BAB 2
TINJAUAN PUSTAKA
2.1 Diabetes Melitus 2.1.1 Definisi
Diabetes melitus (DM) merupakan suatu kelompok penyakit metabolik yang ditandai dengan hiperglikemia kronik dengan gangguan metabolisme karbohidrat, lemak dan protein, yang terjadi karena kelainan sekresi insulin, disfungsi insulin atau keduanya. Hiperglikemia kronik pada DM berhubungan dengan kerusakan jangka panjang, disfungsi atau kegagalan beberapa organ tubuh, terutama mata, ginjal, saraf, jantung, dan pembuluh darah (WHO, 1999; Purnamasari, 2009; Amod et al., 2012; ADA, 2014).
2.1.2 Klasifikasi
DM dapat diklasifikasikan menjadi empat kategori klinis, yaitu (PERKENI, 2011; ADA, 2014);
1. Diabetes melitus tipe 1, ditandai dengan adanya defisiensi insulin absolut akibat destruksi sel pankreas yang dapat disebabkan oleh autoimun maupun idiopatik.
2. Diabetes melitus tipe 2, ditandai dengan adanya defisiensi insulin relatif atau resistensi insulin.
3. Diabetes melitus tipe lain.
4. Diabetes melitus pada kehamilan (gestasional).
2.1.3 Gejala klinis dan diagnosis DM
diagnosis, namun apabila tidak ditemukan gejala khas DM, maka diperlukan dua kali pemeriksaan glukosa darah abnormal.
Tabel 2.1 Kriteria diagnosis Diabetes Melitus
No Kriteria Diagnosis
1 Gejala klasik DM + glukosa plasma sewaktu ≥ β00 mg/dl (11,1 mmol/L)
Glukosa plasma sewaktu merupakan hasil pemeriksaan sesaat pada suatu hari tanpa memperhatikan waktu makan terakhir. 2 Atau, Gejala klasik DM + glukosa plasma puasa ≥ 1β6 mg/dl (7,0
mmol/L)
Puasa diartikan pasien tidak mendapatkan kalori tambahan sedikitnya 8 jam.
3 Glukosa plasma 2 jam pada TIGO (Test Toleransi Glukosa Oral) ≥ β00 mg/dl (11,1 mmol/L)
TTGO dilakukan dengan standar WHO, menggunakan beban glukosa yang setara dengan 75 gram glukosa anhidrus yang dilarutkan ke dalam air
Hasil pemeriksaan glukosa darah 2 jam setelah pembebanan dibagi menjadi 3 yaitu (Purnamasari, 2009) :
1. < 140 mg/dl Normal
2. 140 - <200 mg/dl Toleransi Glukosa Terganggu 3. ≥ β00 mg/dl DM
2.2 Anatomi dan Histologi Koklea
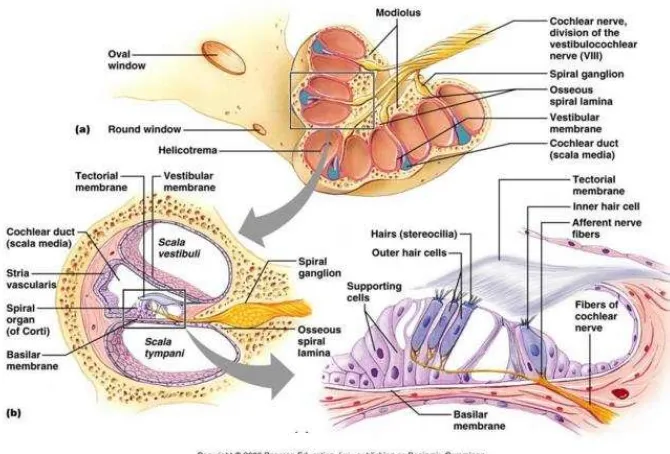
Koklea merupakan saluran tulang yang menyerupai cangkang siput dan bergulung 2,5 sampai 3 kali putaran, panjangnya kurang lebih 35 mm dengan sumbu panjang dari arah anterior ke posterior. Pusat koklea disebut modiolus, dan terletak di depan vestibulum. Koklea bersama dengan organ vestibuler berada dalam tulang temporal, dan merupakan salah satu tulang paling keras dalam tubuh manusia. Koklea bersama organ vestibuler sering disebut dengan labirin (Moller, 2006; Pawlowsky, 2004; Weber & Khariwala, 2014).
segitiga tersebut dikenal dengan nama membran basilaris yang menjadi dasar dari organ korti (Gambar 2.1) (Moller, 2006; Weber & Khariwala, 2014).
Gambar 2.1 Anatomi Koklea dan Organ Korti
Koklea pada telinga dalam mengandung sel-sel yang berperan terhadap persepsi suara. Koklea terdiri dari labirin tulang, dimana didalamnya terdapat labirin membran. Termasuk di dalam labirin tulang adalah kapsul otik yang merupakan batas luar dari koklea dan modiolus yaitu tabung tulang yang membentuk sumbu pusat koklea dan mengandung serat saraf auditori dan sel-sel ganglionnya. Stria vaskularis dan ligamentum spiralis terdapat dekat dengan tulang sepanjang dinding lateral koklea. Organ Corti, yang mengandung sel rambut (3 sel rambut luar dan 1 sel rambut dalam) sebagai sel sensoris dan sel penyokong, berbentuk spiral pada membran basilaris (Nagashima et al., 2005).
terdapat kanalis Rosental yang berisi ganglion spiralis dan berhubungan dengan modiolus (Moller, 2003; Guyton & Hall, 2006; Gillespie, 2006). Skala vestibuli dan skala timpani adalah labirin tulang dari koklea yang berisi cairan perilimfe. Skala vestibuli dan skala timpani saling berhubungan di helikotrema pada apeks koklea. Pada bagian basis koklea skala vestibuli berakhir di foramen ovale dan skala timpani pada foramen rotundum. Skala media yang berisikan cairan endolimfe berada di antara skala vestibuli dan skala timpani (Lonsbury, Martin & Luebke, 2003; Moller, 2003; Guyton & Hall, 2006).
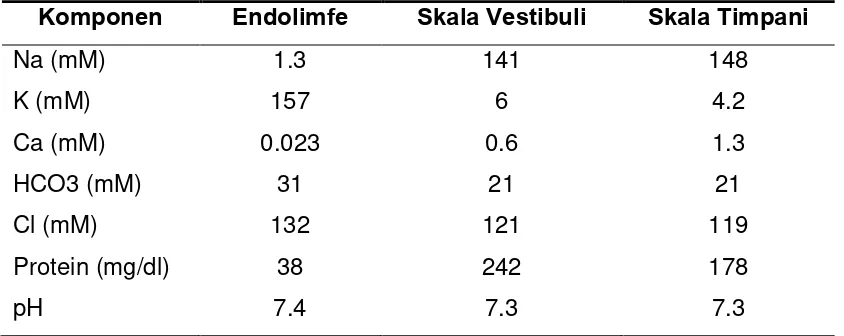
Cairan perilimfe memiliki komposisi ion yang mirip dengan cairan cerebrospinalis (CSF) dan juga mirip dengan cairan ekstraseluler, dengan konsentrasi natrium (Na+) tinggi dan kalium (K+) rendah. Sedangkan pada endolimfe, memiliki komposisi ion yang hampir sama dengan cairan intraseluler yaitu konsentrasi natrium (Na+) rendah dan kalium (K+) yang tinggi (Tabel 2.2.) (Lonsbury, Martin & Luebke, 2003; Gillespie, 2006).
