Model Hidrolik yang Tepat untuk Perubahan Pemanjangan Daun pada Kondisi Fluktuasi Penguapan dan Status Air Tanah
Fahroziah Assyifa, Ganjar Abdillah Ammar, Isna Muflihah Mulyaningrum, Maria Benita Pratiwi, Rahma Hanifah, Sutra Eci Hutari
Sekolah Ilmu dan Teknologi Hayati Institut Teknologi Bandung
Abstrak
Tanaman mengalami perubahan kebutuhan evaporatif dan kandungan air tanah yang lebih lanjutnya akan mengubah status air dan pertumbuhan. Sebagai kontradiksi dari hipotesis hidraulik, laju elongasi daun (LER) mengalami penurunan di pagi hari dan dapat kembali ke nilai semula jika rehidrasi tanah dilakukan. Laju elongasi daun mengalami peningkatan yang lebih cepat dibandingkan dengan laju transpirasi dan potensial air pada daun (waktu paruh tipikal selama 30 menit dibandingkan 1-2 jam). Penurunan LER di pagi hari dimulai ketika cahaya redup dan transpirasi yang rendah diikuti dengan pembukaan stomata pada daun yang teriluminasi cahaya matahari langsung, yang merepresentasikan fraksi kecil dari luas penampang daun. Model simulasi pada jagung (Zea mays) menyiratkan bahwa penemuan ini bersifat kompatibel dengan hipotesis hidrolik. Fluks air yang kecil menuju lubang stomata bernilai cukup untuk menurunkan potensial air pada xilem dan jaringan yang tumbuh, sehingga menyebabkan penurunan LER secara cepat dibandingkan dengan penurunan air pada jaringan dewasa akibat kapitansi hidrolik. Model ini juga menggambarkan pola pertumbuhan di siang hari atau ketika rehidrasi tanah dilakukan. Perubahan konduktansi hidrolik pada tanaman sebagian berlawanan dengan transpirasi. Secara keseluruhan, mekanisme dan model yang direpresentasikan merupakan komponen penting bagi toleransi terhadap kekeringan pada tuntutan evaporatif dan kelembapan tanah yang berubah secara fluktuatif.
Kata kunci: Evaporatif; Elongasi; Hidrolik; Fluktuatif; Simulasi
Abstract
Plants are constantly facing rapid changes in evaporative demand and soil water content, which affect their water status and growth. In apparent contradiction to a hydraulic hypothesis, leaf elongation rate (LER) declined in the morning and recovered upon soil rehydration considerably quicker than transpiration rate and leaf water potential (typical half-times of 30 min versus 1-2 h). The morning decline of LER began at very low light and transpiration and closely followed the stomatal opening of leaves receiving direct light, which represent a small fraction of leaf area. A simulation model in maize (Zea mays) suggest that these findings are still compatible with a hydraulic hypothesis. The small water flux linked to stomatal aperture would be sufficient to decrease water potentials of xylem and growing tissues, thereby causing a rapid decline of stimulated LER, while the simulated water potential of mature tissues decline more slowly dua to a high hydraulic capitance. The model also captured growth patterns in the evening or upon soil rehydration. Changes in plant hydraulic conductance partly counteracted those of traspiration. Overall, the mechanisms and model presented here may be an essential component of drought tolerance in naturally fluctuating evaporative demand and soil moisture.
1. PENDAHULUAN
Leaf Elongation Rate (LER)
merupakan salah satu bentuk respon tumbuhan yang dipengaruhi oleh air dan penguapan. Hal ini dibuktikan dari tumbuhan-tumbuhan monokotil (Hsiao et al., 1970; Ben Haj Salah dan Tardieu, 1997; Munns et al., 2000). Penguapan yang terjadi pada kondisi normal bergantung pada perubahan cahaya, suhu, dan embun. Penguapan pada kondisi normal akan mempengaruhi kandungan air dalam tanah. Penguapan pada tanah serta penyerapan air oleh tumbuhan dan air hujan akan mempengaruhi pergerakan air. Pergerakan air pada tumbuhan akan memiliki dampak dalam pertumbuhan daun.
Dalam beberapa penelitian yang telah dilakukan pada tumbuhan monokotil, fluktuasi Leaf Elongation Rate (LER) yang cepat disebabkan oleh proses hidrolik yang terjadi pada tumbuhan dan variasi konduktivitas dari aktivitas hidrolik tersebut (ehlert et al., 2009). Selain itu, fluktuasi juga terjadi karena perubahan gradien potensial antara xylem dan jaringan meristem. Faktor lain yang mempengaruhi fluktuasi LER adalah struktur dinding sel pada zona tumbuh yang berubah karena kekurangan air. Kekurangan air ini juga disebabkan oleh Asam Absisat (ABA) yang dikombinasikan dengan hormon lain.
Hal yang dianalisa dan dipelajari pada jurnal yang dipilih adalah analisa variasi waktu Leaf Elongation Rate dari tumbuhan Zea mays. Analisa ini dilakukan pada transisi waktu antara subuh dan pagi, antara siang dan malam, dan saat proses rehidrasi dari tanaman
yang kering. Perlakuan-perlaukan yang ditentukan ini mendasar pada literatur (Parent et al., 2009; Tardieu et al., 2010; Welcker et al., 2011) yang berfokus pada potensial air dan konduktivitas hidrolik. Literatur yang menjadi patokan sudah diuji menggunakan model.
