KEEFEKTIFAN FUNGI MIKORIZA ARBUSKULA
DALAM MENINGKATKAN HASIL DAN ADAPTASI
CABAI (
Capsicum annuum
L.) PADA TANAH
BERCEKAMAN ALUMINIUM
DWIANA WASGITO PURNOMO
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI DISERTASI
DAN SUMBER INFORMASI
Dengan ini saya menyatakan bahwa disertasi “ Keefektifan Fungi Mikoriza Arbuskula dalam Meningkatkan Hasil dan Adaptasi Cabai (Capsicum annuum L.) pada Tanah Bercekaman Aluminium” adalah karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apapun kepada perguruan tinggi manapun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebut dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir disertasi ini.
Bogor, Februari 2008
ABSTRACT
Dwiana Wasgito Purnomo. The Effectiveness of Arbuscular Mycorrhizal Fungus to Increase Yield and Adaptation of Chili (Capsicum annuum L.) in Ultisol with Aluminum Stress. Under supervision of Bambang S. Purwoko, Sudirman Yahya, Sriani Sujiprihati and Irdika Mansur.
The objectives of the research were: to identify genotypes of chili tolerant and sensitive to Aluminum (Al) by root screening test, and agronomical-characters screening test, to obtain compatible, effective and efficient species of Arbuscular Mycorrhizal Fungus (AMF) for yield improvement and adaptation of chilli against Al-stress in Ultisol , and to explain adaptation and tolerance-mechanisms of chili inoculated with AMF to Al stress in Ultisol. The research was conducted in University Farm of IPB, Cikabayan, Bogor from September 2005 to March 2007. Screening was performed to 20 genotypes of chili using Ultisol and various Al-saturation, 0.77, 15.92, 31.96, 60.85 and 83.48% using limestone of 18.33, 13.75, 9.16, 4.58 and 0 g CaCO3/kg soil, respectively. Genotypes were identified as tolerant and as sensitive to aluminum as assessed by using developmental characters and yield under condition without Al stress and Al stressed. Compatibility test was carried out for 4 indigenous AMF genotypes in Ultisol, namely Glomus manihotis, Glomus etunicatum, Gigaspora margarita and Acaulospora sp. They have been inoculated to tolerant and aluminum sensitive chili genotypes. The compatible AMF species was tested further to 2 tolerant genotypes and 2 sensitive genotypes which were planted in media without Al-stress and with Al-stress condition. The results showed that responses of chili genotypes to Al-stress varied among genotypes in screening test and evaluation process. There was positive correlation and consistency between responses of relative root length in screening test and responses of fruit weight harvested in evaluation test. Selection based on the root length and fruit weight harvested in screening and evaluation methods produced tolerant genotypes: PBC 619, Jatilaba, Cilibangi 5 and Jayapura, and sensitive genotypes: Cilibangi 3, Helm, PBC 549 and Tit Bulat. Based on its infectivity and effectivity, AMF species Gigaspora margarita was the most compatible to chili. Inoculation of Gigaspora margarita effectively decreased negative influence of Al-stress by increasing plant height, shoot dry weight, number of fruit harvested, fruit length, weight per fruit and fruit weight harvested. Inoculation of Gigaspora margarita to sensitive genotypes was more advantageous than to tolerant genotypes. Sensitive genotypes adapted to Al-stress if they had been inoculated with Gigaspora margarita. In Al-stress condition, inoculation of Gigaspora margarita to sensitive genotypes increased weight of harvested fruits up to 94.49% in Cilibangi 3 and 80.37% in Helm. Adaptation ability of tolerant genotypes against Al-stress developed as external ability by decreasing Aluminum absorption. Adaptation process in sensitive genotypes to Al-stress was more likely by increasing of N and P contents using Gigaspora margarita as symbion.
ABSTRAK
Dwiana Wasgito Purnomo. Keefektifan Fungi Mikoriza Arbuskula dalam Meningkatkan Hasil dan Adaptasi Cabai (Capsicum annuum L.) pada Tanah Bercekaman Aluminium. Dibawah Bimbingan : Bambang S. Purwoko, Sudirman Yahya, Sriani Sujiprihati dan Irdika Mansur.
Penelitian ini bertujuan untuk mengidentifikasi genotipe cabai yang berpotensi toleran dan peka Al melalui metode penapisan panjang akar dan evaluasi karakter agronomi, mendapatkan jenis fungi mikoriza arbuskula (FMA) yang kompatibel dan efektif untuk perbaikan hasil dan adaptasi cabai terhadap cekaman Al pada tanah ultisol, serta menguraikan mekanisme toleransi cabai yang bermikoriza terhadap cekaman Al pada tanah tanah ultisol. Penelitian dilaksanakan di Kebun Percobaan Cikabayan IPB, Bogor dan berlangsung dari bulan September 2005 sampai Maret 2007. Penapisan 20 genotipe cabai menggunakan media tanah ultisol dengan berbagai tingkat kejenuhan Al, yaitu 0.77 , 15.92, 31.96, 60.85 dan 83.48%, yang diperoleh dengan pemberian kapur masing-masing sebanyak: 18.33, 13.75, 9.16, 4.58 dan 0 g CaCO3/kg tanah. Genotipe yang termasuk kriteria toleran dan peka Al masing-masing sebanyak 4 genotipe kemudian dievaluasi berdasarkan karakter pertumbuhan dan hasil pada kondisi tanpa cekaman Al dan tercekam Al. Uji kompatibilitas dilakukan terhadap 4 jenis FMA indigen tanah ultisol, yaitu Glomus manihotis, Glomus etunicatum, Gigaspora margarita dan Acaulospora sp yang diinokulasi pada genotipe cabai yang toleran dan peka Al. Jenis FMA yang kompatibel diuji keefektifannya terhadap 2 genotipe toleran dan 2 peka yang ditanam pada kondisi normal dan tercekam Al. Hasil penelitian menunjukkan bahwa tanggap genotipe cabai terhadap cekaman Al bervariasi antar genotipe baik pada percobaan penapisan maupun evaluasi. Terdapat konsistensi dan korelasi positif antara tanggap panjang akar relatif pada percobaan penapisan dan tanggap bobot buah panen pada percobaan evaluasi. Hasil seleksi berdasarkan panjang akar dan bobot buah panen pada metode penapisan dan evaluasi diperoleh genotipe toleran, yaitu genotipe PBC 619, Jatilaba, Cilibangi 5 dan Jayapura; serta genotipe peka, yaitu Cilibangi 3, Helm, PBC 549 dan Tit Bulat. Berdasarkan infektivitas dan efektivitasnya, jenis FMA Gigaspora margarita lebih kompatibel dengan tanaman cabai. Inokulasi Gigaspora margarita efektif mengurangi pengaruh buruk akibat cekaman Al melalui peningkatan tinggi tanaman, bobot kering tajuk, jumlah buah panen, panjang buah, bobot per buah dan bobot buah panen. Genotipe peka lebih diuntungkan dengan adanya inokulasi Gigaspora margarita dibandingkan genotipe toleran. Genotipe peka dapat beradaptasi terhadap cekaman Al jika bersimbiosis dengan Gigaspora margarita. Pada kondisi tercekam Al, inokulasi Gigaspora margarita pada genotipe peka meningkatkan bobot buah panen sebesar 94.49% pada Cilibangi 3 dan 80.37% pada Helm. Kemampuan adaptasi terhadap cekaman Al pada genotipe toleran dilakukan secara eksternal dengan mengurangi penyerapan Al. Sementara itu, adaptasi genotipe peka terhadap cekaman Al lebih melibatkan peningkatan kandungan N dan P melalui simbiosis dengan Gigaspora margarita.
© Hak Cipta milik IPB, tahun 2008
Hak Cipta dilindungi Undang-undang.
1. Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumber.
a. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik atau tinjauan suatu masalah.
b. Pengutipan tidak merugikan kepentingan yang wajar IPB.
KEEFEKTIFAN FUNGI MIKORIZA ARBUSKULA
DALAM MENINGKATKAN HASIL DAN ADAPTASI
CABAI (
Capsicum annuum
L.) PADA TANAH
BERCEKAMAN ALUMINIUM
DWIANA WASGITO PURNOMO
Disertasi
sebagai salah satu syarat untuk memperoleh gelar Doktor pada
Departemen Agronomi dan Hortikultura
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Penguji Luar Komisi :
Pada Ujian Tertutup : Dr. Ir. Anas D. Susila, MS
(Staf Pengajar pada Depertemen Agronomi dan Hortikultura, Faperta IPB)
Pada Ujian Terbuka : Dr. Ir. Yusdar Hilman, MS
(Kepala Pusat Penelitian dan Pengembangan Hortikultura, Badan Litbang Pertanian, Departemen Pertanian)
Dr. Ir. Trikoesoemaningtyas, M.Sc
Judul Disertasi : Keefektifan Fungi Mikoriza Arbuskula dalam Meningkatkan Hasil dan Adaptasi Cabai (Capsicum annuum L.) pada Tanah Bercekaman Aluminium.
Nama : Dwiana Wasgito Purnomo
NIM : A 361020161
Disetujui Komisi Pembimbing
Prof. Dr. Ir. Bambang S. Purwoko, M.Sc Ketua
Prof. Dr. Ir. Sudirman Yahya, M.Sc Anggota
Dr. Ir. Sriani Sujiprihati, MS Anggota
Dr. Ir. Irdika Mansur, M.For.Sc Anggota
Diketahui
Ketua Program Studi Agronomi
Dr. Ir. Munif Ghulamahdi, MS
Dekan Sekolah Pascasarjana IPB
Prof. Dr. Ir. Khairil A. Notodiputro, MS
PRAKATA
Alhamdulillahirabbil alamin, segala puji bagi Allah SWT. Atas izin dan petunjuk Allah yang maha rahman dan rahim, penulis dapat menyelesaikan penelitian dan penyusunan disertasi dengan judul: “ Keefektifan Fungi Mikoriza Arbuskula dalam Meningkatkan Hasil dan Adaptasi Cabai (Capsicum annuum L.) pada Tanah Bercekaman Aluminium”.
Pemberdayaan lahan-lahan marginal perlu mendapat perhatian di masa yang akan datang. Hal ini disebabkan kemajuan pembangunan sering mengorbankan lahan-lahan pertanian yang produktif sehingga luasannya semakin sempit. Salah satu jenis tanah yang tergolong marginal dan cukup potensial untuk pengembangan budidaya cabai adalah tanah Ultisol. Namun adanya kelarutan aluminium yang tinggi pada tanah Ultisol merupakan kendala utama yang sering membatasi pertumbuhan tanaman, disamping memang tingkat kesuburan tanahnya sangat rendah.
