L
Journal of Experimental Marine Biology and Ecology 249 (2000) 13–27
www.elsevier.nl / locate / jembe
Exopolysaccharide production by the epipelic diatom
Cylindrotheca closterium: effects of nutrient conditions
a ,* b a
Natascha Staats , Lucas J. Stal , Luuc R. Mur
a
Laboratory for Microbiology /ARISE, University of Amsterdam, Nieuwe Achtergracht 127, 1018 WS Amsterdam, The Netherlands
b
Netherlands Institute of Ecology, Centre for Estuarine and Coastal Ecology, P.O. Box 140, 4400 AC Yerseke, The Netherlands
Received 10 June 1999; received in revised form 27 January 2000; accepted 1 February 2000
Abstract
During the stationary phase of a batch culture of the epipelic diatom Cylindrotheca closterium, accumulation of exopolysaccharides and intracellular carbohydrates was observed. When nitrogen was added to the culture in the stationary phase, growth was resumed and the accumulation of exopolysaccharides was delayed. This indicated that nitrogen depletion caused cessation of growth, and stimulated exopolysaccharide accumulation. Exopolysaccharide accumulation was also stimulated when cells were either resuspended in medium lacking N or P, or when they were inoculated in medium with low concentrations of N or P. Growth was not immediately affected by low N or P concentrations. S depletion only resulted in exopolysaccharide accumulation when growth was affected. Si or Fe depletion did not stimulate exopolysaccharide accumulation, even when growth rates were lowered. Apparently, stimulation of exopolysaccharide accumulation is dependent on the type of nutrient depletion. Intracellular storage carbohydrates did not accumulate when cells were incubated at low N or P concentrations. Cells grown with ammonium as nitrogen source produced more carbohydrates (both extracellular and intracellular) than cells grown with nitrate as nitrogen source, indicating that both exopolysaccharides and intracellular carbohydrates accumulated as a result of overflow metabolism. 2000 Elsevier Science B.V. All rights reserved.
Keywords: Cylindrotheca closterium; Diatoms; Exopolysaccharides; Microphytobenthos; Nutrients
1. Introduction
Surficial sediments of tidal mudflats are often colonized by epipelic diatoms which can produce large amounts of exopolysaccharides. These exopolysaccharides form a
*Corresponding author. Present address: Max Planck Institute for Marine Microbiology, Celsiusstrasse 1, 28359 Bremen, Germany. Tel.: 149-421-2028-634; fax: 149-421-2028-580.
14 N. Staats et al. / J. Exp. Mar. Biol. Ecol. 249 (2000) 13 –27
matrix that may increase the resistance of the sediment bed towards erosion (Holland et al., 1974; Dade et al., 1990; Yallop et al., 1994; Paterson, 1997). It seems that the degree of stabilization is related to the amount of exopolysaccharide present in the sediment (Sutherland et al., 1998). Hence, it is of interest to understand the factors that control the production of exopolysaccharide.
Secretion of polysaccharides by epipelic diatoms is involved in locomotion, facilitat-ing adhesion of the cell to a substratum (Edgar and Pickett-Heaps, 1984; Lind et al., 1997; Wang et al., 1997; Wetherbee et al., 1998). However, other factors may also control exopolysaccharide production. For instance, in a previous investigation we demonstrated that oxygenic photosynthesis is required for polysaccharide secretion in the epipelic diatom Cylindrotheca closterium (Ehrenb.) Reimann and Lewin (Staats et al., 1999b). In the present study the effects of different nutrient conditions on exopolysaccharide production by C. closterium are investigated.
Much work has been done on the effect of nutrients on extracellular carbohydrate production by pelagic diatom species. Several reports indicate that it is influenced by growth rate and type of nutrient depletion (Myklestad and Haug, 1972; Myklestad, 1977; Waite et al., 1995). However, little is known with respect to the effect of nutrient conditions on polysaccharide secretion by epipelic diatoms. It is generally assumed that growth of benthic diatoms is not limited by nutrients, because nutrient concentrations in
´
pore water are generally high (Cadee and Hegeman, 1974; Admiraal et al., 1982; Sundby et al., 1992). However, in the thin layer of diatoms on the sediment surface biomass may be highly concentrated, and therefore nutrients may temporarily become depleted (Admiraal, 1977; Flothmann and Werner, 1992). Indeed, evidence is accumulat-ing that at high population densities, growth of benthic diatoms may become nutrient limited, especially by nitrogen (Sullivan and Daiber, 1975; Van Raalte et al., 1976; Hillebrand and Sommer, 1997).