Tabel 2.2 Komposisi Cairan Koklea
Komponen Endolimfe Skala Vestibuli Skala Timpani
Na (mM) 1.3 141 148
K (mM) 157 6 4.2
Ca (mM) 0.023 0.6 1.3
HCO3 (mM) 31 21 21
Cl (mM) 132 121 119
Protein (mg/dl) 38 242 178
pH 7.4 7.3 7.3
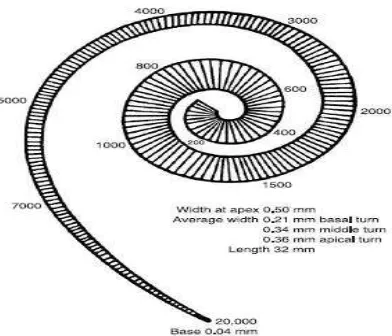
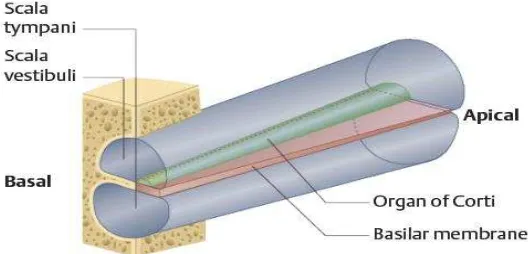
Membran basilaris adalah struktur fibrosa yang berlapis-lapis dari lamina spiral pars osseus ke ligamentum spiralis. Elastisitas membran basilaris bervariasi di sepanjang koklea dari kekakuan dan kelebarannya. Membran basilaris tampak kaku dan sempit di daerah basis koklea dan tampak lebih fleksibel dan luas di daerah apeks koklea (Gambar 2.3) (Moller, 2003; Pawlowsky, 2004; Gillespie, 2006).
Gambar 2.2 Lebar Membran Basilaris dari Basal ke Apeks
Gambar 2.3. Model Membran Basilaris dengan Organ Corti
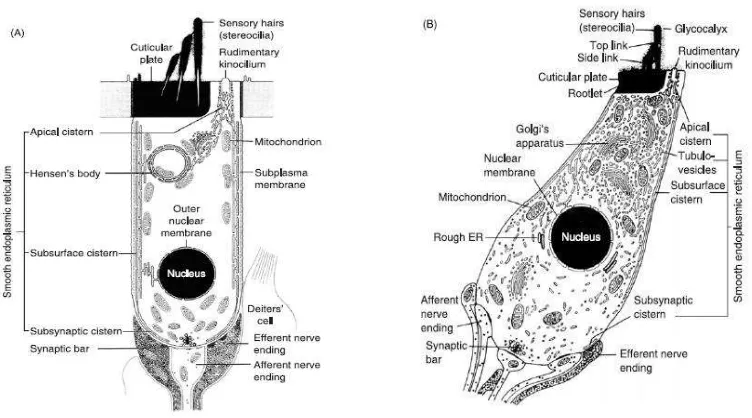
Sel rambut merupakan sel sensoris yang menghasilkan impuls saraf dalam menanggapi getaran membran basilaris. Di organ Corti terdapat 1 deret sel rambut dalam dan 3 deret sel rambut luar. Ada sekitar 4.000 sel rambut dalam dan 12.000 sel rambut luar (Pawlowsky, 2004; Gillespie, 2006). Bentuk dari sel rambut dalam seperti botol dan ujung sarafnya berbentuk piala yang menyelubunginya, sedangkan bentuk dari sel rambut luar seperti silinder dan ujung sarafnya hanya pada basis sel (Gambar 2.4) (Moller, 2003; Pawlowsky, 2004).
Gambar 2.4 Skema Potong Lintang Sel Rambut Luar (A) dan Sel Rambut Dalam (B)
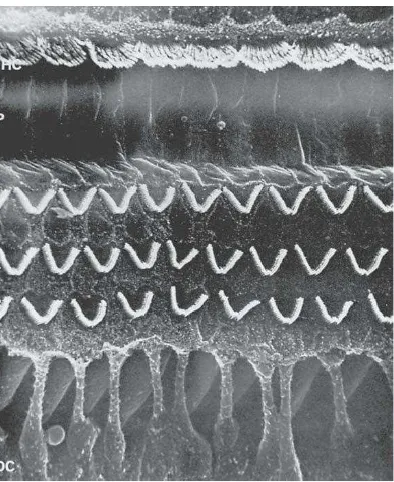
Sel rambut dalam dan luar ini memegang peranan penting pada perubahan energi mekanik menjadi energi listrik. Fungsi sel rambut dalam sebagai mekanoreseptor utama yang mengirimkan sinyal saraf ke neuron pendengaran ganglion spiral dan pusat pendengaran, sedangkan fungsi sel rambut luar adalah meningkatkan atau mempertajam puncak gelombang berjalan dengan meningkatkan aktivitas membran basilaris pada frekuensi tertentu. Peningkatan gerakan ini disebut cochlear amplifier yang memberikan kemampuan sangat baik pada telinga untuk menyeleksi frekuensi, telinga menjadi sensitif dan mampu mendeteksi suara yang lemah (Gillespie, 2006).
Gambar 2.5 Sel Rambut Luar dan Dalam Dilihat dengan Mikroskop Elektron
Pada bagian ujung dari stereosilia terdapat filamen aktin yang terpilin, filamen tersebut nantinya akan dikenal sebagai tip link (Gillespie, 2006). Tip link menghubungkan ujung stereosilia dengan ujung stereosilia yang lain. Bagian basal dari sel rambut diliputi oleh dendrit dari neuron ganglionik spiralis yang terletak pada bagian modiolus (Gillespie, 2006). Selain sel rambut dalam dan luar, komponen utama organ Corti yang lain adalah 3 lapis penyokong (sel Deiters, Hensen, Claudius). Membran tektorial dan kompleks lamina retikularis lempeng kutikular (Pawlowsky, 2004). Sel-sel pendukung yang mengelilingi sel rambut luar adalah sel Deiters dan sel pilar luar. Sel pilar luar berada di sisi modiolar dari sel rambut luar baris pertama dan diantara sel rambut luar baris pertama dengan kedua. Sel Deiters berada diantara sel rambut luar baris dua dengan tiga dan di sisi lateral dari sel rambut luar baris tiga. Gabungan dari sel rambut luar dengan sel Deiters dan sel pilar luar menciptakan sebuah penghalang yang kuat antara endolimfe dan perilimfe (Moller, 2003; Pawlowsky, 2004; Moller, 2005; Gillespie, 2006).
permukaan lamina retikuler dari organ Corti. Membran tektoria kontak langsung dengan sel rambut luar. Sedangkan untuk sel rambut dalam tidak berkontak secara langsung dengan membran tektorial (Moller, 2003).
2.3 Fisiologi Pendengaran
Getaran suara dihantarkan lewat liang telinga dan telinga tengah ke telinga dalam melalui footplate dari stapes, menimbulkan suatu gelombang yang berjalan di sepanjang cairan koklea yang akan menggerakkan membran basilaris dan organ Corti. Puncak gelombang yang berjalan di sepanjang membran basilaris yang panjangnya 35 mm tersebut, ditentukan oleh frekuensi gelombang suara. Hal ini berakibat melengkungnya stereosilia, dengan demikian menimbulkan depolarisasi sel rambut dan menciptakan potensial aksi pada serabut-serabut saraf pendengaran yang melekat padanya. Di sinilah gelombang suara mekanis diubah menjadi energi elektrokimia agar dapat ditransmisikan melalui saraf kranialis ke-8 (Moller, 2006; Gacek, 2009).
Serabut-serabut saraf koklearis berjalan menuju inti koklearis dorsalis dan ventralis. Sebagian besar serabut inti melintasi garis tengah dan berjalan naik menuju kolikulus inferior kontralateral, namun sebagian serabut tetap berjalan ipsilateral menuju kompleks olivarius superior. Penyilangan selanjutnya pada inti lemniskus lateralis dan kolikulus inferior. Dari kolikulus inferior jaras pendengaran berlanjut ke korpus genikulatum dan kemudian ke korteks pendengaran (area 39-40) pada lobus temporalis (Weber & Khariwala,2014; Gacek, 2009).
