L
Journal of Experimental Marine Biology and Ecology 248 (2000) 79–104
www.elsevier.nl / locate / jembe
Drifting algal mats as an alternative habitat for benthic
invertebrates:
Species specific responses to a transient resource
a,b ,* a b
Joanna Norkko , Erik Bonsdorff , Alf Norkko
a ˚ ˚
¨
Huso Biological Station and Department of Biology, Abo Akademi University, FIN-20520 Abo, Finland b
National Institute of Water and Atmospheric Research, P.O. Box 11-115, Hamilton, New Zealand Received 1 September 1999; received in revised form 13 January 2000; accepted 18 January 2000
Abstract
Patchy occurrences of benthic drift algae (i.e. loose lying macroalgal mats) may increase habitat complexity on normally bare soft bottoms, but at the same time, extensive amounts of drifting algal mats are known to stress the benthic fauna. This paper presents results of the first detailed study of the fauna associated with drift algal mats in the northern Baltic Sea. In order to assess the importance of drifting algae as an alternative habitat for benthic fauna, benthic drift algal mats
˚
were sampled on shallow (2–9 m) sandy soft bottoms in the outer archipelago of the Aland Islands (Finland). Species composition, abundance and biomass of the macrofauna associated with algal mats were recorded. The results show that drifting algae at times can harbour very high abundances of invertebrates (up to 1116 individuals / g algal dryweight), surpassing invertebrate densities recorded in seagrass communities. The algal fauna varied between sites and over time, and factors such as ambient benthic fauna, exposure to wind-wave disturbance, depth, and algal coverage and condition influenced the invertebrate community composition of the algal mats. Abundance increased while individual biomass of the animals decreased over time (summer season; July–October). A series of laboratory experiments were conducted in order to test the ability of a few important benthic species to move up into, and survive in a drifting algal mat.
Macoma balthica, Hydrobia spp., Nereis diversicolor and Bathyporeia pilosa were used in the
experiments, and significant differences in their survival and mobility within drifting algae were recorded. This study shows that benthic species differ significantly in their ability to utilise the algal mats, with mainly opportunistic and mobile taxa such as Hydrobia spp., Chironomidae and Ostracoda benefiting from the algae, whereas infaunal species such as M. balthica and B. pilosa are negatively affected. The occurrence of eutrophication induced drifting macroalgal mats has increased significantly during the last decade in the northern Baltic Sea. Hence, the importance of drifting algae as a stress factor and as an alternative habitat for benthic fauna increases. 2000 Elsevier Science B.V. All rights reserved.
*Corresponding author. Fax: 164-7-856-0151. E-mail address: [email protected] (J. Norkko)
Keywords: Baltic Sea; Benthic macrofauna; Eutrophication; Macroalgae; Oxygen deficiency
1. Introduction
Benthic drift algae may increase habitat complexity on normally bare soft bottoms (Langtry and Jacoby, 1996; Norkko, 1997; Raffaelli et al., 1998), thereby changing the settings for interspecific interactions (Norkko, 1998). High abundances of invertebrates have been recorded in sublittoral accumulations of drifting macroalgae and seagrass, and the highest abundances of macrofauna recorded in marine benthic communities were found in accumulations of algal detritus off the West Coast of the USA (Vetter, 1994, 1995). As opposed to other physical structures which add a three dimensional aspect to benthic habitats, benthic drift algae constitute a highly mobile structure. Although ephemeral both in time and space, benthic drift algae have been shown to function as an alternative habitat for mobile benthic macrofauna, providing food and refuge from predators (Gore et al., 1981; Kulczycki et al., 1981; Virnstein and Carbonara, 1985; Virnstein and Howard, 1987; Holmquist, 1994, 1997; Vetter, 1995, 1998; Tzetlin et al., 1997; Norkko, 1998). High abundances of macrofauna have also been recorded in intertidal (Nicholls et al., 1981; Soulsby et al., 1982; Kirkman and Kendrick, 1997) and pelagic (Lenanton et al., 1982; Kingsford and Choat, 1985; Kingsford, 1992; Shaffer et
´
al., 1995; Ingolfsson, 1998) drift algal mats.
The effects of drift algal mats on infauna of normally bare soft bottoms depend on the spatial and temporal extent of algal coverage. Patchy occurrences of drift algae may be beneficial to the benthic community and increase diversity (Pihl et al., 1996; Norkko, 1997; Raffaelli et al., 1998), but at the same time high amounts of drift algae are known to stress benthic infauna, the result often being an impoverished community with an altered species composition (Nicholls et al., 1981; Soulsby et al., 1982; Thrush, 1986; Hull, 1987; Bonsdorff, 1992; Everett, 1994; Peterson et al., 1994; Bonsdorff et al., 1995; Norkko and Bonsdorff, 1996a,b; Holmquist, 1997). When algae cover extensive areas the habitat is homogenised and oxygen deficiency induced, resulting in decreased diversity in the benthic community. The seasonal and often sudden occurrence of drifting algae will have varying effects on the behaviour, habitat use and survival of benthic species, depending on their functional group, mobility and tolerance to hypoxia (Norkko, 1998). If conditions under the algae are stressful, mobile benthic species could potentially avoid these by moving up into the overlying algae. Alternatively, mobile species could actively colonise patches of drift algae in search of food and refuge from predators and thus benefit from the additional physical structure and food drifting algae provide for the benthos.
Excessive growth of macroalgae is a well-known symptom of eutrophication (Morand and Briand, 1996; Schramm and Nienhuis, 1996; Raffaelli et al., 1998), and due to the increasing eutrophication of the Baltic Sea (Elmgren, 1989; Cederwall and Elmgren, 1990; HELCOM, 1990; Rosenberg et al., 1990; Bonsdorff et al., 1991, 1997; Nehring
¨
¨
10–15 years (Makinen and Aulio, 1986; Kautsky, 1991; Bonsdorff et al. 1997). When these algae are detached from their substratum they are transported to the sublittoral and deposited on relatively shallow (,15 m) soft bottoms as increasing amounts of drifting algae, covering ever increasing areas in late summer (Olafsson, 1988; Bonsdorff, 1992; Norkko, 1997). Drift algae occur both as small patches and as large homogenous mats, at times covering up to several hectares (Norkko and Bonsdorff, 1996b). Small amounts of drifting algae have always been present in this region, but the amounts recorded in recent years in the archipelago area are a new, eutrophication induced phenomenon. The effects of the algae are therefore changing from stochastic to more predictable, and the algae have the potential to significantly alter the structure and function of this benthic system (Norkko, 1997).
No spatial and / or temporal studies of species composition and abundance of the macrofauna associated with benthic drift algal mats have been conducted in the Baltic Sea. The aim of this study was therefore to describe and quantify the invertebrate
˚
macrofauna occurring in benthic drift algal mats in the archipelago of the Aland Islands, northern Baltic Sea. We also experimentally tested differences in response to algal mats in species representing different functional and taxonomic groups. We wanted thus to investigate the potential value of benthic drift algal mats as habitat for benthic fauna. This was accomplished through sampling of algal mats in the field and through a series of laboratory experiments.
