KARAKTERISASI MOLEKULER TANAMAN PADI
NIPPONBARE TRANSGENIK
35S::OsERA
1 YANG
TOLERAN KEKERINGAN
SISKA KARTIKA
DEPARTEMEN BIOKIMIA
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI SKRIPSI DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA*
Dengan ini saya menyatakan bahwa skripsi berjudul Karakterisasi Molekuler Tanaman Padi Nipponbare Transgenik 35S::OsERA1 yang Toleran Kekeringan adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Penelitian ini didanai oleh BB-Biogen atas nama Dr. Tri Joko Santoso, S.P, M.Si. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir skripsi ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, September 2013
Siska Kartika
ABSTRAK
SISKA KARTIKA. Karakterisasi Molekuler Tanaman Padi Nipponbare Transgenik 35S::OsERA1 yang Toleran Kekeringan. Dibimbing oleh POPI ASRI KURNIATIN dan BUDI SANTOSA.
Kekeringan merupakan salah satu faktor yang mengakibatkan pertumbuhan dan perkembangan tanaman padi terhambat. Salah satu cara untuk mengatasi masalah tersebut adalah dengan penggunaan tanaman padi transgenik yang toleran kekeringan. Keberhasilan transformasi gen OsERA1 pada tanaman padi genotip Nipponbare memerlukan konfirmasi lebih lanjut mengenai toleransi terhadap cekaman kekeringan. Penelitian ini bertujuan mendapatkan genotip padi Nipponbare transgenik OsERA1 toleran kekeringan, mengetahui jumlah salinan gen, dan membandingkan biji padi isi dan hampa dengan kontrolnya. Evaluasi toleransi kekeringan menggunakan metode Standard System Evaluation for Rice
(IRRI 1996). Hasil evaluasi diperoleh semua genotip transgenik toleran terhadap kekeringan. Hasil karakterisasi molekuler dengan hibridisasi southern
menunjukkan tanaman padi transgenik memiliki 1-3 kopi transgen. Hasil cekaman kekeringan menghambat pertumbuhan tanaman, yaitu menurunkan jumlah biji isi dan meningkatkan kehampaan. Genotip 18.5.4 dan 6.4.10 dinilai toleran terhadap kekeringan. Genotip 18.5.4 memiliki jumlah biji isi per malai 36 biji dan genotip 6.4.10 memiliki persentase biji hampa 38.03%.
Kata kunci: gen OsERA1, hibridisasi southern, kekeringan, padi transgenik
ABSTRACT
SISKA KARTIKA. Molecular Characterization of Drought Tolerant Nipponbare Transgenic 35S::OsERA1 Rice Plants. Supervised by POPI ASRI KURNIATIN and BUDI SANTOSA.
Drought is one of the factors that resulted in the growth and development of rice plants inhibited. One way to overcome this problem is use of transgenic rice plants that are tolerant to drought. The success of the transformation using
OsERA1 gene in Nipponbare genotype rice requires further confirmation of tolerance to drought stress. The aim of this study was to obtain transgenic Nipponbare genotype rice contain gene 35S::OsERA1 drought tolerant, know the number of gene copies, and compare the contents and empty seeds with control. The evaluation of drought the tolerance was performed using Standard System Evaluation for Rice method (IRRI 1996). All tested transgenic genotypes were tolerant to drought. Molecular characterization using southern hybridization showed that transgenic rice plants had 1-3 copies of transgen. Drought stress inhibited plant growth, which lowers the amount of seed and increasing the void content. 18.5.4 and 6.4.10 genotypes are considered tolerant to drouht. 18.5.4 genotype have the number of grains per panicle contents of 36 seeds and 6.4.10 genotype have a 38.03% percentage of empty seeds.
Skripsi
sebagai salah satu syarat untuk memperoleh gelar Sarjana Sains
pada
Departemen Biokimia
KARAKTERISASI MOLEKULER TANAMAN PADI
NIPPONBARE TRANSGENIK
35S::OsERA
1 YANG
TOLERAN KEKERINGAN
SISKA KARTIKA
DEPARTEMEN BIOKIMIA
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM INSTITUT PERTANIAN BOGOR
Judul Skripsi : Karakterisasi Molekuler Tanaman Padi Nipponbare Transgenik
35S::OsERA1 yang Toleran Kekeringan Nama : Siska Kartika
NIM : G84090038
Disetujui oleh
Popi Asri Kurniatin, S.Si.,Apt., M.Si Pembimbing I
Dr Ir Budi Santosa, MP Pembimbing II
Diketahui oleh
Dr Ir I Made Artika, MAppSc Ketua Departemen
PRAKATA
Puji dan syukur penulis panjatkan kepada Allah SWT atas rahmat dan karunia-Nya sehingga karya ilmiah ini berhasil diselesaikan, serta shalawat dan salam semoga tercurahkan pada Rasulullah SAW. Karya ilmiah yang berjudul
“Karakterisasi Molekuler Tanaman Padi Nipponbare Transgenik 35S::OsERA1 yang Toleran Kekeringan” ini ditujukan sebagai salah satu syarat untuk memperoleh gelar sarjana sains di Departemen Biokimia yang dilaksanakan dari bulan Januari sampai dengan Juni 2013 di Laboratorium Biologi Molekuler dan Rumah kaca, Balai Besar Penelitian dan Pengembangan Bioteknologi dan Sumberdaya Genetik Pertanian, Cimanggu-Bogor.
Penulis mengucapkan terima kasih kepada Popi Asri Kurniatin, S.Si., Apt., M.Si selaku pembimbing I dan Dr Ir Budi Santosa, MP selaku pembimbig II atas bimbingan dan arahan yang telah diberikan. Terima kasih penulis juga sampaikan kepada Dr Tri Joko Santoso SP.M.Si, Bapak Iman, Bapak Unang, Mira Sitepu, dan teman-teman semuanya yang senantiasa memberikan bimbingan, ilmu, dorongan, inspirasi, motivasi, dan arahan, serta kepada ayah, ibu, serta seluruh keluarga, atas segala doa dan kasih sayangnya.
Penulis menyadari masih banyak kekurangan dalam karya ilmiah ini. Oleh karena itu, penulis mengharapkan kritik dan saran yang membangun untuk perbaikan di masa mendatang. Semoga karya ilmiah ini bermanfaat.
