Bird assemblages of an Iberian cereal steppe
A. Delgado
a,1, F. Moreira
b,∗aLiga para a Protecção da Natureza, Estrada do Calhariz de Benfica, 187, P-1500 Lisbon, Portugal
bCentro de Ecologia Aplicada ‘Prof. Baeta Neves’, Instituto Superior de Agronomia, Tapada da Ajuda, P-1399 Lisbon Codex, Portugal
Received 22 January 1999; received in revised form 29 June 1999; accepted 12 August 1999
Abstract
The cereal steppes of Castro Verde (Portugal) support many bird species of high conservation value on which agricultural practices can have significant impacts. The aim of this study was to describe the seasonal variations in bird assemblages using the agricultural landscape mosaic and to explore the potential ecological implications of changes in land use on bird populations. Bird densities and habitat variables were measured in four habitat types (cereal fields, fallow land, stubbles, and ploughed land) during 1997/1998, and detrended correspondence analysis used to summarise information on the structure of bird assemblages across habitats in the winter, breeding and post-breeding periods.Pluvialis apricariaL.,Vanellus vanellus
L.,Alauda arvensisL. andAnthus pratensisL. were abundant during winter. During the breeding season, bird assemblages
were quite distinct across land use types, with species typical of cereal fields (e.g.Miliaria calandraL.), fallow land (e.g.
Melanocorypha calandraL.) and ploughed land (e.g.Anthus campestrisL.). After the breeding season, the overlap in bird
assemblages increased again, withM. calandra,M. calandraL. andCiconia ciconiaL. being common. Species used different
habitats along the year, showing the importance of maintaining the traditional agricultural landscape for preserving the diversity of bird species. The potential impacts of several types of agricultural changes (e.g. agricultural intensification, agricultural abandonment and increase in stocking levels) on the bird populations were discussed on the basis of the results obtained. Any change in the current farming system will probably lead to a decrease in steppe bird diversity. ©2000 Elsevier Science B.V. All rights reserved.
Keywords:Landscape; Cereal cultivation; Bird assemblage structure; Impacts of agricultural changes
1. Introduction
Low-intensity farming systems of Europe have a high bird diversity (Tucker, 1997) and, among these, the pseudosteppes of the Iberian Peninsula
∗Corresponding author. Tel.:+351-1-3616080;
fax:+351-1-3623493.
E-mail address:[email protected] (F. Moreira).
1Present address: Centro de Ecologia Aplicada ‘Prof. Baeta Neves’, Instituto Superior de Agronomia, Tapada da Ajuda, P-1399 Lisbon Codex, Portugal.
hold a large proportion of species with unfavourable conservation status (Suárez et al., 1997; Tucker, 1997). Iberian pseudosteppes include semideserts, ‘páramos’ and cereal steppes (Mart´ınez and Purroy, 1993; Teller´ıa et al., 1988a, b). The most common pseudosteppe in western Spain and Portugal is the cereal steppe, created in flat, open landscapes by the extensive cultivation of cereals on a rotational ba-sis, resulting in a mosaic of cereal fields, stubbles, ploughed and fallow land (usually used as sheep pas-tures) (Suárez et al., 1997; Moreira, 1999). Cereal steppes constitute a stronghold for several threatened
bird species, such as the great bustard,Otis tardaL.,
the little bustard, Tetrax tetrax L., the sandgrouse, Pteroclesspp. and the lesser kestrel,Falco naumanni
Fleischer (Tucker and Heath, 1994; Suárez et al., 1997).
Cereal steppes represent an economically marginal farming system, with yields of less than half the aver-age European Union yields (Suárez et al., 1997). Con-sequently, land use of cereal steppes has been chang-ing in the last decades because of agricultural intensi-fication (especially irrigation), land abandonment and afforestation (Tucker and Heath, 1994; Suárez et al., 1997). Thus, this landscape is threatened by habitat degradation and loss, which can have significant im-pacts on the bird populations (e.g. Baldock, 1991; Diaz et al., 1993; Santos, 1996). The present Common Agri-cultural Policy (CAP) of the European Union promotes the maintenance of such farmed landscapes through agri-environmental regulation based on the compen-sation of farmers for maintaining agricultural prac-tices allowing the conservation of threatened species (Robson, 1997).
Several authors (e.g. Baldock et al., 1993; Beaufoy et al., 1994; Telleria et al., 1994; Suárez et al., 1997) have drawn attention to the fact that the scientific basis for establishing practical management actions aimed at maintaining or increasing the bird popula-tions are extremely scarce. Many studies of birds of pseudosteppes have focused on particular species (e.g. Mart´ınez, 1994), season (e.g. Mart´ınez and Purroy, 1993; Mart´ınez and De Juana, 1996) or type of land-scape (e.g. Teller´ıa et al., 1988a; Hódar, 1996a, b), but few (e.g. Teller´ıa et al., 1988b in central Spain) have attempted to characterise the seasonal use by bird as-semblages of the various components of the mosaic cover created by the extensive cultivation of cereals. This type of data is, however, needed for adequately evaluating the consequences of changes in agricultural practices on bird populations.
The objectives of this paper were:
1. to describe the seasonal variation in land use of an agricultural mosaic of the largest cereal steppe in Portugal;
2. to describe the bird assemblages using each land-use category in this mosaic; and
3. to explore the implications of changes in agricul-tural practices on both the agriculagricul-tural landscape mosaic and the bird populations.
2. Methods
2.1. Study area
The study site was an area of 5240 ha located approximately at the centre of the cereal steppe of Castro Verde (ca. 37◦43′N, 7◦57′W), the most
impor-tant area for steppe birds in Portugal (Grimmet and Jones, 1989). The traditional agricultural system used in this region is as follows: each farm is divided into parcels, each of which lies under cereal cultivation for two years, after which the land is left fallow for 2–3 years. The parcel is then ploughed to re-initiate the rotation cycle. Fallow land is generally used as pasture for sheep. The altitude ranges from 150 to 240 m, the annual average temperature from 15.5 to 16.0◦C, and the annual rainfall from 500 to 600 mm.
