L
Journal of Experimental Marine Biology and Ecology 248 (2000) 35–52
www.elsevier.nl / locate / jembe
Underground mating in the fiddler crab Uca tetragonon: the
association between female life history traits and male
mating tactics
a ,* b c d
Tsunenori Koga , Minoru Murai , Seiji Goshima , Sombat Poovachiranon
a
Department of Biology, Faculty of Science, Kyushu University, Fukuoka 812-8581, Japan
b
Tropical Biosphere Research Center, Sesoko Station, University of the Ryukyus, Okinawa 905-0227,
Japan
c
Department of Biology and Aquaculture, Faculty of Fisheries, Hokkaido University, Hakodate 041-0821,
Japan
d
Phuket Marine Biological Center, P.O. Box 60, Phuket 83000, Thailand
Received 25 April 1998; received in revised form 6 January 2000; accepted 13 January 2000
Abstract
Brood size and other life-history traits of females affect male investment in mating. Female Uca
tetragonon, producing relatively small broods, were attracted to the burrows of males for
underground mating (UM) while carrying eggs. Most UM females released larvae and ovulated new broods during the pairing, averaging 3.9 days. While a female was incubating one brood, another brood was developing within the ovaries because the females were feeding adequately during incubation. These findings suggest that in U. tetragonon, a small-brood species, females increase the total number of broods produced by breeding continually. In contrast, in large-brood species, feeding by ovigerous females is relatively brief and not enough to prepare the next brood during incubation, inducing temporal separation between incubation and brood production. Unlike females in other ocypodids where females with large broods remain in the breeding burrows of males, most female U. tetragonon left the male after UM. Wandering in female U. tetragonon after the pairs separate may occur because their small broods are adequately protected by an abdominal flap. Relative brood size probably determines the vulnerability of the incubated broods to the females’ surface behavior. Hence, male reproductive success in large-brood species may decrease greatly if males expel their mates after ovulation, although this is not necessarily so in small-brood species. Whether the male drives away the female or not may depend on which behavior within either small- or large-brood species yields the greater male reproductive success. In U. tetragonon some females extruded eggs in their own burrows after surface mating as well as in males’
*Corresponding author. Present address: Ecosystem Study Laboratory, Environmental Biology Division, National Institute for Environmental Studies, 16-2 Onogawa, Tsukuba, Ibaraki 305-0053, Japan. Tel.: 181-298-50-2470; fax: 181-298-50-2493.
E-mail address: [email protected] (T. Koga)
burrows after UM. It was unclear whether females chose a male with a larger burrow as an UM mate unlike several large-brood species. Burrows of both UM males and ovigerous females in U.
tetragonon were relatively smaller than those in some large-brood species, indicating that
incubation of small broods does not require large burrows. Rather than benefits of UM by female choice, wandering resulting from intersexual conflict, and sperm competition may explain why some females mate in males’ burrows in this small-brood species. 2000 Elsevier Science B.V. All rights reserved.
Keywords: Life history traits; Mating tactic; Brood size; Fiddler crab; Underground mating; Burrow size
1. Introduction
In ocypodid crabs, relative brood size, one of the main traits in female life history, has been used to explain inter-specific variation in feeding activity of females during incubation, intervals of consecutive brood production and location of mating and incubation (Christy and Salmon, 1984; Salmon, 1987; Henmi, 1989). A trade-off between brood size and reproductive frequency occurs in intertidal ocypodids. Female crabs, including fiddlers, lay eggs that adhere to setae on the pleopods and are retained and brooded in a mass beneath the broad abdomen of the female. Relative brood size determines whether the broods are well protected by abdominal flaps of the females during incubation, which affects the feeding activity of the ovigerous females.
Christy and Salmon (1984) suggested that production of large broods coevolved with underground incubation. Female Uca pugilator, U. lactea, Ilyoplax pusilla and Scopi-mera globosa, with large broods live in upper tidal flats and rarely feed while incubating (Christy, 1978; Murai et al., 1987; Henmi and Kaneto, 1989). These females are probably subjected to heat stress and desiccation when they feed on the surface (Salmon, 1987; Henmi, 1989). In contrast, female U. vocans vomeris and Macrophthalmus japonica with small broods, live in the lower tidal flats and feed actively while incubating (Salmon, 1984; Henmi, 1989). That is, female reproductive patterns in the ocypodids can be considered as follows. In large-brood species, females output a relatively large number of eggs in a single brood but cannot bear broods continuously probably because the possible vulnerability of the large brood to heat stress and desiccation does not allow them to feed enough during their incubation to develop the next brood internally. In contrast, in small-brood species, females output relatively few eggs in each brood but can bear broods continuously probably because the protected small brood allows them to feed enough during incubation to develop the next brood internally (see Henmi, 1989).
