PENDAHULUAN
Latar BelakangBahan pakan sumber protein di Indonesia sangat banyak macamnya dan beragam kualitasnya. Untuk menyusun satu macam ransum biasanya digunakan beberapa macam bahan. Bila dilihat dari segi proteinnya, maka ransum yang disusun hanya berdasarkan kadar protein saja menjadi kurang akurat. Hal ini dapat dilihat di lapang untuk ransum yang berbeda namun mengandung kadar energi dan protein yang sama, sering kali menghasilkan tingkat produktivitas yang berbeda-beda. Ransum dengan kadar protein yang sama bisa jadi memiliki tingkat fermentabilitas, ketahanan protein terhadap degradasi dalam rumen, kecernaan protein oleh enzim pencernaan pascarumen dan sintesis protein mikroba yang berbeda-beda. Oleh karena itu kadar protein ransum yang tinggi tidak dapat menjamin bahwa ransum tersebut berkualitas.
Pemberian protein ransum hendaknya didasarkan pada banyaknya protein yang dapat diabsorbsi (NRC, 1989). Pada ruminansia jumlah protein yang dapat diabsorbsi bergantung pada pasokan protein mikroba dan protein ransum. Oleh karena itu perlu dikembangkan tolok ukur mutu protein yang ada keterkaitannya dengan pasokan protein tersebut. Mutu protein ransum ditentukan oleh banyak faktor antara lain: (i) Produksi amonia, karena amonia rumen diperlukan untuk mendukung sintesis mikroba rumen. Pada ruminansia, protein yang dikonsumsi sebagian akan didegradasi dalam rumen menjadi asam amino dan selanjutnya mengalami deaminasi menjadi amonia (NH3) dan sebagian lainnya akan lolos dari perombakan rumen. Amonia sebagai sumber nitrogen (N) diperlukan untuk sintesis de novo asam amino oleh mikroba rumen. Kadar amonia cairan rumen yang mendukung sintesis mikroba rumen berkisar antara 4-14 mM (Satter dan Slyter 1974; Preston dan Leng 1987; Sutardi 1994). (ii) Tingkat degradasi protein dalam rumen, yang merupakan indikator besarnya protein yang memasok protein asal ransum (Orskov et al. 1980). Daya degradasi protein ransum di dalam rumen sangat bervariasi bergantung pada komposisi, struktur fisik dan kimia, pengolahan bahan baik secara kimia, fisik maupun biologis (Madsen dan Hvelplund 1985), (iii) Kecernaan protein oleh
pepsin HCl (AOAC 1980; Calsamiglia dan Stern 1995; Habib et al. 2001), sebagai pemasok asam amino pascarumen. Protein ransum yang tak terdegradasi dalam rumen dengan kecernaan oleh enzim pencernaan pascarumen yang tinggi diperlukan untuk menyediakan protein ransum bagi induk semang. Hal ini sehubungan dengan pasokan asam amino untuk ternak dengan tingkat produksi tinggi tidak cukup jika hanya mengandalkan pasokan yang berasal dari protein mikroba. Sebaliknya protein ransum yang tak terdegradasi dalam rumen, tetapi tidak dapat dicerna oleh enzim pencernaan pascarumen akan dikeluarkan melalui feses. Oleh karena itu tingkat ketahanan protein ransum terhadap degradasi dalam rumen sekaligus kecernaan protein oleh enzim pencernaan pascarumen menjadi penting untuk diperhitungkan dalam menentukan kualitas protein ransum ruminansia. (iv) Produksi basa purin cairan rumen (Zinn dan Owens 1986; Chen dan Gomez 1992; Obispo dan Dehority 1999) untuk melihat besarnya sintesis mikroba rumen, yang merupakan komponen pemasok protein asal mikroba rumen. Semakin tinggi basa purin yang dihasilkan menunjukkan semakin tinggi nilai hayati dari protein ransum. Mikroba rumen merupakan sumber pemasok N yang utama bagi ruminansia karena mampu mencukupi kebutuhan N sebesar 40 – 80% (Sniffen dan Robinson 1987).
Pengukuran mutu protein ransum perlu mempertimbangkan cara yang mudah, cepat dan murah serta mempunyai relevansi tinggi dengan retensi nitrogen dan pertumbuhan, sehingga tolok ukur mutu protein tersebut bisa digunakan untuk menduga produktivitas. Pengukuran dengan menggunakan hewan percobaan (in vivo) dapat diketahui secara langsung besarnya retensi nitrogen maupun pertumbuhannya, akan tetapi dibutuhkan biaya yang mahal dan membutuhkan waktu lama. Pengukuran secara in vitro dan in sacco dapat dilakukan dalam waktu yang cepat, sampel yang dibutuhkan sedikit, kondisi relatif homogen dan dapat dikontrol serta biaya lebih murah.
Untuk menjawab hal tersebut dilakukan serangkaian percobaan yang menyangkut pengkajian beberapa faktor yang mempengaruhi masing-masing tolok ukur, baik secara in vitro, in sacco dan in vivo. Parameter mutu protein ransum in vitro meliputi produksi amonia, kecernaan pepsin dan produksi purin, sedangkan secara in sacco dapat diukur tingkat degradasi protein dalam rumen.
