HUBUNGAN ARSITEKTUR TAJUK DENGAN
FOTOSINTESIS, PRODUKSI DAN KANDUNGAN
MINYAK JARAK PAGAR
(Jatropha curcas L.)
INCE RADEN
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
▸ Baca selengkapnya: tajuk tesis syariah
(2)PERNYATAAN MENGENAI DISERTASI DAN
SUMBER INFORMASI
Dengan ini saya menyatakan bahwa disertasi “Studi Arsitektur Tajuk Jarak Pagar (Jatropha curcas L.) Hubungannya dengan Fotosintesis, Produksi dan Kandungan Minyak” adalah karya saya sendiri dengan arahan Komisi Pembimbing dan belum diajukan dalam bentuk apapun kepada Perguruan Tinggi manapun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir disertasi ini.
Bogor, Januari 2009
Ince Raden
ABSTRACT
INCE RADEN. Shoot Architecture and Its Relation to Photosynthesis,
Production, and Seed Oil Content of Physic Nut (Jatropha curcas L.). Under supervisory of Bambang Sapta Purwoko as a chairman, Hariyadi, Munif Ghulamahdi, and Edi Santosa as members of the advisory committee.
Experiments on physic nut were conducted (1) to analyze branching and shoot pattern, and to observe flowering at various shoot architectures, (2) to determine physiological characteristics, leaf phyllotaxis and leaf morphology at canopy for determining reference’s leaf for photosynthetic measurement (3) to study shoot architecture with special emphasis on the number of primary branches that support optimum growth, production and oil content, and (4) to study shoot architecture by managing primary and secondary branches to support growth, production and oil content.
First experiment consisted of four treatments; those were stem pruned at height of 20 cm (T20), 30 cm (T30), 40 cm (T40) from soil surface, and control without stem pruning (T0). Second experiment observed morphological and physiological character of leaf with single factor, i.e., leaf age. Third experiment consisted of ten treatments, i.e., control, T20= stem was pruned at height of 20 cm from soil surface and without control on number of primary branches, T20-2= pruned at 20 cm and two primary branches, T20-3= pruned at 20 cm and three primary branches, T30= pruned at 30 cm and without control on number of primary branches, T30-2= pruned at 30 cm and two primary branches, T30-3= pruned at 30 cm and three primary branches, T40= pruned at 40 cm and without control on number of primary branches, T40-2= pruned at 40 cm and two primary branches, and T40-3= pruned 40 cm and three primary branches. Fourth experiment consisted of five treatments, those were K= control, TbP-2S = without control on number of primary branches and two secondary branches, 2P-2S= two primary branches and two secondary branches, 2P-3S= two primary branches and three secondary branches, 3P-3S= three primary branches and three secondary branches.
respectively. The high productivity was concomitant with highest photosynthesis rate on treatment T40 (8.10 µmole CO2/m2/s). Similarly, controlling number of secondary branches increased production on physic nut, however, this role applicable only if the number of primary branch was maintained more than three (TbP-2S). Under this circumstance, the treatment produced seed 151.92 g/ plant equal to 0.380 ton per ha (≈ 117.52 kg crude oil per ha) with photosynthesis rate reaching 9.64 µmole CO2/m2/s.
RINGKASAN
INCE RADEN. Hubungan Arsitektur Tajuk dengan Fotosintesis, Produksi dan Kandungan Minyak Jarak Pagar (Jatropha curcas L.). Dibimbing oleh Bambang Sapta Purwoko selaku ketua komisi , Hariyadi, Munif Ghulamahdi, dan Edi Santosa sebagai anggota komisi pembimbing
Pemangkasan dilakukan untuk mengatur ukuran dan bentuk pohon sesuai dengan tipe pertumbuhan dan produksi yang diinginkan, meningkatkan tunas terminal, memperbaiki kualitas buah dengan pendekatan keseimbangan pertumbuhan vegetatif dan reproduktif, memperbaiki penetrasi cahaya ke dalam kanopi sehingga cahaya tersebut dapat digunakan untuk pengembangan tunas bunga, fruit set dan pertumbuhan buah.
Untuk membentuk arsitektur tajuk diperlukan pemahaman yang mendasar tentang fisiologi tanaman, bagaimana tanaman tersebut tumbuh dan merespon tipe intervensi melalui pemangkasan sehingga dapat mengubah pertumbuhan vegetatif dan reproduktif untuk menghasilkan buah sesuai dengan karakter yang diinginkan. Pola sistem percabangan, pembungaan, pembuahan, kapasitas fotosintesis, penyebaran dan distribusi daun sebagai “source”, potensi produksi biji dan kandungan minyak berdasarkan arsitektur tajuk yang diintervensi melalui pemangkasan perlu dipelajari dalam upaya meningkatkan produksi dan kandungan minyak tanaman jarak pagar.
Penelitian dilakukan dalam empat percobaan, (1) menganalisis pola percabangan, model tajuk (menggambarkan geometri tajuk) dan mengobservasi pembentukan bunga pada arsitektur tajuk tanaman jarak pagar, (2) studi karakteristik fisiologi dan morfologi daun berdasarkan posisi daun dan umur daun pada kanopi cabang tanaman jarak pagar dalam kaitannya dengan kemampuan fotosintesis, terutama dalam menentukan nomor daun terbaik sebagai referensi (3) menemukan arsitektur tajuk yang memiliki jumlah cabang yang dapat mendukung pertumbuhan dan meningkatkan produksi serta hasil minyak jarak pagar melalui : pengujian pengaruh arsitektur tajuk berdasarkan tinggi pangkasan batang utama dan jumlah cabang primer yang dipelihara terhadap pertumbuhan, produksi, dan kandungan minyak, dan (4) menemukan arsitektur tajuk yang memiliki jumlah cabang primer dan sekunder yang dapat meningkatkan pertumbuhan, produksi dan kandungan minyak serta menemukan laju fotosintesis berdasarkan arsitektur tajuk yang dibentuk.
Batang utama yang dipangkas dapat meningkatkan jumlah cabang primer. Cabang pada tanaman jarak pagar secara alami (kontrol) terbentuk melalui 2 cara, yaitu sebelum tanaman berbunga dan sesudah tanaman berbunga. Proyeksi posisi cabang menunjukkan bahwa cabang primer tanaman jarak pagar pada posisi batang utama terdistribusi secara spiral dan bunga terletak pada terminal apeks.
tertinggi dicapai pada minggu ke-6, yaitu 42.14 per mm2 dan bagian bawah minggu ke-9 (238.59 per mm2). Daun mulai berfotosintesis sejak umur 1 minggu hingga umur 14 minggu, setelah itu daun mengalami senesen. Laju fotosintesis maksimum yang ditemukan pada penelitian ini, yaitu pada daun umur 6 minggu atau daun yang berada pada posisi daun 11 sampai 13 dari ujung pucuk cabang, yaitu 8.99 µmol CO2/m2/s. Oleh karena itu, daun ke-11 sampai 13 atau daun umur 6 minggu setelah terbentuk dapat dijadikan referensi untuk mengevaluasi laju fotosintesis.
Batang utama yang dipangkas secara umum dapat meningkatkan jumlah cabang primer yang dapat mencapai 6.7 (T30) lebih banyak dibandingkan kontrol (5.2). Peningkatan jumlah cabang akibat pemangkasan batang memberikan pengaruh terhadap peningkatan diameter batang, jumlah daun, luas daun total, tetapi menurunkan diameter cabang dan panjang cabang per tanaman jarak pagar.
Terjadi kecenderungan bahwa semakin banyak jumlah cabang, jumlah daun dan luas daun memberikan dampak terhadap peningkatan intersepsi cahaya, kandungan klorofil a, klorofil b, dan klorofil total dan antosianin. Selain itu memberikan pengaruh positif terhadap produksi buah, jumlah biji, serta laju fotosintesis arsitektur tajuk. Tinggi pangkasan 30 sampai 40 cm dengan jumlah cabang primer 3 atau lebih (6 cabang primer) dapat meningkatkan produksi jarak pagar. Produksi tahun pertama mencapai 323.81 g/tanaman atau 0.810 ton/ha pada T40 dan 320.11 g/tanaman atau 0.802 ton/ha pada T30-3 dengan minyak yang dihasilkan masing-masing 244.56 kg/ha dan 276.61 kg/ha. Hal ini mengindikasikan tinggi pangkasan dan jumlah cabang tersebut dapat direkomendasikan untuk meningkatkan produksi dan kandungan minyak jarak pagar. Laju fotosintesis tertinggi dicapai pada perlakuan T40, yaitu 8.10 µmol CO2/m2/s dan yang terendah terjadi pada perlakuan T20-2, yaitu 4.71 µmol CO2/m2/s.
Jumlah cabang sekunder 2 dapat meningkatkan produksi jarak pagar bila jumlah cabang primer yang dipelihara lebih dari 3 cabang (TbP-2S) dengan potensi produksi 151.92 g per tanaman atau 0.380 ton per ha dengan hasil minyak 110.83 kg per ha. Adapun laju fotosintesis yang dicapai pada perlakuan TbP-2S, yaitu 9.64 mol CO2/m2/s. Berdasarkan hasil tersebut, pada perlakuan jumlah cabang primer tidak dibatasi (7.3 cabang) dengan jumlah cabang sekunder dua dapat meningkatkan produksi jarak pagar. Jika hasil produksi dan minyak yang diperoleh pada tahun pertama pada percobaan 3 dan percobaan 4 diperbandingkan maka produksi dan hasil minyak per hektar percobaan 3 lebih tinggi dibandingkan percobaan 4. Hal ini berhubungan dengan pembentukan kerangka pohon yang memerlukan waktu dan energi.
Kandungan (rendemen) minyak biji jarak pagar tidak dipengaruhi secara nyata oleh perlakuan pemangkasan batang utama, jumlah cabang primer, maupun jumlah cabang sekunder. Kandungan minyak (rendemen) rata-rata 32.44 % yang berkisar pada angka 30.49 % - 34.43 %, akan tetapi produksi minyak per satuan luas (ha) yang dicapai oleh perlakuan T40 dan T30-3 mencapai nilai tertinggi, berturut-turut 276.61 kg/ha dan 244.56 kg/ha.
© Hak cipta milik Institut Pertanian Bogor, tahun 2009 Hak cipta dilindungi Undang-Undang
1. Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencamtumkan atau menyebutkan sumber :
a. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik atau tinjauan suatu masalah
b. Pengutipan tidak merugikan kepentingan yang wajar IPB
HUBUNGAN ARSITEKTUR TAJUK DENGAN
FOTOSINTESIS, PRODUKSI DAN KANDUNGAN
MINYAK JARAK PAGAR
(Jatropha curcas L.)
