MARBLING DAGING DOMBA
HIDAYATI
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
BOGOR
2015
Dengan ini saya menyatakan bahwa disertasi berjudul “Identifikasi Keragaman Gen Lecithin Cholesterol Acyltransferase dan Lipoprotein Lipase serta Hubungannya dengan Kualitas Marbling Daging Domba” adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apapun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir disertasi ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, Januari 2015
Hidayati NRP D161100011
dan Lipoprotein Lipase serta Hubungannya dengan Kualitas Marbling Daging Domba. Dibawah bimbingan CECE SUMANTRI, RONNY RAHMAN NOOR dan RUDY PRIYANTO
Indonesia memiliki beberapa rumpun domba lokal yang memiliki keragaman fenotip dan genetik yang tersebar di beberapa wilayah. Keragaman genetik yang dimiliki oleh masing-masing individu atau masing-masing rumpun merupakan sumber daya genetik yang dapat digunakan dalam peningkatan dan pengembangan domba lokal di masa yang akan datang. Direktorat Jenderal Peternakan telah melaporkan peningkatan populasi domba secara Nasional selama 5 tahun terakhir (2011-2014) yaitu 10.725.000 ekor; 11.791.000 ekor; 13.420.000 ekor; 14.926.000 ekor dan 15.716.000 ekor secara berturut-turut. Peningkatan populasi domba ini merupakan suatu potensi untuk memenuhi permintaan daging merah yang terus meningkat dari tahun ke tahun. Namun konsumsi daging domba dan kambing di Indonesia masih rendah yaitu baru sekitar 5% dari total kebutuhan daging atau setara dengan 0.24 g/kapita/tahun. Masih rendahnya konsumsi daging kambing dan domba karena harga daging kambing yang relatif tinggi, persepsi masyarakat bahwa daging domba tidak sehat karena memiliki kandungan kolesterol dan lemak jenuh yang tinggi dan adanya bau khas yang sulit dihilangkan. Keberadaan lemak pada bagian intramuskuler atau dikenal dengan marbling merupakan lemak yang memiliki arti penting dalam penilaian terhadap daging masak karena berpengaruh terhadap keempukan, juiciness dan flavour terutama dalam pembuatan steak serta lemak pada bagian ini tidak dapat dipisahkan langsung karena merupakan bagian dari otot berbeda dengan lemak pada bagian lainya. Keragaman gen-gen fungsional yang berasosiasi dengan kualitas marbling merupakan suatu hal penting dalam mengembangkan domba yang aman untuk dikonsumsi diantaranya dengan eksplorasi keragaman gen LCAT dan gen LPL melalui metode sekuensing dan selanjutnya keragaman yang muncul dihubungkan dengan kualitas marbling untuk menemukan penanda genetik yang dapat digunakan dalam seleksi (Marker Assisted Selection) dalam pengembangan domba lokal di masa yang akan datang
Hasil penelitian pertama menemukan 3 SNPs baru gen LCAT ekson 6 pada domba lokal Indonesia yaitu pada posisi basa c.742 C>T; c.770 T >A dan c.882 C>T. Kombinasi 3 SNPs membentuk sembilan diplotipe. Mutasi transisi sitosina menjadi timina c.742 merupakan synonymous mutation (Ala>Ala); mutasi transversi timina menjadi adenina c.770 dan mutasi transisi sitosina menjadi timina c.882 merupakan non-synonymous mutation mengakibatkan perubahan asam amino phenylalanina>isoleusina dan valina>alanina. Ketiga SNPs yang ditemukan merupakan SNPs baru yang belum dilaporkan pada populasi domba lainnya. Keragaman gen LCAT ekson 6 ditemukan pada rumpun domba ekor tipis sumatera, domba ekor tipis jawa, domba ekor gemuk jawa, domba garut, domba lembah palu dan domba pulau rote dan tidak ditemukan pada rumpun domba kissar dan domba sumbawa.
Hasil penelitian kedua menemukan 3 SNPs baru gen LPL bagian 5’UTR dan ekson 1 yaitu insersi pada posisi basa g.26>C/G, g.27>G dan mutasi pada
c.192T>C. Insersi g.26>C/G dan insersi g.27>G merupakan frameshift mutation dengan frekuensi munculnya SNPs pada kedua titik mutasi ini relatif rendah pada domba garut dan domba ekor tipis sumatera. Mutasi transisi pada posisi basa c.192 T>C merupakan non synonymous mutation karena perubahan basa mengakibatkan perubahan asam amino valina>alanina (Val>Ala). Mutasi pada c.192 bersifat polimorfik pada domba garut dan monomorfik pada domba ekor tipis sumatera.
Penelitian tahap tiga bertujuan untuk mengasosiasikan keragaman gen LPL c.192 dengan kualitas marbling domba garut. Asosiasi keragaman genotipe gen LPL dan kualitas marbling dianalisis menggunakan analisis sidik ragama satu arah dan uji beda nyata terkecil untuk uji lanjut. Hasil penelitian menunjukkan bahwa perbedaan genotipe gen LPL mempengaruhi asam lemak heneikosanoat, dimana genotipe TT (0.04%) memiliki kandungan heneikosanoat lebih tinggi dibandingkan genotipe CC (0.03%) dan genotipe CT (0.02%). Asam lemak heneikosanoat merupakan asam lemak minor di alam, namun keberadaan asam lemak heneikosanoat harus diwaspadai karena keberadaan asam lemak jenuh rantai panjang dengan atom karbon ganjil ini mengindikasikan terjadinya akumulasi propionat pada ternak ruminansia. Propionat merupakan asam lemak terbang hasil fermentasi rumen pada ternak ruminansia yang diberi pakan konsentrat tinggi dan mengindikasikan terjadinya defisiensi biotin. Biotin dibutuhkan oleh ternak ruminansia dalam proses konversi propionat menjadi methyl malonyl CoA yang dibutuhkan dalam proses perpanjangan asam lemak dengan menyumbangkan dua atom karbon. Defisiensi biotin dapat menghambat pembentukan methyl malonyl CoA mengakibatkan munculnya asam lemak rantai panjang dengan atom karbon ganjil. Selain itu keberadaan asam lemak henekosanoat dapat mengoksidasi asam lemak omega, namun mekanisme pastinya belum diketahui.
Rasio asam lemak tidak jenuh ganda (ALTJG): asam lemak jenuh (ALJ), kandungan kolesterol, nilai desirable fatty acid (DFA), nilai indeks atherogenicity dan rasio (stearat + oleat): palmitat marbling, merupakan indikator bagi daging domba yang sehat dikonsumsi. Hasil analisis menunjukkan bahwa daging domba garut relatif masih aman dan sehat dikonsumsi karena kandungan kolesterol rendah (6.10%-8.91%), nilai indeks atherogenicity berkisar 0.965-1.180, rasio (stearat + oleat): palmitat berkisar 1.984 – 2.330, nilai desirable fatty acid 42.360 – 47.720 kecuali rasio ALTJG : ALJ lebih rendah (0.062-0.065) dari rekomendasi 0.40.
Acyltransferase gene and Lipoprotein Lipase gene and its association with marbling lamb quality on Indonesian local sheep. Supervised by CECE SUMANTRI, RONNY RAHMAN NOOR and RUDY PRIYANTO.
Indonesia local sheep breeds was scattered in several provinces and its own phenotypic and genetic characteristic and variation. The existing genetic variation within and among groups can be utilized to improve their productivity. Directorate general of animal husbandry has reported increasing of sheep population last five years (2010-2014) i.e. 10.725.000 heads; 11.791.000 heads; 13.420.000 heads; 14.926.000 heads and 15.716.000 heads respectively. However, sheep and goat meat consumption in Indonesia was still low, only about 5% of the total demand for meat, equivalent to 0.24 g/capita/ year. The low consumption of lamb is caused relatively by high cholesterol and saturated fatty acid of lamb. The consumers are concerned about health factors, due to strong statements from the medical profession that lamb may contain too much saturated fatty acid and trans monounsaturated fatty acid. Marbling is important on the assessment meat because effected the tenderness, juiciness and flavor, especially in steak. This fat could be separated from muscle. The genes exploration of Lecithin Cholesterol Acyltransferase and Lipoprotein Lipase polymorphisms by sequencing method and its association with marbling quality is an important to Marker Assisted Selection (MAS) can be utilized to development and enhanced the genetic quality of Indonesian local sheep.
The results of the first study show three novel SNPs of LCAT gene exon 6 in Indonesian local sheep, at base c.742 C>T; c.770 T>A and c.882 C>T. The combination of three SNPs was formed nine diplotypes. The transition mutation of cytosine into thymine is synonymous mutation c.742 (Ala>Ala); the transversion mutation of adenine to thymine at c.770 and transition mutation thymine to cytosine c.882 were a non-synonymous mutations and resulted of changed phenylalanine>isoleucine and valine>alanine. So far there is no published studies describing three novel SNPs. Polymorphisms of LCAT gene exon 6 were found in sumatran thin tail ed sheep, javanese thin tail ed sheep, javanese fat tail ed sheep, garut sheep, lembah palu sheep and rote island sheep and neither in kissar sheep and sumbawa sheep.
The results of the second study were showed three novel SNPs of LPL gene i.e. the insertion at base g.26>C/G, g.27>G and transition mutation at base c.192T>C. Insertion g.26.C/G and g.27>G were a frameshift mutation with appearance frequency relatively low in both sumatran thin tail ed sheep and garut sheep. Mutations in the base c.192 T>C was non synonymous mutation and changed of valine>alanine (Val> Ala). This mutation is monomorphic on sumtran thin tail and polymorphic on garut sheep.
The third study showed the polymorphisms of LPL gene at c.192T>C on garut sheep was associated with heneicosanoic acid, whereas TT genotype (0.04) had higher than CC (0.03) and CT (0.02). Heneicosanoic is a minor fatty acid in nature, but the presence of heneicosanoic (21:0) affect the meat quality. The
propionic acid and biotin deficiency. Propionate is one of the volatile fatty acids of rumen fermentation. Conversion propionate into methyl malonyl CoA requires biotin. The methyl malonyl CoA was required for elongated fatty acid process by providing two carbon atoms. Biotin deficiency could inhibit the formation of methyl malonyl CoA and caused odd long-chain fatty acids. In addition, the presence of fatty acids heneicosanoic acid could oxidize omega fatty acids, but the exact mechanism is still unknown.
