Maturation in Douglas-fir: I. Changes in stem, branch and
foliage characteristics associated with ontogenetic aging
GARY A. RITCHIE and JAMES W. KEELEY
Weyerhaeuser Company, 505 North Pearl Street, G.R. Staebler Forest Resources Research Center, Cen-tralia, WA 98531, USA
Received February 23, 1994
Summary
Two experiments were conducted to characterize changes associated with ontogenetic aging in coastal Douglas-fir (Pseudotsuga menziesii (Mirb.) Franco) and to identify possible maturation ‘‘markers’’ for this species. In the first experiment (Experiment 1), scions of ages 1, 4 and 9 years were collected from four seed zones in field progeny tests and grafted onto 1-year-old rootstocks. In Experiment 2, scions from five full-sib families of ages 1 and 10 years were collected from one progeny test and similarly grafted. Grafts for both experiments were planted in the field in a completely randomized design. The grafts were measured after two and six growing seasons. In Experiment 1, graft diameter, number of terminal bud flushes per year, number of branches, and branch length decreased with age. Plagiotropic angle and needle weight increased. All variables except needle length had a significant age × seed zone interaction. After 6 years, internodal stem diameters, numbers of nodal and internodal branches, and length and diameter of internodal branches decreased with age, and there were age × seed zone interactions with most variables. In Experiment 2, graft height and diameter, number of flushes, number and size of lateral branches, needle length and weight decreased with age. After 6 years, height and diameter, size and number of nodal and internodal branches, and leaf chlorophyll concentration (meas-ured in April) decreased, but there were relatively few age × family interactions. An analysis based on traits that were significantly affected by age, but that did not interact with seed zone or family, indicated that main stem diameter, nodal branch length and nodal branch diameter were the most consistent and reliable maturation markers.
Keywords: juvenility, maturation, phase change, Pseudotsuga menziesii.
Introduction
The phenomenon of maturation (also called phase change or ontogenetic aging) in plants has been extensively studied and reviewed (e.g., Doorenbos 1965, Borchert 1976, Hackett 1985, 1992, Greenwood 1992, Greenwood and Hutchison 1993). Briefly, as perennial plants age they exhibit irreversible morphological and physi-ological changes. These often include reduced growth rate and branch vigor (Sweet 1973, Greenwood 1984), diminished apical dominance (Wareing 1987), increased leaf width and surface area (Steele et al. 1989), increased propensity to flower (Greenwood 1984, Zimmerman et al. 1985), decreased stomatal conductance, and increased chlorophyll content and net photosynthesis (Hutchison et al. 1990, Gross-nickle and Russell 1993). Many of these traits vary independently of one another and appear to be under some degree of hormonal control (Zimmerman et al. 1985), and at least some are associated with changes in gene expression (Greenwood 1992).
of the Pacific Northwest coniferous forest biome. Substantial investments are being made in Douglas-fir regeneration systems, based on both seed and vegetative propa-gation. Maturation state is an important consideration in both cases. It is a key determinant of flowering and hence seed production, and also strongly affects success of vegetative propagation both in vitro and ex vitro, which is impeded as maturation progresses (Clark 1981, Smith 1986, Wareing 1987). Nevertheless, little is known about the morphological consequences of maturation in Douglas-fir.
Here we report results of a study aimed at identifying morphological maturation ‘‘markers’’ in Douglas-fir. These markers will be used to assess the maturation state of vegetative propagules growing in the field.
Materials and methods
The experimental approach was to collect scions of similar genetic origin from field progeny tests of different ages. Each scion was grafted onto a juvenile rootstock and its performance was evaluated over 6 years as a function of physiological age and origin. In the first experiment (Experiment 1), scions were collected from trees of ages 1, 4 and 9 years from planting (+1 = years from seed) that were growing as field controls (checks) at three progeny test sites in western Washington. In Experiment 2, scions were collected from one progeny test site on which full-sib crosses were being evaluated. Material was taken from four families and two age classes: 1 year and 10 years after planting. Thus, in Experiment 1 there were three age classes with considerable genetic variability, whereas in Experiment 2 there were two age classes and genetic variability was tightly controlled.