Penggunaan genotipe toleran Al dan pemanfaatan simbiosis dengan fungi mikoriza arbuskula (FMA) dapat dipertimbangkan untuk mengatasi kendala cekaman Al pada tanah Ultisol. Beberapa genotipe cabai toleran Al telah diseleksi dalam penelitian ini, yaitu PBC 619, Jatilaba, Cilibangi 5, Jayapura dan Marathon. Genotipe peka juga dapat digunakan, namun perlu aplikasi FMA yang kompatibel dan efektif untuk meningkatkan hasil pada tanah Ultisol.
Penelitian dan penulisan disertasi ini dapat diselesaikan karena peran dan dukungan berbagai pihak. Oleh karena itu penulis sampaikan penghargaan dan terima kasih yang sebesar-besarnya kepada semua pihak.
Pertama-tama kepada Bapak Prof. Dr. Ir. Bambang S. Purwoko, M.Sc sebagai ketua komisi pembimbing dalam penyusunan disertasi ini. Bimbingan beliau yang intensif, cermat dan terarah, memberikan tuntunan kepada penulis cara berpikir analitis dan sistematis. Selain itu beliau juga banyak memberikan tuntunan tentang disiplin, tawakal dan kesabaran sehingga penulis termotivasi dalam menyelesaikan disertasi ini.
sebesar-besarnya karena telah memberikan bimbingan intensif, motivasi, informasi dan kritik yang sangat berharga dalam penyelesaian disertasi ini.
Penulis sampaikan juga penghargaan dan terima kasih kepada Bapak Dr. Ir. Anas D. Susila, M.Si sebagai penguji luar komisi pada ujian prakualifikasi dan ujian tertutup serta Bapak Dr. Ir. Yusdar Hilman MS dan Ibu Dr. Ir. Trikoesoemaningtyas, M.Sc keduanya sebagai penguji luar komisi pada ujian terbuka. Pertanyaan dan saran yang disampaikan Bapak dan Ibu sangat berarti dalam perbaikan penulisan disertasi ini.
Kepada Rektor Universitas Negeri Papua (Unipa) dan Dekan Fakultas Pertanian dan Teknologi Pertanian Unipa, penulis sampaikan terima kasih karena telah memberikan izin untuk melanjutkan studi di IPB.
Kepada Rektor Institut Pertanian Bogor (IPB), Dekan Sekolah Pascasarjana IPB, Staf Pengajar, Staf Administrasi dan Staf University Farm, penulis sampaikan terima kasih karena telah memberikan kesempatan, bimbingan ilmu dan pelayanan yang baik selama melaksanakan studi di IPB. Penulis bangga bisa menjadi bagian dari keluarga besar IPB.
Ucapan terima kasih penulis sampaikan kepada Departemen Pendidikan Nasional yang telah memberikan beasiswa melalui dana DUE-Like batch 3.
Khususnya kepada orang tua penulis Ayah H. Saryono SW (Alm) dan Ibu Hj. Mudjidjatin, serta istri tercinta Ir. Iva Purnomo, M.Si, Ibu mertua Ny. Sundari, Ayah mertua Bpk. Amir Nanza (Alm) serta keluarga : Om Setyo, Mbak Yayuk, Dik Anto, Dodo, Toto, Ita, Ikhlas, dan keluarga saudara ipar : Dik Mirna, Dik Asrul, Alma, Nia, dan semua saudaraku terima kasih atas doa, cinta kasih dan dukungannya.
Kepada keluarga rekan Charlie, Pak Takdir, Mas Anton, Mas Yulius, Pak Irba, Pak Imam, rekan Jhon Marwa dan semua pihak yang telah membantu baik berupa tenaga, saran atau kesediaannya meminjamkan literatur, penulis ucapkan banyak terima kasih. Akhirnya semoga disertasi ini dapat bermanfaat bagi pengembangan ilmu pengetahuan.
Bogor, Januari 2008
RIWAYAT HIDUP
Penulis dilahirkan di Kabupaten Brebes pada tanggal 21 April 1968 sebagai anak kedua dari tujuh bersaudara, dari pasangan Ayah H. Saryono Siswowardoyo (Alm.) dan Ibu Hj. Mudjidjatin. Pada tahun 1995 penulis menikah dengan Ir. Iva Purnomo, M.Si.
Pendidikan dasar sampai menengah pertama di selesaikan di SD Negeri I dan SMP Negeri I Nabire, Papua. Penulis melanjutkan ke pendidikan menengah atas di SPMA Negeri Manokwari, Papua Barat. Pendidikan Sarjana diselesaikan pada tahun 1992 di Fakultas Pertanian Universitas Cenderawasih Manokwari. Pada tahun 1999, penulis mendapat kesempatan melanjutkan studi Magister Sains di Program Studi Agronomi, Sekolah Pascasarjana IPB Bogor dan lulus pada tahun 2002. Pada tahun 2002 di tempat yang sama penulis melanjutkan ke Program Doktor pada Program Studi Agronomi.
DAFTAR ISI
Halaman
DAFTAR TABEL ………... xiii
DAFTAR GAMBAR ………...………... xvi
PENDAHULUAN ………... 1
Latar Belakang ………. 1
Tujuan Penelitian ………. 4
Hipotesis ... 4
TINJAUAN PUSTAKA ………. 6
Permasalahan pada Tanah Ultisol ... 6
Pengaruh Cekaman Aluminium pada Tanaman ... 7
Adaptasi Tanaman terhadap Cekaman Aluminium ... 9
Struktur Umum Fungi Mikoriza Arbuskula ... 12
Keragaman dan Manfaat FMA bagi Tanaman ... 14
Syarat Tumbuh Tanaman Cabai ... 17
PENAPISAN GENOTIPE CABAI (Capsicum annuum L.) TOLERAN ALUMINIUM BERDASARKAN PERBEDAAN PANJANG AKAR PADA FASE VEGETATIF ... 19
Abstrak ... 19
Abstract ... 19
Pendahuluan ... 20
Bahan dan Metode ... 21
Hasil dan Pembahasan ... 25
Simpulan ... 32
EVALUASI GENOTIPE CABAI (Capsicum annuum L.) BERDASARKAN TANGGAP PERTUMBUHAN DAN HASIL UNTUK TOLERANSI TERHADAP ALUMINIUM ... 33
Abstrak ... 33
Abstract ... 33
Pendahuluan ... 34
Bahan dan Metode ... 35
Hasil dan Pembahasan ... 38
Simpulan ... 47
KOMPATIBILITAS JENIS FUNGI MIKORIZA ARBUSKULA DENGAN CABAI (Capsicum annuum L.) ... 48 Abstrak ... 48
Abstract ... 48
Pendahuluan ... 49
Bahan dan Metode ... 50
Hasil dan Pembahasan ... 54
TANGGAP PERTUMBUHAN DAN HASIL TERHADAP INOKULASI FUNGI MIKORIZA ARBUSKULA PADA CABAI (Capsicum annuum
L.) YANG MENGALAMI CEKAMAN ALUMINIUM ... 58
Abstrak ... 58
Abstract ... 58
Pendahuluan ... 59
Bahan dan Metode ... 60
Hasil dan Pembahasan ... 63
Simpulan ... 75
TANGGAP FISIOLOGI TERHADAP INOKULASI FUNGI MIKORIZA ARBUSKULA PADA CABAI (Capsicum annuum L.) YANG MENGALAMI CEKAMAN ALUMINIUM ... 76
Abstrak ... 76
Abstract ... 76
Pendahuluan ... 77
Bahan dan Metode ... 78
Hasil dan Pembahasan ... 84
Simpulan ... 92
PEMBAHASAN UMUM ... 93
SIMPULAN DAN SARAN ... 99
Simpulan ... 99
Saran ... 99
DAFTAR PUSTAKA ... 100
DAFTAR TABEL
Nomor
Teks
Halaman 1 Identifikasi FMA indigen tanah podsolik pada lahan pertanaman
padi gogo dan ubi kayu (Iriani 2003) ... 14 2 Jumlah kebutuhan kapur CaCO3 untuk memperoleh kejenuhan
aluminium tanah ultisol yang berbeda ... 24 3 Karasteristik sifat tanah ultisol asal Gajrug, Lebak Banten sebelum
dan sesudah pemberian kapur ... 26 4 Nilai kuadrat tengah dari hasil analisis ragam pengaruh genotipe
cabai dan tingkat kejenuhan Al pada panjang akar, bobot kering akar dan bobot kering tajuk ... 27 5 Tanggap panjang akar, bobot kering akar dan bobot kering tajuk
terhadap tingkat kejenuhan Al pada tanah Ultisol ... 28 6 Pengelompokan genotipe cabai untuk toleransi terhadap cekaman
Al berdasarkan nilai panjang akar relatif ... 29 7 Nama genotipe cabai hasil penapisan berdasarkan panjang akar
pada fase vegetatif yang dievaluasi untuk toleransi terhadap cekaman Al ... 35 8 Tanggap tinggi tanaman terhadap cekaman Al pada cabai yang
ditanam di tanah Ultisol ... 39 9 Tanggap bobot kering tajuk terhadap cekaman Al pada cabai yang
ditanam di tanah Ultisol ... 39 10 Tanggap jumlah buah total terhadap cekaman Al pada cabai yang
ditanam di tanah Ultisol ... 41 11 Tanggap jumlah buah panen terhadap cekaman Al pada cabai
yang ditanam di tanah Ultisol ... 41 12 Tanggap panjang buah terhadap cekaman Al pada cabai yang
ditanam di tanah Ultisol ... 42 13 Tanggap bobot per buah terhadap perlakuan genotipe cabai dan
kondisi cekaman pada tanah Ultisol ... 43 14 Tanggap bobot buah panen terhadap cekaman Al pada cabai yang
ditanam di tanah Ultisol ... 44 15 Konsistensi tingkat toleransi Al antara hasil penapisan karakter
panjang akar dan evaluasi karakter agronomi ... 45 16 Perbedaan jumlah propagul infektif pada beberapa inokulum FMA 54 17 Derajat infeksi berbagai jenis FMA pada akar cabai ... 55 18 Nilai kuadrat tengah dari hasil analisis ragam pengaruh genotipe
19 Perbedaan bobot kering akar, bobot kering tajuk dan nisbah tajuk-akar pada genotipe cabai yang diinokulasi berbagai jenis FMA .... 57 20 Pengaruh cekaman Al terhadap derajat infeksi Gigaspora
margarita pada beberapa genotipe cabai ... 64 21 Perbedaan panjang akar akibat cekaman Al pada beberapa
genotipe cabai ... 65 22 Perbedaan tinggi tanaman akibat cekaman Al pada beberapa
genotipe cabai ... 66 23 Perbedaan bobot kering tajuk akibat cekaman Al pada beberapa
genotipe cabai ... 67 24 Perbedaan jumlah buah panen akibat cekaman Al pada beberapa
genotipe cabai ... 68 25 Perbedaan panjang buah akibat cekaman Al pada beberapa
genotipe cabai ... 68 26 Perbedaan bobot per buah akibat cekaman Al pada beberapa
genotipe cabai ... 69 27 Perbedaan bobot buah panen akibat cekaman Al pada beberapa
genotipe cabai ... 69 28 Peningkatan tinggi tanaman, bobot kering tajuk, jumlah buah
panen, panjang buah, bobot per buah dan bobot buah panen oleh inokulasi Gigaspora margarita pada kondisi cekaman Al yang berbeda. ... 70 29 Tanggap jumlah buah panen akibat inokulasi Gigaspora margarita
pada genotipe cabai yang tercekam Al ... 72 30 Tanggap bobot buah panen akibat inokulasi Gigaspora margarita
pada genotipe cabai yang tercekam Al ... 72 31 Pengaruh cekaman Al terhadap derajat infeksi Gigaspora
margarita pada genotipe toleran dan peka ... 84 32 Perbedaan kandungan asam malat, sitrat dan oksalat dalam akar
dan tajuk akibat cekaman Al pada genotipe toleran dan peka ... 85 33 Kandungan asam malat, sitrat dan oksalat dalam akar dan tajuk
akibat inokulasi FMA pada kondisi tanpa cekaman Al dan tercekam Al ... 87 34 Perbedaan kandungan aluminium dalam akar dan tajuk akibat
cekaman Al pada genotipe toleran dan peka ... 88 35 Pengaruh inokulasi FMA terhadap kandungan N dan P pada
Lampiran
1 Beberapa karakteristik genotipe cabai yang digunakan dalam percobaan (koleksi Laboratorium Genetika dan Pemuliaan Tanaman Departemen Agronomi dan Hortikultura, Faperta IPB) .. 108 2 Rekapitulasi hasil analisis ragam pada percobaan evaluasi karakter
pertumbuhan dan hasil untuk toleransi terhadap cekaman Al…….. 109 3 Rekapitulasi hasil analisis ragam pada percobaan tanggap
pertumbuhan dan hasil terhadap inokulasi FMA pada genotipe cabai yang mengalami cekaman Al ………. 109 4 Prosedur pengamatan koloni FMA dengan teknik pewarnaan akar
untuk menghitung derajat infeksi akar ... 110 5 Rekapitulasi hasil analisis ragam pada percobaan tanggap fisiologi
DAFTAR GAMBAR
Nomor
Halaman 1 Bagan alur kegiatan penelitian ... 5 2 Pengaruh Al terhadap effluks asam malat melalui saluran protein
(Delhaize & Ryan 1995) ... 9 3 Struktur miselium intraradikal pada FMA: (a) yang membentuk
struktur arbuskula dan vesikula, dan (b) hanya arbuskula (Brundrett et al. 1996) ... 13 4 Grafik penurunan nilai kejenuhan Al tanah ultisol akibat
pemberian kapur CaCO3. ... 23
5 Perbedaan panjang akar cabai umur 6 MST antara genotipe PBC 619 (1) dan Cilibangi 3 (2) pada tanah ultisol dengan tingkat kejenuhan Al berturut-turut 0.77%, 15.92%, 31.96%, 60.85% dan 83.48% ... 31 6 Korelasi antara panjang akar relatif dan bobot buah panen ... 46 7 Pengaruh inokulasi Gigaspora margarita pada genotipe cabai
yang toleran Al (1=PBC 619) dan peka Al (2=Cilibangi 3). Kedua genotipe cabai ditanam pada kondisi tercekam Al (kejenuhan Al= 60.85%) ... 73 8 Peningkatan jumlah buah panen, bobot per buah dan bobot buah
panen karena inokulasi Gigaspora margarita pada genotipe cabai yang mengalami cekaman Al ... 74 9 Panjang akar beberapa genotipe cabai yang bersimbiosis dengan
Gigaspora margarita ... 75 10 Perubahan kandungan total asam organik dalam akar dan tajuk
pada kondisi tanpa cekaman Al dan tercekam Al. ... 86 11 Kandungan Al dalam akar maupun tajuk akibat inokulasi FMA
PENDAHULUAN
Latar Belakang
Cabai (Capsicum annuum L) merupakan salah satu komoditas sayuran yang bernilai ekonomi tinggi. Hal ini terlihat dari areal pertanaman cabai yang menempati areal terluas diantara tanaman sayuran yang diusahakan di Indonesia. Areal pertanaman cabai pada tahun 2005 seluas 103 531 ha dan pada tahun 2006 meningkat menjadi 113 079 ha atau menempati 18.27% dari total luas pertanaman sayuran (Departemen Pertanian 2007). Bertambahnya luas areal tersebut dilakukan untuk memenuhi kebutuhan cabai dalam negeri.
Kebutuhan cabai terus meningkat sejalan dengan bertambahnya jumlah penduduk dan berkembangnya industri makanan, kosmetik, serta farmasi yang menggunakan cabai sebagai bahan baku. Data statistik menunjukkan bahwa konsumsi cabai mencapai 4.65 kg per kapita per tahun, jika diasumsikan penduduk yang mengkonsumsi cabai berumur 15 tahun ke atas sebanyak 170 juta maka diperkirakan kebutuhan cabai dalam negeri sebesar 790 500 ton per tahun. Di lain pihak, produksi cabai secara nasional baru mencapai 736 019 ton dengan produktivitas sebesar 6.51 ton/ha (Departemen Pertanian 2007), sehingga produksi cabai perlu ditingkatkan untuk memenuhi kebutuhan di dalam negeri.
Sentra produksi cabai di Indonesia sekitar 80% masih berada di Pulau Jawa, namun ketersediaannya pada masa yang akan datang tidak lagi dapat diandalkan dari Pulau Jawa. Hal ini disebabkan semakin berkurangnya lahan-lahan pertanian di Pulau Jawa akibat dikonversi menjadi lahan-lahan-lahan-lahan non pertanian, sehingga perluasan areal pertanaman cabai di luar Pulau Jawa perlu mendapat perhatian.. Lahan kering di luar Pulau Jawa yang potensial untuk lahan pertanian luasnya mencapai 132.88 juta hektar atau 92.31% dari total luas lahan kering di Indonesia (Hidayat & Mulyani 2002). Namun, 33.58% dari luas lahan tersebut atau seluas 44.62 juta ha merupakan jenis tanah Ultisol yang tersebar di Sumatra, Kalimantan, Sulawesi, Maluku, serta Papua.
sehingga menurunkan ketersediaan hara P (Ralalage et al. 1995; Baligar et al. 1997). Selain itu, kelarutan Al pada pH kurang dari 4.5 didominasi bentuk Al 3+ yang dapat menghambat pertumbuhan akar sehingga menurunkan kemampuan akar dalam menyerap hara mineral dan air (Marschner 1995; Rout et al. 2001).
Masalah cekaman Al pada tanah Ultisol dapat diatasi dengan memperbaiki kondisi tanah melalui pengapuran (Naidu et al. 1990; Idris 1995; Nurlaeny et al. 1998), namun pendekatan ini tidak ekonomis karena dibutuhkan dalam jumlah yang banyak dalam aplikasinya. Pada daerah-daerah dengan sarana transportasi terbatas dan jauh dari sumber industri kapur akan terkendala dengan tingginya biaya pengadaan. Penggunaan genotipe yang adaptif terhadap cekaman Al dapat dijadikan alternatif karena lebih praktis (Samac & Tesfaye 2003; Bakhtiar et al. 2007). Genotipe adaptif Al dapat tumbuh dan berproduksi dengan baik pada tanah Ultisol karena adanya peningkatan sintesis asam-asam organik untuk mendetoksifikasi Al (Ma 2000; Watanabe et al. 2006; Enggarini & Marwani 2006). Asam-asam organik akan membentuk kompleks Al-asam organik, sehingga mengurangi toksisitas Al pada tanaman (Jones & Brassington 1998; Ma et al. 2001).
3 hanya sampai fase vegetatif. Oleh karena itu hasil penapisan berdasarkan panjang akar perlu dievaluasi lebih lanjut sampai periode panen.
Perbaikan daya adaptasi tanaman terhadap cekaman abiotik juga dapat dilakukan dengan memanfaatkan simbiosis antara tanaman dan fungi mikoriza arbuskula (FMA). Pemanfaatan FMA telah banyak diketahui mampu meningkatkan adaptasi tanaman terhadap cekaman abiotik, seperti pada cabai terhadap cekaman hara P maupun kekeringan (Haryantini & Santoso 2001; Rahayu et al. 2002), serta jagung, kedelai, dan kacang tunggak terhadap cekaman Al pada tanah ultisol (Nurlaeni et al. 1996; Hanum 2004; Rohyadi et al. 2004). Fungi mikoriza arbuskula memberikan keuntungan pada tanaman melalui ekspansi hifa eksternalnya sampai ke lapisan subsoil sehingga meningkatkan kapasitas penyerapan hara dan air (Cruz et al. 2004).
Fosfat adalah unsur hara utama yang diserap tanaman dengan bantuan mikoriza (Joner & Johansen 2000; Rohyadi et al. 2003). Inokulasi FMA pada cabai dapat meningkatkan penyerapan P sebesar 30.95% di tanah andisol (Haryantini & Santoso 2001). Selain itu, koloni FMA dapat menginduksi aktivitas fosfatase asam dan sintesis asam organik sehingga dapat memperbaiki kondisi rizosfir (Joner et al. 1995; Joner & Johansen 2000).
Potensi pemanfaatan tanah Ultisol untuk budidaya cabai dapat diketahui melalui serangkaian penelitian untuk mendapatkan genotipe yang adaptif terhadap cekaman Al serta jenis FMA yang kompatibel dan efektif dalam perbaikan hasil dan adaptasi cabai pada tanah Ultisol. Penanaman genotipe yang adaptif terhadap cekaman Al dan pemanfaatan jenis FMA yang efektif diharapkan dapat meningkatkan produktivitas tanaman cabai pada tanah Ultisol. Adapun alur penelitian secara lengkap disajikan pada Gambar 1.
Tujuan Penelitian Penelitian ini bertujuan untuk :
1. Mengidentifikasi genotipe cabai yang toleran dan peka Al secara cepat melalui pengamatan perbedaan panjang akar pada fase vegetatif.
2. Mengevaluasi genotipe hasil penapisan uji hayati akar berdasarkan karakter pertumbuhan dan hasil sehingga diperoleh genotipe yang diindikasikan adaptif terhadap cekaman Al.
3. Mendapatkan FMA yang kompatibel dengan tanaman cabai.
4. Menguji keefektifan FMA dalam meningkatkan hasil dan kemampuan adaptasi cabai pada tanah masam berkadar Al tinggi.
5. Menguraikan mekanisme adaptasi terhadap cekaman Al pada tanaman cabai yang bermikoriza.
Hipotesis
1. Melalui penapisan panjang akar pada fase vegetatif dan evaluasi pertumbuhan dan hasil, dapat diperoleh genotipe yang adaptif terhadap cekaman Al.
2. Terdapat perbedaan kemampuan kolonisasi akar antar jenis FMA dengan cabai.
5
Keterangan :
= garis pemanfaatan data
= garis target/hasil yang diharapkan
KEEFEKTIFAN FUNGI MIKORIZA ARBUSKULA DALAM MENINGKATKAN HASIL DAN ADAPTASI CABAI PADA TANAH BERCEKAMAN ALUMINIUM
Tahap 1: Identifikasi kemampuan adaptasi genotipe cabai terhadap cekaman Al. 1a. Penentuan tingkat kejenuhan Al
tanah ultisol dengan pengapuran 1b. Penapisan pada fase vegetatif 1c. Evaluasi karakter pertumbuhan
dan komponen hasil
Tahap 2. Pengujian kompatibilitas fungi mikoriza arbuskula (FMA) dan cabai.
2a. Penafsiran jumlah propagul infektif
2b. Pengujian daya infeksi FMA
Tahap 3. Pengujian efektifitas FMA untuk perbaikan hasil dan kemampuan adaptasi cabai pada tanah ultisol.
3a. Pengujian terhadap tanggap pertumbuhan dan hasil 3b. Pengujian terhadap tanggap
fisiologi
- Informasi karakter
pertumbuhan akar dan tajuk, serta komponen hasil untuk seleksi terhadap cekaman Al - Diperoleh genotipe toleran
dan peka Al
Jenis FMA yang kompatibel dengan cabai
- Peningkatan kemampuan adaptasi melalui perbaikan pertumbuhan dan hasil oleh FMA
- Informasi karakter fisiologi yang berkaitan dengan mekanisme adaptasi terhadap cekaman Al
PENINGKATAN PRODUKTIVITAS CABAI PADA TANAH ULTISOL
Permasalahan pada Tanah Ultisol
Jenis tanah yang berordo Ultisol merupakan salah satu jenis tanah yang cukup potensial digunakan sebagai lahan pertanian, namun karena tingkat kesuburan tanahnya rendah menyebabkan tanah tersebut masih termarjinalkan. Luas tanah ultisol mencapai 45.794 juta ha atau 24.3% dari total luas lahan kering di Indonesia (Hidayat & Mulyani 2002). Sebaran tanah Ultisol tersebut terdapat di Kalimantan (47.91%), Sumatra (20.68%), Papua (16.63%), Sulawesi (9.39%), Maluku (2.71%) dan Jawa+Bali+Nusa Tenggara (2.68%). Konsepsi pokok dari tanah Ultisol adalah tanah yang telah mengalami proses hancuran lanjut (ultimate) dan pencucian berat oleh curah hujan yang tinggi, berwarna merah kuning, berpenampang > 2m, dan terdapat lapisan argilik dari akumulasi liat (Subagyo et al. 2000). Basa-basa akan tercuci dari kompleks jerapan sehingga tinggal Al dan H sebagai kation dominan yang menyebabkan tanah bereaksi masam. Oleh karenanya tanah Ultisol banyak tersebar luas di wilayah tropis basah.
7 misalnya AlOH2+ pada pH 4.5-5.0, Al(OH)2+ pada pH 5.5-6.0, dan Al(OH)4- atau
Al(OH)3 pada pH di atas 6.0 (Marschner 1995).
Adanya kelarutan Al yang tinggi dapat menyebabkan keracunan pada akar sehingga pertumbuhan akar menjadi terhambat dan akhirnya menurunkan kemampuan akar dalam menyerap hara mineral dan air (Matsumoto et al. 1996; Samuel et al. 1997). Selain itu kelarutan Al yang tinggi pada tanah Ultisol dapat juga menyebabkan ketersediaan hara P menurun. Pada reaksi tanah yang masam, ion-ion Al bebas akan memfiksasi ion fosfat membentuk hidroksi fosfat yang tidak larut, akibatnya sering terjadi defisiensi hara tersebut (Ralalage et al. 1995; Rout et al. 2001).
Pengaruh Cekaman Aluminium pada Tanaman
Cekaman Al yang sering terjadi pada tanah Ultisol merupakan penghambat utama bagi pertumbuhan tanaman. Gejala umum yang paling nyata terlihat bila keracunan Al adalah terhambatnya pertumbuhan dan perpanjangan akar, baik akar primer, akar lateral, maupun bulu akar. Tanaman yang keracunan Al mempunyai akar yang pendek, percabangan sedikit, akar adventif lebih banyak tumbuh pada pangkal akar, serta akar primer berkembang melebar ke arah apikal meristem sehingga terlihat gemuk (Rout et al. 2001). Pertumbuhan akar yang demikian sulit melakukan penetrasi ke lapisan sub soil menyebabkan penyerapan hara dan air menjadi lebih rendah (Marschner 1995).
Kelarutan Al sebagai Al3+ merupakan bentuk yang sangat beracun dan dapat merusak akar serta menghambat penyerapan hara mineral (Delhaize & Ryan 1995; Samuel et al. 1997). Bentuk lain seperti Al(OH)2+ juga beracun bagi tanaman terutama kedelai, disamping itu bentuk monomer secara bersamaan dapat lebih menghambat pertumbuhan akar dibandingkan secara individual (Alva et al. 1986; Bushamuka & Zobel 1998). Kerusakan akar akibat keracunan Al juga dilaporkan pada beberapa tanaman, seperti padi (Sivaguru & Paliwal 1993; Bakhtiar et al. 2007), sorgum (Tan et al. 1993), barley (Matsumoto et al. 1996), gandum (Samuel et al. 1997) dan jagung (Bushamuka & Zobel 1998).
membran plasma, selanjutnya Al akan berikatan dengan gugus P pada DNA sehingga menghambat pembelahan sel (Delhaize & Ryan 1995; Matsumoto 1988; Matsumoto et al. 1996). Selain itu, Al dapat juga menggantikan Ca pada ikatan Ca-pektat serta menstimulir sintesis lignin di dalam dinding sel, akibatnya terjadi kekakuan dinding sel dan pemanjangan sel-sel akar menjadi terhambat (Blamey et al. 1993; Watanabe & Okada. 2005). Kalsium sangat dibutuhkan untuk menjaga stabilitas membran dan perkembangan sel. Penggantian Ca oleh Al akan menye-babkan kerusakan pada membran plasma dan terjadi kebocoran sel (Matsumoto et al. 1996; Rengel 1997).
Akumulasi Al pada jaringan akar akan menentukan tingkat toleransi genotipe tanaman terhadap cekaman Al dan berkorelasi dengan tingkat kerusakan akar. Pada genotipe toleran, kandungan Al yang terakumulasi pada jaringan akar umumnya lebih rendah dibandingkan genotipe peka (Delhaize & Ryan 1995; Ma 2000). Hasil deteksi visual penetrasi Al dengan metode pewarnaan hematoksilin pada akar kedelai dari genotipe Yellow (toleran Al ) menunjukkan bahwa akumulasi Al terdapat pada permukaan ujung akar sehingga kerusakan yang terlihat hanya pada bagian tersebut. Sementara itu, pada genotipe Lumut (peka Al) terlihat adanya akumulasi Al sampai ke bagian akar yang lebih tua sehingga kerusakan akar tampak pada jaringan yang lebih dalam dan ke arah jaringan yang lebih tua (Sopandie et al. 2003). Sementara itu, pada tanaman yang sangat peka seperti barley, kerusakan dapat mencapai korteks maupun epidermis dan tidak terbatas pada jaringan meristematik saja (Matsumoto et al. 1996).
Gejala kerusakan akar dapat terlihat dalam waktu yang relatif singkat setelah Al diserap oleh akar. Aluminium dapat menyebabkan kerusakan akar saat masih berada di dinding sel tanpa menunggu masuk ke dalam sel (Delhaize & Ryan 1995). Pada tanaman ercis, perpanjangan akar mulai terhambat setelah 24 jam terkena cekaman Al (Matsumoto et al. 1996). Akar tanaman gandum yang mengalami cekaman Al selama 6 jam memperlihatkan gejala kerusakan akar yang tidak dapat balik (Sasaki et al. 1994).
9 reaksi tanah yang masam, ion-ion Al bebas akan mengfiksasi ion fosfat membentuk hidroksi fosfat yang tidak larut, akibatnya sering terjadi defisiensi hara tersebut (Naidu et al. 1990; Ralalage et al. 1995).
Adaptasi Tanaman terhadap Cekaman Aluminium
Adanya sifat toleransi terhadap kelebihan Al merupakan faktor utama untuk adaptasi tanaman pada tanah masam. Secara umum, kemampuan adaptasi tanaman terhadap cekaman Al dilakukan melalui 2 mekanisme, yaitu : (1) mekanisme eksternal, yaitu sistem adaptasi dengan cara mencegah Al agar tidak masuk ke dalam sel (Pineros et al. 2002), dan (2) mekanisme internal, yaitu sistem adaptasi dengan cara mengurangi toksisitas Al di dalam sel tanaman (Ma 2000; Watanabe et al. 2006). Pada mekanisme eksternal, detoksifikasi Al terjadi di apoplas, sedangkan mekanisme internal terjadi di simplas (Rout et al. 2001).
Tanaman dengan sistem mekanisme toleransi eksternal memiliki kemam-puan untuk melakukan penolakan terhadap Al (eksklusi Al) dari tempat peka di akar. Permeabilitas membran plasma yang selektif terhadap Al dan kapasitas tukar kation (KTK) apoplas yang rendah merupakan bentuk toleransi dengan cara mengeksklusi Al (Taylor 1988). KTK apoplas tinggi akan meningkatkan pertukaran kation-kation polivalen sehingga kemungkinan terjadinya akumu-lasi Al menjadi lebih besar (Matsumoto et al. 1996).
Gambar 2. Pengaruh Al terhadap effluks asam malat melalui saluran protein (Delhaize & Ryan 1995)
2 1
Luar pH 4.0 – 5.0 Sitoplasma
Keseimbangan
Malat-Al Malat
2-K+
Al3+
Saluran Protein pH 7.0
Selain itu, sistem toleransi eksternal dapat juga dilakukan tanaman melalui eksudasi asam-asam organik yang mampu mengkelat Al, sehingga terjadi immobilisasi Al di apoplas dan menurunkan kelarutan Al di daerah rizosfir (Ma 2000). Proses efluks asam organik dari dalam sel akar dapat dilakukan dengan tiga cara, yaitu efluks secara pasif melalui membran bilayer, efluks melalui saluran protein pada membran plasma, dan efluks secara aktif karena adanya gradien potensial elektro-kimia yang diatur oleh H+-ATPase (Jones & Brassington 1998). Pada beberapa tanaman, eksudasi asam-asam organik sebagai respon terhadap cekaman Al sebagian besar diefluks melalui saluran protein (Gambar 2). Menurut Delhaize & Ryan 1995, pengaruh Al terhadap efluks asam malat melalui saluran protein melibatkan tiga mekanisme, yaitu (1) Al berinteraksi langsung dengan saluran protein, sehingga meningkatkan masa waktu pembukaan saluran protein, (2) Al berinteraksi dengan reseptor spesifik pada permukaan membran atau dengan membran yang selanjutnya melalui serangkaian kurir sekunder di sitoplasma akan mengubah aktivitas saluran protein, dan (3) Al masuk ke sitoplasma, kemudian dapat secara langsung mengubah aktivitas saluran protein dengan cara menempel saluran dari dalam, atau secara tidak langsung melalui suatu transduksi sinyal. Di daerah rizosfir, asam-asam organik yang diketahui mempunyai afinitas yang tinggi terhadap logam bervalensi tiga seperti Al3+ dan Fe3+ akan membentuk kompleks Al-asam organik atau Fe-asam organik sehingga mengurangi kelarutan logam-logam tersebut (Jones & Brassington 1998).
11 Peningkatan pH rizosfir pada tanah masam juga merupakan salah satu bentuk mekanisme toleransi eksternal terhadap cekaman Al. Kemampuan tanaman untuk menaikkan pH dapat mengurangi pengaruh buruk Al melalui proses hidrolisis dan polimerasi Al menjadi bentuk yang kurang beracun (Taylor, 1988). Keseimbangan penyerapan kation-anion, terutama amonium (NH4+) dan
nitrat (NO3-), merupakan salah satu parameter yang penting dalam sistem
pengaturan pH rizosfir. Pada tanaman kedelai, naiknya pH berkaitan dengan kemampuan genotipe toleran untuk menyerap nitrat lebih banyak dibandingkan amonium. Hal ini juga disertai dengan tingginya aktivitas spesifik NR (nitrat reduktase) pada akar genotipe toleran (Sopandie 1999). Nitrat yang diserap tanaman akan direduksi menjadi asam-asam organik dan menyebabkan ekskresi OH- sehingga pH rizosfir meningkat (Haynes 1990).
Kemampuan tanaman untuk mendetoksifikasi Al dalam sitosol merupakan ciri spesies tanaman yang mempunyai sistem mekanisme toleransi internal. Tanaman mampu mempertahankan proses metabolismenya, walaupun Al telah masuk ke dalam sitosol. Mekanisme ini dapat berupa pengkelatan Al di sitosol, kompartementasi Al ke dalam vakuola, sintesis protein pengikat Al dan evolusi enzim yang toleran Al (Taylor 1988).
Peningkatan akumulasi asam-asam organik pada tanaman yang toleran Al mengindikasikan adanya peranan asam-asam organik dalam mendetoksifikasi Al dalam jaringan tanaman. Detoksifikasi Al oleh asam organik dapat dilakukan dengan cara mengkelat Al pada kelompok karboksilat dari molekul asam organik membentuk kompleks Al-asam organik yang stabil sehingga tidak menyebabkan keracunan bagi tanaman (Ma 2000). Pada akar tanaman jagung yang toleran terdapat akumulasi asam malat dan trans-akonitat yang diduga dapat mendetoksifikasi Al dalam sitosol (Suhayda & Haug 1986). Sementara itu, pada tanaman Hydrangea macrophylla detoksifikasi Al di sitosol dilakukan oleh asam sitrat (Ma et al. 1997).
aktivitas transport H+ tonoplas mempunyai peranan yang penting dalam pertukaran proton dengan Al yang dikompartementasi di vakuola.
Pada tanaman gandum, peningkatan aktivitas beberapa enzim merupakan salah satu bentuk toleransi terhadap Al. Slaski et al. (1996) melaporkan adanya peningkatan enzim-enzim glukosa-6-fosfat dehidrogenase dan 6-fosfoglukonat dehidrogenase pada genotipe gandum yang toleran Al. Kedua enzim tersebut terlibat dalam biosintesis lintasan pentosa fosfat yang diduga dapat berperan dalam sistem toleransi tanaman terhadap cekaman Al.
Struktur Umum Fungi Mikoriza Arbuskula
Mikoriza merupakan suatu struktur khas pada sistem perakaran yang terbentuk karena adanya simbiosis mutualistik antara fungi (myces) dan akar (rhiza) dari tumbuhan tingkat tinggi. Bentuk struktur khas mikoriza dapat dibedakan berdasarkan cara infeksinya pada perakaran tanaman inang, yaitu : (1) endomikoriza, merupakan struktur mikoriza yang terbentuk sampai ke dalam sel korteks akar, (2) ektomikoriza, merupakan struktur mikoriza pada lapisan luar akar yang bentuknya berupa jala hartig; dan (3) ektendomikoriza, merupakan struktur mikoriza yang tidak hanya dapat membentuk jala hartig di permukaan akar, tetapi dapat menembus sel korteks (Smith & Read, 1997). Salah satu simbion fungi yang banyak membentuk struktur endomikoriza pada tanaman pertanian adalah fungi mikoriza arbuskula (FMA). Secara taksonomi FMA termasuk ke dalam kelas Zygomycetes, ordo Glomales yang terbagi dalam tiga famili yaitu Gigasporaceae, Glomaceae, dan Acaulosporaceae. Masing-masing famili terbagi atas dua genus, secara berurutan adalah Gigaspora, Scutelospora, Glomus, Sclerocyctis, Aculospora, dan Entrophospora.
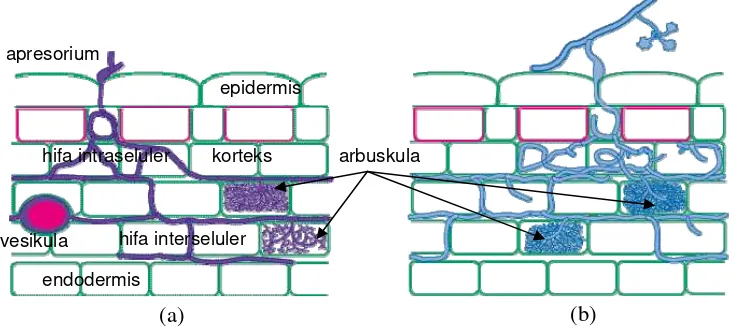
13 simbion, sedangkan vesikula berfungsi sebagai organ penyimpan cadangan makanan. Tidak semua jenis FMA membentuk kedua struktur tersebut, seperti jenis Glomus, Sclerocystis, Acaulospora, dan Entrophospora dapat membentuk struktur arbuskula maupun vesikula, sedangkan jenis lainnya seperti Gigaspora dan Scutellospora hanya membentuk struktur arbuskula (Brundrett et al. 1996). Struktur miselium intraradikal dari FMA pada akar tanaman dapat dilihat pada Gambar 3.
Gambar 3. Struktur miselium intraradikal pada FMA: (a) yang membentuk struktur arbuskula dan vesikula, dan (b) hanya arbuskula (Brundrett et al. 1996).
Secara umum proses koloni FMA pada akar tanaman melewati empat tahap, yaitu (1) induksi perkecambahan spora dan pertumbuhan hifa, (2) kontak antara hifa dan permukaan akar yang menyebabkan pengenalan dan pembentukan apresorium, (3) penetrasi hifa ke dalam akar, dan (4) perkembangan struktur hifa interseluler (Bonfante & Perotto 1995). Koloni FMA pada akar tanaman inang diawali dengan kontak hifa dan pelekatan pada akar, setelah 2-3 hari terjadi pembentukan apresorium yang membengkak. Perkembangan selanjutnya akan terjadi pengecilan diameter hifa membentuk ujung yang agak runcing, dan kemudian diikuti dengan ekspansi hifa memasuki lumen sel dan berkembang diantara rongga-rongga interselluler korteks. Hifa interselluler kemudian berkembang membentuk struktur percabangan (arbuskula) dan pada beberapa cendawan dapat juga membentuk struktur oval (vesikula). Meskipun
vesikula
arbuskula epidermis
korteks
endodermis apresorium
hifa intraseluler
(a) (b)
perkembangan arbuskula dan vesikula terlihat menembus dinding sel korteks, namun kenyataannya membran plasmanya tidak rusak, tetapi berkembang mengelilingi bentuk arbuskula atau vesikula membentuk kompartemen apoplastik baru yang disebut bidang kontak (Smith & Smith 1996).
Keragaman dan Manfaat FMA bagi Tanaman
[image:31.612.119.482.453.650.2]Fungi mikoriza arbuskula merupakan tipe mikoriza yang paling banyak mendapat perhatian, karena diketahui dapat bersimbiosis dengan sekitar 80% spesies tanaman (Brundrett et al. 1996). Keragaman FMA dapat ditemukan pada berbagai ekosistem, baik yang masih utuh maupun yang telah mengalami perubahan. Eksplorasi FMA pada daerah perkebunan kelapa sawit telah berhasil diidentifikasi jenis-jenis FMA, antara lain Acaulospora tuberculata, A. foveata, A. mellea , A. scrobiculata, Glomus fluegianum, G. rubiforme, G. sinuosum, dan G. globisporum (Widiastuti & Kramadibrata, 1993). Sementara itu, eksplorasi pada bukit pasir di daerah pantai banyak ditemukan FMA dari jenis Glomus, Sclerocystis, Acaulospora, dan Gigaspora (Setiadi 2000). Pada tanah masam podsolik juga telah diidentifikasi jenis-jenis FMA indigen pada pertanaman padi gogo dan ubi kayu (Tabel 1).
Tabel 1. Identifikasi FMA indigen tanah podsolik pada lahan pertanaman padi gogo dan ubi kayu (Iriani 2003)
Jenis pertanaman
Jenis CMA indigen yang ditemukan
Kerapatan (spora/g tanah) Padi gogo Acaulospora laevis
Glomus agregatum Glomus etunicatum Gigaspora margarita Glomus fasciculatum
23 14 12 10 4 Ubi kayu Glomus agregatum
Gigaspora margarita Glomus manihotis
Glomus etunicatum Acaulospora laevis Glaziella spp.
Glomus macrocarpum
31 25 16 9 7 3 3
15 Bioteknologi Kehutanan Pusat Studi Hayati dan Bioteknologi IPB (Setiadi 2000). Koleksi berbagai isolat FMA dapat dijadikan sumber plasma nutfah yang sangat potensial dan dapat digunakan untuk kegiatan penelitian maupun kepentingan lainnya. Beberapa isolat yang telah dibuat biakan murninya (monosenic) adalah : Glomus manihotis (INDO-1), Acaulospora tuberculata (INDO-2), Glomus mossea (INDO-3), Acaulospora delicata (INDO-4), Acaulospora scrobiculata (INCO-1), Acaulospora sp (INCO-2), Gigaspora rosea (FL.105), Gigaspora margarita (FL.1-5), Glomus etunicatum (FL.126), dan Glomus sp(BIO-4).
Secara alami terdapat asosiasi mikoriza antara fungi dan tanaman dalam bentuk simbiosis mutualisme. Berdasarkan aspek fungsional, simbiosis mutualisme dikatakan berhasil apabila kedua simbion dapat memanfaatkan fungsi simbiosis secara maksimal. Indikator keberhasilan tersebut dapat terlihat dari pertumbuhan dan perkembangan kedua simbion. Manfaat fungsional yang diperoleh FMA dapat dilihat dari adanya pembentukan struktur arbuskula dan atau vesikula di dalam sel-sel akar serta produksi spora yang tinggi. Perkembangan FMA dan produksi spora membutuhkan energi yang diperoleh melalui penyerapan C organik dari tanaman inang (Smith & Read 1997). Sementara itu, tanaman inang dapat memanfaatkan fungsi simbiosis berupa hara mineral dan air yang penyerapannya dibantu oleh FMA sehingga pertumbuhan dan hasil tanaman meningkat.
Transfer hara antara fungi dan tanaman terjadi dalam struktur arbuskula. Fungi menyerap carbon dalam bentuk karbohidrat terlarut kemudian dengan cepat mengkonversinya menjadi bentuk lipid dan glikogen, yang selanjutnya disimpan di dalam struktur hifa maupun vesikula (Brundrett et al. 1996). Sukrosa dan glukosa merupakan bentuk utama karbohidrat terlarut yang ditranslokasikan ke akar tanaman yang bermikoriza (Bago et al. 2000).
bersimbiosis dengan cabai adalah Gigaspora margarita (Haryantini & Santoso 2001), dan Glomus agregatum (Rahayu et al. 2002).
Adanya simbiosis dengan FMA telah banyak diketahui mampu memper-baiki hara tanaman inang melalui penyerapan hara dan air yang pada akhirnya akan meningkatkan pertumbuhan dan hasil tanaman. Inokulasi FMA pada cabai dapat meningkatkan serapan P (Haryantini & Santoso 2001) dan meningkatkan adaptasi terhadap kekeringan (Rahayu et al. 2002). Pada tanah Ultisol, inokulasi FMA juga dapat meningkatkan serapan P pada jagung (Nurlaeni et al. 1996), meningkatkan adaptasi kedelai terhadap cekaman kekeringan (Hapsoh et al. 2005) maupun cekaman ganda Al dan kekeringan (Hanum 2004). Fungi mikoriza arbuskula yang menginfeksi sistem perakaran tanaman inang akan memproduksi jalinan hifa eksternal yang dapat tumbuh secara ekspansif dan menembus lapisan subsoil sehingga meningkatkan kapasitas akar dalam penyerapan hara dan air. Volume tanah yang dapat dijelajahi oleh 1 cm akar yang bermikoriza dapat mencapai 12-15 cm3 atau 6-15 kali lebih banyak dibandingkan akar tanaman tidak bermikoriza yang hanya dapat menjelajahi sekitar 1-2 cm3 (Cruz et al. 2004).
Fosfat merupakan unsur hara utama yang dapat diserap oleh tanaman yang bermikoriza (Bolan 1991). Secara fungsional, FMA dapat membantu penyerapan hara terutama P melalui tiga tahap, yaitu (1) P diserap oleh hifa dari larutan tanah, (2) translokasi P melalui hifa, dan (3) transfer P melewati interfase ruang fungi (Cruz et al. 2004). Pergerakan P anorganik secara difusi dari larutan tanah ke rizosfir sangat lambat sehingga terjadi daerah deplesi P di sekitar perakaran. Adanya hifa eksternal memungkinkan tanaman dapat mengeksploitasi P di luar daerah deplesi (Joner et al. 1995). Pada tanaman bermikoriza daerah deplesi P di sekitar perakaran akan semakin luas, karena eksploitasi P oleh hifa eksternal cukup tinggi (Bolan 1991).
17 Selain itu, FMA juga membantu penyerapan hara N, K dan Mg yang mobil, serta unsur mikro seperti Cu, Zn, Mn, B dan Mo (Smith & Read 1997). Inokulasi FMA pada tanaman kedelai mampu meningkatkan serapan hara N, P, dan K serta dapat memperbaiki toleransi tanaman terhadap cekaman ganda Al dan kekeringan (Hanum 2004).
Kondisi tanah yang terdegradasi umumnya tingkat kesuburan tanahnya rendah, dan ada yang mengandung unsur yang beracun bagi tanaman. Pemanfaatan FMA telah banyak dilaporkan mampu meningkatkan toleransi tanaman terhadap tanah masam (Clark 1997; Cuenca et al. 2001), dan mampu bertindak sebagai bio-remediator pada tanah yang tercemar logam berat seperti tembaga, nikel, seng dan timbal (Shetti et al. 1995; Setiadi 2000).
Fungsi FMA dalam meningkatkan toleransi tanaman terhadap Al belum dibahas secara mendalam, namun beberapa penelitian mengindikasikan bahwa FMA dapat memperbaiki kondisi rizosfir sehingga meningkatkan daya adaptasi tanaman terhadap cekaman Al pada tanah masam. Bentuk mekanisme tersebut diduga melibatkan induksi sintesis asam-asam organik dan aktivitas fosfatase asam, serta perubahan pH rizosfir. Hifa eksternal dari FMA sangat aktif menyerap nitrat sehingga menginduksi peningkatan pH rizosfir (Bago et al. 1996). Selain itu, banyak ditemukan Al dalam sel dari micelium FMA yang diinokulasikan pada Clusia multifloria (Cuenca 2001), namun bagaimana Al dikompartementasikan dalam sitoplasma belum diketahui. Beberapa jenis FMA yang diketahui dapat berperan pada kondisi tanah masam dan konsentrasi Al yang tinggi antara lain: Glomus manihotis, Gigaspora sp, dan Acaulospora sp (Clark 1997).
Syarat Tumbuh Tanaman Cabai
menghambat pembungaan. Suhu optimum untuk pertumbuhan vegetatif adalah 160 - 200 C (Sumarni, 1996).
Sementara itu Rubatzky & Yamaguchi (1996), menyatakan bahwa intensitas curah hujan yang sesuai untuk pertumbuhan tanaman cabai adalah 600 – 1.200 mm/tahun. Tanaman cabai kurang tahan terhadap curah hujan yang tinggi, terutama pada saat berbunga, karena bunga akan gugur. Curah hujan yang merata sepanjang tahun berpengaruh baik, karena tidak terjadi kekurangan ataupun kelebihan air selama pertumbuhannya.
Penanaman cabai umumnya dilakukan dengan cara pindah tanaman (transplanting), namun ada juga yang dilakukan dengan cara sebar langsung tanpa penyemaian. Media persemaian yang baik adalah campuran antara pupuk kandang dan tanah dengan perbandingan 1: 1 (Rubatzky & Yamaguchi 1997 ). Pada sistem transplanting, bibit yang berumur 3-4 minggu setelah semai dapat dipindahkan ke lapang (Sumarni 1996). Jarak tanam yang biasa digunakan yaitu 60 – 80 cm untuk jarak antara baris, dan 40 – 50 cm untuk jarak dalam baris. Jarak tanam yang terlalu rapat mengakibatkan tanaman tumbuh terlalu tinggi, sehingga mudah patah oleh angin. Jumlah pupuk yang diberikan tergantung pada kebutuhan tanaman dan ketersediaan hara dalam media tumbuh. Cabai yang ditanam dalam rumah kaca menggunakan polibag membutuhkan jumlah pupuk yang lebih banyak dibandingkan bila ditanam di lapangan. Tanaman cabai di lapangan memerlukan pupuk kurang lebih 170 – 220 kg N/ha, dan 120 kg P2O5/ha
serta kalium ditambahkan apabila ketersediaannya kurang (Rubatzky & Yamaguchi 1997). Pada budidaya cabai di lapangan menggunakan mulsa polyethylene dan irigasi tetes membutuhkan pupuk untuk memperoleh hasil yang optimum sebanyak 200 kg N/ha, 100 kg P2O5/ha dan 150 kg K2O/ha (Alviana
PENAPISAN GENOTIPE CABAI (
Capsicum annuum
L.)
TOLERAN ALUMINIUM BERDASARKAN PERBEDAAN
PANJANG AKAR PADA FASE VEGETATIF
Abstrak
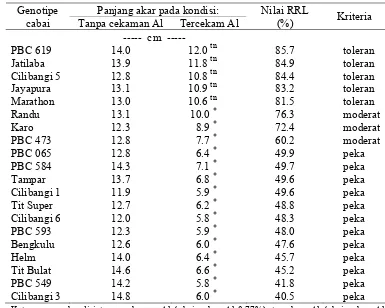
Penelitian ini bertujuan menentukan tingkat kejenuhan Al untuk penapisan genotipe cabai terhadap cekaman Al pada tanah Ultisol, serta mengidentifikasi secara cepat genotipe cabai yang toleran dan peka Al pada fase vegetatif. Penelitian dilaksanakan di Kebun Percobaan Cikabayan IPB dari bulan September 2005 sampai Januari 2006, dengan menggunakan polibag yang bermedia tanah ultisol dari Gajrug, Lebak, Banten. Sebanyak 20 genotipe cabai diuji pada kejenuhan Al: 0.77 , 15.92, 31.96, 60.85 dan 83.48%, yang diperoleh dengan pemberian kapur berturut-turut sebanyak: 18.33, 13.75, 9.16, 4.58 dan 0 g CaCO3/kg tanah. Penentuan toleransi tanaman terhadap Al didasarkan pada nilai panjang akar relatif (relative root length= RRL), dengan membandingkan panjang akar pada kondisi tercekam Al dengan kondisi tanpa cekaman Al. Hasil penelitian menunjukkan bahwa berdasarkan tingkat keragaman yang tinggi, maka tingkat kejenuhan Al 60.85% ditetapkan sebagai kondisi tercekam Al dalam penapisan genotipe cabai untuk toleransi Al pada media tanah Ultisol. Penapisan berdasarkan nilai RRL menghasilkan 5 genotipe yang termasuk kriteria toleran, yaitu PBC 619, Jatilaba, Cilibangi 5, Jayapura dan Marathon; 3 genotipe moderat yaitu Randu, Karo dan PBC 473; serta 12 genotipe peka, yaitu Cilibangi 3, PBC 549, Tit Bulat, Helm, Bengkulu, PBC 065, PBC 593, Tampar, Tit Super, Cilibangi 6, PBC 584 dan Cilibangi 1.
Kata kunci : penapisan, cekaman aluminium, pertumbuhan akar, cabai, Ultisol
Abstract
The aims of the research were to determine index of Al saturation causing Al stress in chili and to identify chili genotypes tolerant to Al using root bioassay. The research was carried out in University Farm of IPB in Cikabayan from September 2005 to January 2006. The media was Ultisol from Gajrug (Lebak, Banten) in polybags. Twenty genotypes of chili were tested at different level of Al saturation (0.77, 15.92, 31.96, 60.85 and 83.48%) by adding lime also in different treatments (18.33, 13.75, 9.16, 4.58 and 0 g CaCO3/kg soil, respectively). The tolerance to aluminum was determined by relative root length (RRL) value,
i.e. the root length at Al-stress condition were compared to that without Al-stress condition. Screening results showed that 5 genotypes were potentially tolerant to Al, namely PBC 619, Jatilaba, Cilibangi 5, Jayapura dan Marathon; 3 genotypes were moderate (Randu, Karo and PBC 473); and 12 genotypes were sensitive to Al (Cilibangi 3, PBC 549, Tit Bulat, Helm, Bengkulu, PBC 065, PBC 593, Tampar, Tit Super, Cilibangi 6, PBC 584 and Cilibangi 1).
Pendahuluan
Tanah Ultisol yang tersebar luas di Kalimantan, Sumatra, Papua, Sulawesi
dan Maluku sangat potensial dikembangkan untuk pertanaman cabai. Pada tanah
tersebut terdapat beberapa kendala yang sering ditemukan secara serempak dan
saling berinteraksi dalam mempengaruhi pertumbuhan tanaman. Adanya kelarutan
aluminium (Al) yang tinggi pada tanah ultisol merupakan kendala utama yang
dapat membatasi pertumbuhan tanaman (Idris 1995 ; Marschner 1995). Pengaruh
aluminium terhadap tanaman dapat terlihat jelas dengan terhambatnya
pertumbuhan akar. Tanaman yang keracunan aluminium mempunyai akar yang
pendek dan terlihat gemuk karena pertumbuhan dan perpanjangan akar primer
maupun lateral terhambat. Pertumbuhan akar yang demikian sulit melakukan
penetrasi ke lapisan subsoil sehingga penyerapan hara dan air menjadi lebih
rendah (Marschner 1995; Ma et al 2001).
Upaya untuk mengurangi kendala yang terdapat pada tanah ultisol dapat
dilakukan dengan pemberian kapur. Pengapuran dapat memperbaiki kondisi
tanah sehingga pertumbuhan tanaman menjadi lebih baik. Jumlah kapur yang
diberikan sebaiknya didasarkan pada daya netralisir untuk meniadakan pengaruh
racun dari aluminium. Pengapuran dapat menekan kelarutan Al, Fe, dan Mn serta
menaikkan pH tanah dan ketersediaan unsur seperti P, Ca dan Mo (Idris 1995).
Namun demikian, pendekatan ini kurang ekonomis karena dalam aplikasinya
dibutuhkan kapur dalam jumlah yang banyak, sehingga pada daerah-daerah
dengan sarana transportasi terbatas dan jauh dari industri kapur akan terkendala
dengan tingginya biaya pengadaan. Pada penelitian ini kapur digunakan untuk
memperoleh tingkat kejenuhan Al yang berbeda pada tanah ultisol.
Alternatif lain agar tanah ultisol dapat dimanfaatkan untuk lahan pertanian
adalah penggunaan tanaman yang toleran terhadap cekaman aluminium.
Toleransi tanaman terhadap cekaman aluminium merupakan faktor penting untuk
adaptasi pada tanah ultisol. Keragaman genotipe cabai di Indonesia cukup tinggi,
namun selama ini baru dimanfaatkan untuk perbaikan daya adaptasi terhadap
cekaman biotik seperti penyakit antraknosa dan virus mosaik, sedangkan untuk
21
banyak dilakukan. Oleh karena itu, seleksi terhadap genotipe yang tersedia perlu
dilakukan untuk memperoleh genotipe yang toleran.
Identifikasi perbedaan karakter pertumbuhan akar merupakan salah satu
peubah yang dapat digunakan dalam seleksi toleransi terhadap cekaman Al,
karena akar merupakan target utama kerusakan oleh Al. Pada beberapa tanaman,
metode cepat untuk menapis genotipe yang toleran terhadap cekaman Al dapat
dilakukan dengan mengamati perbedaan panjang akar pada fase vegetatif (Sasaki
et al.1994; Samuel et al. 1997; Hanum 2004; Bakhtiar et al. 2007).
Penelitian ini bertujuan untuk menentukan tingkat kejenuhan Al untuk
penapisan genotipe cabai terhadap cekaman Al pada tanah ultisol, serta
mengidentifikasi secara cepat genotipe cabai yang toleran dan peka Al melalui
pengamatan perbedaan karakter panjang akar pada fase vegetatif .
Bahan dan Metode Tempat dan Waktu
Penelitian dilaksanakan di kebun percobaan Cikabayan, University Farm,
Institut Pertanian Bogor (IPB), sedangkan analisis tanah dilakukan di
Laboratorium Tanah Balai Besar Sumber Daya Lahan Bogor. Waktu pelaksanaan
penelitian dari bulan September 2005 sampai Januari 2006.
Bahan Penelitian
Bahan tanaman cabai yang diuji terdiri atas 20 genotipe berasal dari
koleksi laboratorium Genetika dan Pemuliaan Tanaman Departemen Agronomi
dan Hortikultura, Faperta IPB, yaitu PBC 065, PBC 473, PBC 549, PBC 584,
PBC 593, PBC 619, Cilibangi 1, Cilibangi 3, Cilibangi 5, Cilibangi 6, Marathon,
Jatilaba, Tit Super, Tit Bulat, Karo, Tampar, Helm, Bengkulu, Randu, dan
Jayapura. Beberapa karakteristik dari genotipe tersebut dapat dilihat pada Tabel
Lampiran 1.
Bahan lainnya adalah tanah ultisol dari Gajrug, Kabupaten Lebak, Banten,
kapur CaCO3, pupuk Urea, SP36, KCl, dan. Karakteristik sifat tanah Ultisol
Gajrug mempunyai pH 4.2, kandungan Al-dd 30.08 me/ 100 g tanah dengan
Metode Penelitian
Penelitian ini merupakan penelitian faktorial dengan 2 faktor perlakuan,
dan disusun menggunakan rancangan acak lengkap (RAL) 3 ulangan. Faktor
pertama adalah 20 genotipe cabai, yaitu : PBC 065, PBC 473, PBC 549, PBC 584,
PBC 593, PBC 619, Cilibangi 1, Cilibangi 3, Cilibangi 5, Cilibangi 6, Marathon,
Jatilaba, Tit Super, Tit Bulat, Karo, Tampar, Helm, Bengkulu, Randu, dan
Jayapura. Faktor kedua adalah tingkat kejenuhan Al, yaitu : 0.77, 15.92, 31.96,
60.85 dan 83.48%. Tingkat kejenuhan Al tersebut diperoleh dengan
menambahkan kapur CaCO3 masing-masing sebanyak 18.33, 13.75, 9.16, 4.58
dan 0 g kapur CaCO3/kg tanah (Tabel 3). Penelitian ini terdiri atas 20 x 5 x 3 =
300 satuan percobaan, dimana setiap satuan percobaan terdiri atas 2 polibag.
Model linier rancangan yang digunakan adalah :
Yijk = µ + αi + βj + (αβ)ij + εijk
Dimana,
Yijk = hasil pengamatan dari perlakuan genotipe cabai ke i dan tingkat
kejenuhan Al ke j pada ulangan ke k
µ = nilai rataan umum
αi = pengaruh perlakuan genotipe cabai ke i
βj = pengaruh perlakuan tingkat kejenuhan Al ke j
(αβ)ij = pengaruh interaksi antara genotipe cabai ke i dan tingkat kejenuhan
Al ke j
εijk = pengaruh galat percobaan dari perlakuan genotipe cabai ke i dan
tingkat kejenuhan Al ke j pada ulangan ke k i = 1, 2, ..., 20
j = 1, 2, ..., 5
k = 1, 2, 3
Data yang diperoleh diuji secara statistik dengan analisis ragam untuk
mengetahui pengaruh perlakuan yang diberikan. Untuk mengetahui perbedaan
panjang akar pada kondisi tercekam Al dengan tanpa cekaman Al dilakukan uji t
pada taraf 5%. Pengelompokan tingkat toleransi terhadap cekaman Al didasarkan
pada nilai panjang akar relatif (relative root length = RRL) dengan mengacu pada kriteria yang telah dimodifikasi dari Matsumoto et al. (1996), yaitu :
Toleran : jika nilai RRL > 50% dan panjang akar pada kondisi tercekam Al
tidak berbeda nyata dengan kondisi tanpa cekaman Al.
Moderat : jika nilai RRL > 50% dan panjang akar pada kondisi tercekam Al
23
Peka : jika nilai RRL ≤ 50% dan panjang akar pada kondisi tercekam Al
berbeda nyata dengan kondisi tanpa cekaman Al.
Nilai panjang akar relatif (RRL) diperoleh dari persamaan:
Pelaksanaan Penelitian
Penentuan tingkat kejenuhan Al. Tingkat kejenuhan Al yang berbeda pada tanah ultisol dapat diprediksi melalui percobaan pendahuluan tentang
pemberian kapur CaCO3 dengan takaran 0, 5, 10, 15, dan 20 g kapur CaCO3/kg
tanah. Tanah ultisol yang telah diayak, ditimbang sebanyak 3 kg bobot kering
udara dan dicampur secara merata dengan kapur, kemudian dimasukkan ke dalam
polibag untuk diinkubasi selama 1 bulan (Idris 1995). Setelah diinkubasi
dilakukan analisis tanah terhadap unsur K, Ca, Mg, Na, Al, H dan kejenuhan Al.
Kebutuhan kapur untuk menurunkan kejenuhan Al yang diinginkan ditentukan
dengan analisis regresi.
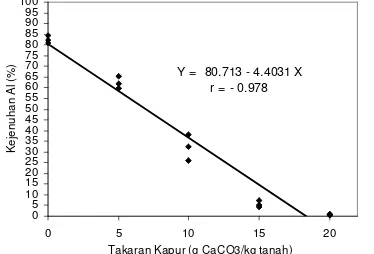
Hasil analisis regresi yang menggambarkan hubungan antara pemberian
kapur (X) dan kejenuhan Al (Y) diperoleh persamaan Y = 80.713 – 4.4031 X, r =
- 0.978 (Gambar 4).
Y = 80.713 - 4.4031 X r = - 0.978
0 5 10 15 20 25 30 35 40 45 50 55 60 65 70 75 80 85 90 95 100
0 5 10 15 20
Takaran Kapur (g CaCO3/kg tanah)
[image:40.612.205.392.471.598.2]K ej enuhan A l ( % )
Gambar 4. Grafik penurunan nilai kejenuhan Al tanah ultisol akibat pemberian kapur CaCO3.
Berdasarkan persamaan regresi di atas, maka diperoleh kejenuhan Al
tanah ultisol masing-masing sebesar 0.77, 15.92, 31.96, 60.85 dan 83.48%
dengan pemberian kapur seperti yang tercantum pada Tabel 2. panjang akar pada kondisi tercekam Al
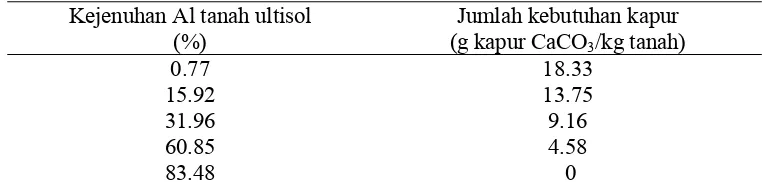
Tabel 2. Jumlah kebutuhan kapur CaCO3 untuk memperoleh kejenuhan
aluminium tanah ultisol yang berbeda
Kejenuhan Al tanah ultisol (%)
Jumlah kebutuhan kapur (g kapur CaCO3/kg tanah)
0.77 18.33 15.92 13.75 31.96 9.16 60.85 4.58
83.48 0
Pemberian kapur sebanyak 18.33, 13.75, 9.16, 4.58 dan 0 g CaCO3/kg
tanah kemudian digunakan sebagai standar untuk memperoleh tingkat kejenuhan
Al seperti pada perlakuan, yaitu kejenuhan Al 0.77, 15.92, 31.96, 60.85 dan
83.48% (analisis tanah dapat dilihat pada Tabel 3).
Persiapan media tanam. Tanah dari lapangan dibersihkan dari sisa-sisa tanaman dan kotoran kemudian digemburkan dan dikeringanginkan selama satu
minggu. Tanah yang telah kering diayak dengan ayakan yang berukuran 2 mm,
kemudian ditimbang sebanyak 3 kg bobot kering udara dan dicampur secara
merata dengan kapur sesuai dengan standar tersebut. Campuran tanah dan kapur
dimasukkan ke dalam polibag, kemudian diinkubasi selama 1 bulan (Idris 1995).
Pembibitan dan penanaman. Pembibitan dilakukan di tray persemaian yang mempunyai 72 lubang yang berisi media campuran tanah dan pupuk kascing
(organik) dengan perbandingan 1:1. Bibit yang telah berumur 4 minggu setelah
semai, kemudian dipindahkan ke dalam polibag sebanyak 1 bibit per polibag.
Pemeliharan. Pemeliharaan meliputi pemupukan, penyiraman, penyiangan, serta pengendalian hama dan penyakit. Pupuk diberikan dengan
dosis 250 kg N/ha, 150 kg P2O5/ha dan 200 kg K2O/ha atau setara dengan 0.82 g
urea/polibag, 0.63 g SP36/polibag dan 0.50 g KCl/polibag. Aplikasi pupuk
dilakukan sebanyak dua kali dengan cara disebar dalam alur melingkar,
pemupukan pertama dilakukan sehari sebelum bibit ditanam dengan nisbah N
(1/2)+P(1)+K(1/2), sedangkan pemupukan kedua dilakukan 3 minggu setelah
bibit ditanam dengan nisbah N (1/2)+P(0)+K(1/2). Penyiraman dilakukan setiap
hari jam 08.00 sesuai kapasitas lapangan. Kebutuhan air untuk mencapai
kapasitas lapangan ditentukan dari kadar air tanah kapasitas lapangan (35.25%)
25
570.5 ml atau disetarakan menjadi 570 ml/polibag. Untuk melindungi tanaman
cabai dari serangan hama dan penyakit dilakukan penyemprotan pestisida jika
diperlukan. Insektisida yang digunakan berbahan aktif Deltamethrin dengan
konsentrasi 50 mg/l air dan fungisida berbahan aktif Mankozeb dengan
konsentrasi 1.6 g/l air (Haryantini & Santoso 2001).
Pengamatan
Media tanah Ultisol yang digunakan dalam penelitian dianalisis untuk
mengetahui karasteristik sifat tanahnya pada kondisi sebelum maupun sesudah
inkubasi kapur. Analisis tanah dilakukan di Laboratorium Penelitian Kimia dan
Uji Tanah, Balai Besar Sumber Daya Lahan, Bogor. Hasil analisis dapat dilihat
pada Tabel 3.
Pengamatan karakter panjang akar, bobot kering akar dan bobot kering
tajuk dilakukan pada umur 6 minggu setelah bibit ditanam, yaitu :
1. Pengamatan terhadap panjang akar dilakukan dengan cara mencabut tanaman
secara perlahan dengan menyiram air terlebih dahulu agar mudah
mencabutnya dan akar tidak putus. Akar dibersihkan dari tanah menggunakan
air mengalir kemudian akar diukur dari pangkal hingga ujung akar.
2. Pengamatan terhadap bobot kering akar dilakukan dengan cara mencabut
tanaman dan akarnya dibersihkan dari tanah, kemudian dipotong pada bagian
pangkal akar. Bagian akar dikeringanginkan terlebih dahulu, kemudian
dimasukkan dalam oven pada suhu 70oC selama 2-4 hari.
3. Pengamatan terhadap bobot kering tajuk dilakukan dengan cara memotong
bagian pangkal akar tanaman. Bagian atas tanaman dikeringanginkan terlebih
dahulu, kemudian dimasukkan dalam oven pada suhu 70oC selama 2-4 hari.
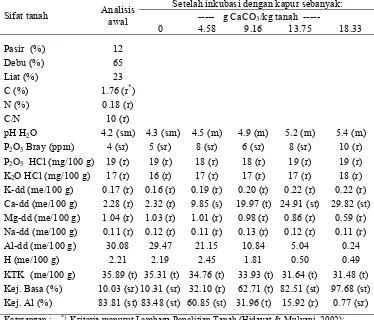
Hasil dan Pembahasan Karakteristik Sifat Tanah
Hasil analisis tanah sebelum pengapuran (Tabel 3) menunjukkan bahwa
tanah ultisol asal Gajrug mempunyai reaksi tanah yang sangat masam (pH 4.2),
kandungan bahan organik rendah dengan nisbah C/N tergolong rendah.
Kandungan P potensialnya rendah, P tersedia sangat rendah dan K potensial
rendah, namun KTK termasuk tinggi. Kejenuhan Al pada tanah tersebut
tergolong sangat tinggi (83.81%) dengan kandungan Al dapat dipertukarkan
sebanyak 30.08 me/100 g. Kondisi tanah dengan pH kurang dari 5 dan kejenuhan
[image:43.612.129.503.198.518.2]Al yang tinggi merupakan ciri tanah yang tercekam Al (Hidayat & Mulyani 2002).
Tabel 3. Karasteristik sifat tanah ultisol asal Gajrug, Lebak Banten sebelum dan sesudah pemberian kapur
Setelah inkubasi dengan kapur sebanyak: --- g CaCO3/kg tanah --- Sifat tanah Analisis
awal
0 4.58 9.16 13.75 18.33
Pasir (%) 12
Debu (%) 65
Liat (%) 23
C (%) 1.76 (r*)
N (%) 0.18 (r)
C/N 10 (r)
pH H2O 4.2 (sm) 4.3 (sm) 4.5 (m) 4.9 (m) 5.2 (m) 5.4 (m) P2O5 Bray (ppm) 4 (sr) 5 (sr) 8 (sr) 6 (sr) 8 (sr) 10 (r) P2O5 HCl (mg/100 g) 19 (r) 19 (r) 18 (r) 18 (r) 19 (r) 19 (r) K2O HCl (mg/100 g) 17 (r) 16 (r) 17 (r) 17 (r) 17 (r) 18 (r) K-dd (me/100 g) 0.17 (r) 0.16 (r) 0.19 (r) 0.20 (r) 0.22 (r) 0.22 (r) Ca-dd (me/100 g) 2.28 (r) 2.32 (r) 9.85 (s) 19.97 (t) 24.91 (st) 29.82 (st) Mg-dd (me/100 g) 1.04 (r) 1.03 (r) 1.01 (r) 0.98 (r) 0.86 (r) 0.59 (r) Na-dd (me/100 g) 0.11 (r) 0.12 (r) 0.11 (r) 0.13 (r) 0.12 (r) 0.11 (r) Al-dd (me/100 g) 30.08 29.47 21.15 10.84 5.04 0.24 H (me/100 g) 2.21 2.19 2.45 1.81 0.50 0.49 KTK (me/100 g) 35.89 (t) 35.31 (t) 34.76 (t) 33.93 (t) 31.64 (t) 31.48 (t) Kej. Basa (%) 10.03 (sr) 10.31 (sr) 32.10 (r) 62.71 (t) 82.51 (st) 97.68 (st) Kej. Al (%) 83.81 (st) 83.48 (st) 60.85 (st) 31.96 (t) 15.92 (r) 0.77 (sr)
Keterangan : *) Kriteria menurut Lembaga Penelitian Tanah (Hidayat & Mulyani 2002); sr=sangat rendah; r=rendah; s=sedang; t=tinggi; st= sangat tinggi; sm=sangat masam; m=masam
Pemberian kapur menurunkan kejenuhan Al dan kandungan aluminium,
serta menaikkan kejenuhan basa, kandungan kalsium, pH tanah, dan P tersedia
(Tabel 3). Berkaitan dengan cekaman Al, maka pemberian kapur sebanyak 18.33,
13.75, 9.16, 4.58 dan 0 g CaCO3/kg tanah terlihat dapat menghasilkan tingkat
kejenuhan Al yang berbeda, yaitu masing-masing sebesar 0.77, 15.92, 31.96,
60.85 dan 83.48%. Kejenuhan Al tersebut sesuai untuk lingkungan penapisan
genotipe toleran Al yang menggunakan media tanah ultisol dengan tingkat
27
Penentuan Tingkat Kejenuhan Al untuk Penapisan
Kondisi tanah percobaan sesuai digunakan untuk lingkungan penapisan
genotipe toleran Al karena tingkat kejenuhan Al yang dibuat melalui pemberian
kapur terlihat berpengaruh sangat nyata terhadap karakter yang diamati (Tabel 4).
Hasil analisis ragam menunjukkan adanya interaksi antara genotipe dan tingkat
kejenuhan Al sehingga penapisan genotipe toleran Al harus dilakukan pada
lingkungan target sesuai dengan tingkat kejenuhan Al. Selain itu, perbedaan
genotipe cabai menunjukkan perbedaan karakter yang diamati. Hal ini
mengindikasikan adanya keragaman antara 20 genotipe yang diuji sehingga dapat
[image:44.612.125.514.330.401.2]digunakan sebagai bahan tanaman untuk percobaan penapisan.
Tabel 4. Nilai kuadrat tengah dari hasil analisis ragam pengaruh genotipe cabai dan tingkat kejenuhan Al pada panjang akar, bobot kering akar dan bobot kering tajuk
Nilai kuadrat tengah pada pengamatan: Sumber keragaman
Panjang akar Bobot kering akar Bobot kering tajuk
Genotipe cabai (G) 22.691 ** 0.189 ** 2.753 **
Tingkat kejenuhan Al (A) 431.844 ** 1.831 ** 26.504 **
Interaksi G x A 3.437 ** 0.016 * 0.154 tn
Keterangan : * = berpengaruh nyata; ** = berpengaruh sangat nyata; tn = tidak nyata