PEMANFAATAN PENCIRI GEN К-KASEIN UNTUK SELEKSI
PADA SAPI DAN KERBAU
HASANATUN HASINAH dan EKO HANDIWIRAWAN
Pusat Penelitian dan Pengembangan Peternakan Jl. Raya Pajajaran Kav E-59, Bogor 16152
ABSTRAK
Kasein merupakan salah satu protein yang paling banyak ditemukan di dalam susu. Polimorfisme gen kasein susu telah diduga berhubungan dengan perbedaan komposisi nutrisi susu, prosesing dan kualitas dan juga dengan karakteristik produksi. Apabila hubungan itu dapat ditemukan dan cukup erat dan hubungan itu merupakan sifat khas dari seluruh populasi, maka dapat digunakan untuk seleksi sebagai indikator produktivitas. Dengan penciri genetik tersebut potensi produksi ternak dapat diketahui secara lebih dini dan lebih efisien. Polimorfisme genetik κ-kasein telah dapat dideteksi pada level protein maupun DNA, pada sapi yang telah diidentifikasi umumnya terdiri dari dua alel yaitu A dan B dengan menggunakan teknik PCR-RFLP dan enzim restriksi Hind III dan Taq I. Alel B dilaporkan sangat menguntungkan untuk produksi susu tetapi mungkin bersifat resesif karena tidak ada perbedaan antara genotipe AA dan AB. Alel B juga bertanggung jawab untuk protein susu dan persen protein susu yang tinggi, tetapi tidak berpengaruh pada persen lemak susu. Hasil-hasil penelitian tersebut menunjukkan bahwa penciri genetik gen κ-kasein pada sapi dapat dipergunakan sebagai alat bantu dalam menseleksi produksi susu dan komposisi susu. Beberapa laporan hasil penelitian pada beberapa bangsa kerbau menunjukkan bahwa frekuensi alel pada gen κ-kasein berbeda dengan yang ditemukan pada sapi. Beberapa peneliti melaporkan tidak ditemukannya alel A pada kerbau, keseluruhan melaporkan monomorfik, baik dengan teknik PCR-RFLP maupun SSCP. Alel yang ditemukan pada penelitian tersebut adalah hanya alel B, keseluruhan individu kerbau bergenotipe BB. Hasil penelitian tersebut menunjukkan bahwa dengan tidak adanya polimorfisme pada lokus gen κ-kasein tidak memungkinkan mengetahui pengaruh alel terhadap produksi dan komposisi susu pada kerbau. Dengan demikian penciri PCR-RFLP (dengan enzim restriksi Hind III dan Hinf I) dan SSCP tidak dapat dipergunakan sebagai penciri genetik sebagai alat bantu seleksi pada kerbau. Masih diperlukan penelitian lebih jauh mengenai hal ini untuk kerbau Indonesia atau pencarian penciri genetik lain yang menunjukkan polimorfisme dan berhubungan erat dengan produksi dan komposisi susu untuk dapat dipergunakan sebagai alat seleksi. Kata Kunci: Penciri genetik, κ-kasein, seleksi, sapi, kerbau
PENDAHULUAN
Susu dan produk susu telah dikenal sebagai bahan makanan yang bergizi tinggi dan sangat penting untuk kebutuhan manusia karena mengandung zat yang sangat diperlukan oleh tubuh seperti protein, karbohidrat, lemak, vitamin dan mineral. Susu juga dapat dibuat antara lain menjadi produk olahan asal susu, seperti susu bubuk, keju dan lain-lain. Komposisi kandungan utama susu bervariasi di antara spesies, tetapi semua susu mengandung kandungan nutrisi yang sama. Di dalam spesies faktor genetik dan kondisi lingkungan mempengaruhi komposisi susu (OTAVIANO et
al., 2005). Oleh karena itu, peningkatan
produktivitas ternak dapat dilakukan melalui perbaikan faktor lingkungan yang meliputi pemberian pakan, perawatan, lingkungan atau
melalui perbaikan genetik. Kelompok ternak yang baik dapat diperoleh dengan cara seleksi terhadap kemampuan produksi dan komposisi susu yang dihasilkan, yang merupakan sifat yang mempunyai nilai ekonomi tinggi.
