HASIL DAN PEMBAHASAN
Hasil
A. HASIL ISOLASI DAN POLA KERAGAMAN PLAK FAGE EPEC K1.1 Hasil isolasi fage yang diambil dari sampel limbah cair rumah tangga di sepanjang Babakan Raya Darmaga Bogor pada mulanya diperoleh 9-18 plak serta adanya beberapa zona bening atau zona hambat lainnya di pinggir agar cawan yang ditumbuhi kultur EPEC K1.1. Plak yang terbentuk dari suatu kultur bakteri yang ditumbuhkan di cawan petri merupakan satu parameter penting dari adanya fage pada siklus litik. Plak tersebut terlihat bening yang menandakan adanya zona kerusakan sel.
(a) (b)
(c) (d)
Gambar 2 Pola keragaman plak fage. Isolat FB1 (a), FB2 (b), FB3 (c), FB4 (d) (masing-masing plak diberi tanda lingkaran).
Fage hasil isolasi dipilih menjadi 4 isolat fage secara random. Masing-masing isolat fage diberi nama FB1, FB2, FB3, dan FB4. Masing-Masing-masing isolat fage tersebut satu sama lain memiliki pola keragaman plak yang berbeda, seperti terlihat pada Gambar 2. Pola keragaman plak terlihat dari ukuran dan bentuk zona
bening dari masing-masing fage. Fage FB4 cenderung memiliki ukuran lebih besar dan lebih beragam daripada ketiga isolat fage lainnya (Gambar 2d).
B. HASIL PENENTUAN KISARAN INANG
Kisaran inang dari fage yang terisolasi spesifik terhadap bakteri EPEC K1.1. Hal ini terlihat dari pembentukan plak yang hanya tampak pada kultur EPEC K1.1 sedangkan pada E. coli non patogen tidak tampak adanya pembentukan plak (Gambar 3b).
(a) (b)
Gambar 3 Kisaran inang fage. Plak muncul pada fage yang diinfeksikan dengan EPEC K1.1 (a) (diberi tanda lingkaran); sedangkan pada E. coli non patogen tidak muncul adanya plak (b).
C. HASIL KUANTIFIKASI FAGE
Dari 100 µl kultur EPEC yang diinfeksikan dengan 100 µl fage menghasilkan plak yang terbentuk (PFU) dengan jumlah atau kuantitas seperti yang terlihat pada Tabel 1.
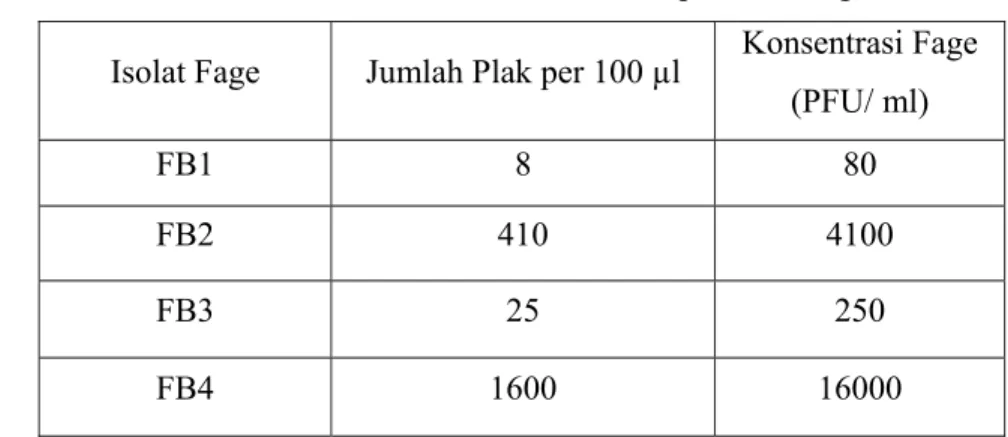
Tabel 1 Hasil kuantifikasi dari keempat isolat fage
Isolat Fage Jumlah Plak per 100 µl Konsentrasi Fage
(PFU/ ml)
FB1 8 80 FB2 410 4100 FB3 25 250 FB4 1600 16000
D. HASIL KARAKTERISASI PROTEIN FAGE
Kadar protein masing-masing fage berbeda satu sama lain. Pada Tabel 1.. terlihat bahwa FB 4 memiliki kadar protein paling tinggi dibanding fage yang lain sebanyak 222.5 µg/ ml sedangkan FB1, FB2 dan FB3 secara berurutan memiliki konsentrasi protein sebesar 97.5; 82.5 dan 170µg/ ml.