2. Methods
2.1. Field study
2.1.1. Study area
The field study was conducted in the western and northwestern parts of the outer ˚
archipelago of the Aland Islands, northern Baltic Sea (608N, 208E, Fig. 1). Nine sites (A–I) were chosen on shallow (2–9 m) sandy bottoms, with exposure to wind-wave disturbance varying from semiexposed to exposed. Oxygen saturation near the bottom is usually high at these sites, but local hypoxia and anoxia have been recorded in connection with extensive occurrences of drifting algal mats (Bonsdorff, 1992). Salinity varies between 5.5 and 6.2‰. The benthic fauna has been extensively studied in this area (see Bonsdorff and Blomqvist, 1993, for a review). At site A the effects of drifting algal mats on infauna have been thoroughly investigated (Bonsdorff, 1992; Norkko and Bonsdorff, 1996a,b; Norkko, 1997) and at sites B, F and H the invertebrate fauna
¨ associated with Zostera marina meadows has been studied (Mattila, 1995; Bostrom and Bonsdorff, 1997).
˚
Biomasses of drifting algal mats recorded in the Aland Island archipelago during 1990–1997 (July–August, the time of maximum occurrences) ranged from 50 to 830 g
2 2
dryweight (dwt) per m , with average values of approximately 220 g dwt per m ¨
Fig. 1. Map of the study area in the northern Baltic Sea (608N, 208E), with study sites A–I.
Pilayella littoralis (L.) Kjellman and Dictyosiphon foeniculaceus (Hudson) Grev. ¨
(.95% of the biomass, Norkko and Bonsdorff, 1996a; Holmstrom, 1998).
2.1.2. Field methods
cylinder was used for each sample and the cylinders were transported to the laboratory in seawater. In the laboratory the algal samples were flushed out of the cylinders and preserved in 4% hexamine [hexamethylenetetramine; (CH ) N ]-buffered formalin.2 6 4
Sieving the samples is not practical in the case of filamentous algae as the animals easily get trapped in the algae. The algae were sorted under a dissection microscope at 153
magnification and species composition, abundance and biomass (wwt) of the algal macrofauna were recorded. Presence of meiofaunal taxa such as Nematoda, Copepoda and Turbellaria was noted. Also for each sample, the dominant algal species and state of decomposition (condition) of the algae were noted, and their dwt estimated (24 h at 608C). The results are subsequently reported as abundance or biomass per g algal dwt. Number of species is given per replicate sample.
As drifting algal mats occur on shallow sandy soft bottoms, we wanted to compare the algal fauna with the resident benthic fauna of these sediments. Five to six replicate
2
benthic core samples (diameter 4.7 cm, area sampled 17.3 cm , depth 10 cm) were obtained from seven of the sites (excluding sites C and E) at the same time as the algal samples. The benthic samples were obtained from bare sand at least 10 m away from the nearest algal mat. Samples were preserved in 4% hexamine-buffered formalin, washed on a 0.5 mm mesh sieve and sorted under a dissection microscope at 153 magnifica-tion, and analysed as for the algal fauna. Results for the benthic fauna, including number of species, abundance and biomass, are given per core sample.
On each sampling occasion the algal coverage of the sea-floor at each study site was estimated visually by diving over the site. Algal coverage was then allocated to one of three categories (15small patches, ,25% coverage; 25large patches, 25–75% coverage; 3575–100% coverage). On shallow sandy bottoms drift algae can cover large areas; Norkko and Bonsdorff (1996a,b) estimated the drift algae to cover an area of
¨
2 hectares at site A for a 6 week period in July–August and Holmstrom (1998) reported maximum occurrences of drift algae in August, with algae covering 80% of the sandy bottom at site A.
To facilitate comparison with benthic fauna, abundance of algal fauna was related to the areal coverage of algal mat by estimating the biomass of algal mats. On nine different sampling occasions a core (diameter 18.9 cm) was pushed vertically through the algal mat into the sediment and dry weight of the algae was measured (24 h at 608C). To convert abundances and biomasses of the algal fauna to areal units for comparisons
2
with the benthic fauna a conversion factor of 100 g algae per m was used and then related to the area of the benthic core used. This value was chosen as a conservative value, as it is close to the lower limit of the standard deviation of the actual values
2
recorded (1816104 g dwt / m ).
2.2. Laboratory experiments
2.2.1. Test organisms
invertebrates studied represent different functional groups (Bonsdorff and Blomqvist, 1993; Bonsdorff and Pearson, 1999).
The bivalve Macoma balthica (L.) is an important species in the Baltic Sea ˚
(Segerstrale, 1962; Bonsdorff et al., 1995). It is classified as a semimobile suspension feeder / surface detritivore (Bonsdorff and Blomqvist, 1993) and is tolerant to hypoxia (Dries and Theede, 1974). M. balthica has pelagic larvae and settling coincides both
˚ temporally and spatially with mass occurrences of drifting algal mats in the Aland archipelago (Bonsdorff et al., 1995). Two size classes (3–5 mm and 10–15 mm) of M. balthica were used in the experiments. The mudsnails Hydrobia spp. are common on sandy as well as hard bottoms and they also live associated with vegetation. These mudsnails are very mobile, feed mainly on detritus, and are continuously moving to fresh patches of food. They are classified as mobile surface detritivores. Shell length of
¨ Hydrobia used in the experiments was 3–4 mm. Nereis diversicolor O. F. Muller is a
˚
polychaete common on shallow soft bottoms in the Aland archipelago. It is mobile, omnivorous, eating small invertebrates, algae and detritus, and can function as predator,
¨
surface detritivore, and suspension feeder (Ronn et al., 1988; Nielsen et al., 1995). N. diversicolor is tolerant to hypoxia, anoxia, and even hydrogen sulphide (Vismann, 1990), and is commonly found under algal mats (Norkko and Bonsdorff, 1996b). Length of N. diversicolor used in the experiments was 40–50 mm. The amphipod Bathyporeia
˚ ¨
pilosa Lindstrom is common on exposed sandy bottoms in the Aland archipelago. It swims actively at night and is tolerant to short-term hypoxia (Mettam, 1989; Sandberg, 1994). B. pilosa feeds on benthic diatoms and detritus, and it is classified as a mobile carnivore / surface detritivore. Length of B. pilosa used in the experiments was 3–5 mm.
2.2.2. Experimental set-up
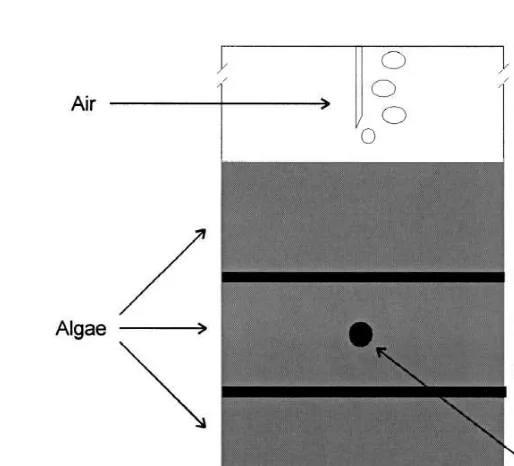
Drift algae for the laboratory experiments were collected from site B (Fig. 1) 1 day prior to the start of each experiment. The algae, consisting mainly of the brown algae E. siliculosus, P. littoralis, and D. foeniculaceus, were rinsed in running fresh water to remove larger macrofauna and then checked to ensure the rinsing was successful. M. balthica and N. diversicolor were collected from site B and B. pilosa from site H using an Ekman–Birge grab and Hydrobia spp. were obtained from algae collected at site B. The aquaria for the laboratory experiments were 50 cm high39.1 cm diameter plexiglass cylinders (volume 3.25 l, Fig. 2). Each aquaria had a 5 cm layer of air-dried and sieved (mesh size 0.5 mm) sand as sediment. Each species was tested separately and the five experiments were conducted using: (1) ten M. balthica (5 mm size class); (2) five M. balthica (10 mm size class); (3) 50 Hydrobia spp.; (4) five N. diversicolor; and (5) 20 B. pilosa in the replicate aquaria. The number of animals in the experimental aquaria corresponded to densities recorded in the field. The animals were acclimatised for 24 h before the experiments were started by adding the algae. To the algal treatment
2
(five replicates) a total 13.0 g wwt algae (corresponding to 2000 g wwt / m or 400 g
2
dwt / m ) was added in three layers separated by a thin net (mesh size 1–2 cm). Each
2
Fig. 2. Schematic detail of experimental aquarium; algal treatment. Diameter of cylinder 9.1 cm and actual height of cylinder 50 cm, thickness of each algal layer 4 cm.