2.4 Gangguan Pendengaran pada Diabetes Melitus
Gangguan pendengaran sensorineural pada penderita DM dikaitkan dengan mikroangiopati pada telinga dalam, degenerasi neuronal dan diabetik ensefalopati, meskipun temuan mengenai hal tersebut masih sering diperdebatkan dan inkonsisten. Gangguan pendengaran pada penderita DM dapat juga merupakan akibat dari kekacauan metabolisme glukosa serta peningkatan stres oksidatif (Kakarlapudi, Sawyer & Staecker, 2003; Aladag et al., 2009).
Angiopati diabetik memiliki karakteristik berupa proliferasi endotelial, akumulasi glukoprotein pada lapisan intima pembuluh darah, dan penebalan membran basal kapiler serta pembuluh - pembuluh darah kecil. Selain itu juga dijumpai penebalan dan fibrosis dinding kapiler serta penyempitan lumen arteri auditorius interna (Maia & de Campos, 2005; Malucelli et.al., 2012). Penelitian mengenai hal ini sudah dilakukan, baik pada hewan coba maupun pada manusia. Penelitian oleh Fukushima et.al (2006) yang mempelajari tulang temporal manusia penderita DM menunjukkan adanya penebalan membran basilaris dan stria vaskularis, terutama pada dinding pembuluh darah stria vaskularis bagian basal selain itu juga didapati hilangnya sel rambut luar yang signifikan.
Penelitian pada hewan coba baik menggunakan obat diabetogenik seperti alloxan dan streptozotocin, atau dengan pankreatektomi total atau subtotal menghasilkan hal yang relatif sama berupa penebalan dinding pembuluh darah modiolus (Costa, 1967), penebalan dinding pembuluh darah stria vaskularis (Smith et al., 1995), dan hilangnya sel rambut luar (Raynor et al., 1995; Triana et al., 1991).
yang menebal, pengurangan aliran darah karena penyempitan pembuluh darah, dan degenerasi sekunder saraf vestibulokoklear yang menyebabkan neuropati (Malucelli et.al., 2012).
Angiopati terjadi terutama di stria vaskularis dan pada ligamentum spiralis. Studi pada tikus yang di DM-kan menunjukkan bahwa gangguan pendengaran disebabkan terutama oleh pengurangan jumlah sel ganglion spiral dan yang kedua oleh edema di stria vaskularis. Beberapa penulis, juga berpendapat bahwa gangguan pendengaran terjadi karena keterlibatan jalur pendengaran sentral dan bukan karena angiopati koklea. Adanya atrofi neuron ganglion spiralis dan demielinasi saraf vestibulocochlear juga didapatkan pada pasien diabetes. Hal ini menunjukkan bahwa demielinisasi juga merupakan bentuk cedera awal pada saraf perifer penderita DM. Pengamatan melalui mikroskop menunjukkan demielinasi nervus auditorius akibat degenerasi selubung myelin dengan perubahan minor pada akson dan fibrosis perineurium; atrofi parah pada ganglion spiral dengan hilangnya sel di koklea, dan pengurangan pada jumlah serabut saraf pada lamina spiralis. Temuan lainnya berupa pengurangan jumlah sel ganglion dalam nukleus dorsal dan ventral koklea, kehilangan sel ganglion pada nukleus olivarius superior, kolikulus inferior, dan korpus genikulatum medial. Tak ada perubahan langsung yang berkaitan dengan DM terlihat pada sentral pendengaran di lobus temporal (Malucelli et.al., 2012).
Fukushima et al. (2006) meneliti efek DM terhadap koklea manusia dan menyimpulkan bahwa pasien DM tipe 1 mungkin mengalami mikroangiopati koklea dan degenerasi dinding lateral koklea serta sel-sel rambut. Metabolisme glukosa secara signifikan mempengaruhi telinga dalam. Baik kadar gula yang rendah maupun tinggi dapat mempengaruhi fungsi telinga dalam. Pasien dengan gangguan metabolisme glukosa mungkin memiliki gejala gangguan pendengaran, vestibular, atau campuran keduanya.
perubahan kecil kadar gula mempengaruhi fungsi telinga dalam. Gangguan metabolisme telinga dalam, baik akibat pelepasan insulin oleh pankreas atau perubahan reseptor membran sel, cenderung mengakibatkan pergeseran kalium dari endolimfe ke perilimfe dan sebaliknya pada natrium, dimana mekanisme tersebut memicu timbulnya vertigo, tinitus, dan gangguan pendengaran (Raynor et al., 1995; Triana et al., 1991).
Penurunan pendengaran terutama terjadi pada frekuensi tinggi. Hal ini kemungkinan berkaitan dengan kurangnya glikogen jaringan sebagai sumber energi pada penderita DM. Proses transduksi pada organ korti
2. Sel-sel rambut di daerah basal lebih panjang sehingga untuk dapat meneruskan rangsangan ke serabut-serabut saraf memerlukan energi lebih besar.
3. Potensial endolimfatik pada bagian basal lebih tinggi sehingga memerlukan energi lebih banyak.
4. Skala timpani pada bagian basal lebih besar sehingga kebutuhan akan sumber energi eksternal (glukosa) dan oksigen lebih besar. Meskipun sel sel rambut dapat menggunakan substrat selain glukosa (seperti glutamate, piruvat, atau fumarat) untuk mempertahankan potensial endolimfatik, namun glukosa merupakan substrat yang paling efektif.
2.5 Respon Stres Oksidatif Seluler
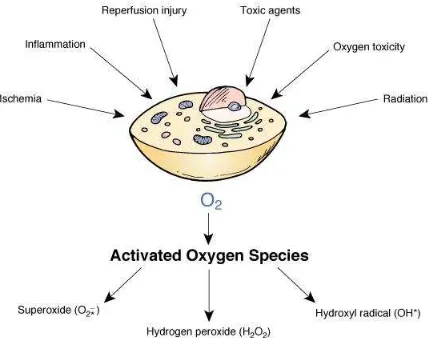
sangat reaktif serta cenderung melepaskan atau menerima elektron dari jaringan sekitarnya (Sarma, Mallick & Ghosh, 2010). Di dalam tubuh organisme, pembentukan radikal bebas dapat berasal dari metabolisme molekul oksigen. Reaksi-reaksi metabolisme pada umumnya merupakan reaksi oksidasi reduksi. Oksidasi adalah reaksi yang melepaskan elektron, sedangkan reduksi adalah reaksi yang menerima elektron (Sies, 1997). Selain proses metabolisme, radikal bebas juga dapat berasal dari faktor eksternal lainnya (Gambar 2.6).
Gambar 2.6. Sumber Radikal Bebas
Dalam metabolisme aerobik, radikal bebas terpenting yang terdapat dalam tubuh adalah derivat oksigen atau oksiradikal atau yang disebut
juga dengan Reactive Oxygen Species (ROS). Oksigen memiliki sifat yang unik, yakni lebih mudah bereaksi dengan melepaskan 1 elektron daripada
2 elektron. Oksigen dapat direduksi sempurna menjadi air dalam
mitokondria melalui 4 tahap reaksi penambahan 1 elektron sebagai berikut
(Granot & Kohen, 2004):
O2 + e- O2•- (radikal anion superoksida)
O2•- + e- + 2H+ H2O2 (hidrogen peroksida)
H2O2 + e- + H+ H2O + OH• (radikal hidroksil) βH• + e- + H+ H
2O (air)
Apabila oksigen hanya tereduksi sebagian, maka terbentuklah radikal
radikal anion superoksida (O2•-), hidrogen peroksida (H2O2), radikal hidroksil (OH•), radikal peroksil (ROO•-), dan lain-lain (Evans & Halliwell, 1999).