Some ocypodid females mate in male-defended burrows (underground mating, UM) or on the substrate surface (surface mating, SM), which has been considered to depend on the relative brood size of females of different species (Christy and Salmon, 1984; Henmi, 1989). Christy and Salmon (1984) suggested that UM generally occurs in large brood species like U. pugilator and SM generally occurs in small-brood species like U.
vocans. In order to breed successfully, female U. pugilator prefer burrows located high
incubation and persist longer than those closer to the water (Christy, 1983). It may be very costly for females to prepare such burrows by themselves. Inter-male competition occurs for access to breeding sites of females, i.e. breeding burrows. In large-brood species, males occupy high locations in tidal flats and prepare burrows there through male–male combats seem to enjoy high mating and reproductive success through female choice (Christy, 1980, 1983; reviewed in Christy, 1987a). In contrast, in small-brood species, competition for burrows does not seem to be important for males to access females probably because females breed in their own burrows and feed during incubation. These females may not need special burrows for incubation (Christy and Salmon, 1984; Salmon, 1984).
However, we need to reconsider mating systems of the ocypodids because some new aspects have been found. For example, U. beebei, U. lactea and S. globosa, large-brood species, perform SM as well as UM and females often breed in their own burrows independent of males and males’ burrows (Christy, 1987b; Murai et al., 1987; Koga, 1998). In this study, we add further information on mating systems of the ocypodids and review them.
Ocypodid species producing small broods may also engage in UM involving postcopulatory guarding which guarantees a high paternity rate by the guarding male, because of last-male precedence in sperm competition in the ocypodids (Diesel, 1991; Koga et al., 1993). Uca tetragonon inhabits on exposed lower intertidal, sandy / or mud-flats on shores fringed with mangrove forest, coral rubble or rock (Frith and Frith, 1977). U. tetragonon, the species we have studied, belong to the same subgenus
Thalassuca (Crane, 1975) as U. vocans which bear relatively small broods, but U.
tetragonon mates underground in the burrows of males (Murai et al., 1995). In one experiment, ovigerous females were invited into males burrows and produced new broods there (Goshima et al., 1996). Thus, the females are expected to have life history traits common to species with relatively small broods, such as U. vocans and M. japonica (Salmon, 1984; Henmi, 1989).
burrow for the next UM and increase male reproductive success. Hence, male ocypodids may invest relatively more (e.g., courtship waving and a burrow) for a single UM in large-brood than in small-brood species (e.g., courtship waving), i.e. relative brood size of female ocypodids may affect male mating tactics.
Large burrows may be suitable for ovulation and incubation of female ocypodids in large-brood species. Male U. lactea and S. globosa possess larger burrows for UM than do females of these species (Goshima and Murai, 1988; Koga and Murai, 1997). Female U. pugilator and U. annulipes choose an UM mate based on features of the males burrows (Christy, 1983; Backwell and Passmore, 1996). Female U. annulipes prefer smaller and more compact burrows, not larger burrows. However, the burrow that is used for mating has to be large enough for the pair to stay together and mate.
In general, we would expect a mating burrow to be larger on average than a burrow used by a single crab. In U. tetragonon, burrow size of UM males may be larger than that of females but females may not choose males for UM based on the size of males’ burrows, if the brood size is small and females feed but do not stay underground during incubation. In small-brood species, females may have relatively smaller burrows during incubation than those in large-brood species, because they don’t need large burrows for incubating small broods. Again, in small-brood species, UM males may simply provide their mates with relatively smaller burrows than in large-brood species.
Female U. tetragonon perform SM (Crane, 1975) and UM (Murai et al., 1995) as do female U. beebei, U. lactea and S. globosa (Christy, 1987b; Murai et al., 1987; Koga and Murai, 1997). In advance of UM, however, U. tetragonon females leave their burrows, wander on the surface and find their UM mates there, which may involve predation risk to the females (Koga et al., 1998). Why do females of this species sometimes bother with UM when they can SM and extrude eggs and incubate in their burrows? They may benefit by choosing UM mates with breeding burrows of high quality that may be difficult for them to make, as in U. pugilator and U. annulipes (Christy, 1983; Backwell and Passmore, 1996). Otherwise, females may be aggressively displaced by males and wander, as in U. lactea and U. rosea (Murai et al., 1987, 1996), presumably increasing the chance for males to attract females into the formers’ burrows for UM. This would guarantee paternity of the males through sperm competition (Diesel, 1991; Koga et al., 1993).
internally; (3) while carrying eggs, most females are attracted to mate in male burrows and new broods are laid shortly after pairing; and (4) most UM females leave the mating burrow after bearing new broods, unlike in other UM species with large broods. Second, we investigate the burrow size of reproductive crabs and show that in U. tetragonon (1) burrows of UM males are larger than those of SM females but, unlike in some large-brood species, female choice of larger burrows is unclear; and (2) burrows of UM males and SM females during incubation are relatively smaller than those of UM males, and of females ovulating in their own burrows after SM in some large-brood species. Third, we mention the possible reasons why female U. tetragonon sometimes mate and breed in males’ burrows while the females can breed in their own burrows after SM.