2. MATERIAL DAN METODE
2.1 Kondisi Pertumbuhan Tanaman pada Rumah Kaca
Tanaman ditumbuhkan pada kolom polivinil klorida (diameter 0.23 m dan tinggi 0.40 m) yang mengandung campuran 40:60 (v/v) tanah liat yang terfiltrasi (diameter partikel 0.1-4 mm) dan kompos organik. Suhu udara dan kelembaban relatif diukur pada tanaman setiap 30 detik menggunakan sembilan buah sensor (HMP35A; Vaisala Oy). Temperatur dari zona meristematik diukur dengan termokopel tembaga yang konstan (diameter 0.2 mm) yang disisipkan di antara pelepah daun untuk 2 dan 3 dari 6 tanaman setiap eksperimennya. PPFD diukur setiap 30 detik dengan menggunakan 9 buah sensor (LI-190SB dari Li-Cor dan SOLEMS 01/02/012). Perbedaaan tekanan uap air dari meristem ke udara ditentukan setiap saat sebagai perbedaan tekanan uap air di antara keadaan tersaturasi pada suhu meristem dan tekanan uap air di udara. pada temperatur meristem dan aliran tekanan uap pada udara. Seluruh data yang berhubungan terhadap iklim dirata-ratakan dan disimpan setiap 15 menit pada penyimpan data (Campbel Scientific LTD-CR10X Wiring Panel).
otomatis setiap 15 menit. Perbedaan berat dihubungkan terhadap perubahan kandungan air pada tanah, setelah dilakukan koreksi untuk meningkatkan rata-rata biomassa tanaman sebagai fungsi dari fase fenologis dan efek perpindahan transduser. Kurva pelepasan air pada tanah diperoleh dengan mengukur potensial air pada tanah dari sampel tanah dengan perbedaan kandungan air pada jangkauan nilai 0.4 – 0.2 g g-1 (WP4-T Dewpoint Meters; Decagon Devices), sehingga potensial air rata-rata pada tiap kolom dapat diukur tiap 15 menit sekali. Pada eksperimen yang dilakukan dalam kondisi ketersediaan air yang baik, substrat tanah yang dijaga pada kapasitas retensi (penyimpanan) dengan pengairan yang baik oleh larutan Hoagland termodifikasi. Larutan yang sama digunakan dalam eksperimen dalam keadaan kekurangan air (-0.15 dan -0.6 MPa). Seluruh data terkait iklim dirata-ratakan dan disimpan setiap 15 menit melalui penyimpan data. Tambahan cahaya digunakan selama musim dingin untuk menjaga proses fotoperiodik selama 12 jam terang dan 12 jam gelap dan densitas fluks foton fotosintesis lebih dari 400 umol m-2 s-1.
2.2 Kondisi Pertumbuhan Tanaman dalam Kondisi Hidroponik
Biji jagung dikecambahkan di cawan petri pada 20°C dalam gelap dan udara jenuh. Setelah 4 hari, bibit ditempatkan dalam tabung dengan serat mineral dan ditransfer ke rumah kaca dengan akar yang terus diaerasi pada sebuah larutan. Komposisi larutan adalah sebagai berikut: 0,25 mM CaSO4, 0,8 0,04 mM Fe-EDDHA, dan 2,5 mM MES, pH 5,5 - 6. Larutan hidroponik diperbaharui setiap hari ketiga atau keempat, dan pH dikontrol setiap hari. Perlakuan defisit air yang diterapkan dengan mengganti larutan nutrisi standar oleh nutrisi larutan yang mengandung manitol (Sigma-Aldrich), sesuai dengan potensial air 20,15 MPa, diperiksa dengan osmometer tekanan uap (Vapro 5520; Wescor). Perlakuan manitol diaplikasikan pada hari terakhir pengukuran. Data lingkungan diukur dan disimpan seperti di atas.
2.3 Pengukuran Tanaman
LER diukur dengan perubahan transduser secara rotatik (601-1045 Full 360o Smart Position Sensor; Spectrol Electronics). Katrol telah diletakkan pada sensor yang membawa benang tembaga ke ujung daun dan untuk mengimbangi berat 20-g. LER dirata-ratakan dan disimpan setiap 3 menit pada kebanyakan eksperimen dan 5-15 menit pada beberapa eksperimen setelah koreksi dari efek temperatur pada panjang benang. Pada tiap eksperimen, pengukuran dimulai ketika ujung dari keenam daun muncul diatas gulungan dan bertahan hingga muncul daun ke-8. Periode ini dihentikan ketika LER bernilai stabil. LER diekpresikan pada waktu termal, melalui hari ekuivalen pada suhu 20oC sebagai efek langsung dari fluktuasi temperatur.
mana lapisan atas disiram air sehingga kandungan air bernilai sama pada kolom yang diuji. Laju transpirasi dirata-ratakan dan disimpan setiap 15 menit. Luas daun dikalkulasikan dari panjang dan lebar setiap daun dari 60 tanaman setiap hari kedua. Model logistik dicocokkan pada bagian waktu dari luas setiap daun. Luas daun diperoleh setiap hari dari penentuan luas dari seluruh daun pada tanaman.