The results indicate that the lamb of garut sheep is relatively safe and healthy to be consumed because the cholesterol content still low (6.10%-8.91%), atherogenicity index of fatty acid was 0.965 -1.180, the ratio of (stearic + oleic): palmitic was 1.984 - 2.330, and index of desirible fatty acid is 42.360 - 47.720. They were still within the recommended range, except for PUFA: SFA ratio was lower (0.062-0.065) than recomended 0.40.
© Hak Cipta milik IPB, tahun 2015
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB
Dilarang mengumumkan dan memperbanyak sebagian atau seluruh karya tulis ini dalam bentuk apapun tanpa izin IPB.
MARBLING DAGING DOMBA
HIDAYATI
Disertasi
Sebagai salah satu syarat untuk memperoleh gelar Doktor
pada
Program Studi Ilmu dan Teknologi Peternakan
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
BOGOR
2015
Penguji pada Ujian Tertutup: 1. Dr Ir Dedi Duryadi Solihin, DEA
Staf Pengajar FMIPA IPB
2. Dr Tuti Suryati, SPt MSi
Staf Pengajar Fakultas Peternakan IPB
Penguji pada Ujian Terbuka : 1. Dr Ir Dedi Rahmat, MSi
Staf Pengajar Fakultas Peternakan UNPAD 2. Prof (R) Dr Ir Ismeth Inounu
Peneliti Utama Puslitbangnak Kementerian Pertanian RI
dan karunia-Nya sehingga disertasi ini dapat diselesaikan. Topik kajian yang diteliti dalam penelitian yang dilaksanakan dari bulan Oktober 2012 sampai dengan Agustus 2014 adalah “Identifikasi Keragaman Gen Lecithin Cholesterol Acyltransferase dan Lipoprotein Lipase serta Hubungannya dengan Kualitas Marbling Daging Domba”. Kedua gen ini memiliki peranan dalam transportasi lemak dan deposisi lemak di dalam tubuh. Karya ilmiah ini disusun sebagai salah satu syarat untuk memperoleh gelar Doktor pada program studi Ilmu dan Teknologi Peternakan, Sekolah Pascasarjana, Institut Pertanian Bogor.
Penulis menyadari bahwa penelitian dan penyusunan disertasi ini tidak akan terlaksana dengan baik tanpa bantuan dan dukungan dari banyak pihak. Oleh karena itu ucapan terima kasih penulis haturkan kepada komisi pembimbing Bapak Prof Dr Ir Cece Sumantri, MAgrSc selaku ketua, Bapak Prof Dr Ir Ronny Rahman Noor, MRurSc dan Bapak Dr Ir Rudy Priyanto selaku anggota yang telah banyak meluangkan waktu, bimbingan, dorongan semangat dan masukan, dalam penelitian maupun penulisan disertasi ini. Ucapan terima kasih juga penulis haturkan kepada Kepala Laboratorium Genetika Molekuler Ternak Bapak Prof Dr Ir Muladno, MSA berserta staf (Eryk Andreas, SPt MSi, Isyana, S Pt dan Selvi) yang telah memberikan izin dan kesempatan kepada penulis untuk melaksanakan penelitian di Laboratorium Genetika Molekuler Ternak. Ucapan yang sama juga penulis haturkan kepada Kepala Laboratorium Terpadu IPB Baranang Siang dan staf (Bu Ani dan Mbak Rita). Kepada Dr Ir Salundik, MSi selaku ketua Program Studi ITP serta staf (Bu Ade dan Mbak Okta) terima kasih atas pelayanan administrasi yang ramah selama penulis menempuh studi.
Ucapan terima kasih juga penulis sampaikan kepada Direktorat Jenderal Pendidikan Tinggi Kementerian Pendidikan Nasional atas Beasiswa BPPS 2010-2013 dan Bantuan Biaya Pendidikan tahun 2014 dari Direktorat Pendidikan Tinggi Islam, Kementerian Agama RI. Terlaksananya penelitian ini juga tidak terlepas dari bantuan biaya penelitian dari Lembaga Penelitian dan Pengembangan Universitas Islam Negeri Sultan Syarif Kasim Riau melalui SK Rektor Nomor: 988/R/2013 untuk itu penulis haturkan banyak terima kasih kepada mantan Rektor UIN Suska Riau Prof Dr HM Nazir dan mantan Kepala LPPM UIN Suska Riau Bapak Drs Husni Thamrin MS. Ucapan terima kasih juga penulis sampaikan kepada mantan Dekan Fakultas Pertanian dan Peternakan UIN Suska Riau Ibu Ir Eniza Saleh, MSi yang telah memberikan bantuan biaya publikasi melalui DIPA Fakultas Pertanian dan Peternakan UIN Suska Riau tahun 2014.
Ucapan terima kasih juga disampaikan kepada Isnandar Putra, SPt dan drh Hanif dari Dinas Peternakan Provinsi Sumatera Barat yang telah banyak membantu dalam pengambilan sampel domba ekor tipis sumatera begitu juga kepada Holfinaldi, SPt atas luangan waktu untuk survey lokasi penelitian. Berikut kepada Bapak Saharudin dan peternak domba di Kecamatan Koto Tangah Padang juga diucapkan terima kasih. Ucapan terima kasih juga penulis sampaikan kepada Bapak Dr Ir Agus Susanto, MSc beserta istri atas persaudaraan dan diskusi molekulernya selama ini. Kepada ibu Ir Sri Rahayu, MS juga diucapkan terima kasih atas bantuan materi penelitian domba garut. Semoga Allah SWT membalas segala kebaikan yang telah Bapak dan Ibu berikan selama ini.
Ungkapan terima kasih yang tulus penulis haturkan kepada yang tercinta Ayahnda H Heldi Syair dan Ibunda Hj Wasni Nawas, kakak/abang, abang/kakak ipar serta seluruh keluarga besar atas segala doa, kasih sayang, dukungan baik moril maupun materil serta motivasi yang diberikan sehingga disertasi ini dapat diselesaikan. Ucapan terima kasih juga penulis sampaikan kepada suami Edwin Perwira, ST MSc M Eng dan kedua putriku Raisha Adiva Khalila dan Naira Aliya Rafifa, terima kasih atas izin kuliah, doa, pengertian, kasih sayang dan dukungan yang diberikan selama ini. Berikut ucapan terima kasih juga penulis tujukan kepada semua pihak yang telah membantu yang tidak dapat penulis sebutkan satu per satu, untuk itu penulis memohon maaf yang sebesar-besarnya.
Semoga karya ilmiah ini bermanfaat.
Bogor, Januari 2015
DAFTAR ISI
DAFTAR TABEL DAFTAR GAMBAR DAFTAR LAMPIRAN 1 PENDAHULUAN 1 Latar Belakang 1 Perumusan Masalah 2 Tujuan Penelitian 3 Manfaat Penelitian 3 Hipotesis 42 IDENTIFIKASI KERAGAMAN GEN LECITHIN CHOLESTEROL
ACYLTRANSFERASE PADA DOMBA LOKAL INDONESIA 5
Pendahuluan 6
Bahan dan Metode 8
Hasil dan Pembahasan 11
Simpulan 19
3 IDENTIFIKASI KERAGAMAN GEN LIPOPROTEIN LIPASE PADA
DOMBA EKOR TIPIS SUMATERA DAN DOMBA GARUT 20
Pendahuluan 21
Bahan dan Metode 25
Hasil dan Pembahasan 27
Simpulan 32
4 ASOSIASI KERAGAMAN GEN LIPOPROTEIN LIPASE c.192
DENGAN KUALITAS MARBLING DAGING DOMBA GARUT 33
Pendahuluan 34
Bahan dan Metode 35
Hasil dan Pembahasan 38
Simpulan 46
5 PEMBAHASAN UMUM 47
6 SIMPULAN DAN SARAN 50
DAFTAR PUSTAKA 51
xviii
DAFTAR TABEL
2.1 Diplotipe gen LCAT ekson 6 domba lokal Indonesia 12 2.2 Frekuensi genotipe, frekuensi gen, nilai heterosigositas dan nilai
chi kuadrat gen LCAT pada posisi basa c.742 domba lokal Indonesia 17 2.3 Frekuensi genotipe, frekuensi gen, nilai heterosigositas dan nilai
chi kuadrat gen LCAT pada posisi basa c.770 domba lokal Indonesia 17 2.4 Frekuensi genotipe, frekuensi gen, nilai heterosigositas dan nilai
chi kuadrat gen LCAT pada posisi basa c.882 domba lokal Indonesia 18 3.1 Diplotipe sekuens bagian dari 5’UTR dan ekson 1 gen LPL domba
ekor tipis sumatera dan domba garut 30
3.2 Frekuensi genotipe dan frekuensi gen LPL domba ekor tipis sumatera
dan domba garut 31
3.3 Nilai heterosigositas dan nilai chi kuadrat gen LPL domba ekor tipis
sumatera dan domba garut 32
4.1 Kandungan lemak, kolesterol dan komposisi asam lemak domba garut
pada genotipe gen LPL yang berbeda 38
4.2 Kandungan lemak dan kolestrol beberapa breed domba 39 4.3 Kandungan ALJ, ALTJT, ALTJG dan rasio ALTJG:ALJ beberapa breed