The site was located in the Meso-Mediterranean bioclimatic stage (Rivas-Mart´ınez, 1981), tree cover being restricted to some plantation of olive trees (Olea europaeaL.). Areas of scrub (Cistus ladanifer
L.) occurred in rocky outcrops with no agricultural use.
2.2. Land use
The availability of the different land-use classes was assessed by mapping land use in the whole study area (scale 1 : 25 000) in November 1996, using aerial photography and field checks of all farms. From then until August 1997, a monthly survey was made to register land-use changes. The area with each land use type was measured with a planimeter. In each season, i.e. winter (November to February), spring (breeding season, i.e. March to May), and summer (post-breeding, i.e. June to August) the percentage of the area with each land use was calculated by averag-ing the monthly estimates. Nine land uses were con-sidered i.e. three types of cereals (wheatTriticumspp., oatsAvenasp. and barleyHordeumsp.), two types of
2.3. Bird counts
Bird counts were carried out within patches of a given land use type, using the Jarvinen and Vaisa-nen (1975) line transect sampling. Observations were recorded within, and beyond, a main belt of 25 m on both sides of the observer. Only birds within the sam-pled patch of habitat were registered.
Transects were defined in the land cover map in or-der to account for two basic conditions: (1) a mini-mum length of 500 m, and (2) assuring a sample size of at least 10 transects in each land-use type.
Counts were made during the first 3 h after sun-rise. Mean travel speed (±SD) was 1.94±0.39 km/h
(n=358). Transect length ranged from 500 to 1125 m
(mean=782±165 m).
Cereal field transects were surveyed monthly and these data were grouped in the following time peri-ods: winter (December to February), breeding (March to May) and post-breeding (June to August). Fallow field transects were counted in winter (in Novem-ber and again in January), breeding (April) and post-breeding (July). Ploughed land and stubble tran-sects were counted twice in their season of occurrence (respectively breeding and post-breeding).
2.4. Habitat variables
Data on vegetation height and percentage cover were collected in fallows and cereal fields in Novem-ber (DecemNovem-ber for cereal fields), January, April and July. Along each transect used for bird counts, the height of the vegetation was measured over the first 100 m of the transect at 10 sites (every 10 m) with a ruler to the nearest cm. The percentage cover in fallows was measured at 5 sites (20 m distance) along the transect. At each site, a 10-m tape was stretched in a random direction and the length of the tape with vegetation cover noted to the nearest 5 cm. For cereal fields, due to difficulties (potential crop damage) in stretching the tape, a quadrat of 50×50 cm2was used
for estimating visually the cover of vegetation (Hays et al., 1981).
2.5. Data analyses
In half of the birds species considered for data analyses (11 out of 22), no individuals were
regis-tered within the main belt in at least one season. Thus, data for each transect were pooled (sum of de-tections within, and beyond, the belt) and expressed as (birds km−1). Species occurring in<5% of all the
counts were not analysed. Counts of both species of the genusGaleridaoccurring in the area (thekla lark, G. theklae Brehm, and crested lark, G. cristata L.)
were pooled, due to difficulty in identifying some individuals.
Data for each transect were analysed grouped into season. Winter and post-breeding counts were log transformed to minimise the influence of large flocks in multivariate analysis (see below). When more than one count was made in each transect, data were aver-aged prior to transformation. For the breeding season the maximum count was used for species using the habitat for nesting and the average for species using the habitat for feeding (e.g. white stork,Ciconia cico-niaL., and cattle egrett,Bubulcus ibisL.), as it was
as-sumed that the maximum count was the best estimate of breeding densities whereas the average would be the best indicator of intensity of use as feeding habitat. Detrended correspondence analysis (DCA) (Gauch, 1982), was used to summarise the information on bird assemblages across the several components of the landscape. This ordination technique involves the derivation of new axes that maximally account for the structure of the points in a multi-dimensional space, making possible the reduction of dimensionality with minimal loss of information. These new axes max-imise the correspondence between habitats and birds species, so that transects with similar bird assemblage composition are placed together, as are species with similar patterns of abundance across transects. DCA was carried out using the program CANOCO(Ter
Braak, 1987), with the option for detrending by second-order polynomials.
For habitat variables, each land-use type was char-acterised by the average vegetation height and cover.
Non-parametric tests (Siegel and Castellan, 1988) were used to compare:
1. transect coordinates of each habitat type in the axes of DCA;
2. bird densities across habitats; and
probability of incorrectly rejecting one or more true null hypothesis. Analyses were performed using the SPSS software (Norusis, 1992).
3. Results
3.1. Land use and habitat variables
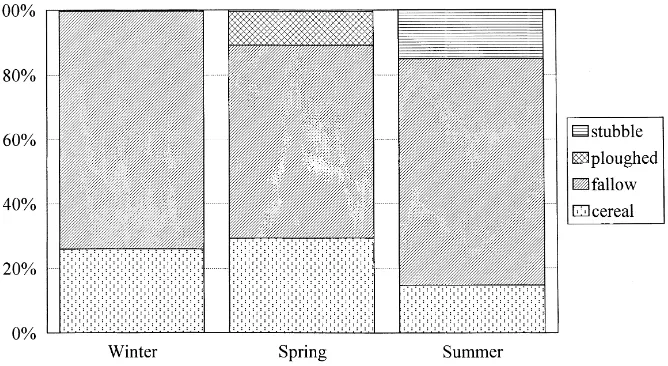
Seasonal variations in the agricultural landscape mosaic are shown in Fig. 1. During winter, the agricul-tural mosaic consisted of cereal fields (26% of the land use) and fallows (73%), 27% of which were recent (re-sulting from the stubbles of the previous summer). In spring, the landscape included ploughed fields (10% of the land cover), rapidly covered with vegetation and classified as fallows in the following summer. In sum-mer, after harvest stubbles averaged 15% of the land cover. Several cereal fields were not harvested due to the low seed production caused by adverse weather conditions in that year.
