www.elsevier.comrlocaterapplanim
Aggression and social spacing in light horse
ž
Equus caballus mares and foals
/
J.W. Weeks
a,), S.L. Crowell-Davis
a, A.B. Caudle
a, G.L. Heusner
ba
College of Veterinary Medicine, The UniÕersity of Georgia, Athens, GA 30602, USA
b
Department of Animal and Dairy Science, The UniÕersity of Georgia, Athens, GA 30602, USA Accepted 8 December 1999
Abstract
Aggression and social spacing were studied in 14 light horse mares and their foals living at pasture. Focal samples were collected on each mare–foal dyad for 6 to 10.5 h from 2 months of foal age until weaning at approximately 4 months of age. Observations on foals continued until approximately 6 months of age for 7.5 to 10.5 h per foal. Every 2 min the identities of all individuals within 5 m were recorded. All occurrences of agonistic behavior, and the participants, were recorded during the focal samples. In addition, during feeding of supplemental grain, all occurrences of agonistic behavior by all subjects were recorded. Significant correlations were found between mare rank and the rank of foals both prior to and after weaning. Before weaning, the rank of the foal was significantly correlated with birth order. No significant correlation between birth order and foal rank was found for the post-weaning hierarchy. An animal’s gender had no significant effect on foal rank or the choice of preferred associate. Both prior to and after weaning, foals associated preferentially with the foal of their dam’s most preferred associate. In addition, significant positive correlations were found between rank of mares and foals and the rate at which they directed aggression to other herd members.q2000 Elsevier Science B.V. All rights
reserved.
Keywords: Horse; Social ontogeny; Sex differences; Dominance relationships
1. Introduction
Since the introduction of the concept of social dominance or ‘‘peck-order’’ ŽSchjelderup-Ebbe, 1922 , a great deal of debate has ensued revolving around the.
)Corresponding author.
0168-1591r00r$ - see front matterq2000 Elsevier Science B.V. All rights reserved. Ž .
Ž .
concept Bernstein, 1981; Drews, 1993; Dewsbury, 1994 . The debate involves several key issues of dominance, the first being the actual definition of dominance. Dominance
Ž . Ž
has been defined as aggressiveness Wilson, 1975 ; a trait that conveys rank
Baen-. Ž .
ninger, 1981 ; priority of access to resources Wilson, 1975 ; and peck order, in which Ž
there is a consistent unidirectional pattern of agonistic interactions Schjelderup-Ebbe, .
1922 . The concept of peck order has been melded into a modified peck order that takes Ž into account more types of agonistic interactions and is less species-specific Drews,
. 1993 .
It is usually assumed that dominant animals accrue many benefits from their status. Dominant animals usually have a priority access to preferred resources such as food and mates. However, there are now some questions as to the costs and benefits of being a high ranking animal. Recent studies have shown that dominant animals have higher
Ž .
levels of stress hormones than lower ranking animals Creel et al., 1996 . These elevated cortisol levels have been noted in female mongooses and wild dogs. The chronic elevation of stress hormones could lead to a shorter life span for high ranking animals, as well as decreased reproductive ability. Therefore, the benefits of being a dominant animal may not come without a cost. Further research will determine what these costs may be.
Even though there exists a great deal of disparity in reference to the concept of dominance, there is no debate about the existence of dominance relationships in many socially living species. The study of the many facets of dominance in numerous species has created a wealth of data on the subject.
It is well established that feral horses, when in groups of two or more, develop and Ž
maintain dominance relationships Tyler, 1972; Clutton-Brock et al., 1976; Berger, 1977; Wells and vonGoldschmidt-Rothschild, 1979; Rubenstein, 1981; Waring, 1983; Ellard and Davis, 1989; Rutberg and Greenberg, 1990; Araba and
Crowell-.
Davis, 1994 . These dominance relationships can take many forms. Dominant stallions Ž
may gain exclusive breeding capabilities over younger or subordinate stallions Feist and .
McCullough, 1976; Miller, 1981; Waring, 1983 . Particular bands of horses can be dominant over other bands. These dominant bands have priority of access to resources
Ž .
such as water, food and resting places Tyler, 1972; Feist and McCullough, 1976 .
Ž .
Within each band there exists a dominance hierarchy Waring, 1983 . Feral horses are seen most often in harem bands in which the stallion may be the dominant individual ŽFeist and McCullough, 1976; Miller, 1981 , though this is not always the case Houpt. Ž
.
and Keiper, 1982 . Among the mares of the harem, there is normally a dominance
Ž .
hierarchy Tyler, 1972; Houpt and Keiper, 1982 . This hierarchy is maintained through Ž
agonistic interactions and is relatively stable Wells and vonGoldschmidt-Rothschild, .
1979 . Most changes in the dominance hierarchy among feral populations are due to
Ž .
changes in the younger age classes Wells and vonGoldschmidt-Rothschild, 1979 . Colts, upon reaching reproductive maturity, will often leave the natal herd and either
Ž .
join a bachelor herd or remain solitary Waring, 1983 . Fillies also will disperse and take
Ž .
up membership in other harem bands Waring, 1983 .
Domestic herds under human management also form stable dominance hierarchies ŽHoupt and Wolski, 1980; Ellard and Crowell-Davis, 1989; Araba and Crowell-Davis,
.
as well, though these resources are often not limited. Dominant individuals often Ž
maintain control over supplemental feed longer than subordinate horses Houpt and .
Wolski, 1977; Ellard and Crowell-Davis, 1989 and will often displace lower ranking individuals for access to food, water, or resting sites.
There are many factors that may influence the dominance rank an individual attains within its herd. Dominance rank has been found to correlate with age in feral horses ŽTyler, 1972; Clutton-Brock et al., 1976; Wells and vonGoldschmidt-Rothschild, 1979;
. Rubenstein, 1981; Keiper and Sambraus, 1986; Rutberg and Greenberg, 1990 , in
Ž .
Przewalski horses Feh, 1988; Keiper, 1988; Keiper and Receveur, 1992 , and in horse Ž
populations managed by humans Ellard and Davis, 1989; Araba and Crowell-.
