Hormon dan Perkembangan
Tumbuhan
Kelompok:
1. Annisa Balqis
2. Enung Azizah Mulyawati 3. Mulyani
Instructive roles for hormones
in plant development
DAVID ALABADÍ, MIGUEL A. BLÁZQUEZ*, JUAN CARBONELL, CRISTINA FERRÁNDIZ
and MIGUEL A. PÉREZ-AMADOR.
Instituto de Biología Molecular y Celular de Plantas (UPV-CSIC), Universidad Politécnica de Valencia, Valencia, Spain
Hormon Tumbuhan
→ pengatur metabolisme, pertumbuhan, dan perkembangan.
→ tidak selalu seperti hormon metazoa. Salah
satunya karena hormon tumbuhan tidak disintesis di organ tertentu, kemudian di transfer menuju
organ target.
→ bisa disintesis di semua sel, dan kerjanya sangat sering ditemukan pada sel dimana ia dibuat.
→ Hormon pd tumbuhan sering disebut sebagai zat pengatur tumbuh – fungsinya tak spesifik
proses fisiologi tertentu. Misal auksin untuk penambahan laju pembelahan sel, etilen u/ pematangan buah dll.
→ Namun, beberapa bukti menunjukkan bahwa hormon berperan sebagai sinyal instruktif bagi tumbuhan.
→ Yang lebih menarik lagi, sebagian peran mirip dengan apa yang terjadi pada hormon peptida & protein selama perkembangan hewan.
Hormon pada tumbuhan tidak sekedar berperan sebagai sinyal transduser atau messenger, tapi juga mengatur secara umum.
Contohnya adalah pengaturan bagi kemungkinan tahapan perkembangan.
Gradien morfogen: Auxin on the Move
Auksin sangat penting untuk perkembangan tumbuhan. Distribusi asimetris dari auksin, memiliki beberapa efek:
● Polarisasi embrio
● Diferensiasi vaskular
● Pola perakaran dan batang ● Tropisme dan percabangan
Contoh auksin: IAA (Indole acetic acid), IBA (Indol butiric acid).
IAA (Indoleacetic acid)
IAA merupakan hormon auksin aktif pada tumbuhan. IAA bersifat:
- molekul organik kecil - mudah larut
- mampu melintasi membran
- terdistribusi asimetrik (adanya PAT/Polar auxin
Bagaimana Auksin Bergerak?
1. Floem : meristem & daun muda → floem → organ sink (akar).
2. Transport antar sel yang memerlukan protein
carrier.
PAT – Polar auxin transport: dikendalikan oleh
protein transport PINFORMED (PIN). Pada
Proses Sinyalisasi Auksin → Transkripsi
Proses Sinyalisasi Auksin → Transkripsi
Komponen-Komponen
● Gen Aux/IAA: membentuk protein berumur
pendek. 29 macam di Arabidopsis. Pada
promoter IAA, ditemukan juga AuxRE → sensitif auksin (bisa merespon).
● ARF: Auxin Response Factor - faktor
transkripsi yang mengikat AuxRE → memicu ekspresi gen responsif auksin. Mampu
Auksin pada Perkembangan Embrio
NB: Perubahan aliran auksin di atur oleh ekspresi gen dan polarisasi PIN transporter.
Mengapa Meristem tetap Eksis?
SAM pada Arabidopsis dipertahankan oleh gen
identitas meristem:
● WUSCHEL (WUS)
● CLAVATA 1-3 (CLV1-3)
● KNOTTED1-like Homeobox (KNOX)
→ Fungsi:
WUS: pembesaran stem cell (meristem), CLV repress WUS → ukuran sel SAM konstan.
Mengapa Meristem tetap Eksis?
Selain itu, ada SHORTMERISTEMLESS (STM) dan BREVIPEDICELLUS (BP) → cegah
meristem menjadi primordia daun.