2. Methods
The study was carried out during daytime low water on a sandy–muddy tidal flat in Ao Tang Khen, Phuket, Thailand (lat 78449N, long 988259E) from September 1990 to March 1991 and from September to December 1995. To ascertain the reproductive conditions of mating females, we dug up 29 UM females shortly after pairing had occurred. Another 36 UM females were captured after the pairs had separated. The sex that left the UM burrow first was recorded. In cases where the male left, the female continued to use the mating burrow during incubation. In cases where the female left, the latter did not use the burrow but wandered and found a new burrow for herself. If there were wandering females with new broods, we assumed that some of them had left the males burrows after mating and ovulation, while others may have been evicted soon after breeding in their own burrows after SM. We randomly captured 144 wandering females, which were brought to the laboratory, and their carapace width (CW) was measured to the nearest 0.1 mm. We classified the crabs as ovigerous (OV) or nonovigerous (NO). For ovigerous females, egg developmental stages were determined microscopically, and placed into one of four categories (Goshima et al., 1996) for each brood. Among females with Stage I eggs, those with eggs comprising only one to four cells, were also recorded.
To determine the breeding frequency and surface activity of individual females, we found and removed OV females from burrows. The measured females of known egg stages (Stages I, II and III) were then marked with unique combinations of color spots on the dorsal surface of their carapace and released into these burrows. The incubation period of this species was 28.3 days (range, 25 to 31 days) under laboratory conditions (Stages I: 8.3 days; II: 9.9 days; III: 5.6 days; and IV: 3.9 days, according to Goshima et al., 1996). On the basis of these results we determined when we should start recapturing the marked females. These marked females were recaptured to check for respawning 24, 10 and 4 days after their initial release when they were active on the surface. If they had not respawned, we repeated such releases and recaptures. These were terminated after a female was found to be carrying a new brood.
every day. We also determined the average duration between UM and pair separation after ovulation (n533).
To determine the developmental stage of eggs within the ovaries, we randomly collected 5 NO and 24 OV females (four, seven, eight and five females with Stage I, II, III and IV eggs, respectively). We measured the diameter of ten eggs per ovary per female by dissecting the ovaries under a microscope. Ovarian weight, relative to body weight was obtained after drying the tissues at 608C for 48 h. Eggs were removed from the pleopods to determine dry brood weight relative to dry body weight.
Female U. tetragonon extrude eggs in the male’s burrow after UM (Goshima et al., 1996; this study) or her own burrow after SM (this study). Larger burrows may be more suitable as female reproductive sites. To compare the size of burrows of UM males and SM females, burrow casts were taken with plaster after UM males had plugged the entrance of their burrows and within several minutes of the completion of SM by resident females. All the females were ovigerous. The maximum length, depth, and width of terminal chamber and entrance, and volume of the casts were measured in the laboratory. The width of the casts was calculated as a square root of the product of length multiplied by the width of the largest part of the terminal chamber of the burrow cast. The width of the narrowest point near the entrance of the casts was measured as the width of the entrance. Some burrow casts of males rejected by females as UM mates were also measured (see below for details).
To test whether females prefer some features for UM, we compared body size (CW) and width of burrow chamber between rejected and chosen males as UM mates (the chamber width was conspicuously wider for UM males than for SM females; see Results). We observed interactions between wandering females and courting males. After the waving display of a male, the female entered and soon left the male’s burrow if she rejected him as an UM partner. If she chose him as an UM partner, she stayed in the burrow and the male plugged the entrance. Five wandering females left one male’s burrow and one female rejected two males’ burrows before choosing an UM mate. For the female who rejected two males, parameters of body size and the chamber width of the rejected males’ burrows were averaged for comparison with those of the chosen male.
The minimum size of females that paired in plugged burrows was 16-mm CW. For the purpose of analysis, we assumed this to be the minimum size for UM.
We compared size of breeding burrows between species with large and small broods. In small-brood species, females may not need large burrows during incubation for their small broods unlike OV females in large-brood species. Then, males may provide their UM mate with relatively small burrows in small-brood species unlike males in large brood species. Female S. globosa enlarge their burrows for breeding after ovulation; the resulting burrows are larger than those at the time of SM (Koga and Murai, 1997; unpublished data).
3. Results
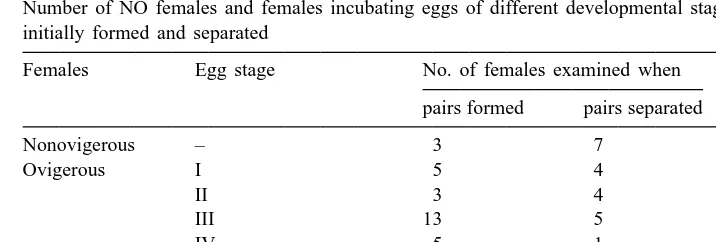
Table 1
Number of NO females and females incubating eggs of different developmental stages, captured when pairs initially formed and separated
Females Egg stage No. of females examined when
pairs formed pairs separated
NO and OV females at various stages of egg development entered male burrows to pair for mating, with 90% of them being ovigerous (Table 1). When they separated from the male, 42% of UM females had new broods (Table 1). The average duration between UM and the separation of the pairs after ovulation was 3.961.8 (S.D.) days, with a range of 1 to 9 days (n533). Since Stages III and IV take 5.6 and 3.9 days on average (see Methods), respectively, females with Stage I or II and some with Stage III eggs when they paired underground did not spawn before the sexes separated. The ovaries of OV females with Stage III and IV eggs were riper than those of the NO females and the other OV females (z52.81, P,0.01 for egg size in ovaries; z52.41, P,0.02 for relative ovary weight; Mann Whitney U-test; Fig. 1). Hence, because there were few NO females among those captured at pairing, most females spawning in the burrows of males were OV females with Stage III or IV eggs.