Davis, 1994 . In contrast, several studies have not found a significant correlation
Ž . Ž
between age and rank in feral Berger, 1977 and managed populations Houpt et al., .
1978; Haag et al., 1980 .
A horse’s body size is also thought to influence its dominance rank. Either height, Ž weight or both have been found to correlate significantly with dominance rank Tyler, 1972; Berger, 1977; Houpt and Wolski, 1977; Ellard and Crowell-Davis, 1989; Rutberg
. Ž
and Greenberg, 1990 . As with age, these correlations are not found universally Houpt .
and Keiper, 1982; Houpt et al., 1978; Haag et al., 1980 .
An integral part of the creation and maintenance of dominance relationships is aggression. Significant positive correlations have been found between aggression and
Ž .
dominance rank Houpt and Wolski, 1980; Houpt et al., 1978; Haag et al., 1980 . Inverse correlations have been found between aggression rates and dominance rank in
Ž .
foals Araba and Crowell-Davis, 1994 .
The rank of a foal’s dam may also influence the dominance rank a foal achieves. Two studies have reported a significant correlation between the dominance rank of mares
Ž
within a mare herd and that of the foals born into the herd Houpt and Wolski, 1980; .
Araba and Crowell-Davis, 1994 . It is not know whether this influence is genetic, environmental or a combination of both of these possibilities.
It is not surprising that there are so many contrasting reports of the correlations between individuals and their dominance status. There is no established norm for the measurement of dominance hierarchies in horses. Some hierarchies may be based solely
Ž
on the occurrence of threats Wells and vonGoldschmidt-Rothschild, 1979; Keiper and .
Receveur, 1992 , while others use more complex requirements of interactions, using Ž
submissive responses as well Ellard and Crowell-Davis, 1989; Araba and Crowell-Davis, .
1994 . Additionally, all of the studies were conducted on different types of horses ranging from feral to human managed. Many different breeds of domestic horses were used as well. As there is little to no data reporting the similarities or differences in behavior of different breeds of horses, it is assumed that their behavior is similar.
The dominance rank of an individual can influence the animals with whom it associates. A horse tends to associate with other individuals that are similar to it in age
Ž .
and rank Clutton-Brock et al., 1976; Ellard and Crowell-Davis, 1989 . One or more
Ž .
preferred associates can usually be identified for herd members Estep et al., 1993 . Preferred associates also appear to be chosen by gender. Foals will tend to preferentially
Ž
associate with other foals of the same gender Crowell-Davis et al., 1986; Araba and .
This study was designed to assess the determinants of dominance and preferred association in light breed mares and foals. Similar studies have previously been
Ž
conducted to assess the determinants of dominance in draft horses Ellard and
Crowell-. Ž .
Davis, 1989; Araba and Crowell-Davis, 1994 and ponies Asa et al., 1979 . Studies involving light breed horses have been of Thoroughbreds or groups of various breeds ŽHoupt and Wolski, 1980; Houpt et al., 1978 and were not conducted in the field.. Instead, these studies determined dominance relationships from the results of paired feeding tests. The following study used observations of spontaneous behavior in the field to assess dominance and preferred associate relationships among light breed mares and foals. The following hypotheses were tested:
I. The dominance rank of the foal in the foal herd correlates with that of its dam in the mare herd, but not with its order of birth into the herd.
Ž .
II. The dominance rank of the foal correlates with its size height or weight , but not gender.
III. Foals associate most often with the foal of their dam’s most preferred associate. IV. The foal’s most preferred associate is more likely to be of the same gender than of the opposite gender.
V. Foals are more likely to aggress toward other foals than toward their most preferred associate per unit time spent near them.
VI. The rank of the foal positively correlates with its rate of giving aggression. VII. The rank of the foal is negatively correlated with the rate at which its aggression is ignored.
VIII. Males and females differ in the type of aggression they most often display toward other foals.
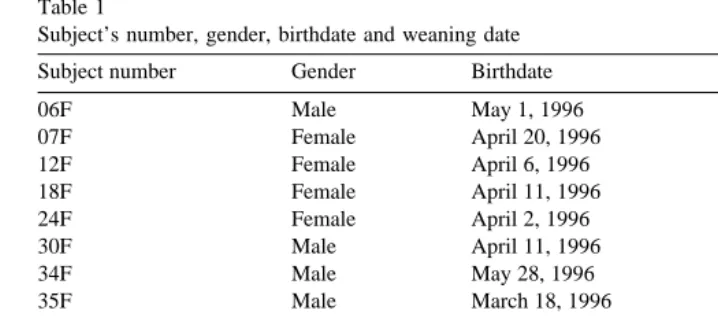
2. Subjects and study site
The subjects of this study were 14 light breed mares and their foals. The herd consisted of 10 Quarter horses, two Arabians, one Hanovarian, and one Saddlebred. Among the foals there were several fillies, four colts, two early pre-pubertal castrates ŽEPC and one pre-pubertal castrate. The early pre-pubertal castrates were gelded at. approximately 1 month of age. The pre-pubertal castrate was gelded at approximately 4 months of age. The mares existed together as a herd for almost 12 months prior to the birth of the first foal. Four to six weeks before a mare’s expected foaling date each mare was moved to an individual foaling pen. The foaling season started March 16 and lasted
Ž . until the birth of the last foal on May 28, 1996 Table 1 .
Observations were conducted at the Smithonia Breeding Center in Colbert, GA. The Ž herd was maintained on a 10-ha pasture. The pasture is composed of fescue Festuca
. Ž . Ž
sp. , common bermudagress Cynodon dactylon , coastal bermudabrass Cynodon
dacty-. Ž .
lon L. Pers. and rye grass Lolium perenne . Observations were also conducted in a
small paddock to which mares were brought daily. Mares and foals were brought to the Ž .