Bogor, September 2013
DAFTAR ISI
DAFTAR TABEL vi
DAFTAR GAMBAR vi
DAFTAR LAMPIRAN vi
PENDAHULUAN 1
METODE 2
Waktu dan Tempat Penelitian 2
Bahan 2
Alat 3
Prosedur Penelitian 3
HASIL 5
PEMBAHASAN 10
SIMPULAN DAN SARAN 14
Simpulan 14
Saran 15
DAFTAR PUSTAKA 15
LAMPIRAN 17
DAFTAR TABEL
1 Klasifikasi tanggap tanaman terhadap kekeringan (daun menggulung
pada fase vegetatif) 3
2 Klasifikasi tanggap tanaman terhadap kekeringan (daun mengering
pada fase vegetatif) 4
3 Klasifikasi tanggap kesembuhan tanaman setelah perlakuan pemulihan 4 4 Skor hasil pengujian daun menggulung, daun mengering, dan skor
kesembuhan tanaman padi Nipponbare transgenik (generasi T2) dan tanaman Nipponbare kontrol dengan perlakuan kekeringan pada fase
vegetatif 6
5 Hasil analisis dan pola integrasi gen sisipan (OsERA1) pada generasi kedua (T2) pada kultivar Nipponbare menggunakan gen hptII untuk PCR dan pelacak hpt untuk hibridisasi southern 8 6 Jumlah biji isi dan biji hampa tanaman padi Nipponbare transgenik dan
Nipponbare kontrol setelah perlakuan kekeringan 9
DAFTAR GAMBAR
1 Penampilan tanaman padi Nipponbare transgenik generasi T2 dan
Nipponbare kontrol setelah uji kekeringan. 6
2 Hasil amplifikasi PCR beberapa tanaman padi Nipponbare transgenik
menggunakan primer hptII. 7
3 Hasil amplifikasi PCR beberapa tanaman padi Nipponbare transgenik
menggunakan primer OsERAI-R. 7
4 Profil hibridisasi southern 16 genotip padi Nipponbare transgenik. 8
DAFTAR LAMPIRAN
1 Selisih bobot tanah per hari pada perlakuan cekaman kekeringan 17 2 Hasil pengukuran konsentrasi dan kemurnian DNA daun padi 18 3 Hasil amplifikasi PCR tanaman padi transgenik menggunakan primer
OsERAI-R 19
4 Hasil amplifikasi PCR tanaman padi transgenik menggunakan primer
hptII 20
PENDAHULUAN
Tanaman padi (Oryza sativa L.) merupakan penghasil beras dan menjadi salah satu komoditas pangan utama. Beras merupakan sumber utama gizi dan energi bagi lebih dari 90% penduduk Indonesia (BPS 2011). Saat ini, penyediaan pangan terutama beras menjadi prioritas utama pembangunan nasional untuk memenuhi kebutuhan penduduk Indonesia (Abdullah et al. 2007; Litbang Pangan 2012). Akan tetapi, usaha akan pemenuhan kebutuhan padi tersebut sering terhambat oleh beberapa kendala, di antaranya adalah cekaman abiotik. Cekaman abiotik yang umum dialami oleh tanaman padi adalah cekaman terhadap kekeringan (Rachmawati 2006; Pratama 2010). Pemanasan global menyebabkan berkurangnya ketersediaan air sehingga terjadi kekeringan yang berpengaruh dalam peningkatan hasil produksi tanaman padi (Suardi 2002; Makarim 2009).
Kekeringan merupakan salah satu faktor yang berdampak terhadap produksi tanaman. Pada tanaman padi, kekeringan mengakibatkan pertumbuhan dan perkembangan tanaman terhambat. Akibat kekeringan tanaman dapat kehilangan hasil sampai 50% (Jongdee et al. 2006). Hasil penelitian di rumah kaca menunjukkan terjadi penurunan hasil padi rata-rata 52.3% pada lingkungan tercekam kekeringan dibanding padi yang tumbuh pada lingkungan normal (Sammaullah dan Darajat 2001).
Usaha untuk meningkatkan produktivitas tanaman padi toleran kekeringan dapat dilakukan melalui rekayasa genetika. Bioteknologi atau teknologi molekuler memiliki peranan penting dalam menghasilkan kultivar dengan sifat unggul baru. Penggunaan varietas unggul merupakan cara paling ekonomis untuk mengatasi masalah kekeringan yang diharapkan dapat menjaga produksi dan kestabilan hasil padi (Soedarini & Patricia 2006). Hal yang penting dalam pengembangan ini adalah memahami mekanisme toleransi kekeringan karena karakter kekeringan dikendalikan oleh banyak gen. Gen-gen tersebut ada yang ekspresinya tergantung asam absisat (ABA) dan ada yang tidak (Yamaguchi-Shinozaki dan Shinozaki 2005).
Asam absisat memiliki peranan penting dalam proses selular, di antaranya perkembangan biji, dormansi, germinasi, pertumbuhan vegetatif, penutupan stomata, dan respon terhadap cekaman. Asam absisat merupakan fitohormon yang berperan di dalam membuka dan menutupnya stomata untuk mengurangi kehilangan air pada tumbuhan saat terjadi transpirasi. Sejumlah gen pada tanaman telah diketahui dapat meningkatkan dan menurunkan respon terhadap fitohormon, terutama hormon asam absisat. Salah satunya adalah gen Enhanced Response to ABA1 (ERA1) yaitu gen yang berperan dalam meningkatkan sensitifitas sel penjaga pada stomata terhadap asam absisat. Gen ERA1 pertama kali diisolasi dari tanaman model Arabidopsis thaliana. Gen ERA1 mengkode sub unit β dari enzim farnesiltransferase yang berperan dalam respon tanaman terhadap hormon asam absisat (Pei et al. 1998; Wang et al. 2005). Padi mutan era1 diidentifikasi berdasarkan pada peningkatan dormansi biji dan kadar asam absisat yang sangat sedikit. Padi mutan era1 menunjukkan adanya peningkatan toleransi terhadap cekaman kekeringan melalui pengurangan intensitas membuka stomata dan berkurangnya tingkat layu tanaman (Ziegeholfer et al. 2000).
2
pada kondisi tercekam dengan kondisi optimal atau normal. Hall (1993) menyatakan bahwa toleransi terhadap kekeringan dapat diukur dari produktivitas relatif antara genotip yang satu dengan genotip yang lain pada saat mengalami cekaman kekeringan yang sama.
Laboratorium Biologi Molekuler, Balai Besar Penelitian dan Pengembangan Bioteknologi dan Sumberdaya Genetik Pertanian (BB-BIOGEN) sedang mengembangkan padi yang toleran terhadap cekaman abiotik berupa kekeringan dengan melakukan rekayasa genetik terhadap gen ERA1. Gen ERA1 dari tanaman padi (Oryza sativa-ERA1 yang kemudian disingkat OsERA1) sebelumnya telah berhasil diklon ke dalam vektor kloning pGEM-T easy oleh Peneliti di Laboratorium Biologi Molekuler BB-BIOGEN. Selain itu, gen OsERA1 juga telah berhasil disisipkan pada pCAMBIA 1301 yang kemudian ditransformasi ke dalam kalus padi Nipponbare dengan menggunakan bantuan vektor Agrobacterium tumefaciens dan promoter 35S sehingga terbentuk tanaman padi Nipponbare transgenik OsERA1. Penelitian juga telah dilakukan terhadap tanaman padi Nipponbare transgenik OsERA1 generasi T1 yang toleran terhadap cekaman kekeringan. Oleh karena itu, perlu dilakukan penelitian lanjutan terhadap gen
OsERA1 pada generasi T2 dengan melakukan uji toleransi kekeringan dan
southern blot untuk mengetahui kestabilan gen OsERA1 di dalam tanaman padi Nipponbare transgenik tersebut.