The aquaria were covered with black plastic to the height of the algae in order to simulate light conditions prevailing under a more extensive algal mat. In control treatments the aquaria were covered only to the height of the sediment surface. Both algal treatments and controls were aerated and oxygen saturation above, in and under the algae was measured before the addition of algae, and then again after 12, 24, 48 and 72 h, using Winkler titration. Water samples in and under algae were obtained using a syringe to pierce silicon filled holes in the walls of the aquaria (Fig. 2). The experiments were conducted over 72 h with a 12:12 h light / dark period. When the experiments were terminated, the algal layers were retrieved separately, and survival and position (in sediment, on sediment or in any of the three layers of algae) of the animals were recorded.
2.3. Data analysis
2.3.1. Univariate
component (J ) were calculated for all replicate algal and benthic samples. For comparisons of algal and benthic fauna and for statistical analysis of data from the laboratory experiments the Mann–Whitney U-test was used. All values are given as mean6standard deviation (x6S.D.).
2.3.2. Multivariate
To describe temporal and spatial patterns in the invertebrate community of the drift algae, abundance data was analysed using correspondence analysis (CANOCO; ter Braak, 1988) and using the Bray–Curtis similarity index followed by nonmetric multidimensional scaling (MDS) ordination (Kruskal and Wish, 1978; PRIMER; Clarke and Warwick, 1994). Analyses were carried out on raw and transformed data (presence / absence, fourth root transformation), but only solutions using raw data are presented here, as we were interested in differences in abundances as well as species composition. Correspondence analysis and MDS ordinations gave very similar results and therefore only the correspondence analysis ordination plots are presented here. Canonical correspondence analysis (ter Braak, 1988) and BIOENV (Clarke and Ainsworth, 1993) were used to examine relationships between structure of faunal communities and environmental data. Again analyses carried out on raw data are presented. Environmen-tal parameters used in canonical correspondence and BIOENV analyses included time (date), depth, algal biomass, algal coverage, algal condition, exposure to wind-wave disturbance, and longitude and latitude of the sites. In addition, correspondence analysis scores for ambient benthic fauna at each site were used as an environmental factor in a canonical correspondence analysis, in order to investigate the influence of the structure of benthic fauna on the structure of algal fauna.
3. Results
3.1. Field study
3.1.1. Algal fauna: community composition and abundances ˚
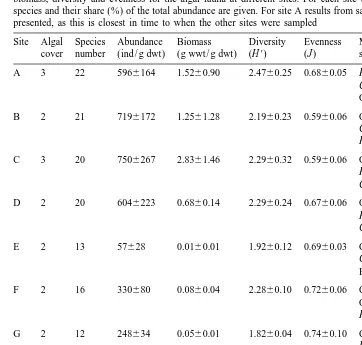
Our results show that drifting algal mats in the Aland archipelago at times can harbour high abundances of invertebrate macrofauna (Table 1). Twenty-six taxa were found in the algal mats and 20 taxa in the sediments (Table 2). The numerically most abundant taxa in algal mats were Hydrobia spp., Chironomidae, Ostracoda and newly settled larvae of Cerastoderma glaucum (Table 1, Fig. 3). Other taxa such as Oligochaeta, Hydrachnidae and Gammarus spp. were present in 100, 85.5, and 74.7% of the samples respectively, but never in high densities. The abundance of copepods and nematodes was not estimated, but they were present in 97.6 and 94.0% of the samples respectively.
3.1.2. Spatial variation of the algal fauna
Table 1
Algal cover (150–25%, 2525–75%, 3575–100% cover) and total number of species, mean abundance, biomass, diversity and evenness for the algal fauna at different sites. For each site the three most abundant species and their share (%) of the total abundance are given. For site A results from sampling occasion A2 are presented, as this is closest in time to when the other sites were sampled
Site Algal Species Abundance Biomass Diversity Evenness Most abundant Abund. cover number (ind / g dwt) (g wwt / g dwt) (H9) (J ) species (%)
Table 2
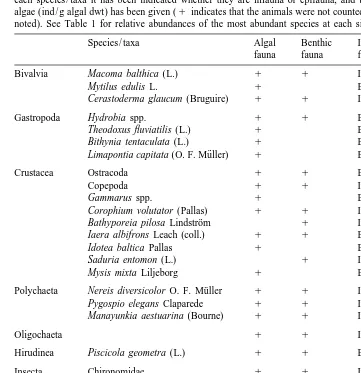
Taxa recorded in algae and in sediments, all sites and sampling occasions included (83 samples in total). For each species / taxa it has been indicated whether they are infauna or epifauna, and their mean abundance in algae (ind / g algal dwt) has been given (1 indicates that the animals were not counted, only presence / absence noted). See Table 1 for relative abundances of the most abundant species at each site
Species / taxa Algal Benthic In / epi Abundance
fauna fauna fauna in algae
Bivalvia Macoma balthica (L.) 1 1 In 0.561.0
Mytilus edulis L. 1 Epi 3.466.7
Cerastoderma glaucum (Bruguire) 1 1 In / epi 64.7683.7
Gastropoda Hydrobia spp. 1 1 Epi 61.7675.9
Theodoxus fluviatilis (L.) 1 Epi 0.962.4
Bithynia tentaculata (L.) 1 Epi 0.160.2
¨
Limapontia capitata (O. F. Muller) 1 Epi 3.465.9
Crustacea Ostracoda 1 1 Epi 139.26141.7
Copepoda 1 1 In / epi 1
Gammarus spp. 1 Epi 7.1613.6
Corophium volutator (Pallas) 1 1 In 0.160.2
¨
Bathyporeia pilosa Lindstrom 1 In / epi 0
Iaera albifrons Leach (coll.) 1 1 Epi 1.262.2
Idotea baltica Pallas 1 Epi 1.663.6
Saduria entomon (L.) 1 In / epi 0
Mysis mixta Liljeborg 1 Epi 1
¨
Polychaeta Nereis diversicolor O. F. Muller 1 1 In 0.0260.1
Pygospio elegans Claparede 1 1 In 0.0360.2
Manayunkia aestuarina (Bourne) 1 1 In 0.160.4
Oligochaeta 1 1 In 22.4619.9
Hirudinea Piscicola geometra (L.) 1 1 Epi 0.361.1
Insecta Chironomidae 1 1 In / epi 132.06156.8
Trichoptera 1 Epi 0.361.5
Arachnida Hydrachnidae 1 1 In / epi 15.8618.6
Nematoda 1 1 In 1
Turbellaria 1 1 In / epi 1
Nemertinea Prostoma obscurum Schultze 1 1 In / epi 0.961.5
Hydrozoa Hydra sp. 1 1 Epi 22.5632.2
Fig. 3. Relative abundance of the five most dominant species at all sites. The degree of exposure to wind-wave disturbance at the sites has been indicated.