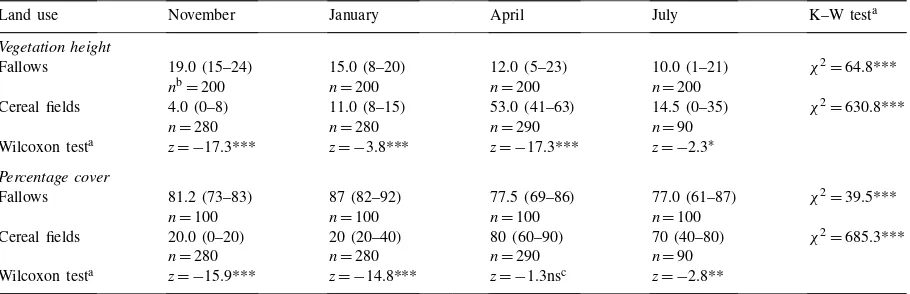
Table 1 shows the seasonal variation of vegeta-tion height and cover in fallows and cereal fields. Most comparisons (only one exception) yielded sig-nificant differences between habitats and seasons (for each habitat). In fallows, vegetation height and cover decreased progressively from winter to sum-mer. For cereal fields, there was a peak in both these
Fig. 1. Average proportion of the land use of the Castro Verde study area occupied in each season by fallow land, cereal fields, stubbles and ploughed fields.
variables during spring, when the plants were fully developed.
3.2. Bird assemblage structure
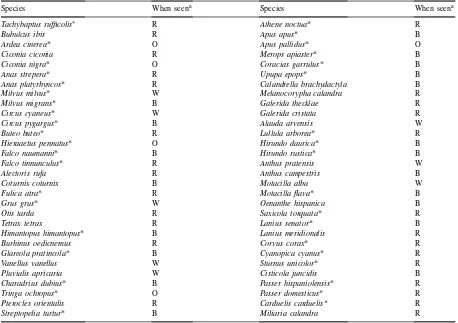
The 60 bird species observed during the field work are listed in Table 2. Of these, only 22 occurred in >5% of the transects.
3.2.1. Winter
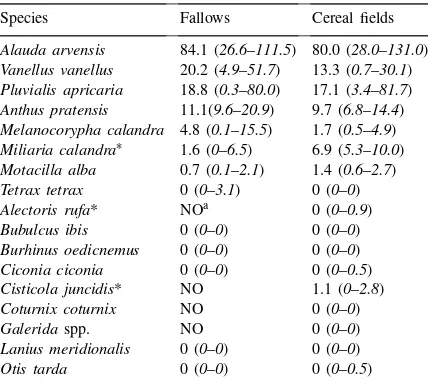
The four commonest species wintering in both, fallow land and cereal fields were skylark, Alauda arvensis L., golden plover, Pluvialis apricaria L.,
lapwing, Vanellus vanellus L., and meadow pipit, Anthus pratensis L., all reaching median densities
>10 birds km−1 (Table 3). The results of the DCA
(Fig. 2) suggest that there were differences in bird assemblage structure between these two components of the landscape, in spite of some overlap. There were significant differences between fallow and cereal transect coordinates in both the first (Wilcoxon test,
z= −4.06,p<0.001) and the second axis (Wilcoxon
test, z= −3.07, p<0.01). The first axis separated a
group of species which were particularly abundant in some cereal fields, i.e. red-legged partridge,Alectoris rufa L., quail, Coturnix coturnix L., Galerida spp.
and fan-tailed warbler,Cisticola juncidisRafinesque.
Table 1
Seasonal variation in median (with 25% and 75% quartiles), vegetation height (cm) and cover (%) in fallow land and cereal fields in Castro Verde
Land use November January April July K–W testa
Vegetation height
Fallows 19.0 (15–24) 15.0 (8–20) 12.0 (5–23) 10.0 (1–21) χ2=64.8***
nb=200 n=200 n=200 n=200
Cereal fields 4.0 (0–8) 11.0 (8–15) 53.0 (41–63) 14.5 (0–35) χ2=630.8***
n=280 n=280 n=290 n=90
Wilcoxon testa z= −17.3*** z= −3.8*** z= −17.3*** z= −2.3∗
Percentage cover
Fallows 81.2 (73–83) 87 (82–92) 77.5 (69–86) 77.0 (61–87) χ2=39.5***
n=100 n=100 n=100 n=100
Cereal fields 20.0 (0–20) 20 (20–40) 80 (60–90) 70 (40–80) χ2=685.3***
n=280 n=280 n=290 n=90
Wilcoxon testa z= −15.9*** z= −14.8*** z= −1.3nsc z= −2.8**
∗p
<0.05, **p<0.01, ***p<0.001.
aResults of the Kruskall–Wallis (K–W) test and Wilcoxon test are presented. bn=sample size.
cp> 0.05.
with several fallow transects (stone curlew,Burhinus oedicnemus L., and Tetrax tetrax) and one species
(Bubulcus ibis) occurring in high density in two
ce-real transects. The remaining species were near to the origin of the plot, showing no particular pattern of association with any of the land-use types. A univari-ate comparison of species abundances between fallow land and cereal fields (Table 3) yielded significant differences for A. rufa (Wilcoxon test, z= −2.88, p<0.01), C. juncidis (Wilcoxon test, z= −4.74,
p<0.001) and corn bunting, Miliaria calandra L.,
(Wilcoxon test, z= −3.5, p<0.001), all more
abun-dant in cereal fields.