Penelitian ini bertujuan mendapatkan genotip padi Nipponbare transgenik generasi T2 yang mengandung gen OsERA1 yang toleran kekeringan, untuk mengetahui jumlah salinan gen OsERA1 pada genotip padi Nipponbare transgenik, dan membandingkan biji isi dan biji hampa antara Nipponbare transgenik dengan Nipponbare kontrol. Manfaat penelitian adalah genotip padi Nipponbare transgenik OsERA1 yang toleran kekeringan dapat digunakan sebagai sumber plasma nutfah padi dan dapat digunakan sebagai tetua persilangan untuk merakit varietas unggul padi.
METODE
Waktu dan Tempat Penelitian
Penelitian dilaksanakan dari Januari - Juni 2013. Tempat pelaksanaannya di rumah kaca dan laboratorium Biologi Molekuler, Balai Besar Penelitian dan Pengembangan Bioteknologi dan Sumberdaya Genetik Pertanian, Jalan Tentara Pelajar No.3A, Cimanggu-Bogor.
Bahan
Bahan-bahan yang digunakan dalam penelitian antara lain benih padi Nipponbare transgenik dan Nipponbare kontrol, nitrogen cair, bufer ekstraksi,
RNase, kalium asetat, natrium asetat, isopropanol dingin, primer hpt forward, primer hpt reverse, 10x bufer PCR, larutan MgCl2, dNTPs, enzim Taq DNA polimerase, ddH2O, agarosa, 1x bufer TAE, marker 1 kb ladder, enzim restriksi
EcoRI, plasmid pCambia OsERA1, Digoxigenin-11-dUTP (DIG-11-dUTP), 5x
3 larutan 20x SSC, larutan washing, larutan antibodi, larutan developer, larutan fixer, substrat CDP-Star, dan Amersham Hyperfilm.
Alat
Alat yang digunakan dalam penelitian meliputi, mikropipet, tabung
eppendorf (0.5 mL, 1.5 mL, dan 2 mL), spin, ice maker, coolbox, waterbath, inkubator, stirer, autoklaf, vorteks, pH meter, oven, sentrifus, neraca analitik Kern
770, nanodrop Thermo Fisher Scientific 2009, PCR DNA Engine Tetrad Peltier Thermal Cycler 225, chemidoc gel system, microwave, shaker, dan tangki elektroforesis.
Prosedur Penelitian
Analisis Tanaman Padi Transgenik terhadap Kekeringan
Benih padi Nipponbare kontrol dan Nipponbare transgenik ERA 6.4, 11.7, 18.2, 18.5, dan 18.8 (generasi T2) masing-masing sebanyak 30 biji dikecambahkan pada cawan petri yang telah dilapisi kertas saring dan diberi air secukupnya. Benih padi yang telah tumbuh dipindahkan ke dalam media tanah dalam bak plastik untuk bibit di rumah kaca, kemudian dipindahkan ke dalam ember yang berisi 8 kg tanah, 4 liter air, dan 2 gram pupuk NPK dengan 1 tanaman per ember. Tanaman padi dikelompokkan menjadi tiga kelompok yaitu tanaman padi Nipponbare kontrol tanpa perlakuan kekeringan, tanaman padi Nipponbare kontrol dengan perlakuan kekeringan, dan tanaman padi Nipponbare transgenik dengan perlakuan kekeringan. Penyiraman dilakukan satu atau dua hari sekali. Tanaman Nipponbare transgenik yang dipindahkan ke dalam ember berjumlah 43 tanaman yang terdiri atas 18 ulangan tanaman dari genotip ERA 6.4, 2 ulangan tanaman dari genotip ERA 11.7, 8 ulangan tanaman dari genotip ERA
18.2, 11 ulangan tanaman dari genotip ERA 18.5, dan 4 ulangan tanaman dari genotip ERA 18.8.
Perlakuan cekaman kekeringan dilakukan pada saat tanaman akan memasuki fase berbunga dengan dihentikan penyiramannya. Selama dilakukan perlakuan kekeringan diambil beberapa sampel tanah untuk mengetahui kadar airnya. Pengamatan respon tanaman dilakukan dengan pemberian skoring berdasarkan gejala daun menggulung dan gejala daun mengering. Setelah itu, dilakukan pemulihan tanaman dengan cara menyiram. Tanaman yang masih tumbuh kemudian dilakukan penilaian kesembuhan. Skoring dilakukan mengikuti metode Standard Evaluation System for Rice (IRRI 1996, Silitonga et al. 2003) seperti tertera pada Tabel 1, 2, dan 3:
Tabel 1 Klasifikasi tanggap tanaman terhadap kekeringan (daun menggulung pada fase vegetatif)
Skor/Skala Keterangan
0 Daun-daun sehat
3 Daun-daun melipat (berbentuk huruf V)
5 Daun betul-betul kuncup (membentuk huruf U)
7 Ujung-ujung daun bersentuhan (bentuk O)
4
Tabel 2 Klasifikasi tanggap tanaman terhadap kekeringan (daun mengering pada fase vegetatif)
Skor/Skala Keterangan
0 Tidak ada gejala
1 Ujung daun sedikit mengering
3 Ujung-ujung daun mengering sampai ¼ panjang pada hampir
semua daun
5 ¼ sampai ½ dari semua daun betul-betul kering
7 Lebih dari 2/3 dari semua daun betul-betul kering
9 Tanaman mati
Tabel 3 Klasifikasi tanggap kesembuhan tanaman setelah perlakuan pemulihan
Skor/Skala Keterangan
DNA genomik dari tanaman padi diekstraksi dengan metode isolasi DNA mengikuti metode Dellaporta et al. (1983) yang telah dimodifikasi. Metode ini dilakukan melalui tiga tahap yaitu ekstraksi DNA, pemurnian DNA, dan pemekatan DNA. Preparasi ekstrak sel dimulai dengan daun padi digerus menggunakan liquid nitrogen (LN) sampai halus, kemudian dimasukkan ke dalam tabung korning 50 mL, dan ditambahkan bufer ekstraksi serta larutan 20% SDS. Pemurnian dan pemekatan DNA dilakukan dengan penambahan kalium asetat 5M, natrium asetat 3M, RNase, dan isopropanol dingin. Pelet DNA dilarutkan dengan bufer TE (Tris EDTA) 0.1x sebanyak 50 μL. Selanjutnya kemurnian dan konsentrasi DNA diukur dengan nonodrop (Thermo Fisher Scientific 2009).
Amplifikasi PCR
Hasil isolasi DNA yang telah ditentukan konsentrasinya selanjutnya diamplifikasi dengan mesin PCR DNA Engine Tetrad Peltier Thermal Cycler. Analisis molekuler dengan PCR di tahap awal menggunakan primer spesifik hpt
(hygromycin phospho transferase) yang mengamplifikasi bagian gen higromisin yang merupakan marka seleksi pada plamid pCAMBIA yang disisipkan. Total
5 Selain itu, untuk deteksi transgen secara molekuler juga digunakan primer spesifik untuk gen OsERA1 yang dikombinasikan dengan primer untuk promoter, yaitu dengan kombinasi pasangan primer 35S-F/OsERA1-R. Amplifikasi PCR
dilakukan dengan total volume reaksi yaitu 20 μL menggunakan program ERA1 -N50 dengan siklus pradenaturasi pada 95oC selama 3 menit, denaturasi pada 94oC selama 1 menit 30 detik, penempelan primer pada 55oC selama 1 menit 30 detik, dan pemanjangan primer pada 72oC selama 2 menit. Siklus kedua sama seperti siklus pertama, sampai terjadi 34 siklus. Pemanjangan primer terakhir dilakukan pada 72oC selama 10 menit dan inkubasi pada 15oC selama 15 menit.
Elektroforesis DNA
Produk hasil PCR selanjutnya diseparasi menggunakan gel agarosa 1% di dalam 1x bufer TAE (Tris HCl-asam asetat-EDTA). Pewarnaan gel dilakukan dengan menambahkan GelRed (BIOTIUM) pada loading dye yang akan dimasukkan ke dalam sumur bersama produk hasil PCR. Kemudian pita-pita DNA divisualisasi menggunakan chemidoc gel system.
Hibridisasi Southern
Hibridisasai southern dilakukan berdasarkan metode Trijatmiko et al. (2011) menggunakan pelabelan Digoxigenin -11-dUTP (DIG-11-dUTP). Kegiatan terdiri atas beberapa tahap, yaitu pemotongan DNA menggunakan enzim restriksi
EcoRI, pelabelan marker, transfer DNA yang sudah dipotong dengan enzim restriksi ke membran Hybond-N+, pelabelan pelacak, hibridisasi membran dengan pelacak yang telah dilabel, dan deteksi sinyal dengan larutan CDP star.
Pengamatan Biji Hasil Perlakuan Kekeringan
Tanaman padi Nipponbare transgenik dan Nipponbare kontrol setelah uji kekeringan diamati jumlah biji isi dan jumlah biji hampa untuk mengetahui produktivitasnya.
HASIL
Analisis Tanaman Padi Transgenik terhadap Kekeringan
Bibit tanaman padi Nipponbare transgenik yang berasal dari lima genotip dilakukan uji terhadap cekaman kekeringan selama 7-10 hari tertera pada Tabel 4. Tanaman Nipponbare kontrol dengan perlakuan kekeringan memiliki skor kesembuhan tanaman 9 yaitu tanaman mati semua, sedangkan semua genotip tanaman padi Nipponbare transgenik memiliki skor kesembuhan 1 berarti tanaman masih dapat tumbuh dengan baik. Tanaman padi Nipponbare transgenik memiliki skor daun menggulung 9 yaitu daun menggulung ketat, sedangkan nilai skor daun mengering bervariasi berkisar antara 3-7 berarti tanaman padi Nipponbare transgenik generasi T2 masih mengalami segregasi.
6
yang disertai dengan pertumbuhan daun yang segar dan malai-malai bermunculan. Pertumbuhannya hampir menyerupai tanaman Nipponbare kontrol normal dengan pengairan optimal, sedangkan tanaman Nipponbare kontrol dengan perlakuan kekeringan menunjukkan tidak terbentuknya tanda-tanda pertumbuhan karena batang tanaman rebah, daun kering, dan tidak munculnya malai.
Tabel 4 Skor hasil pengujian daun menggulung, daun mengering, dan skor kesembuhan tanaman padi Nipponbare transgenik (generasi T2) dan tanaman Nipponbare kontrol dengan perlakuan kekeringan pada fase vegetatif
Genotip Jumlah Tanaman
Fase Vegetatif
Kesembuhan Daun
Menggulung
Daun Mengering Nipponbare
Perlakuan 5 9 9 9
ERA 6.4 18 9 3-7 1
ERA 11.7 2 9 7 1
ERA 18.2 8 9 3-7 1
ERA 18.5 11 9 5-7 1
ERA 18.8 4 9 7-7 1
Keterangan: skor 1 = 90-100% tanaman sembuh, skor 3 = 70-89% tanaman sembuh, skor 5 = 40-69% tanaman sembuh, skor 7 = 20-39% tanaman sembuh, skor 9 = 0-19% tanaman sembuh.
Gambar 1 Penampilan tanaman padi Nipponbare transgenik generasi T2 dan Nipponbare kontrol setelah uji kekeringan
Analisis Molekuler dengan PCR
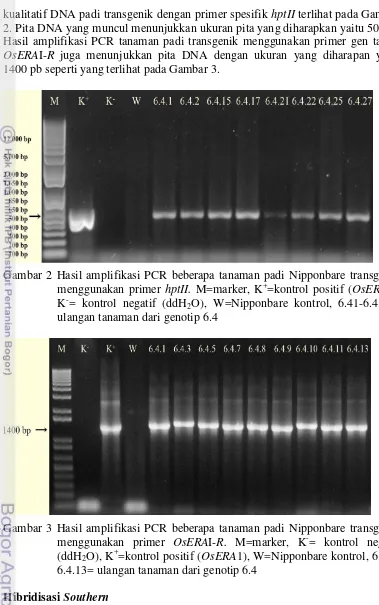
7 kualitatif DNA padi transgenik dengan primer spesifik hptII terlihat pada Gambar 2. Pita DNA yang muncul menunjukkan ukuran pita yang diharapkan yaitu 500 pb. Hasil amplifikasi PCR tanaman padi transgenik menggunakan primer gen target
OsERAI-R juga menunjukkan pita DNA dengan ukuran yang diharapan yaitu 1400 pb seperti yang terlihat pada Gambar 3.
Gambar 2 Hasil amplifikasi PCR beberapa tanaman padi Nipponbare transgenik menggunakan primer hptII. M=marker, K+=kontrol positif (OsERA1), K-= kontrol negatif (ddH2O), W=Nipponbare kontrol, 6.41-6.4.27= ulangan tanaman dari genotip 6.4
Gambar 3 Hasil amplifikasi PCR beberapa tanaman padi Nipponbare transgenik menggunakan primer OsERAI-R. M=marker, K-= kontrol negatif (ddH2O), K+=kontrol positif (OsERA1), W=Nipponbare kontrol, 6.4.1-6.4.13= ulangan tanaman dari genotip 6.4
Hibridisasi Southern
8
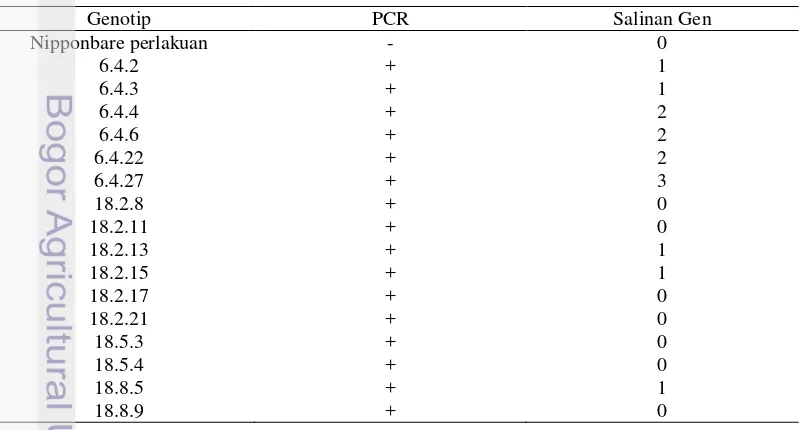
beberapa genotip memiliki 1-3 salinan transgen seperti terlihat pada Gambar 4. Genotip 6.4.2, 6.4.3, 18.2.13, 18.2.15, dan 18.8.5 memiliki 1 salinan transgen, genotip 6.4.4, 6.4.6, dan 6.4.22 memiliki 2 salinan transgen, dan genotip 6.4.27 memiliki 3 salinan transgen, sedangkan genotip 18.2.8, 18.2.11, 18.2.17, 18.2.21, 18.5.3, 18.5.4, dan 18.8.9 tidak menghasilkan salinan transgen, begitupun dengan genotip kontrol.