The central question of this paper was whether exopolysaccharide production was affected by nutrient conditions. When benthic diatoms are grown in batch culture, polysaccharide secretion increases during the transition from exponential to stationary growth (Bhosle et al., 1995; Sutherland et al., 1998; Staats et al., 1999a). It was therefore of interest to know which factor induced the transition to stationary growth, and the associated exopolysaccharide production in cultures of C. closterium, e.g. light limitation or a nutrient starvation. And, in case of a nutrient starvation, which nutrient depletion caused stationary growth? A related question, whether the type of nutrient influenced polysaccharide secretion, was also considered. Finally, the effect of different inorganic nitrogen sources (ammonium or nitrate) on polysaccharide secretion was investigated.
2. Materials and methods
2.1. Organism and culture conditions
Haren, The Netherlands). Cells were grown in Kester medium (Kester et al., 1967). The medium had a salinity of 33 PSU. The diatoms were cultivated on a substrate of purified sea sand (Merck, Darmstadt, Germany) in 300 or 1000 ml glass Erlenmeyers at 208C.
22 21
Cultures were illuminated at an incident photon irradiance of 60 mmol m s , provided by fluorescent tubes (Philips TLE 32 / 33) with a light–dark cycle of 12:12 h. Cells were harvested by gently shaking the culture, until all cells were in suspension. The sand was allowed to sediment and the cell suspension was poured off. Axenity of the cultures was checked by plating on agar as well as by direct microscopic observation.
2.2. Induction of stationary phase
An experiment was carried out to assess whether stationary growth was induced by shortage of light or by nutrient depletion. A culture entering the stationary phase was divided into three portions. One portion was incubated at a photon irradiance of 100
22 21
mmol m s , another portion received a pulse of nutrients yielding the original medium concentration at day 0, and the third portion served as a control and was
22 21
incubated at 60mmol photons m s and no addition of nutrients.
1
Nitrogen in the medium was supplied in the form of NH , and therefore it was4 1
supposed that an increase in pH might result in a rapid decrease in NH4 concentration caused by dissociation into NH gas. In order to verify whether stationary phase was3
induced by nitrogen depletion, a culture entering the stationary phase was divided into two portions. One portion was given a pulse of NH Cl (final concentration: 0.5 mM),4
the other portion served as a control and was not treated. Samples were taken in triplicate from a single culture. The experiments were repeated independently, results from one culture are presented.
2.3. Nutrient depletion experiments
To test the effect of different nutrient depletions on exopolusaccharide accumulation, cells were grown in complete medium for 7 days, followed by harvesting and resuspension in medium devoid of either Fe, Si, P, S or N. Resuspension in complete medium served as a control. Cells that were resuspended in medium without Si were grown in polycarbonate vessels, and without sand substratum. After 4 days of incubation in the depleted medium, part of the culture was harvested and analyzed and the remainder was resuspended in fresh medium (depleted of the same inorganic nutrient). After another 4 days of incubation cultures were analyzed. In addition, cell density was measured 2 days after each resuspension. Samples were taken in triplicate from two independent cultures for each treatment.
To examine the influence of low N or P supply on exopolysaccharide accumulation, cells were inoculated in medium with low concentrations of N (95.5 mM NH Cl) or P4
*
(7.7 mM NaH PO H O). These concentrations were calculated to approximate the2 4 2 21
16 N. Staats et al. / J. Exp. Mar. Biol. Ecol. 249 (2000) 13 –27
standard medium (Staats et al., 1999b). Thus, it was ensured that stationary growth was induced by the nutrient provided in a low concentration. In the control the regular medium concentrations were used, which were 0.5 mM N and 50mM P. Samples were taken in triplicate from a single culture. The experiment was repeated independently, results from one culture are presented.