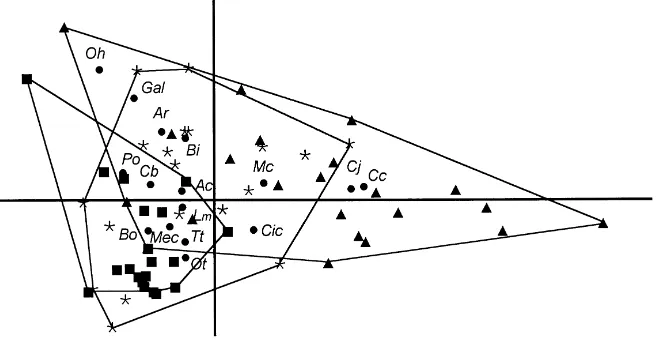
3.2.2. Breeding season
Each of the habitats had a rather distinctive bird assemblage (Table 4) as clearly shown in the DCA results (Fig. 3), with significant differences between cereal, fallow and ploughed land transect coordinates in the three axes (Kruskal–Wallis test,p<0.01). The
patches of ploughed land had a high abundance of short-toed lark, Calandrella brachydactyla Leisler
(24 birds km−1) and relatively high densities
(com-pared to other habitats) of black-eared wheatear,
Oenanthe hispanica L., B. oedicnemus, and tawny
pipit, Anthus campestris L. Fallows were
charac-terised by the abundance of calandra lark, Melano-corypha calandra (19 birds km−1) and Tetrax tetrax
(10 birds km−1), as well as a group of other species
attaining higher densities in fallows than in other habitats:B. ibis,Ciconia ciconiaandOtis tarda.
Cereal fields had a high abundance ofMiliaria ca-landra(36 birds km−1),C. juncidis(12 birds km−1),T. tetrax(10 birds km−1), andC. coturnix(2.3 birds km−1).
This habitat was the only one having exclusive species (A. pratensis, A. arvensis,A. rufaand southern grey
shrike,Lanius meridionalisTemminck).
Comparison of species abundances across fallows, ploughed land and cereal fields (Table 4) yielded sig-nificant differences (Kruskal–Wallis test,p<0.05) for
most species.
3.2.3. Post-breeding
The structure of bird assemblages was more variable after the breeding season (Fig. 4 and Table 5) in spite of significant differences between cereal fields, fallow and stubbles transect coordinates (for the first axis, Kruskal–Wallis test,χ2=22.7,p<0.001; for the
sec-ond axis, Kruskal–Wallis test,χ2=11.8,p<0.01).
Fallows were characterised by the abundance of
Melanocorypha calandra(4.0 birds km−1),C. ciconia
(1.7 birds km−1) andL. meridionalis(1.4 birds km−1).
Table 2
Bird species observed in the study area from November 1996 to August 1997
Species When seena Species When seena
Tachybaptus rufficolis∗ R Athene noctua* R
Bubulcus ibis R Apus apus* B
Ardea cinerea* O Apus pallidus* O
Ciconia ciconia R Merops apiaster* B
Ciconia nigra* O Coracias garrulus* B
Anas strepera* R Upupa epops* B
Anas platyrhyncos* R Calandrella brachydactyla B
Milvus milvus* W Melanocorypha calandra R
Milvus migrans* B Galerida thecklae R
Circus cyaneus* W Galerida cristata R
Circus pygargus* B Alauda arvensis W
Buteo buteo* R Lullula arborea* R
Hieraaetus pennatus* O Hirundo daurica* B
Falco naumanni* B Hirundo rustica* B
Falco tinnunculus* R Anthus pratensis W
Alectoris rufa R Anthus campestris B
Coturnix coturnix B Motacilla alba W
Fulica atra* R Motacilla flava* B
Grus grus* W Oenanthe hispanica B
Otis tarda R Saxicola torquata* R
Tetrax tetrax R Lanius senator* B
Himantopus himantopus* B Lanius meridionalis R
Burhinus oedicnemus R Corvus corax* R
Glareola pratincola* B Cyanopica cyanus* R
Vanellus vanellus W Sturnus unicolor* R
Pluvialis apricaria W Cisticola juncidis B
Charadrius dubius* B Passer hispaniolensis* R
Tringa ochropus* O Passer domesticus* R
Pterocles orientalis R Carduelis carduelis* R
Streptopelia turtur* B Miliaria calandra R
aO, occasional; W, exclusively during winter; B, breeding season; and R, all year round.
∗Species occurring in
<5% of the counts, and hence not included in the analyses.
The main feature of cereal fields was the abundance ofC. juncidis(6.1 birds km−1), along with the
occur-rence ofC. ciconia(1.3 birds km−1),L. meridionalis
(0.5 birds km−1) andC. coturnix(0.4 birds km−1).
Species typical of both, fallow land ( Melanoco-rypha calandra) (2.4 birds km−1) and cereal fields
(Miliaria calandra) (0.8 birds km−1) occurred in
stubbles. L. meridionalis (0.8 birds km−1) also
at-tained high abundance in comparison with other seasons.
A univariate comparison of species abundance across habitat types (Table 5) yielded significant differ-ences (Kruskal–Wallis test, p<0.05) for
Melano-corypha calandra,B. oedicnemus andC. ciconia, all
more abundant in fallows, and for C. juncidis and C. coturnix, more abundant in cereal fields.
4. Discussion
4.1. Assumptions and data limitations
Open landscapes with a flat topography were studied, in which it was assumed that differences in bird detectability across land-use types did not have a significant influence on the results. The ex-ception could be the well-developed cereal fields during the spring census, where detectability could be reduced. Nevertheless, the fact that during this season there was a clear relationship across habi-tats between the abundance index (birds km−1)
Table 3
Winter relative densities (birds km−1) in fallow land (n=20 tran-sects) and cereal fields (n=29 transects) in Castro Verde, with
median and inter-quartile distance
∗Species with significant differences between fallow and cereal
field densities (Wilcoxon test). aSpecies not observed.
most species suggests that detectability was not a problem.
Landscape-scale effects, which were not taken into account, could influence the composition of bird as-semblages mainly through (1) patch size and edge ef-fects, and (2) a different scale of perception by species that select landscape configurations rather than spe-cific patches (Forman, 1997; Farina, 1998). The for-mer problem was minimised by the fact that the plots sampled (with the exception of fallows, which were the matrix of the landscape), had roughly the same size and shape and no distinct edges, such as hedgerows or tree lines. Also, some larger body sized species (e.g.
B. ibis, O. tarda and C. ciconia) explored the
land-scape at a larger scale, and caution should be taken when interpreting the results obtained.