Bagian waktu dari variabel iklim, pemanjangan daun, dan laju transpirasi diperoleh dari 23 eksperiman yang diklasifikasikan ke tipe skenario terbatas. Kluster dari waktu diperoleh dalam tiga langkah menggunakan bahasa R. Data mentah diklasifikasikan dengan transisi siang/malam dan fotoperiodik setiap eksperimen: Transisi hari yang cepat pada siang dan malam terjadi ketika musim semi atau gugur karena tambahan cahaya diberikan, di mana pada musim semi atau panas memiliki transisi lambat karena kehadiran tambahan cahaya; (2) Tersedianya air tanah yang berbeda antara eksperimen telah diklasifikasikan menjadi pemberian air yang baik (-0.1 – 0 MPa), defisit air ringan (-0.2 - -0.1 MPa) dan defisit air (-0.6 - -0.5 MPa); (3) permintaan evaporatif esensial yang bergantung pada PPFD dan VPD yang diklasifikasikan pada kondisi cerah (PPFD > 1000 umol m-2 s-1 dan VPD > dirata-ratakan dan dipertimbangkan sebagai satu kali perulangan pada analisis lebih lanjut untuk memperoleh nilai rata-rata dan interval kepercayaan yang baik. Waktu paruh dari variasi LER, potensial
air daun, dan laju transpirasi selama perolehan kembali dikalkulasi setelah interpolasi menggunakan fungsi R spline. Nilai maksimum dan minimum dimasukkan pada fungsi spline. Waktu paruh dikalkulasi dengan waktu sebagai nilai fungsi dari rata-rata nilai minimum dan.Potensial air diukur pada daun tanpa perluasan dengan ruang tekanan (Soil Moisture Equipment). Potensial air dari daun yang ditutupi diukur setelah penutupan dilakukan dengan plastik transparan untuk menghindari penguapan dan penutupan dengan pembungkus aluminium untuk menghindari kenaikan suhu. Daun disiapkan selama malam dan siang untuk pengukuran. Konduktansi stomata diukur setiap menit menggunakan analisator gas (CIRAS-2; PP Systems). Dua analisator gas digunakan secara simultan, 1 diletakkan pada bagian daun yang terkena sinar matahari dan satu yang lain berlawanan sisi dari penerimaan cahaya difusi. Konduktansi stomata juga diukur dalam eksperimen ke-5 (hidroponik) pada daun untuk menerima sinar matahari secara langsung menggunakan odometer difusi (AP4; Delta-T Devices) yang dikalibrasi setiap 30 menit.
Seluruh konduktansi hidrolik tanaman dikalkulasi dari nilai kehilangan air setiap tanaman, dibagi dengan perbedaan potensial air antara tanah dan daun. Potensial transpirasi dan air tanah dikalkulasi setiap 15 menit. Nilai rata-rata dari potensial air di daun diperoleh dari pengukuran pada 5 menit terakhir berdasarkan langkah setiap waktu 15 menit. Konduktivitas hidrolik akar diukur pada tanaman secara hidroponik di dalam
diisi serat kapas dan diukur beratnya. Potensial osmosis dari getah dan larutan bernutrisi diukur dengan tekanan uap osmometer (Vapro 5520; Wescor). Konduktivitas hidrolik akar pada gradien osmotik (Lp) dikalkulasi.
Lpos=J∗
(
πsap−πsol)
−1∗A−1…(1)
J: fluks air melalui akar (mg.m-2.s-1)
πsap danπsol : Potensial osmotik dari sampel getah dan larutan bernutrisi A: Area dari sistem akar (m2)
2.4 Eksperimen Pembasahan Berulang pada Kondisi Terkontrol
Tanaman dengan garis keturunan sel B73 ditumbuhkan di dalam rumah kaca dengan defisit potensial air sebesar -0,4 Mpa. Ketika tujuh buah daun muncul, tanaman dipindahkan pada malam hari ke ruang pertumbuhan dengan penguapan yang tinggi (VPD = 2,8 kPa, 28C, PPFD = 600 µmolm2s1. Tanaman dibiarkan bertranspirasi pada kondisi tersebut selama 6 jam. Potensial air pada keenam daun diukur menggunakan ruang bertekanan dan LER, kemudian dimonitor setiap 5 menit menggunakan transduser perpindahan rotasional. Pada saat t=0, tanaman disiram kembali untuk meretensi kapasitas dan ditempatkan dalam kondisi gelap pada VPD senilai 0,8 kPa agar proses transpirasi berhenti. LER dan potensial air pada daun diukur setiap 3 menit, diikuti dengan penyiraman kembali setelah 5 jam kemudian. Eksperimen ini dilakukan secara triplo.
2.5 Model Transfer Air
Menurut (Tardieu dan Davies 1993), model ini berjalan pada setiap satu menit.Air mengalir di antara kompartemen ψsoil hingga ψr, ψxyl, dan
ψbundle. ψsoil mewakili potensial air tanah sekitar akar pada malam hari, hal ini setara dengan potensial air saat dini hari. ψr merupakan potensial air di permukaan akar lebih luar. Resistensi Rsp (MPas tanaman mg-1) antara kedua kompartemen dihitung menurut (Gardner, 1960), dengan d dan r adalah jarak antara akar-akar yang bersebelahan dan radius akar, serta k( ) adalahϴ konduktivitas hidrolik tanah sebagai fungsi dari konten air tanah.
ln(d 2
r2)
4ᴨk(θ)…(2)
percobaan rehidrasi. Fluks Jxc (mg plant -1s-1) antarkompartemen dan ψbundle
dihitung mengikuti gradient potensial air antara kedua kompartemen dengan resistensi Rc, yang bergantung pada PPFD dengan cara yang sama seperti pada Rp. Berikut persamaan diferensial untuk menghitung potensial air pada sel CCEL dan Jxc.
