TINJAUAN PUSTAKA
Karakteristik Geminivirus
Morfologi, Taksonomi dan Molekuler
Geminivirus merupakan salah satu kelompok virus patogen tumbuhan terbesar. Nama geminivirus berasal dari karakteristik morfologi partikel virus berupa isometrik kembar yang selalu berpasangan (twinned-geminate) dengan ukuran berkisar 20-30 nm. Secara genetik geminivirus memiliki genom berupa DNA utas tunggal (ssDNA) yang berbentuk sirkuler (Harrison & Robinson 1999).
Dhar & Singh (1996) dan Harison & Robinson (1999) menjelaskan bahwa geminivirus diklasifikasikan ke dalam famili Geminiviridae yang dibagi ke dalam tiga genus yaitu Mastrevirus, Curtovirus, dan Begomovirus yang didasarkan atas perbedaan kisaran inang, serangga vektor dan genomnya. Mastrevirus adalah geminivirus yang menginfeksi tanaman inang monokotil, ditularkan oleh serangga vektor wereng daun dan memiliki struktur genom monopartit. Curtovirus, menginfeksi tanaman dikotil dengan vektor dan struktur genom sama dengan genus pertama. Begomovirus, menginfeksi tanaman dikotil dan ditularkan oleh kutukebul (Bemisia tabaci) memiliki struktur genom monopartit atau bipartit. Menurut Van Regenmortel (2000) selain ketiga genus tersebut, ada satu genus lainnya yang termasuk kedalam famili Geminiviridae yaitu Topocuvirus yang menginfeksi tanaman dikotil, ditularkan oleh wereng pohon dan memiliki genom monopartit.
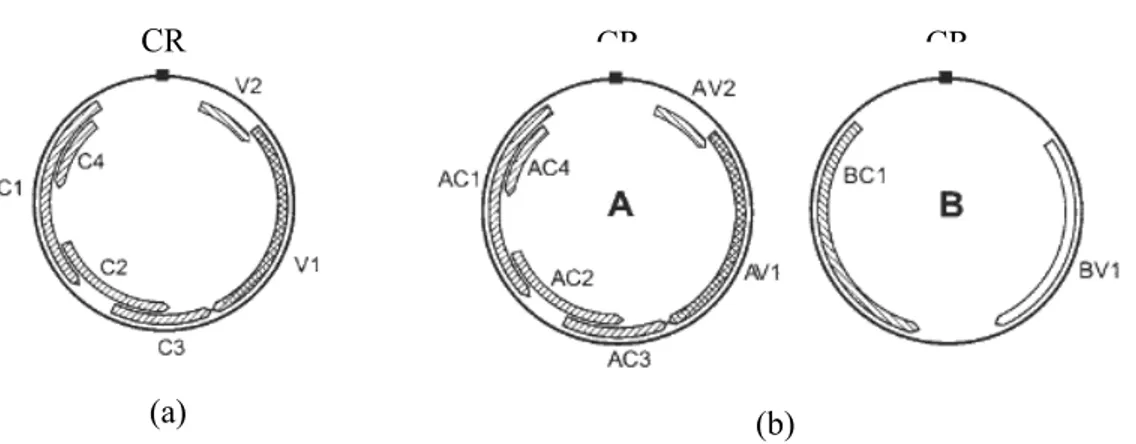
Diantara genus-genus tersebut diatas, Begomovirus merupakan genus dengan jumlah anggota terbesar. Genus Begomovirus terdiri dari virus-virus dengan genom bipartit atau monopartit. Sebagian besar anggota genus Begomovirus memiliki genom bipartit yang terdiri dari dua molekul DNA utas tunggal sirkuler yang berbeda yaitu DNA A dan DNA B dengan masing-masing berukuran 2,7-2,8 kb. Begomovirus dengan genom monopartit, semua gennya terletak pada satu DNA utas tunggal sirkuler yang berukuran 2,8 kb (Gambar 1). Komponen DNA geminivirus baik monopartit maupun bipartit mengandung gen-gen yang menyandikan protein dengan fungsi yang khusus (Tabel 1). Gen penyandi protein selubung virus merupakan daerah genom yang mempunyai
runutan DNA dengan derajat kesamaan yang tinggi antara anggota geminivirus dalam satu genus (Rojas et al. 1997).
Tabel 1 Jenis dan fungsi gen Begomovirus
Monopartit Bipartit Protein dan fungsi
V1 AV1 Protein selubung virus (coat protein), berperan dalam penyebaran virus, pergerakan virus di dalam inangnya dan berperan dalam penularan yaitu melindungi partikel virus dari degradasi pada saat masuk sistem pencernaan kutukebul (Briddon et al. 1989; Morin et al. 2000; Hull 2002; Harrison & Robinson 1999)
V2 AV2 Movement protein (MP), berperan dalam pergerakan virus dalam tanaman terinfeksi (Hull 2002; Harrison & Robinson 1999)
C1 AC1 Replication-associated protein (Rep), berperan dalam proses replikasi virus (Desbiez et al. 1995; Hull 2002)
C2 AC2 Transcriptional activator protein (TrAP), protein yang terlibat dalam pengaktifan transkripsi dari promoter protein selubung. Protein ini ditemukan pada inti dan berperan dalam patogenisitas virus (van Wezel et al. 2001)
C3 AC3 Replication enhancer protein (REn), protein ini berinteraksi dengan protein C1 dan meningkatkan akumulasi DNA virus (Hanleybowdoin et al. 2000) C4 AC4 Berinteraksi dengan C1 dan V2, berperan dalam
penentu gejala dan terlibat dalam inisiasi pembelahan sel (Krake et al. 1998), pergerakan DNA virus dari sel ke sel (Rojas et al. 2001), mematahkan mekanisme pertahanan tanaman (van Wezel et al. 2002),
- BV1 Nuclear shuttle protein (NSP) dan menyandikan virion DNA B (Hull 2002; Salati 2002)
- BC1 Movement protein (MP), berperan dalam pergerakan virus di dalam tanaman terinfeksi (Hull 2002; Salati 2002).
Gambar 1 Organisasi genom Begomovirus: Genom monopartit (a), Genom bipartit (b) (Hull 2002)
Daerah Penyebaran dan Kisaran Inang
Geminivirus memiliki daerah penyebaran yang sangat luas terutama di daerah tropis dan subtropis yang dapat mendukung perkembangan kutukebul dengan baik. Dilaporkan bahwa di Meksiko, Venezuela, Amerika Serikat (Florida), dan beberapa negara di Amerika Tengah serta Karibia infeksi geminivirus mengakibatkan hancurnya industri tomat (Polston & Anderson 1997). Infeksi geminivirus isolat cabai di Texas dan tomato yellow leaf curl virus di Israel menyebabkan kehilangan hasil 100% (Stenger et al. 1990; Pico et al. 1996). Di Indonesia geminivirus pertama kali diketahui menyerang tanaman tembakau di Bojonegoro, Jawa Timur yang menyebabkan kerusakan sebesar 30% (Trisusilowati 1990). Infeksi geminivirus juga telah terdeteksi pada tanaman cabai di daerah Jawa Barat, Daerah Istimewa Yogyakarta dan Kalimantan Selatan (Rusli et al. 2000; Sulandari et al. 2001). Geminivirus telah menyebabkan epidemi penyakit di beberapa sentra produksi cabai di Indonesia terutama di Jawa mulai tahun 2000 sampai 2003.