INCE RADEN
Disertasi
Sebagai salah satu syarat untuk memperoleh gelar Doktor pada
Program Studi Agronomi
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Judul Disertasi : Hubungan Arsitektur Tajuk dengan Fotosintesis, Produksi, dan Kandungan Minyak Jarak Pagar (Jatropha curcas L.)
Nama Mahasiswa : Ince Raden
Nomor Pokok : A361050081
Program Studi : Agronomi (AGR)
Disetujui :
Komisi Pembimbing
Prof. Dr. Ir. Bambang Sapta Purwoko, M.Sc. Dr. Ir. Hariyadi, MS Ketua Anggota
Dr. Ir. Munif Ghulamahdi, MS Dr. Edi Santosa, SP., M.Si
Anggota Anggota
Diketahui :
Ketua Program Studi Dekan Sekolah Pascasarjana Agronomi
Dr. Ir. Munif Ghulamahdi, MS. Prof. Dr. Ir. Khairil Anwar Notodiputro M.Sc
PRAKATA
Puji syukur penulis panjatkan kehadirat Allah SWT yang telah
melimpahkan rahmat dan petunjuk-Nya sehingga disertasi yang merupakan salah
satu syarat untuk memperoleh gelar Doktor pada Sekolah Pascasarjana di Institut
Pertanian Bogor dapat penulis selesaikan dengan baik.
Karya Ilmiah yang dihasilkan penulis yang diterbitkan sebagai bagian dari
disertasi adalah : Karakteristik Daun Jarak Pagar (Jatropha curcas L.)
Hubungannya dengan Fotosintesis. Artikel tersebut diterbitkan di Buletin
Agronomi Vol. XXXVI No. 2 Agustus 2008.
Penulis menyadari bahwa keberhasilan tersebut merupakan bimbingan dan
bantuan yang tulus dan ikhlas dari berbagai pihak, untuk itu pada kesempatan ini
penulis ingin menyampaikan rasa terima kasih yang setulus-tulusnya kepada :
1. Prof. Dr. Ir. Bambang Sapta Purwoko, MSc. Selaku ketua komisi
pembimbing, Dr. Ir. Hariyadi, MS., Dr. Ir. Munif Ghulamahdi, MS., dan
Dr. Edi Santosa, SP., MSi. Masing-masing selaku anggota komisi
pembimbing yang telah memberikan bimbingan dan arahan selama
melakukan penelitian dan penyusunan disertasi.
2. Pimpinan beserta staf Institut Pertanian Bogor yang telah berkenan untuk
menerima penulis sebagai mahasiswa Program Doktor
3. Koordinator Kopertis Wilayah XI Kalimantan yang telah mengizinkan dan
merekomendasikan penulis untuk melanjutkan pendidikan ke Program
Doktor.
4. Rektor beserta staf Universitas Kutai Kartanegara yang telah
mengizinkan penulis untuk melanjutkan pendidikan ke Program Doktor.
5. Tim BPPS Dikti 2005 yang telah memberikan bantuan beasiswa selama
mengikuti pendidikan program Doktor di Institut Pertanian Bogor.
6. Pemerintah Kabupaten Kutai Kartanegara, Pemerintah Propinsi
Kalimantan Timur yang telah membantu penulis untuk biaya penelitian
7. Tim Hibah Bersaing Dikti tahun 2008 yang telah memberikan bantuan
8. Pengelola University Farm dan lahan Kebun Percobaan di Cikabayan, para
laboran di Lab Fisiologi IPB, RGCI, dan PAU IPB.
9. Orang tua, mertua, istri (Dra. Saoda Nur) dan anak-anak tercinta (Raudhia
Zahra, Nurul Azmi Afifah, dan Mohamad Farras Arhab), beserta keluarga
kandung penulis: Ir. Asma Intje Gani, MSi, Drs. Muktasim, Ince Moh.
Hasan, Ince Nurfaida, SE., dan Ince Moh. Ikbal, ST. dan keluarga besar
penulis yang telah memberi motivasi, bantuan moril maupun materi
kepada penulis.
10.Rekan-rekan mahasiswa Sekolah Pascasarjana khususnya Ir. Bambang
Budi Santoso, MSc., Ir. Iskandar Lapanjang, MP., Ir. Abdul Haris Badrun,
MSi, Ir. Thamrin, M.P. serta teman-teman Program Studi Agronomi untuk
segala bantuan dan diskusinya
11.Semua pihak yang telah banyak membantu dan mendukung penulis selama
mengikuti pendidikan di IPB yang tidak dapat penulis sebutkan satu
persatu.
Semoga bimbingan dan segala bentuk bantuan yang telah diberikan dari
semua pihak mendapatkan nilai ibadah yang diterima oleh Allah
SWT…Amin.
Bogor, Januari 2009
Penulis,
RIWAYAT HIDUP
Penulis dilahirkan di Poso pada tanggal 8 September 1967 dari ayah Drs.
H. Mansur Intje Gani dan Ibu Hj. Zaitun Abdul Samad. Penulis merupakan anak
ketiga dari enam bersaudara. Tahun 1996 penulis menikah dengan Dra. Saoda Nur
dan hingga saat ini dikaruniai 3 orang anak, yaitu Raudhia Zahra, Nurul Azmi
Afifah, dan Mohammad Farras Arhab.
Tahun 1993 penulis menyelesaikan pendidikan sarjana di Program Studi
Budidaya Pertanian, Fakultas Pertanian, Universitas Tadulako Palu. Tahun 1997
penulis melanjutkan pendidikan ke Program Magister di Universitas Padjadjaran
Bandung pada Bidang Ekofisiologi Tanaman dan menamatkannya pada tahun
1999. Selanjutnya, penulis melanjutkan Pendidikan Program Doktor di Sekolah
Pascasarjana Institut Pertanian Bogor sejak tahun 2005 pada Program Studi
Agronomi. Beasiswa pendidikan pascasarjana diperoleh dari BPPS Dikti sejak
tahun 2005.
Penulis bekerja sebagai staf pengajar kopertis wilayah XI Kalimantan
dipekerjakan di Fakultas Pertanian Universitas Kutai Kartanegara di Tenggarong,
Kalimantan Timur sejak tahun 1994.
Selama mengikuti program S3, penulis menjadi Wakil Ketua Pengurus
Forum Mahasiswa Pascasarjana Agronomi periode 2005-2006, Koordinator
Bidang pada Forum Wacana Mahasiswa Pascasarjana IPB periode 2006-2007.
Ketua Forum Mahasiswa Asal Kalimantan Timur periode 2006-2007. Karya
Ilmiah yang dihasilkan penulis yang telah dan akan diterbitkan adalah : (1)
Pengaruh Alelopati Jarak Pagar (Jatropha curcas L.) terhadap perkecambahan
benih Jagung, Tomat dan Padi Gogo, (2). Karakteristik Daun Jarak Pagar
(Jatropha curcas L.) Hubungannya dengan Fotosintesis. (3). Pengaruh
Pemangkasan Batang Utama dan Jumlah Cabang Primer yang Dipelihara terhadap
Pertumbuhan, Produksi dan Kandungan Minyak Jarak Pagar (Jatropha curcas L.)
Artikel (1) dan (2) berturut-turut telah diterbitkan di Buletin Agronomi April
2008 dan Agustus 2008. Sementara artikel (3) masih dalam tahap telaah reviewer
DAFTAR ISI
Halaman PRAKATA... DAFTAR ISI... DAFTAR TABEL... DAFTAR GAMBAR... DAFTAR LAMPIRAN... I. PENDAHULUAN Latar Belakang………... Tujuan Umum Penelitian... Tujuan Khusus Penelitian... Kegunaan Penelitian... Ruang Lingkup Penelitian...II. TINJAUAN PUSTAKA
Biologi dan Ekologi Jarak Pagar... Fisiologi Pemangkasan... Pembentukan Arsitektur Tajuk Melalui Pemangkasan... Faktor-Faktor yang Mempengaruhi Laju Fotosíntesis………..…...
III. ANALISIS PERCABANGAN DAN MODEL TAJUK JARAK PAGAR
IV. KARAKTERISTIK DAUN JARAK PAGAR DAN HUBUNGANNYA DENGAN FOTOSINTESIS ABSTRACT…………..………... PENDAHULUAN………... Latar Belakang…...……… Tujuan……… BAHAN DAN METODE……….. Waktu dan Tempat………... Metode Percobaan………... Peubah yang Diamati.………... HASIL DAN PEMBAHASAN... Posisi Daun... Perkembangan Daun... Kandungan Klorofil... Stomata Daun…... Laju Fotosintesis... SIMPULAN...
V. PENGARUH TINGGI PANGKASAN BATANG UTAMA DAN JUMLAH CABANG PRIMER YANG DIPELIHARA TERHADAP PERTUMBUHAN, PRODUKSI DAN KANDUNGAN MINYAK JARAK PAGAR (Jatropha curcas L.)
VI. PENGARUH PENGENDALIAN JUMLAH CABANG PRIMER DAN JUMLAH CABANG SEKUNDER TERHADAP PERTUMBUHAN, PRODUKSI DAN KANDUNGAN MINYAK JARAK PAGAR
ABSTRACT…………..………... PENDAHULUAN………... Latar Belakang…...………. Tujuan………. BAHAN DAN METODE……….... Waktu dan Tempat………... Bahan dan Alat... Metode Percobaan... ... Pelaksanaan Percobaan……... Peubah yang Diamati... HASIL DAN PEMBAHASAN... Komponen Vegetatif... Intersepsi Cahaya ... Kandungan Klorofil dan Antosianin Daun... Kerapatan Stomata... Laju Fotosintesis... Komponen Generatif... Produksi Buah dan Biji... Kandungan Minyak dan Air... Korelasi Peubah Pertumbuhan dan Produksi... Simpulan...
PEMBAHASAN UMUM... Arsitektur Tajuk dan Sistem Percabangan... Perkembangan Daun dan Fotosintesis... Pertumbuhan Vegetatif, Klorofil, dan Intersepsi Cahaya... Pertumbuhan Generatif, Tanah dan Iklim... Kandungan dan Hasil Minyak...
DAFTAR TABEL
Halaman
1. Karakteristik jumlah cabang, diameter batang, sudut cabang, dan diameter cabang primer akibat pemangkasan pucuk ………...