Dengan Vsat (mm3) adalah volume daun pada keadaan jenuh dan VRes adalah volume air residu -0,3MPa, dan α dan n
adalah parameter persamaan Van Genuchten yang dimasukkan pada kurva tekanan volume. Jxc dihitung sebagai perbedaan dalam Vcell antardua waktu yang berbeda untuk proses optimalisasi pada solusi persamaan diferensial. Pada setiap waktu i, fluks yang melalui akar dan xylem adalah jumlah dari transpirasi fluks J dan Jxc.
Untuk menghitung konduktansi stomata, konsentrasi ABA dalam getah xylem ([ABA]xyl), fluks air, akar, dan potensial air di sisi transpirasidari potensial air tanah, intensitas cahaya, dan VPD udara. Konduktansi stomata dihitung sebagai berikut.
β
[
ABA]
xylexp(δΨbundle)¿ ¿
g s=gsmin+αexp¿
Dengan gsmin adalah konduktansi stomata minimum yang dihasilkan dari konduktansi kutikula dan konduktansi melalui stomata tertutup. α, β, dan δ adalah parameter yang diusulkan. Fluks transpirasi J dihitung dengan persamaan
Penman Monteith untuk
mengekspresikan laju transpirasi per tanaman. [ABA]xyl dihitung sebagai berikut (Tardieu dan Davies, 1993).
[
ABA]
xyl=−aΨr/(J+b) ....(6) LER sebagai laju maksimum yang dipengaruhi oleh ln[ABA]xyl (Tardieu et al., 2010) dan ψxyl secara linear(parameter a_lerdanc_ler).aba
LER=aler+min(0,−a¿ln[ABA]xyl
(
1+c¿ler× Ψxyl)
) ...(7)Tumbuhan jagung bergaris keturunan sel B73 ditumbuhkan pada greenhouse dengan defisiensi air (Potensial air tanah -0.3 MPa). Pada tahap seven-visible-leaf, tumbuhan dipindahkan ke growth chamber pada siang hari dengan nilai VPD=2.8 kPa, 28oC, PPFD= 600 molm-2s-1.Tumbuhan tersebut didiamkan selama 6 jam selama potensial air daun dihitung dengan
pressure chamber dan nilai LER pada daun keenam dipantau setiap 5 menit dengan rotational displacement
transducer. Saat t=0, tumbuhan diberikan
air dan disimpan dalam tempat gelap pada nilai VPD 0.8 kPa. Nilai VPD tersebut akan menghentikan transpirasi. LER dan potensial air daun dihitung setiap 3 menit untuk 5 jam rehidrasi. Eksperimen ini diulang tiga kali.
3. HASIL
3.1 Penurunan LER tidak cocok untuk transpirasi tanaman di pagi hari
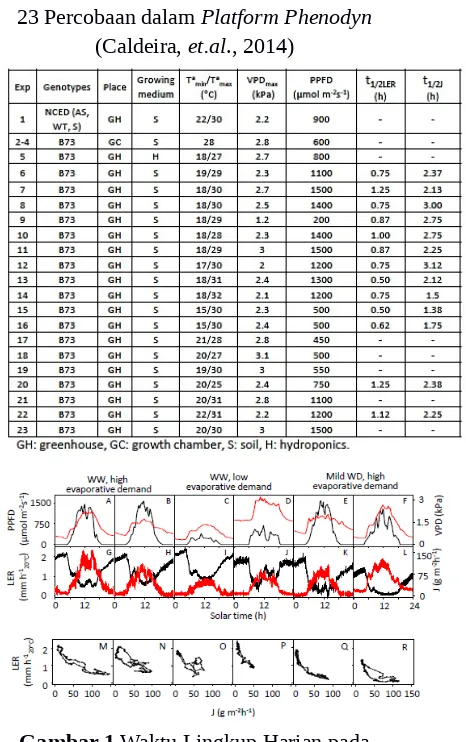
terbitnya matahari sekitar pukul 4.30 hingga 07.00 pagi. Transpirasi yang berlawanan dengan LER menghasilkan depresi harian dan laju transpirasi yang lebih tinggi dengan defisit air yang sangat rendah seperti yang ditunjukkan pada Gambar 1, Gambar 2 dan Gambar 3. Namun, penurunan LER pada transisi antara siang dan malam jauh lebih cepat dari laju transpirasi, dengan rata-rata setengah kali dari 0,8 ± 0,3 dan 2.3 ± 0,5 jam pada pagi hari dari 14 percobaan, seperti yang ditunjukkan pada Tabel 1 dan Gambar 3. Hal ini berlangsung pada waktu dimana laju transpirasi dapat diabaikan (5 ± 3 g m-2 h -1) karena rendahnya nilai densitas flux
foton pada fotosintesis (PPFD) dan defisit tekanan uap air (YPD; 10 ± 3μmol m-2 s-1dan 0,75 m-2 h-1 ± 0,2 kPa). Menariknya, hal tersebut juga terjadi pada hari dengan tingkat penguapan yang rendah, ketika LER menurun secara drastis pada peningkatan laju transpirasi yang sangat lambat seperti yang ditunjukkan pada Gambar 1, I-J. Hubungan transpirasi dan LER adalah nonlinier pada pagi dan sore hari seperti yang ditunjukkan pada Gambar 1 dan Gambar 2. Oleh karena itu, perubahan pada LER dianggap sebagai laju transpirasi independen atau sangat sensitif terhadapnya.