Beberapa variasi genetik ditemukan pada sebagian besar protein susu dan menunjukkan pengaruh pada produksi susu dan komposisi susu serta secara tidak langsung akan mempengaruhi hasil pengolahan susu seperti keju. Polimorfisme yang ada pada protein susu disebabkan oleh variasi genetik dan varian tersebut diwariskan berdasarkan hukum Pewarisan Mendel sederhana non dominan (OTAVIANO et al., 2005). Variasi tersebut pada umumnya dapat dideteksi melalui teknik elektroforesis berdasarkan perbedaan struktur atau muatan listrik molekul.
Kasein merupakan salah satu protein dalam susu yang diketahui jumlahnya di dalam susu paling banyak. Polimorfisme gen kasein susu telah dihubungkan dengan perbedaan komposisi susu, prosesing dan kualitas (MCLEAN, 1987) dan juga dengan karakteristik
produksi (LIN et al., 1986).
Apabila hubungan itu dapat ditemukan dan cukup erat dan hubungan itu merupakan sifat khas dari seluruh populasi, maka dapat digunakan untuk seleksi sebagai indikator produktivitas (WARWICK et al., 1990). Penciri indikator sifat produksi tersebut dapat dimanfaatkan untuk melakukan seleksi dengan lebih efisien. Seleksi dapat dilakukan pada saat ternak masih belum menunjukkan kemampuan produksinya, akan tetapi dengan penciri genetik tersebut potensi produksi ternak dapat diketahui secara lebih dini.
Penelitian dalam usaha menentukan hubungan antara perbedaan biologis atau polimorfisme dengan sifat produksi dari ternak telah banyak dilakukan. Produksi susu dunia saat ini umumnya dipenuhi dari ternak sapi dan kerbau, demikian pula di Indonesia oleh karena itu dalam makalah ini dibahas pemanfaatan penciri gen kasein sebagai alat seleksi pada ternak tersebut.
KOMPOSISI SUSU SAPI DAN KERBAU
Produksi susu di Indonesia sekitar 577,6
ribu ton (DIREKTORAT JENDERAL
PETERNAKAN, 2006) yang sebagian besar dihasilkan oleh sapi perah dan hanya sebagian kecil yang diproduksi ternak kerbau. Sapi
perah yang utama di Indonesia adalah sapi Friesian Holstein (FH).
Di Indonesia, rata-rata produksi susu sapi FH/PFH berkisar antara 2500-3500 kg per laktasi dengan kadar lemak 3,7% (SYARIEF dan
SUMOPRASTOWO, 1990). Sementara itu,
HARDJOSUBROTO (1994) mengemukakan bahwa lebih dari 5% produksi susu di dunia berasal dari ternak kerbau. Di India, 60% konsumsi susu berasal dari susu kerbau. Produksi susu di India tercatat rata-rata 4 sampai 7 kg per hari selama masa laktasi 285 hari. Di Pakistan dilaporkan bahwa produksi susu kerbau Nili/Ravi sebesar 1.925 kg dengan masa laktasi selama 282 hari. Secara umum produksi susu kerbau lumpur di Indonesia adalah rendah, sekitar 2 liter/ekor/hari (COCKRILL, 1974 yang disitasi SIREGAR et al.,
1997).
Susu kerbau, seperti halnya dengan susu sapi dapat diproses menjadi berbagai macam produk seperti keju, mentega, es krim, yoghurt dan buttermilk. Untuk membuat 1 kg keju dibutuhkan 8 kg susu sapi, tetapi dengan susu kerbau hanya 5 kg, sedangkan untuk membuat 1 kg mentega yang dengan susu sapi dibutuhkan 14 kg, dengan susu kerbau hanya membutuhkan 10 kg.
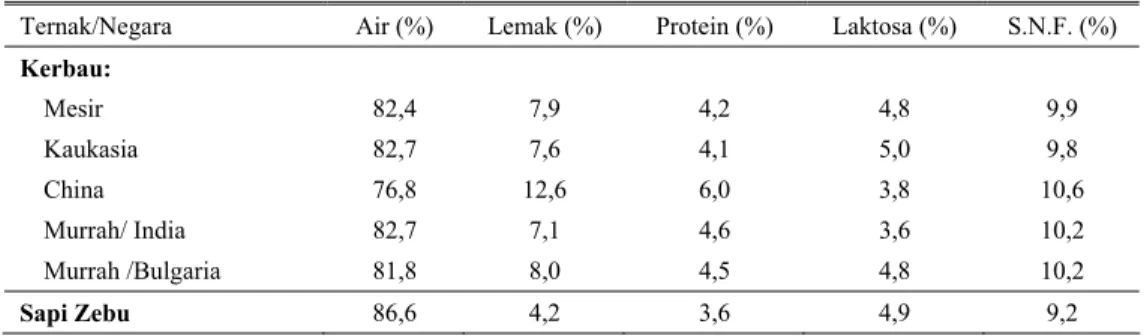
Susu dari spesies yang berbeda secara umum mengandung penyusun yang sama tetapi mempunyai komposisi yang bervariasi dan tergantung pada banyak faktor. Tabel 1 memperlihatkan variasi komposisi air susu kerbau di berbagai tempat dan sapi Zebu.
Tabel 1. Komposisi susu kerbau dan sapi Zebu
Ternak/Negara Air (%) Lemak (%) Protein (%) Laktosa (%) S.N.F. (%) Kerbau: Mesir 82,4 7,9 4,2 4,8 9,9 Kaukasia 82,7 7,6 4,1 5,0 9,8 China 76,8 12,6 6,0 3,8 10,6 Murrah/ India 82,7 7,1 4,6 3,6 10,2 Murrah /Bulgaria 81,8 8,0 4,5 4,8 10,2 Sapi Zebu 86,6 4,2 3,6 4,9 9,2
Perbedaan terbesar komposisi susu kerbau dan sapi adalah pada kandungan lemak. Pada sapi kandungan lemak antara 3 – 5%, sedangkan pada kerbau kandungan lemak berkisar antara 7 – 12% tetapi bisa juga mencapai 13%. Sementara itu, kandungan protein, laktosa dan abu agak lebih tinggi pada susu kerbau dibandingkan pada susu sapi (Tabel 1).
Komposisi susu dipengaruhi oleh beberapa faktor termasuk bangsa, variasi genetik dalam bangsa, kesehatan, lingkungan, manajemen, stadium laktasi, pakan dan umur (ECKLES et
al., 1957, LAMPERT, 1975; BATH et al., 1985). Protein Susu
Susu mengandung sejumlah protein yang jumlahnya berkisar antara 2,8-4,0% (ECKLES et
al., 1957) dan menurut SOEPARNO et al. (2001) protein dalam susu terdiri atas kasein (80%), laktalbumin (18%) dan laktoglobulin (0,05 - 0,07%).
Kasein merupakan komplek senyawa protein dengan garam Ca, P dan sejumlah kecil Mg dan sitrat sebagai agregat makromolekul yang disebut kalsium fosfo-kaseinat atau misel kasein (ESKIN et al., 1990). Kasein dapat
dipresipitasi oleh asam atau enzim rennin dan presipitasi kasein oleh rennin ini merupakan dasar untuk pembentukan curd dalam keju (BATH et al., 1985).
Kasein terdapat dalam susu sebagai suatu suspensi koloidal partikel-partikel kompleks yang disebut misel (SOEPARNO, 1992). Kasein
terdiri dari tiga komponen yaitu α-kasein, β-kasein dan δ-β-kasein. Alfa-β-kasein dan β-β-kasein terbentuk di dalam kelenjar susu atau ambing sedang δ-kasein mula-mula ditemukan di dalam aliran darah kemudian masuk ambing lalu bergabung dengan kompleks α-kasein dan dikenal sebagai κ-kasein (LAMPERT, 1975).
κ-kasein adalah protein susu yang menyusun sekitar 12 - 15% dari total kasein pada susu sapi dan bertindak sebagai stabilisasi, yaitu mempertahankan seluruh kompleks kasein dalam suspensi koloidal yang memberikan warna putih susu (SOEPARNO,
1992). Jumlah dan tipe κ-kasein persentasenya berbeda-beda tergantung pada individu sapi itu sendiri (NG-KWAI-HANG et al., 1991).
Seleksi menggunakan Penciri Genetik
Teknik genetika molekuler dapat memberikan hasil yang lebih menguntungkan untuk program pemuliaan karena dapat menentukan potensi seekor ternak, sebelum fenotipenya diketahui. Menurut HALEY (1995),
penciri DNA menunjukan ada dua kemungkinkan aplikasi program seleksi untuk ternak yaitu kombinasi dua alel atau lebih bangsa dan seleksi alel dalam satu bangsa. Hal ini memberikan peluang untuk mempergunakan genotipe κ-kasein sebagai alat bantu dalam pelaksanaan seleksi pada sapi dan kerbau.
Pada Sapi Perah
Gen kasein sapi meliputi sebuah fragmen sepanjang 200 kb di kromosom 6, yang dirangkai secara tandem dengan urutan : α-s1, β, α-s2, dan κ (LIEN dan ROGNE, 1993).
Khusus gen κ-kasein meliputi sebuah fragmen sepanjang 13 kb yang dibagi ke dalam 5 ekson (ALEXANDER et al., 1998).
Polimorfisme genetik κ-kasein telah dapat dideteksi pada level protein maupun DNA. Perbedaan kedua alel tersebut adalah alel B mempunyai isoleusin dan alanin berturut-turut pada posisi 136 dan 148 urutan asam amino sebagai pengganti treonin dan aspartat pada alel A (MIRANDA et al., 1993).
Hubungan antara alel A dan alel B κ-kasein dengan produksi susu, lemak susu dan protein susu telah banyak dilaporkan dengan sedikit perbedaan disebabkan oleh perbedaan bangsa, populasi dan metode analisis. Alel B dilaporkan sangat menguntungkan untuk produksi susu tetapi mungkin bersifat resesif karena tidak ada perbedaan antara genotipe AA dan AB. Alel B juga bertanggung jawab untuk protein susu dan persen protein susu yang tinggi, tetapi tidak berpengaruh pada persen lemak susu. LIN et al. (1992) dan MCLEAN
(1987) menyatakan bahwa genotipe κ-kasein BB berhubungan dengan proses pembuatan keju. SABOUR et al. (1993) menyatakan
beberapa variasi genetik pada kasein susu tersebut mempengaruhi komposisi susu serta hasil pengolahan susu seperti keju. Genotipe AB dilaporkan menunjukkan produksi susu yang lebih tinggi dibanding genotipe AA dan
BB, tetapi pada laporan lain disebutkan bahwa genotipe BB lebih tinggi dibandingkan kedua genotipe lain (MAO et al., 1992). Hasil-hasil penelitian tersebut menunjukkan bahwa penciri genetik gen κ-kasein pada sapi dapat dipergunakan sebagai alat bantu dalam menseleksi produksi susu dan komposisi susu.
Seleksi terhadap alel B mulai dilakukan terhadap pejantan IB sapi perah di Kanada. Seleksi pejantan IB diarahkan ke pejantan dengan genotipe BB, karena sifat-sifat baik seperti heat stability dan curd yang padat serta kandungan protein yang tinggi terutama kasein susu yang dibutuhkan oleh industri keju (SABOUR et al., 1993).
EENENNAAM dan MEDRANO (1991) melaporkan penggunaan susu dari sapi yang bergenotipe κ-kasein BB mempengaruhi hasil/proses pembuatan keju, waktu koagulasi rennet lebih pendek, bentuk curd lebih padat dan produk keju yang dihasilkan lebih bagus dibandingkan dengan penggunaan susu yang berasal dari sapi yang bergenotipe κ-kasein AA. Pengaruh ini berhubungan dengan misel kasein susu yang ukurannya bervariasi antara sapi yang membawa genetik κ-kasein yang berbeda.
Misel yang kecil mengandung proporsi κ-kasein lebih baik daripada misel yang besar. Susu yang berasal dari sapi yang membawa genotipe κ-kasein AA mengandung banyak misel yang besar, ini dikarenakan oleh rendahnya proporsi κ-kasein dalam fraksi kasein pada susu dari κ-kasein AA. κ-kasein BB dalam susu penyebaran ukuran misel lebih homogen, curd saling berikatan didalam proses pembuatan keju (EENENNAAM dan MEDRANO,
1991).
Alel A dan B terdapat pada Bos Taurus dan
Bos indicus, frekuensinya hampir sama pada
semua bangsa kecuali untuk FH (produksi susu) dan Jersey (produksi lemak) mempunyai frekuensi B masing-masing 0,32 dan 0,77 (BARROSO et al., 1997). Alel A banyak terdapat pada sapi Friesian, Ayrshire, Red Danish dan Zebu India, sedangkan alel B banyak ditemukan pada sapi Jersey, Normande, dan Zebu Afrika. Sapi potong umumnya mempunyai alel B. MITRA et al. (1998) dengan teknik PCR-RFLP (Polymerase
Chain Reaction-Restriction Fragment Length Polymorphism) telah menemukan alel A dan B
pada sapi Sahiwal dengan menggunakan enzim restriksi Hind III, Hinf I dan Taq I dengan frekuensi alel B sebesar 0,16. Pada sapi Sahiwal ditemukan genotipe AA dan genotipe AB sedangkan genotipe BB pada sapi ini tidak terdeteksi.