Table 1
Subject’s number, gender, birthdate and weaning date
Subject number Gender Birthdate Weaning date
06F Male May 1, 1996 September 27, 1996
07F Female April 20, 1996 September 20, 1996
12F Female April 6, 1996 September 11, 1996
18F Female April 11, 1996 August 24, 1996
24F Female April 2, 1996 August 26, 1996
30F Male April 11, 1996 September 11, 1996
34F Male May 28, 1996 August 26, 1996
35F Male March 18, 1996 August 19, 1996
45F Male April 6, 1996 September 27, 1996
53F Female April 30, 1996 September 11, 1996
57F Male April 22, 1996 September 27, 1996
00F Female April 27, 1996 September 27, 1996
37F Male April 22, 1996 September 20, 1996
100F Female March 16, 1996 August 2, 1996
measured. The small paddock consisted of a mostly dirt substrate. Mares and foals were brought from the large field to the small paddock approximately five times a week until early July. After this time, they were brought to the small paddock one to two times a week. Mares and foals would spend several hours in the paddock each time they were housed there.
Foals were weaned at approximately 4 months of age. Therefore, foals were not weaned at the same time, though they were sometimes weaned in groups of two or three. Throughout the weaning process, weaned foals remained in the pasture with unweaned foals and their mothers. The dams of weaned foals were moved to another pasture on the premises. The practice of weaning foals at 4 months of age is a standard practice at the Smithonia Breeding Center.
Ž
The caretakers distributed grain twice daily between 0830 and 0900 and between .
1700 and 1730 in buckets approximately 3 m apart. Throughout the study there were
Ž . Ž .
between 11 and 18 buckets from which 14 post-weaning to 28 pre-weaning animals fed. Distribution of food was not under the researcher’s control.
3. Methods
Observations were made on mares and foals from the middle of June until the last foal was weaned in September. Data were recorded for foals post-weaning until approximately 6 months of age.
3.1. Pre-weaning data collection
Ž .
Multiple 30-min random focal samples Altman, 1974 were conducted on each
Ž .
during times when supplemental feed was given. Only agonistic data were recorded during supplemental feedings. Within each 30-min sample session the following types of data were recorded: agonistic encounters, non-aggressive encounters and spatial relation-ship.
3.1.1. Agonistic encounters:
All agonistic encounters involving the focal mare and foal were recorded. For each encounter the initiator of the aggression and the recipient of the aggression were recorded. A description of the action of the aggressor and the reaction of the recipient were also recorded.
The following behavioral patterns were scored as aggressive:
1. Head threat: The extension of the aggressor’s head and neck towards another individual while laying the ears against its head.
2. Bite threat: The aggressor’s ears were laid back. Its mouth was opened and a biting motion was made while it moved its head or whole body toward another animal, but no contact was made.
3. Bite: The aggressor’s ears were laid back and its teeth were closed on some body part of another animal.
4. Kick threat: Any of the three following behavioral sequences were scored as a threat to kick:
a. The aggressor, with its ears laid back, made a rapid movement so as to place its hindquarters near another animal.
b. The aggressor, with its ears laid back, raised one hind limb and held it in the air while in a position so as to potentially strike another.
c. The aggressor, with its ears laid back, rapidly struck with one or both hind limbs towards another animal, but no contact was made.
5. Kick: With its ears laid back, one or both hindlimbs of the aggressor were projected outward rapidly and struck another animal.
6. Strike threat: The aggressor’s ears were laid back and its head and shoulders were oriented toward another individual. One or both forelimbs moved outward and forward toward the other animal but no contact was made.
7. Strike: The aggressor’s ears were laid back, and one or both forelimbs moved outward and struck the body of another animal.
8. Chase: With its ears laid back, the aggressor pursued an individual, for at least one full stride.
The following behavioral patterns were scored as being submissive responses to one of the previously mentioned aggressive behaviors:
1. Head turn: The recipient of the aggression turned its head away from the aggressor. The recipient’s limbs did not move.
3.2. Non-aggressiÕe behaÕior
The following behavioral pattern was recorded as a ‘‘win or lose’’ even though no aggression occurred:
1. Approach and retreat: This pattern was scored whenever one individual immedi-ately moved away from an animal that had just moved to within 2 m of it and had failed to exhibit an aggressive behavior.
Only interactions in which there was a clear winner and loser were used to calculate the dominance hierarchy. In these instances, the animal that approached, threatened aggression, or aggressed was scored as a winner. The animal that responded either with a head turn, avoidance, or retreat was scored as a loser. Approach and aggressive behavior that did not elicit a submissive response was not used in the calculation of the dominance hierarchy. All aggressive behavior was used to calculate rates of aggression. An animal was considered subordinate to another animal if it lost five encounters and won none. If an animal won one or more encounters with a particular foal, then, in addition to the required five losses, that foal had to lose another two encounters for each win in order to be scored as the subordinate foal. If a dyad existed that did not meet the above-mentioned criteria, the foal that lost the greater number of encounters was scored subordinate.
In order to determine the rank order among mares and foals, these dyadic relation-ships were evaluated, such that the highest ranking mare or foal submitted to the least number of mares or foals. The foal rank order was compared to the rank order of the
Ž .
dams, using Spearman’s rank order correlation coefficient rho SPSS to test for maternal influence on the rank of the foals. A correlation test was also performed on foal
Ž .
rank and foal birth order using Spearman’s rho. The wither height in inches and weight Žin grams of the foals were also subject to a Spearman’s rho correlation test. Body size.
Ž .
measurements were taken 1 week prior to weaning approximately 4 months of age and
Ž .
during the last week of the study approximately 6 months of age . A Mann–Whitney test was used to test for any differences in rank among colts and fillies.
Rates of aggression were calculated for individual mares and foals by dividing the focal animal’s total number of aggressive interactions by the total number of hours of observation in which agonistic behaviors were recorded. These aggression rates were broken down into rate of aggression given, rate of aggression received, rate of received aggression that was ignored and the rate of aggression given by the focal animal that was ignored by the recipient. All of the specific rates of aggression were calculated in the same manner as the overall rate of aggression.
Aggression was also categorized as a percentage of aggression administered up the dominance hierarchy for each individual as well as the entire herd. The rates of aggression given, aggression received, and aggression ignored were all tested for correlation with the rank of the individual using a Spearman’s rho. Mares and foals were also ranked in accordance to their respective rates of giving and receiving aggression. These ranks were tested for correlation using the Spearman rank order correlation.
ANOVA was performed to test for any effects of gender on the type of aggression most often displayed. All correlations were performed one-tailed as the direction of the correlation was predicted prior to collection of data.
3.3. Spatial relationships
The third category of data involved the focal animal’s spatial relationships with other animals. Every 2 min during each 30-min focal session the following data were recorded:
1. The identities of all animals within 1 m of the focal animal.
2. The identities of all animals greater than 1 m away but within 5 m of the focal animal.
3. The identities of all animals greater than 5 m away but within 10 m of the focal animal.
These spatial relations were used to determine each mare and foal’s most preferred associate. The most preferred associate was defined as the animal that was observed within 5 m of the focal animal the greatest percentage of 2-min samples. A t-test for paired samples was used to test for an effect of the foal’s gender on the gender of its most preferred associate. Only fillies and colts were used for this test as the small number of early pre-pubertal castrates in the study group precluded their use in these analyses.
In order to test whether or not a foal preferentially associated with the foal of its dam’s most preferred associate, the probability of the focal foal associating with the foal of the dam’s most preferred associate if association was random was determined. This was tested using a t-test for paired samples. Physical proximity rates were calculated for mares and foals by taking the total number of animals that were observed within 10 m of the focal animal and dividing by the total number of hours in which spatial data was recorded. These rates were then correlated with rank in mares and foals. A correlation test was also run between the sociability rates of mares and the sociability rates of their foals to test for any maternal influence on sociability. Aggression rates to preferred associates were calculated and a t-test for paired samples was used to test for any differences in aggression towards other foals and preferred associates.
4. Results
Mares were observed for a total of 121.75 h. Foals were observed for 121.75 h pre-weaning and 118.67 h post-weaning. The dominance hierarchies found for mares
Ž .
and foals were not perfectly linear, but did approach linearity Tables 2–4 . Foal rank in Ž .
the dominance hierarchy pre-weaning Table 5 was significantly correlated to the foal
Ž .
rank in the post-weaning dominance hierarchy rs0.66, ps0.005 . The foal rank
Ž . Ž .
Table 2
Field observations of mare dominance–subordinance interactions. Each score represents the number of win–loss interactions of that pair. Mares appear in order of their rank
Winners
45 37 12 24 06 07 100 35 18 30 00 53 57 34
45 X 0 1 0 0 0 0 20 0 1 0 20 0 0
37 48 X 1 0 0 0 0 0 0 0 0 0 0 0
12 21 29 X 1 0 0 0 0 0 0 0 0 0 0
24 19 8 37 X 1 0 0 0 0 1 0 0 0 0
06 11 10 6 11 X 0 0 4 1 4 0 0 0 0
07 9 10 11 12 23 X 1 0 6 0 0 0 0 0
100 20 8 11 9 7 8 X 6 1 0 0 0 0 0
35 0 10 6 7 1 7 15 X 0 0 0 0 0 0
18 7 3 3 10 1 0 6 15 X 5 0 0 0 0
30 11 9 9 11 6 8 16 4 6 X 3 0 0 0
00 9 6 6 2 9 18 8 8 4 17 X 1 0 0
53 2 11 6 7 8 12 9 4 8 2 9 X 22 0
57 31 11 11 1 5 11 6 5 12 11 14 1 X 1
34 3 4 5 5 3 6 3 1 4 4 4 7 3 X
Ž .
The post-weaning foal rank Table 6 was significantly correlated with mare rank Žrs0.60, ps0.011 . Birth order Table 7 was significantly correlated with foal rank. Ž .
Ž . Ž .
pre-weaning rs0.66, ps0.005 but not with the post-weaning foal rank p)0.05 . Height was significantly negatively correlated with foal rank both pre and post-weaning Žrs y0.574, ps0.033; rs y0.633, ps0.019 . Weight was not significantly.
Table 3
Field observations of pre-weaning dominance–subordinance interactions. Each score represents the number of win–loss interactions of that pair. Foals appear in order of rank
Winners
100F 45F 12F 07F 24F 30F 35F 06F 37F 00F 57F 53F 18F 34F
100F X 1 0 2 0 1 0 0 0 0 0 0 0 0
L45F 2 X 0 6 2 0 0 0 1 0 0 0 0 0
O12F 1 4 X 0 1 1 2 2 0 0 0 1 0 0
S07F 0 2 1 X 0 0 0 1 0 2 1 0 0 0
E24F 1 6 2 0 X 0 1 0 1 0 0 0 0 0
R30F 0 0 0 2 0 X 1 1 0 0 0 0 0 0
S35F 4 0 6 1 0 4 X 0 0 0 0 0 0 0
06F 1 2 2 1 0 1 0 X 2 0 0 0 0 0
37F 2 1 1 0 0 0 0 0 X 0 0 0 0 0
00F 1 2 1 0 1 1 0 1 0 X 0 0 0 0
57F 1 5 0 1 1 0 2 1 0 1 X 0 1 0
53F 1 1 2 1 1 0 0 0 0 0 3 X 2 1
18F 3 4 3 0 2 1 0 0 1 1 0 1 X 0
Table 4
Field observations of post-weaning dominance–subordinance interactions. Each score represents the number of win–loss interactions of that pair. Foals appear in order of their rank
Winners
06F 45F 30F 100F 12F 00F 37F 35F 24F 07F 18F 57F 34F 53F
06F X 1 0 0 0 1 0 2 0 0 0 0 0 0
45F 13 X 0 12 1 1 0 0 0 7 0 0 0 0
30F 12 16 X 0 0 0 1 0 0 0 0 0 0 0
100F 7 1 1 X 0 0 0 1 0 3 0 0 0 0
12F 6 11 3 1 X 2 1 0 1 0 0 0 0 1
00F 15 20 7 2 11 X 1 3 5 0 0 0 0 0
37F 6 22 1 6 2 6 X 0 1 0 0 2 0 0
35F 5 12 4 6 0 3 1 X 1 1 1 1 0 0
24F 10 15 5 2 3 4 6 7 X 1 1 2 0 0
07F 6 14 1 0 0 3 5 5 1 X 0 2 0 0
18F 6 10 5 8 11 9 7 5 15 9 X 0 2 0
57F 8 22 3 7 10 9 3 12 12 5 23 X 2 0
34F 5 13 3 2 8 3 2 6 5 3 9 3 X 0
53F 1 7 3 1 8 7 9 4 11 9 11 11 2 X
correlated with rank at any time, though the correlation did approach significance Žrs0.46, ps0.08 prior to weaning. Gender had no effect on the dominance rank. obtained by the foal either pre or post-weaning. Foal rank was not significantly correlated with weaning order.
Aggression rates were calculated for mares and foals. The rank of a mare was
Ž .
significantly correlated rs0.93, ps0.000 with the rate at which the mare gave
Ž .
aggression to other herd members Table 8 . Mare rank was also significantly,
Table 5
Pre-weaning rank order of the foals compared to the post-weaning rank order of the foals. The foal are listed in descending order of rank
Pre-weaning Post-weaning
100F 06F
45F 45F
12F 30F
07F 100F
24F 12F
30F 00F
35F 37F
06F 35F
37F 24F
00F 07F
57F 18F
53F 57F
18F 34F
Table 6
Mare rank order compared to the pre-weaning rank order and the post-weaning rank order of the foals. The mares and foals are listed in descending order of rank
a b
Mare rank Pre-weaning foal rank Post-weaning foal rank
45 100F 06F
37 45F 45F
12 12F 30F
24 07F 100F
06 24F 12F
07 30F 00F
100 35F 37F
35 06F 35F
18 37F 24F
30 00F 07F
00 57F 18F
53 53F 57F
57 18F 34F
34 34F 53F
a
Spearman’s rank correlation coefficients0.70, p-0.003.
b
Spearman’s rank correlation coefficients0.60, p-0.01.
Ž .
negatively correlated rs y0.55, ps0.02 with the rate of receiving aggression. Prior to weaning, the rate at which a foal gave aggression was significantly, positively
Ž .
correlated with rank rs0.93, ps0.000 , while the rate at which a foal received aggression was not significantly correlated with rank. The data for foals post-weaning
Table 7
The pre-weaning rank order and post-weaning rank order of foals compared to their order of birth into the
Ž .
herd. The foals are listed in descending order of birth oldest at the top and rank
c d
Pre-weaning rank order Post-weaning rank order Birth order
100F 06F 100F
45F 45F 35F
12F 30F 24F
07F 100F 12F, 45F
24F 12F 30F, 18F
30F 00F 07F
35F 37F 37F, 57F
06F 35F 00F
37F 24F 53F
00F 07F 06F
57F 18F 34F
53F 57F
18F 34F
34F 53F
c
Spearman’s rank correlation coefficients0.055, p-0.055.
d
Table 8
Mare’s listed in order of rank, aggressions received per hour, aggression given per hour and percentage of aggression given up hierarchy
e f
Mare rank Aggression received Aggression given Percentage up
45 3.29 14.98 0
37 3.84 9.33 0
12 5.51 12.22 0
24 7.14 8.11 0
06 3.69 4.86 0
07 6.13 5.96 0
100 8.48 7.76 4
35 5.94 8.65 47
18 5.41 4.54 23
30 6.84 4.00 5
00 7.49 2.55 13
53 9.30 2.70 72
57 10.67 2.22 81
34 4.84 0.09 0
e
Receives aggression rate. Spearman’s rank correlation coefficients0.55, p-0.02.
f
Gives aggression rate. Spearman’s rank correlation coefficientsy0.93, p-0.01.
Ž .
was similar to that of the mares Table 9 . The foal’s post-weaning rank was
signifi-Ž .
cantly, positively correlated rs0.67, ps0.004 with the rate at which the foal aggressed toward others. The rate at which foals received aggression post-weaning was
Ž .
also significantly, negatively correlated rs y0.67, ps0.004 with rank in the
Table 9
Foal’s post-weaning rank order, aggressions received per hour, aggression given per hour and percentage of aggression given up the hierarchy
g h
Foal rank Aggression received Aggression given Percentage up
06F 0.23 7.96 0
45F 1.96 12.00 0.7
30F 0.76 2.29 0
100F 0.46 5.54 34
12F 1.50 4.73 2
00F 3.81 3.46 6
37F 2.08 1.73 11
35F 1.02 2.33 19
24F 0.93 3.62 25
07F 1.27 3.69 41
18F 3.15 2.95 6
57F 7.15 2.54 46
34F 2.65 0.92 0
53F 4.18 0.44 0
g
Receives aggression rate. Spearman’s rank correlation coefficients0.68, p-0.004.
h
hierarchy. Mare rank for giving aggression was positively correlated with the rank of the
Ž .
foals for giving aggression both prior to weaning rs0.722, ps0.002 and after
Ž .
weaning rs0.508, ps0.032 . A foal’s rank for giving aggression pre-weaning and a Ž
foal’s rank for giving aggression post-weaning were also positively correlated rs0.790, .
ps0.000 . The mare herd administered an average of 19% of all aggression up the hierarchy. Similarly, the foal herd post-weaning administered 15% of all aggression up the hierarchy. Aggression rates per subordinate were not significantly correlated with rank in either the mares or the foals post-weaning. The mare and foal ranks for receiving aggression were never significantly correlated.
The manner in which an individual ignores aggression may also be affected by the rank of the two individuals involved. Aggression rates were calculated for the focal ignoring aggression directed at it and for aggression given by the focal animal that is ignored by the recipient. Prior to weaning, the rank of a foal approached a significant
Ž .
correlation rs0.45, ps0.054 with the rate at which its aggression was ignored by the recipient, i.e., aggression given by a higher ranking foal was less likely to be ignored than the aggression given by a lower ranking foal. This correlation was not significant
Ž .
for the foals after they were weaned p)0.05 . The rate at which an individual ignored aggression was not significantly correlated with the rank of a foal either pre- or
Ž .
post-weaning p)0.05 . There was no significant correlation between rank and either rate of ignoring aggression in the mares. Fillies and colts did not differ in the type of
Ž .
aggression exhibited p)0.05 .
Ž . Ž
Sociability rates Table 10 of mares and foals were significantly correlated rs0.67, .
ps0.005 prior to weaning, but not after weaning. Rank of an individual did not influence its sociability rate in any group.
Gender was found to have no effect on a foal’s choice of most preferred associate. This lack of effect was evident in foals prior to as well as after the foals were weaned. The foals had a preferential association with the foal of their dam’s most preferred
Table 10
Sociability rates of mares, pre-weaning foals and post-weaning foals
i
Subject ID number Mares Pre-weaning Post-weaning
06 26.70 72.00 96.38
Ž . associate. This association was significant prior to weaning ts y3.69, ps0.003 and
Ž .
approached significance for the foals post-weaning ts y2.10, ps0.056 . Foals were not more likely to aggress to preferred associates than to the other members of the herd Žp)0.05 ..
Mares and foals were both observed to share supplemental feed when it was offered. Mares were observed to share a feed bucket with their own foals, other mares, and foals other than their own. Foals were also observed sharing feed buckets both prior to and after weaning. Foals were observed to share a feed bucket with up to three other individuals.
5. Discussion
Dominance rankings found for both the mare and foal herds were not perfectly linear, but did approach linearity. The finding that the dominance ranking of the foals was significantly correlated with that of the mares is in accordance with the findings of
Ž .
Araba and Crowell-Davis 1994 for draft horses. This influence appears in light as well as heavy breeds of horses. The fact that the foal’s rank was significantly correlated with its dam’s rank both prior to and after weaning implies that the influence of the dam is relatively constant over time.
Several other mammalian species exhibit a similar matrilineal dominance system in which the offspring come to rank immediately below their mothers. This phenomenon is most often seen with female offspring. This type of hierarchical system has been
Ž
reported in several species of macaques Kawai, 1958; Missakian, 1972; Sade, 1972; DeWaal, 1977; Silk et al., 1981; Netto and Van Hoof, 1986; Paul and Kuester, 1987;
. Ž . Ž
Chapais, 1988 , Vervet monkeys Horrocks and Hunte, 1983 , and baboons Hausfater,
. Ž .
1975; Hausfater et al., 1982; Johnson, 1987 . Jenks et al. 1995 also reported maternal rank to be influential in spotted hyenas.
How the dam influences the foal’s rank is still unclear at this time. This influence could stem from some undetermined genetic factor, as dominance itself cannot be inherited since it is a learned relationship between two individuals. However, there may be one or multiple factors that influence an animal’s ability to be dominant that is heritable. The finding that the aggressive rankings of mares and foals are significantly correlated lends support to the idea that there may be heritable behavioral predisposi-tions in these animals. This heritability of traits would most likely be a result of maternal influence as opposed to paternal influence as the majority of the foals in this study were sired by the same stallion and acquired varying roles in the dominance hierarchy.
Active maternal participation in offspring social encounters has not yet been studied. Until that time, the degree of behavioral influence exerted by a mare will not be known. Lastly, the maternal influence could be a combination of both genetic and environ-mental factors. This will not be clarified until researchers employ embryo transfer or cross-fostering of foals as a means of potentially manipulating dominance. Additionally, studies involving orphaned or rejected foals would be of benefit.
Birth order was significantly correlated with foal dominance rank prior to weaning. This effect was not seen after the foals were weaned. This finding is in accord with the
Ž
findings of other researchers using the same methodology in a draft breed Araba and .
Crowell-Davis, 1994 . The lack of correlation between age and rank in foals post-wean-Ž
ing is contrary to many published findings Tyler, 1972; Clutton-Brock et al., 1976; Wells and vonGoldschmidt-Rothschild, 1979; Rubenstein, 1981; Rutberg and
Green-. berg, 1990 .
The lack of significant correlation between age and rank after weaning can be the result of several factors. There was a 3-month age span among the foals, the first being born in the beginning of March and the last foal being born in the middle of May. Therefore, prior to weaning, the foal members of the herd varied in size and motor development. This developmental differential would make some individuals in the herd inexperienced with the social workings of the herd as well as less able to perform all aggressive acts. After the foals were weaned, the size and developmental differences were diminished. Therefore, these factors would not affect the creation and maintenance of dominance relationships as much after the foals were weaned. Age appears to have more of an influence on dominance in feral populations than in human managed
Ž
populations Tyler, 1972; Clutton-Brock et al., 1976; Wells and vonGoldschmidt-Rothschild, 1979; Rubenstein, 1981; Rutberg and Greenberg, 1990; Araba and
Crowell-. Davis, 1994 .
Ž .
Similar to previously published results Araba and Crowell-Davis, 1994 , a foal’s dominance rank was not affected by its gender. Sexual dimorphism is minimal in this species, and females have the same capacity for giving aggression as do males. Given this reasoning, a gender effect on dominance would not be expected. The negative correlation between size and foal rank only adds to the confusion about the influence of size on dominance in horses. Many studies have found significant correlations between
Ž . Ž
some component of size height andror weight and rank Tyler, 1972; Houpt and
. Ž
Wolski, 1980; Ellard and Crowell-Davis, 1989 while others have not Haag et al., 1980; .
Houpt and Keiper, 1982 . Intuitively, size should not be a major factor in a herd of similar breed horses as they all share similar body characteristics and are not very sexually dimorphic. However, there still exist reports of significant correlations between body size and dominance rank. These discrepancies could be attributed to differences in methodology or body size measurements, e.g., weighed on a scale versus subjective opinions of the researchers.
hierarchy. These findings are in accordance with those of other researchers using Ž
different breeds of horses Houpt and Wolski, 1980; Houpt et al., 1978; Haag et al., .
1980; Araba and Crowell-Davis, 1994 . This result did not occur in the foals prior to weaning. The overall rates of aggression for foals prior to weaning were low. Thus, any rank influences on rates of aggression may not have been easily detected. The rates of aggression for the foals after weaning were much higher, making rank influences on aggression easier to detect in the latter sample.