domba 40
4.4 Profil asam lemak marbling domba garut berdasarkan genotipe gen LPL
yang berbeda (%) 42
4.5 Kandungan lemak, kolesterol dan profil asam lemak marbling domba garut berdasarkan genotipe gen LPL yang berbeda (mg/100g) 43 4.6 Profil asam lemak pada breed domba lainnya 44
DAFTAR GAMBAR
1.1 Alur kerangka berpikir penelitian 3
2.1 Peranan LPL dan LCAT dalam transport lemak dan kolesterol dalam tubuh
ternak 7
2.2 Struktur gen LCAT, terdiri atas 6 ekson (I, II, III, IV, V dan VI) dan
5 intron 7
2.3 Sekuen amplikon gen LCAT (gen bank GQ.150556.1) 10 2.4 Hasil amplifikasi PCR gen LCAT ekson 6 domba lokal Indonesia 12 2.5 Aligment sekuens gen LCAT ekson 6 domba lokal Indonesia 13 2.6 Potongan sekuens gen LCAT ekson 6 yang menunjukkan terjadinya 14
mutasi pada posisi basa c.742, c.770 dan c.882
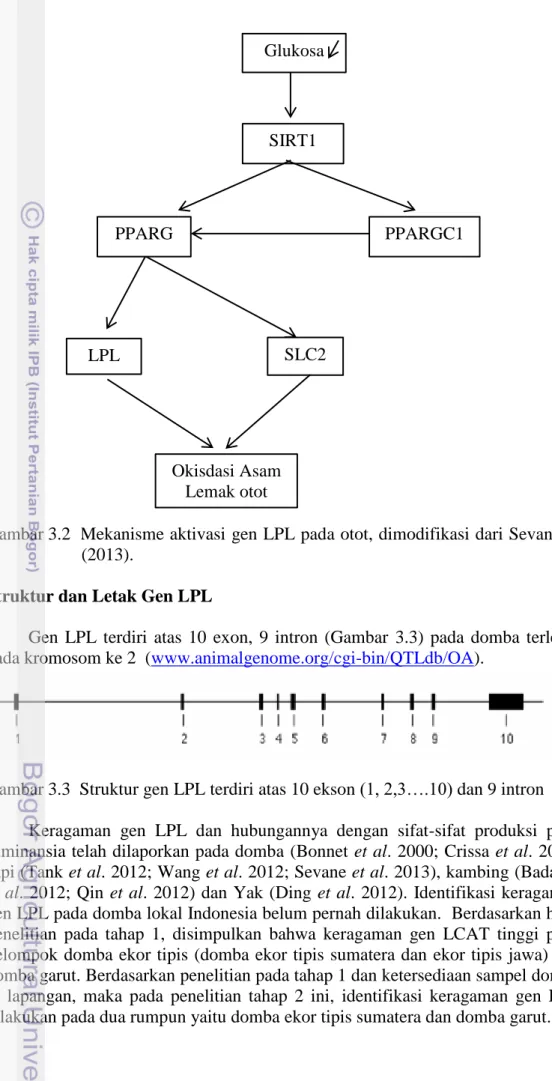
3.1 Transpor lemak ke jaringan pada ternak ruminansia dan non ruminansia 23
3.2 Mekanisme aktivasi gen LPL pada otot 24
3.3 Struktur gen LPL, terdiri atas 10 ekson (1,2, 3…10) dan 9 intron 25 3.4 Amplikon sekuen gen LPL (gen bank X.68308.1) 24
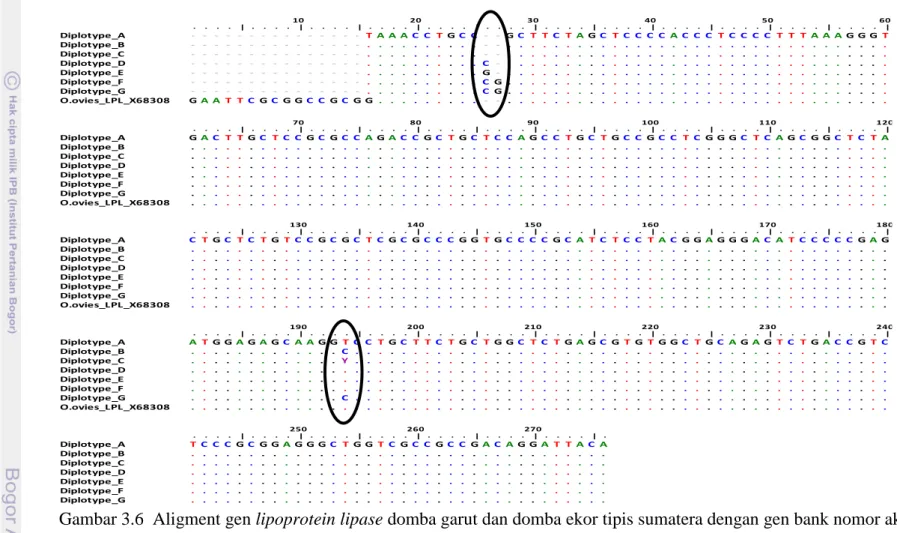
3.5 Hasil amplifikasi PCR gen LPL daerah 5’UTR dan bagian ekson 1 27 3.6 Aligmnet gen lipoprotein lipase domba garut dan domba ekor tipis
sumatera dengan gen bank nomor akses X.68308.1 28 3.7 Parsial sekuens gen LPL yang menunjukkan insersi pada posisi basa
c.26 dan c.27 29
3.8 Aligment sekuens protein gen LPL domba garut dan domba ekor tipis
sumatera 30
3.9 Parsial sekuens gen LPL yang menunjukkan adanya mutasi pada posisi
c.192 T>C pada domba garut 31
4.1 Otot longissimus dorsi 36
4.2 Nilai desirable fatty acid (DFA), atherogenicity index dan rasio
(stearat+oleat): palmitat marbling domba garut 41
DAFTAR LAMPIRAN
1 Ovis aries lecithin-cholesterol acyltransferase (LCAT) gene, exons 2, 3, 6
dan partial cds gen bank: GQ150556.1 58
1 PENDAHULUAN
Latar BelakangPeningkatan konsumsi daging di Indonesia diikuti dengan permintaan masyarakat akan penyediaan daging ASUH (Aman, Sehat, Utuh dan Halal), ramah lingkungan dan dijamin keberlanjutannya yang memiliki daya saing dan sesuai dengan kebutuhan pasar domestik. Konsumsi daging domba dan kambing di Indonesia masih sangat rendah, yaitu sekitar 5% dari total kebutuhan daging atau setara dengan 0.24 g/kapita/tahun (Inounu 2011). Direktorat Jenderal Peternakan telah melaporkan peningkatan populasi domba secara Nasional selama 5 tahun terakhir (2011-2014) yaitu 10.725.000 ekor; 11.791.000 ekor; 13.420.000 ekor; 14.926.000 ekor dan 15.716.000 ekor secara berturut-turut. Peningkatan populasi domba ini merupakan suatu potensi untuk memenuhi permintaan daging merah yang terus meningkat dari tahun ke tahun. Masih rendahnya konsumsi daging kambing dan domba karena harga daging yang relatif tinggi dan berkembang persepsi pada masyarakat bahwa daging kambing dan domba memiliki kandungan kolesterol tinggi, kandungan asam lemak jenuh tinggi dan bau khas yang sulit dihilangkan.
Lemak di dalam tubuh ternak ditemukan pada jaringan adipose, hati, plasma, sel darah merah, otot rangka serta otak dan jaringan syaraf (Leat 1983). Jaringan adipose ditemukan dalam bentuk lemak visceral, lemak subkutan, lemak intermuskular dan lemak intramuskuler (marbling) (Kauffman dan Breidenstein 1994). Lemak intramuskuler merupakan butiran-butiran lemak yang ditemukan di dalam otot (Aberle et al. 2001). Lemak intramuskuler memberikan pengaruh terhadap penilaian konsumen terhadap daging karena dapat mempengaruhi keempukan, juiciness dan flavor (Miller 1994). Disisi lain keberadaan lemak di dalam makanan juga merupakan poin penting dalam memilih jenis makanan yang akan dikonsumsi terutama bagi penderita penyakit-penyakit tertentu.
Konsumsi daging yang mengandung asam lemak jenuh (ALJ) tinggi terutama laurat dan miristat dapat mengakibatkan otot rentan terhadap resistensi insulin sehingga timbul hiperinsulinemia atau meningkatkan produksi kolesterol oleh hati (Cheeke dan Dierenfeld 2010). Begitu juga mengkonsumsi daging dengan kandungan kolesterol tinggi berdampak timbulnya atherosclerosis atau penebalan pembuluh darah ke jantung. Rasio asam lemak tak jenuh ganda (ALTJG): ALJ yang tinggi dan kandungan kolesterol rendah pada daging domba dibutuhkan sebagai pangan fungsional bagi penderita penyakit-penyakit tertentu.
Deposisi lemak pada jaringan ditentukan oleh keseimbangan proses-proses yang berlangsung di dalam tubuh meliputi lipogenic, lipolitic, transpor asam lemak dan jumlah asam lemak yang digunakan. Keseimbangan proses-proses tersebut ditentukan oleh jumlah lemak yang dimakan, sintesis de novo asam lemak, sintesis triasilgliserol, degradasi lipid dan proses transpor asam lemak (Zhao et al. 2010).
Distribusi lemak, komposisi asam lemak dan tipe serabut otot, sekitar 35% dipengaruhi oleh genetik (Williams 2008). Karamichou et al. (2006) menyatakan bahwa nilai heritabilitas komposisi asam lemak marbling domba scottish blackface berkisar sedang sampai tinggi, mengindikasikan bahwa seleksi pada tetua dapat meningkatkan performans asam lemak pada turunannya.
2
Seleksi terhadap kualitas marbling menggunakan metode konvensional sangat sulit dan relatif mahal sehingga dalam program seleksi berdasarkan fenotipe, kualitas marbling tidak umum dilakukan. Alternatif yang dapat dilakukan adalah melalui seleksi genom. Seleksi genom diawali dengan penemuan kandidat gen yang berasosiasi dengan sifat-sifat yang diinginkan. Salah satunya melalui eksplorasi single nucleotide polymorphisms (SNPs) untuk mengetahui genotipe ternak dan selanjutnya dihubungkan dengan sifat-sifat yang dijadikan sebagai kriteria seleksi. SNP merupakan variasi sekuen DNA yang muncul ketika satu nukleotida (A, T, C atau G) berbeda dari sekuen pada umumnya.
Guo et al. (2014) melaporkan bahwa kualitas marbling sapi dan domba ditentukan oleh poligen dan ekspresinya dipengaruhi oleh faktor lingkungan seperti penyakit dan pakan. Keberadaan dan aktivitas enzim dalam tubuh ternak disandikan oleh gen. Diantara enzim yang berperan terhadap kandungan kolesterol dan profil asam lemak adalah lecithin cholesterol acyltransferase (LCAT) dan lipoprotein lipase (LPL).
Lecithin cholesterol acyltransferase adalah soluble enzyme yang mampu mengonversi kolesterol dan lecitin menjadi ester kolesterol dan lisolecitin pada permukaan high density lipoprotein (HDL) dan berperan penting dalam metabolisme lipoprotein, terutama dalam proses reverse cholesterol transport. Enzim ini disintesis di hati tapi sirkulasinya pada plasma darah sebagai komplek komponen HDL. Ketiadaan enzim ini menyebabkan akumulasi kolesterol bebas pada jaringan daging dan darah.
Lipoprotein lipase adalah enzim kunci yang berperan utama pada metabolisme dan transpor lipoprotein serta memberikan pengaruh penting pada level trigliserida darah, mengontrol partisi trigliserida pada jaringan adipose dan otot sehingga meningkatkan penyimpanan lemak atau menyediakan energi dalam bentuk asam lemak untuk pertumbuhan otot (Ren et al. 2002; Dunner et al. 2013).