2.4. Nitrogen source experiment
To investigate the role of N source on exopolysaccharide accumulation, cells were
1 2
grown with either NH4 (0.5 mM) or NO3 (5.9 mM). Samples were taken in triplicate from three independent cultures for each treatment.
2.5. Isolation of exopolysaccharides
The cell suspension was centrifuged for 15 min at 20 000 g and 108C. The pellet was extracted with water for 1 h at 308C. This extract was subsequently centrifuged for 15 min at 20 000 g and 108C. Of several methods tested, this was the most efficient method for extraction of exopolysaccharides (Staats et al., 1999a). The extract was not contaminated with intracellular carbohydrates since no cell lysis occurred during the extraction. Furthermore, microscopic observations confirmed that all extracellular material was removed from the cells (Staats et al., 1999a). Therefore, this method was efficient in separating intra- and extracellular material. Polymers were isolated from the culture medium and from the warm water extract by overnight precipitation in cold (2208C) 80% (v / v) ethanol. The precipitated polysaccharides were dried under a flow of nitrogen gas and subsequently stored dry at 2208C before analysis. For total exopolysaccharide amounts, the sum of the amounts in the medium and water extracts was taken.
2.6. Analyses
¨ ¨
Cells were counted by Coulter counter and by microscope using a Burker–Turk counting chamber (a minimum of 300 cells was counted). Cell protein was determined on freeze-dried cell pellets with the Lowry method (Herbert et al., 1971), using bovine serum albumin as reference. As a biomass parameter, protein concentration was preferred to cell density because cells became smaller during batch growth (Staats et al., 1999a). Only in experiments in which treatments resulted in different cell protein contents, cell density was used as a biomass parameter.
Intracellular carbohydrates were determined on freeze-dried cells from which all exopolysaccharides had been removed, applying the extraction procedure as described above. Exopolysaccharides were analyzed with the phenol / H SO assay, which includes2 4
both hexoses and pentoses (Dubois et al., 1956). Glucose was used as standard. The phenol / H SO assay was not suitable for analysis of whole algal cells due to strong2 4
carbohydrate consisted mainly of hexoses (Staats et al., 1999a) these methods yielded comparable results.
Soluble reactive phosporous was analyzed using the method of Murphy and Riley (1962). Ammonium was analyzed by the method of Kempers and Kok (1989).
3. Results
3.1. Induction of stationary phase
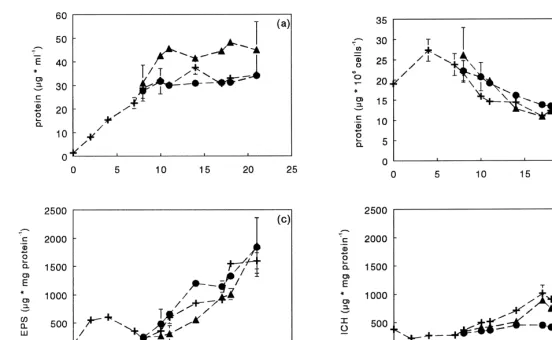
In the culture to which the concentrated mixture of nutrients was added at the beginning of the stationary phase, growth was resumed and continued for approximately more 3 days (Fig. 1a). Protein content of the cells was comparable in all treatments (Fig.
22 21
1b). Incubation at a higher photon flux density (100 mmol m s ) had no effect on
22 21
growth compared to the control (60mmol m s ) (Fig. 1a). Three days after addition of the nutrient mixture, the culture entered the stationary phase again. There was a lag in exopolysaccharide production in the nutrient treated culture compared to the control (Fig. 1c). The nutrient enriched culture started to accumulate exopolysaccharides after day 11, which coincided with the onset of stationary phase. Intracellular carbohydrates also accumulated (Fig. 1d), but at a lower rate than exopolysaccharides.