In 73% of the 15 events of UM followed by ovulation, the female left, and the males remained at the burrow, indicating that the females had begun to wander off just after their ovulation. In 86% of 21 UM events in which the pair separated with no ovulation,
2
the females left the burrow. The difference was not significant (x 50.248, P.0.5). Pooled data demonstrate that females left the burrows in 81% of the 36 UM, significantly more often than the males (P,0.001 by binomial test).
Eighty-five percent of 144 wandering females were ovigerous and 12.5% had newly extruded eggs (one to four cells). These females with new eggs may have been UM females which had recently left mating burrows after ovulation.
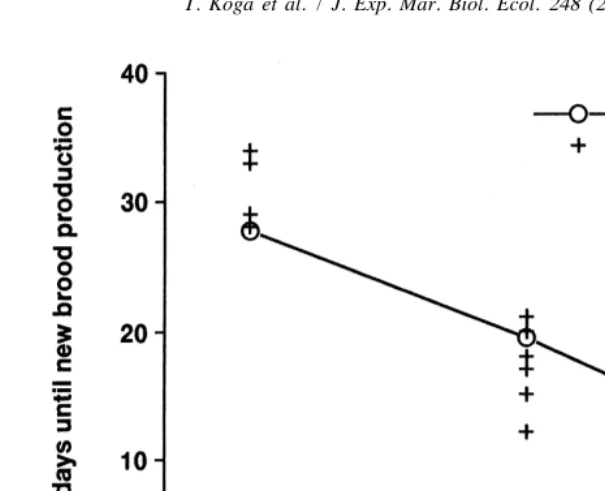
Of eight marked females with Stage I eggs, four were lost and four had Stage I eggs again after 28 to 34 days (mean6S.D.531.062.9, Fig. 2). Six marked females with Stage II eggs and six marked females with Stage III eggs carried new broods after 12 to 21 days (17.263.3) and after 5 to 13 days (7.863.1), respectively (Fig. 2). The average number of days required to develop new broods corresponds approximately to the values expected from the laboratory work of Goshima et al. (1996), indicating that females laid new broods immediately after larval release.
Fig. 1. (a) Mean diameter of eggs in ovaries, and (b) ovary weight relative to body weight; for nonovigerous and ovigerous females carrying Stage I, II, III and IV eggs, respectively. The number ‘3’ in (a) represents three females with identical diameter of the eggs in ovaries (or, one symbol represents three samples).
of 22, 20 out of 28 and 21 out of 22 days. She extruded the second and third broods in her burrow after SM. For these five broods, the females spent 62.4620.7% (mean6S.D.) of the available day foraging during incubation.
Fig. 2. Number of days until next brood production for ovigerous females carrying Stage I, II and III eggs. Laboratory data were based on Goshima et al. (1996) and field data are from the present study.
However, the width of terminal chambers in UM males’ burrows was larger than that for SM females’, which seems to contribute to the difference in volume of the burrows between UM males and SM females (Table 2). In contrast, depth, length and the entrance width of SM females’ burrows were quite similar to those of UM males (Table 2).
There was no significance in CW between rejected and chosen males for UM (Table 3). The width of the chambers in the successful males’ burrows seemed to be larger than
Table 2
a
Body (carapace width, CW) and burrow size of UM males and SM females in U. tetragonon
UM males SM females t P After B / D
(6S.D.) (6S.D.) correction
CW (mm) 20.662.4 19.462.2 1.697 0.097 n.s.
Volume (ml) 76.0627 50.0617 3.839 0.0004 sig.
Width of chamber (mm) 31.264.9 26.264.4 3.562 0.0009 sig.
Depth (mm) 87.2616.8 87.3616.7 20.18 0.986 n.s.
Length (mm) 149.0644.0 136.9622.3 1.150 0.257 n.s.
Width of entrance (mm) 17.362.3 15.962.7 1.869 0.069 n.s.
a
Table 3
Differences in average CW and width of burrow chamber between males rejected and chosen males by female
a
U. tetragonon for UM (chosen2rejected)
Male characters Difference in Paired t P
average
CW (mm) 20.3 0.272 0.797
Chamber width (mm) 3.0 21.905 0.115
a
Sample size was 6.
that in the unsuccessful ones, but the difference was not significant, perhaps because of the small sample size (Table 3).