Identifikasi keragaman gen LCAT dan LPL diperlukan dalam rangka mencari penanda genetik (genetic marker) yang dapat digunakan sebagai Marker Assisted Selection (MAS) untuk menghasilkan dan mengembangkan domba-domba lokal yang mampu menghasilkan daging yang aman dan sehat untuk dikonsumsi.
Perumusan Masalah
Kualitas lemak daging mempengaruhi penilaian konsumen terhadap daging masak. Pada umumnya lemak mengandung gliserol ester, kolesterol, phospholipid dan asam lemak. Lemak disusun atas rangkaian asam lemak yang dikelompokkan ke dalam 3 kategori berdasarkan ikatan rangkap yang dimilikinya yaitu asam lemak jenuh (ALJ), asam lemak tak jenuh tunggal (ALTJT) dan asam lemak tak jenuh ganda (ALTJG). Timbunan lemak pada otot ditentukan oleh keseimbangan dari proses yang berlangsung dalam tubuh. Keseimbangan proses ini juga dipengaruhi oleh jumlah lemak yang dimakan.
Proses pembentukan lemak dan deposisi lemak di dalam tubuh dipengaruhi oleh peran serangkaian enzim yang saling bekerja sama di dalam tubuh. Enzim disandikan oleh gen penyandi. Setiap mutasi yang muncul pada suatu gen dapat mempengaruhi fungsi dari protein yang disandikan dan atau tidak dapat menghasilkan protein yang disandikan. Keaneka ragaman rumpun domba lokal Indonesia berpotensi memiliki gen-gen potensial yang perlu dieksplorasi lebih
lanjut. Eksplorasi keragaman gen-gen potensial yang berpengaruh terhadap kualitas marbling sapi ciamis dan sapi PO telah dilaporkan Hilmia (2013) melalui keragaman gen steoryl CoA desaturase, namun keragaman gen belum mempengaruhi profil asam lemak. Eksplorasi terhadap keragaman gen-gen potensial terhadap kualitas marbling domba lokal Indonesia belum pernah dilaporkan selama ini. Diantara gen yang diduga berhubungan dengan kualitas marbling adalah LCAT dan LPL. Alur kerangka berpikir penelitian disajikan pada Gambar 1.1.
Gambar 1.1 Alur kerangka berpikir penelitian
Tujuan Penelitian Tujuan penelitian ini adalah:
1. Mengidentifikasi keragaman gen LCAT pada domba lokal Indonesia
2. Mengidentifikasi keragaman gen LPL pada domba ekor tipis sumatera dan domba garut
3. Menganalisis hubungan perbedaan genotipe gen LPL terhadap kualitas marbling otot longissimus dorsi domba garut.
Manfaat Penelitian Keluaran dari penelitian ini adalah;
1. Menginformasikan keragaman gen LCAT dan gen LPL pada domba lokal Indonesia sebagai salah satu upaya dalam mengeksplorasi sumber daya genetik domba lokal Indonesia yang dapat dijadikan sebagai data base dalam pengembangan domba lokal Indonesia di masa yang akan datang.
2. Ditemukannya penanda genetik penentu kualitas lemak intramuskuler daging domba yang dapat dijadikan sebagai Marker Assisted Selection (MAS) dalam menghasilkan ternak domba dengan kualitas lemak intramuskuler yang baik.
4
3. Dalam jangka panjang dapat membentuk breed domba lokal yang menghasilkan daging yang menyehatkan.
Hipotesis Hipotesis dalam penelitian ini adalah:
1. Adanya keragaman genotipe gen LCAT dan gen LPL pada domba lokal Indonesia.
2. Keragaman genotipe gen LCAT dan gen LPL akan memberikan pengaruh yang berbeda terhadap kandungan lemak total, kolesterol dan profil asam lemak otot longissimus dorsi domba garut.
2 IDENTIFIKASI KERAGAMAN GEN LECITHIN CHOLESTEROL
ACYLTRANSFERASE PADA DOMBA LOKAL INDONESIA
Abstrak
Lecithin cholesterol acyltransferase (LCAT) adalah soluble enzyme yang mampu mengonversi kolesterol dan lesitin menjadi ester kolesterol dan lisolesitin pada permukaan high density lipoprotein dan berperan dalam metabolisme lipoprotein. Penelitian ini bertujuan untuk mengeksplorasi single nucleotide polymorphisms (SNPs) gen LCAT pada domba lokal Indonesia. Total 118 DNA genom domba lokal Indonesia, terdiri dari domba ekor tipis sumatera (43 ekor); domba garut (19 ekor); domba ekor tipis jawa (17 ekor); domba ekor gemuk jawa (6 ekor), domba pulau rote (7 ekor); domba kissar (7 ekor); domba sumbawa (10 ekor) dan domba lembah palu (9 ekor) digunakan dalam penelitian ini. Amplifikasi DNA genome menggunakan Polymerase Chain Reaction pada fragment ekson 6 gen LCAT menghasilkan amplikon dengan panjang 250 bp dan metode direct sequencing digunakan untuk mengidentifikasi keragaman sekuens. Hasil sekuens dianalisis menggunakan software Bioedit dan MEGA 5.2. Kemudian sekuens disejajarkan dengan metode Clustal W dan selanjutnya diBLAST dengan gene bank nomor akses GQ.150556.1. Hasil penelitian menunjukkan ditemukan tiga SNPs baru yaitu pada posisi basa c.742 C>T, c.770 T>A dan c.882 C>T. Substitusi sitosina menjadi timina c.742 merupakan synonymous mutation; timina menjadi adenina c.770 dan sitosina menjadi timina c.883 merupakan non synonymous mutation. Kata kunci: Domba, gen LCAT, PCR, sekuens, SNPs
Abstract
Lecithin Cholesterol Acyltransferase (LCAT) was a soluble enzyme that converted cholesterol and lecithin to cholesteryl esters and lysolecithins on the surface of high density lipoprotein and played in lipoprotein metabolism. The research was aimed to explore single nucleotide polymorphisms (SNPs) of LCAT gene in Indonesian local sheeps. A total of 118 genomic DNA of Indonesian local sheeps used in this study, consisted of sumatran thin tail ed sheep (43 heads); garut sheep (19 heads); javanese thin tail ed sheep (17 heads); javanese fat tail ed sheep (6 heads), rote island sheep (7 heads); kissar sheep (7 heads); sumbawa sheep (10 heads) and lembah palu sheep (9 heads). Polymerase chain reaction was used to amplify genomic DNA for exon 6 (250 bp) and direct sequencing method was used to identify polymorphism sequences. The results of sequence were analyzed with BioEdit and MEGA 5.2 software. The sequences were aligned with Clustal W method and BLAST sequence obtained from Gene Bank with accession number GQ 150556.1. The results were showed three novel SNPs, i.e. c.742C>T, c.770 T>A dan c.882 C>T. Substitution cytosine to thymine c.742 is a synonymous mutation; thymine to adenine c.770 and cytosine to thymine c.882 are non synonymous mutation.
6
Pendahuluan
Domba merupakan salah satu ternak potong yang memiliki nilai penting bagi masyarakat Indonesia terutama dalam menyediakan daging segar, hewan kurban (Inounu 2011), sosial, budaya dan sumber gen yang digunakan dalam memperbaiki mutu ternak lokal melalui persilangan diantara ternak lokal atau dengan bangsa eksotik lainnya (Sumantri et al. 2007). Menurut FAO (2002), ternak lokal penting dilindungi karena telah beradaptasi dengan lingkungan setempat, dapat bertahan pada kondisi pakan seadanya dan lebih tahan terhadap penyakit dan parasit.
Domba lokal Indonesia memiliki keragaman yang tinggi berdasarkan karakter morfometrik (Sumantri et al. 2007), keragaman DNA mikrosatelit (Sumantri et al. 2008a dan Jakaria et al. 2012). Domba merupakan salah satu ternak potong di Indonesia. Berkembangnya persepsi di masyarakat bahwa daging domba mengandung kolesterol dan kandungan asam lemak jenuh (ALJ) tinggi, harga yang relatif mahal, bau daging yang kurang diminati dan sulit untuk dihilangkan merupakan faktor-faktor yang diduga sebagai penyebab rendahnya konsumsi daging domba secara Nasional yaitu sekitar 5% atau setara dengan 0.24 g/kapita/tahun. Konsumsi makanan dengan kandungan kolesterol dan asam lemak jenuh tinggi ditenggarai sebagai pemicu penyakit jantung. Kromhout et al. (1995), telah melaporkan bahwa konsumsi makanan mengandung asam lemak jenuh dan kolesterol yang tinggi merupakan faktor penting penentu angka penderita penyakit jantung pada 9 negara.
Kandungan lemak tubuh ternak meningkat di antara awal pertumbuhan sampai saat dipotong dan proporsi asam lemak juga turut berubah (Wood et al. 2008). Kandungan ALJ dan asam lemak tak jenuh tunggal (ALTJT) meningkat sangat cepat seiring dengan peningkatan lemak, berbeda dengan asam lemak tak jenuh ganda (ALTJG) cenderung menurun sehingga rasio ALTJG:ALJ juga akan menurun (Smeet et al. 2004). Berbeda dengan daging dengan kandungan lemak sedikit (lean), proporsi ALTJG mayor lebih tinggi (Wood et al. 2008). Beberapa penelitian menunjukkan bahwa kandungan kolesterol dan asam lemak diatur oleh gen-gen fungsional.
Lecithin Cholesterol Acyltransferase (LCAT) adalah enzim kunci yang berperan dalam katabolisme ekstra seluler dari lipoprotein darah, disintesis di hati dan disekresikan ke dalam plasma darah. Enzim ini bekerja mengonversi kolesterol dan lesitin menjadi ester kolesterol dan lisolesitin pada permukaan high density lipoprotein (HDL), terutama dalam proses reverse cholesterol transport (Kaplanova et al. 2010; Crisa et al. 2010; Qiao et al. 2010). Defisiensi enzim ini ditenggarai dapat mengakibatkan akumulasi kolesterol bebas di dalam darah dan jaringan (Klein et al. 1993; Kaplanova, et al. 2010; Qiao et al. 2010).