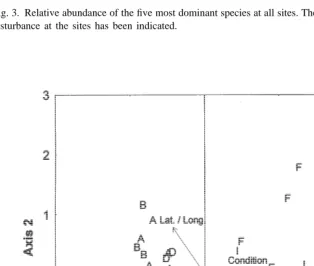
different sites, explaining 83% of the variability shown in Fig. 4 (P50.001). BIOENV indicated that depth, exposure and algal cover were the most important factors influencing the algal fauna, explaining 49% (r50.7) of the variability in the algal fauna. To investigate the influence of benthic fauna on the composition of algal fauna at each site, a correspondence analysis was carried out on raw data from the benthic samples (using mean abundance values from each site), with sample scores from this analysis included as environmental data in a canonical correspondence analysis with forward selection. This canonical correspondence analysis showed that the composition of benthic fauna explained 18% of the variability in the algal fauna (P50.03) and that 63% of the variability in algal fauna was explained by a combination of composition of benthic fauna, algal cover, algal condition and exposure.
3.1.3. Temporal changes in algal fauna
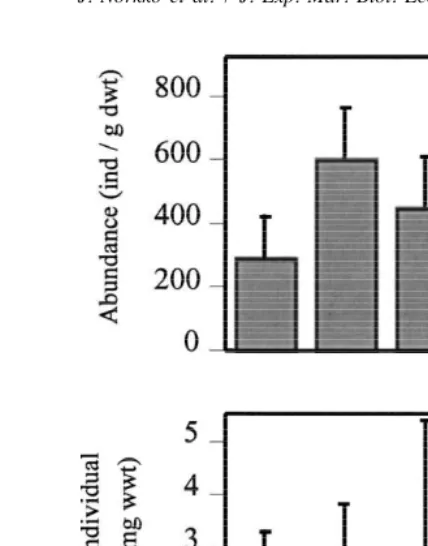
At site A drifting algal mats were sampled five times during 1996 (sampling occasions A1–A5). Sampling began when algal mats first started appearing in July, and continued until they began disintegrating in early October. Algal biomass peaked at A2 /A3 and algal coverage varied between 50 and 100%, decreasing towards the end of the sampling period. The algae also became more fragmented over time.
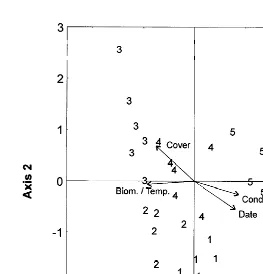
No significant differences in number of species or total biomass were noted between sampling dates (Kruskall–Wallis H-test, P.0.05), but multiple comparison tests showed that total abundance was significantly higher at A5 than at A1 (P,0.01). In general, total abundance increased over time, whereas the average individual biomass decreased (Fig. 5). Correspondence analysis ordination indicated that the faunal community changed over time, with 59% of the variability explained by the first two axes (Fig. 6). Canonical correspondence analysis suggested date, algal cover and algal condition to be the most important factors influencing faunal composition over time, and 81% of the variability was explained by the first two canonical axes (P50.001). Fig. 6 indicates that, of the environmental variables measured, high algal biomass and cover seem to be most influential at times A2–A4 (time of maximum occurrences), whereas the condition of algae (more fragmented) influenced the fauna at A5.
Fig. 5. Total abundance and the average individual biomass of the animals at different sampling occasions (A1–A5; July–October) at site A. Algal biomass peaked at A2 /A3 and subsequently algal coverage decreased and the algae became more fragmented.
During sorting of algal samples the condition of algae had been recorded, and the sites were subsequently grouped in to sites / sampling occasions with (Group I) fresh algae (B, C, E, A1, A2, A3, A4) or with (Group II) more fragmented and degraded algae (D, F, G, H, I, A5). Number of species and total biomass (Mann–Whitney U-test, P,0.0001), as well as diversity (P50.0125) were significantly higher in fresh algae (Table 3), whereas no significant differences were recorded with respect to abundance and evenness (P.0.05). As Fig. 4 shows, also the multivariate analyses of different sites indicated that algal condition influences the algal fauna.
3.1.4. Comparison of benthic and algal fauna
Generally, the algal fauna was dominated by Hydrobia spp., C. glaucum, Ostracoda and Chironomidae (Fig. 3, Table 1), whereas the benthic fauna was dominated by M.
balthica, Oligochaeta, Pygospio elegans, B. pilosa, as well as Hydrobia spp.,
Fig. 6. Ordination diagram based on canonical correspondence analysis of the temporal variation of the algal fauna at site A (sampling occasions 1–5). The important environmental factors are represented by arrows. The arrows for temperature and algal biomass overlap.
total abundance, biomass and diversity of the benthic fauna and multiple comparison tests (P,0.05) showed that abundance was significantly higher at site F than at sites G and H, biomass was significantly lower at site G than at sites A and B, and diversity was significantly higher at site B than at site F.
In order to compare community parameters of algal and benthic fauna in general,
Table 3
a Comparison of community parameters of algal fauna in fresh (Group I) and degraded (Group II) algae
Species Abundance Biomass Diversity Evenness
(per sample) (ind / g dwt) (g wwt / g dwt) (H9) (J )
Group I 15.062.9 5756341 1.2061.06 2.2560.32 0.6360.09
Group II 11.863.4 5136337 0.2860.26 2.0460.39 0.6660.14
P-value ,0.0001 0.5186 ,0.0001 0.0125 0.2324
*** N.S. *** * N.S.
a
Table 4
a Comparison of community parameters of algal fauna and benthic fauna
Species Abundance Biomass Diversity Evenness
(per sample) (ind / core) (g wwt / core) (H9) (J ) Algal fauna 12.363.9 82.9655.4 0.09560.139 2.0860.36 0.6660.11 Benthic fauna 5.961.6 34.4627.6 0.15960.269 1.5860.54 0.7260.19
P-value ,0.0001 0.0001 0.5156 ,0.0001 0.0526
*** *** N.S. *** N.S.
a
Number of species, total abundance, total biomass, diversity and evenness as mean (x6S.D.) of all benthic and all corresponding algal samples, and statistical differences (Mann–Whitney U-test) between these two. Levels of significance given as *** (P,0.001) and N.S. (not significant, P.0.05).
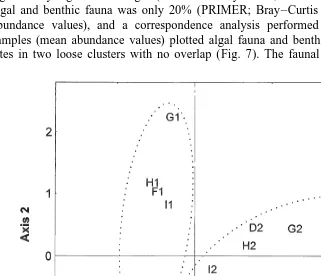
mean values of all benthic samples were compared with mean values of corresponding algal samples (Table 4). Number of species, total abundance, and diversity were significantly higher in the algae (Mann–Whitney U-test, P#0.001). Similarity between algal and benthic fauna was only 20% (PRIMER; Bray–Curtis similarity index, mean abundance values), and a correspondence analysis performed on algal and benthic samples (mean abundance values) plotted algal fauna and benthic fauna from different sites in two loose clusters with no overlap (Fig. 7). The faunal community of drifting
algae thus differs significantly from the resident benthic community of shallow sandy soft bottoms in the northern Baltic Sea.
3.2. Laboratory experiment
3.2.1. Aquaria conditions
During the experiments water temperature was 19.061.18C, salinity 5.760.1‰ and pH 7.9060.02 above the algae and in the controls. Oxygen saturation decreased rapidly at the sediment surface under the algae, reaching hypoxic conditions only 6–12 h after addition of algae (Fig. 8). In the algae oxygen saturation decreased to intermediate levels (35–75% O ; Fig. 8), and fluctuated depending on the structure and density of the algal2
mat. At the termination of the experiments the water under the algae was anoxic and there was a strong smell of H S in more than half of the algal replicates. At the same2
time oxygen saturation above algae was consistent at 100%.