Ion-ion logam diketahui dapat mengkatalis reaksi pembentukan radikal
bebas. Ion-ion logam tersebut misalnya Fe, Cu, Mn, Cr, Ni, V, Zn dan Al.
Proses oksidasi yang dikatalisasi oleh ion-ion logam melalui 2 mekanisme
yaitu reaksi ion-ion logam dengan hidroperoksida atau dengan molekul
lipid (Reische, et al., 2008).
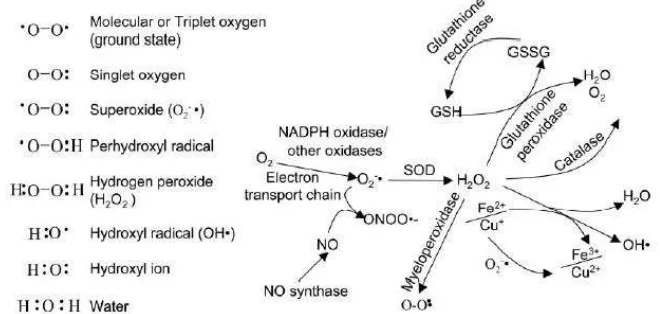
ROS dimetabolisme melalui reaksi reduksi-oksidasi seluler dan
dibentuk secara alamiah sebagai produk sampingan dalam proses
metabolik aerobik normal serta dinetralisir oleh scavenger enzimatik berupa antioksidan endogen alamiah dalam tubuh, meliputi superoxide dismutase (SOD), catalase (CAT) dan glutathione peroxidase (GPx), yang berguna sebagai mekanisme proteksi terhadap produksi ROS (Evans &
Halliwell, 1999). Berbagai molekul ROS serta reaksi pembentukan dan
detoksifikasinya dapat dilihat pada gambar 2.7 berikut:
Gambar 2.7 Molekul ROS serta Reaksi Formasi dan Detoksifikasinya
Dalam keadaan normal, ROS berada dalam keadaan seimbang
dengan antioksidan endogen alamiah tubuh. Ketidakseimbangan antara
kadar antioksidan dan ROS maupun ketidakmampuan antioksidan untuk
menghambat produksi ROS berlebih akan menyebabkan terjadinya stres
oksidatif yang mampu merusak sel (Sies, 1997). ROS dihasilkan dalam
berguna sebagai molekul sinyalisasi untuk fungsi seluler normal guna
mengendalikan homeostasis sel dan jaringan, pembelahan, migrasi dan
kontraksi sel serta produksi mediator-mediator (Evans & Halliwell, 1999;
Le Prell, et al., 2007; Poirrier, et al., 2010; Uchida, et al., 2011; Rewerska,
et al., 2013).
Radikal bebas dianggap berkontribusi terhadap kejadian berbagai
penyakit termasuk penyakit Alzheimer (Christen, 2000), penyakit
Parkinson (Wood-Kaczmar, Gandhi & Wood, 2006), diabetes (Giugliano,
Ceriello & Paolisso, 1996; Davi, Falco & Patrono, 2005), artritis reumatoid
(Hitchon & El-Gabalawy, 2004), dan penyakit motor neuron neurogeneratif
(Cookson & Shaw, 1999).
Radikal bebas yang menyebabkan kerusakan oksidatif pada DNA
diketahui dapat menyebabkan terjadinya kanker. Beberapa enzim
antioksidan seperti SOD, CAT, GPx, GR, GST, dan lain-lain mampu
melindungi DNA dari stres oksidatif. Terdapatnya polimorfisme pada
enzim-enzim ini berhubungan dengan kerusakan DNA dan kemudian
risiko individu terhadap kerentanan terjadinya kanker (Khan, et al., 2010).
Mekanisme pertahanan lini pertama terhadap ROS adalah
menghilangkan ROS atau mengubahnya menjadi radikal bebas yang
kurang toksik. Hal ini diperankan oleh enzim SOD yang mampu mengubah
radikal anion superoksida (O2•-) menjadi hidrogen peroksida (H2O2)
dengan proses dismutasi. Dismutasi merupakan istilah yang mengacu
kepada tipe reaksi khusus dimana 2 reaksi yang sama namun berlawanan
terjadi pada 2 molekul yang terpisah. Enzim SOD mampu mengambil 2
molekul radikal anion superoksida (O2•-) lalu melepaskan elektron ekstra
pada salah 1 molekul dan menempatkannya pada molekul lainnya,
sehingga jumlah elektron yang dimiliki oleh salah 1 molekul menjadi
berkurang lalu membentuk molekul oksigen normal, sedangkan molekul
lainnya memiliki elektron ekstra. Molekul yang memiliki elektron ekstra
kemudian secara cepat mengambil 2 ion hidrogen untuk membentuk
hidrogen peroksida (H2O2) (Evans & Halliwell, 1999; Goodsell, 2007).
(H2O) dan oksigen (O2) oleh CAT. Enzim GPx akan turut membantu CAT
dalam mengkonversi hidrogen peroksida (H2O2) dan glutathione tereduksi
(GSH) menjadi molekul air (H2O) serta glutathione teroksidasi (GSSG).
Proses konversi yang dilakukan oleh CAT dan GPx ini dianggap sebagai
mekanisme pertahanan lini kedua. Guna menyelesaikan siklus reaksinya,
glutathione reductase (GR) kemudian akan mereduksi glutathione
teroksidasi (GSSG) dengan bantuan NADPH dan ion hidrogen menjadi
glutathione tereduksi (GSH) dan NADP+. Rangkaian reaksi tersebut di atas dapat dilihat pada reaksi kimia dan Gambar 2.12 di bawah ini:
SOD : 2O2•- + 2H+ O2 + H2O2
tubuh, ditemukan hampir pada semua sel aerobik dan cairan ekstraseluler,
yang merupakan enzim antioksidan endogen terbanyak di dalam tubuh
dan sebagian besar terdapat pada organ hati. SOD termasuk ke dalam
famili enzim metalloenzyme dengan ramifikasi (percabangan) yang meluas, memiliki gugus prostetik yang berbeda-beda, lokasi intraseluler
yang bervariasi dan heterogenitas jaringan yang besar (Cayuela, 1995;
Nurhayati, Kisnanto & Syaifudin, 2011; Johnson & Giulivi, 2005).
SOD merupakan kelas enzim yang berkaitan erat dengan katalisis
pemecahan 2 molekul radikal anion superoksida (O2•-) menjadi hidrogen
peroksida (H2O2) dan oksigen (O2) dengan kecepatan 104 kali lebih tinggi
dibandingkan proses dismutasi spontan pada pH fisiologis (Bannister,
Bannister & Rotilio, 1987; Cayuela, 1995; Tarhan & Tuzmen, 1998; Zelko,
Mariani & Folz, 2002).
SOD pertama kali diisolasi oleh Mann dan Keilin pada tahun 1938 yang
melakukan pemurnian terhadap protein yang terdapat pada darah dan
fungsi yang tidak diketahui dan disebut sebagai eritrocuprein atau
hemocuprein atau sitocuprein. Temuan radikal anion superoksida (O2•-)
sebagai substrat SOD pertama kali diteliti oleh Linus Pauling pada tahun
1930-an. Kemudian, Brewer (1967) menamakan SOD dengan indofenol
oksidase atau trazolium oksidase saat melakukan analisis protein SOD
dengan gel pati menggunakan teknik fenazin-tetrazolium. Reaksi kimia
dan peranan katalitik SOD terhadap proses dismutasi radikal anion
superoksida (O2•-) menjadi hidrogen peroksida (H2O2) dan oksigen (O2)
pertama kali ditemukan oleh Joe McCord dan Irwin Fridovich pada tahun
1969 (McCord & Fridovich, 1988; Lucchetti-Miganeh, et al., 2011).