Metabolisme Kolesterol dan Peran Lecithin Cholesterol Acyl Transferase Kolesterol di dalam tubuh berasal dari pakan dan sintesis de novo pada hati dan jaringan lainnya. Kolesterol ditemukan dalam bentuk kolesterol bebas atau kombinasi kolesterol dengan asam lemak rantai panjang (long chain fatty acid) dalam bentuk kolesterol ester. Lipoprotein memiliki peranan utama dalam metabolisme kolesterol (Cheeke dan Dierenfeld 2010). Reaksi LCAT terhadap lipoprotein melewati beberapa tahapan yaitu dimulai dengan pengikatan enzim
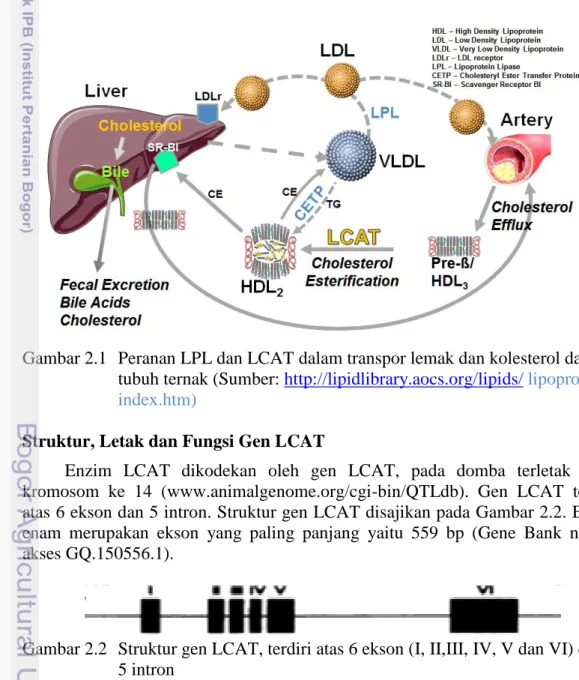
terhadap permukaan lipid, diikuti dengan aktivasi LCAT oleh apolipoprotein, pengikatan substrat lipid dan tahapan katalitik yang mengakibatkan peningkatan produk lipid (Jonas 2000). Kolesterol masuk ke jaringan melalui low density lipoprotein (LDL) dan dipindahkan dari jaringan oleh HDL melalui mekanisme reverse cholesterol transport dengan bantuan enzim LCAT (Cheeke dan Dierenfeld 2010). Saat HDL menerima kolesterol dari membran sel, kolesterol bebas mengalami esterifikasi oleh LCAT. Terdapat dua mekanisme yaitu HDL memindahkan kolesterol bebas dan ester kolesterol ke hati dan kolesterol bebas dan ester kolesterol dibentuk menjadi very low density lipoprotein (VLDL) melalui CETP (Cholesterol Ester Transfer Protein). VLDL yang mengalami hidrolisis dengan perantaraan LPL menjadi LDL. Kolesterol LDL dibuang dari darah melalui interaksi dengan reseptor LDL. Kejadian ini memungkinkan kolesterol masuk ke dalam sel jaringan (Cheeke dan Dierenfeld 2010). Mekanisme peranan LPL dan LCAT dalam transpor lemak dan kolesterol dalam tubuh ternak dijelaskan pada Gambar 2.1.
Gambar 2.1 Peranan LPL dan LCAT dalam transpor lemak dan kolesterol dalam tubuh ternak (Sumber: http://lipidlibrary.aocs.org/lipids/ lipoprot/ index.htm)
Struktur, Letak dan Fungsi Gen LCAT
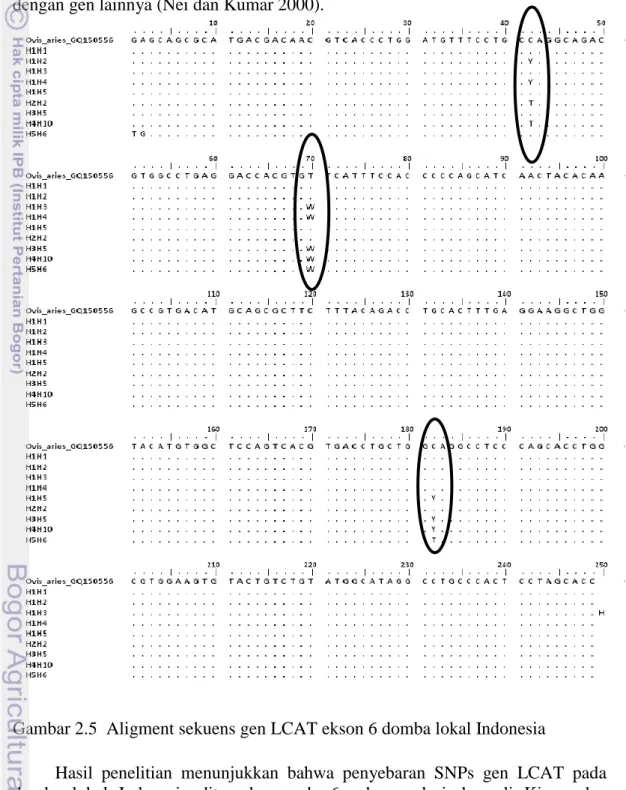
Enzim LCAT dikodekan oleh gen LCAT, pada domba terletak pada kromosom ke 14 (www.animalgenome.org/cgi-bin/QTLdb). Gen LCAT terdiri atas 6 ekson dan 5 intron. Struktur gen LCAT disajikan pada Gambar 2.2. Ekson enam merupakan ekson yang paling panjang yaitu 559 bp (Gene Bank nomor akses GQ.150556.1).
Gambar 2.2 Struktur gen LCAT, terdiri atas 6 ekson (I, II,III, IV, V dan VI) dan 5 intron
8
Beberapa penelitian telah melaporkan adanya keragaman gen LCAT pada ternak yang berhubungan dengan sifat-sifat produksi yaitu pada babi (Kaplanova et al. 2010; Qiao et al. 2010); domba (Crisa et al. 2010; Moioli et al. 2012). Crisa et al. (2010) menemukan adanya satu SNP pada intron 2 g.181 T>C dan 2 SNPs pada exon 6 c.806 G>A dan c.1075 T>C. Mutasi ini berhubungan negatif dengan asam lemak jenuh ganda C18:2 (asam lemak linoleat) dan berhubungan positif
dengan produksi dan kandungan asam lemak jenuh stearat pada susu domba (Crisa et al. 2010; Moioli et al. 2012). Qiao et al. (2010) melaporkan satu SNP pada posisi intron 1 g.266 C>G berhubungan sangat nyata dengan beberapa karakteristik karkas pada babi.
Single nucleotide polymorphism merupakan variasi yang muncul pada sekuen DNA ketika satu nukleotida (A, T, C atau G) berbeda dari sekuen pada umumnya. SNP digunakan untuk mengetahui keragaman suatu gen dan frekuensi genotipe yang muncul pada suatu populasi ternak. Asosiasi keragaman SNP dengan sifat-sifat yang memiliki nilai ekonomis dan memiliki arti penting untuk menemukan penanda genetik (genetic marker) yang dapat digunakan dalam seleksi genom.
Seleksi genom digunakan untuk memprediksi nilai pemuliaan (breeding value) untuk sifat-sifat kuantitatif pada ukuran populasi yang kecil dan akurasi terhadap Estimated Breeding Value (EBV) rendah. Seleksi genom dapat ditingkatkan melalui eksplorasi SNPs pada kandidat lokus dan memperkirakan pengaruh SNPs pada populasi yang berbeda (Dunner et al. 2013).
Langkah awal yang dapat dilakukan adalah melakukan eksplorasi dan identifikasi keragaman gen-gen potensial yang diasumsikan berhubungan dengan sifat-sifat yang diinginkan. Eksplorasi keragaman gen LCAT pada domba lokal Indonesia merupakan upaya untuk menemukan Marker Assisted Selection (MAS) untuk memproduksi dan mengembangkan domba lokal Indonesia di masa yang akan datang.
Bahan dan Metode Tempat dan Waktu
Pengambilan sampel darah domba ekor tipis sumatera dilakukan di Kecamatan Koto Tangah Kota Padang, Sumatera Barat pada bulan Oktober 2012 dan pengambilan sampel darah domba garut dilakukan di Kandang B Fakultas Peternakan IPB Bogor pada bulan Juni 2013. Ekstraksi dan amplifikasi PCR dilakukan di Laboratorium Genetika Molekuler Ternak Fakultas Peternakan IPB dari bulan Oktober 2012 s/d Juni 2013. Identifikasi keragaman segmen gen LCAT ekson 6 dilakukan dengan metode direct sequencing dengan mengirimkan sampel ke First Base Laboratory Juni s/d Agustus 2013.
Materi
Sampel darah domba lokal Indonesia sebanyak 57 sampel darah terdiri dari 43 sampel domba ekor tipis sumatera berasal dari Kecamatan Koto Tangah Padang, Sumatera Barat dan 14 sampel domba garut (Jawa Barat) diambil pada bagian vena jugularis menggunakan venojact. Darah disimpan dalam tabung 5 ml dan ditambahkan ethanol absolut. Selain itu juga digunakan 61 koleksi DNA domba Laboratorium Genetika Molekuler Ternak Fakultas Peternakan IPB terdiri dari domba garut, Jawa Barat (garut-margawati tipe pedaging 5); ekor tipis
jawa-Jawa Barat (Jonggol, 7; MT Farm Bogor, 10); ekor gemuk jawa-jawa-Jawa Timur (Situbundo, 6); Pulau Rote (7); Kissar (7); Sumbawa-Nusa Tenggara Barat (10) dan Lembah Palu (9). Total DNA yang diuji dalam penelitian ini adalah 118 sampel domba lokal Indonesia.
Ekstraksi DNA
Ektraksi DNA menggunakan metode Phenol-chloroform technique (Sambrook et al. 1989) yang telah dimodifikasi. Sebanyak 200 µL sampel darah dipindahkan ke dalam tabung 1.5 mL lalu ditambahkan 1000 µL delution water, divortex dan didiamkan selama ± 20 menit pada suhu ruang. Selajutnya proses presipitasi dengan cara disentrifugasi pada kecepatan 8.000 rpm selama 5 menit. Larutan supernatan yang terbentuk dibuang dan ditambahkan 200 µl 1 x sodium tris EDTA (1 x STE), 40 µl sodium dosesil sulfate (SDS) 10% dan 20 µl Proteinase K (5 mg/ml). Larutan selanjutnya dikocok perlahan pada suhu 550C selama 2 jam. Kemudian ditambahkan larutan phenol 400 µl, 400 µl chloroform:isoamyl alcohol = 24:1 (CIAA) dan 40 µL NaCl 5 M. Tabung selanjutnya dikocok pada suhu ruang selama 1 jam dan disentrifugasi selama 5 menit. Larutan bening selanjutnya dipindahkan ke tabung 1.5 ml yang baru dan ditambahkan 800 µl ethanol absolute, 40 µL NaCl 5 M, dan disimpan pada suhu -20oC selama semalam. Selanjutnya larutan disentrifugasi pada kecepatan 12.000 rpm selama 5 menit, supernatan yang terbentuk dibuang lalu tabung didiamkan dalam keadaan terbuka sampai alkohol yang ada pada tabung hilang. Selanjutnya ditambahkan 100 µL TE (Tris EDTA) 80% pada tabung tersebut dan disimpan pada suhu -200C.