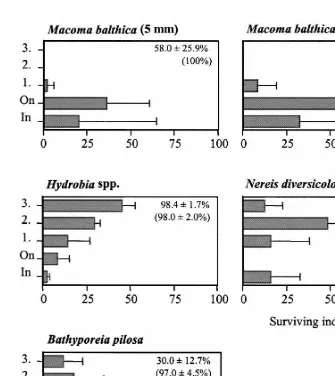
3.2.2. Survival and migration of test organisms in algae
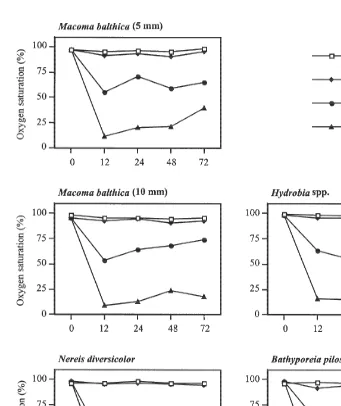
Survival was good in all control treatments in all experiments (Fig. 9) and the animals remained buried in the sediment. Survival was significantly lower (Mann–Whitney
U-test) in algal treatments than in controls for the 5 mm M. balthica (P50.0367) and
for B. pilosa (P50.0090). The position of the animals was recorded at the termination of the experiments (Fig. 9), and together with survival data, these results were used as a measure of a species ability to move up into a drifting algal mat. In all experiments, the vast majority of all the animals (83615%) had emerged from the sediment in the algal treatment, indicating that algae stress infauna and induce an escape response.
The majority of M. balthica (both size classes) emerged on to the sediment surface, but only a few moved into the algae, and none moved any higher than the first algal layer. M. balthica (10 mm) showed 100% survival in the algal treatment, indicating a higher tolerance to hypoxia and H S compared to juvenile M. balthica (5 mm,2
58.0625.9% survival). Hydrobia spp. showed high survival (98.461.7%) and mobility in algae, and most of the mudsnails were found in the second and third algal layers, where oxygen concentrations were higher. Survival of N. diversicolor was high (92.0611.0%), and these polychaetes seemed to move freely in the algae, showing no preference for any particular layer. Survival of B. pilosa was low (30.0612.7%), and the few surviving individuals were found in the upper algal layers (Fig. 9).
4. Discussion
4.1. The invertebrate community of drifting algal mats
Fig. 8. Oxygen saturation in the controls and in the algal treatments (above, in, and under the algae) in the experiments.
¨
Fig. 9. Position of the test organisms at the termination of the experiments; in the sediment, on the sediment or in the algae (layer 1, 2 or 3). At the start of the experiment all animals were in the sediment. The x-axis represents the proportion of the percentage surviving individuals found in each layer. In the upper right corner of each graph the survival of the animals in the algal treatment and the control (in brackets) are given (x6S.D.).
migrate into algal mats. Thus the reduced abundances often recorded under algae could partly be a result of animals moving up into algae and not only a result of mortality. Our results still indicate that not all benthic species are capable of successfully entering the algae. This would lead to mortality, unless they are mobile enough to horizontally escape from an incoming algal mat or tolerant to the hypoxia induced under drift algae. Most species comprising the sediment communities at the study sites were found in the algae (Table 2), but for example the bivalve M. balthica, the spionid polychaete Pyg.
elegans and the amphipod B. pilosa, which were amongst the dominant benthic species,
were only rarely found in the algae. It is, however, important to remember that the benthic fauna in the study area might previously, to an unknown extent, have been impoverished by drifting algal mats, affecting the interpretation of our results.
The algal mat communities at exposed sites differed from the communities at more sheltered sites (Figs. 3 and 4, Table 3). As exposure to wind-wave disturbance will change the structure and stability of the algal mats by fragmenting the algae, drift algae at sheltered sites probably provide a more stable habitat both in time and space. Also over time a succession in the faunal community was recorded (Fig. 6), with different species dominating at different times. The recorded decrease in average individual biomass (Fig. 5) may either be a consequence of juveniles being recruited to the algae, e.g., as observed for Hydrobia spp. or a consequence of a shift in species composition towards smaller sized taxa in more fragmented algae (e.g. from Hydrobia spp. to Chironomidae and Oligochaeta, Fig. 3). Both Hydrobia spp. and Chironomidae have short enough life cycles to potentially reproduce during the period the drift algae are present. The occurrence of drifting algae is an annually repeated phenomenon, and algae are generally broken down and decomposed every year. The invertebrate fauna found in algae probably play an important role in this process, facilitating breakdown by fragmenting the algae and making them more available for microorganisms. The habitat provided by drift algae thus changes as the algae are colonised by microorganisms and invertebrates and become fragmented (Snelgrove et al., 1996). The fate of algal mats that drift away before they are broken down has not been investigated in this area.
4.2. Benefits of drift algae to benthic macrofauna
Benthic drift algae add a complex three-dimensional structure to normally bare sandy soft bottoms and patchy occurrences of benthic drift algae increase habitat complexity (Vetter, 1995; Langtry and Jacoby, 1996; Norkko, 1997; Raffaelli et al., 1998) and can to some extent increase the area available for colonisation. Benthic drift algae can provide refuge from predators (Heck, 1979; Gore et al., 1981; Heck and Thoman, 1981; Kulczycki et al., 1981; Wilson et al., 1990; Norkko, 1998; Vetter, 1998), but they may also enhance prey availability if infauna is stressed by the algae and therefore emerge from the sediment (Norkko and Bonsdorff, 1996a,c).
Drifting algae, invertebrates living in the algae and / or microorganisms growing on the algae can provide a valuable food resource for higher trophic levels. Detached algae thus provide an important input of allochthonous organic material to the benthic community
¨
1997; Soares et al., 1997). Vetter (1995) recorded very high secondary production in accumulations of algal detritus. On the basis of the high abundances recorded in our study, production in the algal mats also appears to be high compared to surrounding sediments, although this is a transitory phenomenon.
Drifting algae can also affect the distribution (or redistribution) of fauna as some species can utilise algae as a transport mechanism through rafting (Highsmith, 1985; Holmquist, 1994). The number of C. glaucum has increased at site A during the last 10 years and this could be a result of larvae settling in drifting algal mats at the site or being transported to the site with drifting algae. Larvae of C. glaucum normally settle in attached algal belts before the secondary spat fall to the bottom. In this way, patches of drifting algae could function as a ‘‘stepping-stone’’ for recolonisation of sediments that have previously been defaunated by excessive amounts of drift algae. Movements of benthic drift algae could thus influence the distribution of species associated with the algae, as Kingsford (1992) showed for pelagic drift algae.
4.3. Benthic drift algae as a habitat; species specific responses of benthic macrofauna
Drifting algal mats are a highly mobile and seasonal structure in a state of slow degradation, governed by the availability of algae from the littoral zone and physical forces such as wind, currents and wave action (Kitching, 1937; Williams Cowper, 1978; Virnstein and Carbonara, 1985; ‘Olafsson, 1988; Bell et al., 1995; Vetter, 1995; Norkko and Bonsdorff, 1996a). Being seasonal and mobile, algal mats are insecure habitats both in time and space. A single drifting algal mat may also be highly heterogeneous with regards to oxygen levels, and algal composition, condition, thickness, size and
´
patchiness (Escartın and Aubrey, 1995), all of which may vary both vertically and horizontally within the mat. Our laboratory experiments showed high oxygen levels above algae, intermediate and fluctuating levels in the algae, and hypoxic to anoxic conditions under the algae (Fig. 8). Similar patterns have been recorded in the field (Thybo-Christesen et al., 1993; Norkko and Bonsdorff, 1996a) and hypoxia, anoxia and hydrogen sulphide are often induced under algal mats (Williams Cowper, 1978; Nicholls
¨
et al., 1981; Thrush, 1986; Sundback et al., 1990; Everett, 1994; Norkko and Bonsdorff, 1996a,b,c). The low oxygen concentrations prevailing under the algal mat would thus favour the algal mat as a habitat if the benthic fauna can migrate up into it (Nicholls et al., 1981).