Tabel 2 Konsentrasi protein keempat isolat fage
Isolat Fage Konsentrasi Protein
(µg/ ml)
FB1 97.5 FB2 82.5 FB3 170 FB4 222.5
Analisis Bobot Molekul Protein Fage
Hasil analisis bobot molekul protein fage dengan menggunakan Sodium
Dodecyl Sulphate-Poly Acrilamide Gel Electrophoresis (SDS-PAGE) terlihat
pada Gambar 4. Analisis pada SDS-PAGE dilakukan setelah terlebih dahulu menentukan kadar protein fage dengan menggunakan metode Bradford (1976). Dengan volume larutan fage yang sama pada masing-masing sumur gel menunjukkan bahwa FB3 menampakkan pita protein pada ukuran 51.8 kDa; 42.6 kDa; 34.6 kDa; dan 30.8 kDa sedangkan FB4 menampakkan pita protein pada
ukuran 40.3 kDa dan 35.9 kDa. Pada FB 1 dan FB2 tidak menampakkan adanya pita protein, mungkin karena rendahnya kadar protein fage tersebut.
Gambar 4 Kisaran berat molekul protein fage pada SDS-PAGE . 1) Marker. 2) Isolat FB4, dengan ukuran pita protein sebesar 40.3 kDa (a) dan 35.9 kDa (b). 3) Isolat FB3, dengan ukuran pita protein sebesar 51.8 kDa
(c), 42.6 kDa (d), 34.6 kDa (e), 30.8 kDa (f).
E. HASIL PENGAMATAN MORFOLOGI FAGE DENGAN TEM
Analisis morfologi dengan menggunakan TEM pada FB4 menunjukkan kepala fage yang berbentuk heksagonal ikosahedral. Lebar kepala fage sebesar 81.56 nm dan panjangnya sebesar 103.11 nm, seperti terlihat pada Gambar 5.
(a) (b)
Gambar 5 Morfologi fage FB 4, perbesaran 80000x (a), kepala fage yang diperbesar menunjukkan bentuk heksagonal ikosahedral (b). Tanda ( ) menunjukkan kepala.
1 2 116.0 kDa 66.2 kDa 45.0 kDa 35.0 kDa a b c d e f 3
F. EFEKTIFITAS FAGE MELISIS EPEC K1.1
Infeksi fage terhadap jumlah pertumbuhan EPEC K1.1 selama 24 jam dengan selang waktu rata-rata 5 jam, terlihat pada Gambar 6. Kurva ini menunjukkan penurunan jumlah populasi sel EPEC K1.1 pada 1 jam pertama dan semakin tampak penurunannya setelah 5 jam berikutnya. Hal ini tampak berbeda dengan perlakuan kontrol yang baru menunjukkan penurunan jumlah populasi sel EPEC K1.1 setelah 10 jam. Penambahan FB4 pada kultur EPEC K1.1 yang
diinkubasi pada suhu 370 C selama 5 jam dapat menurunkan jumlah populasi
EPEC K1.1 sebanyak 22% dan 84 % setelah inkubasi selama 24 jam.
(a) (b)
Gambar 6 Kurva efektifitas lisis sel EPEC K1.1 oleh FB4. EPEC K1.1 yang diinfeksi oleh FB4 (a) dan yang tanpa fage (sebagai kontrol) (b)
selama 24 jam.