Amplifikasi Gen LCAT Ekson 6 dengan Polymerase Chain Reaction
Sampel DNA diamplikasi menggunakan mesin PCR GeneAmp PCR sistem 9700 dan Master Cycler Personal 22331 Eppendorf, menggunakan primer yang dirancang sendiri menggunakan program Primer Blast dari NCBI (http://www.ncbi.nlm.nih.gov/tools.primer-blast) dengan referensi yang digunakan adalah gen bank nomor akses GQ.150556.1. Primer forward yang digunakan adalah F’5- GAGCAGCGCATGACGACAACG-3’ dan primer reverse F’5- AGGTGCTAGGAGTGGGCAGGC-3’. Posisi primer forward dan primer reverse pada sekuens GenBank GQ.150556.1 mengapit pada fragment ekson 6 gen LCAT pada posisi basa ke 701 sampai 950 dengan panjang product 250 bp (Gambar 2.3).
Setiap reaksi PCR terdiri dari 50 ng (2 – 3 µl) DNA template, 0.25 µM primer forward and reverse, 12.5 µL Dream Tag Green Master Mix dari Thermo Scientific #K1081 ditambah dengan dH2O hingga 25 µL. Kondisi mesin PCR
diatur dengan suhu denaturasi awal pada 950C selama 5 menit dan diikuti dengan 35 siklus yang terdiri dari denaturasi pada suhu 950C selama 30 detik, annealing pada suhu 620C selama 45 detik dan ekstensi pada suhu 720C selama satu menit. Ekstensi akhir dilakukan pada suhu 720C selama 5 menit.
Hasil dari PCR kemudian dielektroforesis menggunakan gel agarose (1.5% agarose/0.5x TBE) menggunakan 200 ng/mL ethidhium bromide (EtBr). Penentuan panjang produk menggunakan ladder marker 100 bp. Elektroforesis berlangsung pada tegangan 100 volt selama 30 menit. Selanjutnya visualisasi gel menggunakan UV transilluminator untuk mengetahui berhasil atau tidaknya
10
proses amplifikasi primer pada gen target. Selanjutnya produk PCR dikirim ke First Base Laboratory Singapore untuk analisis direct sequencing menggunakan Dideoxy Sequencing ABI 3730 XL Automated DNA Sequencer.
Gambar 2.3 Amplikon gen LCAT (gen bank nomor akses GQ.150556.1) dengan panjang 250 bp mengapit sekuens gen LCAT c.701-c.950 merupakan bagian dari ekson 6. Primer ditandai dengan garis bawah, bold dan italic.
Analisis Data
Hasil sekuens fragmen gen LCAT ekson 6 dianalisis menggunakan BioEdit (Hall 2011) dan MEGA versi 5.2 (Kumar et al. 2004) menggunakan metode Clustal W. Selanjutnya hasil sekuens disejajarkan menggunakan BLAST (Basic Local Aligmnet Search Tool) dengan sekuens Gene Bank nomor akses GQ.150556. POP GENE ver.1.31 (Yeh et al. 1999) software untuk menghitung frekuensi gen, frekuensi genotipe, uji chi kuadrat dan nilai heterosigositas pada setiap titik mutasi.
Rumus untuk menghitung frekuensi gen dan frekuensi genotipe menurut Nei dan Kumar (2000) yaitu;
;
Keterangan:
xi = Frekuensi gen ke-i
xj = Frekuensi gen ke-j
= Jumlah sampel dengan genotipe ii = Jumlah sampel dengan genotipe ij
N = Jumlah sampel
Frekuensi genotipe dihitung menggunakan rumus:
;
Keterangan:
= Frekuensi genotipe ke-ii = Frekuensi genotipe ke-jj = Frekuensi genotipe ke-ij
Penyimpangan frekuensi genotipe yang muncul dari keseimbangan Hardy- Weinberg dianalisis menggunakan chi square test ( ) berdasarkan rumus Nei dan Kumar (2000);
∑
Keterangan:
= chi square test
O = Frekuensi genotipe sampel yang diamati E = Frekuensi genotipe harapan
Nilai heterosigositas pengamatan (Ho) dan harapan (He) dihitung
menggunakan POPGENE 32 versi 1.31 software (Yeh et al. 1999):
∑ ∑ ∑ ∑ Keterangan: = Heterosigositas observasi = Heterosigositas harapan = Ukuran populasi efektif
= Frekuensi genotipe AiAj, populasi ke-k
Hasil dan Pembahasan
Single Nucleotide Polymorphisms Gen LCAT Ekson 6 Pada Domba Lokal
Indonesia
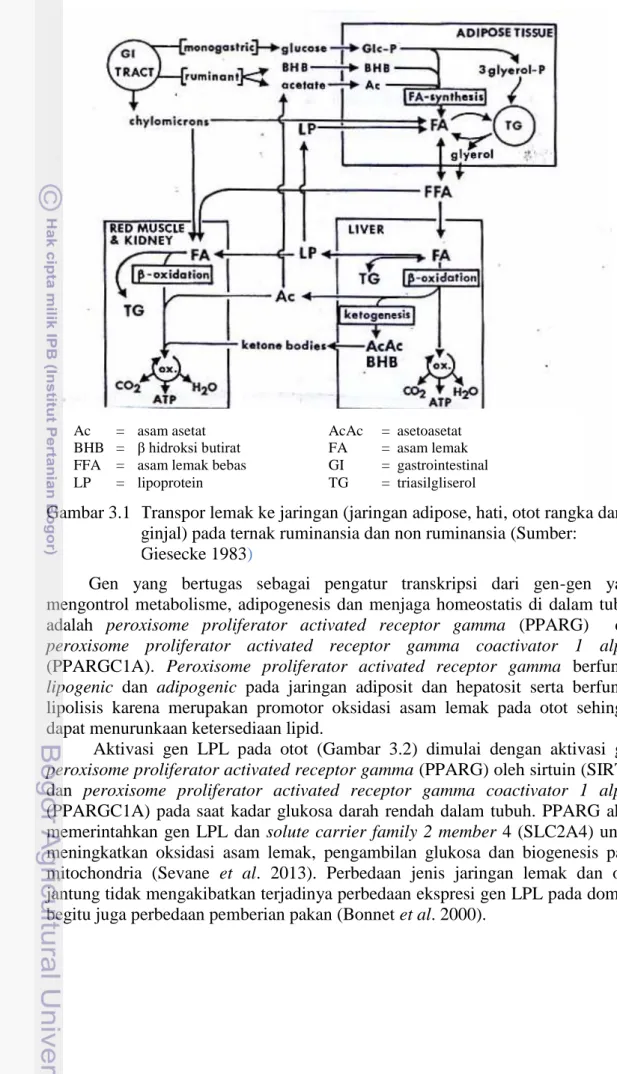
Hasil amplifikasi DNA pada fragmen gen LCAT (Gambar 2.4), menghasilkan amplikon dengan panjang 250 bp, yang mengapit sekuens gen LCAT pada posisi basa ke 701 sampai 950 bp merupakan sebagian dari ekson 6 (Lampiran 1). Hasil aligment 118 sekuens gen LCAT domba lokal Indonesia dengan sekuens gen bank nomor akses GQ.150556.1 (Lampiran 1), menunjukkan 3 titik mutasi baru yaitu pada posisi basa c.742 C>T, c.770 T>A dan c.882 C>T. Tiga SNPs ini membentuk 9 diplotipe (Gambar 2.5) dan tersebar pada 8 rumpun domba lokal Indonesia. Tiga diplotipe dengan frekuensi yang tinggi adalah diplotipe H1H1 (66.10%), H1H3 (16.10%) dan H1H5 (5.08%) (Tabel 2.1). Tiga SNPs yang ditemukan dalam penelitian ini merupakan SNPs baru yang belum pernah dilaporkan oleh peneliti lain. Keragaman gen LCAT pada domba lokal Indonesia bersifat polimorfik pada 6 rumpun yaitu domba ekor tipis sumatera, domba ekor tipis jawa, domba ekor gemuk jawa, domba lembah palu dan domba pulau rote dan monomorfik pada rumpun domba kissar dan domba sumbawa.
Mutasi transisi pada posisi basa c.742, substitusi sitosina menjadi timina merupakan synonymous mutation, tidak merubah asam amino alanina (Ala>Ala) (Gambar 2.6). Mutasi ini ditemukan pada diplotipe H2H2 (4 ekor) dan H4H10(1 ekor), yang tersebar pada domba ekor tipis sumatera (2 ekor); domba garut (2 ekor) dan domba lembah palu (1 ekor). Genotipe heterozigot (CT) ditemukan
12
pada H1H2 dan H1H4 pada domba ekor tipis sumatera (1 ekor) dan domba ekor tipis jawa (4 ekor). Menurut Komar (2007), secara alami, munculnya silent SNP bisa menyandikan sintesis protein dengan sekuens asam amino yang sama, namun dapat mengakibatkan struktur dan fungsi protein yang disandikan berbeda.
Mutasi transversi yang muncul pada posisi basa c.770 yaitu subsitusi timina menjadi adenina dan mutasi transisi pada posisi basa c.882 yaitu subsitusi sitosina menjadi timina (Gambar 7) merupakan non-synonymous mutation, dimana perubahan basa mengakibatkan perubahan penilalanina menjadi isoleusina (Phe>Ile) dan alanina menjadi valina (Ala>Val). Non-synonymous SNPs (nsSNPs), dapat merubah susunan asam amino yang berdampak pada perubahan protein yang terbentuk, pada manusia umumnya berhubungan dengan penyakit keturunan (Bao dan Cui 2005).