Selanjutnya dilakukan analisis relevansi masing-masing tolok ukur terhadap retensi dan pertumbuhan domba dengan analisis regresi.
Tujuan
1 Mengkaji pengaruh sumber protein ransum terhadap utilisasi protein berdasarkan tinjauan beberapa parameter mutu protein.
2 Menentukan parameter mutu protein ransum yang erat kaitannya dengan retensi nitrogen dan pertumbuhan domba.
Manfaat
1 Dengan adanya relevansi antara produksi amonia, degradasi protein dalam rumen, kecernaan protein tak terdegradasi dalam rumen oleh pepsin HCl dan produksi purin dengan retensi nitrogen serta pertumbuhan domba, maka parameter tersebut dapat digunakan sebagai tolok ukur mutu protein ransum.
2 Tolok ukur mutu protein tersebut dapat digunakan sebagai pedoman untuk memilih bahan yang sesuai dalam menyusun ransum guna mencapai pertumbuhan domba seperti yang diharapkan.
Hipotesis
Produksi amonia, degradasi protein dalam rumen, kecernaan protein tak terdegradasi dalam rumen oleh pepsin HCl dan produksi purin dapat digunakan sebagai tolok ukur mutu protein ransum, karena berpengaruh pada retensi nitrogen dan pertumbuhan domba.
Pencernaan dan Metabolisme Karbohidrat serta Lemak pada Ruminansia
Proses pencernaan adalah suatu proses perubahan yang dialami bahan makanan baik secara fisik maupun kimiawi di dalam saluran pencernaan menjadi zat-zat yang lebih sederhana yang dipersiapkan untuk diabsorbsi dan digunakan oleh ternak untuk memenuhi kebutuhannya. Proses pencernaan pada ruminansia lebih kompleks dibandingkan dengan non ruminansia, yaitu meliputi interaksi antara pakan dengan populasi mikroba dan ternak itu sendiri. Proses pencernaan pada ruminansia terjadi secara mekanis (di mulut), fermentatif (oleh enzim yang dihasilkan oleh mikroba rumen) dan hidrolisis (oleh enzim-enzim hewan induk semang). Oleh karena itu ruminansia mempunyai kemampuan yang lebih baik dalam mencerna zat-zat makanan terutama yang berasal dari makanan berserat.
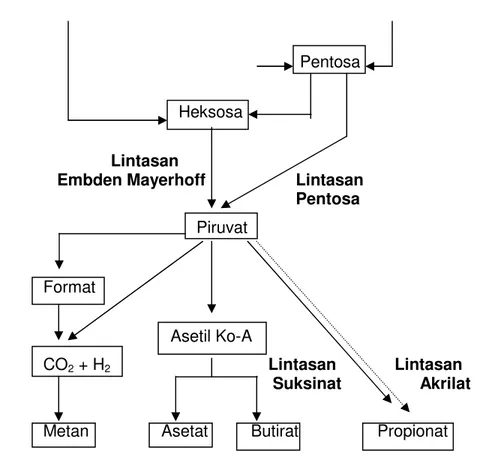
Pakan ruminansia mengandung sejumlah nutrien seperti karbohidrat, protein, lemak, vitamin dan mineral. Karbohidrat merupakan komponen yang mendominasi suatu bahan pakan dan umumnya berupa selulosa, hemiselulosa, pati dan pektin. Hasil pencernaan karbohidrat dalam rumen terutama berupa asam lemak mudah terbang (volatile fatty acid = VFA). Proses pencernaan dan fermentasi karbohidrat dalam rumen menjadi VFA secara skematis disajikan pada Gambar 1. Partikel karbohidrat pakan seperti selulosa, hemiselulosa mengalami hidrolisis oleh enzim â-1-4-glukosidase dari mikroba menjadi sakarida sederhana seperti heksosa, pentosa, dan selobiosa. Karbohidrat yang lain seperti pati, fruktan dan gula-gula mudah larut akan dicerna menjadi maltosa, fruktosa dan glukosa. Hasil pencernaan tersebut segera memasuki proses glikolisis Embden-Meyerhoff untuk menghasilkan piruvat.
Piruvat akan segera dirubah oleh mikroorganisme secara intraseluler menjadi VFA. Komponen VFA yang merupakan produk akhir pencernaan karbohidrat dalam retikulorumen terdiri atas asam asetat, propionat, butirat dan sejumlah kecil valerat serta asam lemak berantai cabang yaitu isobutirat, isovalerat dan 2-metilbutirat (Church dan Pond 1988). Perubahan piruvat menjadi produk akhir VFA dapat melalui beberapa lintasan. Asetat dihasilkan
Pentosa
Heksosa
Lintasan
Embden Mayerhoff Lintasan Pentosa Piruvat Format Asetil Ko-A CO2 + H2 Lintasan Lintasan Suksinat Akrilat Metan Asetat Butirat Propionat
Gambar 1 Skema fermentasi karbohidrat menjadi VFA dalam rumen
(France dan Siddons 1993)
melalui dua jalur yaitu yang pertama melalui pembentukan asetil Ko-A terlebih dahulu, kemudian dirubah menjadi asetil-fosfat kemudian asetat. Jalan yang kedua yaitu dari piruvat dirubah menjadi asetil-fosfat dan format. Format yang dibentuk oleh Methanobacterium diuraikan menjadi CO2 dan H2 yang selanjutnya menghasilkan metan. Pembentukan propionat juga dapat melalui dua jalur, yaitu jalur laktat atau akrilat dan jalur suksinat. Jadi selain VFA, dalam fermentasi karbohidrat juga dihasilkan gas CO2, H2 dan metan yang dikeluarkan dari rumen melalui proses eruktasi.