Morfologi Sel EPEC K1.1 akibat infeksi fage
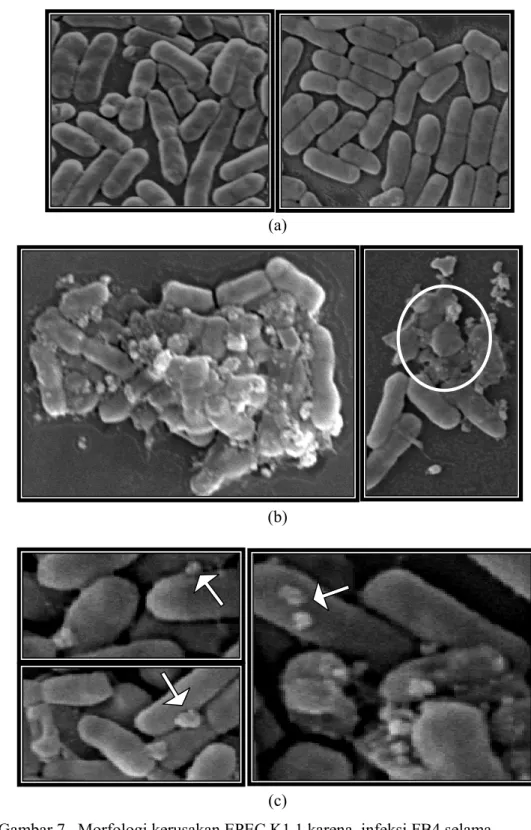
Hasil pengamatan di SEM menunjukkan adanya pengaruh atau efek dari penginfeksian fage FB4 terhadap morfologi sel EPEC K1.1. Berdasarkan hasil pengamatan di SEM, tampak sejumlah besar fage yang menginfeksi sel EPEC K1.1. Fage yang menginfeksi beberapa EPEC K1.1 menyebabkan beberapa EPEC K1.1 menjadi lisis melepaskan banyak partikel fage baru. Pada sel-sel EPEC K1.1 yang tidak diinfeksi oleh fage, sebagai kontrol positifnya (Gambar 7a), tampak morfologi sel yang masih utuh dan rapat. Pengaruh penginfeksian fage FB4 terhadap morfologi sel EPEC K1.1 terlihat pada Gambar 7 dan Gambar 8.
EPEC K1.1 yang Diinfeksikan oleh Fage
0 100 200 300 400 500 600 0 5 10 15 20 25 30 Waktu (jam) P er kir aa n J u m la h K o l
EPEC K1.1 yang tidak Diinfeksikan oleh Fage
0 100 200 300 400 500 600 0 5 10 15 20 25 30 Waktu (jam) P er ki raa n Ju m la h K o l
(a)
(b)
(c)
Gambar 7 Morfologi kerusakan EPEC K1.1 karena infeksi FB4 selama 25 menit. Sel EPEC K1.1 tanpa fage (a); sel EPEC K1.1 yang diinfeksi oleh fage dengan inkubasi 25 menit (b), (c). Tanda panah menunjukkan fage yang menginfeksi EPEC K1.1, tanda lingkaran menunjukkan kerusakan dan lisisnya sel EPEC K1.1.
Hasil pengamatan di SEM, terlihat bahwa FB4 dapat memecahkan/ melisiskan sel EPEC K1.1 dalam waktu 25 menit inkubasi (Gambar 7b). Dalam waktu lebih lama lagi yaitu 30 menit, morfologi sel EPEC K1.1 tampak banyak yang sudah hancur atau lisis melepaskan fage-fage baru kemudian menginfeksi bakteri-bakteri lain. Pada permukaan dinding sel terlihat sudah tidak utuh lagi atau rusak. Hancuran dari sel EPEC K1.1 tampak bergabung dengan partikel fage yang semakin banyak keluar dari sel inang (Gambar 8b).
(a)
(b)
Gambar 8 Morfologi kerusakan EPEC K1.1 karena infeksi FB4 selama 30 menit (a), (b). Tanda panah menunjukkan fage yang menginfeksi EPEC K1.1, tanda lingkaran menunjukkan kerusakan dan lisisnya sel EPEC K1.1.