Tabel 1. Kondisi Lingkungan dan Garis pada
23 Percobaan dalam Platform Phenodyn
(Caldeira, et.al., 2014)
Gambar 1 Waktu Lingkup Harian pada Kondisi Lingkungan, Transpirasi (J) dan Laju Perpanjangan Daun (LER) dan Hubungan LER dan E dalam 6 Kelompok
Percobaan.
(Sumber : Caldeira, et al., 2014)
Gambar 2 Waktu Lingkup Harian pada Kondisi Lingkungan, Transpirasi (J) dan Laju Perpanjangan Daun (LER) dan Hubungan LER dan J dalam 5 Kelompok
Percobaan
Gambar 3 Waktu Paruh Reduksi pada Laju Perpanjangan Daun dan Peningkatan Transpirasi Selama Pagi Hari dalam 13
Percobaan
(Sumber : Caldeira, et al., 2014)
3.2 Perubahan Potensial Daun pada Seluruh bagian Tanaman Lebih Rendah Dibanding pada LER selama Bertambahnya Kebutuhan Evaporasi atau Rehidrasi Tanah
Potensial air pada daun berkurang selama 4 jam dipagi hari pada tanaman yang tumbuh ditanah atau larutan hidroponik (potensial air atau tanah -0,15 MPa; seperti yang ditunjukkan pada Gambar 4 dan 5), lebih rendah dibandingkan penurunan LER. Potensial air pada tanaman yang ditutup dengan plastik dan alumnium foil dengan perlakuan yang sama lebih rendah dibandingan dengan tanaman yang tidak ditutup pada pagi hari namun lebih tinggi siang hari seperti yang ditunjukkan pada Gambar 4B.
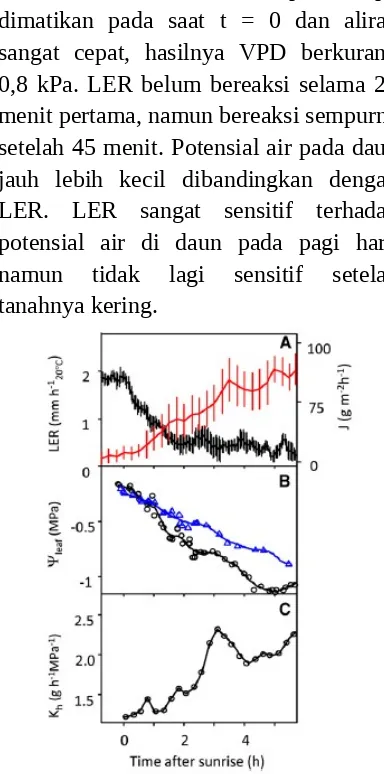
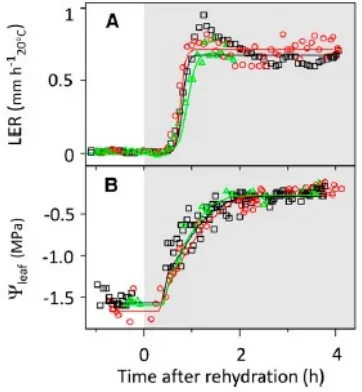
Potensial air dan LER selama penyiraman dianalisis dengan tiga kali percobaan dengan waktu resolusi 5 menit seperti yang ditunjukkan pada Gambar 6. Tanaman yang pertama tumbuh selama 8 jam pada potensial air tanah sebesar -0,6 ± 0,1 Mpa dengan penguapan yang tinggi ( PPFD = 800 μmol m-2 s-1 dan VPD = kPa), hasilnya potensial air di
daun adalah sebesar -1.5 Mpa. Lampu dimatikan pada saat t = 0 dan aliran sangat cepat, hasilnya VPD berkurang 0,8 kPa. LER belum bereaksi selama 25 menit pertama, namun bereaksi sempurna setelah 45 menit. Potensial air pada daun jauh lebih kecil dibandingkan dengan LER. LER sangat sensitif terhadap potensial air di daun pada pagi hari, namun tidak lagi sensitif setelah tanahnya kering.
Gambar 4 Waktu Lingkup LER dan Variabel yang Berhubungan dengan Pertumbuhan
Tanaman dalam tanah (Percobaan 6)
Gambar 5 Waktu Lingkup LER dan Variabel yang Berhubungan dengan Pertumbuhan Tanaman dalam Hidroponik (Percobaan 5)
(Sumber : Caldeira, et al., 2014)
Gambar 6 Waktu Lingkup LER dan
Potensial Air di Daun selama Rehidrasi Tanah dalam 3 Percobaan (Percobaan 2-4)
(Sumber : Caldeira, et al., 2014)
3.3 Peningkatan Konduktansi Hidrolik Tanaman pada Awal Pagi yang
Membatasi Penurunan LER, secara Spesial dalam Kondisi Pemberian Air yang Baik
Konduktansi hidrolik tanaman mengalami peningkatan sepanjang waktu pagi, sesuai dengan penelitian
sebelumnya (Lopez et al., 2003; Sakurai-Ishikawa et al., 2011; Hachez et al., 2012). Pembatasan penurunan LER pada pembukaan stomata konsisten dengan model sehingga tidak dapat diprediksi dengan data eksperimen LER tanpa adanya peningkatan konduktansi tanaman yang cukup besar. Semakin cepat penurunan LER pada tanaman dalam memproduksi ABA menunjukkan konsistensi dengan kontribusi utama dari konduktivitas hidrolik (Parent et al., 2009) dan pembukaan stomata lebih cepat di pagi hari dengan kecenderungan yang berlawanan dalam memproduksi ABA. Dengan cara yang sama, semakin cepat penurunan LER dalam defisit air yang berhubungan dengan rendahnya kontribusi dari konduktivitas hidrolik dari keseluruhan konduktansi total dari tanah ke daun seperti yang diprediksikan model. Pada tanah basah (-0.01 MPa), konduktansi hidrolik tanaman lebih rendah dibandingkan konduktansi hidrolik tanah sehingga berpengaruh dalam pembatasan perpindahan air.