The average (6S.D.) burrow volume of UM males in U. tetragonon (76627 ml, n522) was significantly smaller than that in U. lactea (105622, n515; z53.016, P50.003, U-test) in spite of the larger body size of the former (20.662.4 mm, n522 for U. tetragonon; 14.061.1, n515 for U. lactea; z55.104, P,0.0001, U-test, Fig. 3a). In analysis of regression of body size (x-axis) on burrow volume ( y-axis), the intercept in S. globosa was significantly larger than that in U. tetragonon (F1,505 550.913, P,0.0001, ANCOVA, Systat 5.2), although the slope in S. globosa was not significantly different from that in U. tetragonon (F1,4950.046, P50.830, Fig. 3a), indicating that burrow size of UM males in U. tetragonon was relatively smaller than that in S. globosa. The average (6S.D.) burrow size of OV females after SM in U.
tetragonon (50617 ml, n522) was not significantly different from that in U. lactea
(45623, n513) and S. globosa (3567, n56; F2,3851.514, P50.233, ANOVA), although body size of the females in U. tetragonon (19.462.2 mm, n522) was much larger than that in U. lactea (12.261.3, n513) and S. globosa (10.860.6, n56; F2,38590.142, P,0.0001, ANOVA, Fig. 3b), indicating that burrow size of females in U. tetragonon was relatively smaller than that in U. lactea and S. globosa.
4. Discussion
Fig. 3. Relationships between body size (carapace width) and burrow size (volume) for reproductive crabs in
Scopimera globosa, Uca lactea and U. tetragonon. (a) Males engaging in underground mating (UM) in their burrows. S. globosa: y56.85x252.43, n530, r50.803, P,0.0001; U. tetragonon: y57.36x275.58,
of both sexes in U. tetragonon were relatively smaller than those in some large-brood species.
4.1. Continuous or discontinuous incubation and feeding by OV females
In ocypodid crabs, relatively small broods, which are protected and not visible externally, probably allow the females to feed adequately during incubation to be able to produce their next broods consecutively (Christy and Salmon, 1984; Salmon, 1987; Henmi, 1989). A complementary factor for continuous breeding in the small-brood species is that the females would need much less food for bearing small broods, and then, perhaps they need to feed no more intensely than females in the large-brood species. The relative egg weight to body weight was 8% for U. tetragonon, much smaller than that in other UM species. Relative brood size of U. lactea, I. pusilla and S. globosa is more than 30% (Henmi, 1989; Henmi, unpublished data). Small broods are protected by the abdominal flap until late in the incubation period, while eggs from relatively large broods are exposed as soon as they are laid (Christy and Salmon, 1984). While U. tetragonon incubated one brood, another brood was developing within the ovaries (Fig. 1). Most UM females released larvae and extruded a new brood of eggs onto their pleopods within a few days of pairing. Large females of M. japonica produce small broods every two weeks (Y. Henmi, personal communication). Female M.
japonica and U. vocans vomeris carry broods for 2 weeks (Salmon, 1984; Henmi and
Kaneto, 1989). Female U. tetragonon, for reasons as yet unknown, carry broods for 4 weeks (see also Christy and Salmon, 1984; Murai et al., 1987).
The trade-off between reproductive frequency and relative brood size has been observed in a variety of animals (Stearns, 1992; Andersson, 1994). Field studies on birds have shown that an experimental increase in brood size delays reproduction the following year (Godfray et al., 1991; Lessells, 1991). Heaney and Monagham (1996) also showed in common terns that an experimental increase in brood size delays the production of the next brood even within a reproductive season. Reaka (1979) stated that stomatopods exhibit a trade-off between reproductive frequency and the relative size of the brood, to the extent that large species produced frequent but proportionately small broods, while small species produced infrequent but large broods relative to their body size. Female U. tetragonon and U. vomeris produce relatively small broods every 4 and 2 weeks, respectively, while in species with large broods, there is a temporal separation between consecutive breeding phases (Table 4). Continuous breeding is considered to be very adaptive for crabs bearing small broods with long incubation periods.
Most female U. tetragonon were attracted to male burrows for UM when carrying eggs (Table 1). Since all females are almost always breeding, NO females were few. In contrast, in species with large broods, females remain underground for incubation lasting 2 weeks. Since female U. beebei, U. lactea, S. globosa and U. annulipes feed actively on the surface after their eggs hatch, only NO females of these species are attracted to courting males (Table 4, Backwell and Passmore, 1996).
Koga
et
al
.
/
J.
Exp
.
Mar
.
Biol
.
Ecol
.