Bird community structure varies in time, both in the presence or absence of species and in their relative abundances (Wiens, 1989). The restricted sampling period did not provide information on po-tential yearly variations in assemblage structure, but previous data suggest that during the breeding season the dominant species in each land-use type are fairly constant (Leitão and Moreira, 1996; Moreira, 1999).
Nevertheless, assemblage structure might be more variable during winter (Wiens, 1989).
4.2. Seasonal variation in bird assemblages
The winter overlap in the composition of bird assemblages of fallow land and cereal fields was caused by the widespread abundance of four win-tering species. The observed differences in habitat structure between these two land uses seemed not to influence bird assemblage composition. It is possible that there was enough food available (mainly leaves and seeds for A. arvensisand A. pratensis, and soil
invertebrates forP. apricariaandV. vanellus(Cramp
and Simmons, 1983; Cramp, 1988) in both habitats. Diaz and Teller´ıa (1994) have shown that food abun-dance was not a limiting factor for seed-eating birds wintering in cereal steppes of central Spain, and San-tos and Teller´ıa (1985) argue that invertebrate activity during winter, in the Iberian Peninsula, is sufficient for providing the necessary food resources for birds.
P. apricariaandV. vanelluscould respond to
vegeta-tion structure, as they avoid dense and tall vegetavegeta-tion, but vegetation height during this time of year was be-low the described limits for these species (Cramp and Simmons, 1983). The main differences in wintering species density occurred for species which used ce-real fields as breeding habitat (A. rufa,C. juncidisand Miliaria calandra). These species started to define
breeding territories in cereal fields at the end of win-ter, at a time when vegetation height was increasing.
During the breeding season particular bird species were associated with specific habitats. This suggests that vegetation structure was the main determinant of habitat selection during the breeding season, as reported for many bird species (Cody, 1985; Wiens, 1989). Species attached to small breeding territories probably were not able to explore, e.g. for foraging, other landscape patches, in contrast with other seasons when birds were not attached to breeding territories. Ploughed fields were used as nesting habitat by a group of species with a preference for bare ground and short vegetation, particularly C. brachydactyla (e.g.
Cramp and Simmons, 1983; Cramp, 1988; Moreira and Leitão, 1996). In fallow fields,Melanocrypha ca-landraandT. tetraxwere the more abundant species,
Fig. 2. Plot of the first two axes from detrended correspondence analysis based on the winter data. Transects with similar bird assemblage composition are placed together, as are species with similar pattern of abundance across transects. Eigenvalues are a measure of species spread in the axes. Axis 1, eigenvalue=0.17 (20.7% variance); and Axis 2, eigenvalue=0.12 (14.9% variance). Code for transect habitats:
quadrats, fallow land; and triangles, cereal fields. Codes for species (dots): Aa,Alauda arvensis; Ap,Anthus pratensis; Ar,Alectoris rufa;
Bi,Bubulcus ibis; Bo,Burhinus oedicnemus; Cc,Coturnix coturnix; Cic,Ciconia ciconia; Cj,Cisticola juncidis; Gal,Galeridaspp.; Lm, Lanius meridionalis; Ma,Motacilla alba; Mc,Miliaria calandra; Mec,Melanocorypha calandra; Ot,Otis tarda; Pa,Pluvialis apricaria;
Tt,Tetrax tetrax; and Vv,Vanellus vanellus.
Table 4
Relative densities (birds km−1) during the breeding season in fallow land (n=20 transects), cereal fields (n=29 transects) and ploughed land (n=10 transects) in Castro Verde, with median and inter-quartile distance
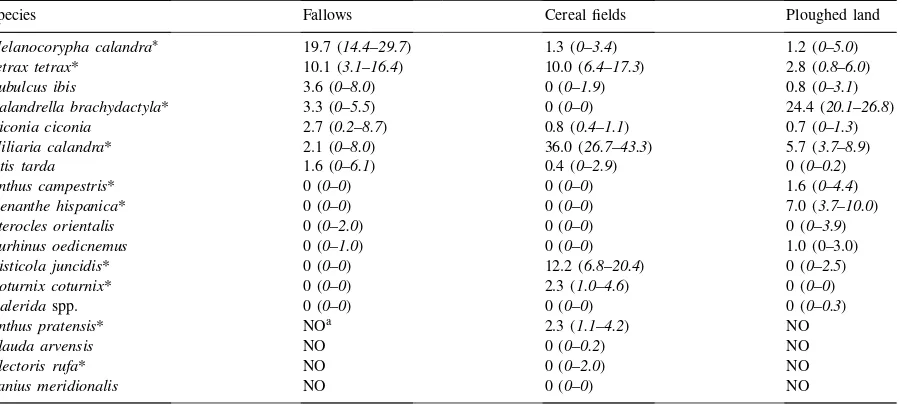
Species Fallows Cereal fields Ploughed land
Melanocorypha calandra∗ 19.7 (14.4–29.7) 1.3 (0–3.4) 1.2 (0–5.0) Tetrax tetrax* 10.1 (3.1–16.4) 10.0 (6.4–17.3) 2.8 (0.8–6.0) Bubulcus ibis 3.6 (0–8.0) 0 (0–1.9) 0.8 (0–3.1) Calandrella brachydactyla* 3.3 (0–5.5) 0 (0–0) 24.4 (20.1–26.8)
Ciconia ciconia 2.7 (0.2–8.7) 0.8 (0.4–1.1) 0.7 (0–1.3)
Miliaria calandra* 2.1 (0–8.0) 36.0 (26.7–43.3) 5.7 (3.7–8.9)
Otis tarda 1.6 (0–6.1) 0.4 (0–2.9) 0 (0–0.2)
Anthus campestris* 0 (0–0) 0 (0–0) 1.6 (0–4.4)
Oenanthe hispanica* 0 (0–0) 0 (0–0) 7.0 (3.7–10.0)
Pterocles orientalis 0 (0–2.0) 0 (0–0) 0 (0–3.9)
Burhinus oedicnemus 0 (0–1.0) 0 (0–0) 1.0 (0–3.0)
Cisticola juncidis* 0 (0–0) 12.2 (6.8–20.4) 0 (0–2.5)
Coturnix coturnix* 0 (0–0) 2.3 (1.0–4.6) 0 (0–0)
Galeridaspp. 0 (0–0) 0 (0–0) 0 (0–0.3)
Anthus pratensis* NOa 2.3 (1.1–4.2) NO
Alauda arvensis NO 0 (0–0.2) NO
Alectoris rufa* NO 0 (0–2.0) NO
Lanius meridionalis NO 0 (0–0) NO
∗Species with significant diferences between habitats (Kruskal–Wallis test).
Fig. 3. Plot of the first two axes from detrended correspondence analysis based on the breeding season data. Axis 1, eigenvalue=0.56
(19.4% variance); and Axis 2, eigenvalue=0.25 (13.4% variance). Code for transect habitats: quadrats, fallow land; triangles, cereal fields;
and diamonds, ploughed land. Codes for species (dots) similar to Fig. 2 with the additional species: Ac,Anthus campestris; Cb,Calandrella brachydactyla; Oh,Oenanthe hispanica; and Po,Pterocles orientalis.
Fig. 4. Plot of the first two axes from detrended correspondence analysis based on the post-breeding season data. Axis 1, eigenvalue=0.36
(16.7% variance); and Axis 2, eigenvalue=0.29 (13.3% variance). Code for transect habitats: quadrats, fallow land; triangles, cereal fields; and asterisks, stubbles. Codes for species (dots) similar to Figs. 2 and 3.
Cereal fields were used mainly by species preferring tall and dense vegetation, such asMiliaria calandra, C. juncidis and C. coturnix (Cramp and Simmons,
1980; Cramp, 1992; Cramp and Perrins, 1994; Mor-eira, 1999; Borralho et al., 1998). Some individuals which had not started to migrate were also recorded. High densities ofT. tetraxwere found in cereal fields
and fallows. Most studies on habitat selection in this
species (Shulz, 1985; Mart´ınez, 1994; Mart´ınez and De Juana, 1996; Moreira and Leitão, 1996; Moreira, 1999) suggest that cereal is not a preferred habitat, but the result obtained might be explained by the fact that due to weather conditions cereal fields did not grow as usual in that particular year.
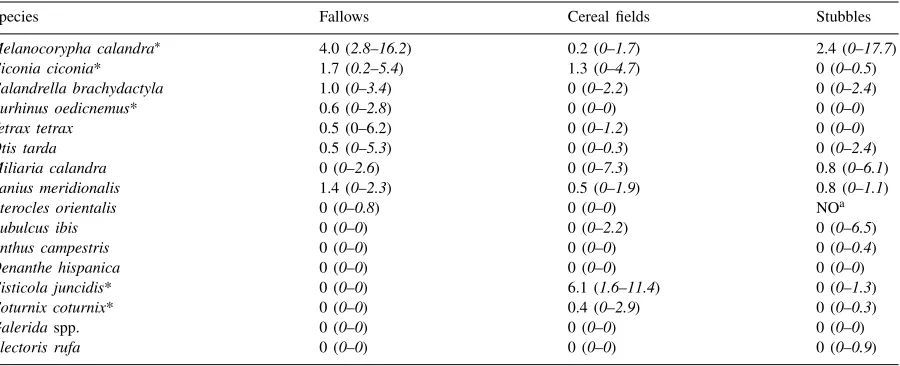
Table 5
Post-breeding relative densities (birds km−1) in fallow land (n=20 transects), cereal fields (n=22 transects) and stubbles (n=20 transects) in Castro Verde, with median and inter-quartile distance
Species Fallows Cereal fields Stubbles
Melanocorypha calandra∗ 4.0 (2.8–16.2) 0.2 (0–1.7) 2.4 (0–17.7) Ciconia ciconia* 1.7 (0.2–5.4) 1.3 (0–4.7) 0 (0–0.5) Calandrella brachydactyla 1.0 (0–3.4) 0 (0–2.2) 0 (0–2.4) Burhinus oedicnemus* 0.6 (0–2.8) 0 (0–0) 0 (0–0)
Tetrax tetrax 0.5 (0–6.2) 0 (0–1.2) 0 (0–0)
Otis tarda 0.5 (0–5.3) 0 (0–0.3) 0 (0–2.4)
Miliaria calandra 0 (0–2.6) 0 (0–7.3) 0.8 (0–6.1)
Lanius meridionalis 1.4 (0–2.3) 0.5 (0–1.9) 0.8 (0–1.1)
Pterocles orientalis 0 (0–0.8) 0 (0–0) NOa
Bubulcus ibis 0 (0–0) 0 (0–2.2) 0 (0–6.5)
Anthus campestris 0 (0–0) 0 (0–0) 0 (0–0.4)
Oenanthe hispanica 0 (0–0) 0 (0–0) 0 (0–0)
Cisticola juncidis* 0 (0–0) 6.1 (1.6–11.4) 0 (0–1.3)
Coturnix coturnix* 0 (0–0) 0.4 (0–2.9) 0 (0–0.3)
Galeridaspp. 0 (0–0) 0 (0–0) 0 (0–0)
Alectoris rufa 0 (0–0) 0 (0–0) 0 (0–0.9)
∗Species with significant diferences between habitats (Kruskal–Wallis test).
aSpecies not observed in the transects.
resource (Teller´ıa et al., 1988a; Diaz and Telleria, 1994), used especially by Miliaria calandra and Melanocrypha calandra.C. coturnix andC. juncidis
remained associated to cereal fields, whereas C. ci-coniaused fallows and non-harvested cereal fields as
feeding grounds.
Most species used several habitats of the agricul-tural landscape mosaic along the annual cycle. The
maintenance of the mosaic agricultural system is, therefore, essential for preserving the diversity of bird species in this landscape. Other authors have also shown the importance of habitat diversity either for specific species (e.g. Mart´ınez, 1994; Salamonard et al., 1996) and the whole bird community (Donazar et al., 1997).