Gambar 4 Profil hibridisasi southern 16 genotip padi Nipponbare transgenik. Lingkaran-lingkaran pada gambar menunjukkan pita DNA hasil
southern
Tabel 5 juga menggambarkan hasil analisis PCR dan southern. Hasil PCR pada 16 sampel genotip Nipponbare transgenik menggunakan primer hptII
menunjukkan hasil positif yang ditandai dengan adanya pita DNA pada gel agarosa. Namun, hasil hibridisasi southern menggunakan pelacak hpt
menunjukkan jumlah salinan gen yang bervariasi. Hal ini menunjukkan bahwa masih terjadi segregasi pada pada tanaman padi Nipponbare transgenik generasi T2.
Tabel 5 Hasil analisis dan pola integrasi gen sisipan (OsERA1) pada generasi kedua (T2) pada kultivar Nipponbare menggunakan gen hptII untuk PCR dan pelacak hpt untuk hibridisasi southern
9
Analisis Jumlah Biji Padi setelah Perlakuan Kekeringan
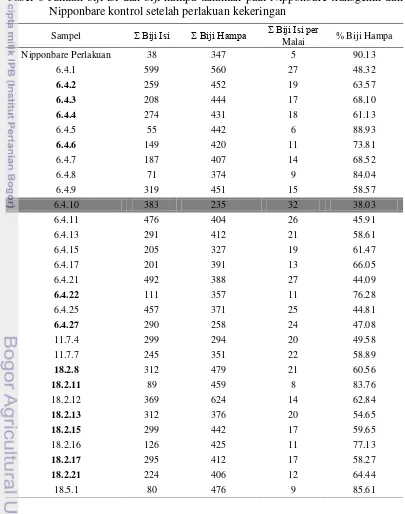
Produktivitas tanaman padi yang baik salah satunya dapat dilihat dari banyaknya biji yang dihasilkan. Jumlah biji isi per malai tanaman padi Nipponbare transgenik generasi T2 pada umumnya lebih tinggi dibandingkan Nipponbare kontrol dengan perlakuan kekeringan, dan genotip 18.5.4 memiliki jumlah biji isi per malai sebanyak 36 biji yang lebih tinggi dari Nipponbare kontrol tanpa perlakuan kekeringan yaitu 34 biji seperti terlihat pada Tabel 6. Tanaman padi Nipponbare transgenik juga menunjukkan persentase biji hampa lebih rendah dari Nipponbare kontrol dengan perlakuan kekeringan, namun tidak lebih tinggi dari Nipponbare kontrol tanpa perlakuan kekeringan.
Tabel 6 Jumlah biji isi dan biji hampa tanaman padi Nipponbare transgenik dan Nipponbare kontrol setelah perlakuan kekeringan
10
Analisis Tanaman Padi Transgenik terhadap Kekeringan
Penggunaan varietas padi yang toleran terhadap kekeringan merupakan cara paling ekonomis untuk mengatasi masalah kekeringan yang diharapkan dapat menjaga produksi dan kestabilan hasil padi. Hasil uji kekeringan telah dilakukan selama 7-10 hari terhadap 43 tanaman padi Nipponbare transgenik dan disertakan tanaman padi Nipponbare kontrol sebagai pembanding. Kadar air pada saat dilakukan cekaman kekeringan menunjukkan semua tanaman berada dalam kondisi tercekam karena kadar air tanah sebesar 18.45% (Lampiran 5) yang nilainya lebih rendah dari pF (kemampuan partikel tanah mengikat air) yaitu sebesar 18.84% (titik layu permanen). Hasil pengamatan daun menggulung fase vegetatif menunjukkan bahwa semua genotip memiliki skor 9 (daun menggulung ketat) (Tabel 4). Hasil pengamatan daun mengering pada fase vegetatif menunjukkan bahwa genotip 6.4, 11.7, 18.2, 18.5, dan 18.8 memiliki skor 3-7 (ujung daun mengering sampai ¼ panjang pada hampir semua daun – lebih dari 2/3 dari semua daun betul-betul kering) (Tabel 4), sedangkan tanaman Nipponbare kontrol dengan perlakuan kekeringan memiliki skor 9 (tanaman mati) (Tabel 4). Variasi skor yang terjadi pada masing-masing genotip tanaman padi Nipponbare transgenik dapat disebabkan adanya segregasi gen OsERA1 dan kemungkinan menyisipkan gen pada tempat yang berbeda.
11 masih dapat tumbuh. Selain itu, varietas tanaman padi Nipponbare kemungkinan memiliki respon yang cukup baik dalam menanggapi kekeringan, sedangkan pada tanaman padi Nipponbare transgenik menunjukkan bahwa gen ERA1 yang disisipkan ke dalam tanaman dapat bekerja dengan baik sehingga tanaman toleran terhadap kekeringan. Penampilan tanaman padi Nipponbare transgenik dan Nipponbare kontrol yang diberikan perlakuan kekeringan terlihat berbeda setelah uji kekeringan. Tanaman transgenik terlihat dapat tumbuh menjadi segar kembali setelah uji kekeringan dibandingkan dengan tanaman kontrol (Gambar 1).
Gen ERA1 yang diintroduksikan ke dalam genom tanaman berperan dalam meningkatkan sensitifitas dari sel penjaga pada stomata terhadap asam absisat. Respon asam absisat pada tanaman saat terjadi cekaman kekeringan akan berlangsung secara cepat sehingga dapat menyebabkan terjadinya penutupan stomata. Hormon asam absisat akan meningkatkan sinyal cascade pada sel penjaga sehingga menghasilkan penutupan stomata dengan menghambat pembukaan stomata (Qiagen 2003). Hal inilah yang menyebabkan terjadinya penutupan stomata pada tanaman padi Nipponbare transgenik ketika kekurangan air akibat cekaman kekeringan sehingga masih dapat melangsungkan siklus hidupnya.
