BAB IV
PENINGKATAN VIRULENSI Spodoptera exigua
NUCLEOPOLYHEDROVIRUS (SeNPV)
Increasing the virulence of Spodoptera exigua nucleopolyhedrovirus (SeNPV) Abstrak
Salah satu kelemahan Spodoptera exigua nucleopolyhedrovirus (SeNPV) untuk dijadikan agens hayati pengendali ulatgrayak bawang (UGB) adalah membutuhkan waktu relatif lama untuk mematikan serangga inangnya. Penelitian ini bertujuan untuk 1) mengetahui pengaruh pakan serangga inang terhadap virulensi SeNPV, 2) mendapatkan bahan perangsang makan (phagostimulant) yang efektif meningkatkan virulensi SeNPV, dan 3) memperoleh konsentrasi asam borat yang optimal sebagai enhancer SeNPV. Hasil penelitian menunjukkan bahwa virulensi SeNPV terhadap UGB yang diberi pakan daun bawang merah, daun bawang daun dan pakan buatan tidak berbeda nyata. Pakan buatan yang digunakan dinilai cocok untuk perbanyakan dan produksi SeNPV. Dari lima jenis bahan yang diduga sebagai phagostimulant, kecap 5% dan sukrosa 5% secara signifikan meningkatkan konsumsi UGB pada pakan buatan dan meningkatkan mortalitas larva yang terinfeksi SeNPV. Penggunaan sukrosa 1 sampai 5% dinilai efektif meningkatkan konsumsi UGB dan virulensi SeNPV di laboratorium. Sedangkan konsentrasi sukrosa 10% cenderung menurunkan aktifitas makan UGB. Asam borat terbukti dapat meningkatkan virulensi SeNPV. Semakin tinggi konsentrasi asam borat yang digunakan, semakin rendah nilai LT50 UGB terinfeksi SeNPV. Akan tetapi penggunaannya harus dibatasi maksimal 5%, untuk menghindari dampak negatif terhadap lingkungan.
Abstract
Spodoptera exigua nucleopolyhedrovirus (SeNPV) is an entomophathogenic virus of onion caterpillar S. exigua larvae commonly used as bioinsecticide. The major limitations of SeNPV for biocontrol of onion caterpillar is it requires long time for the virus to kill the insect host. This research was aimed to: (1) investigate the effect of insect diets on the virulence of SeNPV, (2) find out the material as phagostimulant to increase SeNPV virulence, and (3) determine the optimum boric acid concentra-tion as an enhancer for SeNPV. The results showed that the infectivity of the SeNPV to the larvae that feeding on shallot, leek and artificial diets was not significantly different. This result indicated that artificial diet used was considered suitable for the propagation and production of SeNPV. Soybean sauces 5% and sucrose 5% increased S. exigua consumption on artificial diets and enhanced viral activity. When mixed with polyhedra of SeNPV, 1% to 5% sucrose significantly increased S. exigua consumption and increased the virulence of SeNPV, while 10% sucrose tended to decrease the feeding activity of S. exigua. Boric acid concentrations enhanced the SeNPV virulence. The LT50 of SeNPV was decreased as the concentration of boric acids increased. However, the increase of boric acid concentration should be restricted maximum 5%, for avoiding the negative impacts on the environment. Keywords: phagostimulant, enhancers, UGB, SeNPV, virulence
Pendahuluan
Spodoptera exigua nucleopolyhedrovirus (SeNPV) adalah virus patogen serangga (entomophatogen) yang potensial untuk dikembangkan sebagai bioinsektisida pengendali ulatgrayak bawang (UGB) S. exigua yang ramah lingkungan. Beberapa keunggulan penggunaan NPV dibandingkan dengan insektisida kimia, antara lain: tidak membunuh organisme bukan sasaran, memperbanyak diri dalam tubuh inang dan menyebar melalui transmisi sekunder sehingga dapat mengendalikan hama sasaran berikutnya, tidak mengakibatkan resistensi hama sasaran, tidak meninggalkan residu berbahaya pada makanan, membantu upaya pelestarian musuh alami dan dapat meningkatkan biodiversitas (Young 1989; Lacey et al. 2001; Armenta et al. 2003). Akan tetapi kinerja NPV dalam mematikan inangnya di lapangan masih membutuhkan waktu relatif lama, sehingga serangga yang terinfeksi masih makan dan menimbulkan kerugian (Arifin 1988; Bonning &
Hammock 1996; Dushoff & Greg 2001; Trang & Chaudhari 2002). Salah satu penyebab lambatnya kinerja NPV tersebut disebabkan oleh sedikitnya jumlah polihedra yang termakan oleh serangga inang (Cisneros et al. 2002; Castillejos et al. 2002). Masalah tersebut dapat diatasi dengan dua pendekatan, yaitu: menambahkan perangsang makan (phagostimulant) untuk meningkatkan jumlah inokulum yang dimakan serangga inang, atau menggunakan bahan yang dapat meningkatkan virulensi NPV (Suhas et al. 2009).
NPV dapat menginfeksi serangga inang hanya apabila polihedra termakan oleh serangga inang melalui pakan yang terkontaminasi (Vasconcelos et al. 2002). Salah satu upaya untuk meningkatkan jumlah polihedra yang dikonsumsi adalah dengan menambahkan bahan-bahan yang dapat merangsang makan (phagostimulant) (Farrar et al. 2005; Genc 2006). Menurut Genc (2006) phagostimulant adalah bahan kimia yang memiliki pengaruh pada aktivitas makan serangga. Mcfarlane (1985) menyatakan bahwa, beberapa bahan kimia seperti asam amino, gula, asam lemak, sterol dan vitamin, dapat merangsang makan bagi serangga. Hasil penelitian Ahmad et al. (2001) menunjukkan bahwa komposisi bahan pakan terutama protein dan karbohidrat sangat berpengaruh pada performan larva S. exempta. Lebih lanjut Ahmad & Kamal (2001) menyatakan bahwa diantara jenis karbohidrat, sukrosa memiliki pengaruh yang lebih baik terhadap pertumbuhan larva S. exempta daripada glukosa, fruktosa, mannitol dan maltosa. Hasil penelitian Genc (2006) menegaskan bahwa gula heksosa dan sukrosa merupakan bahan nutrisi utama serangga dan berfungsi sebagai phagostimulant bagi serangga pemakan daun.