HASINAH (2003) dengan teknik PCR-RFLP melaporkan telah mengidentifikasi 3 genotipe κ-kasein pada sapi perah di BPTU Baturaden yaitu AA, AB dan BB, hasil digesti fragmen DNA κ-kasein ukuran 780 bp dengan enzim
Hind III dan Pst I dengan frekuensi alel A dan
alel B berturut-turut sebesar 0,67 dan 0,33, yang berhubungan dengan kualitas susu. Pengaruh ketiga macam genotipe κ-kasein tersebut terhadap kadar lemak dan protein susu tidak menunjukkan perbedaan. Meskipun demikian hasil penelitian secara kuantitatif menunjukkan genotipe BB mempunyai kadar lemak dan protein lebih tinggi dibandingkan AA dan AB.
Pada Kerbau
Kendala dalam seleksi ternak kerbau adalah masih lemahnya identifikasi ternak dan rekording yang dilakukan. Belum terwujudnya identifikasi dan rekording yang baik dikarenakan umumnya kerbau dipelihara oleh peternak dalam jumlah kecil dan dipelihara secara tradisional serta belum cukupnya pemahaman dan kesadaran peternak terhadap mutu genetik ternak yang dipelihara, tidak sebagaimana sapi perah yang telah banyak diusahakan oleh perusahaan peternakan dengan identifikasi dan rekording yang sudah cukup baik. Oleh karena itu seleksi dengan memanfaatkan penciri genetik yang berhubungan dengan sifat produksi dan komposisi susu akan mempermudah dalam melakukan seleksi pada ternak kerbau.
Beberapa laporan hasil penelitian pada beberapa bangsa kerbau menunjukkan bahwa frekuensi alel pada gen κ-kasein berbeda dengan yang ditemukan pada sapi. Sejauh ini beberapa penulis melaporkan bahwa pada kerbau alel A tidak ditemukan. MITRA et al. (1998) dengan teknik PCR-RFLP dan enzim restriksi Hind III dan Hinf I melaporkan bahwa pada bangsa kerbau Murrah dan Nili-Ravi ditemukan monomorfik, yang kesemuanya mempunyai alel B. Demikian pula MERCIER et
al. (1998), DENICOURT et al. (1990) dan
PINDER et al. (1991) telah mempelajari ekson IV dari alel B gen κ-kasein pada kerbau Murrah, Nili-Ravi dan kerbau Mesir dengan teknik RFLP dan memperoleh hasil yang sama. Hasil serupa didapatkan oleh PIPALIA (1999)
dengan menggunakan teknik PCR-RFLP dan menggunakan enzim restriksi Hind III dan Hinf
I pada kerbau Jaffarabadi, Mehsani, Surti dan
Pandharpuri. Keseluruhan sampel pada lokus gen κ-kasein menunjukkan monomorfisme.
Dengan teknik yang berbeda yaitu SSCP (Single Strand DNA Conformation
Polymorphism), OTAVIANO et al. (2005) tidak
menemukan polimorfisme pada semua sampel kerbau betina Murrah yang diuji. Alel yang ditemukan pada penelitian tersebut adalah hanya alel B, keseluruhan individu kerbau bergenotipe BB. Diduga genotipe BB inilah yang menyebabkan kandungan lemak, protein dan curd pada susu kerbau lebih tinggi dari sapi Taurus maupun Zebu.
Hasil penelitian tersebut menunjukkan bahwa dengan tidak adanya polimorfisme pada lokus gen κ-kasein tidak memungkinkan mengetahui pengaruh alel terhadap produksi dan komposisi susu pada kerbau. Dengan demikian penciri PCR-RFLP (dengan enzim restriksi Hind III dan Hinf I) dan SSCP tidak dapat dipergunakan sebagai penciri genetik sebagai alat bantu seleksi pada kerbau. Sejauh ini, belum diketahui pula apakah ditemukan polimorfisme kerbau Indonesia dengan dua penciri tersebut. Jika terdapat variasi produksi dan komposisi susu kerbau lokal Indonesia maka hal tersebut merupakan peluang untuk melakukan seleksi dengan penciri genetik lain yang berhubungan dengan sifat tersebut.