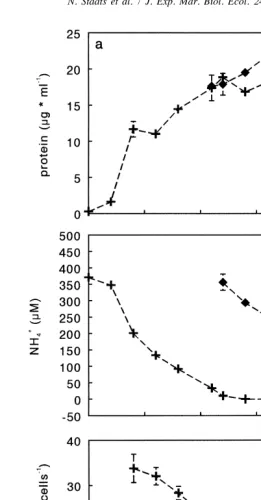
1
The onset of stationary phase in the control culture coincided with depletion of NH4 1
(Fig. 2a,b). The addition of NH4 caused an increase in cell density compared to the control (Fig. 2a).Cell protein content was similar to the control (Fig. 2c), so the increase in protein concentration was not the result of an increase in protein content of the cells.
3.2. Nutrient depletion experiments
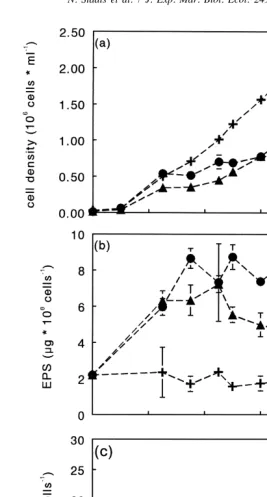
Table 1 shows the growth rates that were obtained when cells were resuspended in medium without Si, S, P, Fe or N. After the first resuspension, none of the treatments exhibited a lower growth rate than the control (resuspended in complete medium). However, when the resuspension step was repeated, all treatments led to substantially lower growth rates than the control (Table 1). Cells transferred to medium without P or N showed much higher rates of exopolysaccharide production than the control (Fig. 3a). In all treatments rates of intracellular carbohydrate accumulation were slightly negative after the first transfer.
18
N
.
Staats
et
al
.
/
J.
Exp
.
Mar
.
Biol
.
Ecol
.
249
(2000
)
13
–
27
Fig. 1. Protein concentration (a), protein content of the cells (b), exopolysaccharide (EPS) concentration (c) and intracellular carbohydrate (ICH) concentration (d) in 22 21
batch cultures of C. closterium, with addition of concentrated medium at day 7 (m), incubation at a PFD of 100mmol m s at day 7 (d) and a control (no 22 21
1
Fig. 2. Protein concentration (a), NH4 concentration (b) and protein content of the cells (c) in batch cultures of 1
20 N. Staats et al. / J. Exp. Mar. Biol. Ecol. 249 (2000) 13 –27 Table 1
21
Growth rates (d ) of C. closterium when resuspended in medium devoid of a specific nutrient. Average of two replicate cultures, standard deviation between brackets
a a
For experimental design: see Materials and Methods section.
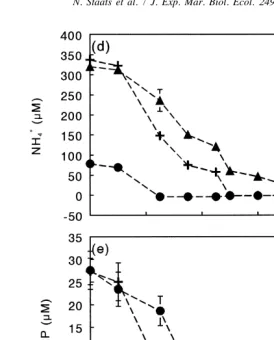
with the depletion of N or P (Fig. 4d,e). During the first 5 days, exopolysaccharide concentrations in the cultures grown at low N or P concentrations had increased markedly. This was not the case in the control culture, in which concentrations remained constant (Fig. 4b). The control culture maintained a high growth rate until day 16, and no increase in exopolysaccharides was observed during that period (Fig. 4a,b). In the ‘low N’ culture exopolysaccharide levels remained more or less constant from day 7 onwards. In the ‘low P’ culture, the concentration of exopolysaccharides decreased after 9 days. Intracellular carbohydrates showed a different pattern from exopolysaccharides (Fig. 4c). In all cultures, concentrations initially remained constant. In the ‘low N’ and ‘low P’ cultures concentrations started to increase after approximately 7 days, whereas in the control culture concentrations remained low. As was observed for exopolysac-charides, the concentration of intracellular carbohydrates in the ‘low N’ culture was higher than in the ‘low P’ culture (Fig. 4c).