4.3. Implications of agricultural changes
Cereal steppes are threatened by agricultural changes (Baldock, 1991; Diaz et al., 1993; Tucker and Heath, 1994; Santos, 1996; Suárez et al., 1997). The results of this paper and other research made in the Castro Verde region can be used for a pre-liminary evaluation of the impact of several types of agricultural changes on bird populations, with the assumption of non-adaptation of bird species to a changing environment.
Cereal fields and stubbles are likely to decrease, to be replaced by pastures and fallow land. As a conse-quence bird species that rely on cereal would probably be reduced, whereas bird species occurring mainly in fallows could be promoted. An increase in stock den-sity would lead to changes in vegetation structure (de-creased height and higher cover by bare ground) due to overgrazing, along with increased trampling of nests and eggs (Ausden and Treweek, 1995; Donazar et al., 1997). Thus, populations ofC. brachydactyla,O. his-panicaandA. campestriscould have an increased area
of available habitat, although with a potentially lower reproductive success.
Homogenisation of the landscape, with a progres-sive increase in vegetation height and shrub cover (mainlyCistusspp.) would occur in abandoned fields.
In a first stage, this would benefit species likeB. oedic-nemus, B. ibis and Galerida spp. (Moreira, 1999),
but the increase in shrub cover would be equivalent to habitat loss for the steppe birds. Santos (1996) described the bird communities associated with suc-cessional stages of vegetation after land abandonment in this region, and showed that Sylvia conspicillata
Temminck was the only species with unfavourable conservation status found on scrubland.
negatively the populations of T. tetrax and M. ca-landra. If intensification increased the frequency of
cereal cultivation, this could lead to a population growth for species more dependent on cereal fields (such asC. juncidisandMiliaria calandra).
5. Conclusions
The bird diversity of the farmed landscape of Castro Verde depends on the maintenance of a diverse agri-cultural mosaic, as promoted by the extensive cultiva-tion of cereals in a rotacultiva-tional system. It is hypothesised that land-use modifications due to agricultural policies will promote changes in bird assemblage structure, namely a loss in steppe bird richness and an increase in specific bird populations, depending on the type of changes. Scientific research in steppic landscapes with varying level of agricultural changes or in sub-regions of Castro Verde where agricultural changes are already taking place would provide tests for these predictions.
Acknowledgements
This research was financed by the LPN project ‘Conservation of steppe birds in Castro Verde region’, partially funded by LIFE programme (Contract 4-3200/95/510) and Fundação Luso-Americana para o Desenvolvimento. We wish to thank Rui Morgado for providing the data on land-use cover. Thanks also due to Pedro Beja, Rui Borralho, Mario Diaz and two anonymous referees for useful comments that signifi-cantly improved the manuscript. This paper is a con-tribution to project PRAXIS XXI/C/AGR/11063/98 “Factores determinates da biodiversidade em pousios de estepes cerealiferas: implicações para definiçäo de normas de gestäo agro-ambiental”.
References
Ausden, M., Treweek, J., 1995. Grasslands. In: Sutherland, W.J., Hill, D.A. (Eds.), Managing Habitats for Conservation. Cambridge University Press, Cambridge, pp. 179–229. Baldock, D., 1991. Implications of EC farming and countryside
policies for conservation of lowland dry grasslands. In: Goriup, P.D., Batten, L.A., Norton, J.A., (Eds.), The Conservation of Lowland Dry Grassland Birds in Europe.JNCC, Peterborough, pp 111–117.
Baldock, D., Beaufoy, G., Bennett, G., Clark, J., 1993. Nature conservation and new directions in the EC Common Agricultural Policy. Institute for European Environmental Policy, London.
Beaufoy, G., Baldock, D., Clark, J., 1994. The nature of farming. Low intensity farming systems in nine European countries. Institute for European Environmental Policy, London. Borralho, R., Stoate, C., Araújo, M., Rito, A., Carvalho, S., 1998.
Factores ambientais que afectaram a ocorrência primaveril de codornizCoturnix coturnixno Baixo Alentejo. In: Costa, L.T., Costa, H., Araújo, M., Silva, M.A., (Eds.), Actas do simpósio sobre aves migradoras na Pen´ınsula Ibérica. SPEA, Évora, pp. 68–72.
Cody, M.L., 1985. Habitat Selection in Birds. Academic Press, San Diego.
Cramp, S., Simmons, K.L.E. (Eds.), 1980. Handbook of the birds of Europe, the Middle East, and North Africa: the Birds of Western Paleartic, vol. II. Oxford University Press, Oxford. Cramp, S., Simmons, K.L.E. (Eds.), 1983. Handbook of the Birds
of Europe, the Middle East, and North Africa: the Birds of Western Paleartic, vol. III. Oxford University Press, Oxford. Cramp, S. (Ed.), 1988. Handbook of the Birds of Europe, the
Middle East, and North Africa: the Birds of Western Paleartic, vol. V. Oxford University Press, Oxford.
Cramp, S. (Ed.), 1992. Handbook of the Birds of Europe, the Middle East, and North Africa: the Birds of Western Paleartic, vol. VI. Oxford University Press, Oxford.
Cramp, S., Perrins, C.M. (Eds.), 1994. Handbook of the Birds of Europe, the Middle East, and North Africa: the Birds of Western Paleartic, vol. VII. Oxford University Press, Oxford. Diaz, M., Naveso, M.A., Rebollo, E., 1993. Respuestas de las
comunidades nidificantes de aves a la intensificación agr´ıcola en cultivos cerealistas de la Meseta Norte (Valladolid–Palencia, España). Aegypius 11, 1–6.
Diaz, M., Teller´ıa, J.L., 1994. Predicting the effects of agriculture changes in central Spanish croplands on seed-eating overwinter birds. Agric., Ecosyst. Environ. 49, 289–298.
