suatu kegiatan kultur jaringan dalam
menghasilkan tunas.
Hasil analisis ragam menunjukkan bahwa komposisi nutrisi tidak berpengaruh nyata
terhadap saat muncul tunas, sedangkan konsentrasi pisang berpengaruh nyata terhadap
rata-rata saat muncul tunas, serta interaksi antara kedua perlakuan berpengaruh sangat
nyata terhadap rata-rata saat muncul tunas. Tampak bahwa komposisi nutrisi dan
penambahan buah pisang menunjukkan kecepatan saat munculnya tunas yang
bervariasi.
Hal ini diduga disebabkan sifat genetik masing-masing plantlet berbeda satu sama
lain mengingat plantlet yang digunakan berasal dari biji hasil persilangan dari dua verietas
anggrek yang berbeda, sehingga setiap individu yang dihasilkan dari proses persilangan
mempunyai potensi berbeda dalam hal kecepatan pembentukan tunas. Hal ini dikuatkan
oleh pendapat Tisdale dan Nelson (1975), bahwa dari beberapa hasil penelitian
PENGARUH MACAM NUTRISI DAN PEMBERIAN EKSTRAK Biosains
Gambar 6. Pengaruh konsentrasi buah pisang terhadap saat muncul tunas plantlet anggrek Dendrobium secara in vitro.
Komposisi nutrisi diketahui memberikan respon tidak berbeda nyata terhadap saat
muncul tunas pada plantlet anggrek. Hal ini diduga disebabkan oleh perbedaan kadar
unsur hara pada masing-masing jenis nutrisi.
3.7. Jumlah Tunas
Jumlah tunas merupakan faktor terpenting dalam multiplikasi tanaman pada kultur
jaringan. Semakin banyak tunas yang terbentuk, dapat dilakukan multiplikasi kultur untuk
mendapatkan tunas-tunas baru dalam jumlah yang semakin banyak juga.
Hasil analisis ragam menunjukkan bahwa penambahan buah pisang pada berbagai
taraf konsentrasi memberikan pengaruh berbeda nyata terhadap jumlah tunas, akan
tetapi macam nutrisi maupun interaksi antara keduanya memberikan pengaruh tidak
berbeda nyata terhadap variabel jumlah tunas. Penambahan buah pisang ternyata justru
menghambat pertumbuhan jumlah tunas Dendrobium. Hal ini sesuai dengan penelitian
Yunus (2007) pada ekplan bawang merah, bahwa diduga karena fungsi auksin yang
cenderung memacu pembentukan dan pertumbuhan akar sehingga efek yang
ditimbulkannya dapat menghambat pembentukan tunas.
0.22a
PENGARUH MACAM NUTRISI DAN PEMBERIAN EKSTRAK Biosains
Komposisi nutrisi tidak berbeda nyata terhadap variabel jumlah tunas. Ini berarti
bahwa komposisi nutrisi yang digunakan dalam kultur Dendrobium tidak mempengaruhi
jumlah tunas yang terbentuk pada eksplan. Penghambatan pertumbuhan jumlah tunas
hanya dipengaruhi oleh penambahan ekstrak pisang. Analisis ragam juga menunjukkan
bahwa tidak ada interaksi antara macam nutrisi dan penambahan buah pisang terhadap
variabel jumlah tunas. Hal ini diduga karena perbedaan status hara pada masing-masing
nutrisi belum mencukupi untuk multiplikasi tunas.
3.8. Saat Muncul Kalus
Kemunculan kalus ditandai dengan adanya perubahan bentuk pada pangkal
eksplant, seperti terjadi pembengkakan pada jaringan yang mengalami kontak dengan
media secara langsung. Pada penelitian ini, perlakuan N3P1 mampu membentuk kalus.
Hal ini dimungkinkan bahwa kandungan auksin eksogen yang terdapat pada buah pisang
dapat digunakan untuk pertumbuhan kalus, sedangkan pada perlakuan N1P0, N2P0 dan
N3P0 sebagian ulangan dapat membentuk kalus pada 2 MST. Diduga kandungan auksin
endogen pada plantlet mampu membentuk kalus. Auksin umumnya ditambahkan ke
dalam nutrisi media untuk menginduksi kalus dari eksplan (George dan Sherrington,
1984). Warna kalus yang terbentuk pada plantlet anggrek Dendrobium adalah hijau
dengan tektur kompak. Semua kalus yang terbentuk dapat tumbuh pada kehidupan
selanjutnya yaitu, dapat tumbuh tunas. Tekstur kalus merupakan salah satu penanda
yang dipergunakan untuk menilai kualitas suatu kalus.
4. KESIMPULAN
Media VW, Gromore dan AB-Mix tanpa penambahan pisang opimal untuk
meningkatkan tinggi tanaman, jumlah daun, dan jumlah tunas angger Dendrobium secara
in vitro. Saat muncul akar tercepat dan jumlah akar terbanyak terdapat pada kombinasi
perlakuan media dengan nutrisi AB-Mix tanpa penambahan pisang. Panjang akar paling
optimal dicapai pada media VW, Gromore, dan AB-Mix dengan penambahan pisang 50
g/l. Penambahan 150 g/l pisang pada media VW, Gromore, dan AB-Mix menghasilkan
saat muncul tunas tercepat.
DAFTAR PUSTAKA
Anonim. 2007. Mineral Bagi Tanaman. http://www.tanimdo.com/abdi4/hal 2701. htm. diakses pada tanggal 3 Juli 2009.
Arditti, J. and R. Ernts. 1992. Micropropagation of Orchids. Irvine. Departement of Developmental and Cell Biology, University of California.