248
(2000
)
35
–
52
47
Table 4
Reproductive characteristics of female ocypodid crabs
Species Relative Temporal Feeding Sex that leaves Reference
clutch separation during burrow after
a
size between clutches incubation UM
Uca pugilator Large Long No Male Christy and Salmon, 1984
U. lactea Large Long No Male Murai et al., 1987
U. beebei Large Long Brief Male Christy, 1987, unpubl., pers. comm.
U. perplexa Large Long Brief Male Nakasone and Okadome, 1981; Nakasone and Murai, 1998
U. thayeri Large Long Usual – Salmon, 1987
Ilyoplax pusilla Large Long No Male Wada, 1981; Henmi, 1989; Henmi and Kaneto, 1989
I. gangetica Large Long No Male Kosuge et al., 1994a
Scopimera globosa Large Long Rare Male Henmi and Kaneto, 1989; Koga, 1995, pers. obs.
U. vocans vomeris Small Brief Usual – Salmon, 1984
U. vocans vocans Small ? Usual ? Nakasone et al., 1983
Macrophthalmus japonica Small Brief Usual – Henmi, 1989; Henmi and Kaneto, 1989; Henmi, pers. comm.
U. tetragonon Small Brief Usual Female Murai et al., 1995; Goshima et al., 1996; present study
a
globosa, rarely feed while incubating (Table 4). Some OV females of U. beebei and U. perplexa, with large broods, feed while ovigerous, but this is restricted to a brief feeding period (Table 4). Most species bearing large broods live in the upper tidal flats and are probably subjected to heat stress and desiccation, which restricts feeding of OV females on the surface (Salmon, 1987; Henmi, 1989).
4.2. Relative brood size and male mating tactics in response to the activity of OV females
Relatively large broods seem to partly restrict ocypodid females from feeding during incubation, even in low tidal flats. I. gangetica, which produces relatively large broods, occupies lower intertidal silty mudflats (Kosuge et al., 1994a). Although this habitat is neither hot nor dry, unlike the upper tidal flats during low tides, the females remain in UM burrows as do other species with relatively large broods living on upper tidal flats. This suggests that crabs with large broods may be susceptible to physical loss of eggs during wandering, regardless of whether females live in a benign environment. In contrast, although female U. thayeri in lower intertidal mudflats produce large broods, OV females in this species feed on the surface during low tides in Florida (Salmon, 1987). Why do female U. thayeri feed during incubation? The U. thayeri population in Florida studied by Salmon had a lower density than that of the population of this species in Trinidad (von Hagen, 1973, cited in Salmon, 1987). The risk of eviction for OV females during surface feeding may therefore be smaller, because the lower density implies a lower rate of interaction [However, virtually nothing is known regarding female breeding characteristics in the Trinidad population; e.g. whether the females feed on the surface during incubation (Salmon, 1987)]. These findings suggest that OV females with large broods holding their own burrows can feed if there is a low risk of eviction. However, Salmon (1987) suggested that females need 4 weeks between breeding bouts, probably because the feeding sallies of OV females are brief. A low risk of eviction may lead to some feeding, but not enough for sequential brood production without a pause between breeding bouts. Carrying large broods may restrict movement on the surface both in the lower and the upper intertidal zone. However, we know very little about the possible causes of egg loss in females incubating on the surface, or about other kinds of selection acting on females and eggs.
bearing relatively small broods with their mating burrows after the females ovulate, because OV females with small broods usually feed on the surface and can endure wandering. Keeping rather than leaving his burrow after ovulation may be adaptive for UM males in small-brood species because they do not have to obtain a new burrow for the next UM.
We argue that male ocypodids invest in an UM event to a different degree between large- or small-brood species in order to yield greater reproductive success. Burrows of UM males in U. lactea and S. globosa bearing large broods were relatively larger than those in U. tetragonon bearing small broods (Fig. 3a), indicating that male U. lactea and S. globosa provided their UM mates with larger burrows than did male U. tetragonon. Some days after expelling the OV females, some U. tetragonon males succeed in inviting a new female into the same burrows for UM (Murai et al., in preparation). However, some male ocypodids in large-brood species like U. pugilator seem to decrease the costs of UM: the male does not leave and find a new burrow after ovulation of his mate but gives a chamber to each mate and continues to stay in his burrow (Christy, 1982).
4.3. Mating modes and sexual conflicts
In most UM in the ocypodids, wandering females enter burrows of courting males (Table 4) and some females choose male mates depending on features of the burrows in which the females extrude eggs and incubate (Christy, 1983; Backwell and Passmore, 1996). This behavior requires females to leave their own burrows and wander on the surface to find males and burrows for UM, which may involve a greater predation risk than SM (Koga et al., 1998).
In some ocypodids, females extrude eggs in their own burrows after SM or in males’ burrows after UM (Christy, 1987a, 1987b; Murai et al., 1987; Koga, 1998; present study). Why do female ocypodids bother with UM when they can perform SM and extrude eggs and incubate in their own burrows? There are three possible reasons: conspicuous benefits of UM for males, a result of eviction from their burrows by males and potential benefits of UM for females. In some ocypodids, both males and females appear capable of multiple mating (Murai et al., 1987; Henmi et al., 1993; present study), but only sperm of the last male to mate fertilizes eggs extruded by a female (Diesel, 1991; Koga et al., 1993). Hence, UM involving postcopulatory guarding of the female mate until her ovulation guarantees paternity of the mating male, but SM not involving guarding has no such guarantee (Koga et al., 1993).
small, from 0 to about 6 encounters for U. musica, and less for U. beebei (most males get no visit and a few get several visits) (John Christy, personal communication). These low rates suggest that males may benefit by ousting neighboring females; they then have a chance to attract them to their burrows. In the present study, one U. tetragonon female was observed to enter and ovulate in the burrow of the male that evicted her before the mating.
Burrows with a large space may help females to protect their broods during ovulation and incubation, especially in large-brood species. In U. lactea and S. globosa, the burrow volume of UM males is relatively larger than in U. tetragonon (Fig. 3a). Female S. globosa enlarge their burrows after ovulation when they extrude eggs there (Koga, unpublished data) and burrows of female U. lactea extruding eggs in their own burrows are relatively larger than those of NO females (Goshima and Murai, 1988, unpublished data). These features indicate that during incubation, female U. lactea and S. globosa need large burrows that may provide special microenvironments for incubation. Hence, in large-brood species, females may benefit by choosing an UM mate based on the male’s burrow size (Christy, 1983; Backwell and Passmore, 1996), although females suffer from wandering in advance of UM. Females may enlarge their burrows to the minimum extent that they need when they ovulate in their own burrows, while they may prefer a male that has the largest burrow among the sampled males when females ovulate in males’ burrows. However, it is unknown whether female S. globosa and U.
lactea prefer the largest burrow when they mate and ovulate in males’ burrows. In U.
tetragonon, a small-brood species, burrow volume of ovigerous females was smaller for their body size than in U. lactea and S. globosa, large-brood species (Fig. 3b).
Female U. tetragonon do not seem to enlarge their burrows after ovulation when they breed in their own burrows. In small-brood species, females may not need large burrows during incubation, because the brood is small and females do not incubate underground unlike females in large-brood species, and feed on the surface during incubation. Besides, it is unclear whether female U. tetragonon choose an UM mate based on the male’s burrow size (Table 3), unlike U. pugilator and U. annulipes (Christy, 1983; Backwell and Passmore, 1996). It is unlikely that female U. tetragonon would benefit by choosing an UM mate based on this criterion. The evidence suggest that whether female ocypodids choose UM mates based on the males’ burrow size may depend on the relative brood size. In U. tetragonon, therefore, females may mate in male-defended burrows not for potential benefits of UM to the females, but as a result of wandering following intersexual conflicts and the clear benefits of UM for males through sperm competition.
Acknowledgements
English used in this manuscript was revised by K. Miller, M.D. Jennions and Egawa Language and Scientific Service. This study was supported by a Grant-in-Aid for International Scientific Research from the Japanese Ministry of Education, Science and Culture (Nos. 01041069 and 06041082). [RW]
References
Andersson, M., 1994. In: Sexual Selection, Princeton Univ. Press, Princeton, p. 599.
Backwell, P.R.Y., Passmore, N.I., 1996. Time constraints and multiple choice criteria in the sampling behavior and mate choice of the fiddler crab, Uca annulipes. Behav. Ecol. Sociobiol. 38, 407–416.
Christy, J.H., 1978. Adaptive significance of reproductive cycles in the fiddler crab Uca pugilator: a hypothesis. Science 199, 453–455.
Christy, J.H., 1980. The mating system of the sand fiddler crab, Uca pugilator. Ph.D. Thesis, Cornell University, Ithaca, New York, pp. 322.
Christy, J.H., 1982. Burrow structure and use in the sand fiddler crab, Uca pugilator (Bosc). Anim. Behav. 30, 687–694.
Christy, J.H., 1983. Female choice in the resource–defense mating system of the sand fiddler crab, Uca
pugilator. Behav. Ecol. Sociobiol. 12, 169–180.
Christy, J.H., 1987a. Competitive mating, mate choice and mating associations of brachyuran crabs. Bull. Mar. Sci. 41, 177–191.
Christy, J.H., 1987b. Female choice and breeding behavior of the fiddler crab Uca beebei. J. Crust. Biol. 7, 624–635.
Christy, J.H., Salmon, M., 1984. Ecology and evolution of mating systems of fiddler crabs (genus Uca). Biol. Rev. 59, 483–509.
Crane, J., 1975. In: Fiddler Crabs of the World. Ocypodidae: Genus Uca, Princeton University Press, Princeton, p. 736.
Diesel, R., 1991. Sperm competition and the evolution of mating behavior in brachyura, with special reference to spider crabs (Decapoda, Majidae). In: Bauer, R.T., Martin, J.W. (Eds.), Crustacean Sexual Biology, Columbia Univ. Press, New York, pp. 145–163.
Frith, D.W., Frith, C.B., 1977. Observation on fiddler crabs (Ocypodidae: genus Uca) on Surin Island, western peninsular Thailand, with particular reference to Uca tetragonon (Herbst). Phuket Mar. Biol. Cent. Res. Bull. 18, 1–14.