Pembahasan
Fage tidak mampu bereplikasi sendiri tanpa adanya sel inang sehingga dalam melakukan pengisolasian fage perlu dicampurkan dengan bakteri sebagai inangnya virus. Metode isolasi fage yang dilakukan ialah dengan metode
enrichment yaitu memperkaya nutrisi pada media pertumbuhan bakteri agar virus
dapat bereplikasi di dalam tubuh bakteri dan dapat dengan mudah diperoleh dalam jumlah yang banyak. Fage dapat diisolasi dan dikultivasi pada biakan bakteri yang muda (pada fase logaritma) dan sedang tumbuh aktif dalam kaldu atau cawan agar. Penggunaan media padat memungkinkan timbulnya plak sehingga dapat dilakukan pendeteksian atau penghitungan virus bakteri (fage). Persyaratan utama bagi isolasi dan kultivasi fage ialah harus adanya kondisi optimum untuk pertumbuhan organisme inangnya (Pelczar & Chan 1988) sehingga pada
penginfeksian fage dengan EPEC K1.1 dilakukan pada suhu 37 0C. Pada proses
pemurnian fage dilakukan dengan penambahan kloroform yang mengandung 1% etanol. Hal ini bertujuan untuk membunuh sisa-sisa bakteri agar diperoleh fage murni tanpa adanya inang. Kennedy et al. (1984) dalam Pierson dan Stern (1986) menyebutkan bahwa penggunaan kloroform itu merugikan bagi beberapa kolifage sehingga penggunaannya bisa diminimalisir dengan memproduksi fage pada suatu media selektif. Penginfeksian terhadap inang yang selanjutnya digunakan, penting dilakukan untuk mendeteksi viabilitas fage setelah penyimpanan. Penentuan jumlah sel ekstraseluler dilakukan dengan sentrifugasi campuran bakteri dengan fage. Jika berdasarkan ukuran fage yang lebih kecil daripada bakteri maka sejumlah fage diharapkan banyak berada pada bagian supernatan.
Hasil isolasi fage dari sampel limbah cair rumah tangga daerah Babakan Raya menghasilkan 9-18 plak dan dari plak tersebut terpilih 4 isolat fage (FB1, FB2, FB3, dan FB4) secara random. Fage yang diinfeksikan ke bakteri EPEC K1.1 (bakteri uji) dikondisikan dalam siklus litik karena pada siklus litik, tahap adsorpsi, penetrasi, sintesis, dan pematangan berlangsung dengan cepat. Pada siklus ini pula partikel virus keluar dari sel yang diserangnya dengan memecahkan sel tersebut sehingga sel inang mati (lisis). Plak yang terbentuk dari suatu kultur bakteri yang ditumbuhkan di cawan petri merupakan satu parameter penting dari
adanya fage pada siklus litik. Plak tersebut terlihat bening yang menandakan adanya zona kerusakan sel. Setiap plak berasal dari satu partikel fage sama seperti setiap koloni berasal dari satu sel bakteri. Satu plak berasal dari satu partikel virus sehingga seluruh partikel virus yang terdapat pada plak tersebut seharusnya juga memiliki sifat genetik yang sama. Pada saat terbentuknya plak, bakteri yang tidak terkena infeksi tersebar di tempat lain di dalam agar cawan dan menghasilkan suatu background yang keruh.
Proses isolasi fage dilakukan dengan berbagai perlakuan sampai dengan pemurnian untuk memastikan suatu populasi fage yang murni tanpa adanya beberapa bakteri termasuk inang bakteri yang tahan fage. Hal ini juga dianggap penting karena populasi fage terdiri atas beberapa strain fage dengan satu karakteristik umum. Masing-masing fage menginfeksi pada inang tertentu sehingga sangat diperlukan tahapan untuk memperoleh strain murni (Goodridge
et al. 2003).
Fage (FB1, FB2, FB3, dan FB4) yang terisolasi, spesifik terhadap EPEC K1.1 karena fage-fage tersebut mampu mereduksi jumlah EPEC K1.1. Keempat isolat fage yang spesifik terhadap bakteri EPEC K1.1 diduga merupakan isolat yang tidak serupa atau tidak identik. Berdasarkan pengamatan secara visual, ukuran plak yang terbentuk dari satu isolat fage tampak berbeda dengan isolat fage yang lain (Gambar 2). Hal ini menandakan fage-fage tersebut berbeda dalam cara melisiskan sel inang. Hasil penelitian Yoon et al. (2007) yang menggunakan fage Pediococcus juga menunjukkan perbedaan ukuran plak antara beberapa fage
Pediococcus. Ukuran lisisan tersebut signifikan dengan jumlah partikel fage yang
dibebaskan dari sel. Rata-rata dari setiap lisisan melepaskan lebih kurang 60 partikel fage tiap sel bakteri (Ellis & Delbruck 1938). Berdasarkan hasil penelitian tersebut, ukuran pembentukan plak masing-masing fage dari hasil menginfeksi sel inang yang sama berkaitan dengan perbedaan cara merespon sel bakteri EPEC terhadap asam nukleat fage yang disisipkan atau kemampuan fage dalam bereplikasi di dalam sel inang. Pola lisis suatu galur bakteri yang diakibatkan oleh tipe fage yang berbeda-beda memberikan petunjuk mengenai bakteri tersebut dan untuk penentuan tipe fage (phage typing) (Pelczar & Chan 1988). Fage FB4 memiliki ukuran zona bening lebih besar dan lebih beragam daripada ketiga isolat
fage lainnya. Hal ini menandakan FB4 memiliki kemungkinan sebagai fage infektif kuat.