01 02 03 04 05 06 07 08 M 09 10 11 12 13 14 15 16
Gambar 2.4 Hasil amplifikasi PCR gen LCAT ekson 6 domba lokal Indonesia dengan panjang produk 250 bp; M= DNA ladder 100 bp; 01,02…..16 = kode sampel domba ekor tipis sumatera
Tabel 2.1 Diplotipe gen LCAT ekson 6 domba lokal Indonesia
Diplotipe Posisi Mutasi Jumlah
individu Frekuensi (%) c.742 c.770 c.882 H1H1 CC TT CC 78 66,10 H1H2 CT TT CC 3 2,54 H1H3 CC AT CC 19 16,10 H1H4 CT AT CC 2 1,69 H1H5 CC TT CT 6 5,08 H2H2 TT TT CC 4 3,39 H3H5 CC AT CT 4 3,39 H4H10 TT AT CT 1 0,85 H5H6 CC AT TT 1 0,85 Total 118 100 SNP pada posisi c.770, tidak ditemukan domba dengan genotipe AA. Heterozigot AT ditemukan pada 5 diplotipe yaitu; H1H3; H3H5; H5H6; H1H4 dan H4H10, tersebar pada rumpun domba ekor tipis sumatera, domba ekor tipis
jawa, domba lembah palu, domba ekor gemuk jawa dan domba garut. Genotipe TT ditemukan pada empat diplotipe yaitu H1H1, H1H2, H1H5 dan H2H2.
SNP pada posisi basa c.882 membentuk 3 genotipe (CC, CT dan TT). Genotip CC ditemukan pada 106 individu dengan 5 diplotipe yaitu H1H1, H1H2, H1H3, H1H4 dan H2H2. Genotipe CT ditemukan pada 3 diplotipe yaitu. H1H5, H3H5 dan H4H10, dimana genotipe TT hanya ditemukan pada satu diplotipe H5H6. Potensi munculnya variasi sekuens nukleotida menghasilkan synonymous mutation dan nonsynonymous mutation bervariasi antara satu gen dengan gen lainnya (Nei dan Kumar 2000).
Gambar 2.5 Aligment sekuens gen LCAT ekson 6 domba lokal Indonesia
Hasil penelitian menunjukkan bahwa penyebaran SNPs gen LCAT pada domba lokal Indonesia ditemukan pada 6 sub populasi, kecuali Kissar dan Sumbawa yang menujukkan monomorfik. Keragaman gen yang tinggi ditemukan
14
pada kelompok domba ekor tipis (domba ekor tipis sumatera dan domba ekor tipis jawa), diikuti domba garut dan domba ekor gemuk jawa. Crissa et al. (2010) melaporkan 3 SNPs pada gen LCAT pada 3 bangsa domba (altamurana, gentile di puglia dan sarda) yaitu pada posisi basa g.181 T>C, c.806 G>A dan c.1075 T>C, dengan keragaman gen yang relatif rendah.
Keterangan: SNP pada posisi basa c.742 C>T merupakan synonymous mutation (Ala>Ala)
Keterangan: SNP pada posisi basa c.770 T>A merupakan non synonymous mutation (Phe>Ile)
Keterangan: SNP pada posisi basa c.882 C>T merupakan non synonymous mutation (Ala>Val)
Gambar 2.6 Potongan sekuens gen LCAT ekson yang menunjukkan terjadinya mutasi pada posisi basa c.742, c.770 dan c.882
Setiap mutasi yang muncul berpengaruh terhadap satu atau lebih peranan protein yang disandikan. Peranan protein yang mungkin dipengaruhi oleh mutasi yang timbul adalah stabilitas atau folding protein, ligand binding protein, katalisis, regulasi dengan allosteric dan mekanisme lainnya serta modifikasi post translational protein (Wang dan Moult 2001; Teng et al. 2008). Namun, peranan 3 titik mutasi gen LCAT ekson 6 pada domba lokal Indonesia belum diketahui. Crissa et al. (2010) menemukan 2 titik mutasi pada ekson 6 yaitu pada posisi basa c.806 (G>A) dan c.1075 (T>C). Mutasi pada posisi basa c.806 merupakan non-synonymous mutation (Asp>Asn), yang berpengaruh terhadap rasio asam lemak tak jenuh C18:2 (linoleat) dan C18:3 (linolenat) pada susu domba (Crissa et al. 2010)
dan menurunkan produksi susu dan berpengaruh terhadap kandungan asam lemak stearat dan oleat (Moili et al. 2012). Mutasi pada bagian intron 1 gen LCAT c.266 G>C pada babi berhubungan dengan peningkatan kandungan kolesterol darah babi (Kaplanova et al. 2010).
Menurut Uchida et al. (1995) bahwa penurunan aktivitas LCAT mengakibatkan penurunana fertilitas pada induk sapi yang berhubungan dengan fatty liver karena esterifikasi kolesterol oleh LCAT, penting untuk transpor kolesterol dari hati ke jaringan peripheral, seperti corpus luteum dan karena kolesterol merupakan sumber bagi sintesis progesteron. Hasil penelitian in vivo menunjukkanbahwa aktivasi LCAT oleh Apo A-I, namun mekanisme pastinya belum diketahui (Rousset et al. 2010)
Insersi adenina menghasilkan frameshift mutation pada posisi basa g.214 pada manusia, mengubah dalam porsi yang besar enzim LCAT yang dihasilkan, yaitu daerah protein dan aktivitas putative lipase (Bender et al. 2007). Dua titik mutasi yang ditemukan pada manusia yaitu mutasi frameshift pada asam amino ke-83 (tirosina) menyebabkan terpotongnya enzim pada asam amino ke-82 yang menyebabkan jika disekresikan enzim tersebut, namun tidak dapat berfungsi secara normal. Perubahan asam amino ke 156 (Tyr>Asn) membentuk amphipathic helix yaitu residu dalam fase hydrophobic menghasilkan pH yang rendah. LCAT berperan penting pada formasi dan pematangan HDL dan pada tahapan intra vascular yaitu pada tahapan reverse cholesterol transpor (Savel et al. 2012). Frekuensi Gen, Frekuensi Genotipe dan Nilai Heterosigositas Gen LCAT Ekson 6 Domba Lokal Indonesia
Keragaman genetik dalam populasi dapat diketahui melalui dua ukuran keragaman yaitu proporsi polimorfisme gen dalam populasi dan rata-rata proporsi individu heterozigot dalam setiap lokus (Nei dan Kumar 2000). Identifikasi keragaman genetik digunakan untuk mengetahui dan melestarikan bangsa-bangsa pada populasi terkait dan kaitannya dengan sifat-sifat tertentu untuk menjamin keamanan dan ketersediaan bahan pangan yang berkesinambungan (Blott et al. 1998); mempelajari genetika populasi dan genetika evolusi (Nei dan Kumar 2000); dapat digunakan dalam menentukan hubungan antar sub populasi yang terfragmentasi dalam suatu species (Hartl dan Clark 1997) dan dapat mendeteksi alel-alel positif yang berhubungan dengan sifat-sifat yang diinginkan (Sumantri et al. 2011).
Keragaman genetik antara sub populasi dapat diketahui melalui persamaan atau perbedaan frekuensi gen atau frekuensi alel diantara sub populasi (Li et al. 2000). Frekuensi gen atau frekuensi alel adalah ukuran frekuensi relatif dari gen atau alel pada suatu populasi (Nei dan Kumar 2000). Frekuensi alel menunjukkan keragaman genetik dari sebuah populasi spesies atau equivalent dengan kekayaan/keragaman gen tersebut pada poolnya.
Hasil analisis menunjukkan keragaman gen yang tinggi pada lokus c.742 (Tabel 2.2), ditemukan pada domba ekor tipis sumatera, domba ekor tipis jawa, domba garut dan domba lembah palu. Frekuensi gen C merupakan gen dominan dengan kisaran antara 88.20% - 98.80% dengan nilai rataan 95.70% dan frekuensi gen T berkisar 1.20 % - 11.80%, yang mengindikasikan lokus tersebut polimorfik (Tabel 2.2). Nilai rataan frekuensi genotipe CC (94.10%) lebih tinggi dibandingkan CT (3.20%) dan TT (2.70%). Genotipe CC merupakan genotipe
16
dominan yang ditemukan pada semua rumpun domba. Frekuensi genotipe lokus SNP c.742 berada pada ketidak seimbangan Hardy-Weinberg pada domba ekor tipis sumatera, domba garut dan domba lembah palu (P<0.01), kecuali pada domba ekor tipis jawa berada dalam keseimbangan Hardy-Weinberg (P>0.05).
Hal ini diperkuat dengan hasil penghitungan nilai heterosigositas yang didapat pada rumpun domba ekor tipis sumatera, garut dan lembah palu, dimana nilai heterosigositas harapan lebih besar dibandingkan nilai heterosigositas pengamatan. Nilai heterosigositas pengamatan lebih kecil dibandingkan nilai heterosigositas harapan mengindikasikan telah terjadi perkawinan berkerabat yang intensif pada populasi tersebut (Tombasco et al. 2003)
Penyimpangan frekuensi genotipe dalam suatu populasi dapat disebabkkan karena adanya mutasi, migrasi, perkawinan terarah, seleksi dan jumlah sampel yang relatif kecil. Suatu populasi berada dalam keseimbangan Hardy-Weinberg jika frekuensi genotipe dan frekuensi gen tetap dari satu generasi ke generasi berikutnya (Nei dan Kumar, 2000; Noor 2004).
Mutasi transversi pada posisi basa c.770, ditemukan 2 genotipe yaitu genotipe AT dan TT kecuali pada domba kissar dan domba sumbawa hanya satu genotipe (TT). Nilai rataan frekuensi genotipe TT (80.70%) lebih tinggi dibandingkan genotipe AT (19.30%) dan AA (0.00%). Tingginya frekeunsi genotipe TT menghasilkan tingginya kisaran frekuensi gen T (76.50% - 94.20%), dimana frekuensi genotipe berada dalam keseimbangan Hardy Weinberg pada lokus SNP c.742 ini sejalan dengan hasil analisis keseimbangan Hardy-Weinberg yang menunjukkan semua populasi berada dalam keseimbangan Hardy-Weinberg (P>0.05) dan nilai heterosigositas observasi dan harapan relatif sama (Tabel 2.3).
