www.elsevier.com/locate/ibmb
Inducible P450s of the CYP9 family from larval Manduca sexta
midgut
Jennitte L. Stevens
1, Mark J. Snyder
2, Josette F. Koener, Rene´ Feyereisen
* Department of Entomology, Forbes 410, PO Box 210036, The University of Arizona, Tucson, AZ 85721, USAReceived 20 August 1999; received in revised form 18 January 2000; accepted 20 January 2000
Abstract
Several related cytochrome P450 cDNAs belonging to the CYP9 family have been cloned from the midgut of larval tobacco hornworms, Manduca sexta. The first P450, CYP9A2, was obtained by RT-PCR using degenerate primers. Northern blot analysis of expression in the midgut using the CYP9A2 probe revealed a significant induction by a variety of chemicals. Diets supplemented with the wild tomato compound 2-undecanone caused a dose-dependent induction which peaked after 48 h. Induction was also observed after addition to the diet of indole-3-carbinol, phenobarbital, 2-tridecanone and xanthotoxin. Neitherα-pinene, clofibrate nor nicotine were effective inducers. The CYP9A2 probe hybridized to two mRNA species, one of 2.0 kb and another of 4.2 kb, suggesting cross-hybridization to other P450 mRNAs. Additional P450 clones of the CYP9 family were then obtained and sequenced. Northern hybridization revealed that the 4.2 kb band also hybridized to CYP9A4 whereas the 2.0 kb hybridized to CYP9A5. Despite being 91% identical, CYP9A4 and CYP9A5 were induced differentially by clofibrate and xanthotoxin. Multiple P450 genes from various families are therefore induced in Lepidoptera in response to plant allelochemicals or xenobiotics. 2000 Elsevier Science Ltd. All rights reserved.
Keywords: Cytochrome P450; 2-Undecanone; Xanthotoxin; Clofibrate; Xenobiotics; Manduca sexta
1. Introduction
Cytochrome P450 monooxygenases are a diverse and widely distributed protein superfamily resulting from numerous gene duplication events from a common ancestor to all present day P450 forms that existed prior to the evolution of eukaryotes (Nelson and Strobel, 1987). Many functions are attributed to cytochrome P450s including the metabolism of a wide variety of both endogenous substrates and xenobiotics. In insects, cytochrome P450s metabolize hormones and phero-mones but are best known for their roles in the metab-olism of insecticides and host plant chemicals (review in Feyereisen, 1999).
* Corresponding author. Tel.:+1-520-621-9598; fax:+ 1-520-621-1150.
E-mail address: [email protected] (R. Feyereisen). 1 Present address: Molecular Biology Institute, University of
Cali-fornia at Los Angeles, Los Angeles, CA 90095-1570, USA.
2 Present address: Bodega Marine Laboratory, University of
Cali-fornia, Davis, PO Box 247, Bodega Bay, CA 94923, USA.
0965-1748/00/$ - see front matter2000 Elsevier Science Ltd. All rights reserved. PII: S 0 9 6 5 - 1 7 4 8 ( 0 0 ) 0 0 0 2 4 - 2
armigera (Ranasinghe et al., 1997) and in H. zea (Li et al., 2000) and of the CYP28 family in Drosophila spec-ies (Danielson et al., 1997) have all been reported to be inducible by host plant chemicals, and in some cases xenobiotics as well. Little is known of the function of CYP9 genes in insects, for which several representatives have been uncovered by the RT-PCR methodology (Dunkov et al., 1996) and more recently by the genome projects of Drosophila melanogaster and Bombyx mori. The first member of this family, CYP9A1 from Heliothis virescens, is constitutively over-expressed in resistant, thiodicarb-selected tobacco budworms (Rose et al., 1997).
In this paper, we report the partial sequence of several new P450s of the CYP9 family isolated from the larval midgut of a herbivore specialist on Solanaceae, the tob-acco hornworm Manduca sexta. We show that these P450s are differentially induced by a variety of plant chemicals and other xenobiotics and that this induction is both time- and concentration-dependent. These newly identified midgut P450s appear to be important compo-nents in the adaptation to dietary chemicals in Mand-uca sexta.
2. Materials and methods
2.1. Insects
Manduca sexta larvae were maintained as previously described (Snyder et al., 1993). Test chemicals were mixed into melted artificial diet (Prasad et al., 1986) and offered to fifth instar larvae within 2 h after ecdysis.
2.2. Primer construction and RT-PCR
The degenerate forward primer DF was designed TACGA(AG)(CT)IGCI(AG)(AT)IAA(CT)CC(CT)GA around the I helix region of the CYP6 insect family (YELAK(V)NPD), residues 317–324 of CYP6B1 Cohen et al., 1992). The degenerate reverse primer DR was designed CCIA(GT)(AG)CA(AG)TTIC(GT)IGGICC around the heme binding region (GPRNC(IL)G, residues 439–445 of the Papilio polyxenes CYP6B1; Cohen et al., 1992). Reverse transcription of midgut mRNA was done using Superscript enzyme at 42°C according to the manufactures’ directions (GIBCO BRL). The PCR cycles for the midgut reverse-transcribed cDNA were 94°C, 1 min; 54°C, 1 min; 72°C, 1 min, for 35 cycles and 2.5 mM Mg2+in the PCR buffer. The PCR products
were gel-purified and cloned into pCR II (Invitrogen). Sequencing of PCR clones was done by the chain ter-mination method of Sanger et al. (1977). The new P450 (CYP9A2) was identified in 2/9 clones following sequence analysis using the NCBI Blast Network Server. Primers were then designed at the 59 and 39 termini of
the CYP9A2 sequence (primer 1, 59 CCAT-GATCTCAGCGTACAGT 39, primer 2, 59 GCACAA-CATCAAACCGATGAC 39). PCR reactions were set up using a 0.15% tridecanone M. sexta midgut cDNA plas-mid library as a template (constructed as described in Snyder et al., 1995), a CYP9A2 primer and a general Sp6 primer, with reaction conditions as previously noted. Products were gel purified and cloned as described above, sequenced and analyzed for CYP9 homology with the NCBI Blast Network Server. 3/10 clones showed a high degree of similarity to CYP9A2. New primers were made based upon these PCR clones (primer 3: 59 CGACCTCCACATCACTCTAAT 39 and primer 4: 59 TACCTTTACATCACTCTTAC 39), which were used for further PCR-based screening, and two products were again sequenced and analyzed in the same way. Two additional P450 sequences were found (CYP9A4 and CYP9A5). Sequences were analyzed by the DNAs-tar suite of programs. Sequence pair distances (Table 1) were obtained by the Clustal method with PAM250 resi-due weight, ktuple 1, gap penalty 3 and window 5 set-tings.
2.3. RNA isolation and Northern hybridization analysis
Chemicals were mixed into melted artificial diet and offered to fifth instar larvae within 2 h after ecdysis. Midguts were dissected free of peritrophic membranes prior to RNA isolation. Total midgut RNA was isolated by the Cesium chloride gradient method (Sambrook et al., 1989). Poly(A)+ RNA was purified by oligo(dT)-cellulose chromatography (Sambrook et al., 1989). For the Northern analysis, 2.5µg of mRNA were loaded per lane and blotted onto Gene Screen nylon membrane (Du Pont NEN). The blots were hybridized at 42°C to the random-primed 32P-labeled M. sexta CYP9A2 and P450
PCR products, respectively, in 50% formamide, 5×SSPE pH 7.4, 5×Denhardt’s solution, and 1% SDS. Final washing was at 65°C for 15 min in 2×SSPE and 2% SDS. The blots were stripped of bound P450 PCR probe and reprobed with actin. The staining intensities of
Table 1
Percent identities of the translated M. sexta midgut CYP9A cDNAs and the H. virescens CYP9A1 sequence (Rose et al., 1997)a
1 2 3 4 5 6 7
exposed films from each blot were determined on a LKB Ultroscan Laser Densitometer and the resulting values were corrected for equal loading by comparison to actin levels.
2.4. Ribonuclease protection assay
A 322-bp ScaI-ScaI fragment of CYP9A5 that over-lapped the translation stop site was subcloned into pBluescript (Stratagene). After linearization of the plas-mid with BamHI, the probe was synthesized with T7 RNA polymerase and 32P-CTP. For CYP9A4, a 425-bp
HindIII/BamHI fragment was also cloned into
pBluescript (Stratagene) and linearized with BamHI. The riboprobe was made with T7 RNA polymerase and 32
P-CTP. RNase protection experiments were performed as described in Ausubel et al. (1994) with slight modifi-cations. Five micrograms of mRNA from control, clo-fibrate-induced, 2-undecanone induced and xanthotoxin-induced M. sexta larvae were used. Protected fragments were separated on a 6% acrylamide/urea gel along with a sequencing reaction used as a standard.
2.5. Genomic library screening
A genomic library of M. sexta was constructed in the lambda DASH vector (Stratagene) from DNA extracted from larval midgut. This library was probed with the CYP9A4 clone using the Genius kit (Invitrogen) as a means of detection of positive plaques. Ten plaques were selected for phage purification and screening.
3. Results
3.1. CYP9A2 cloning and characterization
A combination of degenerate primers for two con-served regions of insect P450 proteins was used to ampl-ify reverse-transcribed midgut mRNA of M. sexta. The PCR products were cloned and sequenced. This resulted in a clone that was 401 bp in length and encoded a read-ing frame of 120 amino acids (Fig. 1) that scored highly with other P450 proteins by BLAST Network searches. Close alignment with known insect CYP4 and CYP6 sequences, however, showed that this new P450 did not belong to either one of these families. This sequence (accession number AF172278) was most closely related to CYP9A1 from Heliothis virescens (50% identity, see Table 1) and was called CYP9A2 by the P450 nomencla-ture committee (D.R. Nelson).
3.2. Induction studies
The insert from the CYP9A2 clone was used as probe in Northern analyses of P450 expression. A typical
Northern analysis of CYP9A2 expression following diet-ary exposure to 0.5% 2-undecanone is shown in Fig. 2. Two mRNA bands of 2.0 and 4.2 kb were found at all time-points after time 0. The 4.2 kb band is larger than those of the few insect P450 mRNAs studied to date. Since CYP9A2 hybridizes to multiple CYP9 mRNAs (as discussed below), the mRNA levels that were measured by this method represent more that one CYP9 P450, and not only CYP9A2. There was no change in mRNA levels in the first hours of 2-undecanone exposure. A signifi-cant induction was then observed in two phases, first a rapid increase between 2 and 4 h, then a more gradual, further increase from 18 to 36 h, which peaked at 48 h and then declined steadily as larvae reached the wander-ing stage (Fig. 3).
The effect of increasing concentrations of 2-unde-canone in the diet is shown in Fig. 4. A significant induc-tion was found from 0.1–0.2% which progressively increased at 0.5% and 1.0% 2-undecanone. Higher levels of 2-undecanone were toxic to the larvae and their effect on mRNA could not be studied. The effect of different types of chemicals added to the diet is given in Fig. 5. Of the allelochemicals found naturally in the diet of M. sexta, both aliphatic ketones, 2-undecanone and 2-tride-canone were shown to increase mRNA levels, whereas nicotine did not. Significant induction of mRNA levels was found by feeding diets containing the plant chemi-cals indole-3-carbinol and xanthotoxin, but notα-pinene (0.1–0.5%). A low concentration of clofibrate (0.1%) was without effect, although the expression levels at 0.5% were consistently, but not significantly, higher than control levels. However, phenobarbital caused a signifi-cant induction of mRNA levels. The two mRNA bands of 2.0 kb and 4.2 kb were found in all analyses with the CYP9A2 probe, except in the hybridizations with control and clofibrate-induced mRNA. In the latter, the 2.0 kb band is not present (see below). In the time-course and dose–response experiments, the intensity of the two bands relative to each other did not change significantly and the relative mRNA levels reflect the sum of the intensity of both bands. The number and identity of the transcripts hybridizing to the CYP9A2 probe is unknown.
3.3. CYP9A4 and CYP9A5 cloning
Fig. 2. Northern blots of midgut mRNA from 5th instar larvae of M. sexta hybridized with the CYP9A2 probe (top) and actin probe (bottom). Larvae were offered an artificial diet containing 0.5% 2-undecanone within 2 h of ecdysis to the fifth instar and midguts were collected at the indicated times for mRNA isolation.
Fig. 3. Time-course of midgut mRNA induction in 5th instar larvae of M. sexta fed 0.5% 2-undecanone measured by Northern blot hybridization with the CYP9A2 probe. Each point represents the mean±1S.D. of 3–4 determinations relative to the time 0 value (just prior to feeding the diet). As discussed in the text, the CYP9A2 probe crosshybridizes with other CYP9 transcripts and the results are indica-tive of induction but do not reveal the number or identity of such tran-scripts.
Fig. 4. Relative mRNA levels in the midgut of 5th instar larvae of
M. sexta, measured by Northern blot hybridization with the CYP9A2
Fig. 5. Relative mRNA levels in the midgut of 5th instar larvae of
M. sexta, measured by Northern blot hybridization with the CYP9A2
probe in response to the addition of various chemicals to the diet. Bars represent the mean±1S.D. of three determinations relative to the trol diet value. Asterisks indicate significant differences from the con-trol diet value. The chemicals were mixed into artificial diet and offered to fifth instar larvae within 2 h after ecdysis and the midguts were dissected 72 h later. As discussed in the text, the CYP9A2 probe cross-hybridizes with other CYP9 transcripts and the results are indica-tive of induction but do not reveal the number or identity of such tran-scripts.
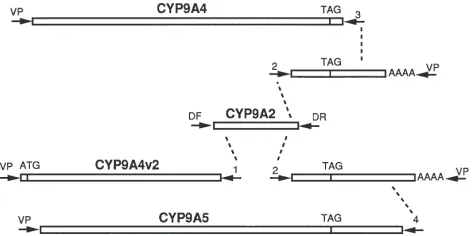
Fig. 6. Cloning strategy for the CYP9A obtained in this study. CYP9A2 was obtained by RT-PCR from midgut mRNA with forward and reverse degenerate primers (primers marked as arrows, DF, DR). The sequence of CYP9A2 directed the synthesis of primers (1 and 2) which were used in PCR reactions with vector primers (VP) on DNA from a midgut cDNA library. The sequence of the partial 39fragments of CYP9A cDNAs directed the synthesis of primers (3 and 4) which were again used in PCR reactions with vector primers (VP) on DNA from a midgut cDNA library, yielding CYPP9A4 and CYP9A5. ATG indicates the start codon, TAG the stop codon of each P450.
CYP9A4v2). Primer 2 gave two different products of about 500 bp that appeared to be the far 39ends of CYP9 P450s. These fragments are too short to merit a specific name, but they appear to be allelic variants of CYP9A4 and CYP9A5, respectively.
Two additional primers were designed from the 39
untranslated regions of these P450 fragments (primers 3 and 4). The reaction with primer 3 gave a 1,611 kb pro-duct that is a nearly full length P450 (missing the
N-terminal first 8 amino acids) named CYP9A4. PCR with primer 4 produced a 1,828 kb product that is also a near full length P450 (missing 14–15 N-terminal amino acids) named CYP9A5. CYP9A4 and CYP9A4v2 differ by 4 nucleotides which cause 3 amino acid substitutions. It is likely that these represent allelic variants of the same gene. CYP9A4 and CYP9A5 differ by 47 amino acids. When CYP9A4 and CYP9A5 are aligned (Fig. 1), CYP9A5 is characterized by an IFGRIFGRI module that causes an insertion of IFGRI respective to the CYP9A1 and CYP9A4 sequences. The nucleotide sequence indi-cates that this insertion is neither a sequencing mistake nor a recent duplication. The M. sexta CYP9 are 46–50% identical to the H. virescens CYP9A1 sequence (Rose et al., 1997). CYP9A4 and CYP9A5 are 80–83% identical to CYP9A2 (Table 1).
3.4. Northern analysis and ribonuclease protection assay
Northern blots of M. sexta midgut mRNA were hybridized with the new CYP9 probes. Fig. 7A shows the two bands of 4.2 kb and 2.0 kb in midgut mRNA from xanthotoxin-fed larvae that hybridize to CYP9A2. For clofibrate, only the 4.2 kb band is seen. Hybridiz-ation to the CYP9A4 probe (Fig. 7B) reveals only the 4.2 kb band after clofibrate and xanthotoxin induction, with only a faint lower band in the xanthotoxin lane. In contrast, hybridization to the CYP9A5 probe (Fig. 7C) reveals only the 2.0 kb band in the xanthotoxin lane. Thus, CYP9A4 is inducible by both clofibrate and by xanthotoxin, and CYP9A5 is inducible by xanthotoxin and not by clofibrate. The CYP9A2 fragment crosshybri-dizes to both CYP9A4 and CYP9A5 under our con-ditions, and this explains the double banding pattern seen in our initial experiments.
This conclusion was further verified through ribonu-clease protection assays (RPA). Riboprobes of 425 bp and 322 bp, respectively, were prepared for CYP9A4 and CYP9A5. Hybridization of these probes to xantho-toxin-induced mRNA and 2-undecanone mRNA resulted in the presence of both a 322 bp and 425 bp protected fragments, while hybridization to clofibrate induced mRNA resulted in the presence of only a 425 bp pro-tected fragment (Fig. 8).
3.5. Genomic library screening with CYP9A4
Fig. 7. Northern blot hybridization of mRNA from midgut of 5th instar larvae of M. sexta hybridized with the CYP9A2 probe (panel A), the CYP9A4 probe (panel B) and the CYP9A5 probe (panel C). The lanes contained mRNA from insects fed a control diet (Cont), a diet containing 0.1% clofibrate (Clof), or a diet containing 0.1% xan-thotoxin (Xant).
the CYP9A2 fragment and with CYP9A4 (data not shown). Bands of approx 20 kb obtained with three restriction enzymes hybridized to both clones and the smallest band that hybridized to both clones was an Sph I fragment of 4.6 kb.
4. Discussion
The genome of plant species has accumulated an extraordinarily diverse set of P450 genes along with O-methyl transferases, isomerases, transferases etc. during evolution. It is thought that the enzymes they encode are necessary, in part, to synthesize the so-called secondary plant chemicals, many of which affect behavior, diges-tion, or more drastically, survival of insect herbivores in
Fig. 8. Ribonuclease protection assays with mRNA from midgut of 5th instar larvae of M. sexta. The riboprobe protected fragment for CYP9A4 is 435 bp long and that of CYP9A5 is 322 bp long. The lanes contained mRNA from insects fed a control diet, a diet contain-ing 0.1% clofibrate, 0.5% 2-undecanone or 0.1% xanthotoxin. Lane 1: CYP9A4 riboprobe with xanthotoxin mRNA; lane 2: CYP9A5 ribop-robe with xanthotoxin RNA. Lanes 3–6, both pribop-robes with 3: control mRNA; 4: clofibrate mRNA; 5: 2-undecanone mRNA and 6: xantho-toxin mRNA.
budworms (Rose et al., 1997) and may thus play a role in pesticide metabolism. In this paper, we provide evi-dence for CYP9 genes being expressed in the midgut of a specialist herbivore in response to the ingestion of plant chemicals and xenobiotics. Our work extends to the CYP9 family some observations that are also valid for members of the CYP6 and CYP4 family.