2. Kerapatan stomata pada bagian atas dan bawah daun berdasarkan posisi daun (umur daun) pada cabang jarak pagar………..
3. Korelasi karakter fisiologi, fotosintesis dengan berbagai faktor lingkungan………...
4. Data hasil analisis sampel tanah tempat penelitian...
5. Data iklim tempat penelitian di kebun Cikabayan IPB, Bogor………...
6. Perkembangan jumlah cabang primer akibat pemangkasan batang utama……….
7. Perkembangan diameter batang (cm) pada berbagai arsitektur tajuk...
8. Perkembangan diameter cabang primer pada berbagai arsitektur tajuk....
9. Perkembangan panjang cabang pada berbagai arsitektur tajuk ...
10. Perkembangan jumlah daun total pada berbagai arsitektur tajuk...
11. Perkembangan luas daun total pada berbagai arsitektur tajuk...
12. Intersepsi cahaya berbagai arsitektur tajuk jarak pagar...
13. Kandungan klorofil a, b, dan total serta antosianin daun pada berbagai arsitektur tajuk jarak pagar...
14. Kerapatan stomata pada arsitektur tajuk jarak pagar...
15. Laju fotosintesis berbagai arsitektur tajuk jarak pagar...
16. Waktu berbunga, persentase cabang primer berbunga dan jumlah buah per tandan...
17. Jumlah buah per tanaman, jumlah biji per tanaman, bobot kering per biji ...
18. Bobot kering biji per tanaman, bobot kering biji per petak, dan bobot kering biji per hektar...
26
43
45
49
50
55
56
57
58
59
59
61
62
64
65
66
67
19. Kandungan minyak dan air biji kering jarak pagar... ...
20. Koefisien korelasi antara peubah pertumbuhan, fisiologi, produksi dan minyak yang dihasilkan pada tanaman jarak pagar ...
21. Penambahan jumlah cabang primer bulan ke-2 sampai ke-10 setelah pangkas batang utama pada arsitektur tajuk jarak pagar...
22. Diameter batang tanaman bulan ke-2 sampai ke-10 setelah pangkas batang utama pada arsitektur tajuk jarak pagar...
23. Diameter cabang primer bulan ke-2 sampai ke-10 setelah pangkas batang utama pada arsitektur tajuk jarak pagar...
24. Diameter cabang sekunder bulan ke-4 sampai ke-10 setelah pangkas batang utama pada arsitektur tajuk jarak pagar ...
25. Panjang cabang bulan ke-2 sampai ke-10 setelah pangkas batang utama pada arsitektur tajuk jarak pagar ...
26. Jumlah daun total bulan ke-2 sampai ke-10 setelah pangkas batang utama pada berbagai arsitektur tajuk jarak pagar ...
27. Luas daun total bulan ke-2 sampai ke-10 setelah pangkas batang utama pada arsitektur tajuk jarak pagar ...
28. Persentase intersepsi cahaya bulan ke-2 sampai ke-10 setelah pangkas batang utama pada arsitektur tajuk jarak pagar ...
29. Kandungan klorofil a, b, dan total serta antosianin daun pada arsitektur tajuk jarak pagar...
30. Kerapatan stomata pada arsitektur tajuk jarak pagar...
31. Laju fotosintesis pada arsitektur tajuk jarak pagar...
32. Waktu berbunga, persentase cabang sekunder berbunga dan jumlah buah per tandan...
33. Jumlah buah per tanaman, jumlah biji per tanaman, dan bobot kering per biji...
34. Bobot kering biji per tanaman, bobot kering biji per petak, dan bobot kering biji per ha...
35. Kandungan minyak dan air jarak pagar ... 69
71
79
79
80
81
81
82
83
83
85
86
87
88
89
89
90
DAFTAR GAMBAR
Halaman
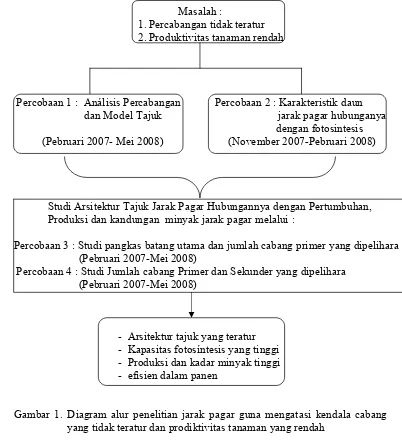
1. Diagram alur penelitian ………
2. Irisan membujur ujung pucuk tampak meristem apical, primordia daun dan primordia tunas samping ...
3. Bagian sebuah dahan yang menunjukkan buku dan beberapa tipe tunas (a), struktur tunas alternate (b), dan struktur tunas opposite (c)...
4. Apikal dormansi (a), pucuk yang tidak dipangkas (b), dan pucuk yang dipangkas (c) ...
5. Cabang terbentuk sebelum berbunga (a) dan cabang terbentuk setelah pucuk batang utama berbunga (b)...
6. Pembungaan dan buah jarak pagar (a) dan ilustrasi pembentukan bunga dan cabang pada tanaman jarak pagar (b) = letak bunga dan buah ...
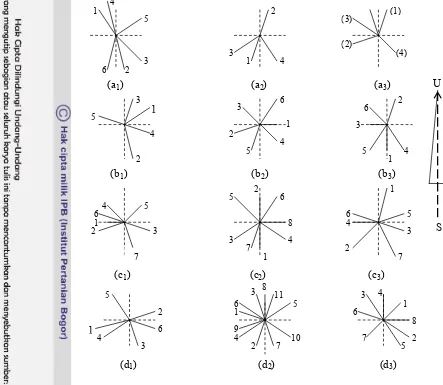
7. Proyeksi posisi cabang jarak pagar yang tanpa pangkas (a1, a2, a3), T20 (b1, b2, b3), T30 (c1, c2, c3), T40 (d1, d2, d3). Angka menunjukkan nomor cabang tampak atas dan garis putus-putus menunjukkan arah utara, selatan, timur dan barat...
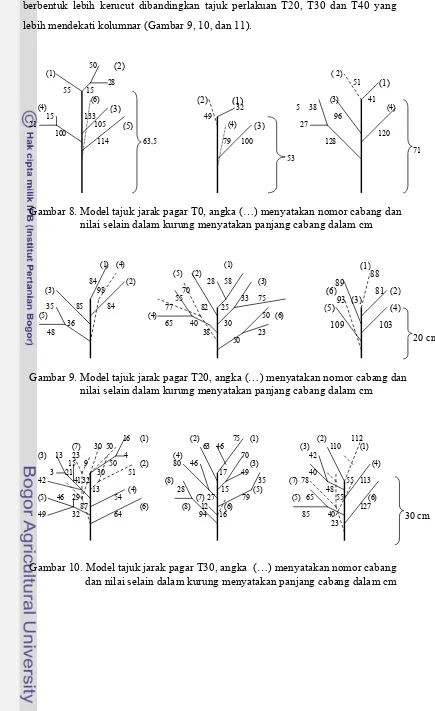
8. Model tajuk jarak pagar T0, angka (...) menyatakan nomor cabang dan nilai selain dalam kurung menyatakan panjang caabang dalam cm ...
9. Model tajuk jarak pagar T20, angka (...) menyatakan nomor cabang dan nilai selain dalam kurung menyatakan panjang cabang dalam cm...
10. Model tajuk jarak pagar T30, angka (...) menyatakan nomor cabang dan nilai selain dalam kurung menyatakan panjang cabang dalam cm...
11. Model tajuk jarak pagar T40, angka (...) menyatakan nomor cabang dan nilai selain dalam kurung menyatakan panjang cabang dalam cm...
12. Jumlah kumulatif daun pada cabang tanaman jarak pagar...
13. Filotaksis daun jarak pagar tampak dari atas... 7
10
12
14
28
29
31
32
32
32
33
37
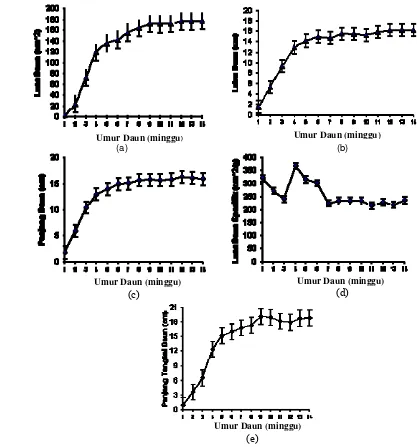
14. Perkembangan luas daun (a), lebar daun (b), panjang daun (c), luas daun spesifik (d), dan panjang tangkai daun ...
15. Perkembangan bobot kering tangkai daun (a) dan bobot kering daun (b) tanaman jarak pagar...
16. Hubungan sudut inklinasi tangkai daun dengan umur daun (a), kehijauan daun dengan umur daun (b) tanaman jarak pagar...
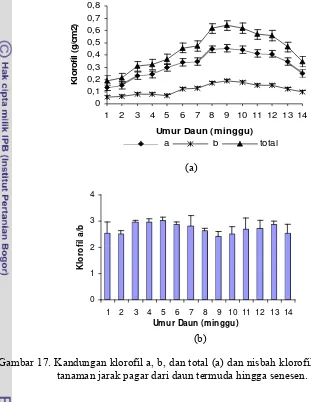
17. Kandungan klorofil a, b, dan total (a) dan nisba klorofil a/b (b) daun tanaman jarak pagar dari daun termuda hingga senesence...
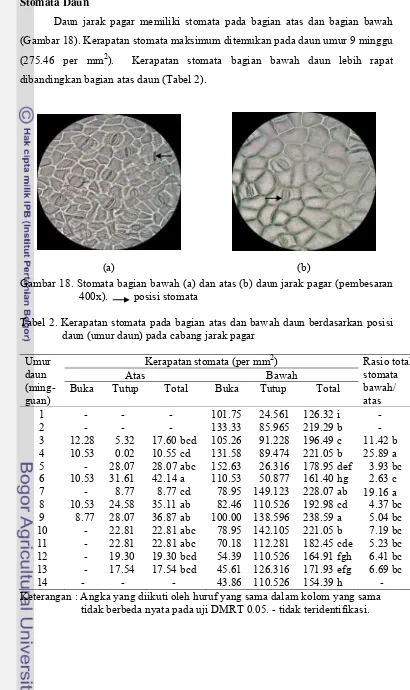
18. Stomata bagian bawah (a) dan atas (b) daun jarak pagar...
19. Laju fotosintesis daun tanaman jarak pagar sejak umur 1 minggu hingga 14 minggu………
20. Data curah hujan, waktu pembibitan, penanaman di lapangan (tnm), waktu pemangkasan batang utama (P.BU), dan pengataman pertama (PP)………...