Geminivirus memiliki kisaran inang yang cukup luas baik pada tanaman budidaya maupun gulma. Tomat, cabai, tembakau, mentimun, terung, ubikayu dan kacang-kacangan adalah inang geminivirus dari tanaman budidaya. Babadotan (A. conyzoides) merupakan gulma yang telah dilaporkan sebagai inang geminivirus di daerah tropis dan subtropis (Sukamto et al. 2005). Gulma lainnya yang dapat menjadi inang geminivirus adalah Sida spp., Macroptilium lathyroides, dan Wissadula amplissima yang ditemukan di Jamaica. Malvastrum
CR CR CR
coromandelianum merupakan gulma yang dapat menjadi inang geminivirus di
Guangdong, Cina (Wu et al. 2007). Achyranthes aspera, Euphorbia heterophylla, Nicandra physaloides, Commelina erecta, Amaranthus spinosus, Erigeron floribundus, A. conyzoides, Bidens pilosa, Sida acuta, Ipomoea batatas, Amaranthus viridis, Portulaca oleracea, Cassia obtusifolia, Euphorbia hirta, Calopogonium mucunoides, Clotalaria retusa, Trianthema portulacastrum, Alternanthera sessilis, Celosia trigyna, Commelina diffusa, Chromolaena odorata, Eclipta prostrata, Synedrella nodiflora, Cassia occidentalis, Spigelia anthelmia, Boerhavia diffusa, Physalis angulata dan Acanthospermum hispidis
adalah 28 spesies gulma yang potensial sebagai sumber tomato yellow leaf curl geminivirus (TYLCV) di Tanzania. H. brevipes, P. floridana, C. juncea, A. conyzoides bunga putih dan ungu adalah gulma yang rentan terhadap geminivirus isolat Segunung pada pengujian di rumah kaca (Sulandari et al. 2006). Demikian pula Brown & Nelson (1988) melaporkan terdapat beberapa spesies tanaman termasuk gulma, famili Solanaceae, Leguminosae, Malvaceae dan Asclepiadaceae yang merupakan inang TYLCV di Meksiko.
Patogenesis dan Gejala Infeksi
Sumber inokulum geminivirus berada pada tanaman inang, sisa-sisa tanaman dan inang alternatif. Secara alamiah geminivirus dapat sampai ke tanaman atau berpindah dari satu tanaman ke tanaman lainnya apabila ada serangga vektor kutukebul (B. tabaci Genn) karena virus ini tidak ditularkan melalui biji ataupun secara mekanik. Setelah virus sampai pada inang yang sesuai maka virus akan melepaskan selubung protein kemudian memanfaatkan DNA tanaman untuk bereplikasi kemudian berpindah dari satu sel ke sel lainnya mengikuti aliran nutrisi dan air tumbuhan sehingga gejala akan bersifat sistemik (Hull 2002).
Gejala yang ditimbulkan geminivirus bervariasi tergantung pada strain virus, jenis tanaman, fase pertumbuhan tanaman dan beberapa faktor lainnya. Gejala infeksi virus pada cabai, berupa daun menggulung, penebalan tulang daun, bercak-bercak klorotik pada daun, klorosis di antara tulang daun, malformasi daun, belang dan menguning (Lotrakul et al. 2000). Menurut Sulandari (2006)
gejala awal yang ditimbulkan pada daun cabai rawit maupun cabai besar berupa penjernihan tulang daun (vein clearing) yang kemudian berkembang menjadi warna kuning, penebalan tulang daun, dan penggulungan daun (cupping). Infeksi lanjut geminivirus menyebabkan daun-daun mengecil, berwarna kuning cerah dan tanaman menjadi kerdil. A. conyzoides yang terinfeksi geminivirus menunjukkan gejala vein clearing atau penjernihan tulang daun (Sukamto et al. 2005). Menurut Haerani & Hidayat (2003) infeksi geminivirus menghasilkan gejala yang beragam pada studi penularan geminivirus asal A. conyzoides terhadap beberapa tanaman Solanaceae (tomat, tembakau dan cabai rawit). Daun tanaman tomat yang terinfeksi mengeriting ke arah bawah, tulang daun menebal, dan tangkai daun melengkung ke bawah. Tanaman tembakau yang terinfeksi menunjukkan gejala berupa daun yang menggulung terutama daun muda, tulang daun menebal dan daun melengkung kearah bawah. Tanaman cabai rawit yang terinfeksi menunjukkan gejala daun yang melepuh, tulang daun menebal dan daun melengkung ke atas.