Upaya untuk meningkatkan virulensi NPV dapat dilakukan beberapa cara, antara lain menambahkan bahan pemicu kinerja (enhancer) (Lacey et al. 2001), mengkombinasikan dengan bahan yang bekerja secara sinergis (Dodin 2001; Samsudin 2001; Arifin 2006; Shapiro & Shepard 2006) atau melalui rekayasa genetik (Treacy 1999). Di antara ketiga cara tersebut yang paling efektif dan efesien untuk dilakukan adalah dengan menambahkan bahan pemicu kinerja (enhancer). Salah satu bahan yang berpotensi sebagai bahan enhancer adalah asam borat (boric acid) (Shapiro & Bell 1982; Chaudhari 1992). Hasil penelitian Morales et al. (1997)
menunjukkan bahwa penambahan asam borat pada Anticarsia gemmatalis NPV (AgNPV) dapat meningkatkan mortalitas larva A. gemmatalis dan memperpendek waktu kematiannya. Sedangkan Cisneros et al. (2002) melaporkan bahwa penambahan 0.5% dan 1% asam borat secara signifikan dapat meningkatkan kematian larva S. frugiferda yang SfNPV.
Penelitian ini bertujuan untuk: 1) mengetahui pengaruh pakan serangga inang terhadap virulensi SeNPV, 2) mendapatkan bahan phagostimulant yang efektif meningkatkan virulensi SeNPV dan 3) memperoleh konsentrasi asam borat yang optimal sebagai enhancer SeNPV.
Bahan dan Metode
Penelitian dilaksanakan mulai bulan Agustus 2009 sampai Januari 2010 di laboratorium Lembaga Pertanian Sehat (LPS) Bogor
Pengaruh Jenis Pakan UGB terhadap Virulensi SeNPV
Penelitian menggunakan rancangan acak lengkap (RAL) dengan 3 perlakuan jenis pakan yaitu 1) daun bawang daun (Allium fistulosum), 2) daun bawang merah (A. ascalonicum L.) dan 3) pakan buatan (lampiran 2). Setiap perlakuan diulang sebanyak 3 kali menggunakan 30 ekor larva UGB instar 3. Serangga uji yang digunakan adalah larva S. exigua instar 3 hasil perbanyakan di laboratorium. Isolat SeNPV yang digunakan adalah suspensi virus murni yang telah diencerkan dengan konsentrasi 1,13 x 108 POB/ml. Perlakuan menggunakan metode kontaminasi pakan (Hunter-Fujita et al. 1998).
Bahan pakan yang diuji disiapkan terlebih dahulu masing-masing seberat 2 gram per wadah. Perlakuan pakan alami menggunakan cawan petri plastik diameter 13 cm yang telah diberi lubang kecil pada bagian tutupnya, sedangkan untuk pakan buatan menggunakan cawan plastik. Aplikasi suspensi virus dilakukan dengan metode pencelupan untuk bahan pakan alami dan diteteskan sebanyak 3 tetes per
wadah untuk pakan buatan. Setelah pakan alami ditiriskan selama 1 jam, pakan tersebut dimasukan ke dalam wadah. Setiap hari dicatat jumlah serangga uji yang mati terinfeksi virus sampai semua serangga uji mati atau menjadi pupa. Persentase mortalitas dihitung berdasarkan rumus Abbott (1925).
Pt = Po – Pk x 100% 100 – Pk
Ket: Po : Persentase kematian larva yang diamati Pt : Persentase kematian larva terkoreksi Pk : Persentase kematian larva pada kontrol.
Pengujian Bahan Perangsang Makan (phagostimulant)
Penelitian dilakukan di laboratorium dengan menggunakan pakan buatan. Bahan yang diduga sebagai perangsang makan yang digunakan adalah: 1) sukrosa (5%), 2) gula putih (5%), 3) molase (5%), 4) kecap (5%), 5) filtrat kunyit (5%), 6) hanya virus (kontrol positif) dan 7) air steril (kontrol pengoreksi). Suspensi virus dengan konsentrasi 1.13 x 108 POB/ml diteteskan ke dalam pakan buatan dalam wadah plastik, kemudian dimasukkan ke dalamnya masing-masing 1 larva instar 3. Setiap perlakuan menggunakan 30 ekor larva dan diulang sebanyak 3 kali. Peubah yang diamati yaitu; 1) jumlah serangga uji yang mati terinfeksi virus, 2) bobot feses larva, dan 3) waktu kematian larva. Persentase mortalitas dihitung berdasarkan rumus Abbott (1925) dan untuk menganalisis aktifitas makan dilihat dari bobot feses yang dikeluarkan larva (Idris & Emelia 2001).
Untuk mengetahui adanya peningkatan kinerja SeNPV akibat dari penambahan bahan-bahan yang diduga sebagai phagostimulant digunakan rumus Peningkatan Kinerja Relatif (PKR), yaitu:
PKR(%) = % mortalitas perlakuan - % mortalitas kontrol positif X 100 % % mortalitas kontrol positif
Untuk menilai efesiensi bahan-bahan yang diuji dalam meningkatkan kinerja SeNPV dihitung dengan menggunakan rumus efesiensi relatif (ER) (Mehrvar et al. 2008), yaitu:
Mortalitas perlakuan Mortalitas kontrol positif
Kinerja bahan yang diduga sebagai phagostimulant terhadap perilaku makan dinilai dengan menggunakan indeks feeding stimulant (IFS) ( Matsuda et al. 1998) dengan rumus:
IFS = Bobot feses perlakuan – bobot feses kontrol positif x 100% Bobot feses perlakuan + bobot feses kontrol positif
Penentuan Konsentrasi Sukrosa yang Efektif Meningkatkan Virulensi SeNPV.
Penelitian dilakukan di laboratorium menggunakan rancangan acak lengkap (RAL) dengan metode kontaminasi pakan (Hunter-Fujita et al. 1998). Konsentrasi sukrosa yang digunakan adalah: 0% (kontrol), 1%, 5%, dan 10% dari suspensi yang berisi polihedra dengan konsentrasi 1,13 x 108POB/ml. Perlakuan dilakukan dengan cara kontaminasi pada permukaan pakan buatan dalam wadah plastik. Setelah diberi perlakuan dimasukan ke dalam masing-masing wadah tersebut 1 ekor larva S. exigua instar 3. Masing-masing perlakuan menggunakan 30 ekor larva dan diulang sebanyak 3 kali.
Variabel yang diamati adalah jumlah dan waktu mortalitas serangga uji. Pengamatan dilakukan sampai semua serangga uji pada kontrol pengoreksi menjadi pupa. Untuk mengetahui adanya peningkatan kinerja SeNPV akibat dari penambahan sukrosa digunakan rumus Peningkatan Kinerja Relatif (PKR), sedangkan untuk menilai efesiensi sukrosa dalam meningkatkan kinerja SeNPV dihitung dengan menggunakan rumus efesiensi relative (ER) (Mehrvar et al. 2008).