21. Alat soxhlet yang digunakan untuk menganalisis kandungan minyak……….
22. Penempatan cuvet portable chamber leaf model ADC Bio scientific Ltd. pada daun jarak pagar yang diukur…...
23. Data curah hujan, waktu pembibitan, penanaman dilapangan (Tnm), waktu pemangkasan batang utama (P.BU), dan pangkas cabang primer (P.CP)………...
24. Posisi dan letak daun tampak dari atas (a) dan tampak samping (b)... 39
40
41
42
43
44
52
53
54
76
95
DAFTAR LAMPIRAN
Halaman
1. Prosedur kerja penentuan kandungan klorofil daun...
2. Prosedur kerja penentuan jumlah stomata...
3. Analisis kandungan minyak jarak pagar metode soxhlet...
4. Prosedur kerja analisis antosianin... 110
111
112
113
I. PENDAHULUAN
Latar Belakang
Salah satu masalah krusial yang dihadapi bangsa Indonesia saat ini adalah
kekurangan pasokan energi. Pasokan energi dalam negeri terutama berbasis fosil
dan minyak bumi mengalami kendala akibat produksi yang lebih rendah
dibanding tingkat konsumsinya yang berakibat ketergantungan pada impor.
Berdasarkan data migas terjadi impor bahan bakar minyak dari 86.6 juta barrel
tahun 2001 dengan nilai 2.6 milyar USD meningkat menjadi 124.8 juta barrel
dengan nilai 5.8 milyar USD tahun 2004. Pada tahun 2007 kebutuhan solar
nasional mencapai 30.40 juta liter. Kebutuhan solar ini akan meningkat pada
tahun 2010 hingga mencapai 34.89 juta liter. Kondisi ini harus diantisipasi
pemerintah dengan pengembangan berbagai sumber energi yang dapat
diperbaharui. Dalam periode 1995-2004, produksi minyak mentah nasional
rata-rata mengalami penurunan 12.06 juta barel per tahun, atau sekitar 2.31% per tahun
(BPS, 2005) dan berdasarkan data Pertamina (2005) persediaan minyak untuk
memenuhi kebutuhan konsumsi Indonesia diperkirakan akan cukup hingga 20
tahun ke depan.
Indonesia memiliki ketersediaan sumber daya genetik tanaman penghasil
minyak nabati yang cukup tinggi yang dapat dimanfaatkan sebagai bahan baku
biodisel. Salah satu diantaranya adalah Jarak Pagar (Jatropha curcas L.). Minyak
jarak pagar (crude jatropha oil) memiliki sifat fisika dan kimia yang sesuai
dengan minyak diesel, sehingga pemerintah memprogramkan untuk mensubtitusi
10 % dari kebutuhan minyak diesel. Tanaman jarak pagar selain dapat dijadikan
sebagai sumber energi alternatif (penghasil minyak nabati non pangan) juga
bermanfaat sebagai obat tradisional, insektisida nabati, tanaman pelindung dan
pencegah erosi / konservasi, serta dapat diolah menjadi pakan ternak, pupuk
organik dan surfaktan (Gubitz et al., 1996).
Jarak pagar merupakan salah satu tanaman yang diunggulkan di Indonesia,
namun demikian, produktivitasnya masih relatif rendah, sehingga perlu upaya
untuk meningkatkan produktivitas menjadi prioritas. Hasil penelitian
bervariasi, yaitu 0.3 kg/pohon atau 833 kg/ha (Heller, 1996), 400 kg/ha/tahun
(Jones dan Miller, 1992), dan 200 kapsul/tanaman atau 0.36 kg/pohon (Hasnam et
al., 2007), dan 880 kg/ha (Santoso et al., 2008). Sementara itu, kandungan minyak
biji (oil content in whole seed) yang dihasilkan oleh berbagai propenan di India
sekitar 33.50 % - 38.42 % (Ginwal et al., 2003), IP-2A 31 % - 32 %, IP-2P 32 %
- 34 %, dan IP-2M 31%-32% (Hasnam et al., 2008).
Perbaikan teknik budidaya untuk meningkatkan produksi jarak pagar di
Indonesia perlu dilakukan karena hingga saat ini teknologi budidaya berdasarkan
kondisi spesifik wilayah Indonesia masih sangat terbatas.
Observasi pendahuluan menunjukkan bahwa tanaman jarak pagar yang
ditanam atau tumbuh secara alami yang berasal dari biji mempunyai karakter (1)
secara alamiah percabangan (tajuk) yang terbentuk tidak teratur dan tidak
produktif, (2) cabang umumnya terbentuk setelah bunga atau buah pertama
terbentuk (memiliki 60 – 70 daun), (3) tinggi pohon mencapai 5-7 meter, (4) tunas
cabang umumnya terbentuk bersamaan dengan perkembangan reproduktif, (5)
bunga muncul pada ujung-ujung pucuk (bunga terminal).
Pembentukan arsitektur tajuk bertujuan untuk mengoptimalkan intersepsi
cahaya dan mengarahkan strategi pertumbuhan dan perkembangan ke arah yang
menguntungkan sehingga produktivitas tinggi dan memudahkan manajemen
kebun. Tujuan ini dapat dicapai dengan mengurangi kompetisi antara organ,
vegetatif dan generatif serta keseimbangan alokasi asimilat yang harus ditunjang
oleh intersepsi dan distribusi cahaya yang baik.
Menurut Curry (1991) hasil tanaman dapat diprediksi dari perkembangan
pertumbuhan tajuk (shoot), intersepsi cahaya dan distribusinya. Model hubungan
distribusi cahaya dengan proses fungsi fisiologi, seperti induksi pucuk bunga,
jumlah bunga menjadi buah (fruit set), dan kualitas buah dapat menjadi nilai yang
berguna untuk mendesain arsitektur tajuk. Selanjutnya dinyatakan bahwa dalam
kenyataannya aplikasi model intersepsi cahaya dan distribusinya dalam
memodifikasi kanopi pohon secara signifikan meningkatkan efisiensi dan kualitas
buah. Hal ini sejalan dengan yang dilaporkan oleh George et al. (1993) dan
Mowat dan George (1994) yang menyatakan bahwa mikroklimat di dalam kanopi
buah (fruit set) dan kualitas buah. Guillermo (2000) menyatakan bahwa intersepsi
photosynthetically active radiation (PAR) selama periode pengisian biji
meningkatkan bobot biji dan konsentrasi minyak pada bunga matahari.
Menurut Costes et al. (2006), untuk menganalisis arsitektur pohon buah
yang berimplikasi pada manajemen pohon dan produksi buah, yang pertama kali
dilakukan adalah mengetahui fenomena pertumbuhan, proses percabangan dan
pembungaan pada kanopi pohon. Selanjutnya, aplikasi analisis arsitektur pada
tanaman difokuskan pada 2 hal, yaitu (1) struktur organ (organ arrangement),
termasuk vegetatif dan organ bunga, (2) cabang buah dan seluruh perilaku pohon.
Kedua hal ini sebagai struktur dasar yang digunakan untuk menginterpretasikan
pengaruhnya pada aspek agronomi secara praktis.
Seni membentuk pohon untuk mengubah tanaman yang berproduksi tinggi
telah dimulai sejak lama pada berbagai tanaman bernilai ekonomi tinggi seperti
kakao, kopi (Halle et al. 1978), tanaman sub tropika apel, pir, peach (Verheij dan
Coronel, 1992) dan famili Rosaceae lain (Ryugo, 1988). Namun untuk tanaman
jarak pagar masih belum banyak diteliti.
Salah satu tindakan agronomis yang dapat dilakukan untuk perbaikan
teknik budidaya tanaman jarak pagar adalah pembentukan arsitektur tajuk
melalui pemangkasan. Pengaturan arsitektur tanaman melalui pemangkasan akan
dapat mengefisienkan ruang tempat tanaman tumbuh dan dapat meningkatkan
produktivitas terutama tanaman yang berbunga di ujung ranting (terminal) atau
hasil tanaman per satuan luas karena tujuan pembentukan arsitektur tajuk untuk
mengatur sistem percabangan, meratakan penerimaan cahaya, menyebarkan
percabangan agar dapat membagi ruang tumbuh secara merata, mempermudah
pengelolaan pohon dan mempermudah penyusunan anggaran kebun serta prediksi
hasil karena ukuran dan bentuk pohon seragam (Widodo, 1995).
Tindakan pembentukan arsitektur tajuk melalui pemangkasan pada
tanaman jarak pagar sangat diperlukan untuk memperoleh tajuk tanaman yang
efisien dalam memproduksi buah, meningkatkan produksi hasil panen,
membentuk struktur fisik tanaman (kanopi) seperti semak atau payung dan
meningkatkan cabang produktif. Semakin banyak cabang produktif yang
pada jumlah cabang terminal tertentu (Mahmud, 2006). Hal ini sejalan dengan
pendapat Ginwal et al. (2003) yang menyatakan terdapat korelasi positif antara
jumlah cabang, jumlah kapsul per tandan dengan produksi tanaman jarak pagar
yang dihasilkan.
Cabang tempat tumbuhnya bunga dan buah jarak pagar (selanjutnya
disebut cabang terminal) sangat ditentukan oleh jumlah cabang primer dan
sekunder yang tumbuh dari batang utama. Oleh karena itu pengaturan arsitektur
tajuk yang berdasarkan jumlah cabang primer dan sekunder yang dipelihara
menjadi penting untuk diteliti agar dapat membentuk arsitektur tajuk yang baik
sehingga tanaman mampu menghasilkan bunga, buah serta kualitas minyak yang
berkualitas. Menurut Ferry (2006) jumlah cabang primer yang perlu dipelihara
antara 3 – 5 cabang sedangkan jumlah cabang sekunder yang perlu dipelihara
sebanyak 3 cabang, Hal ini dilakukan agar setiap pohon jarak pagar mempunyai
40-45 cabang terminal. Berdasarkan laporan Mahmud (2006), di India,
menunjukkan bahwa jumlah cabang terminal yang ideal per tanaman pada
tanaman jarak pagar adalah 40 cabang dan jumlah buah 10-15 per tandan.