Hydrobia ulvae utilise intertidal algal mats both as a food resource and refuge (Soulsby et al., 1982), and this is also likely to be the case in our study. Hydrobia spp. are able to migrate up through the algal mat (Fig. 9), and they rapidly colonise benthic drift algal mats in the field (Norkko and Bonsdorff, 1996a). Similarly, N. diversicolor is not inhibited by the presence of algae; it is tolerant to hypoxia (Vismann, 1990), and it is
¨
detritus. Nevertheless, the number of N. diversicolor found in drift algae in the field in this study was low (Table 2).
Conversely, M. balthica and B. pilosa cannot adapt to a life in drifting algae. M. balthica is adversely affected, as it is incapable of effectively migrating up into the algae (Fig. 9). Other studies have shown M. balthica to be unable to move away horizontally from algal mats (Norkko, 1998) and that drift algae inhibit the bivalves from extending their siphons (Norkko and Bonsdorff, 1996a). As M. balthica is tolerant to hypoxia (LT50 in anoxia 500 h; Dries and Theede, 1974), longer experimental time might have given different results, with more individuals entering the algae (Norkko, 1998; Norkko and Bonsdorff, 1996c). We recorded no B. pilosa in drift algae in the field and survival of the amphipod was poor in the laboratory experiment, even though it is known to tolerate short-term hypoxia (Sandberg, 1994). This indicates that B. pilosa has difficulties adjusting to the complex structure of drifting algal mats. In the laboratory experiments the animals were not given the choice of horizontally moving away from the algae, but even though B. pilosa might be able to escape from algae in the field, an extensive algal cover would significantly decrease the habitat available to this amphipod. The shallow soft bottoms of the northern Baltic Sea are characterised by a pronounced seasonality in production and recruitment (Bonsdorff and Blomqvist, 1993). Both M. balthica and B. pilosa are adversely affected by the algae, as recruitment coincides both spatially and temporally with mass occurrences of drifting algal mats (‘Olafsson, 1988; Bonsdorff et al., 1995). Juvenile M. balthica are also generally more sensitive than adults to stress induced by algae (Fig. 9, Bonsdorff, 1992; Bonsdorff et al., 1995; Norkko and Bonsdorff, 1996c).
The responses of the benthic macrofauna to drifting algae are thus highly species specific, with some species being adversely affected and others being promoted by the algae. The fauna found in the drift algae consists both of species escaping from the adverse conditions prevailing under the algal mat and of species actively colonising the algae, utilising them as a food source or refuge. Mobile, epibenthic species most commonly utilise drifting algae (Nicholls et al., 1981; Everett, 1994; Tzetlin et al., 1997; Norkko, 1998) and according to Gore et al. (1981) and Kirkman and Kendrick (1997) the fauna of accumulations of algae and seagrass mainly consists of detritivores and omnivores. Also, small sized species are more likely to reach high densities in the algae and may therefore better utilise the algae as a means of dispersal, as high numbers render them more likely to succeed in colonising new habitats (Highsmith, 1985). Drifting algae can be an important alternative habitat for epifauna normally associated with seagrass beds (Williams Cowper, 1978; Virnstein and Carbonara, 1985; Virnstein and Howard, 1987; Holmquist, 1997), but in the present study we generally recorded only low numbers of herbivores, e.g., the gastropod Theodoxus fluviatilis, the nudibranch L. capitata, and the isopods Iaera albifrons and Idotea baltica, all of which are
¨ ¨
important inhabitants of attached algal belts in this area (Ostman and Ronnberg, 1991; Kautsky, 1995).
4.4. Effects of ongoing eutrophication
effects of drifting algal mats on benthic communities vary with spatial and temporal extent of algal cover; the algae might be present for just a couple of hours, or up to several months. If the amounts of drifting algae continue to increase in the northern Baltic Sea, the benthic community on shallow sandy bottoms will change from being dominated by long-lived species (e.g. bivalves) to being dominated by opportunistic short-lived species (e.g. chironomids). The effects of increasing nutrient levels in the Baltic Sea will also increase the variability of ecosystem-responses, including those of the benthic macrofauna (Breitburg et al., 1999). For most benthic fauna normally living in or on bare sandy soft bottoms, drift algae can only function as a temporary habitat, as a way of escaping adverse conditions prevailing under the algae, and the effects of drift algae on these species are therefore ultimately detrimental.
In conclusion, benthic drift algal mats may harbour high abundances of invertebrate macrofauna and it is obvious that drift algae may function as a food resource, refuge from predators and transport mechanism for associated macrofauna also in the Baltic Sea. Nevertheless, the responses of benthic macrofauna to drifting algal mats are highly species specific. The hypothetical species to successfully utilise benthic drift algae as a habitat is a highly mobile epibenthic species, a detritivore, tolerant to hypoxia, anoxia and hydrogen sulphide, and has a life history that both spatially and temporally is adapted to the occurrences of drifting algae. The benefits drift algae bring to the benthic fauna (habitat enhancement, food, refuge), can only be considered temporary, as the algal mats are mobile, seasonal and subject to degradation and as the long term effects on the entire system are negative due to degeneration of benthic infauna and seagrass beds. With ongoing eutrophication the occurrence, biomass and the area affected by drifting algal mats is expected to increase in the northern Baltic Sea and their role in structuring benthic communities will therefore also grow.
Acknowledgements
¨
We thank Huso Biological Station for providing excellent working conditions. Judi Hewitt helped with the multivariate analyses of the data and Vonda Cummings, Simon Thrush, Eric Vetter and an anonymous referee offered valuable comments on the
¨ manuscript. This study was supported by the Academy of Finland and the Nordenskiold Foundation. AN was supported by the Academy of Finland (FA 41837), which is gratefully acknowledged. [RW]
References
Bell, S.S., Hall, M.O., Robbins, B.D., 1995. Toward a landscape approach in seagrass beds: using macroalgal accumulation to address questions of scale. Oecologia 104, 163–168.
Blomqvist, E.M., Bonsdorff, E., Essink, K. (Eds.), 1999. Biological, physical and geochemical features of enclosed and semienclosed marine systems, Dev. Hydrobiol., Vol. 135, Kluwer, Dordrecht.
Bonsdorff, E., Blomqvist, E.M., 1993. Biotic couplings on shallow water soft bottoms — examples from the northern Baltic Sea. Oceanogr. Mar. Biol. Annu. Rev. 31, 153–176.
Bonsdorff, E., Pearson, T.H., 1999. Variations in the sublittoral macrozoobenthos of the Baltic Sea along environmental gradients: a functional-group approach. Aust. J. Ecol. 24, 312–326.
Bonsdorff, E., Aarnio, K., Sandberg, E., 1991. Temporal and spatial variability of zoobenthic communities in archipelago waters of the northern Baltic Sea — consequences of eutrophication? Int. Rev. Ges. Hydrobiol. 76 (3), 433–449.
¨
Bonsdorff, E., Norkko, A., Bostrom, C., 1995. Recruitment and population maintenance of the bivalve Macoma balthica (L.) — factors affecting settling success and early survival on shallow sandy bottoms. In: Eleftheriou, A., Ansell, A., Smith, C. (Eds.), Proceedings of the 28th European Marine Biology Symposium, Biology and Ecology of Shallow Coastal Waters, Olsen and Olsen, Fredensborg, Denmark, pp. 253–260.
Bonsdorff, E., Blomqvist, E.M., Mattila, J., Norkko, A., 1997. Long-term changes and coastal eutrophication.
˚
Examples from the Aland Islands and the Archipelago Sea, northern Baltic Sea. Oceanol. Acta 20 (1), 319–329.
¨
Bostrom, C., Bonsdorff, E., 1997. Community structure and spatial variation of benthic invertebrates associated with Zostera marina (L.) beds in the northern Baltic Sea. J. Sea Res. 37, 153–166.
Breitburg, D.L., Sanders, J.G., Gilmour, C.C., Hatfield, C.A., Osman, R.W., Riedel, G.F., Seitzinger, S.P., Sellner, K.G., 1999. Variability in responses to nutrients and trace elements, and transmission of stressor effects through an estuarine food web. Limnol. Oceanogr. 44, 837–863.
Cederwall, H., Elmgren, R., 1990. Biological effects of eutrophication in the Baltic Sea, particularly the coastal zone. Ambio 19 (3), 109–112.
Clarke, K.R., Ainsworth, M., 1993. A method of linking multivariate community structure to environmental variables. Mar. Ecol. Prog. Ser. 92, 205–219.
Clarke, K.R., Warwick, R.M., 1994. Change in Marine Communities: An Approach To Statistical Analysis and Interpretation, Natural Environment Research Council, UK.
Dries, R.R., Theede, H., 1974. Sauerstoffmangelresistenz mariner Bodeninvertebraten aus der westlichen Ostsee. Mar. Biol. 25, 327–333.
Duggins, D.O., Simenstad, C.A., Estes, J.A., 1989. Magnification of secondary production by kelp detritus in coastal marine ecosystems. Science 245, 170–173.
´
Elmgren, R., 1984. Trophic dynamics in the enclosed, brackish Baltic Sea. Rapp. P.-v. Reun. Cons. Int. Explor. Mer. 183, 152–169.
Elmgren, R., 1989. Man’s impact on the ecosystem of the Baltic Sea: energy flows today and at the turn of the century. Ambio 18 (6), 326–332.
´
Escartın, J., Aubrey, D.G., 1995. Flow structure and dispersion within algal mats. Estuar. Coast. Shelf Sci. 40, 451–472.
Everett, R.A., 1994. Macroalgae in marine soft-sediment communities: effects on benthic faunal assemblages. J. Exp. Mar. Biol. Ecol. 175, 253–274.
Gore, R.H., Gallaher, E.E., Scotto, L.E., Wilson, K.A., 1981. Studies on decapod crustacea from the Indian River region of Florida. XI. Community composition, structure, biomass and species–areal relationships of seagrass and drift algae-associated macrocrustaceans. Estuar. Coast. Shelf Sci. 12, 485–508.
Heck, Jr. K.L., 1979. Some determinants of the composition and abundance of total macroinvertebrate species in tropical and temperate turtlegrass (Thalassia testudinum) meadows. J. Biogeogr. 6, 183–200. Heck, Jr. K.L., Thoman, T.A., 1981. Experiments on predator prey interactions in vegetated aquatic habitats. J.
Exp. Mar. Biol. Ecol. 53, 125–134.
HELCOM, 1990. Second periodic assessment of the status of the marine environment of the Baltic Sea, 1984–1988. Background document. In: Baltic Sea Environ. Proc., Vol. 35b, pp. 1–432.
Highsmith, R.C., 1985. Floating and algal rafting as potential dispersal mechanisms in brooding invertebrates. Mar. Ecol. Prog. Ser. 25, 169–179.
Holmquist, J.G., 1994. Benthic macroalgae as a dispersal mechanism for fauna: influence of a marine tumbleweed. J. Exp. Mar. Biol. Ecol. 180, 235–251.
¨
Holmstrom, A., 1998. Occurrence, species composition, biomass and decomposition of drifting algae in the
˚
northwestern Aland archipelago, N. Baltic Sea. MSc Thesis (in Swedish with English summary),
˚
Department of Biology, Abo Akademi University, 69 pp.
Hull, S.C., 1987. Macroalgal mats and species abundance: a field experiment. Estuar. Coast. Shelf Sci. 25, 519–532.
´
Ingolfsson, A., 1998. Dynamics of macrofaunal communities of floating seaweed clumps off western Iceland: a study of patches on the surface of the sea. J. Exp. Mar. Biol. Ecol. 231, 119–137.
Kautsky, H., 1991. Influence of eutrophication on the distribution of phytobenthic plant and animal communities. Int. Rev. Ges. Hydrobiol. 76, 423–432.
Kautsky, U., 1995. Ecosystem processes in coastal areas of the Baltic Sea. PhD Thesis, Stockholm University, 25 pp.
Kingsford, M.J., 1992. Drift algae and small fish in coastal waters of northeastern New Zealand. Mar. Ecol. Prog. Ser. 80, 41–55.
Kingsford, M.J., Choat, J.H., 1985. The fauna associated with drift algae captured with a plankton-mesh purse seine net. Limnol. Oceanogr. 30, 618–630.
Kirkman, H., Kendrick, G.A., 1997. Ecological significance and commercial harvesting of drifting and beach-cast macroalgae and seagrasses in Australia: a review. J. Appl. Phycol. 9, 311–326.
Kitching, J.A., 1937. Studies in sublittoral ecology. III. Recolonisation at the upper margin of the sublittoral region: with a note on the denudation of laminarian forests by storms. J. Ecol. 2, 482–495.
Kruskal, J.B., Wish, M., 1978. Multidimensional Scaling, Sage, Beverly Hills, CA.
Kulczycki, G.R., Virnstein, R.W., Nelson, W.G., 1981. The relationship between fish abundance and algal biomass in a seagrass-drift algae community. Estuar. Coast. Shelf Sci. 12, 341–347.
Langtry, S.K., Jacoby, C.A., 1996. Fish and decapod crustaceans inhabiting drifting algae in Jervis Bay, New South Wales. Aust. J. Ecol. 21, 264–271.
Lenanton, R.C.J., Robertson, A.I., Hansen, J.A., 1982. Nearshore accumulations of detached macrophytes as nursery areas for fish. Mar. Ecol. Prog. Ser. 9, 51–57.
Mattila, J., 1995. Does habitat complexity give refuge against fish predation? some evidence from two field experiments. In: Eleftheriou, A., Ansell, A.D, Smith, C.J. (Eds.), Proceedings of the 28th European Marine Biology Symposium, Biology and Ecology of Shallow Coastal Waters, Olsen and Olsen, Fredensborg, Denmark, pp. 253–259.
¨
Mettam, C., 1989. The life cycle of Bathyporeia pilosa Lindstrom (Amphipoda) in a stressful, low salinity environment. Sci. Mar. 53, 543–550.
Morand, P., Briand, X., 1996. Excessive growth of macroalgae: a symptom of environmental disturbance. Bot. Mar. 39, 491–516.
¨
Makinen, A., Aulio, K., 1986. Cladophora glomerata (Clorophyta) as an indicator of coastal eutrophication. Publ. Water Res. Inst. Finland 68, 160–163.