Gulma yang terinfeksi geminivirus menunjukkan gejala yang bervariasi tetapi gejala yang banyak ditemukan pada gulma adalah penguningan tulang daun (netting). Gejala infeksi geminivirus pada A. conyzoides adalah penjernihan tulang daun atau vein clearing (Sukamto et al. 2005), tulang daun menebal, daun melengkung ke bawah seperti mangkuk terbalik dan pertumbuhan tanaman terhambat (Haerani & Hidayat 2003). M. coromandelianum yang terinfeksi geminivirus di Cina menunjukkan gejala daun keriting, tulang daun menebal dan pertumbuhan terhambat (Wu et al. 2007). Gejala infeksi geminivirus berupa
mosaik kuning pada tulang daun ditemukan pada gulma Croton bonplandianum, A. conyzoides, U. lobata, S. acuminate, S. Rhombifolia, E. alba, A. esculentus di India Timur (Chatterjee et al. 2007).
Penularan Geminivirus
Kajian mengenai penularan geminivirus telah banyak dilakukan oleh para peneliti. Hal ini disebabkan informasi tersebut sangat penting dalam epidemiologi penyakit yang disebabkan oleh geminivirus sebagai salah satu patogen penting yang telah menyebabkan epidemi pada tanaman cabai. Brown & Nelson (1988) melaporkan bahwa penyakit pada tanaman tomat di Meksiko yang disebabkan
oleh geminivirus tidak dapat ditularkan secara mekanik dengan cairan perasan, tetapi melalui serangga vektor. Hal ini didukung juga oleh Hull (2002) yang menyatakan bahwa geminivirus tidak dapat menular secara mekanik atau pun melalui benih, hanya dapat menular dengan cara penyambungan tanaman sakit pada tanaman sehat serta melalui serangga vektor B. tabaci.
Penularan di alam secara alami diketahui hanya melalui vektor B. tabaci sehingga perannya menjadi sangat penting dalam penyebaran geminivirus. Berdasarkan hasil penelitian Rusli et al. (2000) geminivirus isolat Segunung yang ditularkan secara mekanik pada tanaman cabai besar dan cabai rawit tidak menghasilkan gejala sedangkan penularan dengan cara penyambungan menghasilkan gejala dengan kejadian penyakit sebesar 71,4% pada tanaman cabai besar dan 57,1% pada cabai rawit. Keefektifan penularan tertinggi diperoleh pada perlakuan penularan melalui serangga vektor yaitu mencapai 70-80%.
Hubungan antara B. tabaci dan geminivirus berdasarkan lamanya virus bertahan pada vektor bersifat persisten sirkulatif non propagatif yaitu virus tersebut berada dalam tubuh serangga untuk kemudian akan ditularkan pada tanaman sehat melalui proses makan (Harrison & Robinson 1999). Efisiensi penularan geminivirus dengan B. tabaci melalui proses makan sangat dipengaruhi oleh lamanya masa akuisisi serangga tersebut, selain oleh jumlah serangga yang menularkan geminivirus pada tanaman sehat (Rachmawati 2003). Periode akuisisi minimum B. tabaci untuk menularkan TYLCV adalah selama 15 menit dan terus meningkat hingga mencapai tingkat maksimum setelah akuisisi selama 24 jam (Mehta et al.1994). Menurut Aidawati (2006) B. tabaci yang melalui periode makan akuisisi (PMA) dan periode makan inokulasi (PMI) masing-masing selama 15 menit mampu menularkan geminivirus walaupun dengan efisiensi penularan yang berbeda-beda untuk tiap kombinasi biotipe B. tabaci dan strain geminivirus yang berbeda. Pada PMA dan PMI tiga dan enam jam strain geminivirus isolat Bogor menghasilkan efisiensi penularan 80 − 100% dengan masa inkubasi 9 hari. B. tabaci termasuk ke dalam ordo Hemiptera dengan famili Aleyrodidae (Henneberry & Castle 2001). Serangga ini memiliki kisaran inang meliputi berbagai tanaman budidaya dan gulma, dapat berkembang dengan baik di daerah tropis dan subtropis (Kalshoven 1981). Menurut Henneberry & Castle (2001) ada
500 jenis tanaman yang dapat menjadi inang B. tabaci dengan preferensi yang berbeda. Salah satu faktor yang mempengaruhinya adalah permukaan daun, serangga tersebut umumnya memiliki preferensi yang tinggi pada permukaan daun yang berambut (hirsute) dibandingkan dengan daun yang permukaannya tidak berambut (glabrous). Basu (1995) melaporkan ada 540 spesies dari 77 famili tanaman yang dapat menjadi inang B. tabaci. Sementara itu, Bezerra et al. (2004) menyatakan bahwa Basu (1995) hanya melaporkan tanaman inang B. tabaci yang bersifat komersial sehingga masih diperlukan penelitian terhadap tumbuhan non komersial yaitu gulma. Menurut Bezerra et al. (2004) gulma Acanthospernum hispidum paling banyak terinfestasi B. tabaci pada lahan tomat di daerah Brazil dibandingkan dengan Amaranthus reflexus, Datura stramonium dan Euphorbia heterophylla.
Deteksi dan Identifikasi Geminivirus
Deteksi geminivirus secara konvensional dapat dilakukan antara lain melalui pengamatan gejala, uji penularan dengan vektor dan pengamatan partikel dengan mikroskop elektron. Teknik deteksi terhadap virus semakin berkembang seiring dengan berkembangnya metode biologi molekuler. Salah satu teknik tersebut yaitu Polymerase chain reaction (PCR). Menurut Wahyudi (2000) PCR merupakan cara yang sangat peka untuk melakukan deteksi dan identifikasi patogen tanaman. Teknik ini memberikan beberapa kelebihan bila dibandingkan cara konvensional, antara lain tidak diperlukan pembiakan patogen pada media. Hal tersebut sangat menguntungkan untuk patogen yang belum dapat dibiakkan secara in vitro seperti virus.
Teknik PCR banyak digunakan untuk mendeteksi geminivirus pada berbagai tanaman budidaya dan juga gulma. Hidayat et al. (1999) telah berhasil mendeteksi geminivirus asal cabai dari beberapa daerah di Bogor melalui teknik PCR dan restriction fragment length polymorphism (RFLP). Teknik ini juga digunakan Sudiono et al. (2004) untuk mendeteksi geminivirus pada cabai asal Sumatera. Sulandari et al. (2006) menggunakan teknik PCR untuk mendeteksi geminivirus dari enam famili tumbuhan yang terdiri dari 30 jenis tanaman dan gulma. Hallan et al. (1998) berhasil mendeteksi geminivirus pada gulma A. conyzoides, C.
bonpladianum, dan M. coromandelianum yang umumnya tumbuh di daerah pertanian Lucknow, India. Geminivirus pada gulma Sida spp, M. lathyroides dan W. amplissima yang berasal dari Jamaica berhasil dideteksi dan diidentifikasi dengan menggunakan teknik PCR yang dikombinasikan dengan teknik RFLP.