1
Cultures grown with NH4 exhibited a slightly higher growth rate than cultures grown
2
with NO3 as N source (Table 2). Because growth rates were high and cells were not nutrient depleted, absolute rates of exopolysaccharide and intracellular carbohydrate accumulation were relatively low. Both on a cell and on a protein basis, the cultures
1
grown with NH4 accumulated more exopolysaccharides and intracellular carbohydrates
2 1
than the NO3 grown cultures. Because protein content was higher in NH4 grown cells, the differences in carbohydrate concentrations were smaller when expressed on a protein basis than when expressed on a cell basis. The nitrogen source did not have any effect on the ratio exopolysaccharides / intracellular carbohydrates (Table 2).
4. Discussion
In a previous study it was demonstrated that exopolysaccharides in cultures of C.
closterium are mainly produced during the transition from linear to stationary growth
exopolysac-Fig. 3. Accumulation rates of exopolysaccharides (light gray bars) and intracellular carbohydrates (dark grey bars) of C. closterium when grown in medium devoid of Si, S, P, Fe or N. After 7 days of growth in complete medium, cells were transferred to medium lacking one of the nutrients and analyzed after 4 days of incubation (a), followed by a second transfer to fresh nutrient depleted medium and analysis after 4 days (b). Note difference in scale. Average and standard deviation of duplicate cultures.
22 N. Staats et al. / J. Exp. Mar. Biol. Ecol. 249 (2000) 13 –27
Fig. 4. Cell density (a), exopolysaccharide (EPS) concentration (b), intracellular carbohydrate (ICH) 1
concentration (c), NH concentration (d) and soluble reactive phosphorous concentration (SRP) (e) in batch4
Fig. 4. (continued )
limitation were also reported for other diatom species, such as Navicula pelliculosa (Lewin, 1955), Chaetoceros affinis (Myklestad, 1977) and several other planktonic diatom species (Waite et al., 1995).
24 N. Staats et al. / J. Exp. Mar. Biol. Ecol. 249 (2000) 13 –27 Table 2
Growth rates, rates of accumulation of exopolysaccharides (EPS) and intracellular carbohydrates (ICH), 2 1a
protein concentration and the EPS / ICH ratio of C. closterium grown for 7 days with NO3 or NH4
Biomass correction: Biomass correction: EPS accumulation rate 0.14 (0.02) 0.27 (0.03) 5.1 (0.3) 6.9 (0.4) ICH accumulation rate 0.28 (0.02) 0.55 (0.09) 10.3 (1.2) 14.4 (3.0)
6 21
Protein (mg * (10 cells) ) 28.0 (3.6) 38.5 (4.0)
EPS / ICH 0.5 (0.1) 0.5 (0.1)
a
Growth rates and exopolysaccharide and intracellular carbohydrate accumulation rates were calculated over 7 days of exponential growth, protein concentration and the EPS / ICH ratio is based on concentrations analyzed on day 7. Average of three replicate cultures, standard deviation between brackets. EPS and ICH
6 21 21
accumulation rates inmg3(10 cells3day) ormg3(mg protein3day) .
size, to which C. closterium is subject during long term incubations (Staats et al., 1999a). This would also explain why no stimulation of exopolysaccharide production was observed in the control treatment. Of the nutrients studied, only low N and P concentrations caused stimulation of exopolysaccharide production without affecting growth, since S-, Si- and Fe-depleted cultures initially exhibited high growth rates but produced little exopolysaccharide. Similar results have been obtained with the green alga
Micrasterias radiata, which produced more extracellular fibrils when P concentration in
the medium was low, while growth was not limited (Strycek et al., 1992). Similarly,
Eremosphera sp. was able to grow faster at low than at high concentrations of P in the medium, while producing high concentrations of extracellular fibrils (Strycek et al., 1992).
The accumulation of exopolysaccharides, as observed upon transfer to N- or P-depleted medium, was not accompanied by production of intracellular carbohydrates. This was confirmed by the experiment where cells were inoculated with low N or P concentration media, in which intracellular carbohydrate concentrations remained constant during the first 5 days of incubation, when exopolysaccharide accumulation rates were high. Apparently, the induction of polysaccharide secretion by low ambient N or P concentrations did not apply to intracellular carbohydrate accumulation, and intracellular storage and secretion were subject to different controlling mechanisms.