The members of the M. sexta CYP9 family are inducible in the midgut when larvae are fed a diet con-taining indole-3-carbinol, phenobarbital, 2-tridecanone, 2-undecanone, and xanthotoxin (Fig. 5). These results are a logical extension of earlier studies showing induc-tion of P450 enzymatic activity in different insect her-bivores in response to feeding either on host plants or artificial diets containing plant allelochemicals (e.g., Brattsten, 1978; Brattsten et al., 1977, 1984; Yu 1982, 1983). Two to four hours of feeding on 2-undecanone-containing diets are required before a significant increase in M. sexta CYP9 expression is found. This species appears to react more slowly than Spodoptera eridania in which a significant increase in P450 activity was found within 30 min of feeding on diets containing one of several plant chemicals (Brattsten, 1979; Brattsten et al., 1977). In another time-course study of an insect P450 induction, adult house flies (Musca domestica) induced CYP6A1 mRNA level within 1 h of exposure to pheno-barbital (Carin˜o et al., 1992). Ranasinghe et al. (1997) showed that monoterpene induction of CYP6B2 in H. armigera midgut is dose-dependent, with highest induc-tion observed at the highest tested (and probably tolerated) dose. However, their study shows a more tran-sient time-course, with highest increase in mRNA levels by 4 h after feeding, and a subsequent rapid decline. Thus, while the initial phase of rapid induction within 4 h seems a general characteristic, there are differences in the responsiveness of the genes over the longer term. Housefly CYP6A1 expression levels fell after 24 h of continuous phenobarbital exposure (Carin˜o et al., 1992), as with the M. sexta CYP9 in which the mRNA levels peaked at 48 h and then declined though the larvae were still ingesting large amounts of 2-undecanone. This finding expands previous observations of a potential feedback loop in insects that represses P450 gene tran-scription even during continuous consumption of the inducer (Capdevila et al., 1973; Vincent et al., 1985).
Our study also shows that closely related P450s can be induced differentially by chemical inducers: As dem-onstrated by RPA for CYP9A4 and CYP9A5 and as sug-gested for CYP9A2 by Northern blots, several CYP9 genes are induced by 2-undecanone and by xanthotoxin, but whereas CYP9A4 is induced by clofibrate, CYP9A5 is not. Differential induction was noted in our previous study with CYP4 genes in M. sexta (Snyder et al., 1995). The close similarity in CYP9A sequence causes potential problems in the interpretation of Northern blots as shown here and as also discussed by Ranasinghe and Hobbs
(1998) for CYP6B members in H. armigera. RT-PCR methods combined with hybridization may also reveal the presence of sequences closely related to the probe being used, as shown for the same CYP6B subfamily in Helicoverpa zea (Li et al., 2000). The ribonuclease protection assay therefore appears to be a method of choice, as it can distinguish between closely related tran-scripts as demonstrated by Hung et al. (1995a) for the CYP6B1 variants in P. polyxenes.
M. sexta larvae can encounter undecanone and 2-tridecanone in wild tomato, where the mixture of these aliphatic ketones kills neonates (Farrar and Kennedy, 1987). We show that levels of 2-undecanone (0.05%) as low as those found in this plant (0.066%—Farrar and Kennedy, 1987) significantly induces P450. The higher levels of 2-tridecanone in wild tomato (0.368%) also induce CYP9A2 and/or related CYP9 genes. CYP4M1 and CYP4M3 are induced by these compounds in M. sexta larvae (Snyder et al., 1995). The increase in total P450 levels observed by Riskallah et al. (1986) in H. virescens larvae fed wild tomato or 2-tridecanone there-fore represents the combined contribution of selected members of several P450 families, but probably not of all P450s. Interestingly, no induction by nicotine was observed with the CYP9A2 probe, although nicotine induced CYP4M1 and CYP4M3 (Snyder et al., 1995). Consequently, the same species encountering different classes of host plant chemicals will induce a different but sometimes overlapping subset of P450 genes from its repertoire. How can different P450 genes be subject to a common regulation? P450 genes constitute a large paralogous group in the genome, and their diversity results in part from multiple gene duplication events. When the portion of the genome being duplicated encompasses not just the P450 transcription unit but also the entire promoter/enhancer region of the P450 gene, then the two P450 genes resulting from this duplication are likely to have a common regulation. For instance, the recently duplicated CYP6B4 and CYP6B5 genes of Papilio glaucus are both furanocoumarin-inducible (Hung et al., 1996). In P. polyxenes, CYP6B1 and CYP6B3 are both induced by linear furanocoumarins, but CYP6B3 is also induced by angular furanocoumarins (Hung et al., 1995b). Although more divergent in sequence than the CYP6B4/5 pair, the CYP9A2/4 genes appear to have maintained clofibrate and xanthotoxin inducibility, and they are closely linked in the genome, less than 4.6 kb apart from each other. Nonetheless, gene duplication in the P450 superfamily provides not only the opportunity for divergence in substrate specificity, but also for divergence in regulatory attributes, such as induction, tissue or developmental patterns.