Peran Gulma sebagai Sumber Infeksi
Ekologi Gulma
Gulma adalah tumbuhan selain yang dibudidayakan, kehadirannya tidak diinginkan pada lahan pertanian karena dapat menurunkan hasil yang bisa dicapai oleh tanaman budidaya. Batasan gulma bersifat teknis dan plastis. Teknis, karena berkait dengan proses produksi suatu tanaman pertanian. Plastis, karena batasan ini tidak mengikat suatu spesies tumbuhan dengan pengertian bahwa pada tingkat tertentu tanaman budidaya dapat menjadi gulma sebaliknya tumbuhan yang biasanya dianggap gulma dapat pula dianggap tidak mengganggu (Sastroutomo 1990).
Ekologi gulma dalam pertanian berkaitan dengan hubungan populasi gulma dan tanaman budidaya. Perubahan populasi gulma di suatu tempat dipengaruhi oleh beberapa faktor diantaranya, jasad pengganggu pada gulma, karakteristik gulma, faktor lingkungan dan aktivitas manusia (Tjitrosoedirdjo 1984). Pada ekosistem pertanian aktivitas manusia sangat berpengaruh terhadap jumlah dan keanekaragaman populasi gulma. Gulma juga dapat memberikan pengaruh negatif terhadap tanaman budidaya walaupun di pihak lain gulma merupakan suatu komponen yang dapat menjaga keseimbangan alami suatu ekosistem.
Kehadiran gulma pada lahan pertanian sulit untuk dihindarkan. Pertumbuhan gulma dan luas penyebarannya di suatu daerah sangat dipengaruhi oleh keadaan lingkungan, teknik budidaya, dan jenis tanaman yang dibudidayakan. Berdasarkan hasil survei Everaarts (1981) ditemukan ada 35 jenis gulma yang terbagi kedalam 13 familia di sekitar pertanaman sayuran di wilayah dataran tinggi di pulau Jawa, di antaranya adalah Amaranthaceae (Alternanthera philoxeroides, Amaranthus lividus), Caryophyllaceae (Cerastium glomeratum, Drymaria cordata, Drymaria villosa, Spergula arvensis, Stellaria media),
Commelinaceae (C. diffusa), Compositae (A. conyzoides, A. houstonianum, Artemisia vulgaris, G. parviflora, G. quadriradiata, Melampodium perfoliatum), Cruciferae (Capsella bursa-pastoris, Cardamine hirsuta), Fumariaceae (Fumaria rostellata), Oxalidaceae (Oxalis corniculata, O. corymbosa, O. latifolia), Polygonaceae (Polygonum longisetum, P. nepalense, P. persicaria), Portulacaceae (Portulaca oleracea), Rubiaceae (Borreria alata, Richardia brasiliensis), Solanaceae (Datura stramonium, Nicandra physalodes), Cyperaceae (Cyperus rotundus), dan Gramineae (Cynodon dactylon, Digitaria ciliaris, Eleusine indica, Panicum repens, Paspalum paspaloides, Poa annua). Menurut Sembiring (2007) gulma yang ditemukan pada pertanaman cabai di Bengkulu yaitu A. conyzoides, Borreria alata, B. laevis, Cleome rutidosperma, Croton hirtus, Callopogonium muconoides, Mikania micrantha, Mimosa invisa, Oxalis barrelieli, Phyllanthus niruri, Physalis angulata, Stachytarpeta indica, Spigelia anthelmia, Synedrella nodiflora, Axonophus compressus, Eleusine indica, dan Eragnotis tenela.
Identifikasi Gulma
Identifikasi gulma berkaitan dengan pengenalan spesies-spesies gulma berdasarkan sifat-sifat morfologi baik vegetatif maupun generatif. Identifikasi ini dapat menjadi salah satu dasar dalam pengelompokan atau klasifikasi gulma. Salah satu contoh pengelompokan berdasarkan morfologi yang berhubungan dengan pengelolaan gulma adalah berdasarkan jenis daun yaitu berdaun sempit dan berdaun lebar. Gulma berdaun sempit terdiri dari rumput-rumputan dan teki-tekian sedangkan gulma berdaun lebar pada umumnya adalah tumbuhan berkeping dua (dikotil) dan beberapa berkeping satu (monokotil) dengan daun lebar. Ketiga kelompok gulma ini memiliki karakteristik tersendiri yang memerlukan strategi khusus untuk mengendalikannya.