KESIMPULAN
Beberapa variasi genetik ditemukan pada protein κ-kasein dan menunjukkan hubungan dengan produksi susu dan komposisi susu yang secara tidak langsung mempengaruhi hasil pengolahan susu seperti keju. Pada sapi (sapi Taurus dan sapi Zebu) dengan teknik PCR-RFLP telah ditemukan dua macam alel yaitu alel A dan B (sapi dengan genotipe AA, AB dan BB). Beberapa peneliti melaporkan bahwa genotipe BB terkait erat dengan produksi dan komposisi susu. Alel B bertanggung jawab
untuk protein susu dan persen protein susu yang tinggi (mempengaruhi komposisi susu serta hasil pengolahan susu seperti keju), tetapi tidak berpengaruh pada persen lemak susu. Genotipe AB dilaporkan menunjukkan produksi susu yang lebih tinggi dibanding genotipe AA dan BB, tetapi pada laporan lain disebutkan bahwa genotipe BB lebih tinggi dibandingkan kedua genotipe lain. Hasil ini menunjukkan bahwa penciri genetik gen κ-kasein pada sapi dapat dipergunakan sebagai alat bantu dalam menseleksi produksi dan komposisi susu. Berbeda dengan pada sapi, pada kerbau beberapa peneliti dengan teknik PCR-RFLP dan SSCP tidak menemukan polimorfisme pada gen κ-kasein. Sejauh ini, alel yang ditemukan pada kerbau hanya alel B, keseluruhan individu kerbau bergenotipe BB. Dengan demikian belum dapat diketahui pengaruh alel B tersebut terhadap produksi dan komposisi susu kerbau, sehingga penciri genetik tersebut tidak dapat dipergunakan sebagai alat bantu seleksi pada kerbau. Masih diperlukan penelitian lebih jauh mengenai hal ini untuk kerbau Indonesia atau pencarian penciri genetik lain yang menunjukkan polimorfisme dan berhubungan erat dengan produksi dan komposisi susu.
DAFTAR PUSTAKA
ALEXANDER, L. J., A. J. STEWART, A. G. MACHINLAY, T. V. KAPELINSKAYA, T. M.
TKACH andS.I.GORODETSKY. 1998. Isolation and characterization of the bovine kappa-casein gene. Europe Journal Biochemical 178 : 395-401.
BARROSO,A.,S.DUNNER andJ.CANON. 1997. Use of single-strand conformation polymorphisme analysis to perform simple genotyping of bovine κ-casein A, and B variants. J. Dairy Res. 64: 535-540.
BATH,D.L.,F.N.DICKINSON,H.A.TUCKER andR.
D.APPLEMEN. 1985. Dairy Cattle: Principles, Practices, Problems, Profits. Lea and Febiger. Philadelphia.
DENICOURT, G., L. FERRETI, G. ROGNONI and V.
SGARAMELLA. 1990. Restriction fragment length polymorphism analysis of the κ-kasein locus in cattle. Animal Genetics 21 : 107-114.
DIREKTORAT JENDERAL PETERNAKAN. 2006. Statistik
Peternakan. Direktorat Jenderal Peternakan. Jakarta.
ECKLES,C.H.,W.B.COMBS andH.MACY. 1957. Milk and Milk Products. Tata McGraw-Hill Publ. Co., Ltd. Bombay. New Delhi.
EENENNAAM, A. L.V. and J.F.MEDRANO. 1991. Differences in allelic protein expression in the milk of heterozygous κ-casein cows. J. Dairy Sci. 74: 1491-1496.
ESKIN, N. A. M., H. M. HANDERSON and R. J. TOWNSEND. 1990. Biochemistry of Foods.
Academic Press, Inc. New York.
HALEY, C.S. 1995. Livestock QTLs – Bringing home the bacon? Trends Genetics 11: 488-492.
HARDJOSUBROTO, W. 1994. Aplikasi Pemuliabiakan
Ternak di Lapangan. PT Gramedia Widiasarana Indonesia. Jakarta.
HASINAH, H. 2003. Efek genotype kappa kasein dan
paritas terhadap kadar lemak dan protein susu sapi FH di BPTU Baturraden. Thesis. UGM. Yogyakarta.