Perubahan jumlah transkrip PIP
selama pagi hari tersinkronkan dengan konduktansi hidrolik tanaman. Perubahan harian dari ekspresi gen dan atau protein pada sebagian besar PIP tidak berpengaruh pada pemanjangan zona daun (Lopez et al., 2003; Hachez et al., 2008, 2012). Fosforilasi PIP menghasilkan peningkatan aktivitas saluran air seperti yang terjadi di pagi hari sebagaimana ubiquitous dalam kontrol aktivitas PIP (Maurel et al., 2008; Van Wilder et al., 2008). Akuaporin pada saluran mempengaruhi konduktivitas hidrolik pada ketergantungan cahaya termasuk fosforilasi dalam Arabidopsis
thaliana (Prado et al., 2013).
mempengaruhi konduktivitas hidrolik tanaman dalam rehidrasi tanah dan cenderung untuk menurun daripada meningkat selama kekurangan air (Vandeleur et al., 2009)
3.3 Penurunan LER di Pagi Hari Berhubungan dengan Konduktansi Stomata pada Daun yang Teriluminasi
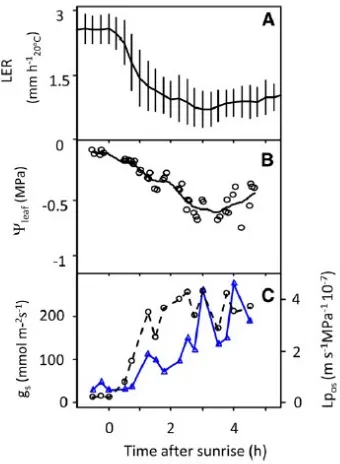
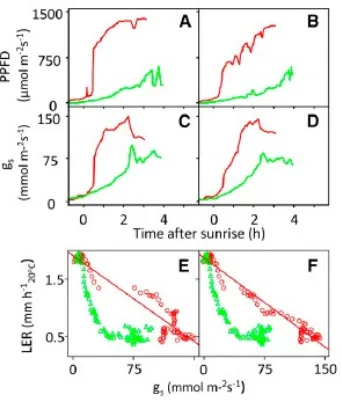
Konduktansi stomata dihitung pada daerah daun baik yang menerima cahaya matahari secara langsung maupun cahaya difusi seperti yang ditunjukkan pada Gambar 9. Konduktansi stomata meningkat secara cepat pada pagi hari, lebih cepat dari laju transpirasi, semakin rendah karena kebutuhan evaporatif yang rendah pada waktu tersebut seperti yang ditunjukkan pada Gambar 8. Stomata terbuka cepat hanya pada daun yang memiliki orientasi untuk menerima cahaya langsung setelah matahari terbit, sedangkan daun hanya menerima difusi cahaya dari stomata yang terbuka secara lambat, sebanding dengan intensitas cahaya yang diterima dari kategori daun seperti yang ditunjukkan pada Gambar 7.
Peningkatan yang sedikit pada laju transpirasi disebabkan oleh pembukaan stomata pada daerah daun di bawah evaporatif yang rendah, namun menyebabkan penurunan LER. Hubungan antara LER dan konduktansi stomata pada daun yang teriluminasi berbanding lurus. Hubungan antara LER dan konduktansi stomata, sangat berbeda dengan perubahan potensial air pada daun dan laju transpirasi. Kesimpulan ini tetap terjadi pada eksperimen dengan evaporatif rendah seperti yang ditunjukkan pada Gambar 7.
Gambar 7 Lingkup Waktu Harian untuk Kondisi Lingkungan, Transpirasi (J) dan
LER dari 6 Kelompok Eksperimen. (Sumber : Caldeira, et al., 2014)
Gambar 8. Korelasi antara LER terhadap Variabel yang Berkaitan dengan Pertumbuhan Tanaman pada Media Hidroponik. A = LER, B = potensial air pada
daun yang terkena cahaya, C = konduktansi stomata.
Gambar 9 Lingkup Waktu PPFD (A dan B) dan Konduktansi Stomata (C dan D) pada
Daun yang Menerima Cahaya Matahari secara Langsung (merah) atau Difusi (hijau)
selama Pagi Hari
(Sumber : Caldeira, et al., 2014)
3.4 Pensimulasian Model Pemisahan Faktor Potensial Air pada Xilem dengan Potensial Air Total Daun
Simulasi klasik pada pemodelan ekspansi daun menggambarkan hubungan LER terhadap faktor transpirasi dan potensial air pada daun. Namun, asumsi ini tidak sesuai dengan pengamatan perubahan fenotipik pada daun Zea mays. Model lainnya menyatakan bahwa nilai LER tidak berkaitan dengan faktor konduktivitas hidrolik, tetapi melibatkan faktor sifat dari dinding sel tumbuhan dan ritme sirkadian. Jika faktor konduktivitas hidrolik dipertimbangkan, maka korelasi antara nilai LER dengan potensial air pada xilem dapat dipertimbangkan. Hal ini mengingat bahwa perubahan konduktansi stomata juga dipengaruhi oleh suplai air ke akar tanaman, di mana potensial air total pada daun bereaksi
lebih lambat karena volume air memberikan nilai kapitansi yang besar.