Gen AS1 (ASYMMETRIC LEAVES1) ada di primordia daun → menghambat ekspresi gen identitas meristem.
Berbagai macam gen di atas juga bekerja pada daun dengan mekanime yang berbeda.
Mekanisme pada Meristem & Primordia
Pola filotaksis dan auksin
Nomor: letak primordia bunga. Tanda * : primordia floral
baru. Garis biru: primordia berkaitan. Warna merah: kadar tertinggi auksin.
Pergerakan auksin pada filotaksis
Auksin di transfer ke bagian pucuk, di primordia (auxin sink). Setelah primordia tumbuh, maka auksin dipompa kembali menuju ke batang,
Auksin dalam Filotaksis
→ Pola asimetris auksin bukan hanya untuk diferensiasi basal-apikal atau pembentukan
meristem pada embrio, tapi arsitektur tumbuhan secara keseluruhan.
→ PAT penting untuk perkembangan
pasca-embrionik. Semisal pembentukan primordia daun. → Penelitian terbaru: dinamika auksin di apeks menunjukkan PIN mengarahkan auksin dalam meristem. Prinsipnya, saat kadar auksin
mencapai ambang batas, PIN melakukan
re-orientasi auksin ke tempat inisiasi organ,
Auksin
danMorfogenesis
Selain pembentukan aksis tanaman, auksin juga berperan untuk gradien morfogenesik pada
beberapa organ: positional information dan pembesaran sel.
Contoh: perkembangan Gynoecium Arabidopsis.
→ perkembangan ada dua, yaitu basal apikal serta transversal. Auksin berperan dalam
Auksin dan Pembentukan Gynoecium
WT: wild type
ett: mutan ETTIN (kelompok ARF)
Hipotesis
Gradien Auksin
Kiri: Normal
Morphogenetic Switch yang
Morphogenetic Switch yang distimulasi oleh Cahaya dan
Hormon
Peran instruktif hormon diantaranya : pilihan rancangan
pertumbuhan yang sesuai
Pilihan ini dipengaruhi cahaya
Ada 2, yaitu
Skotomorfegenesis
Pertumbuhan embrio pada kondisi masih gelap
Embrio teretiolasi, hipokotil yang memanjang, dan
adanya kotiledon yang terlipat pada apical.
Bertujuan untuk melindungi embrio pada awal
Fotomorfogenesis
Ketika embrio mencapai cahaya, arah pertumbuhannya
berubah menjadi fotomorfogenesis
Dicirikan dengan adanya pembukaan kait pada apikal,
pembukaan kotiledon, pengekspresian gen yang terlibat pada perlindungan terhadap kerusakan oksidatif dan
Senesce or develop: hormonal regulation
of fruit set
Keputusan terbentuk atau tidaknya
buah
Faktanya, jalur perkembangan yang normal yang diikuti
oleh gynoecium akan mengarahkannya ke proses
senesce yang menyebabkan kehancuran gynoecium tsb.
Proses senesce ini akan ter represi, hanya jika terdapat
faktor luar berupa sinyal yang dapat memberikan alternative perkembangan buah.
Hormon2 ini mungkin menyerupai sinyal instruktif
yang mengawali program pembentukan buah. Program ini melibatkan sequential effects, atau proses beruntun yang melibatkan berbagai macam hormone.
Fertilisasi bukanlah syarat mutlak untuk terjadinya
perkembangan buah.
Hal tersebut dibuktikan dengan adanya buah2 yang
partenokarpi.
Pengaruh hormone tertentu seperti gad an auksin dapat
menginduksi perkembanagn buah pada emasculated flower atau setelah aborsi dini suatu biji.
Pembentukan buah secara
partenokarpi
Dibuktikan dengan penambahan inhibitor auksin yang
bernama NPA (Naphthylphthalamic acid)
NPA akan memblokir aliran auksin keluar ovarium
sehingga terjadi akumulasi auksin
Peristiwa akumulasi tersebut dapat mentrigger
Bagaimana dengan GA?