Selanjutnya dikatakan jika jumlah cabang terminal per pohon lebih dari 40 cabang
maka jumlah buah per tandan akan berkurang dan ukurannya mengecil sehingga
akan mempengaruhi mutu biji yang dihasilkan.
Berdasarkan hal di atas, kajian model arsitektur tajuk, mekanisme
percabangan dan pembungaan yang terjadi pada tanaman jarak pagar, produksi
dan kandungan minyak biji dengan melakukan intervensi terhadap bentuk
arsitektur tajuk perlu untuk dilakukan dalam memperbaiki teknik budidaya
tanaman jarak pagar.
Tujuan Umum Penelitian
Secara umum penelitian ini bertujuan untuk menemukan arsitektur tajuk
yang memiliki jumlah cabang primer dan sekunder yang dapat mendukung
pertumbuhan, fotosintesis, produksi dan kandungan minyak jarak pagar melalui
Tujuan Khusus Penelitian
1. Menganalisis pola pembentukan cabang, model tajuk, dan mengobservasi
pembentukan bunga pada arsitektur tajuk tanaman jarak pagar.
2. Mengkaji karakteristik fisiologi dan potensi “source” daun secara
morfo-fisiologi berdasarkan posisi dan umur daun pada kanopi cabang tanaman jarak
pagar dalam kaitannya dengan kemampuan fotosintesis, terutama dalam
menentukan nomor daun terbaik sebagai referensi.
3. Mengkaji pengaruh arsitektur tajuk berdasarkan tinggi pangkasan batang utama
dan jumlah cabang primer yang dipelihara terhadap pertumbuhan, produksi
dan kandungan minyak jarak pagar
4. Mengkaji pengaruh arsitektur tajuk berdasarkan jumlah cabang primer dan
sekunder yang dipelihara terhadap pertumbuhan, produksi dan kandungan
minyak jarak pagar
Kegunaan Penelitian
Kegunaan hasil percobaan ditinjau dari konstribusinya untuk
pengembangan ilmu dan pembangunan di bidang pertanian adalah :
1. Menemukan model tajuk dan pola percabangan jarak pagar yang alami dan
dipangkas batang utamanya
2. Menemukan daun referensi untuk mengukur laju fotosintesis pada tanaman
jarak pagar
3. Menemukan tinggi pangkasan batang utama dan jumlah cabang primer yang
optimal terhadap pertumbuhan, produksi dan kadar minyak jarak pagar
4. Menemukan jumlah cabang primer dan sekunder yang optimal terhadap
pertumbuhan, produksi dan kadar minyak jarak pagar
5. Menemukan karakter fisiologi dan fotosíntesis arsitektur tajuk tanaman jarak
Ruang Lingkup Penelitian
Penelitian dilakukan dalam empat percobaan, (1) mengkaji pembentukan
cabang, model tajuk dan mengobservasi pembentukan bunga pada arsitektur tajuk
tanaman jarak pagar, (2) studi karakteristik daun pada arsitektur tajuk jarak pagar
secara morfologi dan fisiologi berdasarkan posisi daun dan umur daun pada
kanopi cabang tanaman jarak pagar dalam kaitannya dengan kemampuan
fotosintesis, terutama dalam menentukan nomor daun terbaik sebagai referensi,
(3) menemukan arsitektur tajuk yang memiliki jumlah cabang yang dapat
mendukung pertumbuhan dan meningkatkan produksi serta hasil minyak jarak
pagar melalui : pengujian pengaruh arsitektur tajuk berdasarkan tinggi pangkasan
batang utama dan jumlah cabang primer yang dipelihara terhadap pertumbuhan,
produksi, dan kandungan minyak, dan (4) menemukan arsitektur tajuk yang
memiliki jumlah cabang primer dan sekunder yang dapat meningkatkan
pertumbuhan, produksi dan kandungan minyak serta menemukan laju fotosintesis
berdasarkan arsitektur tajuk yang dibentuk. Adapun alur penelitian dapat dilihat
Masalah :
1. Percabangan tidak teratur 2. Produktivitas tanaman rendah
Percobaan 1 : Análisis Percabangan Percobaan 2 : Karakteristik daun dan Model Tajuk jarak pagar hubunganya dengan fotosintesis (Pebruari 2007- Mei 2008) (November 2007-Pebruari 2008)
Studi Arsitektur Tajuk Jarak Pagar Hubungannya dengan Pertumbuhan, Produksi dan kandungan minyak jarak pagar melalui :
Percobaan 3 : Studi pangkas batang utama dan jumlah cabang primer yang dipelihara (Pebruari 2007-Mei 2008)
Percobaan 4 : Studi Jumlah cabang Primer dan Sekunder yang dipelihara (Pebruari 2007-Mei 2008)
- Arsitektur tajuk yang teratur - Kapasitas fotosíntesis yang tinggi - Produksi dan kadar minyak tinggi - efisien dalam panen
Gambar 1. Diagram alur penelitian jarak pagar guna mengatasi kendala cabang yang tidak teratur dan prodiktivitas tanaman yang rendah
II. TINJAUAN PUSTAKA
Biologi dan Ekologi Jarak Pagar
Tanaman jarak pagar berasal dari Amerika Tengah dan saat ini menyebar
di seluruh daerah tropik di dunia. Dalam klasifikasinya, tanaman jarak pagar
termasuk divisi Spermatophyta, sub-divisi Angiospermae, klas Dicotyledonae,
ordo Euphorbiales, famili Euphorbiaceae, genus Jatropha, dan spesies Jatropha
curcas L. (Wiesenhutter, 2003; Hariyadi, 2005; Dwary dan Pramanick, 2006;
Prihandana dan Hendroko, 2006). Tanaman perdu dengan tinggi mencapai 5 m
(Heller, 1996; Wiesenhutter, 2003; Ginwal, 2004). Pada kondisi kandungan air
tanah yang baik perkecambahan membutuhkan waktu 10 hari dengan
memunculkan radikula dan empat akar peripheral (Heller, 1996). Percabangan
jarak pagar tidak teratur, batangnya berkayu, silindris dan bila terluka
mengeluarkan getah (Dwary dan Pramanick, 2006). Menurut Heller (1996) dan
Wiesenhutter (2003) jarak pagar termasuk tanaman sukulen yang menggugurkan
daunnya selama musim kering sehingga tanaman ini adaptif pada lahan arid dan
semi-arid.
Daun jarak pagar berupa daun tunggal, berlekuk, bersudut 3 -5, tulang
daun menjari dengan 5 – 7 tulang utama, warna daun hijau (permukaan bagian
bawah lebih pucat dibandingkan permukaan bagian atas), panjang tangkai daun
antara 4 -15 cm. (Tim Jarak Pagar, 2006).
Hasnam (2006) mengemukakan bahwa bunga jarak pagar berupa bunga
majemuk tersusun dalam rangkaian (inflorescence) berumah satu, bunga
berwarna kuning kehijauan, persentase bunga betina 5 – 10 % dari 100 bunga atau
lebih, muncul di ujung batang, masa berbunga bunga betina 3-4 hari, bunga betina
membuka 1-2 hari sebelum bunga jantan, lama pembungaan inflorecence 10-15
hari, bunga menyerbuk dengan bantuan serangga.
Buah disebut buah kapsul, berbentuk bulat telur, panjang buah 2.5 cm,
berwarna hijau ketika masih muda dan kuning setelah matang, terdapat 420
buah dan 1580 biji per kg (Dwary dan Pramanick, 2006). Buah jarak terbagi
menjadi 2-4 ruang yang masing-masing berisi 1 biji yang bentuknya bulat lonjong
kuning mengandung rendemen minyak sekitar 30 – 40% (Pusat Penelitian dan
Perkembangan Perkebunan, 2006); 36 %-38.73 % (Tim Peneliti, 2006); 31-37 %
(Dwary dan Pramanick, 2006)
Pembentukan buah membutuhkan waktu selama 90 hari dari pembungaan
sampai matang (Heller, 1996). Menurut Wiesenhutter (2003) di Cape Verde
produksi mencapai 780 sampai 2,250 kg biji per ha, di India produksi tanpa kulit
biji di atas 12 ton per ha yang dicapai dengan irigasi pada tahun ke enam, di Mali
produksi sekitar 2 – 2.4 ton per ha.
Menurut Heller (1996) jarak pagar beradaptasi baik pada lahan marginal
dengan lahan miskin hara dan curah hujan rendah. Di daerah Amazone jarak pagar
tumbuh baik pada daerah kering dengan rata-rata curah hujan antara 300 – 1000
mm per tahun dan juga dapat tumbuh dengan baik pada curah hujan yang lebih
tinggi dengan aerasi baik. Wiesenhutter (2003) mengemukakan bahwa tanaman
jarak pagar membutuhkan curah hujan 500 – 600 mm per tahun dan di Cape
Verde juga tumbuh baik pada curah hujan 250 mm per tahun dengan kelembaban
yang tinggi dan kondisi kering dapat meningkatkan kandungan minyak pada biji.
Tanaman jarak pagar dapat tumbuh di daerah dataran rendah bahkan
pinggir pantai sampai ketinggian lebih dari 1000 m di atas permukaan laut (dpl),
bahkan menurut Heller (1996) di Fogo, Afrika jarak pagar ditemukan pada
ketinggian 1700 m dpl. Daerah yang optimum untuk pengembangan jarak pagar
adalah daerah dengan ketinggian 0 – 500 m dpl, tanaman ini adaptif dengan suhu
tinggi dan daerah yang menjadi pusat koleksi berbagai provenan di Cape Verde
mempunyai rata-rata suhu tahunan 20-280C.