Analisis Molekuler dengan PCR
Kestabilan integrasi transgen dalam tanaman transgenik dapat diketahui dengan melakukan uji kestabilan gen. Uji stabilitas gen dilakukan dengan mendeteksi transgen pada turunan tanaman transgenik menggunakan teknik
polymerase chain reaction (PCR). Individu-individu putatif transgenik dari genotip-genotip Nipponbare-OsERA1 generasi T2 yang toleran kekeringan kemudian dianalisis secara molekuler untuk mengetahui keberadaan transgen
OsERA1. Peningkatan toleransi terhadap kekeringan berkaitan dengan adanya ekspresi gen faktor transkripsi OsERA1 yang telah terintegrasi ke dalam tanaman. Oleh karena itu, tanaman putatif transgenik masih perlu diidentifikasi secara molekuler untuk mengkonfirmasi keberadaan dan integrasi gen yang diintroduksikan di dalam genom tanaman.
Analisis integrasi awal dilakukan menggunakan primer spesifik hptII
(hygromycin phospho transferase) yang mengamplifikasi bagian gen higromisin. Gen higromisin merupakan marka seleksi yang terdapat pada plasmid pCAMBIA1301 yang disisipkan. Primer hptII mengandung forward sequence: 5’
GAT TCC TTG CGG TCC GAA TG 3’, dan reverse sequence: 5’ TCC GAC CTG ATG CAG CTC TC 3’. Hasil PCR divisualisasi menggunakan elektroforesis gel agarosa 1% (80 volt, 45 menit). Kontrol positif dan kontrol negatif juga disertakan dalam proses PCR. Kontrol positif yang digunakan yaitu plasmid pCAMBIA-ERA1 dan kontrol negatif berupa air. Hasil visualisasi menunjukkan bahwa proses PCR berjalan dengan baik tanpa adanya kontaminasi, artinya 43 tanaman Nipponbare transgenik positif mengandung gen hptII. Tanaman padi yang positif PCR hptII ditandai dengan terbentuknya pita produk PCR berukuran 500 pb (Gambar 2, Lampiran 4a, 4b, 4c).
12
hptII. Primer OsERA1-R telah dikonstruk sebelumnya oleh Tim Peneliti Laboratorium Biologi Molekuler, BB-BIOGEN. Primer tersebut adalah primer spesifik terhadap gen ERA1 sehingga hanya genom tanaman yang tersisipi gen target yang dapat teramplifikasi. Tanaman Nipponbare transgenik yang tidak mengandung gen ERA1 tidak akan teramplifikasi karena tidak adanya sekuen spesifik terhadap primer yang digunakan. Hasil PCR divisualisasi menggunakan elektroforesis gel agarosa 1% menunjukkan adanya produk PCR berupa pita DNA tunggal yang berukuran sekitar 1400 pb (Gambar 3, Lampiran 3a, 3b). Hal ini menunjukkan bahwa semua tanaman Nipponbare transgenik positif mengandung gen ERA1 karena ukuran fragmen DNA tersebut sesuai dengan ukuran fragmen gen ERA1 yaitu sekitar 1400 pb yang terbentuk pada hasil digesti dari vektor rekombinan pGEM-ERA1 yang telah dilakukan dalam penelitian sebelumnya di Laboratorium Biologi Molekuler, BB-BIOGEN.
Tanaman Nipponbare transgenik yang terdeteksi positif PCR hptII dan
OsERA1 pada 43 sampel menunjukkan adanya hubungan dengan analisis terhadap kekeringan. Tanaman Nipponbare transgenik tersebut memiliki nilai kesembuhan tanaman dengan skor 1 (90-100% tanaman sembuh) (Tabel 4). Semua tanaman Nipponbare transgenik tumbuh kembali setelah uji kekeringan dengan adanya pertumbuhan batang, daun, dan malai. Hal ini menunjukkan bahwa tanaman Nipponbare transgenik memiliki gen ERA1 yang telah berhasil disisipkan ke dalam genom tanaman dan gen tersebut dapat berfungsi dengan baik karena toleran terhadap kekeringan sehingga tanaman Nipponbare transgenik dapat tumbuh dan melangsungkan siklus hidupnya.
Analisis Hibridisasi Southern
Metode hibridisasi southern merupakan salah satu cara untuk mengetahui informasi tentang integrasi dan jumlah kopi dari gen yang berhasil masuk ke dalam genom tanaman. Tanaman transgenik yang digunakan untuk analisis
southern yaitu sebanyak 16 sampel yang dipilih berdasarkan gejala tanaman layu dan kemampuannya yang lebih baik dalam menanggapi cekaman kekeringan, yaitu kemampuan dalam mempertahankan kandungan air di dalam tanah yang lebih lama selama proses cekaman (Lampiran 1). Semakin lama tanaman bertahan dalam kondisi tercekam menunjukkan tanaman semakin toleran terhadap kekeringan. Selain itu, pemilihan sampel juga dilakukan berdasarkan konsentrasi dan kemurnian DNA yang tinggi. Pemotongan pita-pita DNA dalam penelitian ini dilakukan menggunakan enzim restriksi EcoRI, dan pelacak hptII.
Analisis hibridisasi southern pada 16 sampel tanaman transgenik OsERA1 menunjukkan bahwa 1 tanaman membentuk 3 pita DNA, 3 tanaman membentuk 2 pita DNA, 5 tanaman membentuk 1 pita DNA, dan tanaman-tanaman lain tidak membentuk pita DNA pada X-ray film yang telah dipaparkan membran nilon
13 positif ketika di deteksi dengan PCR dan negatif ketika di deteksi dengan analisis
southern.
Hasil hibridisasi southern menunjukkan jumlah kopi gen hptII yang terintegrasi di dalam tanaman Nipponbare transgenik bervariasi, berkisar 1-3 kopi (Tabel 5). Pita-pita tersebut terbentuk karena DNA pada padi Nipponbare transgenik yang telah dipotong oleh enzim restriksi EcoRI memiliki sekuen hptII
dan dapat terhibridisasi oleh pelacak hptII. Variasi kemunculan pita dapat disebabkan oleh jumlah salinan gen yang menyisip dalam genom padi tidak sama. Adapun tanaman Nipponbare kontrol tidak menghasilkan pita DNA karena tidak mempunyai gen spesifik. Hal ini menunjukkan bahwa pelacak dan kondisi hibridisasi hanya dapat mendeteksi keberadaan transgen secara spesifik. Dalam penelitian ini dijumpai hubungan yang sistematis antara jumlah transgen yang dimiliki dengan tingkat toleransi kekeringan pada tanaman, yaitu pada genotip 6.4.2, 6.4.3, 18.2.13, 18.2.15, dan 18.8.5. Genotip-genotip tersebut memiliki 1 salinan transgen (Tabel 5) dan memiliki tingkat toleransi kekeringan yang lebih baik dibandingkan dengan genotip lain (Lampiran 1). Hal ini menunjukkan posisi integrasi dari transgen pada genom tanaman padi Nipponbare transgenik menentukan level toleransi terhadap kekeringan dan jumlah salinan transgen (Santosa 2008).