Ekspresi gen GA20ox yang berperan dalam biosintesis GA pada
tanaman kacang-kcangan, meningkat setelah fertilisasi
Peningkatan ini terutama terjadi di dalam ovarium, yang mana
bisa diindikasikan sebagai ‘peran instruktf’
Selanjutnya, pengambilan ovul segera setelah fertilisasi dapat
mencegah perkembangan dan differensiasi buah. Peristiwa ini dapat diasosiasikan dengan berkurangnya sintesis GA
Hubungan antara auksin dengan GA
Akumulasi ga diinduksi oleh auksin
Pemberian auksin 4-chloroindole-3-acetic acid
(4-CI-IAA) pada ovarium kacang menujukkan ekspresi gen yang terlibat dalam biosintesis GA
RESEPTOR FITOHORMON
Persepsi baru: reseptor hormon tumbuhan Angela K. Spartz and William M. Gray
Departemen Biologi Tumbuhan, Universitas Minnesota–Twin Cities,
Reseptor Hormon Tumbuhan
● Fitohormon sebagai pusat regulasi pertumbuhan dan perkembangan tumbuhan ● Hanya sedikit reseptor telah
teridentifikasi
● Mekanisme penghantaran
melalui sinyal kimiawi lingkunganFaktor endogenSinyal
Gen intrinsik
Pertumbuhan dan Perkembangan
Sejarah mengenai Fitohormon
● Julian von Sachs substansi pembentuk
organ spesifik endogen mobil sinyal
● Charles and Francis Darwin fototrofisme.
Pergerakanrumput saat terpapar cahaya secara lateral. Sinyal berasal dari apeks
tumbuhan menginisiasi pemanjangan sel di bagian bawahnya.
Konsep Fitohormon
● Sinyal indole-3-acetic acid (IAA atau auksin)
fitohormon pertama yang teridentifikasi
● Dua komponen : regulator dan reseptor kinase
(kelas protein yang digunakan dalam komunikasi sinyal secara kimiawi)
● Definisi hormon messenger kimiawi yang
memungkinkan terjadinya komunikasi sel
Mekanisme Kerja Hormon
(konsep)
● Fitohormon dlm konsentrasi rendah saja dapat memengaruhi
kontrol perkembangan pembelahan sel, ekspansi, diferensiasi, dan kematian sel.
● Kerja lokal tetapi pengaruh global
● Satu sel dapat merespons banyak tipe hormon, tetapi satu tipe
hormon hanya bekerja pada sel/ jaringan tertentu.
● Fitohormon diproduksi pada keseluruhan bagian tumbuhan
dan bisa memengaruhi sel/jaringan yang berasal dari sel yang sama
● Transpor fitohormon melalui pembuluh atau komunikasi antar
sel
Hormon IAA-model ubiquitin-mediated
● Disintesis dari L-triptofan (wild type) dan glukosinolat
(mutan)
● Berperan dalam diferensiasi vaskular
● Masa induktif oleh gen 4 s.d 30 menit
● Terdapat >29 gen pengkode AUX/IAA pada genom
Arabidopsis
● Bekerja karena adanya gen pengkode dan protein
pembawa AUX/IAA (dalam waktu singkat)
Hormon IAA
1. Konsentrasi auksin di bawah ambang
batas gen responsif auksin ditekan oleh protein Aux /IAA (protein tsb telah
mengalami heterodimerisasi dengan faktor transkripsi ARF).