Kisaran konsentrasi VFA total yang layak bagi kelangsungan hidup ternak adalah 80–160 mM, sedangkan konsentrasi VFA parsial selalu berubah bergantung jenis pakan yang dikonsumsi. Pada pemberian hijauan tinggi, proporsi asetat meningkat, sebaliknya pada pemberian konsentrat tinggi proporsi propionat akan meningkat. Pada pemberian protein ransum tinggi, akan dihasilkan isovalerat dan isobutirat yang tinggi pula (Sutardi 1977). Selanjutnya VFA yang dihasilkan langsung diserap oleh epitelium rumen dan dibawa ke hati melalui sistem porta untuk dimetabolisme lebih lanjut.
Lemak merupakan senyawa organik berminyak yang tidak larut dalam air. Komponen lemak yang paling banyak adalah triasilgliserol yang merupakan bahan bakar utama bagi semua organisme hidup dan juga merupakan komponen utama membran sel yakni tempat terjadinya reaksi-reaksi metabolik (Lehninger 1993). Berbeda dengan ternak non ruminansia, pencernaan lemak pada ruminansia dewasa pertama kali dimulai di dalam retikulorumen, selanjutnya akan mengalami dua proses penting, yaitu lipolisis dan biohidrogenasi. Pertama-tama lemak akan mengalami lipolisis oleh enzim lipase mikroba menghasilkan asam lemak bebas (Free Fatty Acid = FFA), gliserol dan galaktosa. Gliserol dan galaktosa difermentasi lebih lanjut untuk menghasilkan VFA terutama propionat, sedangkan FFA dengan cepat akan dihidrogenasi oleh mikroba menghasilkan produk akhir berupa asam lemak jenuh (Preston dan Leng 1987). Asam lemak rantai pendek (C2-14) dan VFA langsung diserap oleh dinding rumen, sedangkan penyerapan asam lemak jenuh berantai panjang (>C14) hanya terjadi di usus halus.
Pencernaan dan Metabolisme Protein pada Ruminansia
Proses pencernaan protein pada ruminansia termasuk unik, karena melibatkan mikroorganisme di dalam lambung majemuk yang menghasilkan enzim proteolotik dan sangat berperan dalam pencernaan protein. Semua protein dan non proteinnitrogen (NPN) yang masuk ke dalam rumen mengalami hidrolisis oleh enzim proteolitik menjadi oligopeptida dan asam amino. Oligopeptida dan asam amino tersebut merupakan produk antara dan selanjutnya akan dikatabolisme dan dideaminasi menghasilkan VFA, CO2, CH4 dan NH3 (Sutardi 1979; Baldwin dan Allison 1983; McDonald et al. 1988). Proses proteolitik dan deaminasi asam amino menjadi amonia diduga tidak memiliki kontrol metabolik. Hal ini berarti degradasi dan deaminasi terhadap asam amino terus berlangsung meskipun telah terjadi akumulasi amonia yang cukup tinggi.
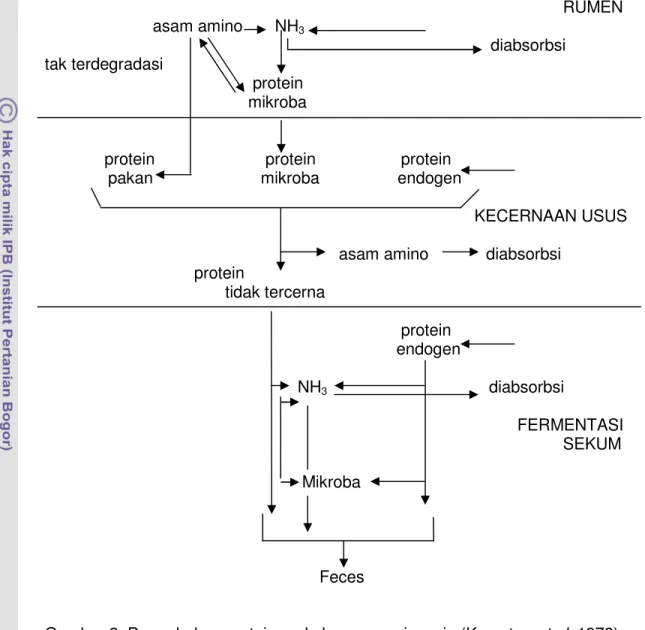
protein pakan protein endogen
FERMENTASI RUMEN asam amino NH3 diabsorbsi tak terdegradasi protein mikroba
protein protein protein pakan mikroba endogen
KECERNAAN USUS asam amino diabsorbsi
protein tidak tercerna protein endogen NH3 diabsorbsi FERMENTASI SEKUM Mikroba Feces
Gambar 2 Perombakan protein pada hewan ruminansia (Kempton et al. 1978) Secara skematis perombakan protein pakan pada ruminansia dijelaskan oleh Kempton et al. (1978) seperti pada Gambar 2. Pada gambar tersebut terlihat bahwa tidak seluruh protein pakan yang masuk ke dalam rumen akan didegradasi oleh mikroba rumen. Ada sebagian protein pakan yang tak terdegradasi dalam rumen dan dapat dicerna oleh enzim pencernaan pascarumen kemudian diserap oleh usus halus.