Pengujian Asam Borat sebagai Pemicu Kinerja (Enhancer) SeNPV
Pengujian asam borat sebagai enhancer dilaksanakan di laboratorium dengan menggunakan rancangan acak lengkap (RAL) dengan metode kontaminasi pakan (Hunter-Fujita et al. 1998). Konsentrasi asam borat yang digunakan adalah: 0% (kontrol), 0,1%, 1%, 5%, dan 10% dari suspensi yang berisi polihedra dengan konsentrasi 1,13 x 108 POB/ml. Setelah diberi perlakuan, dimasukan ke dalam masing-masing wadah tersebut 1 ekor larva S. exigua instar 3. Masing-masing perlakuan menggunakan 30 ekor larva dan diulang sebanyak 3 kali.
Variabel yang diamati adalah mortalitas dan waktu kematian serangga uji. Pengamatan dilakukan sampai semua serangga uji pada kontrol negatif menjadi pupa. Persentase mortalitas dikoreksi berdasarkan rumus Abbott (1925). Untuk menentukan virulensi SeNPV berupa LT50dihitung dengan menggunakan probit analisis. Untuk mengetahui adanya peningkatan kinerja SeNPV akibat penambahan bahan enhancer digunakan rumus Peningkatan Kinerja Relatif (PKR), dan untuk menilai efesiensi asam borat dalam meningkatkan kinerja SeNPV dihitung dengan menggunakan rumus efesiensi relatif (ER) (Mehrvar et al. 2008).
Analisis Data
Semua data yang diperoleh dianalisis menggunakan program SAS. Apabila terdapat perbedaan diantara perlakuan maka dilanjutkan dengan uji jarak berganda (Duncan’s Multiple Range Test) pada taraf nyata α = 0,05.
Hasil dan Pembahasan
Pengaruh Jenis Pakan terhadap Virulensi SeNPV pada S. exigua
Hasil pengujian terhadap tiga jenis pakan UGB, yaitu: daun bawang merah, daun bawang daun dan pakan buatan di laboratorium menunjukkan bahwa pengaruh ketiga jenis pakan tersebut sama (Tabel 4.1). Meskipun pada pengamatan hari ke-4 setelah inokulasi menunjukkan bahwa mortalitas pada pakan buatan lebih tinggi (17,78%) dibandingkan dengan daun bawang merah (7,87%) dan bawang daun (3,70%), akan tetapi mortalita pada pengamatan hari terakhir tidak berbeda nyata.
Hasil penelitian ini menunjukkan bahwa ketiga jenis pakan yang digunakan merupakan pakan yang sesuai untuk pertumbuhan dan perkembangan UGB. Daun bawang merah dan bawang daun merupakan pakan alami utama dari S. exigua di lapangan, sedangkan pakan buatan yang diuji adalah pakan buatan yang digunakan untuk perbanyakan reguler di laboratorium LPS. Dari ketiga jenis pakan tersebut, pakan buatan merupakan jenis yang paling mudah penanganannya dan dapat digunakan untuk menguji bahan-bahan perangsang makan (phagostimulant). Hasil penelitian sebelumnya mengindikasikan bahwa aktifitas virus patogen serangga dipengaruhi oleh pakan serangga inangnya. Hasil penelitian Farrar & Ridgway (2000), menunjukkan bahwa aktifitas Anagrapha falcifera MNPV (AfMNPV) terhadap S. exigua dan Heliverpoa zea dipengaruhi oleh pakan. Aktifitas AfMNPV terhadap S. exigua paling tinggi ketika serangga tersebut diberi pakan tomat (Lycopersicon esculentum) kemudian tanaman kol (Brassica oleracea) dan paling rendah tanaman kapas (Gossypium hirsutum). Aktifitas AfMNPV pada Heliverpoa zea paling tinggi pada pakan jagung (Zea mays), kemudian pada tanaman kacang hijau (Phaseolus vulgaris) dan paling rendah pada tanaman kapas (Gossypium hirsutum).
Kandungan nutrisi dari pakan serangga inang mempengaruhi tingkat prevalensi infeksi virus. Review hasil penelitian yang dilakukan oleh Tanada & Kaya (1993) menunjukkan bahwa kandungan nutrisi pakan, yaitu air, nitrogen dan sukrosa
dapat mempengaruhi prevalensi infeksi virus. Semakin tinggi kandungan nitrogen semakin tinggi prevalensi infeksi virus. Sebaliknya penurunan kandungan air dan sukrosa pada pakan buatan akan meningkatkan mortalitas larva yang terinfeksi virus.
Tabel 4.1. Persentase mortalitas UGB terinfeksi SeNPV pada berbagai jenis pakan Perlakuan
Mortalitas (%)a Hari setelah inokulasi
4 5 6
Daun bawang merah 7,87 + 6,85 ab 37,74 + 4,15 a 58,33 + 8,97 a Daun bawang daun 3,70 + 6,42 b 31,03 + 19,91 a 60,69 + 8,36 a Pakan buatan 17,78 + 5,88 a 39,66 + 7,47 a 60,13 + 4,99 a aRataan pada kolom yang sama yang diikuti oleh huruf yang sama tidak menunjukkan beda nyata (Uji Duncan, α = 0,05).
Waktu kematian UGB terinfeksi SeNPV pada 3 jenis pakan yang digunakan juga tidak berbeda nyata. Sampai hari ke-3 setelah perlakuan belum ada UGB yang mati terinfeksi SeNPV. Hasil ini mengindikasikan bahwa, ketiga jenis pakan yang digunakan tidak berkontribusi dalam mempercepat kinerja SeNPV dalam mematikan UGB di Laboratorium. Goulson et al. (1995) menyatakan bahwa kecepatan virulensi NPV dipengaruhi oleh ukuran serangga bukan dari jumlah konsumsi pakannya. Serangga instar 2 lebih cepat mati daripada instar 3, dan instar 3 lebih cepat dari instar 4, meskipun jumlah konsumsi pakan sebaliknya.
Pertumbuhan UGB pada ketiga jenis pakan yang diuji tidak menunjukkan perbedaan. Hal itu menurut Azidah & Sofian-Azirun (2006) disebabkan ketiga jenis pakan tersebut cocok untuk S. exigua. Sebab umumnya perkembangan, lama hidup dan keperidian serangga pemakan daun, akan dipengaruhi oleh tanaman inangnya.
Pengaruh Perangsang Makan (phagostimulant) terhadap Virulensi SeNPV
Menurut Sutherland (1977) secara umum phagostimulant dibagi menjadi 2 kategori, yaitu bahan nutrisi dan metabolit sekunder. Menurutnya metabolit sekunder yang terdapat dalam tanaman bukan sebagai makanan, tetapi hanya menjadi perangsang bagi serangga sehingga dapat memilih dan menemukan makanan yang cocok atau tanaman inangnya. Farrar & Ridgway (2000) menyatakan bahwa bagian tanaman utuh, juice, ekstrak dan bahan kimia hasil pemurnian dari tanaman dapat mempengaruhi aktifitas patogen terhadap serangga inang. Bahan-bahan dari tanaman tersebut dapat berfungsi sebagai perangsang (enhancer), penghambat (inhibitor) atau tidak berpengaruh.