Scion collection
In Experiment 1, scions were collected from field checks against which the perform-ance of seed orchard families was being evaluated. These checks had been drawn from the same pool of seed for each of the different aged tests, and had the same geographic origin. Seeds used were from Seed Zones 030-05, 041-10, 411-15 and 412-15 (Western Forest Tree Seed Council) in western Washington State. In February 1986, collections were made from the tips of first-order branches located in the upper third of the crown, the most mature portion of the tree (Greenwood et al. 1989). One scion was taken from each of 24 trees in each test. The test sites were located near (1) Brooklyn, (2) Snoqualmie Falls and (3) Kelso, WA. Immediately after collection, the scions were wrapped in moist paper towel, placed in plastic bags, transported to the laboratory on ice and stored at −1 °C until grafting.
Propagation of rootstocks
Seedling rootstocks were propagated during 1985 from seed of the same seed zones (Experiment 1) or families (Experiment 2) as those from which scions were to be collected in 1986. They were grown in plastic containers about 405 mm in diameter by 200 mm depth (Ray Leach cells) during the 1984 growing season in a greenhouse near Rochester, WA. In late summer 1984, they were subjected to a dormancy induction regime consisting of natural photoperiod, mild water stress and fertiliza-tion with a low nitrogen liquid fertilizer. When they had formed overwintering buds they were moved to an unheated greenhouse at Centralia, WA to await grafting.
Grafting
In early March 1986, scions were randomly selected from each treatment combina-tion and grafted to rootstocks that had been propagated from seed of the same seed zone or family from which the scions had been collected. The grafts were made by decapitating the rootstock with a sterile razor blade at a point on the stem roughly equal in diameter to that of the scion base, then cutting a longitudinal slit approxi-mately 2 cm down into the cut end of the stem. The scions were then sharpened to a point on two sides and slipped into the cut forming a V-graft. The grafts were sealed with Parafilm and covered with a small clear plastic envelope to minimize transpira-tion. Graft take was generally good and did not vary across age classes (cf. Green-wood and Hutchison 1993). Grafts were held in a cool, shaded greenhouse and were well healed within 4 weeks.
Field establishment
Experiments 1 and 2 were planted separately, but side by side on a level, well prepared field site at Gilbert Experimental Forest near Chehalis, WA. The soil was a silty loam. Before planting, the site was sprayed with glyphosate and rototilled. Trees were planted on a 1 × 1 m grid in a completely randomized design in mid-April 1986. Between 24 and 30 plants of each treatment combination were planted in Experi-ment 1 for a total of 324, and between 10 and 32 in ExperiExperi-ment 2 for a total of 230. The position and identification of each tree were marked with a metal pin. The site was surrounded by a 3 m high wire fence to prevent browsing. During the second and third summers, weeds were controlled with a string trimmer.
Measurements
In March--April 1992, more detailed measurements were made including: (1) the height above the graft of the terminal bud scar for each of the 1986--1991 growing seasons (retrospective estimate of tree heights), (2) the diameter in the middle of each internode for each of the 1986--1990 increments, (3) the number of nodal branches at each of the 1986--1990 nodes, (4) length and diameter of the longest nodal branch at each of the above nodes, (5) the number of internodal branches for each of the 1986--1991 internodal segments, and (6) in early May, each crown was assessed for bud activity based on a five point scale: 1 = dormant, 2 = swelling, 3 = breaking with bud scales parted, 4 = shoot beginning to elongate, and 5 = shoot partially elongated.
Chlorophyll determination
In March 1992, chlorophyll concentrations were determined on foliage from 20-cm long twig samples collected from one secondary lateral branch from the north side of each tree at the second whorl down from the terminal. The twigs were immediately placed in plastic bags and transported to Centralia, WA on ice in a cooler. They were then held at −1 °C until analyzed for chlorophyll within 5 days of collection. Approximately 30% of the samples from each treatment combination were randomly selected for analysis. Chlorophyll a, chlorophyll b and total chlorophyll were deter-mined spectrophotometrically by the method of Arnon (1949) after extraction in dimethyl sulfoxide (Hiscox and Israelstam 1979). Each determination was made on a pooled sample of 25 apparently healthy needles from the central region of each sample twig.
Data analysis
Both experiments were analyzed with a general linear model analysis of variance (SAS Institute, Cary, NC). The design structure was completely random. The treat-ment structure was a full factorial. Both age and zone in Experitreat-ment 1, and age and family in Experiment 2 were considered fixed effects. Individual means were com-pared by Fisher’s Least Significant Difference (LSD) procedure (Milliken and Johnson 1984).
Results
Experiment 1
Of the 324 grafts planted in the spring of 1986, 99% survived through the first year and 90.5% survived through 6 years. There was no apparent pattern of survival across seed zones or ages. Most of the observed mortality occurred because of graft incompatibility.
termi-nal bud flushes was slightly, but significantly lower in the 9-year-old grafts than in the younger material. Age effects on branch characteristics were particularly strong. Some plagiotropic growth was still evident after 2 years in the field, and was pronounced in the older grafts from two of the four seed zones, although seed zone effects were not statistically significant.
There were no seed zone effects on any of the variables. However, age × seed zone interactions were significant for most variables. These interactions involved both expansion--compression and rank changes of age effects within seed zones (e.g., Figure 1).
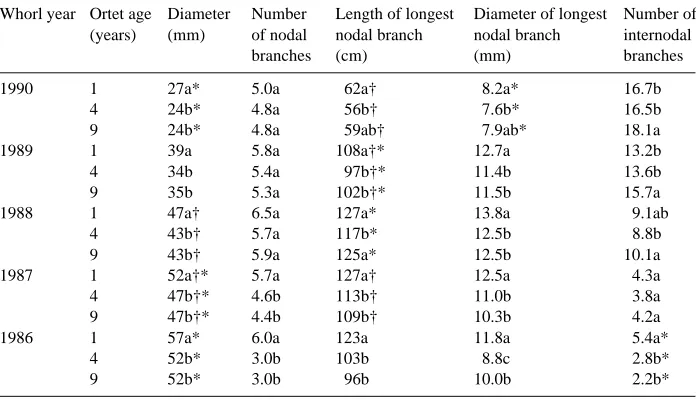
Following 6 years in the field, there were age effects on most measured morpho-logical traits (Table 2). Exceptions were the numbers of nodal branches in the 1987, 1989 and 1990 whorls, and the numbers of internodal branches in the 1987 and 1988 Table 1. Effect of scion age on Douglas-fir grafts after 2 years of field growth. Significant differences (P < 0.05) among means within rows are indicated with different letters according to Fisher’s LSD procedure. Significant age × seed zone interactions are indicated by an asterisk (*). There were no significant seed zone effects.
Variable ANOVA P Scion age at grafting (years)
1 4 9
Graft diameter (mm)* 0.0223 10.1a 8.7ab 8.2b
Graft height (cm)* 0.0169 53.5a 45.3ab 43.5b
No. terminal flushes* 0.0319 3.2a 3.0ab 2.7b
No. branches* 0.0028 20.3a 15.8b 12.8c
Branch length (cm)* 0.0052 362a 251ab 177b
Plagiotropic score* 0.0320 1.23a 1.70a 1.70a
Needle length (mm) 0.4995 22.6a 21.9a 21.5a
Needle weight (g)* 0.9768 4.17a 4.23a 4.25a
whorls. In most cases, the 1-year-old grafts had the highest values, and the 4- and 9-year-old grafts were not significantly different from one another. The 1-year-old grafts consistently had the greatest diameter, with no differences in diameter across whorls or between the 4- and 9-year-old grafts. In the 1986 and 1987 whorls, 1-year-old grafts had more nodal branches than the older grafts, but these differences disappeared by the 1988 whorl. Nodal branch length and diameter were greatest in the 1-year-old grafts and were generally similar in the 4- and 9-year-old grafts. There was a tendency for the youngest grafts to have fewer internodal branches than the older grafts.
Seed zone had less effect on these variables than age. In general, graft height was strongly related to seed zone, whereas branch diameter, length and numbers were not, except for nodal branch length. There were significant interactions between age and seed zone for some variables in some whorl years.
Height growth curves were reconstructed from the graft height data from sequen-tial whorls. Greatest height after six growing seasons was exhibited by grafts originating with the 1-year-old ortets and least from the 4- and 9-year-old material, which were generally not significantly different from each other (Figure 2).
There were no age or seed lot effects on needle dry weight, bud condition, chlorophyll a, chlorophyll b or total chlorophyll concentration, but there was a strong (P = 0.0022) age effect on the chlorophyll a/chlorophyll b ratio. The 1-, 4- and 9-year-old grafts had ratios of 2.39, 2.50 and 2.30, respectively (data not shown). Table 2. Mean values for stem and branch characteristics of Douglas-fir grafts derived from ortets of three ages after 6 years in the field. Different letters within whorl years denote differences among age groups at the 0.05 level as determined by Fisher’s LSD test (P < 0.05). Significant (P < 0.05) seed zone effects are indicated by an asterisk (*); significant age × seed zone interactions are indicated by a †.
Whorl year Ortet age Diameter Number Length of longest Diameter of longest Number of (years) (mm) of nodal nodal branch nodal branch internodal
branches (cm) (mm) branches
1990 1 27a* 5.0a 62a† 8.2a* 16.7b
4 24b* 4.8a 56b† 7.6b* 16.5b
9 24b* 4.8a 59ab† 7.9ab* 18.1a
1989 1 39a 5.8a 108a†* 12.7a 13.2b
4 34b 5.4a 97b†* 11.4b 13.6b
9 35b 5.3a 102b†* 11.5b 15.7a
1988 1 47a† 6.5a 127a* 13.8a 9.1ab
4 43b† 5.7a 117b* 12.5b 8.8b
9 43b† 5.9a 125a* 12.5b 10.1a
1987 1 52a†* 5.7a 127a† 12.5a 4.3a
4 47b†* 4.6b 113b† 11.0b 3.8a
9 47b†* 4.4b 109b† 10.3b 4.2a
1986 1 57a* 6.0a 123a 11.8a 5.4a*
4 52b* 3.0b 103b 8.8c 2.8b*
Experiment 2
After two growing seasons, the 1- and 10-year-old grafts showed large differences in height and diameter across families. The 1-year-old grafts had up to twice the height (Figure 3) and diameter growth (not shown) of the 10-year-old grafts. There were age effects on every variable measured except for plagiotropic angle (Table 3). There was evidence for family effects on graft diameter, branch number and needle charac-teristics only. There were age × family interactions for diameter, bud flushes, and needle weight only.
The 1-year-old grafts exhibited a significantly greater propensity for second and third bud flushing during the second growing season than the 10-year-old grafts (Table 4). The young material had nearly twice the number of lateral branches as the Figure 2. Height growth of Douglas-fir grafts during six field growing seasons. Grafts were collected from 1-, 4- and 9-year-old ortets. Data are pooled across material from four seed zones.
old material and about 20% greater total branch length. Surprisingly, there was only weak evidence for differences in plagiotropic growth between age classes in three of the five families. The needles of young grafts were about 30% longer than those of old grafts and had about 42% greater dry biomass.
After six field growing seasons, 191 of the original 230 grafts survived. Most of the mortality was in the 1-year-old grafts (reason unknown). There were age effects (P < 0.01) on all variables except for 1989 and 1990 branch number and length. There was little evidence for family effects on any variables except for 1988, 1989 and 1990 branch number and 1991 height. There were no age × family interactions except for 1987, 1988 and 1989 height. The 1-year-old grafts exhibited greater height growth than 10-year-old grafts and this difference increased over time (Figure 4). One-year-old grafts also had greater diameters, and more nodal branches with greater length and diameter than 10-year-old grafts (Table 5). In some years they also had more internodal branches.
Table 3. ANOVA P values for effects of ortet age and family on several characteristics of Douglas-fir grafts after 2 years in the field.
Source of variance Age Family Age × Family
Graft diameter 0.0001 0.0195 0.0206
Graft height 0.0001 0.2712 0.0551
No. terminal bud flushes 0.0001 0.2782 0.0261
No. lateral branches 0.0001 0.0008 0.3538
Total branch length 0.0142 0.2058 0.0934
Plagiotropic angle 0.0740 0.2213 0.2181
Needle length 0.0001 0.0111 0.0805
Needle weight 0.0006 0.0254 0.0067
Table 4. Means of measured characteristics of Douglas-fir grafts from five families and two ortet ages after 2 years in the field. Different letters denote differences between ages at the 0.05 level as determined by Fisher’s LSD test.
Family Age Bud flushes Number of Branch Plagiotropic Needle Needle
(years) branches length score length weight
(cm) (mm) (g)
1004 × 1049 1 2.5a 13.3a 18.0a 3.0a 17.0a 2.8a
10 2.3a 8.8b 17.1a 3.7a 15.2a 2.7a
1017 × 1101 1 3.7a 20.5a 27.2a 4.5a 23.1a 4.3a
10 1.7b 8.8b 16.4b 4.3a 9.9b 1.7b
1027 × 1114 1 3.2a 18.3a 22.3a 3.4b 25.6a 7.2a
10 2.2b 10.7b 18.0b 4.1a 16.7b 2.7b
1034 × 1080 1 2.9a 14.8a 26.3a 4.4a 22.5a 3.4a
10 2.4b 9.6b 20.0b 5.2a 20.0a 3.9a
1100 × 1037 1 3.3a 24.0a 23.1a 3.6b 25.5a 4.7a
Stem diameters of the young grafts were consistently about 30% greater than those of the old grafts at every internode. The young material had roughly twice the number of nodal branches as the old material at the 1986 and 1987 nodes, but this difference decreased to about 20% at the 1990 node. Nodal branches of the 1-year-old grafts were consistently about 25% longer, and from 20 to 30% thicker than those of the 10-year-old grafts at each internode. The difference in numbers of internodal branches between ortet ages was about 70% at the 1986 internode, but decreased to about 10% at the 1990 internode.
Ortet age effects were pronounced in most bud and needle characteristics measured (Table 6). There was strong evidence for age effects on all measured variables except chlorophyll b concentration. There was evidence for family effects and age × family Figure 4. Height growth of Douglas-fir grafts during six field growing seasons. Grafts were collected from 1- and 10-year-old ortets. Data are pooled across material from five full-sib families.
Table 5. Mean values for stem and branch characteristics of Douglas-fir grafts derived from ortets of two ages. Different letters denote differences between means at the 0.05 level as determined by Fisher’s LSD test.
Whorl year Ortet age Diameter Number Length of longest Diameter of longest Number of (years) (mm) of nodal nodal branch nodal branch internodal
branches (cm) (mm) branches
1990 1 28a 6.4a 62a 7.3a 15.0a
10 20b 5.0b 47b 6.6b 13.0b
1989 1 40a 6.3a 109a 12.7a 12.2a
10 29b 5.2b 83b 10.0b 12.1b
1988 1 48a 9.1a 121a 13.1a 9.5a
10 34b 5.4b 93b 10.3b 7.0b
1987 1 56a 6.8a 118a 12.1a 3.6a
10 38b 3.3b 77b 8.1b 2.1b
1986 1 59a 5.2a 113a 11.1a 5.6a
interactions only in the cases of bud activity and chlorophyll ratio. Buds of the 1-year-old grafts were more active than those of the 10-year-old grafts when assessed in May. The young grafts also had slightly heavier needles, slightly greater amounts of chlorophyll a and chlorophyll b, and a higher chlorophyll a/chlorophyll b ratio than the old grafts.
Discussion
Maturation is of particular interest in tree species that achieve great size and longevity. Different regions of the crowns of such trees, particularly conifers, often exhibit different maturational states (Greenwood and Hutchison 1993). Generally, branches that are lower on the crown and closer to the stem, even though they might be chronologically older, tend to be more juvenile than those on the upper and outer portions of the crown. This observation prompted Borchert (1976) to argue that morphological and physiological changes during aging may be a direct consequence of increasing size and complexity of the organism.
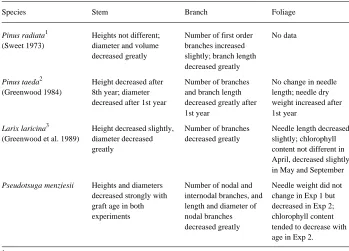
The grafting technique eliminated size and complexity of the organism as well as distance from the root system as possible factors contributing to maturation, because the grafts were of equal size when established and the rootstocks were similar. Therefore, observed morphological and growth differences among material of differ-ent ages were unconfounded with plant size and complexity. This technique was pioneered by Sweet (1973) with radiata pine (Pinus radiata D. Don) and used subsequently by Greenwood (1984) with loblolly pine (P. taeda L.) and Greenwood at al. (1989) with eastern larch (Larix laricina (Du Roi) K. Koch).
Results of the studies by Sweet (1973) and Greenwood and coworkers (Greenwood 1984, Greenwood et al. 1989) are summarized in Table 7 and compared with the results of the present study. In contrast to radiata pine, loblolly pine and eastern larch, Douglas-fir showed a marked reduction in graft height with age. All species exhibited substantial reductions in stem diameter, reflecting a decrease in mean radial tracheid diameter with aging, as demonstrated by Takemoto and Greenwood (1993) in eastern larch. Also common across species was a reduction in the number or length of branches or both. In concert with this, Douglas-fir nodal branches became thinner and shorter as scion age increased. These effects persisted in the field for at least 6 Table 6. Mean values for bud and needle characteristics of Douglas-fir grafts derived from ortets of two ages after 6 years in the field. Measurements were made in March. Different letters denote differences at the 0.05 level as determined by Fisher’s LSD test.
Age Bud Needle dry Chl a Chl b Total Chl Chl a/Chl b
(years) flushes weight (g)1 (mg g−1) (mg g−1)
1 3.22a 0.78a 2.381a 0.999a 3.380a 2.386a
10 2.27b 0.67b 2.138b 0.942b 3.080b 2.269b
years.
Needle traits among species were less consistent than branch and stem traits. Needle dry weight tended to decrease with age in larch and Douglas-fir but increased in loblolly pine. Needle length decreased with age in Douglas-fir in Experiment 2, but not in Experiment 1. Needles of Sitka spruce (Picea sitchensis (Bong.) Carr.) exhibited large changes from age three to age 38 (Steele et al. 1989). Needle length increased up to about age ten, then decreased, whereas width and dry weight increased asymptotically. Most measured traits increased; however, these observa-tions were made on full-grown trees of different ages and so may have been confounded by differences in plant size and complexity, genetics, and other factors. In the field, Douglas-fir grafts from the 9- and 10-year-old ortets often exhibited a darker, more bluish-green foliage than younger material, which tended to appear more yellow-green. Measurements were made to determine the relationship between these differences and chlorophyll concentrations, because Greenwood et al. (1989) found higher chlorophyll contents in mature larch grafts than in juvenile larch grafts. Absolute chlorophyll concentrations for Douglas-fir (Table 6) are in agreement Table 7. Changes in stem, branch and foliage characteristics with increasing physiological age for three conifer species. Data are from scions of different ages grafted to juvenile rootstocks.
Species Stem Branch Foliage
Pinus radiata1 Heights not different; Number of first order No data (Sweet 1973) diameter and volume branches increased
decreased greatly slightly; branch length decreased greatly
Pinus taeda2 Height decreased after Number of branches No change in needle (Greenwood 1984) 8th year; diameter and branch length length; needle dry
decreased after 1st year decreased greatly after weight increased after 1st year 1st year
Larix laricina3 Height decreased slightly, Number of branches Needle length decreased (Greenwood et al. 1989) diameter decreased decreased greatly slightly; chlorophyll
greatly content not different in
April, decreased slightly in May and September
Pseudotsuga menziesii Heights and diameters Number of nodal and Needle weight did not decreased strongly with internodal branches, and change in Exp 1 but graft age in both length and diameter of decreased in Exp 2; experiments nodal branches chlorophyll content
decreased greatly tended to decrease with age in Exp 2.
1 Juvenile grafts were 6 years old, mature grafts were 19 years old; age classes were not genetically
matched; assessed after three years in the field.
2 Graft ages were 1, 4, 8 and 12 years; matched full-sib families; assessed after three years; pot grown. 3 Juvenile grafts were 1 year old, mature grafts were 16-23 years old; half-sib families; assessed after 2
with published data for Norway spruce (Picea abies L. Karst.) (Köstner et al. 1990), Sitka spruce (Lewandowska and Jarvis 1977), red spruce (P. rubens Sarg.) (Rebbeck et al. 1993) and eastern white pine (Pinus strobus L.) (Mann et al. 1980). In Douglas-fir, chlorophyll a, chlorophyll b and total chlorophyll concentrations and the chlorophyll a/chlorophyll b ratio decreased with age. In contrast, Rebbeck et al. (1993) found higher chlorophyll a and lower chlorophyll b concentrations, and a higher chlorophyll a/chlorophyll b ratio in mature red spruce tissues than in juvenile tissues. In larch in April, needle chlorophyll concentrations varied between 3.1 and 3.5 mg g−1 among scion age classes ranging from 1 to 45 years. However, in May, there was a definite increase with age class (Greenwood et al. 1989). These inconsis-tencies may reflect strong seasonal differences in chlorophyll contents, as reported for Norway spruce (Köstner et al. 1990). The lowest seasonal values occurred in April following winter chlorophyll degradation and before the flush of new foliage. In the present study, it is likely that the choice of sampling date influenced the results. There was a weak relationship between bud activity and age. In Experiment 1, the young material tended to produce more terminal bud flushes later in the growing season (lammas growth) than the older material, but there was no difference in bud condition when the trees were scored in May. In Experiment 2, terminal buds exhibited more annual flushes and buds were in a more active state in May.
A key objective of the present study was to identify morphological ‘‘markers’’ that could be used to ascertain the maturation state of young vegetative propagules of Douglas-fir. This has several important practical implications. Mature-behaving trees would be expected to grow more slowly than juvenile trees, but might tend to have more desirable stem form (less taper, fewer and smaller branches) and earlier flowering. If maturation state could be manipulated by vegetative propagation, it might be possible to exploit some of these phenomena (e.g., Menzies and Klomp 1988).
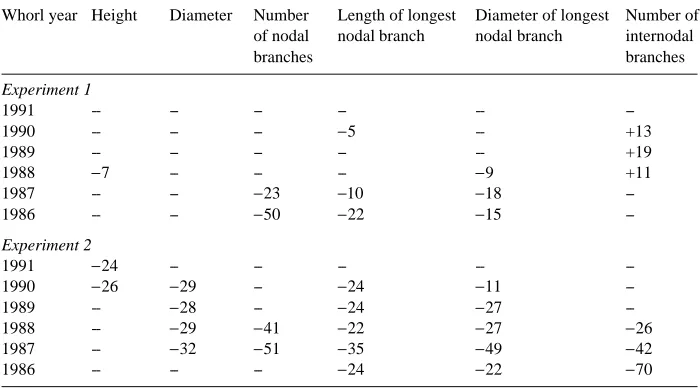
To serve as a suitable marker, a trait must be significantly and unequivocally affected by age, and must be free from age × genotype interactions. The 6-year data from both experiments were examined with respect to these two criteria. Matrices were constructed in which traits meeting the criteria were identified and then displayed as percentage differences between the trait mean for a juvenile and mature plant:
where traitj is the mean value of a particular trait for juvenile plants and traitm is the
mean value of that same trait for mature plants.
smaller diameters than juvenile trees at most internodes. Nodal branch numbers were useful markers only in the older whorls in both experiments. Internodal branches were larger in the mature trees in Experiment 1, but smaller in Experiment 2. In contrast, branch sizes decreased with age in all or most nodes in both experiments.
In both experiments, bud and needle traits declined with age (Table 9). Chloro-phyll a, chloroChloro-phyll b and total chloroChloro-phyll concentrations showed the most consis-tent differences; they each declined between 5 and 10% with aging between 1 and 9 or 10 years.
It appears that the most consistent, least genetically and seasonally confounded maturation markers in Douglas-fir are those traits associated with nodal branches. Specifically, the lengths of the longest nodal branches in juvenile trees (age two from Table 8. Matrix showing stem and branch traits from Experiment 2 that were significantly different (P < 0.05) between ages but did not interact significantly with family. Values show percent increase (+) or decrease (−) in trait mean with age (Equation 1).
Whorl year Height Diameter Number Length of longest Diameter of longest Number of of nodal nodal branch nodal branch internodal
branches branches
Experiment 1
1991 -- -- -- -- --
--1990 -- -- -- −5 -- +13
1989 -- -- -- -- -- +19
1988 −7 -- -- -- −9 +11
1987 -- -- −23 −10 −18
--1986 -- -- −50 −22 −15
--Experiment 2
1991 −24 -- -- -- --
--1990 −26 −29 -- −24 −11
--1989 -- −28 -- −24 −27
--1988 -- −29 −41 −22 −27 −26
1987 -- −32 −51 −35 −49 −42
1986 -- -- -- −24 −22 −70
Table 9. Matrix showing bud and foliar traits from Experiments 1 and 2 that were significantly different (P < 0.05) among ages but did not interact significantly with family or seed zone. Values show percent decreases (−) in trait means with increasing age (Equation 1).
Trait Experiment 1 Experiment 2
Bud condition −10
--Needle dry wt -- −14
Chlorophyll a −9 −10
Chlorophyll b −5 −6
Total chlorophyll −8 −9
--seed) were 20 to 35% greater than in mature trees (age 11 from --seed). Older trees also had 20 to 50% fewer nodal branches on the oldest internodes; however, these differences were confounded with family or seed zone as trees became larger. Main stem diameters were also consistently about 30% greater in juvenile material in Experiment 2.
Although April leaf chlorophyll concentrations showed a consistent, genetically unconfounded decrease with age in both experiments, data from other authors (cited above) indicate strong seasonal changes in these parameters. More research is needed on these variables to confirm their suitability as maturation markers in Douglas-fir.
Acknowledgments
The authors thank Mrs. Patty A. Ward for performing the grafts, Patricia Crank for the chlorophyll analyses, Steven Duke for statistical assistance, D&J Forestry Services, Elma, Washington for providing extensive field measurements, and Drs. M.S. Greenwood and John Frampton for reviewing the original manuscript and offering many useful suggestions. This work was supported entirely by Weyerhaeuser Company.
References
Arnon, D.T. 1949. Copper enzymes in isolated chloroplasts: phenoloxidases in Beta vulgaris. Plant Physiol. 24:1--15.
Borchert, R. 1976. The concept of juvenility in woody plants. Acta Hortic. 56:21--33.
Clark, J.R. 1981. Juvenility and plant propagation. Int. Plant Propagators Soc. Proc. 31:449--453. Doorenbos, J. 1965. Juvenile and adult phases in woody plants. In Encyclopedia of Plant Physiology
XV/1. Ed. W. Rhuland. Springer, Berlin, pp 1222--1235.
Greenwood, M.S. 1984. Phase change in loblolly pine: shoot development as a function of age. Physiol. Plant. 61:518--522.
Greenwood, M.S. 1992. Theoretical aspects of juvenility and maturation. In Mass Production Technol-ogy for Genetically Improved Fast Growing Forest Tree Species. AFOCEL, Bordeaux, France, pp 38--44.
Greenwood, M.S. and K.W. Hutchison. 1993. Maturation as a developmental process. In Clonal Forestry I: Genetics and Biotechnology. Eds. M.R. Ahuja and W.J. Libby. Springer-Verlag, Berlin, pp 14--33. Greenwood, M.S., C.A. Hopper and K.W. Hutchison. 1989. Maturation in larch: 1. Effect of age on shoot
growth, foliar characteristics, and DNA methylation. Plant Physiol. 90:406--412.
Grossnickle, S.C. and J.H. Russell. 1993. Water relations and gas exchange processes of yellow-cedar donor plants and cuttings in response to maturation. For. Ecol. Manage. 56:185--198.
Hackett, W.P. 1985. Juvenility, maturation, and rejuvenation in woody plants. Hortic. Rev. 7:109--155. Hackett, W.P. 1992. Control of maturation in woody species. In Mass Production Technology for
Genetically Improved Fast Growing Forest Tree Species. AFOCEL, Bordeaux, France, pp 45--50. Hiscox, J.D. and G.F. Israelstam. 1979. A method for the extraction of chlorophyll from leaf tissue
without maceration. Can. J. Bot. 57:1332--1334.
Hutchison, K.W., C.D. Sherman, J. Weber, S.S. Smith, P.B. Singer and M.S. Greenwood. 1990. Maturation in larch. II. Effects of age on photosynthesis and gene expression in developing foliage. Plant Physiol. 1308--1315.
Köstner, B., F.-C. Czygan and O.L. Lange. 1990. An analysis of needle yellowing in healthy and chlorotic Norway spruce (Picea abies) in a forest decline area of the Fichtelgebirge (N.E. Bavaria). I. Annual time-course changes in chloroplast pigments for five different needle age classes. Trees 4:55--67. Lewandowska, M. and P.G. Jarvis. 1977. Changes in chlorophyll and carotenoid content, specific leaf
area and dry weight fraction in Sitka spruce in response to shading and season. New Phytol. 79:247--256.
Menzies, M.I. and B.K. Klomp. 1988. Effects of parent age on growth and form of cuttings, and comparison with seedlings. In Workshop on Growing Radiata Pine from Cuttings. New Zealand Forest Research Institute Bulletin 135. Eds. M.I. Menzies, J.P. Aimers and L.J. Whitehouse. Ministry of Forests, Rotorua, pp 18--41.
Milliken, G.A. and D.E. Johnson. 1984. Analysis of messy data. Vol. 1. Designed experiments. VanNos-trand Co., New York, pp 31.
Rebbeck, J., K.F. Jensen and M.S. Greenwood. 1993. Ozone effects of grafted mature and juvenile red spruce: photosynthesis, stomatal conductance, and chlorophyll concentration. Can. J. For. Res. 23:450--456.
Smith, M.A.L. 1986. Influence of juvenility on woody plant growth. Plant Propagator 32:6--7. Steele, M.J., M.P. Coutts and M.M. Yeoman. 1989. Developmental changes in Sitka spruce as indices of
physiological age. I. Changes in needle morphology. New Phytol. 113:367--375.
Sweet, G.B. 1973. The effect of maturation on the growth and form of vegetative propagules of radiata pine. N.Z. J. For. Sci. 2:191--210.
Takemoto, Y. and M.S. Greenwood. 1993. Maturation in larch: age-related changes in xylem develop-ment in the long-shoot foliage and the main stem. Tree Physiol. 13:253--262.
Wareing, P.F. 1987. Phase change and vegetative propagation. In Improving Vegetatively Propagated Crops. Eds. A.A.J. Abbott and R.K. Atkin. Academic Press, London, New York, pp 263--270. Zimmerman, R.H., W.P. Hackett and R.P. Pharis. 1985. Hormonal aspects of phase change and