Amonia sebagai hasil degradasi oleh mikroba rumen akan digunakan untuk sintesis de novo protein mikroba yang selanjutnya menyediakan protein mikroba bagi induk semang. Menurut Cheeke (1998), protein mikroba yang sampai ke abomasum menjadi tersedia untuk induk semang. Protein ransum yang tak terdegradasi dalam rumen bersama protein mikroba akan mengalir ke abomasum terus ke usus halus dan dihidrolisis oleh enzim proteolitik yang dihasilkan oleh ternak dan untuk selanjutnya diserap (Kempton et al.1978 dan Nolan 1993). Protein yang tidak terhidrolisis akan meninggalkan usus dan mengalami fermentasi dalam sekum dan kolon atau diekskresikan melalui feses. Secara umum produk fermentasi dalam sekum dan kolon tidak bermanfaat bagi induk semang (Kempton et al. 1978).
Proses degradasi protein tidak dapat dipandang sebagai suatu proses yang menguntungkan ataupun merugikan. Pada kondisi tertentu proses degradasi diharapkan dapat memenuhi kebutuhan amonia dan peptida untuk pertumbuhan mikroba rumen. Protein mikroba tersebut merupakan sumber pasokan asam amino bagi induk semang (Chen et al. 1992; Verbic et al. 1990; Puchala dan Kulasek 1992). Menurut Sniffen dan Robinson (1987) mikroba rumen dapat mensuplai protein sebanyak 40–80% dari kebutuhan ternak ruminansia. Sebaliknya pada kondisi protein pakan bermutu tinggi, diharapkan laju degradasi protein tidak terlalu tinggi. Oleh karena itu terhadap protein pakan berkualitas tinggi yang fermentabel perlu dilakukan proteksi agar tahan terhadap degradasi dalam rumen.
Pemberian protein ransum yang tak terdegradasi dalam rumen dapat meningkatkan pasokan asam amino ke dalam usus halus dan memperbaiki profil asam amino (Volden 1999). Protein merupakan bahan pakan ternak ruminansia yang cukup mahal harganya, maka perhatian untuk meminimalkan degradasi protein pakan dalam rumen perlu dipertimbangkan (Russel et al. 1992). Lebih lanjut Saricicek (2000) dan Mustafa et al. (2000) menyatakan bahwa untuk mengoptimalkan kemampuan ternak agar dapat berproduksi sesuai potensi genetiknya perlu perhatian pada protein pakan yang tak terdegradasi dalam rumen. Terhadap protein pakan yang tak terdegradasi dalam rumen perlu dievaluasi lebih lanjut, terutama terhadap kecernaan oleh enzim proteolitik di dalam organ pencernaan pascarumen dan keseimbangan asam aminonya. Jadi usaha memacu produksi ternak melalui perbaikan nutrisi protein dapat dilakukan
dengan cara meningkatkan pemberian protein ransum yang tak terdegradasi dalam rumen dan memaksimalkan sintesis protein mikroba. Melalui cara tersebut diharapkan pasokan asam amino untuk diserap oleh usus halus menjadi lebih banyak.
N-NH3 dan Sintesis Protein Mikroba Rumen
Proses perombakan protein oleh mikroba rumen menghasilkan amonia, peptida dan asam amino (Nolan 1993). Amonia, peptida dan asam-asam amino adalah sumber N untuk mikroba rumen. Lebih kurang 82% jenis mikroba rumen dapat menggunakan amonia sebagai sumber N (Sniffen dan Robinson 1987). Sebagian besar bakteri menggunakan amonia untuk sintesis protein tubuhnya, walaupun ada sebagian kecil yang membutuhkan peptida dan asam amino (Leng dan Nolan 1982; Wallace dan Cotta 1998). Hal ini disebabkan karena sebagian mikroba mempunyai sistem transpor untuk mengangkut asam amino ke dalam tubuhnya (Nolan 1993).