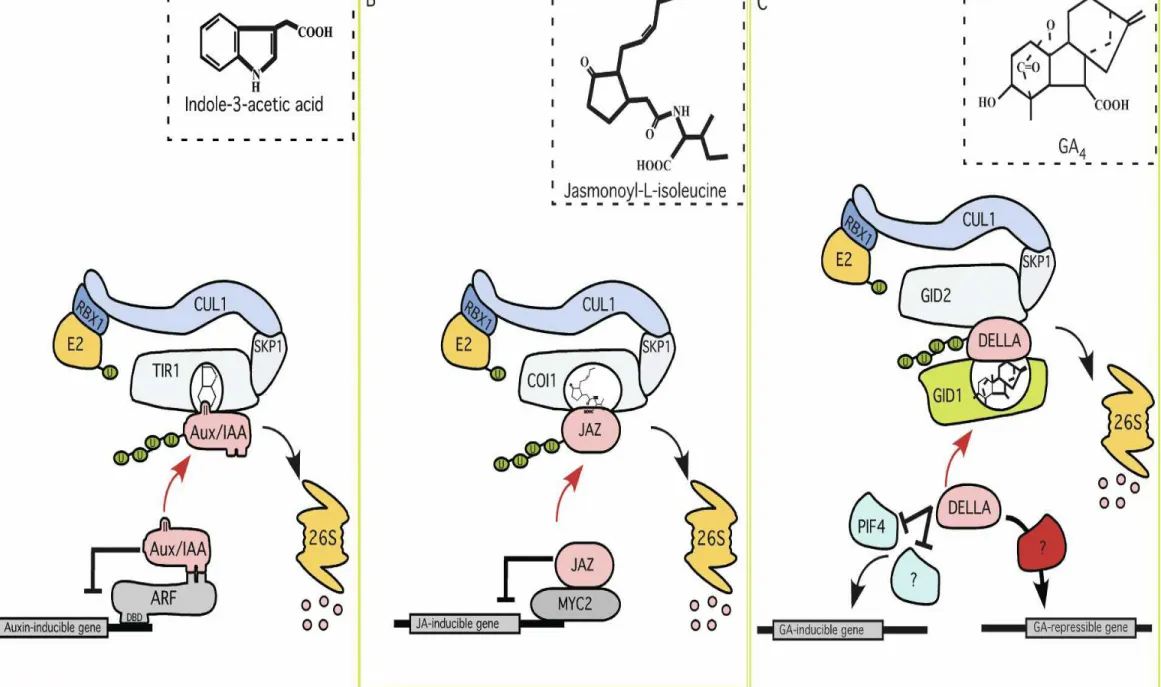
2. Setelah ada stimulus auksin, subunit protein TIR1 F-box (dari subunit SCF-TIR1 ubiquitin-ligase) akan mengikat auksin.
3. Hal tsb memungkinkan perekrutan protein Aux/IAA thd ubiquitinasi kompleks SCF.
4. Degradasi Aux/IAA dilakukan oleh
proteosom 26S shg faktor transkripsi ARF terbebaskan.
Hormon IAA-model ubiquitin-mediated
● Protein AUX/IAA tidak dapat berikatan langsung
dengan DNA pengikatan oleh ARF (Auxin Response factor)
● ARF wilayah promotor gen auksin -responsif
dapat mengaktifkan atau menekan kerja protein lain
● Protein AUX/IAA diketahui dapat menekan
Hormon IAA-model ubiquitin-mediated
TIR1 F-Box Protein : modul substrat pengenalan
subunit protein Skp1–Cullin1–F-box (SCF) ubiquitin-ligase (Petroski and Deshaies 2005)
Protein SCF-TIR1 :
1. Kompleks ikatan TIR1 dan protein SCF
(Scaffolding), berfungsi utk meningkatkan afinitas protein AUX/IAA
2. Menjadi jalur pengenalan ubiquitin-26S
Asam Jasmonat
1. Protein JAZ memiliki regulasi
negatif thd jasmonat menekan aktivitas
transkripsional MYC2.