Berdasarkan informasi tersebut jarak pagar memiliki daya adaptasi yang
cukup luas akan tetapi untuk memperoleh pertumbuhan yang baik disertai
produksi dan mutu yang tinggi, kecukupan air dan unsur hara tanah harus
diperhatikan bila jarak pagar harus ditanam di daerah yang relatif kering dengan
intensitas radiasi yang tinggi karena kondisi kering dapat menyebabkan daun
tanaman gugur dan produktivitasnya menurun. Demikian pula halnya bila jarak
pagar ditanam pada daerah yang curah hujannya tinggi, pembuatan drainase perlu
diperhatikan karena akar tanaman jarak pagar tidak tahan genangan, jika hal ini
Fisiologi Pemangkasan
Syarat mutlak sebagai dasar untuk melakukan pemangkasan adalah harus
memahami aspek fisiologi pertumbuhan tanaman. Ada dua cara tanaman tumbuh
(1) pertumbuhan primer, yaitu peningkatan panjang pucuk (length of shoots) dan
akar yang menyebabkan peningkatan tinggi dan lebar kanopi, (2) pertumbuhan
sekunder, yaitu peningkatan ukuran (thickness) batang dan akar. Kedua tipe
pertumbuhan tersebut membutuhkan pembelahan sel yang diikuti pembesaran dan
diferensiasi sel (Marini, 2003)
Meristem adalah daerah sel membelah. Menurut Marini (2003) ada dua
tipe meristem tanaman (1) meristem apikal, terletak di ujung setiap pucuk (shoot)
dan akar (root) (Gambar 2). Pucuk dan akar memanjang seperti sel yang
tertumpuk satu dengan yang lainnya. Di belakang daerah pembelahan sel terdapat
daerah pembesaran dan diferensiasi sel untuk membentuk berbagai jaringan. (2)
meristem apikal kecil (small apical meristem) disebut axillary meristem
(meristem ketiak) yang membentuk axillary bud (kuncup/tunas ketiak) yang selalu
dorman sampai sebuah daun yang berhadapan dengannya berkembang penuh.
Sebuah tunas ketiak dapat dorman atau berkembang menjadi cabang lateral
(lateral branch) atau bunga (flower). Pada saat tunas vegetatif diiris secara
membujur tampak bahwa, meristem apikal berada di ujung, primordia daun
menjadi daun, meristem ketiak berkembang menjadi tunas ketiak, dan jaringan
prokambial berkembang menjadi kambium.
Gambar 2. Irisan membujur ujung pucuk tampak meristem apikal, primordia daun dan primordia tunas samping (Marini, 2003).
Primordia daun
Meristem apikal
Tunas sangat penting untuk menunjang pertumbuhan vegetatif dan
pertumbuhan reproduktif pohon. Pemangkasan dan pelatihan pohon buah
melibatkan manipulasi tunas. Memproduksi buah menggunakan berbagai teknik,
termasuk pruning untuk memanipulasi pertumbuhan dan pembungaan. Sering
teknik ini mempengaruhi dormansi tunas sehingga pengetahuan tentang tunas dan
dormansi tunas esensial untuk diketahui untuk memahami bagaimana pruning
mempengaruhi pertumbuhan. Selain itu juga penting untuk mengidentifikasi
berbagai tipe tunas pada pohon, terutama tunas bunga dan tunas vegetatif.
Marini (2003) menyatakan bahwa tunas diklasifikasikan dalam empat tipe
(1) klasifikasi berdasarkan kandungan (isi), dikenal tunas vegetatif yang hanya
berkembang menjadi pucuk vegetatif daun dan tunas bunga hanya memproduksi
bunga. Pohon buah batu (apricot dan cherry) menghasilkan tunas vegetatif dan
tunas bunga. Pohon apel dan pear memproduksi tunas vegetatif dan tunas
campuran (mixed bud). Pucuk daun dan bunga muncul dari tunas campuran. (2)
klasifikasi berdasarkan lokasi, tunas terminal terletak pada ujung pucuk. Pada
pohon buah batu (stone) tunas terminal adalah tunas vegetatif. Tunas terminal apel
dan pear selalu vegetatif, walaupun beberapa varietas seperti Rome Beauty secara
terminal memproduksi tunas campuran. Tunas lateral dibentuk dari ketiak daun
yang sering disebut tunas ketiak. Pohon buah stone (batu) tunas lateral dapat
membentuk vegetatif atau bunga. Buku (node) pada tajuk yang berumur satu
tahun mempunyai satu sampai tiga tunas, sebagian dapat membentuk bunga dan
yang lainnya membentuk tunas vegetatif. Tunas bunga berkarakter lebih besar
dengan ujung yang relatif bulat, sedangkan tunas vegetatif lebih kecil. (3)
klasifikasi berdasarkan struktur tunas pada batang, struktur tunas mempengaruhi
struktur cabang pohon buah dan bentuk pohon. Buku terdapat pada batang di
mana daun melekat (Gambar 3a). Pada tanaman apel hanya ada satu daun yang
melekat pada buku, sedangkan pada tanaman peach terdapat tiga daun. Tunas
opposite, ketika dua tunas/cabang, menempati tempat yang berlawanan pada buku
sama. Tunas alternate, ketika hanya ada satu tunas/cabang dari setiap buku dan
tidak ada tunas yang menempati tempat yang sama pada batang, seperti yang satu
(4) Klasifikasi berdasarkan aktivitas, tunas akan dorman ketika tunas tersebut
tidak nyata tumbuh. Ketika ada pemangkasan, tunas yang dorman akan tumbuh.
Gambar 3. Bagian sebuah dahan yang menunjukkan buku dan beberapa tipe tunas (a), struktur tunas alternate (b), dan struktur tunas opposite (c) (Marini, 2003).
Hormon
Hormon adalah zat yang diproduksi dalam jumlah sangat kecil pada satu
bagian tanaman, ditranspor ke bagian lain (Wattimena, 1988, Coombs et al., 1992,
dan Marini, 2003), dan mempunyai efek fisiologi, pertumbuhan dan
perkembangan. Tanaman memproduksi sejumlah hormon yang mengontrol
berbagai aspek pertumbuhan seperti, panjang batang, dormansi tunas dan benih,
pembungaan, fruit set, pertumbuhan dan pemasakan buah, dan respon terhadap
cahaya dan gravitasi. Menurut Marini (2003) hormon promotor adalah giberelin
dan sitokinin dan hormon penghambat adalah auksin dan asam absisik. Hormon
promotor secara umum menyebabkan pertumbuhan tunas, pembelahan dan
perpanjangan sel, dan pertumbuhan batang. Hormon penghambat (inhibitor) selalu
diasosiasikan dengan dormansi, menghambat perkembangan pucuk biji dan tunas,
tetapi dilibatkan dalam induksi tunas bunga. Rasio promotor dan inhibitor lebih
menentukan pertumbuhan tanaman dibandingkan konsentrasi mutlaknya.
Produksi hormon tanaman selalu dikontrol oleh kondisi lingkungan seperti
suhu dan panjang hari. Selanjutnya, dinyatakan bahwa pertumbuhan vegetatif
(a) (b) (c)
Tunas terminal
Tunas lateral
Tunas bunga telah gugur node
selalu diasosiasikan dengan rendahnya rasio inhibitor terhadap promotor dan
dormansi selalu diasosiasikan dengan tingginya rasio inhibitor terhadap promotor.
Dormansi
Dormansi adalah suatu kondisi yang dicirikan dengan berhentinya
pertumbuhan yang sifatnya sementara dan metabolismenya tertekan atau tertahan.
Pada musim dingin pohon tampak tidak tumbuh, tetapi jaringannya tetap hidup
atau aktif, terjadi aktifitas metabolik dan sel berkembang dan berdiferensiasi
secara lambat (Marini, 2003). Selanjutnya Marini (2003) menyatakan para ahli
fisiologi saat ini mendiskripsikan dormansi dalam empat tipe, (1) para-dormansi
terjadi pada dipertengahan dan akhir musim panas ketika tunas tidak tumbuh
sebab inhibitor diproduksi di daun dan tunas terminal menghambat pertumbuhan
tunas ketiak. Para-dormansi dapat diatasi dengan cara menghilangkan daun (leaf
stripping) sepanjang bagian pucuk sehingga tunas ketiak berkembang menjadi
pucuk. Para penangkar bibit (nursery) selalu menggunakan teknik ini untuk
memproduksi pohon dengan cabang lateral. Heading cut dilakukan untuk
menghilangkan sebagian pucuk terminal dan membiarkan beberapa tunas ketiak
tumbuh dan berkembang. (2) ecto-dormansi, terjadi di awal musim gugur sebelum
daun gugur, tanaman tidak tumbuh disebabkan oleh kondisi lingkungan tidak
kondusif untuk pertumbuhan tanaman. Pertumbuhan akan terlihat jika suhu dan
panjang hari cocok (suitable). (3) endo-dormansi terjadi selama musim dingin
sebab tingginya level inhibitor (absisic acid) di dalam tunas. Selama fase
dormansi, pohon tidak akan tumbuh bila kondisi untuk pertumbuhan ideal tidak
terjadi. (4) Eco-dormansi, terjadi di akhir musim dingin pada pertengahan Januari,
setelah persyaratan suhu dingin (chilling) terpenuhi. Pada saat tersebut pohon
tidak tumbuh sebab kondisi tidak mendukung untuk pertumbuhan. Pertumbuhan
akan mulai ketika pohon terekspos pada suhu panas.
Dominansi Apikal
Menurut Cline (2000), dominansi apikal didefinisikan sebagai kendali
yang dilakukan oleh ujung pucuk (shoot apex) terhadap pertumbuhan tunas ketiak
(axillary bud). Konsekuensi morfologinya adalah terhambatnya tunas ketiak
dormancy”. Dominansi apikal berhubungan dengan mekanisme yang dimediasi
oleh auxin dan sitokinin (Cline, 2000) dan status nutrisi pada axillary buds
(Champagnat, 1989).
Dominansi apikal telah dipelajari lebih dari 80 tahun, tetapi mekanismenya
belum dipahami secara jelas, tetapi tampaknya dikontrol oleh konsentrasi relatif
hormon inhibitor dan promotor. Menurut Coombs (1992) dan Marini (2003),
pertumbuhan tunas ketiak dihambat oleh tingginya konsentrasi auksin yang
diproduksi oleh tunas terminal. Auksin bergerak ke bawah pucuk dari sel ke sel
dengan polar, sehingga konsentrasi paling tinggi dekat ujung pucuk. Promotor
diproduksi di akar dan ditransport ke bagian atas pohon. Pertumbuhan tunas
ketiak dapat terjadi pada bagian dasar dari pucuk, di mana konsentrasi hormon
inhibitor secara relatif rendah dan konsentrasi hormon promotor relatif tinggi.
Dominansi apikal dapat dihilangkan dengan cara menghilangkan ujung
pucuk yang berfungsi sebagai sumber auksin (Gambar 4) (Coombs et al., 1992
dan Marini, 2003). Tiga atau empat tunas segera tumbuh di bawah heading cut
selalu berkembang menjadi pucuk. Pinching (memetik pucuk) merupakan sebuah
bentuk dari heading yang akan menginduksi percabangan. Kadang-kandang
dominansi apikal dapat pula dihilangkan dengan penyemprotan pucuk dengan
promotor (giberelin atau sitokinin) sebelum waktu berbunga.