Amonia di dalam rumen tidak seluruhnya digunakan oleh mikroba, sebagian akan diserap melalui dinding rumen. Jumlah amonia yang diserap melalui dinding rumen bergantung pH rumen, semakin tinggi pH (basa) makin besar jumlah amonia yang diserap (Haresign dan Cole 1984). Amonia yang diserap akan dibawa oleh darah menuju ke hati untuk dikonversi menjadi urea. Sebagian urea kembali ke rumen melalui saliva dan sebagian lagi dikeluarkan dari tubuh melalui urin. Pool amonia dalam rumen tidak hanya disuplai oleh proses degradasi protein ransum, tetapi juga oleh sumber N lainnya seperti urea. Urea sudah umum ditambahkan pada pakan yang bermutu rendah. Cheeke (1998) menyatakan bahwa urea merupakan sumber NPN terbesar yang sering ditambahkan dalam pakan dan di dalam rumen akan mengalami degradasi dengan cepat menghasilkan amonia, dengan konsentrasi maksimal dalam 1 jam setelah makan (Broderick dan Wallace 1988). Sementara menurut Kozloski et al. (2000) konsentrasi N amonia rumen meningkat dengan semakin meningkatnya penambahan urea. Peningkatan terjadi hingga 3 jam pertama sesudah pemberian pakan. Berkenaan dengan amonia adalah sumber N utama untuk sintesis protein mikroba maka, konsentrasi amonia dalam rumen perlu diperhatikan.
Sintesis protein mikroba akan mencapai laju optimum pada konsentrasi amonia rumen sebesar 3.57 mM Satter dan Slyter (1974). Preston dan Leng (1987) melaporkan bahwa kadar amonia cairan rumen sangat bervariasi dan bergantung pada kandungan N ransum, dengan kisaran 10.7–14.3 mM. Leng (1991) mendapatkan nilai sebesar 14.29 mM untuk mengoptimumkan aktivitas mikroba rumen pada ternak sapi yang diberi pakan berserat dengan tingkat kecernaan serat dan protein yang rendah. Sementara Mehrez et al. (1977) melalui penelitian in vivo pada ternak domba mendapatkan bahwa konsentrasi amonia yang lebih tinggi, yaitu sebesar 16.78 mM agar aktivitas mikroba dapat optimal. Sutardi (1979) menyatakan bahwa kadar amonia rumen yang mendukung pertumbuhan mikroba adalah 4-12 mM. Pada konsentrasi amonia kurang dari 4 mM proses sintesis protein mikroba sudah terganggu. Pada konsentrasi amonia yang lebih besar membutuhkan ketersediaan karbohidrat yang siap pakai untuk memaksimalkan sintesis protein mikroba. Studi lain menyatakan bahwa kadar amonia yang cukup untuk mencapai efisiensi penggunaan energi dan protein ransum adalah 7-8 mM (Erwanto et al. 1993).
Produksi amonia merupakan petunjuk dari proses degradasi oleh mikroba rumen. Produksi amonia rumen mencapai titik optimal pada saat tiga jam setelah makan (Sutardi 1994; Mirsha et al. 2004). Jika ransum defisien akan protein atau proteinnya tahan degradasi, maka konsentrasi amonia dalam rumen akan rendah dan pertumbuhan mikroba rumen akan lambat yang menyebabkan turunnya kecernaan terutama kecernaan serat (McDonald et al. 1988). Produksi amonia dalam rumen menjadi penting untuk diperhatikan dan terdapat korelasi dengan degradabilitas ransum dalam rumen untuk mendukung sintesis mikroba rumen yang optimum.
Mikroba rumen memberi sumbangan protein yang cukup besar terhadap kebutuhan ternak ruminansia (Sniffen dan Robinson 1987). Untuk mengoptimalkan pertumbuhan mikroba rumen, maka disamping menuntut ketersediaan N yang cukup, pasokan nutrien lainnya sangat dibutuhkan seperti energi, asam amino, mineral dan vitamin. Energi yang dibutuhkan untuk sintesis protein mikroba adalah energi dalam bentuk ATP, sedangkan VFA yang bermanfaat bagi mikroba rumen hanyalah yang bercabang dan dibutuhkan sebagai sumber kerangka karbon. Asam amino berantai cabang sangat mendukung sintesis protein mikroba karena akan mengalami deaminasi dan
dekarboksilasi menghasilkan asam lemak berantai cabang. Erwanto (1995) dan Zain (1999) melaporkan bahwa penambahan asam amino berantai cabang dalam ransum ruminansia mampu memacu pertumbuhan bakteri rumen. Hasil penelitian Kanjanapruthipong et al. (2002) melaporkan bahwa meningkatnya pasokan protein yang tahan degradasi rumen menyebabkan terjadi penurunan sintesis protein mikroba dalam rumen yang diindikasikan oleh menurunnya derivat purin dalam urin.
Meningkatnya pertumbuhan bakteri rumen dapat dilihat dari produksi total purin dalam cairan rumen atau ekskresi alantoin (salah satu derivat purin) di dalam urin. Lebih lanjut dinyatakan bahwa absorbsi purin dari asam nukleat yang didegradasi akan dikeluarkan melalui urin sebagai derivat purin, yaitu hypoxanthin, xanthin, asam urat dan alantoin. Dengan demikian produksi N mikroba rumen dapat diestimasi dari derivat purin yang diekskresi melalui urin (Chen et al. 1990). Secara skematis degradasi purin dan pembentukan derivat purin seperti yang disajikan pada Gambar 3.