2. Setelah pengikatan jasmonat,
SCFCOI1 ubiquitin-ligase mengikat protein JAZ proteolisis terfasilitasi oleh
Asam Jasmonat
● Protein JAZ tidak dapat berikatan langsung dgn
DNA regulator negatif thd aktivitas faktor transkripsi pengikatan oleh MYC2
● MYC2 : faktor transkripsi yang meregulasi
ekspresi gen penginsuksi jasmonat dan menekan ekspresi gen pengkode protein mutan jai3-1 (Chini et al. 2007).
● COI1 F-box protein : reseptor asam jasmonat
meningkatkan stabilitas pengikatan substrat protein-ligan hormonal
Hormon GA
1. Protein DELLA menekan GA regulasi negatif PIF3, PIF4, dan gen induksi GA lain.
2. Protein DELLA membantu ekspresi gen penekan GA (mengkode enzim biosintetis GA dan komponen jalur respons,
termasuk reseptor GID1.
3. Setelah pengikatan GA reseptor GID1 berinteraksi dgn DELLA. Kompleks tsb dikenali oleh SCFGID2 ubiquitin-ligase. Hal tsb merupakan target degradasi
terfsilitasi ubiquitin.
4. Warna panah merah efek respektif reseptor hormon Aux/IAA, JAZ, dan DELLA
Hormon GA
● Fitohormon diterpenoid-tetrasiklik : regulasi
pemanjangan batang, germinasi biji, dan
menginduksi pembungaan (Fleet and Sun 2005).
● GID2 dan SLY1 F-box protein : menyusun
kompleks ubiquitin-ligase bukan reseptor GA
● SLY1 F-box dan SCF/GID3 menambah
stabilitas pengikatan protein DELLA, berfungsi untuk menekan aktifitas respons GA regulator transkripsional negatif
Hormon GA
● Protein DELLA : berinteraksi dengan PIF3 dan
PIF4 mengikat DNA membentuk helix–loop– helix
● Fungsi kompleks DELLA sbg faktor transkripsi
yang meregulasi ekspansi sel sebagai respons cahaya dan GA.
● Region pengikatan DELLA: promotor pencegah
Asam Absisat (ABA)
Berperan dalam regulasi pematangan biji dan dormansi, pertumbuhan akar, senesen daun, dan transisi masa vegetatif-generatif.
Reseptor ABA bekerja pada lokasi sel berbeda Dalam nukleus:
1. Protein FCA pengikat RNA bekerjasama dgn faktor FY ending process mRNA 3’.
Berfungsi : mengontrol wilayah pengkodean FLC inhibitor pembungaan Setelah pengikatan ABA FCA dan FY terpisah terjadi akumulasi FLC
mengkode MADS box menghambat pembungaan. 2. Dalam kloroplas :
Subunit Mg-chelatase H (CHLH) mengikat ABA Berperan sebagai regulator positif
Asam Absisat (ABA)
● FCA (FLOWERING TIME CONTROL PROTEIN A) : reseptor ABA berperan dalam transisi masa vegetatif-generatif.
● G-protein-coupled receptor (GPCR) : persepsi situs pengikatan lokalisasi plasma membran.
● GCR1 dan RGS1 mengkode kandidat GPCR dan secara genetis memengaruhi respons ABA
(Pandey & Assmann 2004; Chen et al. 2006; Pandey et al. 2006).
● GPCR : reseptor ABA pada membran plasma, terdiri dari GCR1, GCR2, dan RGS1. Aktivitasnya belum diketahui.
Asam Absisat (ABA)
GPCR-related protein (GCR2) mengikat ABA. G-protein dan second messanger :
1. (PLD) phospholipase D;
2. (PP2C) protein phosphatase 2C; 3. (MAPK) mitogen-activated kinase; 4. (SnRK) SNF1-related kinase;
5. (PA) phosphatidic acid;
6. (ROS) reactive oxygen species; 7. (NO) nitric oxide.
Kesimpulan
● Peranan fitohormon dalam pertumbuhan dan
perkembangan pertumbuhan difasilitasi oleh reseptor hormon konsep dan sarana