The fact that, at times, there were fewer feed buckets available than there were horses in the field may have affected aggression rates. The competition for access to feed may have increased the aggression levels in this study as compared to levels of aggression found in previous studies. However, mares and foals were observed to share feed buckets both when there were fewer buckets than horses and when an equal number of buckets were present.
Ž . The percentage of aggression given up the hierarchy in the mare herd 19% is
Ž .
slightly higher than that reported by Clutton-Brock et al. 1976 and Wells and Ž .
vonGoldschmidt-Rothschild 1979 , both of whom reported percentages of aggression given up the hierarchy of less than 5%. The percentage of aggression given up the
Ž . Ž .
hierarchy by the foals post-weaning 15% is within the range of that reported 10–20% Ž .
by Wells and vonGoldschmidt-Rothschild 1979 . However, the differences between the present study herd and those previously reported may be accounted for by the fact that
Ž . Ž .
both Clutton-Brock et al. 1976 and Wells and vonGoldschmidt-Rothschild 1979 only scored threats in the creation of their hierarchy and analyses of aggression. This study utilized contact aggression as well as threats. In addition, unlike the studies of
Ž . Ž .
Clutton-Brock et al. 1976 and Wells and vonGoldschmidt-Rothschild 1979 , this study herd was not free-ranging. It is possible that the restrictions of human management are conducive to higher rates of aggression.
The findings that gender had no influence on the type of aggression exhibited by an individual and that foals did not aggress differentially to preferred associates ran contrary to the predictions. Most aggressive encounters were observed at supplemental feedings or near other resources. The fact that most aggression occurred near resources might have influenced the type of aggression exhibited. The most common form of aggression observed was the head threat. This could also be the easiest or least taxing form of aggression available to the horses, especially in a feeding situation. In the feeding situation, engaging in other forms of aggression might take the individual from the feed container, thus increasing its chances of losing access to that feeding area. For this reason, gender differences in aggression may not have been observed.
Physical proximity rates for mares and foals were significantly correlated prior to weaning. This result is not surprising as mares and foals will tend to spend the majority
Ž .
of time near each other. Crowell-Davis 1986 reported that Welsh pony mares and foals spent at least 50% of the time observed within 5 m of each other through week 24. As the foals were often near their dams, they would have had a similar number of other herd members within the same proximity of their dams. This would result in the mares and their foals having similar physical proximity rates.
lessen the stress of isolation from the dam, the foal seeks to remain in close association with its herd mates. This study herd was not weaned simultaneously. Foals were weaned at varying ages with anywhere from one to three foals weaned at the same time. Foals which were weaned earlier were observed spending time near the dams of other foals. This behavior would also create a higher proximity rate for foals after they were weaned.
The choice of preferred associates for the foals was not affected by gender. This is Ž
contrary to results previously reported Davis et al., 1986; Araba and Crowell-.
Davis, 1994 . The foals did, however, preferentially associate with the foal of their dam’s most preferred associate both before and after weaning. These results, along with
Ž .
those of Araba and Crowell-Davis 1994 support the notion that not only does the dam influence the dominance rank of their foal, but they also influence the foal’s choice of preferred associates. Finding this result after weaning indicates that this is a relatively stable influence as well.
The majority of the results found in this study are in concordance with those of Araba Ž .
and Crowell-Davis’ 1994 study of draft mares and foals. This suggests that maternal influence on rank and choice of preferred associates apply to horses in general and not to just a particular breed. Additionally, this study shows that even under conditions of human management, the social structure of mares and their foals comes close to that of their feral and wild counterparts. Most deviations from the social structure of feral horses involve the artificial state of breeding and weaning that human-managed horses are exposed to. The importance of dominance relationships may be magnified in managed herds. As there is not often a scarcity of resources in feral situations, high ranking horses may not incur many advantages from their social status. However, in managed herds that are recipients of supplemental feed, a high position in the herd hierarchy will allow for more time at this preferred resource. Low ranking animals may be completely denied access to supplemental feed.
Foals, both managed or feral, of high ranking mares can gain various advantages over their herd mates. High ranking mares are assumed to be in better body condition due to priority of access to feed, as well as having a decreased chance of bodily injury resulting from the receipt of aggression. An optimal, or close to optimal, body condition may increase lactation in these mares or the quality of milk produced. Additionally, foals of
Ž higher ranking mares can be protected from aggression from other herd members Tyler,
.
1972 . The advantages gained by having a good maternal body condition can translate into an increased survival rate for a foal or a high ranking mare. In managed populations, foals of low ranking mares may not be receiving enough supplemental feed, thus not reaching their nutritional needs for optimal growth rate or development. Herd managers should be aware of the social organization of their herd. Low ranking mares and foals potentially need to be removed from the herd in order to satisfy their nutritional needs.
male offspring of lower ranking mares. If foals of high ranking mares do have a higher degree of reproductive success, this increases the inclusive fitness of the mare. Hence, there are obvious advantages to being a dominant individual in a horse herd.
References
Altman, J., 1974. Observational study of behavior: sampling methods. Behaviour 49, 227–267.
Ž .
Araba, B.D., Crowell-Davis, S.L., 1994. Dominance relationships and aggression of foals Equus caballus . Applied Animal Behaviour Science 41, 1–25.
Ž
Asa, C.S., Goldfoot, D.A., Ginther, O.J., 1979. Socio-sexual behavior and ovulatory cycles of ponies Equus
.
caballus observed in harem groups. Hormones and Behaviour 13, 49–65.
Baenninger, R., 1981. Dominance: on distinguishing the baby from the bathwater. Brain and Behavioral Sciences 4, 431–432.
Berger, J., 1977. Organizational systems and dominance in feral horses in the Grand Canyon. Behavioral Ecology and Sociology 2, 131–146.
Bernstein, I.S., 1981. Dominance: the baby and the bathwater. The Behavioral and Brain Sciences 4, 419–457. Chapais, B., 1988. Experimental matrilineal inheritance of rank in female Japanese macaques. Animal
Behaviour 36, 1025–1037.
Clutton-Brock, T.H., Greenwood, P.J., Powell, R.P., 1976. Ranks and relationships in highland ponies and highland cows. Z. Tierpsychol. 41, 202–216.