It remained unclear why polysaccharide secretion was stimulated at low N or P concentrations but not at low Fe or Si concentrations. The ecological significance of the stimulation of polysaccharide secretion by low ambient N or P concentrations may be that the exopolysaccharide matrix could function as a polyelectrolyte that binds organic
¨
sediment particles (Sundby et al., 1992), thereby influencing availability of both N and P.
Exopolysaccharide secretion may also be the result of overflow metabolism, which is defined as an excess amount of carbon dioxide fixed relative to growth requirements. This is a well known phenomenon in diatoms (Myklestad et al., 1989; Waite et al., 1995). Also in C. closterium, exopolysaccharides accumulated as a result of overflow metabolism. This became apparent when cultures were compared that were grown with
1 2 1
either NH4 or NO3 as nitrogen source. With NH4 as N source, more electrons are
2
available for reduction of CO than with NO2 3 as nitrogen source, which may result in a higher carbohydrate production rate (Turpin, 1991; Smith et al., 1992). However, growth
1 2
on NH4 resulted in a higher protein content of the cells compared with NO3 grown cells. Therefore, only when expressed on a cell basis the difference in carbohydrate accumulation rates reflected differences in electron flow. The two-fold difference in carbohydrate accumulation rates between NH - and NO -grown cells indicated that4 3
1
supplying NH as nitrogen source had a considerable impact on carbohydrate production4
rate. The fact that the ratio exopolysaccharides / intracellular carbohydrates remained
1 2
unaltered with either NH4 or NO3 as nitrogen source suggested that both exopolysac-charides and intracellular carbohydrates were produced as a result of overflow metabolism. In natural populations overflow metabolism (induced by nitrogen limitation) may be a steering factor for exopolysaccharide production, as was calculated by Ruddy et al. (1998). However, the extent to which results from experiments with C. closterium can be extrapolated to the field situation remains to be investigated.
In summary, in batch cultures of C. closterium exopolysaccharide production was enhanced during stationary phase induced by N depletion. However, depletion of other nutrients such as Si or Fe did not lead to enhanced accumulation of polysaccharides. In addition, exopolysaccharide accumulation was enhanced at low, but not growth limiting, concentrations of N or P, respectively. Furthermore, accumulation of both exopolysac-charides and intracellular carbohydrates were subject to overflow metabolism.
Acknowledgements
We thank H. Peletier (RIKZ, Haren, The Netherlands) for providing a culture of C.
closterium. Part of this investigation was financially supported by BOA Research Theme
on Tidal Areas with financial aid from the Netherlands Organization for Scientific Research (NWO), and by the European Commission project INTRMUD, contract MAS3-CT95-0022. This is publication 2544 of the Centre of Estuarine and Coastal Ecology, Yerseke, The Netherlands. [RW]
References
26 N. Staats et al. / J. Exp. Mar. Biol. Ecol. 249 (2000) 13 –27
Admiraal, W., Werner, D., 1983. Utilization of limiting concentrations of ortho-phosphate and production of extracellular organic phosphates in cultures of marine diatoms. J. Plankton Res. 5, 495–513.
Admiraal, W., Peletier, H., Zomer, H., 1982. Observations and experiments on the population dynamics of epipelic diatoms from an estuarine mudflat. Estuar. Coast. Shelf Sci. 14, 471–487.
¨
Armstrong, S.M., Barlocher, F., 1989. Adsorption and release of amino acids from epilithic biofilms in streams. Freshw. Biol. 22, 153–159.
Bhosle, N.B., Sawant, S.S., Garg, A., Wagh, A.B., 1995. Isolation and partial chemical analysis of exopolysaccharides from the marine fouling diatom Navicula subinflata. Bot. Mar. 38, 103–110. Burkholder, J.M., Wetzel, R.G., Klomparens, K.L., 1990. Direct comparison of phosphate uptake by adnate and
loosely attached microalgae within an intact biofilm matrix. Appl. Environ. Microbiol. 56, 2882–2890. ´
Cadee, G.C., Hegeman, J., 1974. Primary production of the benthic microflora living on tidal flats in the Dutch Wadden Sea. Neth. J. Sea Res. 8, 260–291.