Fluks air yang melewati suatu kompartmen dari tumbuhan akan berjalan sesuai dengan perbedaan nilai potensial air, baik dari tanah (ψsoil) menuju akar (ψr), xilem (ψxyl), dan
bundle sheath (ψbundle). Fluks air yang mengalir menuju bundle sheath dapat ditranspirasikan atau ditransfer ke sel daun dewasa, bergantung pada perbedaan nilai potensial air di masing-masing tempat. Rp adalah hambatan jaringan akar, xylem tumbuhan dan protoxilem daun. Diasumsikan berubah terhadap waktu, sesuai dengan Gambar 8 dan 10. Rxl adalah hambatan antara xylem dan
bundle sheaths, Rc adalah hambatan dari
bundle sheaths ke sel daun yang matang
dan berubah terhadap waktu. Konduktansi stomata bergantung pada konsentrasi ABA xilem dan potensial air ketika terevaporasi (ψbundle). LER bergantung pada potensial air xylem (Reymond et al., 2003; Welcker et al., 2011).
Model kontrol stomata berupa biosintesis hormon asam absisat (ABA) dan transfer air bergantung kepada konsentrasi ABA di dalam getah xilem dan potensial air pada titik evaporasi yang bernilai sama dengan ψbundle. Dengan demikian, dapat diasumsikan bahwa nilai LER berhubungan dengan konsentrasi ABA. Melalui model dari pengamatan pada kamar bertekanan (pressure
chamber), terdapat perbedaan yang
evaporasi di pagi hari yang menyebabkan peningkatan konduktansi stomata. Hal ini berlaku hingga nilai kapitansi sel dewasa mencapai nilai maksimum.
Gambar 10 Lingkup Waktu LER Berhubungan dengan Variabel pada
Pertumbuhan Tanaman di Tanah. A menunjukkan LER (hitam) dan laju transpirasi (J; merah). B menunjukkan potensial air pada daun yang tertutup (biru)
dan yang terbuka (hitam) dari cahaya matahari. C menunjukkan konduktansi
hidrolik seluruh tanaman (Kh). (Sumber : Caldeira, et al., 2014) 4. DISKUSI
4.1 Waktu Paruh LER Bersifat Kompatibel terhadap Kontrol Hidrolik dari Pertumbuhan Daun pada Defisit Air
Time courses (lingkup waktu)
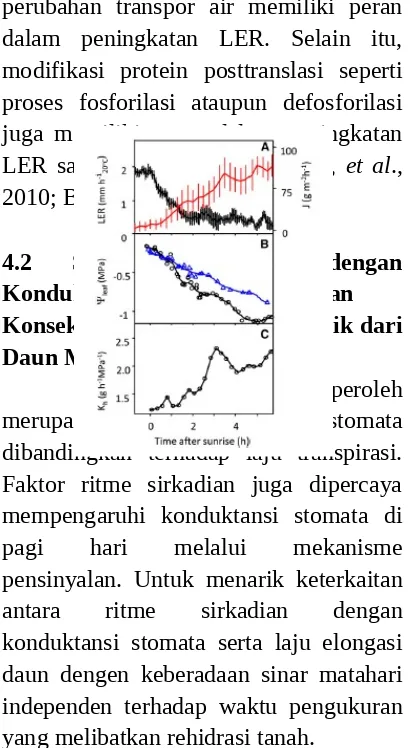
dari LER saat pagi buta dan masa pemulihan setelah rehidrasi tanah atau saat sore hari memiliki nilai waktu paruh sebesar 20-30 menit. Respon yang cepat disebabkan karena efek dari tuntutan penguapan dan perubahan yang cepat dari VPD pada growth chambers. Respon yang cepat ini tidak berhubungan pada efek kekurangan air. Efek ini akan berpengaruh pada waktu yang relatif lama. Variasi waktu dari LER memiliki kesamaan dari konduktansi stomata. Kesamaan dengan konduktansi stomata tersebut mengindikasikan bahwa
perubahan transpor air memiliki peran dalam peningkatan LER. Selain itu, modifikasi protein posttranslasi seperti proses fosforilasi ataupun defosforilasi juga memiliki peran dalam peningkatan LER saat rehidrasi tanah (Novak, et al., 2010; Bonhomme, et al., 2012).
4.2 Sinkronisasi LER dengan Konduktansi Stomata Merupakan Konsekuensi Arsitektur Hidraulik dari Daun Monokotil
Nilai LER yang diperoleh merupakan efek dari pembukaan stomata dibandingkan terhadap laju transpirasi. Faktor ritme sirkadian juga dipercaya mempengaruhi konduktansi stomata di pagi hari melalui mekanisme pensinyalan. Untuk menarik keterkaitan antara ritme sirkadian dengan konduktansi stomata serta laju elongasi daun dengen keberadaan sinar matahari independen terhadap waktu pengukuran yang melibatkan rehidrasi tanah.
bernilai rendah ketika pengukuran pada siang hari dilakukan.