Kepekaan galur bakteri terhadap fage yang menyerangnya berbeda-beda yang diakibatkan variasi molekul reseptor (penghalangan adsorpsi), sistem modifikasi restriksi dalam sel inang, atau sistem ketahanan fage lainnya seperti infeksi abortif (Flynn et al. 2004). Fage melekat ke sel yang peka rangsangan pada lokasi spesifik di dinding sel bakteri (Kudva et al. 1999). Di dalam bakteri gram negatif, sel yang peka rangsangan ialah komponen protein dan lipopolisakarida yang melapisi lapisan selaput sebelah luar termasuk peptidoglikan. Fage tertentu atau sekelompok fage akan melekat ke reseptor spesifik, dan fage berbeda akan melekat ke reseptor yang berbeda. Beberapa bagian struktural bakteri seperti flagella, pilus, kapsul, teichoicacid, LPS, dan OMP dapat menyediakan reseptor spesifik dalam bakteri tertentu (Gurnev et al. 2006). Penentuan kisaran inang dilakukan untuk melihat kespesifisitas inang dan derajat lisis dari fage yang diperoleh. Masing-masing isolat fage memperlihatkan spesifisitas terhadap EPEC K1.1 karena ketika fage yang diperoleh diuji dengan masing-masing isolat EPEC K1.1 dan E. coli non patogen, ternyata kisaran inangnya hanyalah pada isolat EPEC K1.1. Sel EPEC K1.1 tampak dilisiskan oleh fage (Gambar 3a) sedangkan
E. coli non patogen tidak (Gambar 3b). Kespesifikan isolat fage yang diperoleh
terhadap EPEC K1.1 menunjukkan dugaan bahwa di permukaan sel EPEC K1.1 memiliki reseptor-reseptor yang spesifik terhadap fage yang tidak dimiliki oleh E.
coli non patogen.
Adsorpsi partikel-partikel fage terhadap sel-sel bakteri (tahap awal infeksi fage) bergantung pada kehadiran reseptor-reseptor spesifik di dinding sel bakteri (Topley & Wilson 1990). Banyak reseptor-reseptor dinding sel dimiliki oleh galur dan serotipe bakteri yang berbeda. Berdasarkan hasil penelitian Kudva et al. (1999), infeksi fage bergantung pada sifat alami LPS O157. Bakteri Escherichia
coli O157 dapat menghasilkan LPS O157. Jika komplemen E. coli mutan O157-deficient yang ujung gen penyandi LPS 0157 untuk bakteri tersebut dipotong
maka akan membuat bakteri resisten terhadap infeksi fage.
Cara penghitungan yang paling mudah ialah esei bercak/ plak (plaque
terbentuk adalah unik di dalam kemampuan mereka untuk mengidentifikasi dan membeda-bedakan galur inang bakteri. Masing-Masing plak secara teoritis sesuai dengan virus tunggal yang berada pada suspensi awal. Oleh karena itu, konsentrasi suspensi virus diukur oleh banyaknya plak yang biasanya disebut
plaque forming unit (PFU) (Tortora et al. 2006). Penelitian tentang fage seringkali
memerlukan penghitungan fage secara cermat dan teliti. Hasil kuantifikasi atau penghitungan plak yang terbentuk (PFU) (Tabel 1) setelah dilakukan satu kali perbanyakan dalam proporsi yang sama (100 µl kultur EPEC diinfeksikan dengan 100 µl fage) menunjukkan bahwa FB4 memiliki kemampuan infeksi terkuat terhadap EPEC K1.1 dibandingkan ketiga isolat fage yang lain. Kecepatan pembentukan plak ini diduga berkaitan pula dengan kecepatan multiplikasi dari FB4 sehingga lebih efektif dalam menginfeksi sel EPEC K1.1 dibandingkan ketiga isolat lainnya. Hal ini memungkinkan peluang FB4 untuk dapat diaplikasikan dalam biokontrol pencemaran air dan makanan.