Sampai pengamatan hari ke-4 setelah perlakuan tidak ditemukan perbedaan yang nyata pada akumulasi rata-rata persentase mortalitas UGB antara perlakuan dengan kontrol. Perbedaan yang signifikan terjadi mulai hari ke-5, penambahan kecap 5% secara nyata meningkatkan mortalitas UGB terinfeksi SeNPV yaitu 51,21% dibandingkan dengan kontrol positif 31,91%. Pada pengamatan hari ke-6, penambahan sukrosa 5% juga dapat meningkatkan kinerja SeNPV dengan mortalitas rata-rata sebesar 57,17% dibandingkan dengan kontrol sebesar 42,74%. Sementara itu penambahan gula putih, molase dan filtrat kunyit masing-masing 5% sampai hari ke-6 setelah perlakuan pengaruhnya tidak berbeda nyata dengan kontrol (Tabel 4.2). Akan tetapi pada penelitian lapangan yang dilakukan oleh Arifin & Nuzullianti (1999), penambahan molase 20% efektif meningkatkan virulensi Spodoptera litura NPV (SlNPV).
Tabel 4.2. Akumulasi rata-rata persentase mortalitas UGB setelah perlakuan
Perlakuan Mortalitas (%)a
Hari setelah inokulasi
3 4 5 6 Sukrosa 5% 6,20 + 7,42 a 19,38 + 14,92 a 34,99 + 9,73 b 57,17 + 5,77 ab Gula putih 5% 5,57 + 7,31 a 14,17 + 8,46 a 31,51 + 8,12 b 47,07 + 4,79 c Molase 5% 4,55 + 5,62 a 13,47 + 10,98 a 34,95 + 10,70 b 52,96 + 3,67 bc Kecap 5% 7,69 + 15,95 a 24,12 + 9,15 a 51,21 + 13,31 a 64,20 + 8,05 a Filtrat kunyit 5% 5,36 + 6,19 a 13,91 + 7,26 a 25,84 + 7,83 b 44,31 + 6,65 c Hanya virus 2,67 + 3,42 a 18,98 + 11,63 a 31,91 + 8,54 b 42,74 + 8,08 c
aRataan pada kolom yang sama yang diikuti oleh huruf yang sama tidak menunjukkan beda nyata (Uji Duncan, α = 0,05).
Nilai peningkatan kinerja relatif (PKR) dari bahan yang diuji menunjukkan bahwa, kecap dan sukrosa masing-masing 5% dapat meningkatkan kinerja SeNPV berturut-turut 50,21% dan 33,76%. Sementara itu 5% molase, gula dan filtrat kunyit dapat meningkatkan kinerja SeNPV sebesar masing-masing 23,91%, 10,13% dan 3,67% (Tabel 4.3). Meskipun semua bahan yang digunakan dapat meningkatkan kinerja dari SeNPV terhadap UGB, akan tetapi peningkatan yang berbeda nyata secara statistik dengan kontrol terjadi pada nilai PKP lebih dari 30%. Demikian pula dengan nilai efesiensi relatif (ER) meskipun semua nilainya di atas 1, akan tetapi yang berbeda nyata secara statistik apabila nilai ER lebih dari 1,30 (Tabel 4.3). Hasil penelitian Mehrvar et al. (2008) menunjukkan bahwa molase dan gula dapat berfungsi sebagai phagostimulant, sehingga dapat meningkatkan jumlah virus yang termakan dan meningkatkan mortalitas larva.
Tabel 4.3. Nilai peningkatan kinerja relatif (PKR), efesiensi relatif (ER) dan indeks feeding stimulant (IFS) dari bahan-bahan yang diuji
Perlakuan PKR (%)* ER** IFS*** Sukrosa 5% 33,76 1,34 0,37 Gula putih 5% 10,13 1,10 2,00 Molase 5% 23,91 1,24 3,24 Kecap 5% 50,21 1,50 8,97 Filtrat kunyit 5% 3,67 1,04 - 4,67 Hanya virus 0,00 1,00 0,00
* Nilai PKR positif menunjukkan adanya peningkatan kinerja **
Nilai ER lebih dari 1 menunjukkan peningkatan kinerja
***
Nilai positif menunjukkan kinerja feeding stimulant sedangkan nilai negative menunjukkan kinerja antifeedant
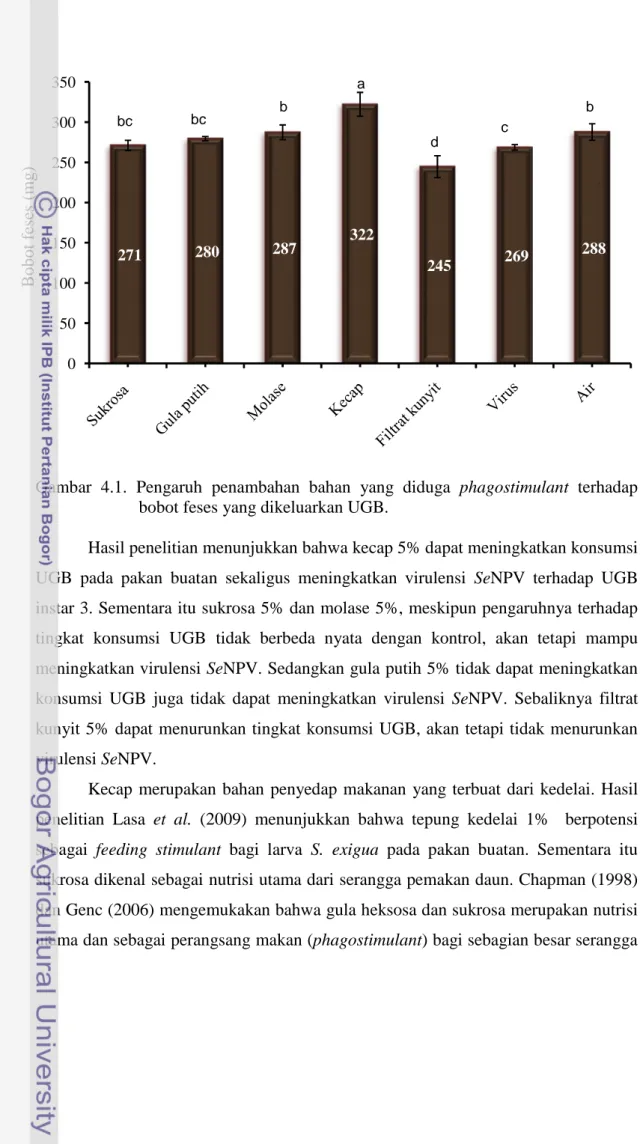
Secara teoritis persentasi mortalitas UGB terinfeksi virus berkorelasi positif dengan tingkat konsumsi pakan. Semakin tinggi tingkat konsumsi pakan serangga inang, maka akan semakin tinggi mortalitasnya. Dari lima jenis bahan yang diuji, hanya kecap 5% yang secara signifikan meningkatkan tingkat konsumsi UGB pada pakan buatan. Rata-rata bobot feses yang dikeluarkan oleh UGB selama tiga hari adalah 322 mg yang berbeda nyata dengan kontrol negatif (hanya air) 288 mg. UGB pada perlakuan sukrosa, gula putih dan molase rata-rata mengeluarkan feses masing-masing: 271, 280 dan 287 mg dan tidak berbeda nyata dibandingkan dengan kontrol negatif yaitu 288 mg. Sementara itu UGB yang diberi perlakuan filtrat kunyit 5% rata-rata mengeluarkan feses seberat 245 mg dibawah kontrol negatif 288 mg dan kontrol positif 269 mg (Gambar 4.1).
Berdasarkan nilai indeks feeding stimulant (IFS) diketahui bahwa sukrosa, gula putih, molase dan kecap memiliki nilai IFS lebih besar dari nol (positif). Hal ini mengindikasikan bahwa keempat bahan tersebut berpotensi sebagai bahan feeding stimulant. Sementara itu filtrate kunyit 5% memiliki nilai IFS dibawah nol (- 4,67) yang mengindikasikan bahwa bahan tersebut berpotensi sebagai antifeedant.
271 280 0 50 100 150 200 250 300 350 B o b o t fe se s (m g ) bc bc
Gambar 4.1. Pengaruh penambahan bahan yang diduga phagostimulant terhadap bobot feses yang dikeluarkan UGB.
Hasil penelitian menunjukkan bahwa kecap 5% dapat meningkatkan konsumsi UGB pada pakan buatan sekaligus meningkatkan virulensi SeNPV terhadap UGB instar 3. Sementara itu sukrosa 5% dan molase 5%, meskipun pengaruhnya terhadap tingkat konsumsi UGB tidak berbeda nyata dengan kontrol, akan tetapi mampu meningkatkan virulensi SeNPV. Sedangkan gula putih 5% tidak dapat meningkatkan konsumsi UGB juga tidak dapat meningkatkan virulensi SeNPV. Sebaliknya filtrat kunyit 5% dapat menurunkan tingkat konsumsi UGB, akan tetapi tidak menurunkan virulensi SeNPV.
Kecap merupakan bahan penyedap makanan yang terbuat dari kedelai. Hasil penelitian Lasa et al. (2009) menunjukkan bahwa tepung kedelai 1% berpotensi sebagai feeding stimulant bagi larva S. exigua pada pakan buatan. Sementara itu sukrosa dikenal sebagai nutrisi utama dari serangga pemakan daun. Chapman (1998) dan Genc (2006) mengemukakan bahwa gula heksosa dan sukrosa merupakan nutrisi utama dan sebagai perangsang makan (phagostimulant) bagi sebagian besar serangga
280 287 322 245 269 bc b a d c
Gambar 4.1. Pengaruh penambahan bahan yang diduga phagostimulant terhadap bobot feses yang dikeluarkan UGB.
Hasil penelitian menunjukkan bahwa kecap 5% dapat meningkatkan konsumsi UGB pada pakan buatan sekaligus meningkatkan virulensi SeNPV terhadap UGB instar 3. Sementara itu sukrosa 5% dan molase 5%, meskipun pengaruhnya terhadap tingkat konsumsi UGB tidak berbeda nyata dengan kontrol, akan tetapi mampu meningkatkan virulensi SeNPV. Sedangkan gula putih 5% tidak dapat meningkatkan konsumsi UGB juga tidak dapat meningkatkan virulensi SeNPV. Sebaliknya filtrat kunyit 5% dapat menurunkan tingkat konsumsi UGB, akan tetapi tidak menurunkan virulensi SeNPV.
Kecap merupakan bahan penyedap makanan yang terbuat dari kedelai. Hasil penelitian Lasa et al. (2009) menunjukkan bahwa tepung kedelai 1% berpotensi sebagai feeding stimulant bagi larva S. exigua pada pakan buatan. Sementara itu sukrosa dikenal sebagai nutrisi utama dari serangga pemakan daun. Chapman (1998) dan Genc (2006) mengemukakan bahwa gula heksosa dan sukrosa merupakan nutrisi utama dan sebagai perangsang makan (phagostimulant) bagi sebagian besar serangga
269 288
b
Gambar 4.1. Pengaruh penambahan bahan yang diduga phagostimulant terhadap bobot feses yang dikeluarkan UGB.
Hasil penelitian menunjukkan bahwa kecap 5% dapat meningkatkan konsumsi UGB pada pakan buatan sekaligus meningkatkan virulensi SeNPV terhadap UGB instar 3. Sementara itu sukrosa 5% dan molase 5%, meskipun pengaruhnya terhadap tingkat konsumsi UGB tidak berbeda nyata dengan kontrol, akan tetapi mampu meningkatkan virulensi SeNPV. Sedangkan gula putih 5% tidak dapat meningkatkan konsumsi UGB juga tidak dapat meningkatkan virulensi SeNPV. Sebaliknya filtrat kunyit 5% dapat menurunkan tingkat konsumsi UGB, akan tetapi tidak menurunkan virulensi SeNPV.
Kecap merupakan bahan penyedap makanan yang terbuat dari kedelai. Hasil penelitian Lasa et al. (2009) menunjukkan bahwa tepung kedelai 1% berpotensi sebagai feeding stimulant bagi larva S. exigua pada pakan buatan. Sementara itu sukrosa dikenal sebagai nutrisi utama dari serangga pemakan daun. Chapman (1998) dan Genc (2006) mengemukakan bahwa gula heksosa dan sukrosa merupakan nutrisi utama dan sebagai perangsang makan (phagostimulant) bagi sebagian besar serangga
pemakan daun. Dari hasil penelitian ini terbukti bahwa bahan phagostimulant dapat meningkatkan virulensi SeNPV. Hasil penelitian Meade & Hare (1993) menunjukkan bahwa jika jenis pakan yang dimakan oleh S. exigua lebih banyak, maka mortalitas akibat B. thuringiensis juga semakin tinggi.
Bahan-bahan alami yang murah dan mudah memperolehnya apabila terbukti mampu meningkatkan tingkat konsumsi UGB pada tanaman inang dan meningkatkan virulensi SeNPV terhadap UGB, maka akan mengurangi biaya aplikasi bioinsektisida SeNPV. Menurut Castillejos et al. (2002) penambahan bahan-bahan phagostimulant dalam formulasi bioinsektisida virus di samping akan meningkatkan keefektifannya juga dapat mengendalikan hama dengan konsentrasi polihedra rendah dan menghemat biaya aplikasi.
Pengaruh Konsentrasi Sukrosa terhadap Peningkatan Virulensi SeNPV.
Hasil penelitian menunjukkan bahwa penambahan sukrosa 1% dan 5% dapat meningkatkan mortalitas UGB secara signifikan dibandingkan dengan kontrol. Sedangkan penambahan sukrosa 0,10% dan 10% pengaruhnya tidak berbeda nyata dibandingkan dengan kontrol (Tabel 4.4). Penambahan sukrosa 10% tidak dapat meningkatkan mortalitas UGB, hal itu disebabkan karena ada sebagian larva yang tidak mengkonsumsi pakan. Hal itu mengindikasikan bahwa kesesuaian pakan serangga memerlukan komposisi yang tepat. Ahmad et al. (2001) menyatakan bahwa, komposisi bahan pakan terutama protein dan karbohidrat sangat berpengaruh pada aktifitas larva Spodoptera exempta.
Tabel 4.4. Pengaruh perlakuan sukrosa terhadap mortalitas UGB terinfeksi SeNPV
SeNPVa+ Sukrosa (%)
Mortalitas (%)b Hari setelah inokulasi
3 4 5 6 0 1,19 + 2,06 b 15,72 + 11,72 a 33,22 + 6,62 ab 44,73 + 6,50 b 0,1 2,38 + 2,07 b 2,60 + 2,05 b 28,33 + 5,95 b 52,69 + 3,21 ab 1 9,57 + 2,33 a 17,54 + 5,31 a 40,63 + 3,04 ab 57,16 + 3,36 a 5 5,11 + 5,17 ab 11,36 + 6,44 ab 42,31 + 10,28 a 63,18 + 1,90 a 10 1,11 + 1,93 b 7,58 + 3,18 ab 27,48 + 7,19 b 46,12 + 9,30 b a
Konsentrasi SeNPV yang digunakan 1,13 x 107POB/ml
b Rataan pada kolom yang sama yang diikuti oleh huruf yang sama tidak menunjukkan beda nyata (Uji Duncan, α = 0,05).
Nilai peningkatan kinerja relative (PKR) dari penambahan larutan sukrosa 0,10%, 1%, 5% dan 10% masing-masing sebesar: 17,81%, 27,79%, 41,24% dan 3,09%. Sementara itu nilai efesiensi relative (ER) dari semua perlakuan konsentrasi sukrosa lebih dari 1. Akan tetapi nilai indeks feeding stimulant (IFS) dari perlakuan konsentrasi sukrosa 10% bernilai negatif yang mengindikasikan bekerja sebagai antifeedant (Tabel 4.5). Data ini mengindikasikan bahwa konsentrasi sukrosa yang direkomendasikan berkisar antara 1 sampai 5%.
Tabel 4.5. Nilai peningkatan kinerja relatif (PKR), efesiensi relatif (ER) dan indeks feeding stimulant (IFS) dari perlakuan sukrosa
SeNPVa+ Sukrosa (%) PKR (%)b ERc IFSd
0 0 1,00 0
0,1 17,809 1,18 0,388
1 27,788 1,28 2,652
5 41,239 1,41 9,825
10 3,094 1,03 -6,418
a Konsentrasi SeNPV yang digunakan 1,13 x 107POB/ml bNilai PKR positif menunjukkan adanya peningkatan kinerja cNilai ER lebih dari 1 menunjukkan peningkatan kinerja
dNilai positif menunjukkan kinerja phagostimulant sedangkan nilai negative menunjukkan kinerja
Pengujian Asam Borat sebagai Pemicu Kinerja (Enhancer) SeNPV
Asam borat atau boric acid (H3BO3) merupakan senyawa kimia turunan dari logam berat boron (B) yang biasa digunakan untuk pembersih, pengawet, antiseptik dan insektisida. Hasil penelitian menunjukkan bahwa, penambahan asam borat sampai 5% tidak berbeda nyata pengaruhnya pada tingkat mortalitas UGB. Peningkatan mortalitas yang berbeda nyata baru terlihat pada perlakuan asam borat 10% (Tabel 4.6). Cisneros et al. (2002) mengemukakan bahwa, perlakuan asam borat saja tidak mengakibatkan kematian pada serangga uji, tetapi akan meningkatkan prevalensi kematian serangga yang terinfeksi virus.
Tabel 4.6. Pengaruh penambahan asam borat terhadap akumulasi mortalitas UGB terinfeksi SeNPV
SeNPVa+ Asam borat (%)
Mortalitas (%)b Hari setelah inokulasi
2 3 4 5 6 0 0 c 15,00 + 8,66 a 25,47 + 8,88 c 52,12 + 5,24 c 67,73 + 9,19 b 0,1 6,87 + 0,19 b 15,82 + 8,76 a 31,86 + 2,33 bc 51,30 + 7,66 c 70,69 + 8,00 ab 1 5,84 + 2,10 b 26,54 + 8,75 a 45,39 + 5,91 a 64,06 + 3,56 b 74,23 + 10,98 ab 5 8,29 + 2,81 ab 28,21 + 10,01 a 44,27 + 10,13 ab 66,54 + 4,95 b 74,02 + 7,66 ab 10 11,23 + 1,83 a 23,56 + 8,69 a 50,54 + 5,37 a 78,34 + 1,46 a 86,48 + 3,62 a a Konsentrasi SeNPV yang digunakan 1,13 x 107POB/ml
bRataan mortalitas terkoreksi (Abort), angka yang diikuti oleh huruf yang sama tidak menunjukkan beda nyata (Uji Duncan, α = 0,05).
Virulensi adalah kemampuan mikroorganisme untuk menghasilkan penyakit yang diekspresikan dalam bentuk median waktu yang dibutuhkan untuk mematikan inang (LT50) atau median konsentrasi yang mematikan (LC50/LD50)(Tanada & Kaya 1993; Hodgson et al. 2001). Nilai LT50 pada perlakuan 1%, 5% dan 10% masing-masing 4,39, 4,33 dan 3,94 hari setelah perlakuan (HSP) berbeda nyata dibandingkan dengan kontrol 5,12 HSP. Hasil ini mengindikasikan bahwa asam borat dapat meningkatkan virulensi SeNPV, semakin tinggi konsentrasi asam borat yang digunakan, semakin tinggi nilai aktifitas relatifnya (AR) (Tabel 4.7). Hasil ini sama dengan penelitian pada Anticarsia gemmatalis NPV (AgNPV) (Morales et al. 1997),
Spodoptera frugiferda NPV (SfNPV) (Cisneros et al. 2002) dan Autographa califor-nica MNPV (AcMNPV) (Jinn et al. 2004).
Tabel 4.7. Nilai LT50dan aktifitas relatif (AR) asam borat pada SeNPV SeNPVa+ Asam borat
(%) LT50(hari) Persamaan regresi AR
b 0 5,12 + 0,14 a y = 14,44x - 23,82 1,00 0,1 4,95 + 0,34 a y = 14,36x - 20,85 1,03 1 4,39 + 0,39 b y = 16,13x - 20,45 1,17 5 4,33 + 0,27 b y = 16,02x - 19,20 1,18 10 3,94 + 0,17 b y = 18,87x - 24,37 1,30
a Konsentrasi SeNPV yang digunakan 1,13 x 107POB/ml b Nilai aktifitas relatif (AR) adalah LT
50kontrol dibagi LT50perlakuan
Da Silva-Cruz et al. (2010) menyatakan bahwa asam borat dapat menginduksi proses kematian sel-sel mesenteron serangga. Hasil penelitian Kilani-Morakchi et al. (2009) menunjukkan bahwa asam borat yang tertelan dapat mengganggu struktur mesenteron kecoa. Sehingga menurut See et al. (2010) asam borat merupakan “racun
perut” yang biasa digunakan untuk mengendalikan serangga urban, seperti kecoa,
semut dan rayap. Oleh karena itu, pengaruh asam borat dalam meningkatkan virulensi SeNPV diduga karena bahan ini mampu mendegradasi matrik peritrofik saluran pencernaan bagian tengah (midgut) serangga, sehingga mengurangi proteksi serangga dari infeksi patogen. Apabila matrik peritrofik hancur, maka virion yang telah terlepas dari badan oklusi akan mudah masuk menginfeksi sel epithelium mesenteron yang kemudian menginfeksi hemolimf dan sel-sel inang lainnya. Guo et al. (2007) melaporkan bahwa aktifitas biologi NPV meningkat ketika dipadukan dengan insektisida chlorfluazuron yang diketahui dapat menghancurkan matrik peritrofik.
Meskipun asam borat secara alami merupakan salah satu nutrisi penting bagi makhluk hidup, akan tetapi pada konsentrasi tinggi dapat bersifat toksik ( Gentz & Grace 2006). Oleh karena itu penggunaannya harus dibatasi pada konsentrasi yang tidak berdampak negatif pada lingkungan dan organisme bukan sasaran. Dari
penelitian ini, konsentrasi asam borat yang dianjurkan adalah antara 1 sampai 5% saja, sebab hasil review yang dilakukan oleh Eisler (1990) menunjukkan bahwa penggunaan asam borat pada konsentrasi lebih dari 10% dapat mematikan lebih dari 40% kecoa (Blatella germanica), 50% lebah madu (Apis mellifera), lebih dari 50% lalat rumah (Musca domestica).
Kesimpulan
Ketiga jenis pakan yang digunakan tidak berpengaruh terhadap kinerja SeNPV dalam mematikan UGB di laboratorium. Kecap dan sukrosa masing-masing 5% berfungsi sebagai phagostimulant yang efektif dapat meningkatkan virulensi SeNPV. Penambahan asam borat pada suspensi SeNPV mempercepat kematian UGB. Untuk menghindari dampak negatif dari penggunaan asam borat, maka konsentrasi asam borat yang dianjurkan adalah antara 1 sampai 5%.
Daftar Pustaka
Abbott WS. 1925. A method of computing the effectiveness of insecticide. J Econ Entomol 18; 265-267.
Ahmad I, Heriyadi S and Anggraeni T. 2001. Nutrient self selection by the armuworm Spodoptera exempta Walker (Lepidoptera: Noctuidae) larvae. Pakistan J. Biol Sci. 4;6: 684-687.
Ahmad I, Kamal M. 2001. Consumption and utilization of complete defined diets covarious carbohydrates by Spodoptera exempta (Lepidoptera; Noctuidae). Biota 6;3: 99-104.
Arifin M. 1988. Pengaruh konsentrasi dan volume nuclear polyhidrosis virus terhadap kematian ulatgrayak kedelai (Spodoptera litura). J Penelitian Pertanian 8;1. Arifin M, Nuzullianti D. 1999. Effectivity of nuclear-polyhedrosis virus (NPV)
bioinsecticide on various feeding stimulant concentrations to soybean cutworm (Spodoptera litura (F.). Seminar Nasional Pertanian Organik, Palembang (Indonesia), 30 Oct 1999.
Arifin M. 2006. Kompatibilitas SlNPV dengan HaNPV dalam pengendalian ulatgrayak dan ulat pemakan polong kedelai. Pengaruh konsentrasi dan volume nuclear polyhidrosis virus terhadap kematian ulatgrayak kedelai (Spolian doptera litura). J Penelitian Pertanian 8;1.
Armenta R, Mertinez AM, Chapman JW, Magallanes R, Goulson D, Caballero P, Cave RD, Cisneros J, Valle J, Castillejos V, Penagos DI, Garcia LF, William T. 2003. Impact of a nucleopolyhedrovirus bioinsecticide and selected synthetic insecticides on the abundance of insect natural enemies on maize in Southern Mexico. J Econ Entomol 96;3: 649-661.
Azidah AA, Sofian-Azirun M. 2006. Fecundity study of Spodoptera exigua (Hubner) (Lepidoptera: Noctuidae) on various host plants. J Entomol 3;3: 261-266. Bonning BC, Hammock BD. 1996. Development of recombinant baculoviruses for
insect control. Annu Rev Entomol 41: 191-210.
Castillejos V, Trujillo J, Ortega LD, Santizo JA, Cisneros J, Penagos DI, Valle J, Williams T. 2002. Granular phagostimulant nucleopolyhedrovirus formulations for control of Spodoptera frugiperda in maize. Biol Contr 24: 300-310.
Chaudhari S. 1992. Formulation of nuclear polyhedrosis virus of Spodoptera litura with boric acid. Ind J Entomol 54: 202-206.
Chapman RF. 1998. The Insects: Structure and Function. 4th Edition. Cambridge University Press. UK. 770 pp.
Cisneros J, Perez JA, Penagos DI, Ruiz J, Goulson D, Caballero P, Cave RD, Williams T. 2002. Formulation of a nucleopolyhedrovirus with boric acid for control of Spodoptera frugiperda (Lepidoptera: Noctuidae) in maize. Biol Cont 23: 87-95.
Da Silva-Cruz A, da Silva-Zacarin ECM, Bueno OC, Malaspina O. 2010. Morphological alterations induced by boric acid and fipronil in the midgut of worker honeybee (Apis mellifera L.) larvae.http://bee-life.eu/en/doc/32/. [akses Januari 2011].
Dodin K, Arifin M dan Harnoto. 2001. Kompatibiltas SlNPV dengan ekstrak biji mimba untuk mengendalikan ulatgrayak pada kedelai. Prosiding Seminar Hasil Penelitian Rintisan dan Bioteknologi Tanaman. 343-347.