Creel, S., Creel, N.M., Monfort, S.L., 1996. Social stress and dominance. Nature 379, 212.
Ž .
Crowell-Davis, S.L., 1986. Spatial relations between mares and foals of the Welsh pony Equus caballus . Animal Behaviour 34, 1007–1015.
Crowell-Davis, S.L., Hoput, K.A., Carini, C.M., 1986. Mutual grooming and nearest-neighbor relationships among foals of Equus caballus. Applied Animal Behaviour Science 15, 113–123.
DeWaal, F.B.M., 1977. The organization of agonistic relations within two captive groups of Java monkeys
ŽMacaca fascicularis . Z. Tierpsychol. 44, 225–282..
Dewsbury, D., 1994. A final word on the inheritance of dominance. Animal Behaviour 48, 984–985.
Ž .
Drews, C., 1993. The concept and definition of dominance in animal behavior. Behaviour 125 3–4 , 283–313.
Ellard, M.E., Crowell-Davis, S.L., 1989. Evaluating equine dominance in draft mares. Applied Animal Behaviour Science 24, 55–75.
Estep, D.Q., Crowell-Davis, S.L., Earl-Costello, S.A., Beatey, 1993. Changes in the social behavior of
Ž .
drafthorse Equus caballus mares coincident with foaling. Applied Animal Behaviour Science 35, 199–213.
Feh, C., 1988. Social behaviour and relationships of Przewalski horses in Dutch semi-reserves. Applied Animal Behaviour Science 21, 71–87.
Feist, J., McCullough, D., 1976. Behavior patterns and communication in feral horses. Z. Tierpsychol. 41, 337–371.
Haag, E.L., Rudman, R., Houpt, K.A., 1980. Avoidance, maze learning and social dominance in ponies. Journal of Animal Science 50, 329–335.
Ž .
Hausfater, G., 1975. Dominance and reproduction in baboons Papio cynocephalus : a quantitative analysis. In: Contributions in Primatology 7 Karger, Basel.
Hausfater, G., Altmann, J.A., Altmann, S., 1982. Long term consequences of dominance relations among
Ž .
female baboons Papio cynocephalus . Science 217, 752–755.
Horrocks, J., Hunte, W., 1983. Maternal rank and offspring rank in Vervet monkeys: an appraisal of the mechanisms of rank acquisition. Animal Behaviour 31, 772–782.
Houpt, K.A., Keiper, R., 1982. The position of the stallion in the equine dominance hierarchy of feral and domestic ponies. Journal of Animal Science 54, 945–950.
Houpt, K.A., Wolski, T.R., 1980. Stability of equine dominance hierarchies and the prevention of dominance related aggression. Equine Veterinary Journal 12, 18–24.
Jenks, S., Weldele, M., Frank, L., Glickman, S., 1995. Acquisition of matrilineal rank in captive spotted hyenas: emergence of a natural social system in peer-reared animals and their offspring. Animal Behaviour 50, 893–904.
Johnson, J., 1987. Dominance rank in juvenile olive baboons, Papio anubis: the influence of gender, size, maternal rank and orphaning. Animal Behaviour 35, 1694–1708.
Kawai, M., 1958. On the system of social ranks in a natural troop of Japanese monkeys: I and II. Primates 1, 111–148.
Ž .
Keiper, R., 1988. Social interactions of the Przewalski horse Equus przewalskii Poliakov, 1881 herd at the Munich Zoo. Applied Animal Behaviour Science 21, 89–97.
Keiper, R.R., Receveur, H., 1992. Social interactions of free-ranging Przewalski horses in semi-reserves in The Netherlands. Applied Animal Behaviour Science 33, 303–318.
Keiper, R.R., Sambraus, H.H., 1986. The stability of equine dominance hierarchies and the effects of kinship, proximity and foaling status on hierarchy rank. Applied Animal Behaviour Science 16, 121–130. Miller, R., 1981. Dominance and breeding behavior in red desert feral horses. Z. Tierpsychol. 57, 340–351. Missakian, E.A., 1972. Genealogical and cross-genealogical dominance relationships in a group of
free-rang-Ž .
ing monkeys Macaca mulatta on Cayo Santiago. Primates 13, 169–180.
Netto, W.J., Van Hoof, J.A.R.A.M., 1986. Conflict interference and the development of dominance
relation-Ž .
ships in immature Macaca fascicularis. In: Else, J.G., Lee, P.C. Eds. , Primate Ontogency, Cognition and Social Behaviour. Cambridge Univ. Press, Cambridge, pp. 291–300.
Ž
Paul, A., Kuester, J., 1987. Dominance, kinship and reproductive value in female Barbary macaques Macaca
.
sylÕanus at Affenberg Salem. Behavioural Ecology and Sociobiology 21, 323–331.
Rubenstein, D.I., 1981. Behavioral ecology of island feral horses. Equine Veterinary Journal 13, 27–34. Rutberg, A.T., Greenberg, S.A., 1990. Dominance, aggression frequencies and modes of aggressive
competi-tion in feral pony mares. Animal Behaviour 40, 322–331.
Ž .
Sade, D.S., 1972. A longitudinal study of social behavior of rhesus monkeys. In: Tuttle, R. Ed. , The Functional and Evolutionary Biology of Primates. Aldine, Chicago, pp. 378–398.
Schjelderup-Ebbe, T., 1922. Beitrage zur sozialpsychologie des haushuhns. Z. Psychol. 88, 225–252. Silk, J.B., Samuels, A., Rodman, P., 1981. Hierarchical organization of female Macaca radiata in captivity.
Primates 22, 84–95.
Tyler, S.J., 1972. The behaviour and social organization of the New Forest ponies. Animal Behavior Monograph 5, 85–196.
Waring, G.H., 1983. The Behavioral Traits and Adaptations of Domestic and Wild Horses, Including Ponies. Noyes Publications, Park Ridge, NJ.
Wells, S.M., vonGoldschmidt-Rothschild, 1979. Social behavior and relationships in a herd of Camargue horses. Z. Tierpsychol. 49, 363–380.