Pengukuran kandungan enzim antioksidan SOD merupakan cara untuk
mengetahui kondisi pertahanan sel terhadap radikal bebas. Aktivitas SOD
bervariasi pada beberapa organ. Aktivitas SOD tertinggi terdapat pada
organ hati, diikuti kelenjar adrenal, ginjal, darah, limpa, pankreas, otak,
paru-paru, usus, ovarium, dan timus. Selain itu, SOD memiliki kecepatan
efisiensi katalitik terbesar dibandingkan enzim-enzim lainnya (Halliwell &
Gutteridge, 1999).
Enzim SOD murni dalam fase kromatografi kolom mempunyai 9
macam asam amino esensial yang dapat ditentukan komposisinya
menggunakan Kromatografi Cair Kinerja Tinggi (HPLC). Fraksi yang murni
mempunyai kadar tirosin dan lisin yang tinggi (163 µmol/ml atau 14.64%
dari total asam amino yang terdeteksi). Namun, asam amino serin, glisin,
alanin, prolin, metionin, sistein, isoleusin dan fenilalanin tidak terdeteksi.
Komposisi asam amino ini dapat berbeda dengan perbedaan sumber
SOD maupun asupan yang diterima oleh sumber, karena asam amino
yang terbentuk di dalam sel hidup sesuai dengan nutrisi yang terdapat
pada sel tersebut. Penurunan aktivitas enzim SOD lebih dari 47% terjadi
pada suhu di atas 50oC. Hal ini menunjukkan bahwa ekstrak enzim SOD
hanya dapat dipergunakan pada suhu dingin untuk menjaga stabilitas
SOD yang terkandung di dalamnya. Enzim SOD paling stabil pada pH 7.5
SOD memiliki banyak isoform yang diklasifikasikan berdasarkan
kofaktor ion logam sisi aktif yang mempengaruhi aktivitasnya dan
distribusinya dalam tubuh, yaitu seperti tembaga (Cu), seng (Zn), mangan
(Mn), besi (Fe) atau nikel (Ni). Perbandingan persentase logam dalam
ekstrak murni dapat menunjukkan jenis kofaktor ion logam yang dominan
dalam SOD (Bannister, Bannister & Rotilio, 1987).
Saat ini, 3 isoform SOD yang berbeda telah diidentifikasi pada
mamalia, dan struktur genom, cDNA, dan proteinnya telah dideskripsikan,
yakni (Zelko, Mariani & Folz, 2002; Miller, 2004; Landis & Tower, 2005;
Nozik-Grayck, Suliman & Piantadosi, 2005; Culotta, Yang & O'Halloran,
2006; Kirsh, 2006; Winarsi, 2007; Miao & St. Clair, 2009; Miller, 2012):
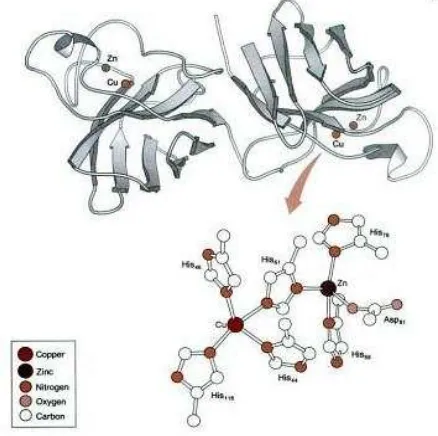
1. Copper/zinc superoxide dismutase [Cu/Zn-SOD (SOD1)]
Cu/Zn-SOD (SOD1) merupakan enzim SOD pertama yang ditemukan
dan berbentuk homodimer, terdapat dalam sitoplasma, kompartemen
nukleus, lisosom dan ruang mitokondria bagian luar serta mengandung
ion logam Cu2+ dan Zn2+ pada sisi aktifnya. Ion logam Cu2+ berperan
aktif secara katalitik, sedangkan ion logam Zn2+ berperan di dalam
menjaga stabilitas enzim tersebut. Cu/Zn-SOD (SOD1) memiliki massa
molekul sekitar 32.000 Da (struktur kimia dapat dilihat pada Gambar
2.13).
2. Manganese superoxide dismutase [Mn-SOD (SOD2)]
Mn-SOD (SOD2) merupakan enzim SOD kedua yang ditemukan dan
berbentuk homotetramer dan mengandung leader peptide yang mampu mengarahkan enzim ini secara khusus ke ruang mitokondria bagian
dalam dan peroksisom, serta mengandung ion logam Mn2+ pada sisi
aktifnya. Mn-SOD (SOD2) memiliki massa molekul sekitar 23.000 Da
(struktur kimia dapat dilihat pada Gambar 2.14).
3. Extracelular superoxide dismutase [EC-SOD (SOD3)]
EC-SOD (SOD3) pertama kali ditemukan oleh Marklund, et al. (1982),
merupakan suatu glikoprotein hidrofobik berbentuk homotetramer dan
asites, cairan sinovial, cairan serebrospinal, dan lain-lain), matriks
selular hati, plasenta dan paru, dengan struktur yang mirip dengan
Cu/Zn-SOD karena mengandung ion logam Cu2+ serta Zn2+ pada sisi
aktifnya. EC-SOD (SOD3) memiliki massa molekul sekitar 135.000 Da
dengan afinitas yang tinggi untuk heparin, heparin sulfat lainnya dan
kolagen tipe I, serta bersifat sensitif terhadap senyawa sianida.
Gambar 2.8 Struktur Kimia Cu/Zn-SOD (SOD1)
Gambar 2.9 Struktur Kimia Mn-SOD (SOD2)
nickel superoxide dismutase (Ni-SOD) yang mengandung logam Ni pada sisi aktifnya. Namun, Fe-SOD ditemukan pada organisme prokariotik dan
bagian dari kloroplas tumbuhan tingkat tinggi, sedangkan Ni-SOD
ditemukan pada organisme prokariotik dan beberapa spesies
Streptomyces (Zelko, Mariani & Folz, 2002; Miller, 2004; Landis & Tower, 2005; Nozik-Grayck, Suliman & Piantadosi, 2005; Kirsh, 2006; Winarsi,
2007).
Proses dismutasi yang dikatalisis oleh SOD sebagai mekanisme
pertahanan lini pertama terhadap radikal anion superoksida (O2•-) dapat
dilihat pada reaksi di bawah ini:
M(n+1)+-SOD + O2•- Mn+-SOD + O2
Mn+-SOD + O2•- + 2H+ M(n+1)+-SOD + H2O2
Keterangan:
M dapat berupa ion logam Cu (n = 2); Mn (n = 2); Fe (n = 2); Ni (n = 2)
Mekanisme yang terjadi pada reaksi yang dikatalisis oleh SOD di atas
disebut juga sebagai ping-pong mechanism atau double displacement reaction. Ciri khas dari reaksi ini adalah terdapatnya substituted enzyme intermediate dimana enzim tersebut mengalami modifikasi secara sementara. Dalam reaksi yang dikatalisis oleh SOD, 1 molekul radikal
anion superoksida (O2•-) akan berikatan dengan enzim M(n+1)+-SOD lalu
memodifikasi enzim tersebut dengan cara mentransfer gugus kimianya
terhadap sisi aktif enzim dan kemudian menghasilkan produk pertama.
Selanjutnya, enzim yang telah termodifikasi tersebut akan mengadakan
reaksi terhadap 1 molekul radikal anion superoksida (O2•-) lain sehingga
menghasilkan kembali (meregenerasi) enzim dengan keadaan seperti
sebelumnya beserta dengan produk kedua. Dalam hal ini, M(n+1)+-SOD
berikatan dengan 1 molekul radikal anion superoksida (O2•-) sehingga
M(n+1)+-SOD mengalami modifikasi ion menjadi Mn+-SOD dan
akan berikatan dengan 1 molekul radikal anion superoksida (O2•-) lain
sehingga akan bereaksi dan menghasilkan kembali (meregenerasi) enzim
M(n+1)+-SOD beserta dengan molekul air (H2O2) sebagai produk kedua
(Bravo, et al., 2001; Akerman & Muller, 2003; Miller, 2004; Yang, et al.,
2004; Patel, 2009).
2.6 Pembentukan Reactive Oxygen Species (ROS) pada DM
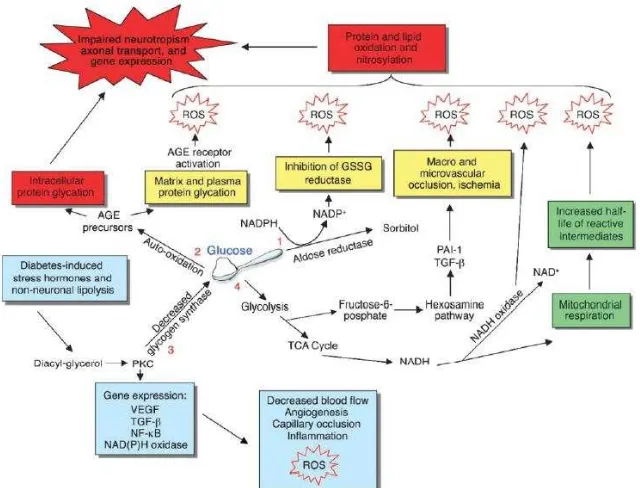
Keadaan hiperglikemia yang berkepanjangan sampai saat ini diakui sebagai faktor utama yang berperan dalam patogenesis terjadinya berbagai komplikasi DM. Hiperglikemia menginduksi sejumlah besar perubahan dalam jaringan pembuluh darah, yang berpotensi menyebabkan aterosklerosis. Beberapa mekanisme utama yang mencakup sebagian besar perubahan patologis yang diamati dalam pembuluh darah penderita DM (Gambar 2.8) yaitu (1) glikosilasi nonenzimatik protein dan lipid, (2) aktivasi protein kinase C (PKC), (3) peningkatan fluks melalui jalur heksosamin (4) peningkatan stres oksidatif, dan (5) peradangan (Aronson, 2008; Vincent et.al, 2004).
Stres oksidatif secara luas dianggap sebagai mekanisme yang mendasari terjadinya aterosklerosis. Kerusakan oksidatif terhadap protein dinding arteri dapat terjadi bahkan pada paparan hiperglikemia jangka pendek. Di antara gejala sisa hiperglikemia, stres oksidatif diperkirakan merupakan mekanisme potensial yang dapat mempercepat terjadinya aterosklerosis. Terdapat hubungan yang erat antara stres oksidatif yang diinduksi hiperglikemia dengan mekanisme kerusakan vaskuler akibat hiperglikemia melalui jalur yang lain seperti pembentukan advanced glycosilation end products (AGEPs), aktivasi PKC, dan peningkatan fluks melalui jalur hexosamine (Gambar 2.9) (Aronson, 2008; Vincent et.al, 2004).
Gambar 2.11 Hubungan Antara Pembentukan Oksidan, Aktivitas Antioksidan dan Stres Oksidatif pada DM
oleh siklus trikarboksilat. Hal ini mengakibatkan tingginya potensial membran mitokondria melalui pompa proton melintasi membran dalam mitokondria. Akibatnya, gradien tegangan yang melintasi membran mitokondria meningkat sampai ambang batas kritis tercapai, dan transpor elektron diblokir. Hal ini meningkatkan waktu paruh koenzim Q (ubiquinone) radikal bebas intermediet, yang mereduksi O2 menjadi superoksida, dan secara nyata meningkatkan produksi superoksida (Aronson, 2008).
Beberapa zat produk advanced glycosylation end products (AGEPs) seperti N(6) – Carboxymethyllysine (CML) dan pentosidin terbentuk dalam reaksi protein dengan glukosa hanya dalam kondisi oksidatif. Dengan demikian, beberapa AGEs yang dihasilkan oleh gabungan proses glikasi dan oksidasi disebut produk glikooksidasi. Setiap struktur AGEPs memiliki mekanisme pembentukannya sendiri dan ketergantungannya sendiri terhadap stres oksidatif. Namun, karena produk glikooksidasi pada protein bersifat ireversibel, disimpulkan bahwa zat tersebut dapat menjadi biomarker akumulasi stres oksidatif pada jaringan. Memang terdapat korelasi kuat antara tingkat produk glikooksidasi dalam kolagen kulit dengan tingkat keparahan komplikasi diabetes yang mengenai retina, ginjal, dan pembuluh darah penyakit (Aronson, 2008).
akibat hiperglikemia dapat menyebabkan aktivasi DAG-PKC dalam jaringan pembuluh darah (Aronson, 2008).
Hal yang juga penting adalah bahwa semua mekanisme disfungsi seluler akibat hiperglikemia tampaknya berhubungan dan semakin meningkat dengan adanya stres oksidatif. Nishikawa et al. telah menunjukkan bahwa over produksi superoksida mitokondria akibat hiperglikemia dapat meningkatkan pembentukan AGEPs, aktivasi PKC, dan aktivitas jalur hexosamine. Penghambatan produksi superoksida melalui overekspresi mangan dismutase (yang dengan cepat mengubah superoksida menjadi H2O2) atau uncoupling protein-1 mencegah kelebihan superoksida akibat hiperglikemia. Secara bersamaan, peningkatan pembentukan AGEPs intraseluler, aktivasi PKC, dan peningkatan pembentukan hexosamin dapat dicegah. Dengan demikian, beberapa mekanisme berkaitan hiperglikemia yang tampaknya tidak saling berhubungan, yang berkontribusi terhadap komplikasi vaskular diabetes, mungkin timbul dari satu proses yaitu kelebihan produksi molekul radikal bebas superoksida (Aronson, 2008).
organik dengan enzim sitokrom P450 pada awalnya menghasilkan anion superoksida (O2-•), yang kemudian secara spontan atau enzimatik (melalui superoksida dismutase) dikonversi ke H2O2. Hidrogen peroksida kemudian direduksi lebih lanjut menjadi radikal hidroksil (OH•) melalui reaksi Fenton dengan adanya ion Cu2+ atau Fe2+. Hidrogen peroksida, merupakan anion superoksida dan radikal hidroksil yang secara umum dikenal sebagai spesies oksigen reaktif (ROS). Secara umum, ROS beracun bagi sel-sel karena kecenderungannya untuk menyebabkan kerusakan makromolekul. Meskipun H2O2 adalah oksidan ringan dan paling kurang reaktif dibandingkan ROS yang lainnya, semua sel aerob dilengkapi dengan berbagai enzim untuk mengeliminasi H2O2 karena H2O2 sangat mudah dikonversi menjadi radikal hidroksil yang sangat reaktif radikal melalui reaksi Fenton.
2.7 Curcuminoid
Curcuminoid adalah kelompok senyawa fenolik yang terkandung dalam rimpang tanaman famili Zingiberaceae antara lain: Curcuma longa syn. Curcuma domestica Val. (kunyit) dan Curcuma xanthorrhiza (temulawak).
Kunyit pertama kali diklasifikasikan secara taksonomi oleh seorang
ilmuwan asal Swedia bernama Carl Linnaeus pada tahun 1753 sebagai
berikut:
Divisi : Spermatophyta
Sub Divisi : Angiospermae
Kelas : Monocotyledoneae
Bangsa : Zingiberales
Suku : Zingiberaceae
Marga : Curcuma
Spesies : Curcuma longa Linnaeus
(Departemen Kesehatan dan Kesejahteraan Sosial RI Badan Penelitian
2.7.1 Sifat kimia dan fisik curcuminoid
Kandungan utama dari curcuminoid adalah curcumin yang memberikan warna kuning. Nama kimia untuk curcumin adalah diferuloylmethane atau
1,6-heptadiene-3,5-dione-1,7-bis (4-hidroksi-3-metoksifenil)-(1E, 6E) dengan rumus molekul C21H20O6 dan Nilai pKa-nya adalah 8.54. Curcumin
mengandung 60-70% karbohidrat, 8.6% protein, 5-10% lemak, 2-7% serat,
3-5% curcuminoid (50-70% curcumin) dan lebih dari 5% minyak esensial dan resin, sisanya mengandung mineral seperti magnesium, besi,
mangan, kalsium, natrium, kalium, timbal, seng, kobalt, aluminium dan
bismuth. Kandungan curcumin di dalam kunyit berkisar 3-4% (Eigner & Schulz, 1999; Joe, Vijaykumar & Lokesh, 2004). Komposisi curcuminoid
sekitar 70% curcumin (curcumin I), 17% demethoxycurcumin (curcumin II), 3% bis-demethoxycurcumin (curcumin III) dan sisanya (10%) disebut dengan cyclocurcumin (curcumin IV) yang memiliki sedikit atau tanpa aktivitas biologis. Curcumin tidak dapat larut dalam air dan eter, tetapi larut dalam etil asetat, etanol, metanol, aseton, asam asetat glasial, dimetilsulfoksida dan benzene. Beberapa peneliti telah membuktikan
sensitivitas curcumin terhadap cahaya, dan sebagai hasilnya
menyarankan bahwa sampel biologis yang mengandung curcumin harus dilindungi dari cahaya. Masalah stabilitas lainnya terjadi ketika curcumin
ditempatkan dalam sistem penyangga fosfat dengan pH 7.2. Dalam
kondisi ini, sebagian besar curcumin (>90%) berdegradasi dalam waktu 30 menit dari penempatan (Araujo & Leon, 2001; Joe, Vijaykumar & Lokesh, 2004; Chattopadhyay, et al., 2004; Sharma, Gescher & Steward, 2005; Aggarwal & Shishodia, 2006; Trujillo, et al., 2013; Yadav, et al., 2013;
Prasad, et al., 2014).
Banyak hasil penelitian menunjukkan bahwa curcumin aman dan tidak toksik bila dikonsumsi oleh manusia. Jumlah curcumin yang aman dikonsumsi oleh manusia adalah 100 mg/hari sedangkan untuk tikus 5 g/hari (Commandeur & Vermeulen, 1996).
Gambar 2.12. Struktur Molekul Komponen Curcuminoid
2.7.2 Target molekuler curcuminoid
Berbagai studi telah berhasil memperlihatkan peranan curcumin dalam memodulasi sejumlah target molekuler, meliputi faktor pertumbuhan,
reseptor faktor pertumbuhan, faktor transkripsi, sitokin, enzim, dan gen
pengatur apoptosis (Gambar 2.22). Meskipun belum diketahui reseptor
asli untuk curcumin, sejumlah molekul tempat melekatnya curcumin telah teridentifikasi, diantaranya serum albumin, 5-LOX, xanthine oxidase,
thioredoxin reductase, zat besi, COX-2, IKK, p-glycoprotein, GST, PKA, PKC, cPK, PhK, autophosphorylation-activated protein kinase, pp60c-src
tyrosine kinase, Ca2+-dependent protein kinase (CDPK), Ca2+-ATPase
retikulum sarkoplasma, reseptor aryl hydrocarbon, sitokrom p450 rat river, Topo II isomerase, reseptor inositol 1,4,5-triphosphate, dan glutathione
Gambar 2.13. Target Molekuler Curcumin
2.7.3 Target penyakit curcumin
Curcumin mempunyai potensi yang baik untuk berbagai penyakit. Pada fase I percobaan klinik mengindikasikan sebanyak 12 g curcumin perhari selama 3 bulan dapat ditoleransi oleh tubuh manusia. Dosis optimum curcumin untuk pengobatan suatu penyakit belum jelas. Data menunjukkan bahwa curcumin memiliki bioavailabilitas yang rendah (Aggarwal, et al., 2006).
mg/kgBB/hari) tidak signifikan secara biologis (Chemoprevention Branch and Agent Development Committee, 1996).
Penelitian mengenai curcumin menunjukkan bahwa toksisitas curcumin tidak signifikan. Pada tikus yang diberi curcumin pada makanannya dengan dosis 0.1-2.0% (0.1-2.7 mmol/kgBB/hari) selama 8 minggu, tidak ada efek pada nafsu makan, peningkatan berat badan, hematologi, kimia serum, atau perubahan histologi saluran pencernaan, hati, limpa dan ginjal yang diperiksa. Penelitian yang serupa, tikus diberikan curcumin hingga 1.000 mg/kgBB/hari per oral selama 3 bulan dan tidak ada efek samping pada pertumbuhan, perilaku, parameter biokimia dan histopatologi (Chemoprevention Branch and Agent Development Committee, 1996).
Laporan ulkus lambung pada tikus yang diberi curcumin sebagai anti inflamasi (ED50 / Effective Dose = 50 mg/kgBB/hari selama 6 hari), curcumin food grade mampu melindungi lambung dari ulkus melalui penurunan asam lambung atau peningkatan sekresi musin (Chemoprevention Branch and Agent Development Committee, 1996). Penelitian pada tikus yang diberi curcumin 600 dan 1.600 mg/kgBB pada usia kehamilan 6-15 hari tidak menimbulkan efek terhadap implantasi, resorpsi dan tidak menimbulkan kematian pada embrio, tidak menimbulkan abnormalitas tulang dan organ dalam. Penelitian jangka panjang pada lebih dari tiga generasi tikus, tidak ada menunjukkan efek teratogenik atau gangguan pada reproduksi (Chemoprevention Branch and Agent Development Committee, 1996).
meneliti peranan curcumin terhadap disfungsi endotelial akibat ROS pada tikus yang diinduksi DM menggunakan dosis curcumin sebesar 300 mg/kgbb, dan mendapatkan hasil yang signifikan dalam hal penurunan produksi ROS pada tikus yang mendapatkan curcumin dibandingkan dengan tikus kontrol.
Dosis curcumin sebagai anti kanker yaitu sampai 10 g/hari (Aggarwal, Kumar & Bharti, 2003). Dosis curcumin sebagai terapi kanker kolorektal yaitu 2.2 g/hari (ekstrak curcuma). Kanker kolorektal stadium akhir diberikan dosis antara 0.45 dan 3.6 g/hari selama empat bulan (Jurenka, 2009; Burgos-Moron, et al., 2010).
Dosis curcumin sebagai anti inflamasi sebanyak 400-600 mg tiga kali sehari (Alter, 2010). Dosis curcumin sebagai anti inflamasi pada tikus 100, 250, 500 dan 1000 mg/kgBB. Daya anti radang minyak atsiri kunyit (senyawa mirip curcumin) 1.2 ml/kgBB secara oral (Salasia, et al., 2002). Ekstrak etanol kunyit dengan berbagai dosis memperlihatkan efek anti inflamasi pada tikus yang diinduksi dengan karagen dimana pada dosis tinggi (1000 mg/kgBB) dapat menekan edema sebesar 78.37%. Pada penelitian tersebut menunjukkan semakin tingginya dosis ekstrak etanol kunyit, jumlah zat aktif yang terkandung di dalamnya semakin tinggi sehingga kemampuannya di dalam menginhibisi edema semakin besar (Rustam, Atmasari & Yanwirasti, 2007).