¨
Nehring, D., Matthaus, W., 1991. Current trends in hydrographic and chemical parameters and eutrophication in the Baltic Sea. Int. Rev. Ges. Hydrobiol. 76 (3), 297–316.
Nicholls, D.J., Tubbs, C.R., Haynes, F.N., 1981. The effect of green algal mats on intertidal macrobenthic communities and their predators. Kieler Meeresforsch., Sonderh. 5, 511–520.
˚
Nielsen, A.M., Eriksen, N.T., Lønsmann Iversen, J.J., Riisgard, H.U., 1995. Feeding, growth and respiration in the polychaetes Nereis diversicolor (facultative filter-feeder) and N. virens (omnivorous) — a comparative study. Mar. Ecol. Prog. Ser. 125, 149–158.
Norkko, A., 1997. The role of drifting macroalgal mats in structuring coastal zoobenthos. PhD Thesis,
˚
Department of Biology, Abo Akademi University, 41 pp.
Norkko, A., 1998. The impact of loose-lying algal mats and predation by the brown shrimp Crangon crangon (L.) on infaunal prey dispersal and survival. J. Exp. Mar. Biol. Ecol. 221, 99–116.
Norkko, A., Bonsdorff, E., 1996a. Rapid zoobenthic community responses to accumulations of drifting algae. Mar. Ecol. Prog. Ser. 131, 143–157.
Norkko, A., Bonsdorff, E., 1996b. Population responses of coastal zoobenthos to stress induced by drifting algal mats. Mar. Ecol. Prog. Ser. 140, 141–151.
‘Olafsson, E.B., 1988. Inhibition of larval settlement to a soft bottom benthic community by drifting algal mats: an experimental test. Mar. Biol. 97, 571–574.
¨ ¨ ˚
Ostman, M., Ronnberg, O., 1991. Effects of ships’ waves on rock-pools in the Aland Archipelago, northern Baltic Sea. Sarsia 76 (1–2), 125–132.
Peterson, C.H., Irlandi, E.A., Black, R., 1994. The crash in suspension-feeding bivalve populations (Katelysia spp.) in Princess Royal Harbour: an unexpected consequence of eutrophication. J. Exp. Mar. Biol. Ecol. 176, 39–52.
Pihl, L., Magnusson, G., Isaksson, I., Wallentinus, I., 1996. Distribution and growth dynamics of ephemeral macroalgae in shallow bays on the Swedish west coast. J. Sea Res. 35 (1–3), 169–180.
Polis, G.A., Hurd, S.D., 1996. Linking marine and terrestrial food webs: Allochthonous input from the ocean supports high secondary productivity on small islands and coastal land communities. Am. Nat. 147 (3), 396–423.
Raffaelli, D., Raven, J., Poole, L., 1998. Ecological impact of green macroalgal blooms. Annu. Rev. Mar. Biol. Oceanogr. 36, 97–126.
Rosenberg, R., Elmgren, R., Fleischer, S., Jonsson, P., Persson, G., Dahlin, H., 1990. Marine eutrophication case studies in Sweden. Ambio 19 (3), 102–108.
¨
Ronn, C., Bonsdorff, E., Nelson, W.G., 1988. Predation as a mechanism of interference within infauna in ¨ shallow brackish water soft bottoms; experiments with an infauna predator, Nereis diversicolor O.F. Muller. J. Exp. Mar. Biol. Ecol. 116, 143–157.
Sandberg, E., 1994. Does short-term oxygen depletion affect predator–prey relationships in zoobenthos? Experiments with the isopod Saduria entomon. Mar. Ecol. Prog. Ser. 103, 73–80.
Schramm, W., Nienhuis, P.H. (Eds.), 1996. Marine Benthic Vegetation: Recent Changes and the Effects of Eutrophication, Springer Verlag, Berlin.
˚
Segerstrale, S.G., 1962. Investigations on Baltic populations of the bivalve Macoma balthica (L). II. What are the reasons for periodic failure of recruitment and the scarcity of Macoma in deeper waters of the inner Baltic? Soc. Sci. Fenn. Commentat. Biol. 24, 1–26.
Shaffer, J.A., Doty, D.C., Buckley, R.M., West, J.E., 1995. Crustacean community composition and trophic use of the drift vegetation habitat by juvenile splitnose rockfish Sebastes diploproa. Mar. Ecol. Prog. Ser. 123, 13–21.
Snelgrove, P.V.R., Grassle, J.F., Petrecca, R.F., 1996. Experimental evidence for ageing food patches as a factor contributing to high deep-sea macrofaunal diversity. Limnol. Oceanogr. 41 (4), 605–614.
Soares, A.G., Schlacher, T.A., McLachlan, A., 1997. Carbon and nitrogen exchange between sandy beach clams (Donax serra) and kelp beds in the Benguela coastal upwelling region. Mar. Biol. 127, 657–664. Sokal, R.R., Rohlf, F.J., 1981. Biometry, Freeman, San Francisco.
Soulsby, P.G., Lowthion, D., Houston, M., 1982. Effects of macroalgal mats on the ecology of intertidal mudflats. Mar. Poll. Bull. 13 (5), 162–166.
¨ ¨ ¨
Sundback, K., Jonsson, B., Nilsson, P., Lindstrom, I., 1990. Impact of accumulating drifting macroalgae on a shallow-water sediment system: an experimental study. Mar. Ecol. Prog. Ser. 58, 261–274.
ter Braak, C.J.F., 1988. CANOCO — a FORTRAN program for canonical community ordination by [partial] [detrended] [canonical] correspondence analysis, principal components analysis and redundancy analysis (version 2.1). Report LWA-88-02, Agricultural Mathematics Group, Wageningen, The Netherlands. Thrush, S.F., 1986. The sublittoral macrobenthic community structure of an Irish sea-lough: effect of
decomposing accumulations of seaweed. J. Exp. Mar. Biol. Ecol. 96, 199–212.
Thybo-Christesen, M., Rasmussen, M.B., Blackburn, T.H., 1993. Nutrient fluxes and growth of Cladophora sericea in a shallow Danish bay. Mar. Ecol. Prog. Ser. 100, 273–281.
Tzetlin, A.B., Mokievsky, V.O., Melnikov, A.N., Saphonov, M.V., Simdyanov, T.G., Ivanov, I.E., 1997. Fauna associated with detached kelp in different types of subtidal habitats of the White Sea. Hydrobiologia 355, 91–100.
Vetter, E.W., 1994. Hotspots of benthic production. Nature 372, 47.
Vetter, E.W., 1995. Detritus based patches of high secondary production in the nearshore benthos. Mar. Ecol. Prog. Ser. 120, 251–262.
Virnstein, R.W., Carbonara, P.A., 1985. Seasonal abundance and distribution of drift algae and seagrasses in the mid-Indian River lagoon, Florida. Aquat. Bot. 23, 67–82.
Virnstein, R.W., Curran, M.C., 1986. Colonization of artificial seagrass versus time and distance from source. Mar. Ecol. Prog. Ser. 29, 279–288.
Virnstein, R.W., Howard, R.K., 1987. Motile epifauna of marine macrophytes in the Indian River lagoon, Florida. II. Comparisons between drift algae and three species of seagrasses. Bull. Mar. Sci. 41 (1), 13–26. Vismann, B., 1990. Sulfide detoxification and tolerance in Nereis(Hediste) diversicolor and Nereis (Neanthes)
virens (Annelida: Polychaeta). Mar. Ecol. Prog. Ser. 59, 229–238.
Williams Cowper, S., 1978. The drift algae community of seagrass beds in Redfish Bay, Texas. Mar. Sci. 21, 125–132.