4.3 Peningkatan Konduktansi Hidrolik Tanaman pada Awal Pagi yang Membatasi Penurunan LER, secara Spesial dalam Kondisi Pemberian Air yang Baik
Konduktansi hidrolik tanaman mengalami peningkatan di pagi hari. Hal ini membatasi pengurangan LER karena pembukaan stomata. Pada tanaman yang memproduksi asam absisat dalam kadar yang relatif rendah, terjadi penurunan LER dalam waktu yang relatif cepat. Hal yang sebaliknya terjadi pada tanaman yang memproduksi asam absisat dalam kadar berlebih. Dengan demikian, dapat dinyatakan bahwa penurunan LER yang berlangsung cepat di bawah ketersediaan air yang hampir cukup dapat dikaitkan
terhadap rendahnya kontribusi
konduktansi hidrolik tanaman terhadap konduktansi total dari tanah ke daun. Pada tanah yang lembap dengan tekanan sebesar -0,01 Mpa, konduktansi hidrolik tanaman bernilai lebih rendah dibandingkan konduktansi hidrolik tanah sehingga pergerakan air terhambat (Draye, et al., 2010). transpirasi. Namun, sebenarnya faktor lain seperti suhu dan cahaya pada ritme sirkadian dapat memberikan pengaruh terhadap faktor genetik, seperti
pembentukan transkrip protein
aquaporin yang terlebih lanjutnya akan mempengaruhi konduktivitas hidrolik tanaman (Cochard, et al., 2007).
4.4. Pemicu Hidrolik dari Perubahan LER tidak Menggarisbawahi
Mekanisme Nonhidrolik pada Zona Elongasi
tuntutan evaporatif yang diberikan akan memberikan hubungan antara mekanisme hidrolik dengan perubahan akhir luas permukaan daun didukung dengan perubahan kondisi lingkungan serta variabilitas genetik. Hal ini terlebih lanjutnya didukung oleh analisis genetik yang menunjukkan komunitas quatitative trait loci antara sensitivitas laju elongasi daun terhadap defisit air tanah dan tuntutan evaporatif, liputan waktu pada penurunan laju elongasi daun di pagi hari dan keselurahan sensitivitas laju elongasi daun terhadap perubahan tuntutan evaporatif, serta kontrol genetik dari laju elongasi daun terhadap luas akhir daun.
6. REFERENSI
Caldeira, C.F., Bosio, M., Parent, B. Jeanguenin, L., Chaumont, F., dan Tardieu, F. 2014. “A Hydraulic Model Is Compatible with Rapid Changes in Leaf Elongation under Fluctuating Evaporative Demand and Soil Water Status.”
Plantphysiol. 164(1), 1718-1730
Cochard, H., Venisse, J.S., Barigah, T.S., Brunel, N., Herbette, S., Guilliot, A., Tyree, M.T., dan Sakr, R. 2007. “Putative Role of Aquaporins in Variable Hydraulic Conductance of Leaves in Response to Light.”
Plant Physiology. 143(1), 122-133
Draye, X., Kim, Y., Lobet, G., Javaux, M. 2010. “Model-Assisted Integration of Physiological and
Environmental Constraints
Affecting the Dynamic and Spatial Patterns of Root Water Uptake from Soils.”Plant Cell Environment. 36(1), 1105-1119
Hachez, C., Veselov, D., Ye, Q., Reinhardt, H., Knipfer, T., Fricke, W., dan Chaumont, F. 2012.
“Short-Term Control of Maize Cell and Root Water Permeability Through Plasma Membrane Aquaporin
Isoforms.” Plant Cell
Environmental. 35(1), 185–198.
Lopez, F., Bousser, A., Sissoëff, I., Gaspar, M., Lachaise, B., Hoarau, dan J., Mahé, A. 2003. “Diurnal Regulation of Water Transport and Aquaporin Gene Expression in Maize Roots: Contribution of PIP2 Proteins.” Plant Cell Physiology. 44(1), 1384–1395.
Maurel, C., Verdoucq, L., Luu, D.T., dan Santoni, V. 2008. “Plant Aquaporins: Membrane Channels with Multiple Integrated Functions.
Annual Revision Plant Biology.
59(1), 595–624
Novak, B., Kapuy, O., Domingo-Sananes, M.R., dan Tyson, J.J. 2010. “Regulated Protein Kinases and Phospatases in Cell Cycle Decisions.” Current Opinions Cell
Biology. 22(2), 801-808
Parent, B., Hachez, C., Redondo, E., Simonneau, T., Chaumont, F., dan Tardieu, F. 2009. “Drought and Abscisic Acid Effects on Aquaporin Content Translate into Changes in Hydraulic Conductivity and Leaf Growth Rate: A Transscale Approach.”
Plant Physiology. 149(2),
2000-2012
Prado, K., Boursiac, Y., Tournaire-Roux, C., Monneuse, J.M., Postaire, O., Da Ines, O., Schäffner, A.R., Hem, S., Santoni, V., dan Maurel, C. 2013. “Regulation of Arabidopsis
Leaf Hydraulics Involves Light-Dependent Phosphorylation of Aquaporins in Veins.” Plant Cell.
Reymond, M., Muller, B., Leonardi, A., Charcosset, A., dan Tardieu, F. 2003. “Combining Quantitative Trait Loci Analysis and An Ecophysiological Model to Analyse the Genetic Variability of the Responses of Leaf Growth to Temperature and Water Deficit.”
Plant Physioogyl. 131(1), 664–675
Tardieu, F., dan Davies, W.J. 1993. “Integration of Hydraulic and Chemical Signalling in the Control of Stomatal Conductance and Water
Status of Droughted Plants.” Plant
Cell Environment. 16(1), 341-349
Welcker C, Sadok W, Dignat G, Renault M, Salvi S, Charcosset A, dan Tardieu, F. 2011. “A Common
Genetic Determinism for