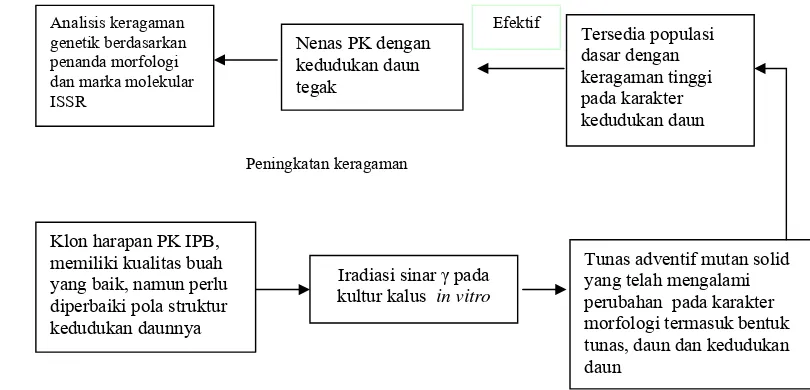
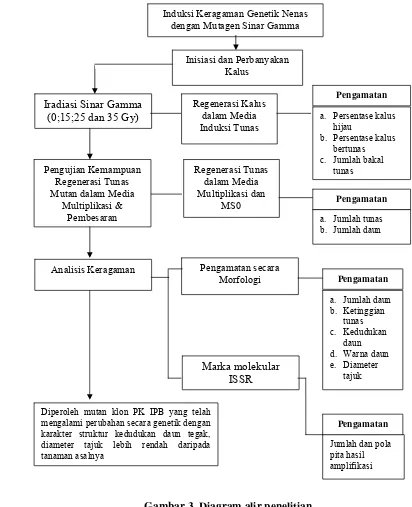
Induksi Keragaman Genetik Dengan Mutagen Sinar Gamma pada Nenas Secara In Vitro
Teks penuh
Gambar
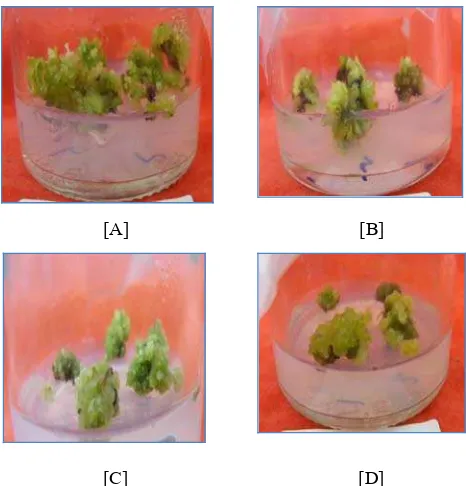
![Gambar 2. [A] Deskripsi tanaman klon PK di lapangan; [B] Karakteristik buah nenas klon PK; [C] Plantlet in vitro nenas klon PK](https://thumb-ap.123doks.com/thumbv2/123dok/713885.361938/25.612.128.508.300.457/gambar-deskripsi-tanaman-lapangan-karakteristik-plantlet-vitro-nenas.webp)

Dokumen terkait
Site didasarkan atas konsep yang telah dipaparkan sebelumnya, yaitu akses menuju site mudah dijangkau oleh pengunjung,hal itu akan memberikan pengaruh terhadp
Alkalisasi pada serat alam adalah metode yang telah digunakan.. untuk menghasilkan serat yang berkualitas
pengolahan produk pisang yang dilakukan pada pisang aroma Mahkota di desa. Gesing, Kecamatan Kandangan, Kabupaten
Set elah pelat ihan selesai, Anda diharapkan dapat m enjawab... Tata cara
2011.. melimpahkan segala Karunia-Nya, sehingga dengan segala keterbatasan yang ada baik waktu, tenaga dan pikiran yang di miliki penyusun. Akhirnya penyusun dapat
Skripsi dengan judul “Perbandingan Jumlah Total Bakteri Feses Sapi Bali Menurut Tingkat Kedewasaan dan Tipe Pemeliharaannya ” diajukan sebagai salah satu syarat
“ Pertama Setiap mahasiswa berhak menjadi anggota perpustakaan dengan cara mendaftar dan mengisi formulir yang telah disiapkan oleh kepala perpustakaan dengan
Pemilu dilaksanakan dengan perangkat peraturan yang mendukung kebebasan dan kejujuran, sehingga dengan adanya undang-undang yang lebih memberi kesempatan kebebasan