Semakin banyak jumlah salinan transgen pada genom tanaman maka akan semakin tinggi persentase pembungkaman gen (Tang et al. 2007). Pembungkaman gen merupakan salah satu fenomena yang menyebabkan terjadinya kegagalan dalam mengekspresikan gen dan terbentuknya dupleks RNA yang mengakibatkan terhalangnya proses translasi sehingga pembentukan protein terganggu (Malik 2005).
Analisis Jumlah Biji Isi dan Biji Hampa Tanaman Padi Nipponbare Transgenik
Teknik transformasi genetik dapat dikatakan berhasil jika ditandai dengan keberhasilan menyisipkan rangkaian gen yang diintroduksikan ke dalam genom tanaman sehingga dapat diekspresikan dan tetap terpelihara dalam seluruh pembelahan sel berikutnya. Oleh karena itu, salah satu upaya untuk menentukan gen tersebut dapat berfungsi dengan benar adalah dengan melihat fenotip tanaman padi tersebut (Santoso 2008). Keberhasilan teknik transformasi genetik dapat dilihat dari fenotip tanaman transgenik yang dibandingkan dengan fenotip tanaman kontrol.
Perlakuan cekaman kekeringan dilakukan pada umur tanaman 40 hari, yaitu pada saat fase vegetatif menuju fase generatif. Kekurangan air pada fase vegetatif menyebabkan pertumbuhan tanaman terhambat, sedangkan fase generatif merupakan fase pertumbuhan yang peka bagi tanaman padi terhadap kekeringan. Kekeringan pada fase tersebut dapat menyebabkan tanaman tidak berkembang atau tidak menghasilkan gabah. Penurunan hasil akibat defisit air setelah fase berbunga terjadi karena penurunan jumlah gabah isi dan meningkatnya kehampaan (Samaullah et al. 1996).
14
genotip tanaman padi Nipponbare transgenik 18.5.4 dinilai lebih toleran terhadap kekeringan. Kekurangan air pada genotip tersebut tidak terlalu berpengaruh terhadap jumlah biji yang dihasilkan. Genotip tanaman padi Nipponbare transgenik yang lain juga menunjukkan jumlah biji isi per malai lebih tinggi dari Nipponbare kontrol yang diberi perlakuan kekeringan dengan jumlah 5 biji per malai (Tabel 6). Penelitian ini mengharapkan hanya tanaman padi transgenik saja yang dapat berproduksi, namun pada Nipponbare kontrol yang diberi perlakuan kekeringan terlihat menghasilkan biji. Hal ini dapat disebabkan sebelum masuk masa perlakuan kekeringan, tanaman sudah memasuki fase pengisian biji sehingga masih dapat menghasilkan biji padi meskipun sedikit.
Hasil pengamatan menunjukkan gabah hampa tidak hanya dihasilkan pada tanaman dengan perlakuan kekeringan, namun juga pada perlakuan pengairan normal. Menurut Yoshida (1981) kondisi kekeringan pada periode 0-2 minggu menjelang pembungaan dapat menyebabkan kehampaan malai. Nipponbare kontrol dengan perlakuan kekeringan menunjukkan persentase biji hampa paling tinggi (90.13%) dibandingkan dengan semua tanaman padi Nipponbare transgenik (Tabel 6). Namun, tanaman Nipponbare transgenik tidak ada yang lebih rendah persentase biji hampanya dari Nipponbare kontrol normal (32.21%). Akan tetapi, tanaman padi Nipponbare transgenik genotip 6.4.10 memiliki persentase biji hampa 38.03% yang dinilai lebih toleran terhadap kekeringan karena persentasenya tidak jauh berbeda dengan Nipponbare kontrol normal. Biji hampa yang dihasilkan oleh tanaman padi Nipponbare transgenik dan Nipponbare kontrol dapat disebabkan perlakuan cekaman kekeringan terjadi pada fase reproduktif (Yoshida 1981) .
Jumlah biji isi dan biji hampa tanaman padi Nipponbare transgenik tidak semuanya lebih unggul dari tanaman Nipponbare kontrol, sehingga dapat dikatakan bahwa tidak semua fenotip dapat dibandingkan dengan jumlah salinan gen yang terintegrasi dalam genom tanaman padi Nipponbare transgenik. Hal ini terlihat pada genotip 18.5.4 meskipun dinilai lebih toleran terhadap kekeringan karena memiliki jumlah biji isi per malai lebih tinggi daripada Nipponbare kontrol tanpa kekeringan, namun tidak memiliki salinan kopi gen. Menurut Kolensik et al. (2004) penentuan fungsi gen dari fenotip yang diekspresikan akan menjadi sulit apabila ada dua atau lebih posisi T-DNA dalam satu genom. Gen tidak hanya elemen-elemen yang memisah dan menghasilkan pengaruh individu yang jelas, tetapi interaksi gen satu dengan yang lainnya memberikan fenotip yang berbeda secara menyeluruh (Sudharmawan 2009).
SIMPULAN DAN SARAN
Simpulan
15 isi per malai lebih tinggi dari Nipponbare kontrol dengan pengairan normal, yaitu 36 biji. Tanaman padi transgenik genotip 6.4.10 juga menunjukkan persentase biji hampa 38.03% yang tidak jauh berbeda dengan Nipponbare kontrol normal dengan persentase 32.21%. Genotip 6.4.10 dan 18.5.4 tersebut dinilai lebih toleran terhadap kekeringan.
Saran
Perlu dilakukan optimasi metode perlakuan kekeringan agar antara fase vegetatif, fase pembungaan, dan fase fase generatif dapat terkontrol dengan baik.
DAFTAR PUSTAKA
Abdullah A, S Ito, dan K Adhana. 2007. Estimate of Rice Consumption in Asian Countries and the World Towards 2050. Jepang: Tottori University.
Allen GJ, Y Murata, SP Chu, M Nafiei, JI Schroeder. 2002. Hypersensitivity of abscisic acid-induced cytosolic calcium increases in the Arabidopsis
farnesyltransferase mutant era1-2. The Plant Cell. 14: 1649-1662.
[BPS] Badan Pusat Statistik. 2011. Statistik Indonesia. Berita Resmi Statistik. Jakarta: BPS.
Dellaporta SI, Jonathan W, James BH. 1983. A plant DNA minipreparation: version II. Plant molecular Biology Reporter 1: 19-21.
Hall AE. 1993. Is dehydration tolerance relevant to genotypic difference in leaf senescence and crop adaptation to dry environments. In: Close TJ and Bray EA (Eds.). Plant responses to cellular dehydration during environmental stress. p: 110. The American Soc. Plant Pathologist. Maryland: Rockville.
[IRRI]. 2010. morfologi dan pertumbuhan padi. [terhubung berkala] http:// www. Knowledgebank. Irri. org/ morph/ morphology.htm [1 Maret 2013].
Jongdee B, Pantuwan G, Fukai S, Fischer K. 2006. Improving drought tolerance in rainfed lowland rice: AN example from Thailand. Agricultural Water Management (80): 225-240.