Protein yang dimiliki oleh keempat isolat fage cenderung kecil. Dalam tiap mililiternya, FB1 hanya memiliki 97.5 µg; FB2 82.5 µg; FB3 170 µg; dan FB4 222.5 µg (Tabel 2). Setelah dijalankan dengan SDS-PAGE, terdapat 2 isolat fage (FB3 dan FB4) yang menunjukkan adanya pita protein. Kisaran protein yang diperoleh antara 30-51 kDa (Gambar 4). Protein fage yang diukur kadar proteinnya tersebut merupakan hasil produksi yang dilakukan berkali-kali. Berdasarkan hasil yang telah diperoleh menunjukkan bahwa ukuran kadar protein fage tidak memberikan adanya signifikansi dengan jumlah partikel fage. Konsentrasi protein yang cenderung lebih kecil diduga disebabkan oleh mudahnya protein fage terdegradasi oleh protease ekstraselular yang dimiliki sel EPEC K1.1. Selain itu, ukuran protein yang kecil mungkin saja disebabkan oleh konsentrasi protein fage EPEC itu sendiri yang memang lebih kecil sehingga walaupun dalam penelitian ini sudah digunakan EDTA 5 mM sebagai tambahan pada media penyimpanan fage yang berfungsi sebagai antiprotease (Waturangi 1999) namun tetap saja kurang berpengaruh. Hal ini hampir serupa dengan penelitian Yoon et
al. (2007), dimana setelah dianalisis dengan SDS-PAGE menunjukkan kisaran
ukuran protein 33-34.5 kDa untuk fage yang menginfeksi Leuconostoc oenos dan 34-37 kDa untuk fage yang menginfeksi P. acidilactici.
Berdasarkan karakteristik isolat FB4 seperti dari ukuran lisisan yang lebih besar; kemampuan dalam menginfeksi sel EPEC K1.1 yang lebih baik; serta lebih tingginya konsentrasi protein FB4 diantara ketiga isolat fage yang lain, maka isolat FB4 yang selanjutnya diamati penampakan morfologinya.
Penampakan morfologi dari fage yang terpurifikasi diperiksa dengan menggunakan Transmission Electron Microscopy (TEM). Pemeriksaan dengan menggunakan TEM dapat memperlihatkan bentuk morfologi fage secara 2 dimensi dengan perbesaran 50000x dan 80000x. Analisis morfologi dengan menggunakan TEM pada FB4 (Gambar 5) menunjukkan morfologi kepala fage yang besar dan berbentuk heksagonal ikosahedral. Lebar kepala fage berukuran sebesar 81.56 nm dan panjang kepala fage sebesar 103.11 nm. Jika hanya berdasarkan ciri-ciri morfologi yang dimiliki isolat fage FB4, maka morfologi fage tersebut hampir serupa dengan banyak fage dari famili Siphoviridae ordo
Caudovirales. Berdasarkan klasifikasi dari the International Committee on Taxonomy of Viruses (ICTVdB Index of Viruses 2000), morfologi fage yang
termasuk famili Siphoviridae memiliki ekor fleksibel, tidak memiliki selaput kontraktil, panjang, heliks dan kepala heksagonal ikosahedral seperti fage λ
enterobacteria pada umumnya. Menurut hasil penelitian Girons et al. (1990),
berdasarkan hasil pengamatan di mikroskop elektron dengan pewarnaan negatif, fage untuk Leptospira terlihat identik secara morfologi dengan kepala polihedral dan ekor kontraktil. Rata-rata diameter kepalanya ialah 85 nm, dengan panjang ekor 100 nm, lebar 25 nm. Morfologi ini serupa dengan banyak fage eubacteria yang masuk dalam kelompok A1 famili Myoviridae (Ackermann & DuBow 1987).