HÖGBERG,M.S.andO.LIND.2003. Milk Production
of Buffalo. In: Buffalo Milk Production. http://www.milkproduction.com/Library/Artic les/
Buffalo_Milk_Production_Chapter_5_Milk_p roduction_of_the_buffalo.htm [2 Juli 2007]. LAMPERT, L. M., 1975. Modern Dairy Product. 3rd
ed. Chemical Publishing Company, Inc. New York.
LIEN, S. and S. ROGNE. 1993. Bovine casein haplotypes number, frequencies and applicability as genetic markers. Animal Genetics 24 : 373-376.
LIN,C.Y.,M.P.SABOUR andA.J.LEE. 1992. Direct typing of milk proteins as an aid for genetic improvement of dairy bulls and cows: A review. Animal Breeding Abstract 60 : 1-10. MAO, I. L., L. G. BUTTAZZONI and R.ALEANDRI.
1992. Effect polymorphic milk protein genes on milk yield and composition traits in Holstein Cattle. Sect. A, Animal Sci. 42 : 1-7. MCLEAN, D. M. 1987. Influence of milk protein
variants on milk composition, yield, and cheese making properties. Animal Genetics 18: 100-102.
MERCIER,J.C.,J.M.CHOBERT andF.ADDEO. 1998.
Comparative study of the amino acids
sequences of the caseinomacropeptides from seven species. FEBS Lett 72 : 208-214. MIRANDA, P, P. ANGLADE, M. F. MAHE’ and G.
ERHARDT. 1993. Biochemical characterization of the bovine genetic κ-casein C dan E variants. Animal Genetics 24: 27-31.
MITRA, A.,P. SCHLEE, I. KRAUSE, J. BLUSCH, T. WEMER, C. R. BALAKRISHNAN and F.
PIRCHNER. 1998. Kappa-casein poly-morphisms in Indian Dairy cattle and buffalo: A new genetic variant in buffalo. Animal Biotechnology 9 (2): 81-87.
NG-KWAI HANG,K.F.,D.ZADWORNY,J.F.HAYES
andU.KUHNLEIN. 1991. Identification of
κ-casein genotype in Holstein sires: Comparison between analysis of milk sample from daughters and direct analysis of semen samples from sires by polymerase chain reaction. J. Dairy Sci. 74: 2410-2415.
OTAVIANO,A.R.,H.TONHATI,J.A.D.SENA and M. F.C.MUÑOZ. 2005. Kappa-casein gene study
with molecular markers in female buffaloes (Bubalus bubalis). Genetics and Molecular Biology 28 (2): 232-241.
PINDER,S.J.,B.N.PERRY,C.J.SKIDMORE andD. SAVVA. 1991. Analysis of polymorphism in the bovine casein genes by use of the polymerase chain reaction. Animal Genetics 22: 11-20.
PIPALIA, D. L. 1999. Genotyping of various buffalo breeds for κ-casein using PCR-RFLP technique. Thesis. Department of Animal Genetics and Breeding. Gujarat Agricultural
University. Anand. openmed.nic.in/2261/01/Dr._D.L.Pipaliya_M.
V.Sc._Thesis.pdf [2 Juli 2007].
SABOUR, M. P., C. Y. LIN, A. KEOUGH, S. M.
MECHANDA and A. J. LEE. 1993. Effects of Selection Practiced on the Frequencies of κ-casein and β-lactoglobulin genotypes in Canadian artivicial insemination bulls. J. Dairy Sci. 76: 274-280.
SIREGAR, A. R., K. DIWYANTO, E. BASUNO,
A. THALIB, T. SARTIKA, R.H. MATONDANG, J. BESTARI, M. ZULBARDI, M. SITORUS,
T. PANGGABEAN, E. HANDIWIRAWAN,
Y. WIDIAWATI dan N. SUPRIYATNA. 1997. Karakteristik performan nutrisi, mikroba rumen, morfologi darah dan dinamika populasi kerbau lumpur di Pulau Jawa. Seminar Nasional Peternakan dan Veteriner 1997. Pulitbang Peternakan. Bogor. Hlm 555-570.
SOEPARNO, 1992. Susu dan Komposisi Susu.
Fakultas Peternakan Universitas Gadjah Mada. Yogyakarta.
SYARIEF, M. Z.danR.M.SUMOPRASTOWO. 1990. Ternak Perah. CV. Yasaguna. Jakarta.
WARWICK, E. J., J. M. ASTUTI dan W.
HARDJOSUBROTO. 1990. Pemuliaan Ternak. Gadjah Mada University Press. Yogyakarta.