Pada iklim temperate, penelitian proses percabangan pohon buah
difokuskan pada pucuk yang berumur satu tahun (one-year-old shoot) pada
periode musim dingin, karena dominansi apikal dan dormansi tunas (bud
dormansi) terjadi pada periode waktu ini.
(a) (b) (c)
Pembentukan Arsitektur Tajuk Melalui Pemangkasan
Pemangkasan bertujuan untuk membentuk pohon kokoh dan tegar,
memperbanyak percabangan (munculnya daun pada ketiak daun dan pucuk
cabang atau batang), menghindari terjadinya dominasi apikal (penekanan
pertumbuhan calon tunas ketiak daun (lateral) oleh ujung ranting yang aktif
tumbuh akibatnya tanaman memanjang), pemilihan tunas baru yang teratur dan
berpola serta meningkatkan jumlah bunga dan buah pada tanaman yang berbunga
terminal sehingga membentuk kerangka pohon yang dapat mendukung
pembungaan dan pertumbuhan buah (Widodo, 1995).
Pemangkasan dan pelatihan bertujuan untuk mengoptimalkan
penangkapan cahaya untuk mencapai produksi biomassa yang tinggi (Jackson,
1980), membuka ruang kanopi untuk menangkap cahaya (Lakso, 1994) sehingga
memperbaiki distribusi cahaya di antara struktur pembuahan (Lakso dan
Corelli-Grappadelli 1992; Wunsche dan Lakso, 2000) dan memperkecil variasi kualitas
buah, mengoptimalkan biomassa yang dipartisikan ke pucuk buah, seperti pada
apel (Lespinasse dan Delort, 1993), alpokat (Thorp and Stowell, 2001), dan
mengurangi ketidakseimbangan antara organ sink, serta menstimulasi
pertumbuhan pada kiwi (Miller et al., 2000).
Secara umum, ada 2 konsep untuk mendefisinikan sistem percabangan.
Pertama konsep ”Organized plan” menjelaskan level hierarchic antara susunan
cabang pada pohon (Costes et al., 2006). Pola hierarchic diperkenalkan untuk
mengindikasikan sebuah hierarchy antara pucuk utama (main shoot) dan
lateralnya secara berturut-turut. Kedua konsep ”excurrent (cabang lateral) versus
decurrent (batang utama tidak dapat dibedakan dengan cabang yang paling
tinggi)” telah diperkenalkan dalam hubungannya dengan dominansi apikal pada
tanaman pohon hutan (Brown et al. 1967). Pola ini merujuk definisi batang utama
menghasilkan cabang lateral atau batang utama yang terbentang tidak dapat
dibedakan dengan cabang yang paling atas (decurrent).
Berdasarkan intensitas pemangkasan dikenal beberapa istilah
pemangkasan diantaranya : Tipping/pinching (memangkas atau memetik pucuk
ranting), cutting back (memangkas sebagian cabang), stubbing (memangkas
cerutu”), dan thinning (penjarangan cabang dengan cara memotong tepat pada
pangkalnya) (Widodo, 1995).
Menurut Widodo (1995) berdasarkan intensitas untuk tujuan pelatihan
tajuk (training) jenis pangkasan di bagi dua, yaitu heading back (pemotongan
batang, cabang atau ranting) dan thinning out (pembuangan cabang untuk
menjarangkan percabangan yang rapat).
Pada prinsipnya perlu ada penghematan bahan fotosintat sewaktu pohon
aktif memproduksi fotosintat, perlu efisiensi sistem jaringan dalam tubuh tanaman
agar hasil asimilasi (fotosintat) yang ada setelah digunakan untuk perawatan
tanaman itu sendiri cukup untuk membentukan bunga dan buah. Efisiensi pada
tanaman ini tidak dengan cara mengurangi fotosintat melainkan dengan menekan
pemborosannya. Caranya dengan memangkas bagian yang bersifat negatif (hanya
menyerap dan tidak menyumbangkan fotosintat sama sekali) atau dengan
mengurangi (memangkas) bagian pengguna fotosintat, seperti daun-daun yang
ternaungi, cabang-cabang yang tidak produktif dan cabang-cabang yang saling
tumpang tindih.
Jumlah cabang primer dan sekunder akan menentukan jumlah bunga, buah
dan biji jarak pagar. Oleh karena itu pemangkasan tajuk yang teratur dan berpola
dengan merujuk pada jumlah cabang primer dan sekunder akan membentuk tajuk
dan cabang yang ideal untuk meningkatkan produktivitas tanaman jarak pagar.
Bunga terminal, seperti jarak pagar, membutuhkan penyiapan tempat
berbunga (bearing unit) yang sebanyak-banyaknya dan diikuti dengan perakaran
pohon yang baik agar dapat menyangga buah yang lebat. Pembentukan tajuk jarak
pagar diperlukan untuk per tanaman agar tajuk tempat keluarnya bunga dan buah
dapat terbentuk, tetapi dengan percabangan yang kompak. Umumnya rumus
pangkas bentuk 3-9-27 memberikan hasil yang terbaik untuk tanaman berbunga
terminal. Setelah tipe tajuk yang cocok untuk menyediakan tempat pembungan
banyak terbentuk, maka pemangkasan selanjutnya hanya berupa pemeliharaan
bentuk dan kebersihan tajuk. Ranting membawa bunga pada pohon yang berbunga
pada terminal perlu dipangkas setelah pemanenan.
Pola tajuk membuka (open center) dapat meningkatkan pemerataan
maksimum. Fotosintesis netto merupakan ukuran produksi asimilat yang
dimanifestasikan sebagai pertambahan bobot bahan kering total atau laju tumbuh
absolut (LTA), laju tumbuh relatif (LTR) dan laju asimilasi bersih atau LAB (net
assimilation rate, NAR) merupakan komponen fisiologi khususnya daun yang
menyumbangkan pertambahan bobot kering dan merefleksikan fungsi bentuk
tajuk dalam proses asimilasi (Lambers, 1987). Analisis pertumbuhan LAB dapat
disederhanakan sebagai pertambahan bobot bahan kering (dry weight basis) per
satuan luas daun sebagaimana tinjauan Wilson (1981). Pengertian LAB yang
sesungguhnya menyatakan hasil CO2 netto (Sitompul dan Guritno, 1995).
Intersepsi cahaya berperan penting terhadap pertambahan asimilat total
dan partisi asimilat ke arah sink (Gifford et al., 1984). Pada tanaman perkebunan,
kakao dan kopi (Ramaiah dan Venkataramanan, 1987 dan zaitun (Stuttle dan
Martin, 1986) partisi bahan kering ke cabang lateral relatif tinggi. Secara teoritis
menurut tinjauan Ryugo (1988) partisi demikian terjadi karena cahaya matahari
pagi yang kaya infra merah mendorong sintesis sitokinin dan menghambat
translokasi karbohidrat dari cabang ke batang karena jaringan kayu cabang-cabang
yang memperoleh training meregang sehingga cabang merupakan sink yang lebih
kuat dibandingkan batang. Pada masa reproduktif cabang merupakan source yang
baik.
Faktor-Faktor yang Mempengaruhi Laju Fotosintesis
Menurut Gardner et al. (1991) laju fotosintesis dipengaruhi oleh
berbagai faktor diantaranya adalah (1) cahaya, (2) karbondioksida (CO2), (3) suhu, dan (4) status air. Radiasi surya yang diterima daun untuk digunakan dalam
fotosintesis hanya fraksi dalam panjang gelombang 400-700 nm yang dikenal
dengan PAR.
Intensitas cahaya tinggi mendukung terjadinya konduktansi stomata
terhadap CO2 sehingga mempunyai pengaruh sangat besar terhadap laju fotosintesis maksimum. Pada intensitas cahaya rendah hampir tidak ada
penyerapan CO2 karena laju penyerapan CO2 melalui fotosintesis lebih rendah dari pada laju evolusi CO2 dari respirasi mitokondria. Hal ini sejalan dengan pendapat Bauer et al. (1997) menyatakan bahwa intensitas cahaya rendah (PAR
pertukaran karbon. Stomata umumnya membuka pada siang hari dan menutup
pada malam hari, hal ini digunakan untuk meningkatkan kinerja fotosintesis. Laju
fotosintesis secara tidak langsung juga dipengaruhi oleh keadaan stomata
(Salisbury dan Ross, 1995. Mohr dan Schopfer (1995) menyatakan bahwa
pembukaan stomata tanaman berkorelasi tinggi dengan laju fotosintesis. Proses
pembukaan stomata secara langsung merupakan fungsi cahaya karena sel penjaga
memiliki klorofil. Kecepatan pembukaan dipengaruhi oleh jenis cahaya, yaitu
cahaya merah dan biru. Berbeda dengan organel fotosintesis yang memerlukan
cahaya merah untuk laju optimum, stomata lebih membutuhkan cahaya biru untuk
pembukaan stomata. Perbedaan kepekaan antara fotosintesis dengan pembukaan
stomata diduga dipengaruhi oleh karakter klorofil sel penjaga (Salisbury dan Ross,
1995).
Pada cahaya penuh, faktor tahanan stomata pada sebagian besar tanaman
bukan merupakan faktor pembatas laju difusi CO2, namun lebih dikendalikan oleh reaksi enzimatis dalam kloroplas (Mohr dan Schopfer, 1995). Konduktansi
stomata adalah jumlah CO2 yang dapat masuk melalui hambatan stomata, semakin kecil hambatannya akan semakin besar konduktansinya. Semakin banyak jumlah
stomata, konduktansi per satuan luas daun akan semakin tinggi demikian juga
semakin lebar bukaannya (Mohr dan Schopfer, 1995). Selanjutnya dinyatakan
bahwa stomata merupakan satu-satunya jalan untuk fiksasi CO2. Santrucek dan Sage (1996) menduga bahwa penurunan konduktansi stomata pada intensitas
cahaya rendah kemungkinan disebabkan oleh jumlah stomata per satuan luas daun
yang rendah.