in H. zea (Li et al., 2000) and M. sexta (this study). This furanocoumarin also induces P450 and glutathione S-transferase activities in the fall armyworm, Spodoptera frugiperda (Yu, 1984; Kirby and Ottea, 1995). Sf9 cells from this latter species have the necessary complement of trans-acting factors to induce transfected CYP6B1
from P. polyxenes in response to xanthotoxin
(Prapaipong et al., 1994). It will be of interest to deter-mine whether the XRE-xan responsive element (Hung et al., 1996) of the CYP6B genes of Papilio sp. is more widely distributed in Lepidoptera, and amongst different P450 families, including M. sexta CYP9A. The possi-bility remains that xanthotoxin acts not via a receptor, but via a feedback mechanism. This furanocoumarin is a known inhibitor of several midgut P450 activities in larval M. sexta (Zumwalt and Neal, 1993). Perhaps the loss in enzyme activity modifies the titer of an endogen-ous metabolite which then somehow feeds back to induce the expression of P450 genes. However, whereas herbivores which do not normally feed on furanocouma-rin-containing plants (e.g., M. sexta) show limited metabolism of furanocoumarins and are significantly inhibited by the reactive metabolites, specialists such as Papilio polyxenes actively metabolize furanocoumarin to less reactive polar metabolites (Zumwalt and Neal, 1993; Berenbaum, 1999). Thus, since xanthotoxin inducibility is not restricted to furanocoumarin-consuming insects, we would rather favor the hypothesis that the XRE-xan responsive element is widespread in Lepidoptera, and that its linkage to genes of furanocoumarin-metabolizing P450s is the “key innovation” in the adapative radiation in the Papilionidae sought by Berenbaum et al. (1996). Inducibility of CYP9A genes by xenobiotics and plant allelochemicals suggests a role in metabolism of foreign compounds, as do the data of Rose et al. (1997) on CYP9A1 discussed above. However, this certainly does not exclude a physiological function towards endogen-ous substrates. The same observation about our ignor-ance of P450 functions in insects is valid for other P450 families. Members of the CYP4 family have been shown to be inducible in M.sexta larval midgut and fat body by xenobiotics and host plant chemicals (Snyder et al., 1995), an observation that has been ignored yet con-firmed for whole larvae of Drosophila mettleri fed cactus alkaloids or phenobarbital (Danielson et al., 1998). Inter-estingly, members of the CYP4 family in vertebrates are usually thought to be involved in the metabolism of endogenous substrates (Lake and Lewis, 1996). In insects, CYP4C1 expression in the fat body of the cock-roach, Blaberus discoidalis is inducible by starvation and by injection of the hypertrehalosemic hormone (Bradfield et al., 1991). Another cockroach gene, CYP4C7, is a terpenoidω-hydroxylase clearly involved in endocrine regulation of the corpora allata (Sutherland et al., 1998). When the large number (<100) of P450
genes in an insect genome is considered, the need for
new approaches in elucidating their function is under-scored.
Acknowledgements
This work was supported by USDA grants #89-37263-4960 and 92-37302-7789 to R.F. and NSF grant 9631128 to M.J.S., and by NIH grant ES06694. J.L.S. was sup-ported in part by the Undergraduate Biology Research Program at the Univesity of Arizona and by an NIGMS Summer Scholar fellowship.
References
Ausubel, F.M., Brent, R., Kingston, R.E., Moore, D.D., Seidman, J.G., Smith, J.A., Struhl, K., 1994. Current Protocols in Molecular Biology. John Wiley and Sons, Boston.
Berenbaum, M.R., 1999. Animal-plant warfare: molecular basis for cytochrome P450-mediated natural adaptation. In: Puga, A., Wal-lace, K.B. (Eds.) Molecular Biology of the Toxic Response. Taylor and Francis, Philadelphia, pp. 553–571.
Berenbaum, M.R., Favret, C., Schuler, M.A., 1996. On defining key innovations in an adaptive radiation — cytochrome P450s and Papilionidae. Am. Natural. 148, S139–S155.
Bernays, E., Graham, M., 1988. On the evolution of host specificity in phytophagous Arthropods. Ecology 69, 886–892.