Pengujian efektifitas infeksi bakteriofage terhadap sel EPEC K1.1 bertujuan agar dapat diketahui dengan tepat waktu yang dibutuhkan bakteriofage dalam mengontrol berkembangbiaknya EPEC pada tubuh inang. Sebanyak 1000 plak dalam 100 µl fage yang setara dengan 10000 PFU telah memberikan pengaruh terhadap pertumbuhan EPEC K1.1. Pengaruh pemberian fage terhadap pertumbuhan EPEC K1.1 memberikan penurunan populasi sel bakteri EPEC K1.1 pada 1 jam pertama dan semakin tampak menurun setelah 5 jam berikutnya (Gambar 6a). Hal ini berbeda dengan pertumbuhan EPEC K1.1 yang normal
(tanpa diinfeksikan oleh fage) karena populasi sel bakteri EPEC K1.1 baru terjadi penurunan setelah 10 jam (Gambar 6b). Berdasarkan penelitian Hughes et al. (1998), fage dapat mendegradasi exopolysaccharide (EPS) yang dihasilkan oleh biofilm dari spesies bakteri Enterobacter agglomerans dan menginfeksi selnya. Proses perusakan biofilm pada bakteri Enterobacter agglomerans oleh fage merupakan suatu kombinasi penurunan EPS oleh depolymerase dan glycanase yang dihasilkan oleh fage dengan proses penginfeksian fage yang selanjutnya melisiskan sel tersebut. Depolimerase ini tampak berlanjut ke permukaan fage dan mendegradasi polisakarida kapsular bakteri untuk diteruskan ke permukaan sel
bakteri. Penambahan FB4 pada kultur EPEC K1.1 yang diinkubasi pada suhu 370
C selama 5 jam dapat menurunkan jumlah populasi EPEC K1.1 sebanyak 22% dan 84 % setelah inkubasi selama 24 jam. Penurunan awal terhadap jumlah sel bakteri diperkirakan merupakan akibat dari kombinasi aksi enzim polysaccharide
depolymerase atau lisozim dari bakteriofage dalam mendegradasi polimer
lipopolisakarida (LPS) sel EPEC K1.1 dengan proses penginfeksian terhadap suatu proporsi populasi bakteri EPEC K1.1.
Diantara sel-sel bakteri yang telah diinfeksi oleh fage selama lebih dari 3 hari, ditemukan adanya sel-sel bakteri EPEC K1.1 yang masih resisten terhadap pembentukan plak oleh infeksi fage-fage ini. Faktor yang mungkin mengkontribusi ketahanan fage diantaranya perubahan atau kehilangan reseptor (Alisky et al. 1998; Barrow & Soothill 1997).
Seperti pada kolifage lainnya, fage yang spesifik terhadap EPEC K1.1 bersifat litik. Multiplicity of Infection (MOI) yang tinggi akan membantu mempercepat lisisnya sel-sel inang. Multiplicity of Infection yang tinggi diperlukan untuk mengukur bahwa setiap bakteri diinfeksikan sedikitnya oleh satu fage. Berdasarkan hasil penelitian Ma & Lu (2008), pada fage yang menginfeksi
Streptococcus suis diperoleh MOI yang optimal dari fage sebesar 0.1 sehingga
MOI yang optimal dari fage EPEC juga ditentukan sebesar 0.1. Selain MOI, aerasi kultur berperan penting dalam mempercepat induksi fage dan memperbanyak hasil pelisisan bakteri. Proses aerasi dapat memberikan kondisi yang memungkinkan bagi fage untuk berinteraksi dengan bakteri. Perlakuan fage pada
suhu 37 0C tanpa aerasi hanya menghasilkan kematian atau lisis bakteri setelah 2 hari (Ellis & Delbruck 1938).
Pertumbuhan fage dapat dibagi dalam 3 periode, yaitu absorbsi fage terhadap bakteri; pertumbuhan ketika atau di dalam bakteri (periode laten); dan pelepasan fage (terjadi proses lisis) (Ellis & Delbruck 1938). Pada penelitian Yoon et al. 2007, yang meneliti bakteriofage litik yang menginfeksi Pediococcus diperoleh periode eclipse dan latent terjadi pada 29 menit dan 34 menit secara berturut-turut. Ukuran lisisan terhitung rata-rata 12 partikel fage per pusat infeksi. Hasil ini menunjukkan bahwa perkembangan intraselular dimulai tidak lama setelah proses infeksi terjadi. Pada fage yang menginfeksi Streptococcus suis diperoleh satu tahap kurva pertumbuhan fage yang memperlihatkan periode latent pada 20 menit dan periode rise pada 120 menit , serta ukuran lisisan terhitung rata-rata 77 PFU/ sel (Ma & Lu 2008). Berdasarkan hasil penelitian-penelitian tersebut, pertumbuhan fage EPEC juga diperkirakan memiliki suatu periode latent pada kisaran waktu antara 20 sampai 30 menit.