Secara umum laju fotosintesis meningkat secara linear dengan
bertambahnya konsentrasi CO2 dalam ruang antar sel (Ci) pada tingkat konsentrasi CO2 interseluler yang rendah, sebab RUBP (ribulase bisphosphate) tidak menjadi pembatas. Pada tingkat konsentrasi CO2 interseluler tinggi, laju fotosintesis mulai menurun sesuai dengan penurunan kemampuan memproduksi RUBP yang tidak
sebanding dengan meningkatnya penyediaan CO2. Laju pertukaran karbon dapat digunakan untuk menghitung akumulasi bahan kering tanaman. Nilai laju
karbon akan semakin tinggi, demikian pula sebaliknya. Oleh karena itu nilai laju
pertukaran karbon yang tinggi pada tanaman ditandai dengan hasil asimilat karbon
yang relatif lebih tinggi.
Suhu lingkungan yang bertambah akan meningkatkan laju fotosintesis
karena peningkatan aktivitas enzim yang mempertinggi kapasitas pemanfaatan
CO2. Fiksasi CO2 merupakan reaksi yang dikendalikan oleh enzim, dan fiksasi CO2 ini meningkat seiring dengan peningkatan aktifitas enzim akibat meningkatnya temperatur hingga mencapai temperatur yang menyebabkan
terjadinya denaturasi enzim-enzimnya
Keadaan stress air akan mendorong penutupan stomata sehingga
mengurangi difusi CO2 dan konsentrasi Ci yang pada gilirannya menurunkan laju fotosintesis. Hanya sekitar 0.1 % dari jumlah air total digunakan oleh tumbuhan
untuk fotosintesis. Transpirasi meliputi 99 % dari seluruh air yang digunakan oleh
tumbuhan; kira-kira hanya 1 % yang digunakan untuk membasahi tumbuhan,
mempertahankan tekanan turgor, dan memungkinkan terjadinya pertumbuhan.
Pengaruh utama kekurangan air terhadap laju pertukaran karbon, yaitu pada
peningkatan tahanan stomata karena tertutupnya stomata. Bila kekurangan air
semakin parah, tahanan mesofil juga akan meningkat karena adanya kerusakan
permanen pada peralatan fotosintesis (Gardner et al., 1991).
Pertumbuhan merupakan resultante dari integrasi berbagai proses fisiologi
dalam tubuh tanaman bersama dengan faktor luar (Sitompul dan Guritno, 1991).
Menurut Fisher (1984) pertumbuhan merupakan proses total yang mengubah
bahan mentah (CO2, zat-zat mineral, air, dan radiasi matahari) secara kimia dan menambahkannya dalam tanaman. Manifestasi pertumbuhan dinyatakan dalam
peningkatan ukuran secara permanen (Taiz dan Zeiger, 2002).
Menurut Sinclair (1994), ketersediaan cahaya matahari menentukan batas
maksimal hasil tanaman karena radiasi yang diintersepsi menyediakan energi
untuk fotosintesis. Laju perolehan massa pada tanaman tergantung jumlah energi
cahaya yang diabsorpsi oleh tanaman dan efisiensi penggunaannya untuk
menggunakan gas CO2 di atmosfir untuk proses fotosintesis dalam memproduksi bahan kering. Menurut Charles-Edward et al. (1986) jumlah energi cahaya yang
proporsi cahaya datang yang diabsorpsi, yang merupakan fungsi sederhana dari
indeks luas daun dan intersepsi cahaya oleh kanopi yang tergantung dari beberapa
faktor seperti arsitektur kanopi daun, inklinasi dan orientasi komponen daun.
Proporsi energi cahaya terintersepsi diabsorpsi oleh sel-sel di dalam daun yang
mengandung organel fotosintesis aktif, yaitu kloroplas.
Charles-Edward et al. (1986) mengemukakan bahwa proporsi energi
cahaya yang diabsorpsi untuk digunakan pada fotosintesis secara nyata juga
dipengaruhi oleh kerapatan dan distribusi sel-sel kloroplas di dalam volume daun.
Selanjutnya Salisbury dan Ross (1995) dan Jones (1992) mengemukakan bahwa
naungan menyebabkan terjadinya perubahan kandungan klorofil daun. Daun yang
ternaungi akan memiliki kandungan klorofil yang lebih tinggi, terutama klorofil b.
Menurut Elfarisna (2000) peningkatan kandungan klorofil a, klorofil b dan
penurunan rasio klorofil a dan b merupakan salah satu mekanisme adaptasi
tanaman yang mengalami cekaman naungan.
Sebagian besar klorofil terdistribusi dalam daun akan tetapi
penyebarannya tidak merata, banyaknya klorofil pada pangkal daun akan berbeda
dengan ujung, tengah serta kedua tepi daun. Rupp dan Traenkle (1995)
mengemukakan bahwa besarnya kandungan klorofil dipengaruhi oleh umur daun,
kandungan klorofil akan meningkat sejalan dengan bertambahnya umur daun.
Peningkatan tersebut terjadi karena selama pertumbuhan daun dari awal
pembentukannya sampai pada umur tertentu, daun tanaman melakukan biosintesis
klorofil. Akan tetapi peningkatan ini akan terhenti pada saat daun tanaman
mengalami penuaan (Mohr dan Schopfer, 1995) karena penuaan daun akan
menyebabkan degradasi klorofil.
Norman dan Arkerbauer (1991) mengemukakan bahwa akumulasi
pertumbuhan tergantung dari total karbon yang difiksasi oleh fotosintesis. Fraksi
dari karbon tersebut dapat dikonversi ke dalam bobot kering walaupun hanya
sebagian karbon yang difiksasi untuk fotosintesis ada pada bobot kering tanaman
dan sebagian lagi karbon hilang melalui respirasi tanaman.
Charles-Edward (1982) dan Hale dan Orcutt (1987) mengemukakan
bahwa secara umum daun-daun yang tumbuh pada lingkungan dengan tingkat
dibandingkan daun yang tumbuh pada tingkat cahaya yang lebih tinggi. Menurut
Taiz dan Zeiger (2002) dan Salisbury dan Ross (1995) hal ini disebabkan oleh
pengurangan lapisan palisade dan sel-sel mesofil pada daun. Selanjutnya Hale
dan Orcutt (1987) mengemukakan bahwa cara ini untuk mengurangi penggunaan
metabolit dan mengurangi jumlah cahaya yang ditransmisikan dan yang
direfleksikan.
Jika daun terlalu banyak, daun-daun bagian bawah tidak menerima cahaya
yang cukup untuk fotosintesis bersih sehingga daun-daun tersebut hanya berfungsi
sebagai sink. Jika indeks luas daun kumulatif mencapai level yang sangat tinggi,
respirasi daun-daun bagian bawah akan seimbang dengan fotosintesis daun-daun
bagian atas, akibatnya laju asimilasi bersih dan laju tumbuh tanaman menurun
sampai nol. Sitompul dan Guritno (1995) mengemukakan bahwa semakin tinggi
kerapatan di antara daun akan menyebabkan semakin sedikit cahaya yang sampai
ke lapisan daun bawah. Nilai indeks luas daun (ILD) > 1 menggambarkan adanya
saling menaungi di antara daun pada lapisan bawah tajuk serta mendapat cahaya
yang kurang sehingga menyebabkan laju fotosintesis yang lebih rendah
dibandingkan yang tidak ternaungi. Akan tetapi nilai ILD < 1 tidak berarti tanpa
naungan karena tergantung pada posisi dan bentuk daun.
Pada prinsipnya tanaman secara fisiologis dan morfologis mampu
beradaptasi pada kondisi intensitas cahaya rendah. Kemampuan adaptasi tanaman
ini secara indigenus karena adanya mekanisme di dalam tanaman akibat pengaruh
lingkungan. Tanaman yang ternaungi mengandung klorofil a dan b empat sampai
lima kali lebih banyak per unit volume kloroplas dan mempunyai rasio klorofil b/a
yang lebih besar dibandingkan tanaman cahaya penuh (Lawlor, 1987).
Individu daun dan kanopi daun tanaman berfungsi sebagai penangkap
cahaya, mengabsorbsi cahaya yang datang dan mengubahnya ke dalam bentuk
energi kimia yang stabil dan dapat disimpan. Energi cahaya yang dapat diserap ini
digunakan untuk fotosintesis untuk menghasilkan karbohidrat. Jika karbohidrat ini
hanya disimpan di jaringan daun maka struktur tanaman non fotosintetik atau
struktur yang fotosintesis rendah seperti akar, batang, dan bunga tidak dapat
berkembang atau berkembang sangat lambat. Oleh karena itu, karbohidrat harus
metabolisme sehingga terjadi proses pertumbuhan dan produksi tanaman. Wright
(1989) mengemukakan bahwa fotosintat hasil fotosintesis kanopi merupakan
sumber karbohidrat yang akan ditranslokasikan ke organ buah, batang, daun, dan
akar. Jumlah alokasi karbohidrat ke masing-masing organ tersebut tergantung dari
aktivitas organ spesifik. Pada tanaman, biji merupakan sink paling kuat, diikuti
daging buah, pucuk dan daun yang sedang tumbuh, kemudian kambium, akar dan
organ penyimpanan lainnya. Oleh karena itu pada saat pertumbuhan buah akan
terjadi peralihan arah pergerakan hasil fotosintesis. Selanjutnya Cline (1997)
mengemukakan bahwa pada tanaman pohon, batang pun merupakan sink terhadap
asimilat yang dihasilkan cabang lateral dan kekuatan ini dikendalikan oleh suatu
III. ANALISIS PERCABANGAN DAN MODEL TAJUK JARAK
PAGAR (
Jatropha curcas
L.)
Analysis of branches and shoot model of Jatropha curcasL.
Abstract
The objective of this research was to analyze pattern of branching, shoot model, and flower form on Jatropha curcasL. This research was conducted using Randomized Complete Block Design with treatment of pruning height from soil i.e., (T0) control (without pruning), T20 (20 cm from soil), T30 (30 cm from soil), and T40 (40 cm from soil). The results showed that branching pattern of Jatropha was spiral. Branch grow from terminal bud to have the character of dichotom. Pruning increased number of primary branches as compared to control. Control plant (T0) shoot model was conical and the pruned plant was columnar. Flowering of Jatropha was terminal. The next flower from the new branch needed ten to seventeen leaves to support. If branching was vigor, three to four inflorescence (flower or fruit) emerged in the same branch. Branching of Jatropha was formed through two ways i.e., (1) before plant flowering, (2) after plant flowering
Key words : height of pruning, shoot model, pattern of branching
PENDAHULUAN
Latar Belakang
Arsitektur tanaman merupakan hasil dari aktivitas meristem. Menurut Bell
(1991), bentuk tanaman berbeda karena perbedaan organ morfologi dan
konstruksi organisasinya. Semua organ tanaman terbentuk dari sel