Godfray, H.C.J., Partridge, L., Harvey, P.H., 1991. Clutch size. Ann. Rev. Ecol. 22, 409–429.
Goshima, S., Murai, M., 1988. Mating investment of male fiddler crabs, Uca lactea. Anim. Behav. 36, 1249–1251.
Goshima, S., Koga, T., Murai, M., 1996. Mate acceptance and guarding by male fiddler crabs Uca tetragonon (Herbst). J. Exp. Mar. Biol. Ecol. 196, 131–142.
Heaney, V., Monagham, P., 1996. Optimal allocation of effort between reproductive phases: the trade-off between incubation costs and subsequent brood rearing capacity. Proc. R. Soc. Lond. B 263, 1719–1724. Henmi, Y., 1989. Reproductive ecology of three ocypodid crabs. II. Incubation sites and egg mortality. Ecol.
Res. 4, 261–269.
Henmi, Y., Kaneto, M., 1989. Reproductive ecology of three ocypodid crabs. I. The influence of activity differences on reproductive traits. Ecol. Res. 4, 17–29.
Henmi, Y., Koga, T., Murai, M., 1993. Mating behavior of the sand bubbler crab Scopimera globosa. J. Crust. Biol. 13, 736–744.
Koga, T., 1995. Movements between microhabitats depending on reproduction and life history in the sand-bubbler crab Scopimera globosa. Mar. Ecol. Prog. Ser. 117, 65–74.
Koga, T., 1998. Reproductive success and two modes of mating in the sand-bubbler crab Scopimera globosa. J. Exp. Mar. Biol. Ecol. 229, 197–207.
Koga, T., Murai, M., 1997. Size-dependent mating behaviours of male sand bubbler crab Scopimera globosa: alternative tactics in the life history. Ethology 103, 578–587.
Koga, T., Backwell, P.R.Y., Jennions, M.D., Christy, J.H., 1998. Elevated predation risk changes mating behaviour and courtship in a fiddler crab. Proc. R. Soc. Lond. B 265, 1385–1390.
Kosuge, T., Murai, M., Poovachiranon, S., 1994a. Breeding cycle and mating behaviour of the tropical ocypodid Ilyoplax gangetica (Kemp 1919) (Crustacea Brachyura). Trop. Zool. 7, 25–34.
Kosuge, T., Poovachiranon, S., Murai, M., 1994b. Male courtship cycles in three species of tropical Ilyoplax crabs (Decapoda, Brachyura, Ocypodidae). Hydrobiologia 285, 93–100.
Lessells, C.M., 1991. The evolution of life histories. In: Krebs, J.R., Davies, N.B. (Eds.), Behavioral Ecology: An Evolutionary Approach, Blackwell, Oxford, pp. 32–68.
Murai, M., Goshima, S., Henmi, Y., 1987. Analysis of the mating system of the fiddler crab, Uca lactea. Anim. Behav. 35, 1334–1342.
Murai, M., Koga, T., Goshima, S., Poovachiranon, S., 1995. Courtship and the evolution of underground mating in Uca tetragonon (Decapoda; Ocypodidae). J. Crust. Biol. 15, 655–658.
Murai, M., Goshima, S., Kawai, K., Yong, H.-S., 1996. Pair formation in the burrows of the fiddler crab Uca
rosea (Decapoda: Ocypodidae). J. Crust. Biol. 16, 522–528.
Nakasone, Y., Okadome, H., 1981. The reproductive cycle and young crabs growth of Uca(celuca) lactea
perplexa. In: Yamaguchi, T. (Ed.), Ecological Studies of Coastal Marine and Freshwater Crabs. Report For
the Grant-in-aid For Co-operative Research, 1978–1980, Ministry of Education.
Nakasone, Y., Akamine, H., Asato, K., 1983. Ecology of the fiddler crab Uca vocans vocans (Linnaeus) (Decapoda: Ocypodidae). II. Relation between the mating system and the drove. Galaxea 2, 119–133. Nakasone, Y., Murai, M., 1998. Mating behavior of Uca lactea perplexa Decapoda: Ocypodidae). J. Crust.
Biol. 18, 70–77.
Reaka, M.L., 1979. The evolutionary ecology of life history patterns in stomatopod crustacean. In: Stancyk, W.E. (Ed.), Reproductive Ecology of Marine Invertebrates, University of South Carolina Press, Columbia, pp. 235–260.
Salmon, M., 1984. The courtship, aggregation and mating system of a ‘primitive’ fiddler crab (Uca vocans: Ocypodidae). Trans. Zool. Soc. Lond. 37, 1–50.
Salmon, M., 1987. On the reproductive behavior of the fiddler crab Uca thayeri, with comparisons to U.
pugilator and U. vocans: evidence for behavioral convergence. J. Crust. Biol. 7, 25–44. Stearns, S.C., 1992. In: The Evolution of Life Histories, Oxford University Press, Oxford, p. 249. Wada, K., 1981. Growth, breeding, and recruitment in Scopimera globosa and Ilyoplax pusillus (Crustacea: