PENGARUH SALINITAS TERHADAP PERTUMBUHAN DAN
BIOMASSA SEMAI NON-SEKRESI
Ceriops tagal
DAN
KANDUNGAN LIPID PADA TINGKAT POHON
RAMAYANI 081201030
PROGRAM STUDI KEHUTANAN
FAKULTAS PERTANIAN
PENGARUH SALINITAS TERHADAP PERTUMBUHAN DAN
BIOMASSA SEMAI NON-SEKRESI
Ceriops tagal
DAN
KANDUNGAN LIPID PADA TINGKAT POHON
SKRIPSI
Oleh: RAMAYANI
081201030
PROGRAM STUDI KEHUTANAN
FAKULTAS PERTANIAN
PENGARUH SALINITAS TERHADAP PERTUMBUHAN DAN
BIOMASSA SEMAI NON-SEKRESI
Ceriops tagal
DAN
KANDUNGAN LIPID PADA TINGKAT POHON
SKRIPSI
Oleh: RAMAYANI
081201030/BUDIDAYA HUTAN
Skripsi sebagai salah satu syarat untuk memperoleh gelar sarjana di Fakultas Pertanian
Universitas Sumatera Utara
PROGRAM STUDI KEHUTANAN
FAKULTAS PERTANIAN
Judul : Pengaruh Salinitas Terhadap Pertumbuhan Dan Biomassa Semai Non-Sekresi Ceriops tagal Dan Kandungan Lipid Pada Tingkat Pohon
Nama : Ramayani
NIM : 081201030
Program Studi : Budidaya Hutan
Disetujui oleh Komisi Pembimbing
Mohammad Basyuni S.Hut, M.Si, Ph.D Dr.Ir. Lollie Agustina P. Putri M.Si
Ketua Anggota
Mengetahui,
Siti Latifah, S.Hut, M.Si, Ph.D Ketua Departemen Kehutanan
ABSTRAK
RAMAYANI : Pengaruh Salinitas Terhadap Pertumbuhan Dan Biomassa Semai Non-Sekresi Ceriops tagal Dan Kandungan Lipid Pada Tingkat Pohon, dibimbing oleh MOHAMMAD BASYUNI dan LOLLIE AGUSTINA P. PUTRI.
Pengaruh salinitas terhadap pertumbuhan dan biomassa semai non-sekresi C. tagal dan kandungan lipid pada tingkat pohon. Penelitian ini dilakukan di rumah kaca, Fakultas Pertanian dan laboratorium penelitian Farmasi, Fakultas Farmasi, Universitas Sumatera Utara dari bulan Juli 2011 sampai Juli 2012. Tujuannya adalah untuk mengetahui tingkat pertumbuhan dan biomassa semai C. tagal pada berbagai tingkat salinitas, mengetahui kandungan lipid pada daun dan akar C. tagal pada tingkat pohon. Penelitian menggunakan 5 perlakuan, yaitu salinitas 0%, 0.5%, 1.5%, 2% dan 3%, dengan masing-masing 5 ulangan. Ekstraksi dan analisis lipid dilakukan dengan mengekstraksi daun dan akar dari pohon dewasa yang diambil dari hutan mangrove Pulau Sembilan.
Hasil penelitian menunjukkan semai C. tagal dengan berbagai tingkat salinitas memberikan pengaruh nyata terhadap tinggi, diameter, dan biomassa semai. Pertumbuhan semai C. tagal yang paling baik terdapat pada salinitas 0.5% dengan pertumbuhan tinggi dan diameter semai yaitu 3,24 cm dan 3,65 mm. Berat basah dan berat kering daun, akar dan batang semai C. tagal yang paling tinggi pada salinitas 1.5%. Total lipid di daun dan akar pohon C.tagal adalah sebesar 16,2 mg dan 8,9 mg.
ABSTRACT
RAMAYANI: Effect of salinity on growth and biomass of Semai Non-secretion of Ceriops tagal And Lipid Content in the tree, guided by MOHAMMAD BASYUNI and LOLLIE AGUSTINA P. PUTRI .
Effect of salinity on seedling growth and biomass of non-secretion of C. tagal and lipid content in the tree. The research was conducted in the greenhouse, Faculty of Agriculture and Pharmaceutical research laboratories, Faculty of Pharmacy, University of North Sumatra from July 2011 to July 2012. The goal is to determine the level of growth and seedling biomass C. tagal at different levels of salinity, knowing the lipid content in leaves and roots of C. tagal at tree level. Studies using the 5 treatments, namely salinity 0%, 0.5%, 1.5%, 2% and 3%, with five replicates each. Lipid extraction and analysis performed by extracting the leaves and roots of mature trees taken from the mangrove forest Pulau Sembilan.
The results showed seedling C. tagal with different levels of salinity give real effect to height, diameter, and seedling biomass. Seedling growth of C. tagal best there is at 0.5% salinity with high growth and seedling diameter is 3.24 cm and 3.65 mm. Wet weight and dry weight of leaves, roots and stems of seedlings with C. tagal the highest at 1.5% salinity. Total lipids in leaves and roots of trees are C.tagal of 16.2 mg and 8.9 mg.
RIWAYAT HIDUP
Penulis dilahirkan di Medan pada tanggal 10 Mei 1990 dari pasangan
Bapak Alm. Haryanto dan Ibu Rodiah. Penulis merupakan putri ke satu dari empat
bersaudara.
Lulus dari Sekolah Dasar (SD) Negeri 080929 Medan pada tahun 2002,
pada tahun 2005 lulus dari SMP Negeri 28 Medan, dan pada tahun 2008 lulus dari
SMA Negeri 2 Medan. Pada tahun yang sama penulis di terima menjadi
mahasiswa Fakultas Pertanian Universitas Sumatera Utara (USU) melalui jalur
Ujian Masuk Bersama. Penulis memilih jurusan Budidaya Hutan, Program Studi
Kehutanan.
Selama mengikuti kuliah, penulis pernah menjadi Asisten Praktikum Hasil
Hutan Non Kayu tahun 2010, Asisten Praktikum Klimatologi Hutan tahun 2011,
Asisten Praktikum Silvika tahun 2011 dan 2012, Asisten Inventarisasi Hutan
tahun 2011, Asisten Praktik Pengenalan Ekosistem Hutan (P2EH) tahun 2011.
Penulis melakukan Praktik Pengenalan Ekosistem Hutan (P2EH) di Hutan
Dataran Tinggi Gunung Sinabung dan TWA Deleng Lancuk Kabupaten Karo
tahun 2010. Penulis melaksanakan Praktik Kerja Lapang (PKL) di HTI PT.
KATA PENGANTAR
Puji dan syukur penulis panjatkan kehadirat Allah SWT, atas segala
rahmat dan karunia-Nya sehingga penulis dapat menyelesaikan skripsi yang
berjudul “Pengaruh Salinitas Terhadap Pertumbuhan Dan Biomassa Semai
Non-Sekresi Ceriops tagal Dan Kandungan Lipid Pada Tingkat Pohon”.
Pada kesempatan ini penulis mengucapkan terima kasih kepada orangtua
penulis yang telah membimbing, mendidik dan memberikan semangat serta
mendukung penulis untuk doa dan materil. Penulis juga mengucapkan terima
kasih kepada ketua komisi pembimbing Mohammad Basyuni, S.Hut, M.Si, Ph.D
dan anggota komisi pembimbing Dr.Ir. Lollie Agustina P. Putri M.Si
yang terus membimbing dan mengarahkan penulis dalam penyelesaian skripsi ini.
Penulis menyadari bahwa skripsi ini masih jauh dari kesempurnaan, baik
dari segi materi maupun teknik penulisan. Oleh sebab itu, penulis sangat
mengharapkan saran dan kritik yang membangun dari para pembaca demi
penyempurnaan skripsi ini.
Akhirnya penulis berharap skripsi ini dapat bermanfaat bagi para
DAFTAR ISI
Hutan Mangrove dan Karakteristiknya ... 4Penyebaran Hutan Mangrove ... 5
Zonasi di Hutan Mangrove ... 5
Taksonomi dan Morfologi Ceriops tagal ... 6
Faktor-Faktor yang Mempengaruhi Pertumbuhan Ceriops tagal ... 7
Biologi Terpenoid pada Mangrove ... 11
Potensi Triterpenoid pada Mangrove ... 13
Mekanisme Toleransi Tanaman Terhadap Cekaman Garam ... 13
METODE PENELITIAN Waktu dan Lokasi Penelitian ... 16
Penanaman ... 16
Pengambilan Sampel dan Ekstraksi ... 16
Gambaran Lokasi Pengambilan Sampel ... 17
Bahan dan Alat Penelitian ... 18
Penanaman ... 18
Ekstraksi dan Analisis ... 18
Analisis Data ... 18
Prosedur Penelitian ... 19
Penyiapan Media Tanam... 19
Berat Basah Akar (g) ... 22
Berat Basah Tajuk (g) ... 22
Berat Basah Batang (g) ... 22
Berat Kering Akar (g) ... 22
Berat Kering Tajuk (g) ... 23
Berat Kering Batang (g) ... 23
Ratio Tajuk dan Akar ... 23
Ratio Batang dan Akar ... 23
HASIL DAN PEMBAHASAN Pengaruh Salinitas Terhadap Pertumbuhan Semai ... 24
Pengaruh Salinitas Terhadap Biomassa Semai ... 27
Ratio Tajuk/Akar dan Ratio Batang/Akar Semai ... 31
Total Lipid dan Kandungan NSL (Nonsaponifiable Lipid) ... 32
KESIMPULAN DAN SARAN Kesimpulan ... 35
Saran ... 35
DAFTAR TABEL
No. Halaman
DAFTAR GAMBAR
No. Halaman
1. Lokasi Pengambilan Sampel Penelitian ... 17
2. Pengaruh salinitas terhadap pertumbuhan Tinggi Semai (A) dan Diamter Semai (B). Data merupakan rata-rata ± SE (n = 13-15). Tanda bintang (*) dan (**) mengindikasikan secara statistik signifikan dari kontrol (0%) pada P< 0.05 dan P< 0.01 dengan uji
Dunnet... 24
3. Rata-rata Jumlah Daun (A) dan Luas Daun Ceriops tagal (B). Data merupakan rata-rata ± SE (n = 13-15) untuk rata-rata jumlah daun dan SE (n= 5) untuk luas daun. Tanda bintang (*) dan (**) mengindikasikan secara statistik signifikan dari kontrol (0%) pada
P< 0.05 dan P< 0.01 dengan uji Dunnet... 26
4. Pengaruh Salinitas terhadap Berat Basah (A) dan Berat Kering (B). Data merupakan rata-rata ± SE (n=5). Tanda bintang (*) dan (**) mengindikasikan secara statistik signifikan dari kontrol (0%) pada
P< 0.05 dan P< 0.01 dengan uji Dunnet... 27
5. Contoh Zonasi Mangrove di Pulau Sembilan Kabupaten Langkat
Sumatera Utara ... 29
6. Rasio Tajuk dan Akar (A), Rasio Batang dan Akar (B). Data merupakan rata-rata ± SE (n=5). Tanda bintang (*) dan (**) mengindikasikan secara statistik signifikan dari kontrol (0%) pada
ABSTRAK
RAMAYANI : Pengaruh Salinitas Terhadap Pertumbuhan Dan Biomassa Semai Non-Sekresi Ceriops tagal Dan Kandungan Lipid Pada Tingkat Pohon, dibimbing oleh MOHAMMAD BASYUNI dan LOLLIE AGUSTINA P. PUTRI.
Pengaruh salinitas terhadap pertumbuhan dan biomassa semai non-sekresi C. tagal dan kandungan lipid pada tingkat pohon. Penelitian ini dilakukan di rumah kaca, Fakultas Pertanian dan laboratorium penelitian Farmasi, Fakultas Farmasi, Universitas Sumatera Utara dari bulan Juli 2011 sampai Juli 2012. Tujuannya adalah untuk mengetahui tingkat pertumbuhan dan biomassa semai C. tagal pada berbagai tingkat salinitas, mengetahui kandungan lipid pada daun dan akar C. tagal pada tingkat pohon. Penelitian menggunakan 5 perlakuan, yaitu salinitas 0%, 0.5%, 1.5%, 2% dan 3%, dengan masing-masing 5 ulangan. Ekstraksi dan analisis lipid dilakukan dengan mengekstraksi daun dan akar dari pohon dewasa yang diambil dari hutan mangrove Pulau Sembilan.
Hasil penelitian menunjukkan semai C. tagal dengan berbagai tingkat salinitas memberikan pengaruh nyata terhadap tinggi, diameter, dan biomassa semai. Pertumbuhan semai C. tagal yang paling baik terdapat pada salinitas 0.5% dengan pertumbuhan tinggi dan diameter semai yaitu 3,24 cm dan 3,65 mm. Berat basah dan berat kering daun, akar dan batang semai C. tagal yang paling tinggi pada salinitas 1.5%. Total lipid di daun dan akar pohon C.tagal adalah sebesar 16,2 mg dan 8,9 mg.
ABSTRACT
RAMAYANI: Effect of salinity on growth and biomass of Semai Non-secretion of Ceriops tagal And Lipid Content in the tree, guided by MOHAMMAD BASYUNI and LOLLIE AGUSTINA P. PUTRI .
Effect of salinity on seedling growth and biomass of non-secretion of C. tagal and lipid content in the tree. The research was conducted in the greenhouse, Faculty of Agriculture and Pharmaceutical research laboratories, Faculty of Pharmacy, University of North Sumatra from July 2011 to July 2012. The goal is to determine the level of growth and seedling biomass C. tagal at different levels of salinity, knowing the lipid content in leaves and roots of C. tagal at tree level. Studies using the 5 treatments, namely salinity 0%, 0.5%, 1.5%, 2% and 3%, with five replicates each. Lipid extraction and analysis performed by extracting the leaves and roots of mature trees taken from the mangrove forest Pulau Sembilan.
The results showed seedling C. tagal with different levels of salinity give real effect to height, diameter, and seedling biomass. Seedling growth of C. tagal best there is at 0.5% salinity with high growth and seedling diameter is 3.24 cm and 3.65 mm. Wet weight and dry weight of leaves, roots and stems of seedlings with C. tagal the highest at 1.5% salinity. Total lipids in leaves and roots of trees are C.tagal of 16.2 mg and 8.9 mg.
PENDAHULUAN
Latar Belakang
Mangrove merupakan salah satu ekosistem yang paling produktif di bumi
dibandingkan dengan ekosistem lainya (Clough et al., 2000). Pentingnya hutan
mangrove telah diakui bagi ekosistem global, namun terdapat sedikit informasi
yang menjelaskan mengapa tanaman mangrove dapat tumbuh di lingkungan
salinitas yang tinggi, terutama yang berasal dari mangrove Indonesia. Menurut
karakteristik morfologinya dalam manajemen garam, tanaman mangrove dibagi ke
dalam dua kelompok besar (Scholander et al., 1962). Kelompok pertama adalah
spesies yang mensekresi garam (jenis sekresi/secreting species) yang memiliki
kelenjar garam di daunnya atau rambut garam untuk menghilangkan kelebihan
garam. Yang kedua adalah spesies non-sekresi (non-scereting species) yang tidak
memiliki fitur morfologi tersebut untuk ekskresi kelebihan garam (Scholander et
al., 1962; Tomlinson, 1986). Dengan demikian, hutan mangrove merupakan
model tanaman yang ideal untuk mempelajari mekanisme toleransi garam pada
tingkat seluler.
Mangrove terkenal kaya sebagai sumber senyawa triterpenoid dan
fitosterol (isoprenoid) (Koch et al, 2003; Basyuni et al, 2007). Salah satu
kemampuan yang sangat tampak yaitu spesies mangrove dapat tumbuh dalam
Basyuni et al., 2007, 2009). Senyawa-senyawa tersebut berfungsi sebagai pertahanan secara kimia bagi dirinya (Williams, 1999).
Penelitian sebelumnya menunjukkan bahwa triterpenoid memainkan
peran penting untuk melindungi mangrove dari cekaman garam (Oku et al., 2003;
Basyuni et al., 2007, 2009, 2011). Meskipun demikian, sedikit studi yang
difokuskan pada toleransi garam dan komposisi triterpenoid di hutan mangrove
Sumatera Utara. Oleh karena itu penelitian ini dilakukan untuk melihat pengaruh
pemberian berbagai macam konsentrasi garam (salinitas) terhadap pertumbuhan
dan biomassa semai mangrove non-sekresi yaitu Ceriops tagal, serta ekstraksi
dan analisis komposisi lipidnya yang diambil dari Pulau Sembilan, Langkat,
Sumatera Utara.
Tujuan Penelitian
Penelitian ini bertujuan untuk :
1. Untuk mengetahui tingkat pertumbuhan dan biomassa semai C. tagal pada
berbagai tingkat salinitas.
2. Untuk mengetahui gambaran posisi C. tagal dalam zonasi mangrove di
Pulau Sembilan.
3. Untuk mengetahui kandungan lipid pada daun dan akar C. tagal pada
tingkat pohon.
Hipotesis Penelitian
Ada perbedaan tingkat pertumbuhan dan biomassa semai C. tagal pada
Manfaat Penelitian
1. Hasil penelitian ini diharapkan dapat menjadi rekomendasi bagi masyarakat
yang mengusahakan pembibitan C. tagal untuk program rehabilitasi agar dapat
memperoleh bibit C. tagal yang pertumbuhannya lebih baik pada berbagai
tingkat salinitas.
2. Pengetahuan tentang lipid pada jenis C. tagal diharapkan dapat berkontribusi
dalam estimasi dan perhitungan tingkat sedimentasi bahan organik di
TINJAUAN PUSTAKA
Hutan Mangrove dan Karakteristiknya
Mangrove adalah tumbuhan berkayu yang tumbuh diantara daratan dan
lautan di daerah tropis dan subtropis. Tumbuhan mangrove memiliki kemampuan
khusus untuk beradaptasi dengan kondisi lingkungan yang ekstrim, seperti kondisi
tanah yang tergenang, kadar garam yang tinggi serta kondisi tanah yang kurang
stabil. Dengan kondisi lingkungan seperti itu, beberapa jenis mangrove
mengembangkan mekanisme yang memungkinkan secara aktif mengeluarkan
garam dari jaringan lewat kelenjar daun. Sementara jenis yang lainnya
mengembangkan sistem perakaran untuk membantu memperoleh oksigen bagi
sistem perakarannya. Dalam hal yang lain, beberapa jenis mangrove berkembang
dengan buah yang sudah berkecambah sewaktu masih di pohon induknya
(vivipar), seperti Kandelia, Bruguiera, Ceriops dan Rhizophora
(Tomlinson, 1986; Kathiresan and Bingham, 2001).
Selanjutnya Tomlinson (1986) membagi flora mangrove menjadi tiga
kelompok, yakni: (1) Flora mangrove mayor (flora mangrove sejati), yakni flora
yang menunjukkan kesetiaan terhadap habitat mangrove, berkemampuan
membentuk tegakan murni dan secara dominan mencirikan struktur komunitas,
secara morfologi mempunyai bentuk-bentuk adaptif khusus (bentuk akar dan
viviparitas) terhadap lingkungan mangrove, dan mempunyai mekanisme fisiologis
dalam mengontrol garam. Contohnya adalah Avicennia, Rhizophora, Bruguiera,
Ceriops, Kandelia, Sonneratia, Lumnitzera, Laguncularia dan Nypa. (2) Flora
mangrove sejati minor, yakni flora mangrove yang tidak mampu membentuk
komunitas, contohnya Excoecaria, Xylocarpus, Heritiera, Aegiceras. Aegialitis,
Acrostichum, Camptostemon, Scyphiphora, Pemphis, Osbornia dan Pelliciera. (3)
Asosiasi mangrove, contohnya adalah Cerbera, Acanthus, Derris, Hibiscus,
Calamus, dan lain-lain.
Penyebaran Hutan Mangrove
Indonesia merupakan negara yang kaya, Indonesia mempunyai hutan
mangrove yang terluas didunia, sebaran terumbu karang yang eksotik, rumput laut
yang terdapat dihampir sepanjang pantai, sumber perikanan yang tidak ternilai
banyaknya. Hutan mangrove atau yang biasa disebut hutan bakau, walaupun
penyebutan hutan bakau itu kurang tepat karena bakau hanya merupakan salah
satu dari jenis mangrove itu sendiri yaitu jenis Rhizopora spp. Hutan mangrove
merupakan tipe hutan yang khas dan tumbuh disepanjang pantai atau muara
sungai yang dipengaruhi oleh pasang surut air laut. Mangrove banyak dijumpai di
wilayah pesisir yang terlindung dari gempuran ombak dan daerah yang landai di
daerah tropis dan sub tropis (FAO, 2007).
Zonasi di Hutan Mangrove
Dalam hal struktur, mangrove di Indonesia lebih bervariasi bila dibandingkan
dengan daerah lainnya. Dapat ditemukan mulai dari tegakan Avicennia marina dengan ketinggian 1 - 2 meter pada pantai yang tergenang air laut, hingga tegakan
campuran Bruguiera- Rhizophora-Ceriops dengan ketinggian lebih dari 30 meter (misalnya, di Sulawesi Selatan). Di daerah pantai yang terbuka, dapat ditemukan
rendah kecuali mangrove anakan dan beberapa jenis semak seperti Acanthus ilicifolius dan Acrostichum aureum (Noor et al, 1999).
Daya adaptasi atau toleransi jenis tumbuhan mangrove terhadap kondisi
lingkungan yang ada mempengaruhi terjadinya zonasi pada kawasan hutan
mangrove. Permintakatan jenis tumbuhan mangrove dapat dilihat sebagai proses
suksesi dan merupakan hasil reaksi ekosistem dengan kekuatan yang datang dari
luar seperti tipe tanah, salinitas, tingginya ketergenangan air dan pasang surut.
Pembagian zonasi kawasan mangrove yang dipengaruhi adanya perbedaan
penggenangan atau perbedaan salinitas meliputi :
1. Zona garis pantai, yaitu kawasan yang berhadapan langsung dengan laut.
Lebar zona ini sekitar 10-75 meter dari garis pantai dan biasanya ditemukan
jenis Rhizophora stylosa, R. mucronata, Avicennia marina dan Sonneratia
alba.
2. Zona tengah, merupakan kawasan yang terletak di belakang zona garis
pantai dan memiliki lumpur liat. Biasanya ditemukan jenis Rhizophora
apiculata, Avicennia officinalis, Bruguiera cylindrica, B. gymnorrhiza, B.
parviflora, B. sexangula, Ceriops tagal, Aegiceras corniculatum, Sonneratia
caseolaris dan Lumnitzera littorea.
3. Zona belakang, yaitu kawasan yang berbatasan dengan hutan darat. Jenis
tumbuhan yang biasanya muncul antara lain Achantus ebracteatus, A.
ilicifolius, Acrostichum aureum, A. speciosum. Jenis mangrove yang tumbuh
adalah Heritiera littolaris, Xylocarpus granatum, Excoecaria agalocha,
Nypa fruticans, Derris trifolia, Osbornea octodonta dan beberapa jenis
asiatica, Cerbera manghas, Hibiscus tiliaceus, Ipomea pes-caprae,
Melastoma candidum, Pandanus tectorius, Pongamia pinnata, Scaevola
taccada dan Thespesia populnea (Pramudji dan Purnomo, 2003).
Pada umumnya, lebar zona mangrove jarang melebihi 4 kilometer, kecuali
pada beberapa estuari serta teluk yang dangkal dan tertutup. Pada daerah seperti
ini lebar zona mangrove dapat mencapai 18 kilometer seperti di Sungai
Sembilang, Sumatera Selatan (Danielsen and Verheugt, 1990) atau bahkan lebih
dari 30 kilometer seperti di Teluk Bintuni, Irian Jaya. Adapun pada daerah pantai
yang tererosi dan curam, lebar zona mangrove jarang melebihi 50 meter. Untuk
daerah di sepanjang sungai yang dipengaruhi oleh pasang surut, panjang
hamparan mangrove kadang-kadang mencapai puluhan kilometer seperti di
Sungai Barito, Kalimantan Selatan. Panjang hamparan ini bergantung pada intrusi
air laut yang sangat dipengaruhi oleh tinggi rendahnya pasang surut, pemasukan
dan pengeluaran material ke dalam dan dari sungai, serta kecuramannya
(Erftemeijer et al., 1989).
Taksonomi dan Morfologi Ceriops tagal
Tengar adalah nama sekelompok
Rhizophoraceae. Dari segi penampilan, tengar mirip denga
umumnya lebih kecil. Deskripsi umum dari Ceriops tagal yaitu pohon kecil atau
semak dengan ketinggian mencapai 25 m. Kulit kayu berwarna abu-abu,
kadang-kadang coklat, halus dan pangkalnya menggelembung (Noor et al., 1999).
Memiliki nama ilmiah Ceriops tagal, tumbuhan ini juga dikenal dengan beberapa
nama lain seperti tangar, tengal, tengah, tingi, palun, parun, bido-bido dan
Klasifikasi Ceriops tagal dapat diuraikan sebagai berikut:
Kingdom : Plantae
Divisi : Magnoliophyta
Kelas : Magnoliopsida
Ordo : Malpighiales
Family : Rhizophoraceae
Genus : Ceriops
Species : Ceriops tagal
Akar banir dari bentukan seperti akar tunjang, kadang membentuk akar
lutut atau akar nafas yang menonjol. Batang berkayu, warna coklat agak jingga.
Bagian bawah batang terdapat banyak akar tunjang dengan panjang ± 50 cm.
Percabangannya banyak dan tidak teratur. Hipokotil berbintil, berkulit halus, agak
menggelembung dan seringkali agak pendek. Leher kotiledon menjadi kuning jika
sudah matang/dewasa. Ukuran hipokotil silindris panjang 4-25 cm dan diameter
8-12 mm.. Daun hijau mengkilap dan sering memiliki pinggiran yang melingkar
ke dalam, ukuran daunnya1-10 x 2-3,5 cm (Noor et al, 1999).
Faktor-Faktor Lingkungan yang Mempengaruhi Pertumbuhan Ceriops tagal Tanah
Jenis tanah yang mendominasi kawasan mangrove biasanya adalah fraksi
lempeng berdebu, akibat rapatnya bentuk perakaran-perakaran yang ada. Fraksi
lempung berpasir hanya terdapat dibagian depan (arah pantai). Nilai pH tanah
dikawasan mangrove berbeda-beda, tergantung pada tingkat kerapatan vegetasi
nilai pH yang tinggi. Nilai pH tidak banyak berbeda, yaitu antara 4,6-6,5 dibawah
tegakan jenis Rhizophora spp (Noor et al., 1999).
Hutan mangrove tanahnya selalu basah, mengandung garam, mempunyai
sedikit oksigen dan kaya akan bahan organik. Bahan organik yang terdapat di
dalam tanah, terutama berasal dari sisa tumbuhan yang diproduksi oleh mangrove
sendiri. Serasah secara lambat akan diuraikan oleh mikroorganisme, seperti
bakteri, jamur dan lainnya. Selain itu juga terjadi sedimen halus dan partikel
kasar, seperti potongan batu dan koral, pecahan kulit kerang dan siput. Biasanya
tanah mangrove kurang membentuk lumpur berlempung dan warnanya bervariasi
dari abu-abu muda sampai hitam (Soeroyo, 1993).
Umumnya tanah yang ditumbuhi mangrove adalah tanah-
tanah yang bertekstur halus, mempunyai tingkat kematangan rendah, mempunyai
kadar garam rendah alkalinitas tinggi, dan sering mengandung lapisan sulfat
masam atau bahan sulfidik (cat clay). Kandungan liat atau debu umumnya tinggi,
kecuali tanah-tanah atau pecahan batu karang. Lapisan gambut dengan kandungan
garam tinggi kadang-kadang ditemukan pada tanah mangrove baik di daerah batu
karang maupun di daerah endapan liat (Mardiana, 2005).
Suhu
Suhu penting dalam proses fisiologis, seperti fotosintesis dan respirasi.
Pada Rhizophora spp., Ceriops spp., Exocoecaria spp., dan Lumnitzera spp., laju
tertinggi produksi daun baru adalah pada suhu 26-28 ºC, untuk bruguiera spp
adalah 27ºC dan Avicennia marina memproduksi daun baru pada suhu 18-20 ºC
Pasang Surut
Pasang surut menentukan zonasi komunitas flora dan fauna mangrove.
Durasi pasang surut berpengaruh besar terhadap perubahan salinitas pada areal
mangrove. Salinitas air menjadi sangat tinggi pada saat pasang naik dan menurun
selama pasang surut. Perubahan tingkat salinitas pada saat pasang merupakan
salah satu faktor yang membatasi distribusi jenis mangrove. Pada areal yang
selalu tergenang hanya Rhizophora mucronata yang tumbuh baik, sedangkan
Bruguiera spp dan Xylocarpus spp jarang mendominasi daerah yang sering
tergenang. Pasang surut juga berpengaruh terhadap perpindahan massa antara air
tawar dengan air laut, dan oleh karenanya mempengaruhi organisme mangrove
(Ansori, 1998).
Salinitas
Kebanyakan tumbuhan memiliki toleransi sangat rendah terhadap salinitas,
sehingga tidak mampu tumbuh di dalam atau di dekat air laut. Hal ini terjadi
karena kebanyakan jaringan makhluk hidup lebih cair daripada air laut, akibatnya
air dari dalam jaringan tumbuhan dapat keluar akibat proses osmosis, sehingga
tumbuhan kekeringan, menjadi layu, dan mati. Lingkungan yang keras ini
menyebabkan diversitas hutan mangrove cenderung lebih rendah daripada
umumnya hutan hujan tropis (Noor et al., 1999).
Kondisi salinitas sangat mempengaruhi komposisi mangrove. Berbagai
jenis mangrove mengatasi kadar salinitas dengan cara yang berbeda-beda.
Beberapa diantaranya secara selektif mampu menghindari penyerapan garam dari
media tumbuhnya, sementara beberapa jenis yang lainnya mampu mengeluarkan
Adaptasi terhadap salinitas umumnya berupa kelenjar ekskresi untuk
membuang kelebih garam dari dalam jaringan dan ultrafiltrasi untuk mencegah
masuknya garam ke dalam jaringan. Tumbuhan mangrove dapat mencegah lebih
dari 90% masuknya garam dengan proses filtrasi pada akar. Garam yang terserap
dengan cepat diekskresikan oleh kelenjar garam di daun atau disimpan dalam kulit
kayu dan daun tua yang hampir gugur. Beberapa tumbuhan mangrove seperti
Avicennia, Acanthus dan Aegiceras memiliki alat sekresi garam. Konsentrasi
garam dalam cairan biasanya tinggi, sekitar 10% dari air laut. Sebagian garam
dikeluarkan melalui kelenjar garam dan selanjutnya diuapkan angin atau hujan.
Hal ini bisa dirasakan dengan mengecap daun tumbuhan mangrove atau bagian
lainnya. Tumbuhan mangrove seperti Bruguiera, Lumnitzera, Rhizophora, dan
Sonneratia tidak memiliki alat ekskresi garam. Untuk itu membran sel di
permukaan akar mampu mencegah masuknya sebagian besar garam dan secara
selektif menyerap ion-ion tertentu melalui proses ultrafiltrasi (Soeroyo, 1993).
Ceriops tagal adalah mangrove yang toleran terhadap garam dengan
kemampuan dapat tumbuh pada kondisi salinitas tinggi dan miskin unsur hara
(Ball, 1988; Gordon, 1993). Kemampuan yang tinggi dalam beradaptasi pada
konsentrasi garam menguntungkan bagi tanaman untuk tumbuh pada tanah yang
tekanan osmotik tanahnya jauh lebih rendah daripada air laut atau pada kondisi
tanah berkadar garam tinggi (Ungar, 1991).
Meskipun mangrove adalah salah satu jenis halophyte, namun semainya
sensitif terhadap stress garam (Tomlinson, 1986; Lin 1997), substrat yang
bergaram mempengaruhi banyak aspek seperti aspek pertumbuhan dan
Downton, 1982). Banyak penelitian yang telah menemukan bahwa semai tumbuh
paling baik di salinitas rendah (25% air laut atau 0,5% konsentrasi garam), di
salinitas tinggi (50%-75% dari air laut atau 1,5%-2% konsentrasi garam) atau
pada keadaan kekurangan garam (salinitas 0% atau air tawar) adalah efek dari
pertumbuhan (Downton, 1982; Clough, 1984; Wang and Lin, 1999). Pertumbuhan
yang lambat di air tawar sering dianggap berasal dari ketidakmampuan halophyte
untuk mengakumulasi bahan ion anorganik dalam jumlah yang cukup untuk
osmoregulasi ketika substrat kekurangan garam (sodium chloride) (Clough, 1984;
Jenning, 1976; Greenway and Munns, 1980; Yeo and Flower, 1980). Beberapa
peneliti mempertimbangkan peristiwa tersebut menjadi ekspresi dari ciri fisiologi
mangrove yang membutuhkan garam (Wang and Lin, 1999), tetapi beberapa
peneliti telah berusaha menjelaskan mekanisme tersebut.
Biologi Terpenoid Pada Mangrove
Tanaman mangrove telah diketahui sebagai sumber bahan phytokimia atau
bahan biologi aktif (Bandaranayake, 2002). Tanaman menghasilkan beragam
metabolit sekunder, yang mana digunakan dalam interaksi dengan lingkungan,
perkembangan terakhir resisten terhadap berbagai macam stress lingkungan
maupun serangan dari luar. Faktor biotik dan abiotik meningkatkan hasil
metabolit sekunder (Sudha and Ravishankar, 2002). Metabolit sekunder
merupakan bahan kimia yang tidak dibutuhkan atau tidak secara langsung
dibutuhkan untuk pertumbuhan atau reproduksi dari tanaman (Bukingham, 2001).
Metabolit sekunder tanaman juga menunjukkan sumberdaya secara luas seperti
Triterpenoid pentasiklik dan phytosterol tersebar luas di tanaman
mangrove (Wannigama et al., 1981; Hogg and Gillan, 1984; Ghosh et al., 1985;
Koch et al., 2003; Basyuni et al., 2007). Karena jarak aktivitas biologi mereka
lebar, isoprenoid merupakan sumberdaya alam yang sangat penting untuk bahan
obat-obatan (Sparg et al., 2004) dan telah lama tanaman mangrove digunakan
sebagai obat tradisional untuk membunuh penyakit (Bandaranayake, 1998).
Karena lipid terdiri dari bagian penting karbon hasil keluaran dari
mangrove (Wannigama et al., 1981; Hogg and Gillan, 1984), pengetahuan tentang
komposisi lipid di mangrove diharapkan dapat berkontribusi terhadap estimasi
dan dasar akumulasi dari sedimentasi bahan organik. Nonsaponifiable lipids
(NSLs) pada dasarnya menunjukkan fraksi lipid sederhana kecuali asam lemak
(saponifiable lipids) setelah dihidrolisi dengan alkalin baru didapatkan total
lipidnya, dan berisi sterol, cincin alkohol yang panjang dan alkana. Pada
umumnya NSL menunjukkan fraksi lipid yang lebih stabil dibandingkan dengan
fraksi lipid saponifikasi, dan mereka lebih tahan terhadap degradasi mikroba
benar-benar telah dipertimbangkan menjadi faktor penting yang relatif untuk
mengontrol penanda diagenetik (Killops and Frewin, 1994; Koch et al., 2005).
Triterpenoid adalah senyawa kimia yang biasa di jumpai pada tanaman tingkat
tinggi terdiri dari NSL sebagai proporsi utama dan telah diidentifikasi dari lapisan
kutikula tanaman mangrove dan jenis tanaman lainnya (Beaton et al., 1955;
Wannigama et al., 1981; Ghosh et al., 1985; Koch et al., 2003). Beberapa
penelitian sebelumnya telah menggunakan terpenoid pentasiklik yang cocok untuk
pengusutan pertama bahan karbon dari tanaman mangrove, mereka lebih stabil
al., 2004; Koch et al., 2005). Demikian analisis terpenoid adalah prasyarat untuk
interpretasi sinyal penanda pada inti sedimen mangrove.
Potensi Triterpenoid Pada Mangrove
Mangrove terkenal kaya sebagai sumber senyawa triterpenoid dan fitosterol
(isoprenoid) (Koch et al, 2003; Basyuni et al, 2007) karena memiliki berbagai
aktivitas biologis, isoprenoidnya dianggap penting sebagai sumberdaya alam yang
potensial untuk senyawa obat (Sparg et al., 2004). Beberapa aktivitas biologi dari
triterpenoid di mangrove telah dilaporkan. Misalnya, ekstrak dari R. apiculata
telah diguanakan sebagai obat tradisional dan biologi, senyawa aktifnya
diindentifikasi sebagai triterpenoid (Kokpol et al., 1990).
Penelitian sebelumnya menduga bahwa triterpenoid mungkin terlibat dalam
perlindungan mangrove dari stress garam (Oku et al., 2003). Demikian pula, telah
dilaporkan bahwa sintesis triterpene dari Arabidopsis thaliana menunjukkan
respon positif terhadap salinitas (kadar garam) (Zwenger and Basu, 2007).
Selain berfungsi sebagai perlindungan terhadap stress garam, triterpenoid
juga dianggap memainkan peran defensif terhadap serangga. Triterpenoid dari
Rhizophora mangle dapat berfungsi sebagai zat pertahanan kimia karena
menunjukkan aktivitas insektisida (William, 1999). Selain itu, publikasi
sebelumnya telah menunjukkan bahwa ekspresi PgTPS dari sintesis terpenoid
meningkat akibat adanya stress garam dalam Panax ginseng (Kim et al, 2008).
Ekspresi gen dari sintesis triterpenoid dari tiga jenis mangrove yaitu K. candel, B.
gymnorrhiza dan R. stylosa meningkat dengan adanya perbedaan tingkat salinitas
Mekanisme Toleransi Tanaman Terhadap Cekaman Garam
Mekanisme sel tanaman untuk beradaptasi terhadap cekaman garam dan
caranya belum banyak dipahami (Yeo, 1998; Munns, 2005). Meskipun
mekanisme toleransi garam pada tanaman tampak kompleks dan bervariasi,
beberapa mekanisme telah dilaporkan seperti penyesuaian tekanan osmotik oleh
akumulasi molekul kecil seperti osmolytes, glisin-betain atau alkohol gula
(Popp, 1984; Sakamoto and Murata, 2000), ekstrusi garam melewati membran
plasma menggunakan ion transporter (Allen et al., 1995), akumulasi garam dalam
vakuola menggunakan tonoplst transporter (Blumwald and Poole, 1987; Mimura
et al., 2003).
Dalam kondisi cekaman garam, tanaman dapat mengubah tingkat metabolit
sekunder seperti triterpenoid atau senyawa fenolik untuk meningkatkan sistem
pertahanan mereka terhadap stress oksidatif (Kim et al, 2008). Membran plasma
telah memainkan peran penting dalam toleransi tanaman untuk cekaman garam.
Komposisi lipid yang terdapat pada membran mengontrol membran permeabilitas
(Mansour et al., 1994, Kim et al, 2008). Pengamatan ini juga sejalan dengan hasil
penelitian sebelumnya bahwa cekaman garam meningkatkan konsentrasi
triterpenoid di akar dan daun tanaman mangrove serta berkontribusi terhadap
METODE PENELITIAN
Waktu dan Lokasi Penelitian Penananam
Penanaman propagul C. tagal dengan perlakuan berbagai konsentrasi
garam selama 6 bulan dilakukan pada 30 Juli 2011 sampai 24 januari 2012 di
rumah kaca Fakultas Pertanian, Universitas Sumatera Utara.
Pengambilan Sampel dan Ekstraksi
Pengambilan sampel daun dan akar C. tagal diambil dari pohon dewasa
yang dilakukan pada 14 April 2012 di Pulau Sembilan, Kabupaten Langkat,
Sumatera Utara. Ekstraksi lipid dan Analisis NSL (Nonsaponifieble Lipids)
dilakukan pada 25 Juni-Juli 2012, di laboratorium Farmasi, Fakultas Farmasi,
Universitas Sumatera Utara.
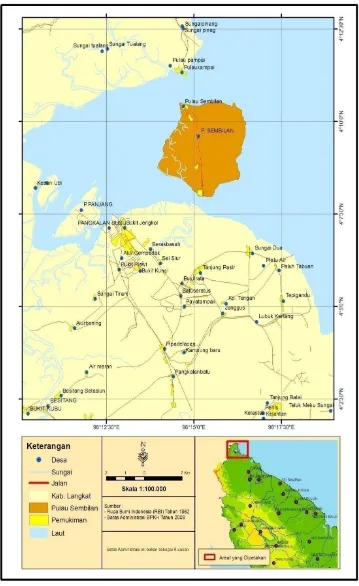
Gambaran Lokasi Pengambilan Sampel
Pulau Sembilan merupakan nama salah satu desa yang berada digugusan
pulau-pulau di Kabupaten Langkat. Desa Pulau Sembilan berdekatan dengan Selat
Malaka dan merupakan salah satu tujuan wisata utama di Kabupaten Langkat.
Pulau Sembilan secara administrasi terletak di kecamatan Pangkalan, Susu
Kabupaten Langkat. Luas Pulau Sembilan 24,00 km2 atau 8.84% dari total luas
kecamatan Pangkalan Susu. Di Pulau ini terdapat hutan mangrove yang
mengelilingi pulau dan tumbuh ekosistem pesisir. Kondisi air tanah masih cukup
baik dimana tidak ditemukan adanya air sumur yang asin atau terkena intrusi air
laut (BPS, 2010). Peta lokasi pengambilan sampel di Pulau Sembilan dapat dilihat
Bahan dan Alat Penelitian Penanaman
Bahan yang digunakan dalam penelitian adalah propagul C. tagal yang
sehat dan matang, bubuk garam komersial (marine salt), pasir dari sungai (tidak
memiliki salinitas), pot plastik, amplop cokelat, dan label.
Alat yang digunakan adalah jangka sorong, penggaris, kamera, oven,
timbangan, cutter, program excel, hand refractometer, image J, program SPSS,
dan alat tulis.
Ekstraksi dan Analisis
Bahan yang digunakan pada penelitian ini adalah daun dan akar pohon
mangrove yang berasal dari jenis C. tagal. Sedangkan bahan kimia dan bahan
lainnya yang digunakan adalah nitrogen cair, klorofom, methanol, hexane, KOH,
ethanol, cholesterol, aluminium foil, kertas tisu.
Alat-alat yang digunakan dalam penelitian ini adalah Salinity refractometer
S/Mill-E (Atago Co. Ltd, Tokyo, Jepang), tabung reaksi untuk mengekstrak daun
dan akar pohon mangrove, rak kultur untuk tempat peletakan tabung reaksi yang
digunakan dalam pengekstrakan, Eyela Evaporator, waterbath, kertas filtrasi No. 2
(Advantec, Tokyo, Jepang).
Analisis Data
Penelitian ini adalah metode analisis dengan Rancangan Acak Lengkap
(RAL) non faktorial dengan 5 perlakuan konsentrasi garam (salinitas) dengan
masing-masing 5 ulangan :
a. Salinitas 0 %
c. Salinitas 1,5 %
d. Salinitas 2 %
e. Salinitas 3 %
Model linear RAL non faktorial
Yij = μ + τi + εij
Di mana : Yij = hasil pengamatan pada perlakuan ke-i dan ulangan ke-j μ = nilai rataan umum ( mean )
τi = pengaruh faktor perlakuan ke-i
εij = pengaruh galat perlakuan ke-i ulangan ke-j i = 1, 2, 3, 4, 5
j = 1, 2, 3, 4, 5
Data dianalisis dengan analisis ANOVA satu arah menggunakan uji
Dunnett untuk perbandingan seluruh perlakuan terhadap kontrol, nilai P< 0.05 dan
P<0,01dipakai sebagai batas untuk menunjukkan pengaruh perlakuan. Uji statistik
dilakukan dengan SPSS versi 16.
Prosedur Penelitian
1. Penyiapan Media Tanam
Propagul dari C. tagal ditanam dalam pot plastik dengan media pasir dan
diberi salinitas bervariasi di bawah sinar matahari alami. Sebuah solusi air laut
disiapkan dengan melarutkan bubuk garam komersial untuk membuat konsentrasi
garam 0%, 0.5%, 1.5%, 2% dan 3% (setara konsentrasi garam air laut). Metode ini
mengacu pada penelitian Basyuni et al., (2009, 2012). Dimana jenis garam yang
dipakai adalah bubuk garam komersial (marine salt). Untuk membuat konsentrasi
salinitas 0.5%, 1.5%, 2% dan 3% dibuat dengan melarutkan 5,66g, 17g, 22,6g,
garam/ massa larutan. Konsentrasi garam pada setiap perlakuan pot diperiksa
seminggu sekali selama percobaan dengan hand refraktometer.
2. Pemilihan Propagul
Propagul C. tagal yang digunakan berasal dari pohon induk yang berumur
5 tahun atau lebih. Propagul yang dipilih sebaiknya telah matang secara fisiologi
dengan warna propagul hijau kecoklatan dan sehat, tidak terserang oleh hama dan
penyakit
3. Penanaman Propagul
Propagul C. tagal yang telah disediakan ditanam ke dalam pot plastik yang
telah berisi media tumbuh yang telah disesuaikan dengan perlakuannya
masing-masing. Kemudian pot plastik diberi tanda/label sesuai dengan perlakuan yang
diberikan.
3. Ekstraksi Lipid
Daun C. tagal sebanyak 2-4 lembar atau 4-6 g akar digerus dengan
Nitrogen cair, kemudian di ekstrak dengan chloroform-methanol 2:1 (CM21),
dinding sel yang berisi kotoran yang tidak larut dalam CM21 disaring dengan
kertas filtrasi No. 2 (Advantec, Tokyo, Jepang) dan yang tersisa adalah ekstrak
lipid di dalam chloroform. Sebagian ekstrak dimurnikan untuk dianalisis
kandungan lipidnya seperti yang digambarkan sebelumnya (Basyuni et al., 2007).
Cairan ekstrak lipid yang pekat dikeringkan kemudian ditimbang dan di dapatkan
berat lipidnya. Sehingga dapat diketahui kandungan total lipid/tisue (mg/g tisue).
4. Analisis NSL (Nonsaponifieble Lipids)
Lipid ekstrak di dalam chloroform (yang telah diketahui berat total
di refluxed selama 10 menit dengan suhu 90º C, ditambahkan 2 ml Hexane (NSL)
kemudian diaduk. Lapisan Hexane dipindahkan kedalam tube yang telah diketahui
beratnya, kemudian cairan di keringkan dengan Nitrogen stream, kemudian
dikeringkan di bawah vakum selama 10 menit, selanjutnya ditimbang berat
NSLnya. sehingga dapat diketahui kandungan NSL/tisue (mg/g tisue) dan
kandungan NSL/total lipid (mg/mg total lipid).
Pengamatan Parameter
Pengamatan dilakukan 6 bulan setelah tanam dan parameter yang diamati
adalah:
1. Pertambahan Tinggi Semai (cm)
Pengambilan data tinggi pertama semai dilakukan setelah 2 bulan
penanaman propagul dan pengambilan data tinggi terakhir setelah 6 bulan tanam
dengan menggunakan penggaris, pada setiap satuan percobaan. Tinggi semai
diukur mulai dari bagian plumula sampai titik tumbuh tertinggi.
2. Pertambahan Diameter Semai (mm)
Pengukuran diameter semai dilakukan pada tanda awal dengan
menggunakan jangka sorong. Pengambilan data diameter dilakukan bersamaan
dengan pengambilan data tinggi semai.
3. Pertambahan Jumlah Daun
Penghitungan jumlah daun dilakukan pada awal munculnya daun mulai
dari pucuk. Pengambilan data dilakukan bersamaan dengan pengambilan data
4. Luas Daun (cm2)
Pengukuran luas daun dilakukan pada akhir pengamatan data. Perhitungan
luas daun menggunakan program komputer. Untuk melakukan perhitungan
terlebih dahulu daun digambar di kertas millimeter blok yang selanjutnya
dilakukan scanning pada gambar tersebut. Setelah di scanning maka gambar
tersebut dihitung dengan program image J (NIH).
5. Berat Basah Akar (g)
Untuk mendapatkan berat basah akar, bagian akar yang baru dipanen
dimasukkan ke dalam amplop dan diberi label sesuai dengan perlakuan.
Ditimbang berat awal akar C. tagal.
6. Berat Basah Tajuk (g)
Untuk mendapatkan berat basah tajuk, bagian tajuk yang baru dipanen
dimasukkan ke dalam amplop dan diberi label sesuai dengan perlakuan.
Ditimbang berat awal tajuk C. tagal.
7. Berat Basah Batang (g)
Untuk mendapatkan berat basah batang, bagian batang yang baru dipanen
dimasukkan ke dalam amplop dan diberi label sesuai dengan perlakuan.
Ditimbang berat awal batang C. tagal.
8. Berat Kering Akar (g)
Untuk mendapatkan berat kering akar, bagian akar dimasukkan ke dalam
amplop dan diberi label sesuai dengan perlakuan. Kemudian akar C. tagal dioven
pada suhu 75ºC sampai berat kering konstan (2-3 hari), lalu ditimbang berat
kering akar C. tagal.
9. Berat Kering Tajuk (g)
Untuk mendapatkan berat kering tajuk, bagian tajuk dimasukkan ke dalam
amplop dan diberi label sesuai dengan perlakuan. Kemudian tajuk C. tagal dioven
pada temperatur 75ºC sampai berat kering konstan (2-3 hari), lalu ditimbang berat
kering tajuk C. tagal.
10.Berat Kering Batang (g)
Untuk mendapatkan berat kering batang, bagian batang dimasukkan ke
dalam amplop dan diberi label sesuai dengan perlakuan. Kemudian batang C.tagal
dioven pada temperatur 75ºC sampai berat kering konstan (2-3 hari), lalu
ditimbang berat kering batang C. tagal.
11. Rasio Tajuk dan Akar
Perhitungan rasio tajuk dan akar dilakukan pada akhir pengamatan.
Perhitungan rasio tajuk dan akar dapat diperoleh dengan menggunakan rumus
sebagai berikut : Rasio =
akar kering Berat
tajuk kering Berat
12. Rasio Batang dan Akar
Perhitungan rasio batang dan akar dilakukan pada akhir pengamatan.
Perhitungan rasio batang dan akar dapat diperoleh dengan menggunakan rumus
sebagai berikut : Rasio =
akar kering Berat
batang kering
HASIL DAN PEMBAHASAN
Pengaruh Salinitas Terhadap Pertumbuhan Semai
Pengaruh salinitas terhadap pertumbuhan semai C. tagal berdasarkan
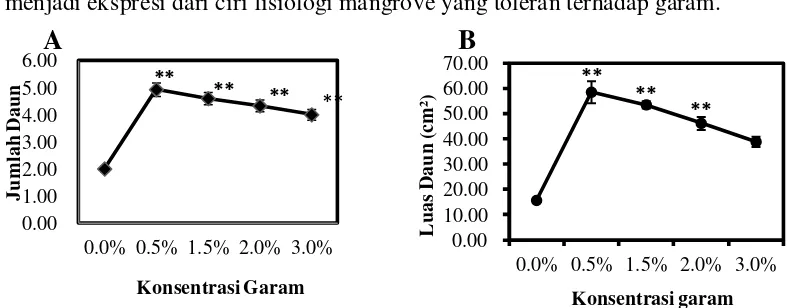
pengukuran tinggi dan diameter tanaman dapat dilihat pada gambar 2A dan B.
Gambar 2. Pengaruh salinitas terhadap pertumbuhan Tinggi Semai (A) dan Diamter Semai (B). Data merupakan rata-rata ± SE (n = 13-15). Tanda (*) dan (**) mengindikasikan secara statistik signifikan dari kontrol (0%) pada P< 0.05 dan P< 0.01 dengan uji Dunnet.
Berdasarkan hasil penelitian yang didapat bahwa konsentrasi salinitas
berpengaruh nyata terhadap pertambahan tinggi semai C. tagal. Semai tertinggi
diperoleh pada pemberian salinitas 0,5%, yaitu 3,24cm. Berdasarkan uji Dunnet
pada P<0,01 berpengaruh nyata terhadap pertumbuhan tinggi semai C. tagal pada
salinitas 0,5%, 1,5%, 2% dan 3% dibandingkan terhadap kontrol. Pertumbuhan
tinggi pada konsentrasi 0,5% menunjukkan kenaikan secara signifikan kemudian
turun dengan pertambahan tingkat salinitas seperti yang terlihat pada Gambar 2A.
Pertumbuhan diameter batang terbesar semai C. tagal terdapat pada
pemberian salinitas 0,5% yaitu 3,65mm. Berdasarkan uji Dunnet pada P<0,01 0.0
Konsentrasi garam
B
pemberian salinitas berpengaruh nyata terhadap pertumbuhan diameter pada
salinitas 0,5%, 1,5% dan 2% dibandingkan terhadap kontrol. Pertumbuhan
diameter semai C. tagal menunjukkan kenaikan yang signifikan pada salinitas
0,5%, 1,5% dan 2% kemudian turun pada salinitas 3% seperti yang terlihat pada
Gambar 2B.
Dari Gambar 2A dan B, semai C. tagal menunjukkan pertumbuhan terbaik pada salinitas 0,5 % (3,24cm, 3,65mm) dan masih tumbuh cukup baik pada
salinitas 2% (2,65cm, 3,38mm) dan bahkan pada salinitas 3% (1,63cm, 2,79mm)
C. tagal masih bisa tumbuh walaupun pertumbuhannya mulai menurun. Hal ini menunjukkan bahwa C. tagal termasuk jenis yang memiliki kemampuan yang cukup tinggi dalam menghadapi salinitas tinggi hal ini sesuai dengan studi Ball
(1988) dan Gordon (1993) yang melaporkan bahwa C. tagal adalah mangrove
yang toleran terhadap garam dengan kemampuan dapat tumbuh pada kondisi
salinitas tinggi (salinitas 3%) dan miskin unsur hara. Kemampuan yang tinggi
dalam beradaptasi pada konsentrasi garam menguntungkan bagi tanaman untuk
tumbuh pada tanah yang tekanan osmotik tanahnya jauh lebih rendah daripada air
laut atau pada kondisi tanah berkadar garam tinggi.
Berdasarkan Gambar 2A dan B pertumbuhan tinggi dan diameter semai
C. tagal. Pertumbuhan tinggi dan diameter yang paling rendah terdapat pada
kontrol (salinitas 0% atau air tawar) yaitu 0,58cm dan 2,61mm. Ini menunjukkan
bahwa C. tagal pertumbuhannya sangat lambat pada air tawar dan hasil ini mengindikasikan bahwa semai C. tagal untuk pertumbuhannya juga membutuhkan konsentrasi garam untuk mengakumulasi bahan ion anorganik, hal
(1980) dan Yeo and Flower (1980) yang melaporkan bahwa pertumbuhan yang
lambat di air tawar sering dianggap berasal dari ketidakmampuan halophyte untuk
mengakumulasi bahan ion anorganik dalam jumlah yang cukup untuk
osmoregulasi ketika substrat kekurangan garam (sodium chloride). Wang and Lin
(1999) juga menyatakan beberapa peneliti mempertimbangkan peristiwa tersebut
menjadi ekspresi dari ciri fisiologi mangrove yang toleran terhadap garam.
kontrol dapat dilihat pada Gambar 3B.
Gambar 3. Rata-rata Jumlah Daun (A) dan Luas Daun Ceriops tagal (B). Data merupakan rata-rata ± SE (n = 13-15) untuk rata-rata jumlah daun dan SE (n= 5) untuk luas daun. Tanda (*) dan (**) mengindikasikan secara statistik signifikan dari kontrol (0%) pada P< 0.05 dan P< 0.01 dengan uji Dunnet.
Pengaruh salinitas terhadap rata-rata jumlah daun dan luas daun C. tagal
seperti yang terlihat pada gambar 3A dan B. Rata-rata jumlah daun tertinggi pada
salinitas 0,5%, yaitu 5 helai. Berdasarkan uji Dunnet pada P<0,01 pemberian
salinitas berpengaruh nyata terhadap rata-rata jumlah daun pada salinitas 0,5%,
1,5%, 2%, dan 3% terhadap kontrol dapat di lihat pada Gambar 3A.
Luas daun tertinggi terdapat pada salinitas 0,5%, yaitu 58,50cm².
Berdasarkan uji Dunnet pada P<0,01 pemberian salinitas berpengaruh nyata
terhadap luas daun pada salinitas 0,5%, 1,5% dan 2% dibandingkan terhadap
kontrol. Luas daun pada konsentrasi 0,5% menunjukkan kenaikan yang signifikan
Gambar 3B. Hal ini menunjukkan laju pertumbuhan daun dan luas daun
berbanding terbalik dengan kenaikan salinitas. Hal ini sesuai dengan hasil
penelitian Kusmiyati et al. (2002) menunjukkan pengelolaan tanah salin dengan
pertambahan luas daun. Laju pertumbuhan daun dan luas akhir daun berbanding
linier dan terbalik (negatif) dengan kenaikan salinitas. Salinitas menurunkan laju
pertumbuhan daun melalui pengurangan laju pembesaran sel pada daun. Haryadi
dan Yahya (1988) menyatakan pengaruh salinitas terhadap pertumbuhan dan
perubahan struktur tanaman yaitu antara lain lebih kecilnya ukuran daun.
Penyerapan hara dan air yang berkurang akan menghambat laju fotosintesis yang
pada akhirnya akan menghambat pertumbuhan tanaman.
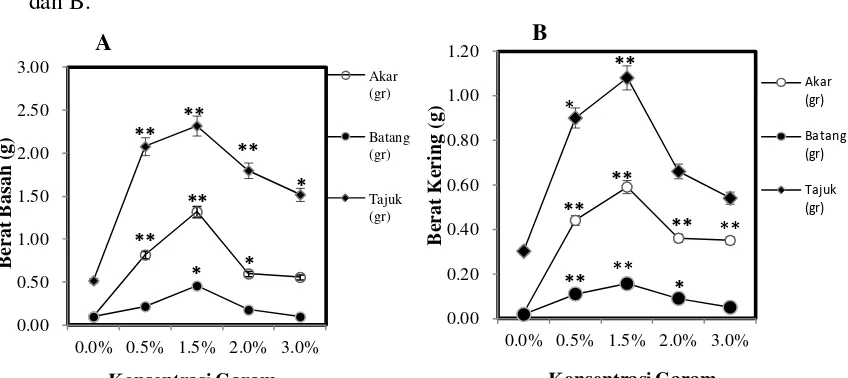
Pengaruh Salinitas Terhadap Biomassa Semai
Berdasarkan penelitian yang dilakukan terdapat kaitan berbagai macam
salinitas terhadap biomassa semai C. tagal seperti yang terlihat pada Gambar 4A
dan B.
Berat basah akar berdasarkan uji Dunnet pada P<0,01 berpengaruh nyata
pada salinitas 0,5% dan 1,5% dan pada P<0,05 salinitas 2% juga berpengaruh
nyata dibandingkan terhadap kontrol dapat dilihat pada Gambar 4A. Berat basah
tertinggi yaitu pada salinitas 1,5% sebesar 1,32g. Sedangkan berat kering akar,
berdasarkan uji Dunnet pada P<0,01 berpengaruh nyata pada salinitas 0,5%,
1,5%, 2% dan 3% dibandingkan terhadap kontrol dapat dilihat pada Gambar 4B.
Berat kering akar tertinggi terdapat pada salinitas 1,5% yaitu 0,59g.
Berat basah batang C. tagal berdasakan uji Dunnet pada P<0,05
berpengaruh nyata pada salinitas 1,5% dibandingkan terhadap kontrol dapat
dilihat pada Gambar 4A. Berat basah batang tertinggi yaitu pada salinitas 1,5%
sebesar 0,38g. Sedangkan berat kering batang, berdasarkan uji Dunnet pada
P<0,01 berpengaruh nyata pada salinitas 0,5 dan 1,5%, sedangkan pada P<0,05
salinitas 2% juga berpengaruh nyata terhadap kontrol dapat dilihat pada Gambar
4B. Berat kering batang tertinggi terdapat pada salinitas 1,5% yaitu 0,16g.
Berat basah tajuk C. tagal berdasarkan uji Dunnet pada P<0,01
berpengaruh nyata pada salinitas 0,5%, 1,5% dan 2%, sedangkan pada P<0,05
salinitas 3% juga berpengaruh nyata dibandingkan terhadap kontrol dapat dilihat
pada Gambar 4A. Berat basah tajuk tertinggi yaitu pada salinitas 1,5% sebesar
2,32g. Berat kering tajuk berdasarkan uji Dunnet pada P<0,01 berpengaruh nyata
pada salinitas 1,5%, sedangkan pada P<0,05 salinitas 0,5% juga berpengaruh
nyata terhadap kontrol dapat dilihat pada Gambar 4B. Berat kering tajuk tertinggi
Berdasarkan dari data berat basah dan berat kering akar, batang dan tajuk
yang didapat bahwa produksi biomassa dari C. tagal dipengaruhi oleh salinitas
0,5% dan 1,5%.
Berdasarkan hasil pertumbuhan tinggi, diameter, dan biomassa semai
C. tagal dapat disarankan bahwa salinitas 0,5 dan 1,5 % adalah konsentrasi
salinitas yang paling baik untuk pertumbuhan semai C. tagal. Dari hasil ini juga
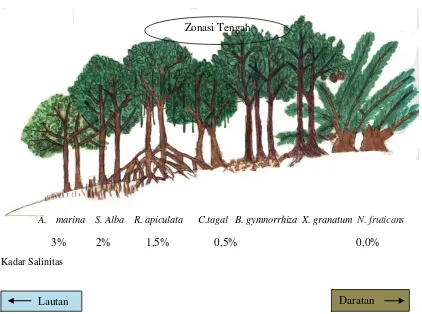
dapat di gambarkan bahwa posisi C. tagal pada zonasi mangrove di Pulau
Sembilan berada di antara jenis mangrove Rhizophora sp dan Bruguiera sp atau
termasuk ke dalam zonasi tengah yaitu zonasi yang berada dibelakang garis pantai
yang akan tergenang apabila terjadi pasang tinggi pada zonasi tersebut, dapat
dilihat pada Gambar 5.
Gambar 5. Contoh Zonasi Mangrove di Pulau Sembilan Kabupaten Langkat Sumatera Utara.
A. marina S. Alba R. apiculata C.tagal B. gymnorrhiza X. granatum N. fruticans
Kadar Salinitas
Lautan Daratan
3% 2% 1,5% 0,5% 0,0%
Zonasi mangrove di Pulau Sembilan tidak jauh berbeda dengan zonasi di
Cilacap Jawa Tengah, berdasarkan studi Noor et al., (1999) yang melaporkan
bahwa posisi C. tagal di dalam zonasi mangrove di Cilacap berada diantara
R. mucronata dan B. cylindrica. Berbeda dengan zonasi mangrove di sungai
Nakama, Jepan. Berdasarkan studi ISME (2001) yang melaporkan bahwa zonasi
mangrove di sungai Nakama, Jepang dari zonasi terluar didominasi oleh jenis
A. marina, dan Soneratia alba, sedangkan zonasi tengah di dominasi oleh
R. stylosa, Kandelia candel, dan Bruguiera gymnorhiza, sedangkan zonasi
belakang di dominasi oleh H. littolaris, dan Pandanus sp, perbedaan antara zonasi
mangrove di sungai Nakama, Jepang dengan zonasi mangrove di Pulau Sembilan
hanya terletak pada zonasi tengah dan zonasi belakang sungai Nakama, Jepang
yaitu pada komposisi jenis mangrove yang menyusun zonasi tersebut seperti jenis
K. kandel dan H.littolaris, Pandanus sp. Jenis tersebut tidak terdapat di mangrove
Pulau Sembilan.
Lebar zonasi di mangrove berbeda-beda tergantung pada intrusi air laut
yang sangat dipengaruhi oleh tinggi rendahnya pasang surut air laut hal ini sesuai
dengan studi Erftemeijer et al (1989) yang melaporkan panjang hamparan ini
bergantung pada intrusi air laut yang sangat dipengaruhi oleh tinggi rendahnya
pasang surut, pemasukan dan pengeluaran material kedalam dan dari sungai, serta
kecuramannya
Meskipun kelihatannya terdapat zonasi dalam vegetasi mangrove, namun
kenyataan di lapangan tidaklah sesederhana itu. Banyak formasi serta zona
vegetasi yang tumpang tindih dan bercampur serta seringkali struktur dan korelasi
Rasio Tajuk/Akar Dan Rasio Batang/Akar Semai
Pengaruh salinitas terhadap rasio tajuk/akar dan rasio batang/akar semai
C. tagal berdasarkan perbandingan berat kering tajuk dan akar (rasio tajuk/akar),
dan berat kering batang dan akar (rasio batang/akar) dapat dilihat pada gambar 2A
dan B.
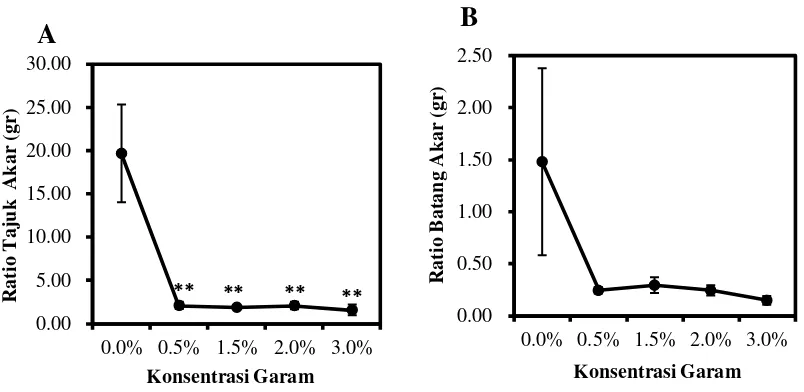
Gambar 6. Rasio Tajuk dan Akar (A), Rasio Batang dan Akar (B). Data merupakan rata-rata ± SE (n=5). Tanda (*) dan (**) mengindikasikan secara statistik signifikan dari kontrol (0%) pada P< 0.05 dan P< 0.01 dengan uji Dunnet.
Rasio tajuk/akar C. tagal dari berbagai tingkat salinitas yang terbesar
adalah pada semai C. tagal salinitas 0% yaitu 19,64 g dan terendah pada semai
C. tagal salinitas 3% yaitu 1,56 g. Rasio tajuk/akar C. tagal berdasarkan uji
Dunnet pada P<0,01 berpengaruh nyata pada salinitas 0,5%, 1,5%, 2%, dan 3%
dibandingkan terhadap kontrol dapat dilihat Gambar 6A. Perbandingan atau rasio
tajuk (pucuk) dan akar sangat menentukan perkembangan selanjutnya terutama
dalam hal produksi. Disini jelas dibutuhkan adanya keseimbangan antara rasio
pertumbuhan daun dengan akar. Artinya agar baik pertumbuhan akar maupun
daun sama-sama tumbuh dan berkembang secara normal dan seimbang tanpa
Rasio batang/akar C. tagal dari berbagai tingkat salinitas yang terbesar
adalah semai C. tagal salinitas 0% yaitu 1,48 g dan terendah pada semai C. tagal
salinitas 0,5% yaitu 0,15 g. Rasio batang/akar berdasarkan uji Dunnet tidak
berpengaruh nyata terhadap kontrol dapat di lihat pada Gambar 6B. Rasio
batang/akar menunjukkan bahwa rerata berat kering akar lebih besar dibanding
berat kering batang. Rasio berat kering batang/akar merupakan karakter fisiologi
yang dapat membantu untuk memahami pertumbuhan relatif batang-akar. Hal ini
berkaitan dengan sinar matahari atau naungan, tanah yang lembab atau tanah yang
kering serta salinitas.
Total Lipid dan Kandungan NSL (Nonsaponifiable Lipid) C. tagal Pada Tingkat Pohon
Total lipid dan kandungan NSL didapat dari hasil ekstraksi daun dan akar
C. tagal. Hasil ekstraksi dapat dilihat pada Tabel 1.
Tabel 1. Total Lipid dan Kandungan NSL pada jenis Ceriops tagal
Jenis Jaringan
Daun 5,0615 16,2 3,5 3,2 0,69 0,22
Akar 6,1512 8,9 0,2 1,45 0,03 0,02
Berdasarkan Tabel 1. di atas hasil ekstraksi dari daun dan akar C. tagal didapatkan kandungan total lipid/jaringan dan NSL/jaringan di daun sebesar
3,2mg/g dan 0,69mg/g. Hasil total lipid jenis C. tagal dari hutan mangrove Pulau Sembilan tidak jauh berbeda dengan kandungan total lipid pada daun dari jenis
P. acidula yang merupakan salah satu jenis mangrove di Okinawa, Jepang yaitu 3,29mg/g jaringan (Basyuni et al,. 2007), tetapi sangat jauh berbeda dengan
Sedangkan di akar sebesar 1,45mg/g dan 0,03mg/g. Dari hasil pada Tabel 1. dapat
dilihat bahwa kandungan total lipid dan NSL ternyata lebih banyak terdapat di
daun daripada di akar. Hal ini mungkin disebabkan oleh fungsi akar pada jenis
mangrove non sekresi yaitu sebagai ultra filter sehingga kandungan lipidnya lebih
sedikit. Dimana membran sel di permukaan akar mampu mencegah masuknya
sebagian besar garam dan bersifat selektif menyerap ion-ion tertentu. Hal ini
sesuai dengan studi Soeroyo (1993) yang melaporkan tumbuhan mangrove seperti
Bruguiera, Lumnitzera, Rhizophora, tidak memiliki alat ekskresi garam. Untuk itu
membran sel di permukaan akar mampu mencegah masuknya sebagian besar
garam dan secara selektif menyerap ion-ion tertentu melalui proses ultrafiltrasi.
Total lipid dan kandungan NSL lebih banyak terdapat di daun bisa saja
disebabkan proses physiologi dari tanaman mangrove jenis non sekresi yang
menghasilkan senyawa metabolit sekunder berupa lipid khusunya NSL agar dapat
mempertahankan diri dari faktor lingkungan baik biotik maupun abiotik. Hal ini
sesuai dengan studi Sudha and Ravishankar (2002) yang melaporkan faktor biotik
dan abiotik meningkatkan hasil metabolit sekunder yang mana digunakan dalam
interaksi dengan lingkungan, perkembangan terakhir resisten terhadap berbagai
macam stress lingkungan maupun serangan dari luar. Dengan demikian, lipid hasil
metabolit sekunder pada membran sel memiliki peranan yang penting dalam
adaptasi tanaman terhadap lingkungan.
Pengetahuan tentang lipid dan kandungan NSL pada jenis C. tagal diharapkan dapat berkontribusi terhadap estimasi dan dasar perhitungan dari
umumnya NSL menunjukkan fraksi lipid yang lebih stabil dibandingkan dengan
fraksi lipid saponifikasi. Hal ini sesuai dengan studi Killops and Frewin (1994),
Koch et al (2005) yang melaporkan pada umumnya NSL menunjukkan fraksi lipid
yang lebih stabil dibandingkan dengan fraksi lipid saponifikasi, dan mereka lebih
tahan terhadap degradasi mikroba dan benar-benar telah dipertimbangkan menjadi
KESIMPULAN DAN SARAN
Kesimpulan
1. Salinitas 0,5% memberikan pengaruh pertumbuhan dan biomassa semai
C. tagal terbaik.
2. Posisi C. tagal pada zonasi mangrove di Pulau Sembilan berada di antara
jenis mangrove Rhizophora sp dan Bruguiera sp atau termasuk ke dalam
zonasi tengah
3. Kandungan total lipid/tisue dan NSL/tisue di daun sebesar 3,2 mg/g dan
0,69 mg/g.
4. Kandungan total lipid/tisue dan NSL/tisue di akar sebesar 1,45 mg/g dan
0,03 mg/g.
Saran
Pembibitan semai C. tagal yang akan digunakan untuk penanaman dan
rehabilitasi hutan mangrove sebaiknya dilakukan pada tingkat salinitas 0,5%, yang
memberikan tingkat pertumbuhan terbaik. Diperlukan penelitian lanjutan untuk
DAFTAR PUSTAKA
Allen, G.J., Wyn-Joens, R.G., Leigh, R.A. 1995. Sodium transport measured in plasma membrane vesicles isolated from wheat genotypes with differing K+/Na+ determination traits. Plant Cell Environ. 18, 105-115.
Ansori, S. 1998. Studi Sifat Fisik dan Pasang Surut Air Laut Terhadap Penyebaran Jenis Rhizophora Hutan Mangrove Pantai Tempora Jatim. Fakultas Kehutanan. Institut Pertanian Malang. Malang.
Ball MC, Farquhar GD. 1984. Photosynthetic and stomatal responses of two mangrove species, Aegiceras corniculatum and Avicennia marina, to Long.
Basyuni, M., Oku, H., Baba, S., Takara, K., Iwasaki, and Oku, H. 2007. Isoprenoids of Okinawan mangroves as lipid input into estuarine ecosystem.. J. Oceanogr.63, 601-608.
Basyuni, M., Oku, H., Tsujimoto, E., Kinjo, K., Baba, S., Takara, K. 2007. Triterpene synthases from the Okinawan mangrove tribe, Rhizophoraceae. FEBS J. 274, 5028-5042.
Basyuni, M., Baba, S., Inafuku, M., Iwasaki, H., Kinjo, K., Oku, H. 2009. Expression of terpenoid synthase mRNA and terpenoid content in salt stressed mangrove. J. Plant Physiol. 166, 1786-1800.
Basyuni, M., Kinjo, Y., Baba, S., Shinzato, N., Iwasaki, H., Siregar, E.B.M., Oku, H. 2011. Isolation of Salt Stress Tolerance Genes from Roots of Mangrove Plant, Rhizophora stylosa Griff., using PCR-based Suppression Subtractive Hybridization. Plant Mol. Biol. Rep. 29, 533-543.
Beaton, J.M., Spring, F.S., Stevenson R., Steward, J.L. 1955. Triterpenoids XXXVII. The constitution of taraxerol. J. Chem. Soc. 2131-2137.
Blumwald, E., Poole, R.J. 1987. Salt tolerance in suspension cultures of sugar beet: induction of Na+/H+ antiport activity at the tonoplast by growth in salt. Plant Physiol. 83, 884-887.
Buckingham, J. 2001. Dictionary of Natural Products on CD-ROM. Chapman & Hall, London. 210 pp.
Clough, B., Tan, D.T., Phuong, D.X., Buu, D.C. 2000. Canopy leaf area index and litter fall in stands of the mangrove Rhizophora apiculata of different age in the Mekong Delta, Vietnam. Aquat. Bot. 66, 311-320.
Downton WJS. 1982. Growth and osmotic relations of the mangrove Avicennia marina as influenced by salinity. Aust J Plant Physiol 9:519-528.
FAO. 2007. The World’s Mangroves 1980–2005. Forest Resources Assessment Working Paper No. 153. Food and Agriculture Organization of The United Nations. Rome.
Ghosh, A., Misra, S., Dutta, A.K., Choudhury, A. 1985. Pentacyclic triterpenoids and sterols from seven species of mangrove. Phytochemistry 24, 1725-1727.
Greenway H, Munns R. 1980. Mechanisms of salt tolerance in non halophytes. Annu Rev Plant Physiol 31:149-190.
Hogg, R.W., Gillan, F.T. 1984. Fatty acids, sterols and hydrocarbons in the leaves from eleven species of mangrove. Phytochemistry 23, 93-97.
Hutching, P. And P. Saenger. 1987. Ecology of Mangrove. University of Queensland Press. Australia.
Jennings DH. 1976. The effects of sodium chloride on higher plants. Biol Rev 51:453-486.
Kathiresan, K. and B. L. Bingham. 2001. Biology of mangrove and mangrove ecosystem. Adv. Mar. Biol. 40, 81-151.
Killops, S.D., Frewin, N.L. 1994. Triterpenoid diagenesis and cuticular preservation. Org. Geochem. 21, 1193-1209.
Kim, Y.J., Ham, A.R., Shim, J.S., Lee, J.H., Jung, D.Y., In J.G., Lee, B.S.,Yang, D.C. 2008. Isolation and characterization of terpene synthase gene from Panax ginseng. J. Ginseng Res. 32,114–119.
Koch, B.P., Rullkotter, J., Lara, R.J. 2003. Evaluation of triterpenoids and sterols as organic matter biomarkers in a mangrove ecosystem in northen Brazil. Wetl. Ecol. Manag. 11, 257-263.
Koch, B.P., Harder, J., Lara, R.J., Kattner, G. 2005. The effect of selective microbial degradation on 111 the composition derived pentacyclic triterpenols in surface sediments. Org. Geochem. 36, 273-285.
Lin P (eds). 1997. Mangrove ecosystem in China (in Chinese, with English abstract). Science Press, Beijing
Mansour, M.M.F., van Hasselt, P.R., Kuiper, P.J.C. 1994. Plasma membrane lipid alternations induced by NaCl in winter wheat roots. Physiol. Plant. 92: 473–478.
Mardiana, S. 2005. Perbedaan Kondisi Fisik Lingkungan Terhadap Pertumbuhan Berbagai Tanaman Mangrove. Fakultas Pertanian Universitas Medan Area. Medan. Jurnal Penelitian Bidang Ilmu Pertanian Volume 3, Nomor 1, April 2005.
Mimura, T., Kura-Hotta, M., Tsujimura, T., Ohnishi, M., Miura, M., Okazaki, Y., Mimura, M., Maeshima, M., Washitani-Nemoto, S. 2003. Rapid increase of vascular volume in response to salt stress. Planta 216, 397-402.
Munns, R., 2005. Genes and salt tolerance: bring them together. New Phytol. 167, 645-663.
Noor, Y, R., M. Khazali, I. N. N. Suryadiputra. 1999. Panduan Pengenalan Mangrove di Indonesia. PKA/WI-IP. Bogor.
Oku, H., Baba, S., Koga, H., Takara, K., Iwasaki, H., 2003. Lipid composition of mangroves and its relevance to salt tolerance. J. Plant Res. 116, 37-45.
Popp, M. 1984. Chemical composition of Australian mangroves II. Low molecular weight carbohydrates. Z. Pflanzenphysiol. 113, 411-421.
Pramudji dan L. H. Purnomo. 2003. Mangrove Sebagai Tanaman Penghijauan Pantai. Pusat Penelitian Oseanografi LIPI. Jakarta : 1 – 6.
Sakamoto, A., Murata, N. 2000. Genetic engineering of glycinebetaine synthesis in plants: current status and implication for enhancement of stress tolerance. J. Exp. Bot. 51, 81-88.
Scholander, P.F., Hammel, H.T., Hemmingsen, E., Garey, W. 1962. Salt balance in mangroves. Plant Physiol. 37, 722-729.
Soeroyo, 1993. Pertumbuhan Mangrove dan Permasalahannya. Buletin Ilmiah INSTIPER. Yogyakarta.
Sparg, S.G., Light, M.E., van Staden, J. 2004. Biological activities and distribution of plant saponins. J. Ethnopharmacol. 94, 219-243.
Sugihara, K., Hanagata, N., Dubinsky, Z., Baba, S., Karube, I. 2000. Molecular characterization of cDNA encoding oxygen evolving enhancer protein 1 increased by salt treatment in the mangrove Bruguiera gymnorrhiza. Plant Cell Physiol. 41, 1279-1285.
Tomlinson P.B. 1986. The Botany of Mangroves. Cambridge University Press; 1986.
Ueda, A., Shi, W., Nakamura, T., Takabe, T. 2002. Analysis of salt-inducible genes in barley roots by differential display. J. Plant Res. 115, 119-130.
Versteegh, G.J.M., SchefuP, E., Dupont, L., Marret, F., Damsté, J.S.S., Jansen, J.H.R. 2004. Taraxerol and Rhizophora pollen as proxies for tracking past mangrove ecosystems. Geochim. Cosmochim. Acta 68, 411-422.
Williams, L.A.D. 1999. Rhizophora mangle (Rhizophoraceae) triterpenoids with insectidal activity. Naturwissenschaften 86, 450-452.
Wang WQ, Lin P. 1999. Influence of substrate salinity on the growth of mangrove species of Bruguiera gymnorrhiza seedling (in Chinese). J Xiamen Univ (Nat Sci) 38 (2): 237-279.
Wannigama, G.P., Volkman, J.K., Gillan, F.T., Nichols, P.D., Johns, R.B. 1981. A comparison of lipid components of the fresh and dead leaves and pneumatophores of the mangrove Avicennia marina. Phytochemistry 20, 659-666.
Yamada, A., Saitoh, T., Mimura, T., Ozeki, Y. 2002. Expression of mangrove allene oxide cyclase enhances salt tolerance in Escherichia coli, yeast, and tobacco cells. Plant Cell Physiol. 43, 903-910.
Yeo AR, Flower TJ. 1980. Salt tolerance in the halophyte Suaeda maritime L. Dum.: evaluation of the effect of salinity upon growth. J Exp Bot 31:1171-1183.
Yeo, A. 1998. Molecular biology of salt tolerance in the context of whole-plant physiology. J. Exp. Bot. Vol. 49, 915–929.