Pengamatan keadaan morfologi sel EPEC K1.1 akibat pengaruh penginfeksian fage pada masa inkubasi 25 menit dan 30 menit dilakukan dengan menggunakan Scanning Electron Microscope (SEM). Scanning electron
microscope yang digunakan adalah JEOL LTD, yaitu mikroskop elektron yang
dapat merekam gambar obyek menjadi elemen terang dan gelap tipe hampa udara rendah. Modelnya ialah model-JSM 5310 LV. Cara kerja dari mikroskop ini adalah pancaran cahaya elektron dengan fokus yang sangat tajam disapukan pada obyek sehingga menghasilkan elektron sekunder. Sinyal-sinyal ini dideteksi terus-menerus selama pancaran cahaya elektron bergerak menyapu permukaan obyek. Sinyal elektron sekunder menghasilkan gambar permukaan morfologi elektron yang terpental kembali lalu menyebar menghasilkan distribusi komposisi dan karakteristik sinar X menghasilkan distribusi elemen yang terdapat pada obyek. Jadi, tujuan pengamatan dengan SEM ialah mengamati seberapa besarnya kerusakan sel EPEC dengan keadaan membran yang rusak dan dinding sel yang terpecahkan akibat diinfeksi oleh fage.
Bakteri EPEC K1.1 yang ditumbuhkan pada kondisi pertumbuhan optimal diinfeksikan dengan fage. Campuran EPEC K1.1 dengan fage tersebut ditunggu
selama 25 menit dan 30 menit sebelum dipreparasi untuk diamati di SEM. Isolat bakteri EPEC K1.1 yang baru ditumbuhkan dan tanpa diinfeksikan oleh fage digunakan sebagai kontrol. Bila suspensi fage diinfeksikan pada suspensi bakteri yang peka, maka partikel fage akan melekatkan diri pada sel menggunakan serabut ekor fage (Gambar 7c). Serabut ekor berkontraksi sehingga terjadi cengkraman bagian paku ekor pada membran sel bakteri (Bayer 1968). Pada permukaan sel terjadi perubahan yang diduga kuat disebabkan daya kerja enzim yang dihasilkan ekor fage dalam merusak dinding sel bakteri sehingga terbentuk pori-pori (Gambar 8a) yang sangat halus atau karena aktivasi enzim inang oleh fage. Proses ini diikuti oleh kontraksi serabut ekor sehingga DNA virus merasuk ke dalam sel melalui pori-pori pada ujung ekor. Selubung protein virus tetap berada di luar sel bakteri (Gambar 7c). Ekor yang diperpanjang adalah suatu pelindung kontraktil yang bertindak sebagai suatu jarum suntik untuk menyemburkan muatan yang ada di kepalanya, seperti material genetik DNA ke dalam sel inang. DNA fage kemudian mengambil alih DNA sel bakteri dan memaksa untuk memproduksi lebih banyak fage. Hal ini memerintahkan sel inang untuk membuat salinan-salinan fage (keturunan di dalam sel). Selanjutnya fage tersebut merusak DNA inang dengan cara melepaskan nukleotida yang bersifat memblok DNA inang dari DNA fage yang disatukan. Material genetik tersebut kemudian dibungkus oleh keturunannya dan kemudian fage dilepaskan ke lingkungan ketika melisiskan sel inang (Wendelschafer-crabb et al. 1975). Hasil pengamatan di SEM memperlihatkan adanya pengaruh atau efek dari penginfeksian fage FB4 yaitu FB4 dapat memecahkan/ melisiskan EPEC K1.1 dalam waktu 25 menit. Dalam waktu lebih lama lagi yaitu 30 menit, sel EPEC terlihat hancur. Hal ini menunjukkan bahwa FB4 secara pasti dapat melisiskan bakteri sasaran EPEC K1.1 resisten tetrasiklin dan ampisilin. Penemuan ini merupakan titik cerah untuk pengendalian bakteri patogen pencemar makanan dan air yang ramah lingkungan.