Ekskresi derivat purin dapat dijadikan indikator dan metode untuk memprediksi biomasa mikroba yang meninggalkan rumen dan dicerna dalam usus halus. Hal ini karena alantoin bersama dengan xanthin dan hipoxanthin merupakan metabolit intermediet dari proses pencernaan bakteri rumen di dalam usus halus yang dikeluarkan melalui urin. Kanjanapruthipong dan Leng (1998) melaporkan bahwa hipoxanthin, xanthin, asam urat dan alantoin yang diekskresikan dalam urin merupakan derivat metabolit dari purin endogen dan eksogen yang didegradasi dalam tubuh. Jumlah asam urat dan alantoin yang diekskresikan dalam urin diketahui mempunyai hubungan dengan pool asam nukleat mikroba dalam rumen (Topp dan Elliot 1965) dan dalam usus halus yang nilainya sebanding dengan derivat purin dalam darah (McAllan 1980). Nilai purin dan derivatnya dapat menggambarkan besarnya protein mikroba yang memasok kebutuhan ternak ruminansia. Alantoin urin sebagai komponen derivat purin mengkontribusi >70% dari totalnya (Orden et al. 2000).
Purin Nukleotida Defosforilasi Purin nukeosida Basa purin Adenin Guanin Adenase Guanase Adenosin Inosin HYPOXANTHIN Xanthin oxidase XANTHIN Xanthin oxidase ASAM URAT Uricase ALANTOIN
Gambar 3 Degradasi purin nukleotida dan pembentukan derivat purin (Chen dan Gomez 1992)
Berdasarkan hasil penelitian Obispo dan Dehority (1999) diketahui kadar purin dari mikroba rumen sebesar 1.88 10-9 ì g setiap koloni. Ogimoto dan Imai (1981) melaporkan bahwa populasi bakteri dalam rumen berkisar 1010-1012 koloni ml-1. Jadi besarnya kadar purin lebih kurang 0.2 mg ml-1 cairan rumen. Perez et al. (1997) melaporkan bahwa basa purin dipengaruhi oleh kadar protein dan karakteristik sumber protein ransum. Besarnya aliran basa purin melalui duodenum ternyata lebih tinggi pada ransum dengan konsentrat tinggi (17.7 mmol hari-1) dibanding ransum dengan konsentrat rendah (12.9 mmol hari-1). Pada kedua level konsentrat suplementasi tepung ikan menghasilkan basa purin terendah (12.3 mmol hari-1) dibandingkan dengan bungkil kedelai (17.25 mmol/h) maupun biji matahari (16.32 mmol hari-1). Komponen alantoin sebagai komponen terbesar dari derivat purin dalam urin menunjukkan pola yang sama. Zain (1999) melaporkan bahwa alantoin urin sejumlah 73 mg hari-1 mampu mendukung pertumbuhan domba sebesar 104 g hari-1.
Pemberian protein pakan yang tidak mudah terdegradasi, akan menurunkan laju fermentasi protein oleh mikroba, sehingga menurunkan pasokan energi dan asam amino mikroba untuk induk semang (Volden 1999). Pada saat sumber protein tak terdegradasi dalam rumen tinggi diberikan, suplemen NPN dibutuhkan untuk menjaga kecukupan level amonia rumen guna mendukung sintesis mikroba rumen. Namun demikian dilaporkan bahwa, dibanding amonia, asam amino dan peptida lebih meningkatkan laju dan jumlah protein bakteri yang disintesis (NRC 1996). Dalam banyak kasus pakan alami mengandung protein terdegradasi yang cukup untuk memenuhi kebutuhan mikroba rumen akan asam amino, peptida dan asam amino bercabang.
Protein Tak Terdegradasi dalam Rumen dan Kecernaan Pascarumen Untuk memaksimalkan produktivitas ternak ruminansia ditinjau dari segi nutrien proteinnya, maka diperlukan pasokan protein yang tak terdegradasi dalam rumen dengan kecernaan oleh enzim pascarumen yang tinggi. Protein mikroba tidak mampu mencukupi seluruh kebutuhan asam amino ruminansia. Mengutip pernyataan Henson et al. (1997), sapi yang berproduksi tinggi membutuhkan sejumlah protein yang bermutu tinggi yang mampu menyediakan asam amino esensial ke saluran pencernaan bagian bawah untuk keperluan laktasi dan fungsi metabolik. Mikroba rumen adalah sumber protein berkualitas tetapi tidak selalu dapat mensuplai jumlah protein metabolisme yang cukup untuk mendukung produksi dan hidup pokok. Protein tidak tercerna dalam rumen dapat meningkatkan aliran asam amino ke saluran gastrointestinal untuk diabsorbsi. Lebih lanjut dinyatakan oleh Sutardi (1979) bahwa sumber protein bagi ruminansia adalah yang memenuhi persyaratan sebagai berikut: (i) mampu menunjang pertumbuhan mikroba rumen yang maksimal, (ii) sebagian besar tahan terhadap degradasi mikroba rumen dan (iii) bernilai hayati atau bernilai
Utilisasi Protein Netto (NPU) tinggi.
Pemenuhan kebutuhan protein ruminansia perlu memperhitungkan jumlah protein pakan yang dapat didegradasi dalam rumen (untuk pertumbuhan mikroba yang optimal) dan jumlah protein ransum yang tak terdegradasi dalam rumen (yang diperlukan untuk melengkapi asam amino asal mikroba rumen). Chen dan Jayasuria (1998) menyatakan bahwa jumlah N yang didegradasi dalam rumen dan dikonversi menjadi protein mikroba menentukan efisiensi
penggunaan ransum oleh ruminansia. Lebih lanjut dinyatakan jika ransum mempunyai efisiensi konversi N yang tinggi, maka protein mikroba lebih banyak diproduksi dan sedikit N yang diekskresikan lewat urin. Adapun terhadap protein pakan yang tak terdegradasi dalam rumen masih diperlukan evaluasi lebih lanjut, terutama terhadap kecernaannya oleh enzim proteolitik di dalam organ pencernaan pascarumen.
Jumlah protein pakan yang mengalami degradasi dalam rumen cukup banyak dan laju degradasi tersebut dipengaruhi oleh kelarutan protein dan laju aliran digesta (Buttery 1976). Derajat ketahanan protein terhadap degradasi oleh mikroba rumen sangat beragam. Madsen dan Hvelplund (1985) telah meneliti degradasi protein dari 38 jenis bahan konsentrat dan 44 jenis hijauan secara in vitro dan in sacco (nylon bag) dan mendapatkan hasil bahwa degradasi protein bahan yang diteliti bervariasi antara 12-90%. Adanya keragaman tersebut memberi peluang kepada kita untuk memilih bahan pakan dengan menyeimbangkan antara bahan pakan yang mudah didegradasi dalam rumen dengan yang tahan degradasi rumen.
Chalupa (1975), Satter dan Roffler (1975) mengklasifikasikan sumber protein atas dasar ketahanan protein terhadap degradasi dalam rumen kedalam tiga kelompok. Bahan pakan sumber protein dengan tingkat ketahanan terhadap degradasi dalam rumen rendah (<40%) adalah kasein, bungkil kedelai, bungkil biji matahari dan bungkil kacang. Sedangkan bahan pakan sumber protein dengan tingkat ketahanan sedang (40-60%) adalah biji kapas, alfalfa yang didehidrasi, biji jagung dan biji-bijian kering bahan pembuat bir, sementara bahan pakan sumber protein dengan tingkat ketahanan tinggi (>60%) adalah tepung daging, corn gluten meal (CGM), tepung darah, tepung bulu, tepung ikan dan beberapa protein yang diproteksi dengan formaldehid.
Peningkatan protein pakan tak terdegradasi dalam rumen tidak selalu meningkatkan produksi. Hal ini dimungkinkan karena protein tersebut memiliki tingkat kecernaan pascarumen yang rendah. Konsekuensinya ketersediaan asam amino untuk dapat diabsorbsi dari usus halus menjadi berkurang (Owen dan Bergen 1983). Hasil penelitian Preston dan Willis (1970) menunjukkan bahwa konsumsi dan pertumbuhan dapat distimulasi dengan pasokan protein pakan tak terdegradasi dalam rumen dari tepung ikan yang ditambahkan pada pakan rendah protein. Hasil serupa juga ditunjukkan dengan pemberian barley
pelet pada domba betina (Orskov et al. 1973). Pada penelitian lain dilaporkan bahwa meningkatnya level protein pakan yang tahan degradasi rumen meningkatkan pertumbuhan bulu pada domba dan terjadi peningkatan pertambahan bobot badan (Litherland et al. 2000).
Hasil Penelitian Pemberian Protein pada Ruminansia
Penelitian terhadap empat macam sumber protein dalam ransum yang disusun isonitrogen telah dilakukan oleh Habib et al. (2001). Hasil yang diperoleh adalah pertambahan bobot badan yang lebih tinggi dari kontrolnya, tetapi tidak menunjukkan perbedaan diantara keempat macam sumber protein. Faktor yang mendukung pertambahan bobot badan tidak seluruhnya dari pasokan protein, tetapi juga melibatkan sumber energi baik karbohidrat maupun lemak. Jadi walaupun pasokan asam amino ke dalam usus berbeda, pertambahan bobot badan yang dihasilkan bisa juga tidak berbeda.
Schlolsser et al. (1993) melaporkan bahwa pemberian bungkil kedelai dan tepung darah pada kambing betina tidak menunjukkan perbedaan total protein yang masuk usus maupun pertambahan bobot badan. Hal yang sama pada sapi dilaporkan oleh Klusmeyer (1990) tidak terdapat perbedaan pengaruh substitusi bungkil kedelai dengan corn gluten meal (CGM) terhadap total protein yang masuk usus. Aliran protein ke pascarumen terdiri atas protein mikroba dan protein yang tidak terdegradasi dalam rumen dan dicerna di dalam usus. Bungkil kedelai sebagai sumber protein yang mempunyai tingkat degradasi lebih tinggi daripada CGM mempunyai dukungan terhadap sintesis protein mikroba yang lebih tinggi, sedangkan CGM dengan tingkat degradasi rendah dalam rumen lebih mampu menyediakan protein ke dalam usus halus.
Kalbande dan Thomas (2001) memberikan substitusi tiga macam ransum dengan tingkat protein degradable tinggi, sedang, dan rendah terhadap protein tahan degradasi. Ransum disusun isonitrogen dan isoenergi. Hasil penelitian menunjukkan bahwa pH dan amonia rumen lebih rendah pada pakan dengan protein tahan degradasi, tetapi VFA total dan parsialnya tertinggi. Perubahan dari profil dan karakteristik fermentasi rumen dipengaruhi oleh jumlah kelarutan dari protein ransum. Konsentrasi N amonia cairan rumen meningkat dengan semakin meningkatnya level protein yang terdegradasi. Ditambahkan oleh Lee
konsentrasi amonia, yang berarti rendahnya kecernaan protein sehingga ketersediaan amonia untuk pertumbuhan mikroba rendah juga. Kanjanapruthipong et al. (2002) menyatakan total bakteri yang hidup dan sintesis protein mikroba yang ditunjukkan dalam bentuk derivat purin dalam urin menurun dengan semakin meningkatnya protein tahan degradasi dalam rumen. Menurunnya bakteri rumen yang diindikasikan oleh jumlah total bakteri hidup dengan semakin meningkatnya kadar protein tahan degradasi dari bungkil kedelai yang diberi formalin dalam pakan diduga akibat lebih rendahnya peptida yang larut dan asam amino yang tersedia untuk asimilasi. Kelebihan dari peptida dan asam amino dari protein degradabel yang berlebihan untuk sintesis protein mikroba dapat digunakan sebagai penghasil ATP.
Evaluasi Mutu Protein Ransum sebagai Penduga Utilisasi Protein Ransum
Evaluasi ransum dapat dilakukan secara fisik, kimiawi dan biologis. Evaluasi fisik ransum dilakukan terhadap ukuran, bentuk, keambaan, daya serap air dan kelarutannya. Evaluasi kimia ransum yang menggambarkan komposisi nutrien, seperti lemak dan protein hanyalah kadarnya saja. Penentuan besarnya kebutuhan nutrien dan pemberian ransum ternak domba di Indonesia masih digunakan standar yang direkomendasikan oleh NRC (1985) dan Kearl (1982). Kebutuhan nutrien yang dimaksud adalah kadar nutrien yang ditentukan berdasarkan analisa laboratorium. Berdasarkan kadar nutrien saja tidak dapat diketahui kualitas dan manfaat ransum bagi ternak yang mengkonsumsi. Kualitas dan manfaat baru dapat diketahui setelah dicobakan pada ternak. Pada ternak ruminansia yang mempunyai lambung majemuk proses pencernaannya berbeda dengan hewan monogastrik. Oleh karena itu pengujian manfaat ransum yang diberikan terhadap ternak yang mengkonsumsi perlu dilakukan.
Evaluasi ransum secara biologis dapat dilakukan secara laboratorium (in vitro dan in sacco) maupun menggunakan hewan percobaan (in vivo). Teknik in vitro mempunyai beberapa keuntungan dibanding teknik in vivo, antara lain dapat dilakukan secara cepat, biaya murah, jumlah sampel yang digunakan sedikit, kondisi relatif homogen dan dapat dikontrol serta dapat mengevaluasi beberapa macam ransum dalam waktu singkat (Church 1979). Pada teknik ini dipergunakan rumen buatan sebagai media (kultur), cairan rumen sebagai
sumber inokulum, larutan buffer untuk mempertahankan pH rumen dengan kondisi dijaga anaerob seolah-olah menyerupai kondisi rumen pada ternak sesungguhnya. Oleh karena itu hasil percobaan in vitro dapat dipergunakan untuk memprediksi percobaan in vivo. Teknik in sacco telah biasa dilakukan untuk mengukur degradasi pakan di dalam rumen, dengan menggunakan hewan berkanula rumen. Tingkat degradasi pakan diukur dari bahan yang hilang pada kantong nilon terhadap bahan awal yang diinkubasikan dalam rumen. Pengukuran didasarkan pada lama inkubasi yang berbeda dan berurutan.
Berdasarkan hal tersebut, maka evaluasi ransum dapat dilakukan dengan teknik in vitro dan in sacco, guna memprediksi mutu ransum yang sesungguhnya bermanfaat bagi ternak yang mengkonsumsinya. Madsen (1985) telah memperbaiki sistem evaluasi protein dengan memperhitungkan tingkat degradasi dan daya cerna nutrien di dalam rumen serta sintesa protein mikroba. Suatu metode pendekatan dilakukan untuk menduga kualitas ransum yang diberikan dan besarnya sumbangan nutrien ransum terhadap penampilan ternak. Salah satu pendekatan untuk mengevaluasi mutu ransum menggunakan tolok ukur mutu protein ransum yang diberikan, yaitu kemampuan menghasilkan amonia (Setter dan Slyter 1974), laju degradasi dalam rumen (Orskov 1982) dan kecernaan protein tak terdegradasi dalam rumen oleh enzim pascarumen (Calsamiglia dan Stern 1995, Muktiani 1994, Habib et al. 2001) dan produksi purinnya (Chen dan Gomez 1992). Terdapatnya korelasi tolok ukur mutu protein ransum tersebut dengan retensi nitrogen, pertambahan bobot hidup dan deposisi protein tubuh, secara tidak langsung dapat digunakan untuk memprediksi produktivitas ternak.