Dushoff J, Greg D. 2001. Evaluating the risks of engineered viruses: modeling pathogen competition. Ecol Appl 11;6: 1602 – 1609.
Eisler R. 1990. Boron hazards to fish, wildlife and invertebrates: a synoptic review. US Fish Wildl. Serv. Biol Rep 85 (1.20).
Farrar RR Jr, Ridgway RL. 2000. Host plant effects on the activity of selected nuclear polyhedrosis viruses against the corn earworm and beet armyworm (Lepidoptera: Noctuidae). Environ Entomol 29;1: 108-115.
Farrar RR Jr, Shapiro M, Shepard BM. 2005. Enhanced activity of the nucleopolyhedrovirus of the fall armyworm (Lepidoptera: Noctuidae) on Bt-transgenic and nonBt-transgenic sweet corn with a fluorescent brightener and a feeding stimulant. Environ Entomol 34;4: 825-832.
Genc H. 2006. General principles of insect nutritional ecology. Trakya Univ J Sci 7;1: 53-57.
Gentz MC, Grace JK. 2006. A review of boron toxicity in insects with an emphasis on termites. J Agric Urban Entomol 23;4: 201-207.
Goulson D, Hail RS, Williams T, Hirst ML, Vasconcelos SD, Green BM, Carty TM, Cory JS. 1995. Transmission dynamics of a virus in a stage-structured insect population. Ecol 76;2: 392-401.
Guo HF, Fang JC, Liu BS, Wang JP, Zhong WF, Wan FH. 2007. Enhancement of the biological activity of nucleopolyhedrovirus through disruption of the peritrophic matrix of insect larvae by chlorfluazuron. Pest Manag Sci 63: 68-74.
Hodgson DJ, Vanbergen AJ, Watt AD, Hails RS, Cory JS. 2001. Phenotypic variation between naturally co-existing genotypes of a lepidopteran baculovirus. Evolut Ecol Research 3:687-701.
Hunter-Fujita FR, Entwistle RF, Evans HF, Crook NE. 1998. Insect Viruses and Pest Management. John Wiley & Sons, Inc., 605 Third Avenue, New York, USA. 620 p.
Idris AB, Emelia O. 2001. Development and feeding behaviour of Spodoptera exigua L. (Lepidoptera: Noctuidae) on different food plants. J Biol Sci 1;12: 1161-1164.
Jinn TR, Kao SS, Liu CI, Tseng HY, Wu TY. 2004. Boric acid as a synergist of Spodoptera exigua and Anagrapha californica nuclear polyhedrosis virus. Formosan Entomol 24: 173-184.
Kilani-Morakchi S, Aribi N, Soltani N. 2009. Activity of boric acid on German cockroaches: analysis of residues and effects on reproduction. African J Biotech 8;4: 703-708.
Lacey LA, Frutos R, Kaya HK, Vail P. 2001. Insect pathogens as biological control agents: do they have a future?. Biol Cont 21: 230-248.
Lasa R, Williams T, Caballero P. 2009. The attractiveness of phagostimulant formulations of a nucleopolyhedrovirus-based insecticide depends on prior insect diet. J Pest Sci 82: 247-250.
Matsuda K, Kaneko M, Kusaka K, Shishido T, Tamaki Y. 1998. Soyasaponins as feeding stimulants to the oriental clouded yellow larva, Coliaserate poliographus (Lepidoptera: Pieridae). Appl Entomol Zool 33;2: 255-258. Mcfarlane JE. 1985. Nutrition and digestive organs. In. Blum MS. Fundamentals of
Insect Physiology Edited. John Wiley & Sons, Inc. 598 p.
Meade T, Hare JD. 1993. Effects of differential host plant consumption by Spodoptera exigua (Lepidoptera: Noctuidae) on Bacillus thuringiensis efficacy. Environ Entomol 22;2: 432-437.
Mehrvar A, Rabindra RJ, Veenakumari K, Narabenchi GB. 2008. Evaluation of adjuvants for increased of HearNPV against Helicoverpa armigera (Hubner) using suntest machine. J Biol Sci 1-8.
Morales L, Moscardi F, Sosa-Gomez DR, Paro FE, Soldorio IL. 1997. Enhanced activity of Anticarsia gemmatalis Hub. (Lepidoptera: Noctuidae) nuclear polyhedrosis virus by boric acid in the laboratory. An. Soc. Entomol. Brasil 26:1; 115-120.
Samsudin. 2001. Keefektifan kombinasi Anagrapha falcifera NPV (AfNPV) dengan Spodoptera litura NPV (SlNPV) untuk mengendalikan ulatgrayak pada kedelai. Prosiding Seminar Hasil Penelitian Rintisan dan Bioteknologi Tanaman. 348-351.
See AS, Salleh AB, Bakar FA, Yusof NA, Abdulamir AS, Heng LY. 2010. Risk and health effect of boric acid. American J Appl Sci 7;5: 620-627.
Shapiro M, Bell RA. 1982. Enhanced effectiveness of Lymantria dispar (Lepidoptera; Lymantriidae) nuclear polyhedrosis virus formulated with boric acid. Ann Entomol Soc Am 75: 346-349.
Shapiro M, Shepard BM. 2006. The gypsy moth (Lepidoptera: Lymantriidae) nucleopolyhedrovirus as a synergist for baculoviruses against beet armyworm, fall armyworm and corn earworm (Lepidoptera: Noctuidae). J Agric Urban Entomol 23;4: 243-251.
Suhas Y, Gopali JB, Patil BV, Lingappa S. 2009. Effect of different adjuvants in enhancing the efficacy of HaNPV against Helicoperva armigera (Hubner) in pigeonpea. Karnataka J Agric Sci 22;3: 502-503.
Sutherland ORW. 1977. Plant chemicals influencing insect behaviour. The New Zealand Entomol 6;3: 222-228.
Tanada Y, Kaya HK. 1993. Insect Pathology. Academic Press. San Diego. California. p. 78-98.
Trang TTK, Chaudhari S. 2002. Bioassay of nuclear polyhedrosis virus (npv) and in combination with insecticide on Spodoptera litura (Fab). Omonrice 10: 45-53. Treacy MF. 1999. Recombinant baculoviruses. In Hall FR & Julius JM. 1999. Biopesticides Use and Delivery. Humana Press. Totowa, New Jersey. 321 – 340.
Vasconcelos SD, Cory JS, Speight MR, Williams T. 2002. Host range structure and baculovirus transmission in Mamestra brassicae L. (Lepidoptera: Noctuidae) larvae: a laboratory examination of small scale epizootics. Neotrop Entomol 31; 3:391-396.
Young SY. 1989. Problems associated with the production and use of viral pesticides. Mem Inst Oswaldo Cruz 84; III: 67-73.