Donazar, J.A., Naveso, M.A., Tella, J.L., Campión, D., 1997. Extensive grazing and raptors in Spain. Farming and Birds in Europe. The Common Agricultural Policy and its Implications for Bird Conservation. Academic Press, San Diego, pp. 117–149.
Farina, A., 1998. Principles and Methods in Landscape Ecology. Chapman and Hall, Cambridge.
Forman, R.T., 1997. Land mosaics. The ecology of landscapes and regions. Cambridge University Press, Cambridge.
Gauch, H.G., Jr., 1982. Multivariate Analysis in Community Ecology. Cambridge University Press, Cambridge.
Grimmet, R.F.A., Jones, T.A., 1989. Important Bird Areas in Europe. ICBP, Cambridge.
Hays, R.L., Summers, C., Seitz, W. (Eds.), 1981. Estimating Wildlife Habitat Variables. — U.S.D.I. Fish and Wildlife Service, FWS/OBS-81/47, Washington, 111 pp.
Hódar, J.S., 1996b. Temporal variations in two shrub-steppe bird assemblages in southeastern Spain: the importance of winter for non-steppe birds. In: Gutiérrez, J.F., Sanz-Zuasti, J. (Eds.), Conservacion de las aves esteparias y su habitat. Junta de Castilla y Léon, Valladolid, pp. 137–151.
Jarvinen, O., Vaisanen, R.A., 1975. Estimating relative densities of breeding birds by the line transect. Oikos 26, 316–322. Leitão, D., Moreira, F., 1996. Estrutura e composição das
comunidades de aves nidificantes na região de Castro Verde. Ciência e Natureza 2, 103–107.
Mart´ınez, C., 1994. Habitat selection by the little bustardTetrax tetraxin cultivated areas of Central Spain. Biological Conserv.
67, 125–128.
Mart´ınez, C., De Juana, E., 1996. Breeding bird communities of cereal crops in Spain: habitat requirements. In: Gutiérrez, J.F., Sanz-Zuasti, J. (Eds.), Conservacion de las aves esteparias y su habitat. Junta de Castilla y Léon, Valladolid, pp. 99-106. Mart´ınez, C., Purroy, F.J., 1993. Avifauna reprodutora en los
sistemas esteparizados ibericos. Ecologia 7, 391–401. Moreira, F., 1999. Relationships between vegetation structure and
breeding bird densities in fallow cereal steppes of Castro Verde, Portugal Bird Study.
Moreira, F., Leitão, D., 1996. A preliminary study on the breeding bird community in fallows of cereal steppes in Southern Portugal. Bird Conserv. Internat. 6, 255–259.
Norusis, M.J., 1992. SPSS for Windows: Professional Statistics. Release 6.
Rice, W.R., 1989. Analysing tables of statistical tests. Evolution 43, 223–225.
Rivas-Mart´ınez, S., 1981. Les étages bioclimatiques de la végétation de la Peninsule Ibérique. An. Jard. Bot. Madrid 37, 251–268.
Robson, N., 1997. The evolution of the Common Agricultural Policy and the incorporation of environmental considerations. In: Pain, D.J., Pienkowski, M.W. (Eds.), Farming and Birds in Europe. The Common Agricultural Policy and its Implications for Bird Conservation. Academic Press, San Diego. pp. 43–78. Salamonard, M., Bretagnolle, V., Boutin, J.M., 1996. Habitat use by Montagu’s harrier, little bustard and stone curlew in Western France: Crop types and spatial heterogeneity. In: Fernández, J., Sanz-Zuasti, J. (Eds.), Conservación de las Aves
Esteparias y su Hábitat. Junta de la Castilla y Leon, Valladollid, pp. 209-220
Santos, C.P., 1996. O abandono dos campos agr´ıcolas e suas implicações nas comunidades de aves nidificantes. Ciência e Natureza 2, 95–102.
Santos, T., Teller´ıa, J.L., 1985. Patrones generales de la distribucion invernal de Passeriformes en la Peninsula Iberica. Ardeola 32 (1), 17–30.
Shulz, H., 1985. Grundlagenforschung zur Biologie der ZwergtrappeTetrax tetrax. Braunschweig.
Siegel, S., Castellan, Jr., 1988. Nonparametric Statistics for the Behavioral Sciences. McGraw–Hill International eds. Suárez, F., Naveso, M.A., De Juana, E., 1997. Farming in the
drylands of Spain: birds of the pseudosteppes. In: Pain, D.J., Pienkowski, M.W. (Eds.), Farming and Birds in Europe. The Common Agricultural Policy and its Implications for Bird Conservation. Academic Press, San Diego. pp. 297–330. Teller´ıa, J.L., Santos, T., Álvarez, G., Saéz-Royuela, C., 1988a.
Avifauna de los campos de cereales del interior de España. In: Bernis, F. (Ed.). Aves de los medios urbano y agr´ıcola en las mesetas españolas, Madrid, Monografias de la Sociedade Española de Ornitologia 2, SEO, pp. 173–317.
Teller´ıa, J.L., Suárez, F., Santos, T., 1988b. Bird communities of the Iberian shrub-steppes. Holarctic Ecol. 11, 171–177. Telleria, J.L.,Santos, T., Diaz, M., 1994. Effects of agricultural
practices on bird populations in the Mediterranean region: the case of Spain. Proceedings Bird Numbers 1992, pp. 57–74. Ter Braak, C.J.F., 1987. CANOCO — A FORTRAN Program
for Canonical Community Ordination by [partial] [detrended] [canonical] correspondence analysis (version 2.1). Wageningen: TNO Institute of Applied Computer Sciences.
Tucker, G.M., 1997. Priorities for bird conservation in Europe: the importance of the farmed landscape. In: Pain, D.J., Pienkowski, M.W. (Eds.), Farming and Birds in Europe. The Common Agricultural Policy and its Implications for Bird Conservation. Academic Press, San Diego, pp. 79–116.
Tucker, G.M., Heath, M.F., 1994. Birds in Europe: Their Conservation Status. Birdlife International, Cambridge. Wiens, J.A., 1989. The Ecology of Bird Communities. Cambridge