Dade, W.B., Davis, J.D., Nichols, P.D. et al., 1990. Effects of bacterial exopolymer adhesion on the entrainment of sand. Geomicrobiol. J. 8, 1–16.
Darley, W.M., 1977. Biochemical composition. In: Werner, D. (Ed.), The Biology of Diatoms, Blackwell Scientific Publications, Oxford, pp. 198–223.
Decho, A.W., 1990. Microbial exopolymer secretions in ocean environments: their role(s) in food webs and marine processes. Oceanogr. Mar. Biol. Ann. Rev. 28, 73–153.
Dubois, M., Gilles, K.A., Hamilton, J.K., Rebers, P.A., Smith, F., 1956. Colorimetric methods for determination of sugars and related substances. Anal. Chem. 28, 350–356.
Edgar, L.A., Pickett-Heaps, J.D., 1984. Diatom locomotion. In: Round, F.E., Chapman, G. (Eds.), Progress in Phycological Research, Vol. 3, Biopress, Bristol, pp. 47–88.
Flothmann, S., Werner, I., 1992. Experimental eutrophication on an intertidal sandflat: effects on micro-phytobenthos, meio- and macrofauna. In: Colombo, G., Ferrari, I., Ceccherelli, V.U., Rossi, R. (Eds.), Marine Eutrophication and Population Dynamics, Olsen and Olsen, Fredensborg, pp. 93–100.
Herbert, D., Phipps, P.J., Strange, R.E., 1971. Chemical analysis of microbial cells. In: Norris, J.R., Ribbons, D.W. (Eds.), Methods in Microbiology, Vol. 5B, Academic Press, London, pp. 209–344.
Hillebrand, H., Sommer, U., 1997. Response of epilithic microphytobenthos of the Western Baltic Sea to in situ experiments with nutrient enrichment. Mar. Ecol. Progr. Ser. 160, 35–46.
Hobson, L.A., Pariser, R.J., 1971. The effect of inorganic nitrogen on macromolecular synthesis by
Thalassiosira fluviatilis Hustedt and Cyclotella nana Hustedt grown in batch culture. J. Exp. Mar. Biol.
Ecol. 6, 71–78.
Holland, A.F., Zingmark, R.G., Dean, J.M., 1974. Quantitative evidence concerning the stabilization of sediments by marine benthic diatoms. Mar. Biol. 27, 191–196.
Kempers, A.J., Kok, C.J., 1989. Re-examination of the determination of ammonium as the indophenolblue complex using salicylate. Anal. Chim. Acta 221, 147–155.
Kester, D.R., Duedall, I.W., Connors, N., Pytkowicz, R.M., 1967. Preparation of artificial seawater. Limnol. Oceanogr. 12, 176–179.
Lewin, J., 1955. The capsule of the diatom Navicula pelliculosa. J. Gen. Microb. 13, 162–169.
Lind, J.L., Heimann, K., Miller, E.A., Van Vliet, C., Hoogenraad, N.J., Wetherbee, R., 1997. Substratum adhesion and gliding in a diatom are mediated by extracellular proteoglycans. Planta 203, 213–221. Murphy, J., Riley, J.P., 1962. A modified single solution method for the determination of phosphate in natural
waters. Anal. Chim. Acta 27, 31–36.
Myklestad, S., 1977. Production of carbohydrates by marine planktonic diatoms. II. Influence of the N / P ratio in the growth medium on the assimilation ratio, growth rate, and production of cellular and extracellular carbohydrates by Chaetoceros affinis var. Willei (Gran) Hustedt and Skeletonema costatum (Grev.) Cleve. J. Exp. Mar. Biol. Ecol. 29, 161–179.
Myklestad, S., Haug, A., 1972. Production of carbohydrates by the marine diatom Chaetoceros affinis var. Willei (Gran) Hustedt I. Effect of the concentration of nutrients in the culture medium. J. Exp. Mar. Biol. Ecol. 9, 125–136.
˚
Myklestad, S., Holm-Hansen, O., Varum, K.M., Volcani, B.E., 1989. Rate of release of extracellular amino acids and carbohydrates from the marine diatom Chaetoceros affinis. J. Plankton Res. 11, 763–773. Paterson, D.M., 1997. Biological mediation of sediment erodibility: ecology and physical dynamics. In: Burt,
Perry, M.J., 1976. Phosphate utilization by an oceanic diatom in phosphorus-limited chemostat culture and in the oligotrophic waters of the central North Pacific. Limnol. Oceanogr. 21, 88–107.
Ruddy, G., Turley, C.M., Jones, T.E.R., 1998. Ecological interaction and sediment transport on an intertidal mudflat I. Evidence for a biologically mediated sediment-water interface. In: Black, K.S., Paterson, D.M. (Eds.), Sedimentary Processes in the Intertidal, Geological Society, Special Publications, London, pp. 135–148.
Smith, G.J., Zimmerman, R.C., Alberte, R.S., 1992. Molecular and physiological responses of diatoms to variable levels of irradiance and nitrogen availability: growth of Skeletonema costatum in simulated upwelling conditions. Limnol. Oceanogr. 37, 989–1007.
Staats, N., De Winder, B., Stal, L.J., Mur, L.R., 1999a. Isolation and characterization of extracellular polysaccharides from the epipelic diatoms Cylindrotheca closterium and Navicula salinarum. Eur. J. Phycol. 34, 161–169.
Staats, N., Stal, L.J., De Winder, B., Mur, L.R., 1999b. Oxygenic photosynthesis as driving process in exopolysaccharide production of benthic diatoms. Mar. Ecol. Progr. Ser. 193, 261–269.
Strycek, T., Acreman, J., Kerry, A., Leppard, G.G., Nermut, M.V., Kushner, D.J., 1992. Extracellular fibril production by freshwater algae and cyanobacteria. Microb. Ecol. 23, 53–74.
Sullivan, M.J., Daiber, F.C., 1975. Light, nitrogen and phosphorus limitation of edaphic algae in a Delaware salt marsh. J. Exp. Mar. Biol. Ecol. 18, 79–88.
Sundby, B., Gobeil, C., Silverberg, N., Mucci, A., 1992. The phosphorus cycle in coastal marine sediments. Limnol. Oceanogr. 37, 1129–1145.
Sutherland, T.F., Grant, J., Amos, C.L., 1998. The effect of carbohydrate production by the diatom Nitzschia
curvilineata on the erodibility of sediment. Limnol. Oceanogr. 43, 65–72.
Turpin, D.H., 1991. Effects of inorganic N availability on algal photosynthesis and carbon metabolism. J. Phycol. 27, 14–20.
Van Raalte, C.D., Valiela, I., Teal, J.M., 1976. Production of epibenthic salt marsh algae: light and nutrient limitation. Limnol. Oceanogr. 21, 862–872.
Waite, A.M., Olson, R.J., Dam, H.G., Passow, U., 1995. Sugar-containing compounds on the cell surfaces of marine diatoms measured using concanavalin A and flow cytometry. J. Phycol. 31, 925–933.
Wang, Y., Lu, J., Mollet, J.-C., Gretz, M.R., Hoagland, K.D., 1997. Extracellular matrix assembly in diatoms (Bacillariophyceae). II. 2,6-Dichlorobenzonitrile inhibition of motility and stalk production in the marine diatom Achnanthes longipes. Plant Physiol. 113, 1071–1080.
Wetherbee, R., Lind, J.L., Burke, J., Quatrano, R.S., 1998. The first kiss: establishment and control of initial adhesion by raphid diatoms. J. Phycol. 34, 9–15.
Wiltshire, K.H., 1992. The influence of microphytobenthos on oxygen and nutrient fluxes between eulittoral sediments and associated water phases in the Elbe estuary. In: Colombo, G., Ferrari, I., Ceccherelli, V.U., Rossi, R. (Eds.), Marine Eutrophication and Population Dynamics, Olsen and Olsen, Fredensborg, pp. 63–70.