Kolesnik T, Szeverenyi I, Bachmann D, Kumar CS, Jiang S, Ramamoorthy R, Cai M, Ma ZG, Sundaresan V, Ramachandran S. 2004. Establishing an afficient Ac/Ds tagging system in rice: large-scale analysis of Ds flanking sequences.
Plant J. 37: 301-314.
Litbang Pangan. 2012. Repositori peningkatan pangan menuju 2020. [terhubung berkala]
http://pangan.litbang.deptan.go.id/index.php?bawaan=download/download_det ail&&id=35. (25 Agustus 2013).
Muliarta IGP, Sudantha LM, Santoso BB. 2012. Daya hasil dan penampilan fenotipik karakter kuantitatif galur-galur F2BC4 padi gogo beras merah. PG 5 Prosiding InSINas. 2012 Nov 29-30; Bandung, Indonesia. Bandung (ID): Insentif Ristek. hlm 1-7.
16
Pei Z, M Ghassemian, CM Kwak, P McCourt, JI Schroeder. 1998. Role of farnesyltransferase in ABA regulation of guard cell anion chanells and plant water loss. Science. 282: 287-290.
Pratama GS. 2010. Analisis Respon Toleransi Padi Nipponbare Transgenik Terhadap Salinitas Tinggi [Skripsi]. Bogor: Departemen Biokimia, FMIPA, Institut Pertanian Bogor.
Qiagen. 2003. GeneGlobe Pathways: Absicid acid. 2003. [terbuhung berkala]. http//:www.qiagen.com/pathways.aspx. (13 Januari 2013).
Rachmawati S. 2006. Studi Perkembangan Perbaikan Sifat Genetik Padi Menggunakan Transformasi Agrobacterium. Jurnal AgroBiogen. 2(1): 36-44. Roche. 2013. Troubleshoot Southern hibridization. [terhubung berkala].
https://www.rocheapplied-science.com/sis/lad/index.jsp?id=LA030400, (10 September 2013).
Samaullah MY, Sutaryo B, Taryat T. 1996. Peluang pemanfaatan genotip padi toleran kekeringan pada daerah terbatas sumber air. Seminar Nasional Masyarakat Konservasi Tanah dan Air. Universitas Brawijaya, Malang.
Sammaullah MJ, Darajat AA. 2001. Toleransi beberapa genotip padi gogo terhadap cekaman kekeringan. Penelitian Pertanian Tanaman Pangan. 20(1): 17-23.
Santosa TJ. 2008. Identifikasi begomovirus Indonesia pada tomat dan analisis diversitas genetik gen AV1 serta pemanfaatannya untuk pengembangan tanaman tahab virus [disertasi]. Bogor: Program Pascasarjana, Institut Pertanian Bogor.
Silitongga TS, Somantri IH, Daradjat AA, Kurniawan H. 2003. Panduan Sistem Karakterisasi dan Evaluasi Tanaman Padi. Departemen Pertanian. Badan Penelitian dan Pengembangan Pertanian. Komisi Nasional Plasma Nutfah. 58h. Soedarini B, Patricia N. 2006. Pangan transgenik: suatu pilihan yang dilematis.
Renai 2: 129-144.
Suardi D. 2002. Perakaran padi dalam hubungaannya dengan toleransi tanaman terhadap kekeringan dan hasil. Jurnal Litbang Pertanian. 21(3): 100-108. Sudharmawan AAK. 2009. Analisis segregasi persilangan varietas padi tahan dan
rentan terhadap cekaman kekeringan. Agroteksos 19: 109-114.
Trijatmiko KR, Olivia N, Slamet-Loedin IH, Kohli A. 2011. Molecular Analysis of Transgenic Plants. New York: Springer.
Wang Y, J Ying, M Kuzma, M Chalifoux, A Sample, C McArthur, T Uchaz, C Sarvas, J Wan, DT Dennis, P McCourt, Y Huang. 2005. Molecular Tailoring of Farnesylation for plant drought tolerance and yield protection. The Plant Journal. 43: 413-424.
Yamaguchi-Shinozaki K, Shinozaki K. 2005. Organization of cis-acting regulatory elements in osmotic and cold stress responsive promoters. Trends in Plant Science. 10: 88-94.
Yoshida S. 1981. Fundamental of rice crop science. IIRRI. Los Banos, Philippines. 269 p
Ziegelhoffer EC, LJ Medrano, EM Meyerowits. 2000. Cloning Arabidopsis WIGGUM gene identifies a role for farnesylation in meristem wdevelopment.
17
LAMPIRAN
Lampiran 1 Selisih bobot tanah per hari pada perlakuan cekaman kekeringan
18 sampel yang dicetak tebal merupakan sampel yang di southern
Lampiran 2 Hasil pengukuran konsentrasi dan kemurnian DNA daun padi
Sampel Konsentrasi (ng/μL) A260/280
Nipponbare kontrol 2124.3 1.87
19 lanjutan
18.5.1 1216.0 1.92
18.5.2 1088.8 1.93
18.5.3 1585.9 1.93
18.5.4 1180.4 1.91
18.5.5 932.2 1.94
18.5.6 990.7 1.93
18.5.7 1042.7 1.93
18.5.8 1876.1 1.91
18.5.10 1157.4 1.93
18.5.11 1943.3 1.91
18.5.15 1227.1 1.93
18.8.5 452.9 1.91
18.8.9 3279.0 1.91
18.8.13 3289.5 1.92
18.8.15 3249.6 1.86
Keterangan: Sampel yang dicetak tebal merupakan sampel yang di southern
Lampiran 3 Hasil amplifikasi PCR tanaman padi transgenik menggunakan primer
OsERAI-R
Keterangan: M=marker, K+=kontrol positif (OsERA1), K-=kontrol negatif (ddH2O), W=Nipponbare kontrol (tipe liar)
(a)
20
Lampiran 4 Hasil amplifikasi PCR tanaman padi transgenik menggunakan primer
hptII
Keterangan: M=marker, K+=kontrol positif (OsERA1), K-=kontrol negatif (ddH2O), W=Nipponbare kontrol (tipe liar)
(a)
(b)
21 Lampiran 5 Data kadar air tanah
Genotip Bobot tanah
(g)
Kadar air
(%)
Basah Kering
Nipponbare 8.33 6.78 18.6
18.2 8.30 6.78 18.3
Rata-rata 18.45
Contoh perhitungan:
Kadar air (%) =
× 100 %
=
× 100 %
22
RIWAYAT HIDUP
Penulis dilahirkan di Cianjur pada tanggal 20 Maret 1990 dari ayah bernama Elan Jaelani dan ibu bernama Teti Rohayati. Penulis merupakan anak pertama dari dua bersaudara. Tahun 2009 penulis menyelesaikan pendidikan Sekolah Menengah Atas di SMA Negeri 1 Cianjur, Jawa Barat dan pada tahun yang sama lolos seleksi masuk dan melanjutkan studi di Institut Pertanian Bogor melalui jalur Undangan Seleksi Masuk IPB (USMI). Di IPB penulis mengambil mayor Departemen Biokimia, Fakultas Matematika dan Ilmu Pengetahuan Alam.