Bradfield, J.Y., Lee, Y.-H., Keeley, L.L., 1991. Cytochrome P450 fam-ily 4 in a cockroach: molecular cloning and regulation by hypertre-halosemic hormone. Proc. Natl. Acad. Sci. USA 88, 4558–4562. Brattsten, L.B., Wilkinson, C.F., Eisner, T., 1977. Herbivore-plant
interactions: mixed function oxidases and secondary plant sub-stances. Science 196, 1349–1352.
Brattsten, L.B., 1979. Ecological significance of mixed-function oxi-dations. Drug Metab. Rev. 10, 35–58.
Brattsten, L.B., Evans, C.K., Bonetti, S., Zalkow, L.H., 1984. Induc-tion by carrot allelochemicals of insecticide-metabolizing enzymes in the Southern armyworm (Spodoptera eridania). Comp. Biochem. Physiol. 77C, 29–37.
Capdevila, J., Morello, A., Perry, A.S., Agosin, M., 1973. Effect of phenobarbital and naphthalene on some of the components of the electron transport system and the hydroxylating activity of house fly microsomes. Biochemistry 12, 1445–1451.
Carin˜o, F.A., Koerner, J.F., Plapp, F.W., Feyereisen, R., 1992. Expression of the cytochrome P450 gene CYP6A1 in the housefly
Musca domestica. ACS. Symp. Ser. 505, 31–40.
Cohen, M.B., Berenbaum, M.R., Schuler, M.A., 1992. A host plant inducible cytochrome P-450 from a host-specific caterpillar: mol-ecular cloning and evolution. Proc. Natl. Acad. Sci. USA 89, 10920–10924.
Danielson, P.B., Macintyre, R.J., Fogleman, J.C., 1997. Molecular cloning of a family of xenobiotic-inducible drosophilid cytochrome P450s: evidence for involvement in host-plant allelochemical resistance. Proc. Natl. Acad. Sci. USA 94, 10797–10802. Danielson, P.B., Foster, J.L., McMahill, M.M., Smith, M.K.,
Fogle-man, J.C., 1998. Induction by alkaloids and phenobarbital of family 4 cytochrome P450s in Drosophila: evidence for involvement in host plant utilization. Mol. Gen. Genet. 259, 54–59.
Dunkov, B.C., Rodriguez-Arnaiz, R., Pittendrigh, B., Ffrench-Con-stant, R.H., Feyereisen, R., 1996. Cytochrome P450 gene clusters in Drosophila melanogaster. Mol. Gen. Genetics 251, 290–297. Farrar, R.R., Kennedy, G.G., 1987. 2-undecanone, a constituent of the
on Heliothis zea and Manduca sexta growth and survival. Entomol. Exp. Appl. 43, 17–23.
Feyereisen, R., 1999. Insect P450 enzymes. A. Rev. Entomol. 44, 507–533.
Gonzalez, F.J., Nebert, D.W., 1990. Evolution of the P450 gene super-family: animal-plant “warfare”, molecular drive, and human gen-etic differences in drug oxidation. Trends Genet. 6, 182–186. Hung, C.F., Prapaipong, H., Berenbaum, M.R., Schuler, M.A., 1995a.
Differential induction of cytrochrome P-450 transcripts in Papilio
polyxenes by linear and angular furanocoumarins. Insect Biochem.
Mol. Biol. 25, 89–99.
Hung, C.F., Harrison, T.L., Berenbaum, M.R., Schuler, M.A., 1995b. CYP6B3: a second furanocoumarin-inducible cytochrome P450 expressed in Papilio polyxenes. Insect Mol. Biol. 4, 149–160. Hung, C.F., Holzmacher, R., Connolly, E., Berenbaum, M.R., Schuler,
M.A., 1996. Conserved promoter elements in the CYP6B gene fam-ily suggest common ancestry for cytochrome P450 monooxygen-ases mediating furanocoumarin detoxification. Proc. Natl. Acad. Sci. USA. 93, 12200–12205.
Kirby, M.L., Ottea, J.A., 1995. Multiple mechanisms for enhancement of glutathione S-transferase activities in Spodoptera frugiperda (Lepidopter: Noctuidae). Insect Biochem. Mol. Biol. 25, 347–353. Lake, B.G., Lewis, D.F.V., 1996. The CYP4 family. In: Ioannides, C. (Ed.) Cytochrome P450, metabolic and toxicological aspects. CRC Press, Boca Raton, pp. 271–297.
Lee, K., Berenbaum, M.R., 1989. Action of antioxidant enzymes and cytochrome P-450 monooxygenases in the cabbage looper in response to plant phototoxins. Arch. Insect Biochem. Physiol. 10, 151–162.
Li, X., Berenbaum, M.B., Schuler, M.A., 2000. Molecular cloning and expression of CYP6B8: a xanthotoxin-inducible cytochrome P450 cDNA from Helicoverpa zea. Insect Biochem. Mol. Biol. 30, 75–84.
Ma, R., Cohen, M.B., Berenbaum, M.R., Schuler, M.A., 1994. Black swallowtail (Papilio polyxenes) alleles encode cytochrome P450s that selectively metabolize linear furanocoumarins. Arch. Biochem. Biophys. 310, 332–340.
Nelson, D.R., 1998. Metazoan cytochrome P450 evolution. Comp. Biochem. Physiol. C 121, 15–22.
Nelson, D.R., Strobel, H.W., 1987. Evolution of cytochrome P-450 proteins. Mol. Biol. Evol. 4, 572–593.
Prapaipong, H., Berenbaum, M.R., Schuler, M.A., 1994. Transcrip-tional regulation of the Papilio polyxenes CYP6B1 gene. Nucl. Acids Res. 22, 3201–3217.
Prasad, S.V., Ryan, R.O., Law, J.H., Wells, M.A., 1986. Changes in lipoprotein composition during larval-pupal metamorphosis of an insect Manduca sexta. J. Biol. Chem. 261, 558–562.
Ranasinghe, C., Hobbs, A.A., 1998. Isolation and characterization of two cytochrome P450 cDNA clones for CYP6B6 and CYP6B7
from Helicoverpa armigera (Hubner): possible involvement of CYP6B7 in pyrethroid resistance. Insect Biochem. Mol. Biol. 28, 571–580.
Ranasinghe, C., Headlam, M., Hobbs, A.A., 1997. Induction of the mRNA for CYP6B2, a pyrethroid inducible cytochrome P450, in
Helicoverpa armigera (Hubner) by dietary monoterpenes. Arch.
Insect Biochem. Physiol. 34, 99–109.
Riskallah, M.R., Dauterman, W.C., Hodgson, E., 1986. Nutritional effects on the induction of cytochrome P-450 and glutathione trans-ferase in larvae of the tobacco budworm Heliothis virescens (F.). Insect Biochem. 16, 491–499.
Rose, R.L., Goh, D., Thompson, D.M., Verma, K.D., Heckel, D.G., Gahan, L.J., Roe, R.M., Hodgson, E., 1997. Cytochrome P450
(CYP)9A1: the first member of a new CYP family. Insect Biochem.
Mol. Biol. 27, 605–615.
Sambrook, J., Fritsch, E.F., Maniatis, T., 1989. Molecular Cloning: A Laboratory Manual. Cold Spring Harbor Laboratory Press, Cold Spring Harbor.
Sanger, F., Nicklen, S., Coulsen, A.R., 1977. DNA sequencing with chain-terminating inhibitors. Proc. Natl. Acad. Sci. USA 74, 5463–5467.
Snyder, M.J., Hsu, E.-L., Feyereisen, R., 1993. Induction of cyto-chrome P450 activities by nicotine in the tobacco hornworm
Mand-uca sexta. J. Chem. Ecol. 19, 2903–2916.
Snyder, M.J., Stevens, J.L., Andersen, J.F., Feyereisen, R., 1995. Expression of cytochrome P450 genes of the CYP4 family in midgut and fat body of the tobacco hornworm Manduca sexta. Arch. Biochem. Biophys. 321, 13–20.
Sutherland, T.D., Unnithan, G.C., Andersen, J.F., Evans, P.H., Murata-liev, M.B., Szabo, L.Z., Mash, E.A., Bowers, W.S., Feyereisen, R., 1998. A cytochrome P450 terpenoid hydroxylase linked to the suppression of insect juvenile hormone synthesis. Proc. Natl. Acad. Sci. USA 95, 12884–12889.
Vincent, D.R., Moldenke, A.F., Farnsworth, D.E., Terriere, L.C., 1985. Cytochrome P-450 in insects. 6. Age dependency and phenobarbital induction of cytochrome P-450, P-450 reductase, and monooxygen-ase activities in susceptible and resistant strains of Musca
dom-estica. Pestic. Biochem. Physiol. 23, 171–181.
Yu, S.J., 1982. Induction of microsomal oxidases by host plants in the fall armyworm Spodoptera frugiperda (J.E. Smith). Pestic. Biochem. Physiol. 17, 59–67.
Yu, S.J., 1983. Induction of detoxifying enzymes by allelochemicals and host plants in the fall armyworms. Pestic. Biochem. Physiol. 19, 330–336.
Yu, S.J., 1984. Interactions of allelochemicals with detoxication enzymes of insecticide-susceptible and resistant fall armyworms. Pest. Biochem. Physiol. 22, 60–68.
Zumwalt, J.G., Neal, J.J., 1993. Cytochromes P450 from Papilio
polxy-enes: adaptations to host plant allelochemicals. Comp. Biochem.