Mutasi transisi pada posisi basa c.882 C>T ditemukan pada rumpun domba ekor tipis sumatera, domba ekor tipis jawa, domba garut dan domba pulau rote. Rataan frekuensi genotipe CC (89.60%) lebih tinggi daripada CT (9.80%) dan TT (0.7%) (Tabel 2.4). Berdasarkan uji keseimbangan Hardy-Weinberg pada empat rumpun domba, frekuensi genotipe berada dalam ketidak seimbangan Hardy-Weinberg (P<0.01) pada rumpun domba ekor tipis sumatera, domba ekor tipis jawa dan domba garut, kecuali pada rumpun domba pulau rote. Hasil ini sejalan dengan hasil penghitungan nilai heterosigositas yang menunjukkan bahwa nilai heterosigositas observasi lebih rendah dibandingkan nilai heterosigositas harapan.
Hasil penelitian menunjukkan bahwa ditemukannya keragaman gen LCAT ekson 6, pada domba lokal Indonesia kecuali pada sub populasi domba kissar dan domba sumbawa. Penyimpangan frekuensi genotipe pada posisi basa c.742 ditemukan pada domba ekor tipis sumatera, domba garut dan domba lembah palu dan pada posisi basa c.882 ditemukan pada domba ekor tipis sumatera, domba ekor tipis jawa dan domba garut.
Tabel 2.2 Frekuensi genotipe, frekuensi gen, nilai heterosigositas dan χ2 test gen LCAT posisi basa c.742 domba lokal Indonesia Sub Populasi Jumlah
Individu
Frekuensi Genotipe Frekuensi Gen Equilibrium χ2 test Ho He CC CT TT C T ETS 43 0.977 0.023 0.000 0.988 0.012 33.185** 0.023 0.111 ETJ 17 0.765 0.235 0.000 0.882 0.118 0.221ns 0.235 0.214 EGJ 6 1.000 0.000 0.000 1.000 0.000 0.000 0.000 0.000 Garut 19 0.895 0.000 0.105 0.895 0.105 24.727** 0.000 0.194 Lembah Palu 9 0.889 0.000 0.111 0.889 0.111 17.067** 0.000 0.209 Pulau Rote 7 1.000 0.000 0.000 1.000 0.000 0.000 0.000 0.000 Kissar 7 1.000 0.000 0.000 1.000 0.000 0.000 0.000 0.000 Sumbawa 10 1.000 0.000 0.000 1.000 0.000 0.000 0.000 0.000 Nilai Rataan 0.941 0.032 0.027 0.957 0.043 0.032 0.091
Keterangan: ETS = ekor tipis sumatera; ETJ = ekor tipis jawa; EGJ = ekor gemuk jawa; ** = berbeda sangat nyata P<0.01 pada taraf α
= 0.01; ns = tidak berbeda nyata P>0.05 pada taraf α = 0.05
Tabel 2.3 Frekuensi genotipe, frekuensi gen, nilai heterosigositas dan χ2 test gen LCAT posisi basa c.770 domba lokal Indonesia Sub Populasi Jumlah
Individu
Frekuensi Genotipe Frekuensi Gen Equilibrium χ2 test Ho He AA AT TT A T ETS 43 0.000 0.116 0.884 0.058 0.942 0.1296ns 0.1160 0.1110 ETJ 17 0.000 0.471 0.529 0.235 0.765 1.3785ns 0.4710 0.3710 EGJ 6 0.000 0.167 0.833 0.083 0.917 0.0000 0.1667 0.1667 Garut 19 0.000 0.316 0.684 0.158 0.842 0.5444ns 0.3158 0.2731 Lembah Palu 9 0.000 0.333 0.667 0.167 0.833 0.2286ns 0.3333 0.2941 Pulau Rote 7 0.000 0.143 0.857 0.071 0.929 0.8000ns 0.5714 0.4396 Kissar 7 0.000 0.000 1.000 0.000 1.000 0.0000 0.0000 0.0000 Sumbawa 10 0.000 0.000 1.000 0.000 1.000 0.0000 0.0000 0.0000 Nilai Rataan 0.000 0.193 0.807 0.097 0.903 0.2345 0.1990
Keterangan: ETS = ekor tipis sumatera; ETJ = ekor tipis jawa; EGJ = ekor gemuk jawa; ns = tidak berbeda nyata P>0.05 pada taraf α = 0.05
Tabel 2.4 Frekuensi genotipe, frekuensi gen, nilai heterosigositas dan χ2 test gen LCAT posisi basa c.882 domba lokal Indonesia Sub
Population n
Frekuensi Genotipe Frekuensi Gen Equilibrium χ2 test Ho He CC CT TT C T ETS 43 0.930 0.070 0.000 0.970 0.030 24.6711** 0,0698 0.2596 ETJ 17 0.824 0.176 0.000 0.912 0.088 6.8174** 0,1770 0.4510 EGJ 6 1.000 0.000 0.000 1.000 0.000 0.0000 0.0000 0.0000 Garut 19 0.842 0.105 0.053 0.895 0.105 5.1390** 0.1053 0.1935 Lembah Palu 9 1.000 0.000 0.000 1.000 0.000 0.0000 0.0000 0.0000 Pulau Rote 7 0.571 0.429 0.000 0.786 0.214 0.3273ns 0.4286 0.3626 Kissar 7 1.000 0.000 0.000 1.000 0.000 0.0000 0.0000 0.0000 Sumbawa 10 1.000 0.000 0.000 1.000 0.000 0.0000 0.0000 0.0000 Nilai Rataan 118 0.896 0.098 0.007 0,949 0.055 0.0931 0.0948
Keterangan: ETS = ekor tipis sumatera; ETJ = ekor tipis jawa; EGJ = ekor gemuk jawa; ** = berbeda sangat nyata P<0.01 pada taraf
α = 0.01; ns = tidak berbeda nyata P>0.05 pada taraf α = 0.05
1
Kecenderungan peternak untuk menjual jantan dewasa yang memiliki performan yang baik sebagai ternak potong, populasi yang tertutup, perkawinan tidak secara acak (selective mating) dan ukuran efektif populasi tidak seimbang diduga sebagai faktor yang menyebabkan rendahnya heterosigositas dalam suatu populasi. Menurut Sumantri et al. (2008b), nilai heterosigositas dipengaruhi oleh jumlah sampel, jumlah dan frekuensi alel serta marka genetik yang digunakan. Tingginya keragaman gen-gen yang dimiliki oleh domba lokal Indonesia telah dilaporkan diantaranya gen calpastatin (Sumantri et al. 2008b; Dagong et al. 2011) dan tidak adanya keragaman gen terdeteksi pada gen myostatin (Sumantri et al. 2011).
Simpulan
Hasil penelitian menemukan 3 SNPs baru gen LCAT ekson 6 pada domba lokal Indonesia yaitu pada posisi basa c.742 C>T; c.770 T>A dan c.882 C>T. Kombinasi 3 SNPs membentuk sembilan diplotipe. Substitusi sitosina menjadi timina c.742 pada posisi kodon ketiga (GCC>GCT) merupakan synonymous mutation (Ala>Ala); subsitusi timina menjadi adenina c.770 pada posisi kodon pertama (TTC>ATC) dan sitosina menjadi timina c.882 (GCA>GTA) pada posisi kodon kedua merupakan non-synonymous mutation mengakibatkan perubahan asam amino fenilalanina>isoleusina dan alanina>valina. Ketiga SNPs yang ditemukan merupakan SNPs baru yang belum dilaporkan pada populasi domba lainnya. Keragaman gen LCAT ekson 6 ditemukan pada rumpun domba ekor tipis sumatera, domba ekor tipis jawa, domba ekor gemuk jawa, domba garut, domba lembah palu dan domba pulau rote dan tidak ditemukan pada rumpun domba kissar dan domba sumbawa.
20
3 IDENTIFIKASI KERAGAMAN GEN LIPOPROTEIN LIPASE
PADA DOMBA EKOR TIPIS SUMATERA DAN
DOMBA GARUT
ABSTRAKLiporotein lipase merupakan enzim kunci yang berperan dalam metabolisme dan transpor lipoprotein sehingga dapat meningkatkan level trigliserida darah. Lipoprotein lipase juga berfungsi mengontrol partisi trigliserida pada jaringan adipose dan otot sehingga dapat meningkatkan penyimpanan lemak dan menyediakan energi dalam bentuk asam lemak untuk pertumbuhan otot. Lipoprotein lipase (LPL) dikodekan oleh gen LPL. Penelitian ini bertujuan untuk mengeksplorasi single nucleotide polymorphisms (SNPs) gen LPL. Total 66 DNA genom digunakan dalam penelitian ini, terdiri dari domba sumatera ekor tipis (50 ekor) dan domba garut (16 ekor). Amplifikasi DNA genom menggunakan metode polymerase chain reaction dan metode direct sequencing digunakan untuk mengidentifikasi keragaman sekuens. Selanjutnya sekuens disejajarkan dengan metode Clustal W dan pensejajaran sekuens dengan gen bank nomor akses X.68308.1 menggunakan program BioEdit dan program MEGA 5.2. Hasil penelitian menunjukkan bahwa ditemukan tiga SNPs baru yaitu pada posisi basa g.26, g.27 dan c.192. Insersi sitosina atau guanina pada basa ke 26 dan insersi guanina pada basa ke 27 merupakan frameshift mutation. Substitusi timina menjadi sitosina pada c.192 adalah non synonymous mutation (Val>Ala), ditemukan pada domba garut.
Kata kunci: direct sequencing, domba, gen LPL, SNPs ABSTRACT
Lipoprotein lipase is a key enzyme that plays in metabolism and transport lipoprotein as well as influence on blood triglyceride levels. Lipoprotein lipase controls triacyl glycerol partitioning between adipose tissue and muscle that increases fat storage or provides energi in the form of fatty acids for muscle growth. Lipoprotein lipase (LPL) is encoded by LPL gene. The research was aimed to explore single nucleotide polymorphisms of LPL gene. A total of 66 genomic DNA of Indonesian local sheeps, consisted of sumatera thin tail (50 heads) and garut (16 heads) sheeps were used in this study. Polymerase chain reaction was used to amplify genomic DNA and direct sequencing method was used to identify polymorphism sequences. The sequences were aligned with Clustal W method and BLAST sequence obtained from Gene Bank with accession number X.68308.1. The results showed three novel single nucleotide polymorphisms at g.26, g.27 and c.192. Insertion cytosine or guanine g.26 and insertion guanine g.27 were frame shift mutation. Substitution thymine to cytosine c.192 was a non synonymous mutation (Val>Ala), it was found in garut sheep. Keywords: direct sequencing, LPL gene, sheep, SNPs,