Menurut Everaarts (1981) dalam mengidentifikasi gulma dapat ditempuh satu atau kombinasi dari sebagian atau seluruh cara-cara seperti, (1) membandingkan gulma tersebut dengan material yang telah diidentifikasi di herbarium, (2) konsultasi langsung dengan para ahli dibidang yang bersangkutan, (3) mencari sendiri melalui kunci identifikasi, (4) membandingkan dengan
determinasi yang telah ada, dan (5) membandingkan dengan ilustrasi yang tersedia.
Peranan Gulma sebagai Inang Alternatif Virus
Gulma sebagai salah satu komponen ekosistem pertanian memiliki pengaruh negatif terhadap tanaman pertanian baik secara langsung maupun tidak langsung. Adanya kompetisi merupakan pengaruh langsung dari keberadaan gulma sedangkan pengaruh tidak langsung adalah peranannya sebagai inang alternatif beberapa patogen (Satroutomo 1990).
Gulma dapat menjadi sumber inokulum penting dalam penyebaran geminivirus terutama apabila ada kesesuaian interaksinya dengan B. tabaci sebagai vektor. Dalam kaitannya dengan penyakit keriting kuning pada cabai, beberapa jenis gulma telah ditemukan dapat menjadi inang alternatif geminivirus. Berdasarkan hal tersebut maka salah satu pengendalian yang dapat dilakukan adalah sanitasi lahan dari gulma dan tanaman volunteer dengan tujuan untuk mengurangi sumber inokulum yang dikombinasikan dengan pengendalian terhadap serangga vektor (Nakhla et al. 1998). Sanitasi gulma dari lahan pertanian juga tidak selamanya memberikan dampak positif terhadap tanaman budidaya, bahkan dapat meningkatkan infeksi suatu penyakit tanaman. Menurut Eberwine (1995) pengendalian gulma johnsongrass yang berperan sebagai inang alternatif maize dwarf mosaic virus pada tanaman jagung baik secara mekanik maupun kimiawi dapat meningkatkan keparahan penyakit maize dwarf mosaic.
Kelompok gulma berdaun lebar dari golongan dikotil sering ditemukan di pertanaman budidaya. Gulma dari kelompok ini memiliki potensi sebagai sumber inokulum geminivirus khususnya Begomovirus (Dhar & Singh 1996; Harison & Robinson 1999; Sukamto et al. 2005).
Pengendalian Penyakit yang Disebabkan Geminivirus
Sampai saat ini belum ditemukan bahan kimia atau cara fisik yang dapat mematikan atau menginaktifkan geminivirus dalam tanaman tanpa mempengaruhi kehidupan tanaman itu sendiri. Oleh karena itu, pengendalian penyakit ini bukan ditujukan untuk menyembuhkan tanaman yang terinfeksi, namun lebih
mengutamakan pada pengelolaan ekosistem yang dapat mencegah dan mengurangi terjadinya infeksi virus pada pertanaman lainnya. Secara alamiah geminivirus tidak menular melalui benih tetapi hanya menular dengan bantuan serangga vektor B. tabaci dari tanaman satu ke tanaman lainnya. Karena itu, pengendalian serangga vektor dan inang alternatif merupakan kunci dalam mengendalikan geminivirus. Inang alternatif dapat menjadi sumber inokulum untuk perkembangan penyakit yang disebabkan oleh geminivirus (Nakhla et al. 1998). Gulma merupakan salah satu inang alternatif yang harus diwaspadai karena kehadirannya pada setiap pertanaman budidaya sulit untuk dihindarkan.