Konsentrasi perasan air kunyit 30% paling efektif dalam memperbaiki kerusakan sel hati pada mencit (Kardena & Winaya, 2011).
Penelitian dengan mengkombinasikan antibiotik dan curcumin pada sedian piringan diffusion assay dengan dosis curcumin 500 µg setiap piringan menghasilkan peningkatan aktivitas dari antibiotik Cefixime, Cefotaxime, Vancomycin dan Tetrasiklin (Moghaddam, et al., 2009).
Curcumin dapat secara bebas melalui membran sel karena sifat lipofilisitasnya. Namun, disebutkan juga bahwa curcumin memiliki kelarutan pada air yang sangat rendah, yaitu hanya 0.6 µg/ml,
dimetabolisme dengan cepat di hati dan dinding usus, serta rentan
rendahnya bioavailibilitas curcumin, sehingga konsentrasinya dalam darah menjadi kurang optimal untuk mencapai efek terapeutik yang diharapkan
(Naksuriya, et al., 2014). Nilai paruh waktu (T½) curcumin pada pemberian intravena (10 mg/kgBB) pada tikus dilaporkan sekitar 28.1 ± 5.6 jam dan pada pemberian oral (500 mg/kgBB) yaitu sekitar 44.5 ± 7.5 jam (Anand, et al., 2007).
2.8. Penggunaan Streptozotocin untuk Membuat Hewan Coba Model Diabetes Melitus
Streptozotocin (STZ) atau streptozosin dengan nama kimia 2-Deoxy-2 - [[(methylnitrosoamino) - carbonyl] amino] – D – glucopyranose (Gambar
2.17) berasal dari Streptomyces achromogenes, merupakan antibiotik spektrum luas yang juga memiliki sifat antineoplastik. STZ bersifat toksik dan menyebabkan proses autoimun terhadap sel pankreas, sehingga banyak dimanfaatkan untuk membuat hewan coba model diabetes melitus. Secara klinis STZ digunakan pada pengobatan karsinoma pankreas (Deeds et al., 2011; Abeeleh et al., 2009; Akbarzadeh et al., 2007).
Gambar 2.17 Struktur Kimia Streptozotocin
Penggunaan STZ sangat bervariasi dan memberikan hasil yang berbeda-beda, baik dalam hal mortalitas maupun keparahan DM yang dihasilkan. Selain itu, sensitivitas STZ juga sangat bervariasi pada tiap-tiap hewan coba yang digunakan. Berbagai metode dan dosis STZ telah dicoba dan disebutkan dalam literatur, tergantung kepada tingkat keparahan diabetes yang diinginkan sesuai protokol eksperimen tertentu. Penggunaan dosis dapat dibagi menjadi 3, yaitu dosis rendah dengan pemberian berulang, dosis sedang sekali pemberian, dan dosis tingggi sekali pemberian. Induksi dengan injeksi tunggal baik secara intravena maupun intraperitoneal dianggap sebagai teknik yang paling banyak digunakan (Deeds et al., 2011).
Dosis yang digunakan untuk menginduksi DM tipe 1 untuk intravena adalah 40-60 mg/kg, sedangkan dosis intraperitoneal adalah lebih dari 40 mg/kg BB. STZ juga dapat diberikan secara berulang, untuk menginduksi DM tipe 1 yang diperantarai aktivasi sistem imun. Untuk menginduksi DM tipe 2, STZ diberikan intravena atau intraperitoneal dengan dosis 100 mg/kg BB pada tikus yang berumur 2 hari kelahiran, pada 8-10 minggu tikus tersebut mengalami gangguan respon terhadap glukosa dan sensitivitas sel terhadap glukosa. Di lain pihak, sel α dan δ tidak dipengaruhi secara signifikan oleh pemberian streptozotosin pada neonatal tersebut sehingga tidak membawa dampak pada perubahan glukagon dan somatostatin. Patofisiologis tersebut identik pada DM tipe II (Szkudelski, 2001; Jackerott et al., 2006; Tormo et al., 2006).
karena aksi STZ dalam mitokondria. Produksi ATP mitokondria yang terbatas selanjutnya mengakibatkan pengurangan secarea drastis nukleotida sel pankreas (Szkudelski, β001).
Peningkatan defosforilasi ATP akan memacu peningkatan substrat untuk enzim xantin oksidase (sel pankreas mempunyai aktivitas tinggi terhadap enzim ini), lebih lanjut meningkatkan produksi asam urat. Xantin oksidase mengkatalisis reaksi pembentukan anion superoksida aktif. Dari pembangkitan anion superoksida, terbentuk hidrogen peroksida dan radikal superoksida. NO dan oksigen reaktif tersebut adalah penyebab utama kerusakan sel pankreas (Szkudelski, β001).
2.9 KERANGKA KONSEP
Keterangan: = Variabel yang diperiksa
Dalam keadaan hiperglikemik terjadi stress oksidatif akibat autooksidasi dan glikolisis. ROS utama yang dihasilkan secara intraseluler adalah superoxide anion (O2•-) yang akan dikonversi menjadi hydrogen peroxide (H2O2) oleh proses dismutasi spontan ataupun reaksi yang dikatalisis oleh antioksidan endogen primer utama yang disebut enzim superoxide dismutase (SOD). H2O2 kemudian akan ditransformasi oleh antioksidan endogen primer lainnya seperti enzim catalase (CAT) menjadi molekul air dan oksigen. Enzim CAT berfungsi untuk mencegah akumulasi H2O2 berlebih sehingga produksi hydroxil radical (OH•) yang dapat memicu terjadinya peroksidasi lipid mampu dihambat sehubungan dengan tidak terdapatnya sistem pertahanan enzim yang mampu menghidrolisis OH• yang terbentuk. OH• dapat menyebabkan terjadinya oksidasi lipid,
Glikolisis >>>
2O2 -
SOD
H2O2
O2 CAT
2H2O + O2
OH-
Peroksidasi lipid
Fe2- Fe3-
MDA
Curcuminoid Hiperglikemia
protein dan DNA dan pada akhirnya menyebabkan kerusakan terhadap struktur sel dengan hasil sampingan berupa malondialdehid (MDA) (Kwiecien, et al., 2002; Liu, et al., 2002; Devasagayam, et al., 2003; Turrens, 2003; Weydert & Cullen, 2010).
Enzim CAT dan GPx berfungsi untuk mencegah akumulasi H2O2 berlebih sehingga produksi hydroxil radical (OH•) yang dapat memicu terjadinya lipid mampu dihambat sehubungan dengan tidak terdapatnya sistem pertahanan enzim yang mampu menghidrolisis OH• yang terbentuk. Curcuminoid mampu menginduksi peningkatan aktivitas seluruh enzim antioksidan endogen dengan cara meningkatkan ekspresi gen enzim tersebut sehingga produksinya menjadi bertambah dan dapat bereaksi dengan radikal bebas lalu mengubahnya menjadi produk yang lebih stabil.
Pada keadaan stress oksidatif, produksi O2•- yang terbentuk akan berlebihan, begitu juga dengan produksi H2O2, sehingga enzim tidak dapat menetralkan semua H2O2 yang terbentuk. Akibatnya, H2O2 secara spontan akan direduksi oleh ion-ion logam transisi bebas seperti Fe2+ dan Cu2+ menghasilkan OH• melalui reaksi Fenton dan Haber-Weiss. Curcuminoid mampu mencegah reduksi spontan H2O2 dengan berkompetisi terhadap ion-ion logam transisi bebas sehingga reaksi Fenton dan Haber-Weiss yang dapat menghasilkan OH• dapat dicegah, akibatnya proses peroksidasi lipid dapat dicegah.
2.10 Hipotesis Penelitian
Berdasarkan latar belakang masalah, landasan teori dan kerangka konseptual tersebut disusun hipotesis penelitian sebagai berikut: