TERH
T
HADAP RA
Termitidae
SE
INS
AYAP Ma
e) PADA T
(Jatrop
MUHAM
EKOLAH
STITUT P
acrotermes
TANAMA
opha curca
MMAD SA
H PASCAS
ERTANIA
BOGOR
2012
s gilvus Ha
AN JARAK
as L.)
AYUTHI
SARJANA
AN BOGO
agen (Isop
K PAGAR
A
OR
Dengan ini saya menyatakan bahwa disertasi dengan judul”Kajian Cendawan Entomopatogen Metarhizium brunneum Petch sebagai Agens Hayati terhadap Rayap Macrotermes gilvus Hagen (Isoptera: Termitidae) pada Tanaman Jarak Pagar (Jatropha curcas L.)” adalah karya saya sendiri dan belum pernah diajukan dalam bentuk apapun kepada perguruan tinggi manapun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan di dalam Daftar Pustaka pada bagian akhir disertasi ini.
Bogor, Januari 2012
Muhammad Sayuthi
MUHAMMAD SAYUTHI.Study of Entomopathogenic Fungus Metarhizium brunneum Petch as Biocontrol Agent Against Macrotermes gilvus Hagen (Isoptera: Termitidae) in Castor (Jatropha curcas L.) Plantantion. Under Supervision of TEGUH SANTOSO, IDHAM SAKTI HARAHAP and UTOMO KARTOSUWONDO.
Macrotermes gilvus Hagen is primary pest of castor (Jatropha curcas L.), as the termites can damage roots and stems. The purposes of this research were: (1) to estimate population size of colony in field stations at KIJP Pakuwon, (2) to study M. gilvus foraging range, (3) to study the symptomatology and lethal time of M. brunneum as biocontrol agent to M. gilvus in the laboratory, (4) to study the effectiveness of the M. brunneum Petch as biological control agents against M. gilvus in KIJP Pakuwon. The study was conducted from November 2009 to October 2010 in the insect pathology laboratory and insect taxonomy laboratory Departement of Plant Protection, Bogor Agricultural University (IPB), Zoology Laboratory LIPI Cibinong and in the KIJP Pakuwon. Laboratory test was designed using Completely Randomized Design (CRD) and the data were subject to probit analysis. Triple mark recapture methods were used to estimate termite population in the field. In the laboratorium the effective density of conidia as biotermiticide was 1,21x106 konidia/mL, and this led to mortality of M. gilvus up to 85,45%, while the density of 1,08x106conidia/mL resulting in mortality of 78,63%. The suspension of fungi at density 1,21x106conidia/mL was poured at each experimental station (150 mL/station). The result showed that in block I (15.210 m2), block II (5.700 m2), block III (27.000 m2), 8, 1 and 15 termite colonies have been detected respectively from which, 150.388, 59.219, and 149.459 individus were found. In block I, the termites maximum foraging range as far as 140,5 m, as compared to 140 m in block III. In all blocks, we noted the significant decrease of termite population after application of M. brunneum, from initial population 359.066 individus to 15.015 individus.
MUHAMMAD SAYUTHI. Kajian Cendawan Entomopatogen Metarhizium brunneum Petch sebagai Agens Hayati terhadap Rayap Macrotermes gilvus Hagen (Isoptera: Termitidae) pada Tanaman Jarak Pagar (Jatropha curcas L.).Di bawah bimbingan TEGUH SANTOSO, IDHAM SAKTI HARAHAP dan UTOMO KARTOSUWONDO.
Rayap Macrotermes gilvus Hagen merupakan salah satuhama penting tanaman jarak pagar yang merusak sistem perakaran pada pangkal batang tanaman, dan mengakibatkan tanaman terluka, patah dan akhirnya mati. Serangan hama ini semakin lama semakin meningkat hingga meluas pada tanaman sehat lainnya.Selama ini pengendalian rayap di KIJP Pakuwon dilakukan dengan menggunakan insektisida yang disiramkan disekitar perakaran tanaman yang dapat menimbulkan dampak negatif terhadap lingkungan.Oleh karena itudiperlukancarapengendalian lain yang ramah lingkungandengan memanfaatkan agens hayati. Cendawan entomopatogen Metarhizium brunneum mempunyai patogenisitas lebih tinggi dibandingkan beberapa cendawan entomopatogen lainuntuk mengendalikan hama ordo Isoptera. Informasi mengenai pemanfaatan cendawan M. brunneum sebagai agens biokontrol yang berpotensi terhadap hama rayap M. gilvuspada tanaman jarak pagar belum pernah dilaporkan. Penelitian ini bertujuan: (1) Menduga ukuran populasi koloni rayap M. gilvus dari setiap koloni pada stasiun pengamatan di KIJP Pakuwon, (2) Mempelajari daya jelajah rayap M.gilvus di KIJP Pakuwon, (3) Mempelajari simtomatologi dan waktu kematian rayap M. gilvussetelah diinfeksi oleh cendawan M. brunneum sebagai biotermitisida di laboratorium, (3) Mempelajari keefektifan cendawan M. brunneum sebagai agens biokontrol terhadap hama rayap M. gilvuspada tanaman jarak pagar di KIJP Pakuwon.Penelitian ini dilakukan di laboratorium Patologi Serangga, Laboratorium Taxonomi Serangga Departemen Proteksi Tanaman, Faperta IPB, Laboratorium Zoologi LIPI Cibinong dan di Kebun Induk Jarak Pagar (KIJP) Pakuwon Sukabumi Jawa Barat,sejak bulanNopember 2009 sampai Oktober 2010. Rancangan acak lengkap (RAL) digunakan untuk penelitian laboratorium sedangkan metode triple mark recapture technique untuk penelitian di lapangan.
Hasil pengamatan menunjukkan bahwa spesies rayap M. gilvussebagai hama dominan di KIJP Pakuwon. Di Blok I dengan luas areal 15.210 m2didapatkan 8 koloni dengan jumah 150.388 individu. Di Blok II dengan luas areal 5.700 m2 didapatkan 1 koloni dengan jumlah 59.219 individu. Di Blok III dengan luas areal 27.000 m2didapatkan 15 koloni dengan jumlah 149.459 individu. Daya jelajah maksimum yang di lepas dari blok II, dan seminggu kemudian diamati pada blok I, sejauh 140,5 m dan pada blok III sejauh 140 m.
©Hak cipta milik IPB, tahun 2012
Hak cipta dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan yang wajar IPB
TERHADAP RAYAP Macrotermes gilvus Hagen (Isoptera:
Termitidae) PADA TANAMAN JARAK PAGAR
(Jatropha curcas L.)
MUHAMMAD SAYUTHI
Disertasi
Sebagai salah satu syarat untuk memperoleh gelar Doktor pada
Program Studi Entomologi-Fitopatologi
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Pagar (Jatropha curcas L.) Nama : Muhammad Sayuthi NRP : A461060031
Disetujui Komisi Pembimbing
Dr. Ir. Teguh Santoso, DEA. Ketua
Dr. Ir. Idham Sakti Harahap, M.Si. Prof. Dr. Ir. Utomo Kartosuwondo, M.S. Anggota Anggota
Mengetahui
Ketua Program Studi Dekan Sekolah Pascasarjana Entomologi / Fitopatologi
Puji syukur penulis panjatkan kehadirat Allah SWT, karena atas berkat dan rahmat-Nyalah, penulis telah dapat menyelesaikan penelitian dan penulisan disertasiyang berjudul “Kajian Cendawan Entomopatogen Metarhizium brunneum Petch sebagai Agens Hayati terhadap Rayap Macrotermes gilvus Hagen (Isoptera: Termitidae) pada Tanaman Jarak Pagar (Jatropha curcas L.)”
Terima kasih penulis ucapkan kepada :
1. Dr. Ir. Teguh Santoso, DEA, selaku Ketua Komisi Pembimbing, serta Dr. Ir. Idham Sakti Harahap, M.Si, dan Prof. Dr. Ir. Utomo Kartosuwondo, M.S, masing-masing sebagai Anggota Komisi Pembimbing, yang selalu mengarahkan, membimbing, memberi saran atau masukan, serta memotivasi, dan memberi bantuan sarana maupun prasarana yang memadai kepada penulis mulai dari penyusunan proposal penelitian dan pelaksanaan penelitian hingga sampai penyusunan disertasi ini.
2. Dekan Sekolah Pascasarjana IPB, Ketua Program Studi Entomologi-Fitopatologi (Dr. Ir. Sri Hendrastuti Hidayat, M.Sc.) dan Ketua Program Studi Mayor Entomologi (Dr. Ir. Pudjianto, M. Si) serta seluruh staf pengajar di Departemen Proteksi Tanaman IPB atas motivasi dan dorongannya kepada penulis untuk menyelesaikan studinya. Kepada Dr. Ir. Pudjianto, M.Si dan Dr. Ir. Haryadi, M.Si, penulis ucapkan banyak terima kasih sebagai penguji luar komisi pada Ujian Prelim Lisan.
3. Kedua orang tua saya tercinta, isteri dan kedua putra saya tercinta, kedua mertua saya tercinta, kakanda saya tercinta (Prof. Dr. Faisal A. Rani, SH, M.Hum dan Keluarga, Dra. Hj. Rosnaini, Dra. Hj. Nurazimah dan Keluarga, Taufik, A.Md dan Keluarga, Hj. Kartini, S.Ag, dan adinda saya tercinta ( Hastuti dan Keluarga, H. Isra Fahlevi, dan dr. Hj. Fitriani) yang selalu memberi dukungan moril maupun materil kepada penulis untuk menyelesaikan Studi Program Doktor di IPB.
4. Rektor, Dekan Fakultas Pertanian Universitas Syiah Kuala yang telah memberi rekomendasi kepada penulis untuk melanjutkan studi Program Doktor di Institut Pertanian Bogor. Staf Pengajar Fakultas Pertanian Universitas Syiah Kuala Banda Aceh (Dr. Ir. Afizar, DAA, Prof. Dr. Ir. Supardi, M.S, Prof. Dr. Ir. Lukman, M.Si. Prof. Dr. Ir. Sabarauddin, M.Agric.Sc, Dr. Ir. Rosmaidar, M.S, Dr. Ir. Husni, M. Agric. Sc, Dr. Ir. Fajri, M.Sc, Dr. Ir. Syamsuddin, M.Si, Iswadi SPd, Muklis SPd dan lain-lain). 5. Staf Pengajar Universitas Iskandar Muda (UNIDA) Banda Aceh (Prof. Dr.
Drs. Syafi,i Ibrahim, M.S, Dr. Drs. Ridwan, M.Si (Almarhum), Ir. Basyir Ahmad, M.S (Almarhum), Alwi Budiman, S.Si M. Si, Ir. Jauhari, MP., Alwi Ibrahim, S.Si., M.Si.
(2006-2008) yang telah banyak membantu penulis dalam mendukung terhadap kebutuhan yang berhubungan dengan penyelesaian studi Program Pendidikan Doktor (S3) di IPB.
8. Teman-teman mahasiswa/i di Laboratorium Patologi Serangga, Departemen Proteksi Tanaman Fakultas Pertanian (Yunimar, Dendi, Rendi, Teguh, Faishol, Iin, Mira, dan lain-lain) atas bantuan dan kerjasamanya yang baik untuk menyelesaikan studi di IPB.
9. Bapak Endang, Bapak Dede, Bapak Slamet, Ibu Aisyah (Teknisi Laboratoriun di Departemen Proteksi Tanaman) yang dengan tulus iklas telah membantu penulis dalam menyelesaikan studi di Departemen Proteksi Tanaman IPB. 10.Teman-teman Mahasiswa Program Pascasarjana Departemen Proteksi
Tanaman IPB (Bambang Supeno, Edi, Syamsuddin, Yusmani, Iwa, Efi Taufik, Susiati, Rita, Fardedi, Husda, Kikidan lain-lain) yang dengan sukarela selalu meluangkan waktu untuk berbagi ilmu melalui diskusiselama studi di sekolah pascasarjana IPB.
Penulis menyadari bahwa masih banyak kekurangan-kekurangan dalam penulisan disertasi ini, oleh karena itu penulis mengharapkan saran atau masukan dari Komisi Pembimbing untuk membantu kesempurnaan penulisan disertasi ini.
Akhir kata penulis mengucapkan terima kasih banyak kepada Bapak Ketua Komisi dan Anggota Komisi Pembimbing, yang selama ini terus membimbing dan memantau perkembangan studi penulis untuk menyelesaikan Program Pendidikan Doktor di IPB. Semoga Allah SWT memberikan balasan amal baik kepada mereka semua dengan balasan kebaikan yang tak terhingga.Akhirnya semoga karya ilmiah ini bermanfaat.
Bogor, Januari 2012
Penulis dilahirkan pada tanggal 23 Nopember 1972 di Aceh Besar, anak ke enam dengan delapan bersaudara dari pasangan H. Abdul Rani Usman dan Hj. Razian Umar. Penulis menempuh Pendidikan Program Doktor (S3) pada Program Studi Entomologi-Fitopatologi di Institut Pertanian Bogor, tahun 2006-2012. Program Magister (S2) ditempuh di Universitas Padjadjaran Bandung pada Program Studi Ilmu Tanaman dengan Bidang Kajian Utama Ekofisiologi Tanaman, tahun 2000-2003. Pendidikan sarjana (S1) ditempuh di Jurusan Hama dan Penyakit Tanaman Fakultas Pertanian, Universitas Syiah Kuala Banda Aceh, tahun 1991-1996.
Penulis adalah staf pengajar Jurusan Hama dan Penyakit Tanaman Fakultas Pertanian Universitas Syiah Kuala Banda Aceh. Penulis menikah dengan Afrida SP, tahun 2002, dan telah dikaruniai dua orang putra yaitu:
DAFTAR TABEL ... xii
DAFTAR GAMBAR ... xiii
DAFTAR LAMPIRAN ... xvi
BAB I PENDAHULUAN ... 1
Latar Belakang ... 1
Tujuan Penelitian ... 3
Manfaat Penelitian ... 3
BAB II TINJAUAN PUSTAKA ... 5
Biologi Rayap ... 5
Polimorfisme ... Pembentukan Koloni ... 6
Siklus Hidup Rayap ... 7
Perilaku Rayap ... 7
Aktivitas Makan ... 8
Aktivitas Kawin ... 8
Daya Jelajah ... 9
Ekologi Rayap ... 9
Pendugaan Ukuran Populasi ... 11
Teknik Tanda Tangkap ... 11
Teknik Penandaan... 12
Pengendalian Rayap ... 13
Cendawan Metarhizium brunneum Petch ... 14
Jarak Pagar (Jatropha curcasL.) ... 16
BAB.III POPULASI KOLONI RAYAP Macrotermes givus HagenDI KIJP PAKUWON SUKABUMI JAWA BARAT ... 17
Abstrak... 17
Abstract ... 17
Pendahuluan ... 18
Bahan dan Metode ... 19
Hasil dan Pembahasan ... 22
Simpulan ... 25
Abstrak ... 27
Abstract ... 27
Pendahuluan ... 28
Bahan dan Metode ... 28
Hasil dan Pembahasan ... 29
Simpulan ... 33
Daftar Pustaka ... 33
BAB V STUDI SIMTOMATOLOGI DAN WAKTU KEMATIAN RAYAP Macrotermes gilvus Hagen (Isoptera:Termitidae) SETELAH INFEKSI OLEH CENDAWAN Metarhizium brunneum Petch ... 35
Abstrak ... 35
Abstract ... 35
Pendahuluan ... 36
Bahan dan Metode ... 37
Hasil dan Pembahasan ... 40
Simpulan ... 47
Daftar Pustaka ... 48
BAB.VI KEEFEKTIFAN CENDAWAN ENTOMOPATOGEN Metarhizium brunneum Petch SEBAGAI BIOTERMITISIDA TERHADAP RAYAP Macrotermes gilvus Hagen DI KIJP PAKUWON SUKABUMI JAWA BARAT ... 51
Abstrak ... 51
Abstract ... 51
Pendahuluan ... 52
Bahan dan Metode ... 53
Hasil dan Pembahasan ... 56
Simpulan ... 57
Daftar Pustaka ... 57
BAB VII.. PEMBAHASAN UMUM ... 59
BAB VIII. SIMPULAN DAN SARAN ... 63
Simpulan ... 63
Saran ... 63
No Halaman 1 Spesies rayap M. gilvus dari stasiun pengamatan di KIJP Pakuwon ... 22
No Halaman
1 Perakitan stasiun pengamatan ... 19
2 Stasiun pengamatan dan pemasangannya di KIJP Pakuwon ... 20
3 Rayap Macrotermes gilvus yang telah diwarnai dengan neutral red 0,25 dan nile blue A 0,05% ... 21
4 Rayap M. gilvus: (a) kasta prajurit mayor, (b) kasta prajurit minor, dan (c) kasta pekrja ... 22
5 Ukuran populasi koloni rayap M. gilvus pada blok I ... 23
6 Ukuran populasi koloni rayap M. gilvus pada blok II ... 24
7 Ukuran populasi koloni rayap M. gilvus pada blok III ... 24
8 Daya jelajah rayap M. gilvus di KIJP Pakuwon pada blok I ... 30
9 Daya jelajah rayap M. gilvus pada blok III di KIJP Pakuwon ... 30
10 Kondisi tanaman jarak pagar (J. curcas) pada blok I ... 31
11 Kondisi tanaman jarak pagar (J. curcas) pada blok III ... 31
12 Pengukuran daya jelajah rayap M. gilvus ... 32
13 Rayap M. gilvus: (a) kasta prajurit dan (b) kasta pekerja ... 37
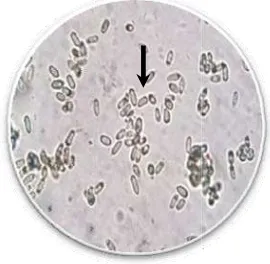
14 Konidia cendawan M. brunneum diamati dengan mikroskop optik pembesaran (400×) ... 38
15 (a) kadaver M. gilvus berubah menjadi gelap setelah diinfeksi oleh konidia M. brunneum (b) miseliumM. brunneum tampak pada bagian tubuh rayap M. gilvus ... 40
16 Pengamatan dengan Scanning Electron Microscope (SEM) pembesaran5000×, yaitu: (a) miselia dari M. brunneum keluar dari internal tubuh inangmelalui lubang alami, (b) miselia hampir menutupi seluruh permukaan tubuh inang ... 41
17 Konidia pada permukaan tubuh rayap M. gilvus diamati menggunakan SEM pembesaran 5000× ... 41
18 Infeksi konidia cendawan M. brunneum terhadap rayap M. gilvus memasuki tahapan destruksi (a) hampir seluruh permukaan tubuh rayap telah tertutupi miselia, dan (b) kadaver rayapkering kehitaman terutama bagian abdomen ... 42
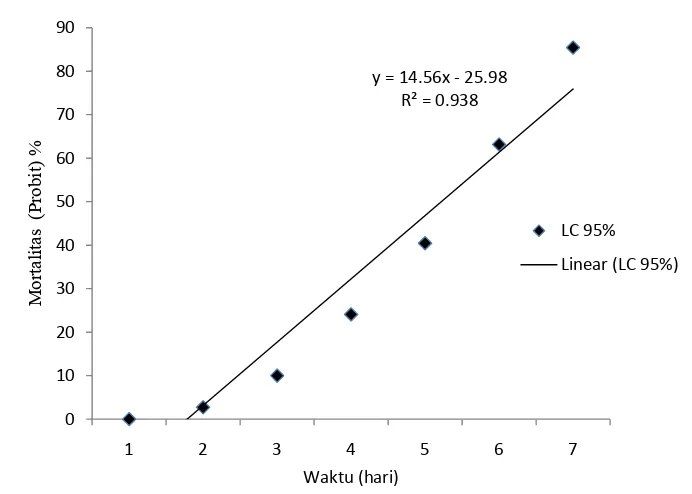
19 Kematian rayap M. gilvus selama 7 hari pengamatanakibat perlakuan konidia cendawan M. brunneum (LC95) ... 46
20 Kematian rayap M. gilvus selama 7 hari pengamatanakibat perlakuan konidia cendawan M. brunneum(LC85) ... 47
21 Populasi koloni Rayap M. gilvus pada blokI ... 55
22 Populasi koloni rayap M. gilvus pada blok II ... 56
No Halaman
1 Bagan alir percobaan ... 71
2 Peta stasiun pengamatan di KIJP Pakuwon ... 72
3 Rekapitulasi data blok I (Percobaan I) ... 73
4 Rekapitulasi data blok II (Percobaan I) ... 75
5 Rekapitulasi data blok III (Percobaan I) ... 75
6 Data hasil pengamatan persentase mortalitas rayap Macrotermes gilvus setelah aplikasi cendawan Metarhizium brunneum dengan kerapatan konidia 1,21×106/mL di laboratorium (Percobaan III) ... 79
7 Data hasil pengamatan persentase mortalitas rayap Macrotermes gilvus setelah aplikasi cendawan Metarhizium brunneum dengan kerapatan konidia 1,08×106/mL di laboratorium (Percobaan III) ... 79
8 Rekapitulasi data blok I(Percobaan IV) ... 82
9 Rekapitulasi data blok II (Percobaan IV) ... 82
BAB I PENDAHULUAN
Latar Belakang
Selama ini Indonesia dikenal sebagai negara pengekspor bahan bakar
minyak dari fosil, namun dengan meningkatnya penduduk dan industri
diperkirakan sepuluh tahun mendatangakan menjadi negara pengimpor bahan
bakar minyak bumi. Oleh karenanya pemerintah perlu memikirkan alternatif
pengganti bahan bakar minyak bukan darifosil tetapi berpotensi untuk
dikembangkan di Indonesia (Hendriadi et al. 2005).
Salah satu sumber energi alternatif yang dapat terbarukan adalah biodisel
dari tanaman jarak pagar (Jatropha curcas L.) yang hanya digunakan sebagai
sumber bahan bakar (Mahmud et al. 2006).Beberapa keunggulan biodisel dari
tanaman jarak pagar yaitu tidak mengandung sulfur, tidak beraroma, dapat
diperbaharui, ramah lingkungan, aman dalam penyimpanan dan transportasi
karena tidak mengandung racun, meningkatkan nilai produk pertanian,
menurunnya ketergantungan suplai minyak dari negara asing, dan mudah terurai
oleh mikroorganisme (Susilo 2006).
Tingkat produktivitas jarak pagar sangat tergantung dari cara
pemeliharaan, lingkungan, sumber benih, ada tidaknya serangan hama dan
penyakit. Produktivitas jarak pagar di berbagai negara, yaitu Nicaragua (5 ton/
ha), Paraguay (4 ton/ha) dan Mali (2,8 ton/ha) (Henning & Reinhard 2000).
Indonesia diperkirakan mampu menghasilkan produktivitas hingga 5 ton biji
kering/ha (Hasnam 2006).
Permasalahan yang dihadapi dalam agribisnis jarak pagar adalah belum
tersedianya varietas unggul, ketersediaan benih sangat terbatas, teknik
budidayanya belum memadai, dan adanya serangan hama dan penyakit (Asbani et
al. 2007). Salah satu hama penting yang merusak tanaman jarak pagar adalah
rayap Macrotermes gilvus Hagen, yang merusak mulai dari akarhingga pada
batang tanaman (Tarumingkeng 2001).
Hasil pengamatan di Kebun Induk Jarak Pagar (KIJP) Pakuwon
jarak pagar mencapai 15 sampai 24% dengan rata-rata 16,33%. Menurut
informasi dari penanggung jawab KIJP Pakuwon,jarak pagar yang ditanamdengan
menggunakan stek ukuran 30 cm di permukaan tanah, 60 sampai 80% terserang
rayap M. gilvus hingga mengalami kematian. Hama ini merusak bagian pangkal
akar hingga batang tanaman dengan membuat tabung kembara dari bahan tanah
yang ditempelkan pada batang tanaman atau dengan cara masuk ke dalam jaringan
tanaman hinggahanyalapisan epidermisyang tersisa. Kondisi ini mengakibatkan
tanaman menjadi patah, roboh dan mengalami kematian. Semakin lama intensitas
serangannya semakin meningkat hingga pada tanaman sehat lainnya. Oleh
karenanya walaupuntingkat serangan hama ini kurang dari 10% tetapi harus
segera dilakukan pengendalian agar tidak menyebar kepada tanaman lain,
sehingga hasil produksi tetap maksimal (Asbani et al. 2007).Selama ini
pengendalian rayap di KIJP Pakuwon menggunakan termitisida sintetik dengan
caradisiramkan sekitar perakaran tanaman yang mengakibatkan pencemaran
lingkungan dan keracunan bagi pengguna (Oka 2005).
Keberadaan agens hayati secara alami yang telah ada di KIJP Pakuwon
kurang memberikan dampak positif terhadap rayap hama M. gilvus. Oleh karena
itu perlu pengendaliancaralain seperti pemanfaatan agens hayati yang mungkin
dapat diterapkan di KIJP Pakuwon.Namun sebelum melakukan pengendalian
terhadap spesieshama ini terlebih dahulu perlu dipelajari ukuran populasi koloni
dan daya jelajah maksimumnya,sehingga populasirayap hama M.gilvus dapat
tereliminasi lebih maksimal.
Dari penelitian sebelumnya dinyatakan bahwa cendawan M. brunneum
memiliki tingkat patogenisitas dan virulensi yang lebih tinggi terhadap serangga
rayap Schedorhinotermes javanicus dibandingkan beberapa spesies cendawan
entomopatogen lain, seperti Metarhizium anisopliae, Beauveria bassiana,
Fusarium oxysporum dan Aspergillus flavus(Desyanti 2007, Ginting 2008).
Penelitian ini mempelajari keefektifan cendawan entomopatogen M. brunneum
dalam menekan ukuran populasi koloni rayap M. gilvus yang menjadi hama
Tujuan Penelitian
Tujuan penelitian ini yaitu:
1. Menduga ukuran populasi koloni rayap Macrotermes gilvus di KIJP
Pakuwon.
2. Mempelajari daya jelajah rayap M.gilvus di KIJP Pakuwon.
3. Mempelajari simtomatologi dan waktu kematian rayap M. gilvussetelah
diinfeksi oleh cendawan M. brunneum sebagai biotermitisida di laboratorium.
4. Mempelajari keefektifan cendawan entomopatogen M. brunneum sebagai
biotermitisida terhadap rayap M. gilvusdi KIJP Pakuwon.
Manfaat Penelitian
Hasil penelitian diharapkan dapat menjadi informasi awal dan sebagai
pedoman dasar untuk penyusunan rekomendasi pengendalian hama rayap
Macrotermes gilvus pada tanaman jarak pagar menggunakan cendawan
BAB II
TINJAUAN PUSTAKA Biologi Rayap
Krishna dan Weesner (1969) menyatakan bahwa rayapdiklasifikasikan ke
dalam 6 Famili(Mastotermitidae, Kalotermitidae, Hodotermitidae,
Rhinotermitidae, Serritermitidae, dan Termitidae). Rayap
tanahMacrotermesgilvus termasuk famili Termitidae sub famili Macrotermitinae,
klasifikasinya adalah sebagai berikut:
filum: Arthropoda
kelas: Insecta
sub-kelas: Pterigota
ordo: Isoptera
famili: Termitidae
sub-Famili: Macrotermitinae
genus: Macrotermes
spesies: Macrotermes gilvus Hagen.
Menurut Krishna dan Weesner (1969) rayap M. gilvushidup berkoloni
yang mempunyaikasta prajurit mayor dan minor.Ciri-ciri kasta prajuritsecara
umumadalah kepala bewarna coklat tua, mandibel berkembang dan berfungsi,
mandibel kiri dan kanan simetris,tidak memiliki gigi marginal, ujung mandibel
melengkung yang berfungsi untuk menjepit. Ujung labrum tidak jelas, pendek dan
melingkar, antena terdiri atas 16-17 ruas. Thapa (1981) dan Tho
(1992)menjelaskan ciri-ciri dari kasta prajuritmayor yaitu kepala bewarna coklat
kemerahan, panjang kepala dengan mandibel 4,80-5,00 mm, lebar kepala
2,88-3,10 mm,antena 17 ruas, ruas ketiga sama panjang dengan ruas kedua dan ruas
ketiga lebih panjang dari ruas keempat. Sedangkan kasta prajurit minor kepala
bewarna coklattua, panjang kepala 1,84-2,08 mm dan lebar 1,52-1,71 mm
sertapanjang kepala dengan mandibel 3,07-3,27 mm. Antena17 ruas, ruas kedua
sama panjangdengan ruas keempat.
Menurut Nandika et al. (2003) rayap M. gilvus banyak tersebar di
berhubungan dengan tanah.Rayap membiakkan cendawan yang berbentuk bunga
karang, serta bangunan-bangunan liat dalam tanah dan untuk menemukan sumber
makanan dengan membuat tabung kembara dari humus atau tanahsebagai jalur
jelajah (Nandika et al. 2003).
Polimorfisme
Polimorfismemerupakan ciri rayap yang hidup secara terorganisir dengan
bentuk, ukuran, dan fungsi yang berbedadalam sebuah koloni, sepertiordo
Isopteraterdiri atas kasta prajurit, kasta pekerja dan kasta reproduktif. Kasta
pekerja bertugas sebagai pencari makan, perawat telur, pembuat dan pemelihara
sarang. Kasta ini pada saat tertentu dapat bersifatkanibalterhadap individu rayap
yang sakit dalam koloninya untuk mempertahankan prinsip efisiensi dan
konservasi energi, serta mengatur keseimbangan koloni (Tarumingkeng 1993,
Tambunan & Nandika 1989).
Kasta prajurit dengan ukuran kepalanyabesar dan mengalami penebalan
pada bagian tersebut serta memiliki mandibelkuat untuk melindungianggota
koloni dari gangguan luar (Tambunan & Nandika 1989). Apabila terjadi gangguan
dari luar, maka kasta prajurit segera menginformasikan kepada anggota kasta
prajurit lain dalam koloninya dengan tanda tertentu, dan semua kasta prajurit
segera menuju sumber gangguan untuk mengatasinya (Harris 2001).
Kasta reproduktif berfungsi untuk bertelur dan jantan membuahi betina,
seperti kasta reproduktifdari rayap Macrotermes spp dapat menghasilkan telur
seminggu setelah melakukan swarming (Harris 1971). Neoten akan muncul bila
kasta reproduktif primer mati atau terpisahdari koloni induk akibat adanya
gangguan luar. Neoten dapat terbentuk beberapa kali dalam jumlah besar sesuai
dengan perkembangan koloni (Richards & Davies 1996).
Pembentukan Koloni
Sebuah koloni rayap dapat terbentuk dari sepasang laron betina dan jantan
dengan melakukan kopulasi, kemudian mencari habitat yang sesuai untuk
membentuk koloni baru (Tarumingkeng 1993). Koloni rayap dapat terbentuk
(2) melalui pemisahan koloni dari koloni utama dengan membentuk kasta
reproduktif suplementer, dan (3) melalui proses migrasi dari sebagian koloni
rayap menuju tempat baru dan koloni yang tertinggal mengembangkan kasta
reproduktif suplementer (Lee & Wood 1971, Harris 1971).
Kasta reproduktif bersayap akan muncul pada musim-musim tertentu,
yang berkumpul dalam koloninya sebelum bersialang (swarming) keluar sarang.
Umumnya beberapa spesies rayap di daerah tropis bersialang pada awal musim
hujan (Lee & Wood 1971). Selama bersialang sepasang imago (jantan dan betina)
bertemu dan segera menanggalkan sayap untuk mencari tempat yang sesuai
(Tambunan & Nandika 1989). Di Amerika Selatan rayap Contrictotermes
cavifronsuntuk membentuk koloni baru melalui fragmentasi koloni dengan
bermigrasi untuk menemukan habitat baru(Krishna & Weesner 1969). Demikian
juga rayap Anoplotermes,Trinervitermes di Afrika (Rismayadi 1999)
danMastotermes darwinensismembentuk koloni baru melaluifragmentasi koloni
(Lee & Wood 1971).
Siklus Hidup Rayap
Rayap mengalami metamorfosis tidak sempurna (paurometabola). Siklus
hidupnya dimulai dari telur, nimfa, dan imago. Nimfa muda yang baru keluar dari
telur dan akan berkembang menjadi kasta pekerja, kasta prajurit, atau alata di
dalam koloninya (Natawigena 1990). Lama siklus hidup rayap dari fase telur
50-60 hari. Ratu rayap Macrotermes sp yang telah berumur 5 tahun mampu
menghasilkan telur hingga 36.000 butir perhari (Hasan 1986). Memasuki instar I
membutuhkan waktu 11-13 hari, instar II (13-18 hari), instar III (16-32 hari),
instar ke IV (30-50 hari), dan instar ke V (14 hari), dan sekali siklus hidup rayap
dibutuhkan waktu 4-6 bulan (Grasse 1984).
Perilaku Rayap
Sebagai serangga sosial, rayap memiliki beberapa prilaku yang khas
(Nandika & Tambunan 1987; Tarumingkeng 2004), yaitu:
Trophallaxis adalahtransfer material (makanan dan protozoa) antara
feedingsedangkan melalui mulut disebut dengan stomodeal feeding. Sifat
trofalaksis merupakan cara memperoleh protozoa flagellatabagi individu yang
baru melakukan ganti kulit (ekdisis), karena pada saat ekdisis integumen
proctodeum tanggal sehingga protozoa simbion yang diperlukan untuk mencerna
selulosa ikut keluar dan diperlukan reinfeksi dengan jalan trophallaxis. Grooming
adalah berkumpul dengan mengosokkan tubuh antara individu dalam sebuah
koloni, dan menjilat bagian tubuhnya yang bertujuan untuk membersihkan diri
dari serangan patogen. Cryptobiotic adalah menyembunyikan dan menghindar
dari cahaya kecuali laron yang menyukai cahaya pada saat swarming. Rayap
hidup dalam tanah dan pada saat mencari makanan dipermukaan tanah,
membentuk tabung kembara dari bahan tanah atau humus. Cannibalistic yaitu
perilaku memakan individu sejenis, seperti kasta prajurit yang lemah tidak dapat
menjaga koloninyasecara efektif, akan dimakan oleh kasta pekerja. Demikian juga
betina dan jantan baik ratu, raja maupun neoten yang tidak mampu memberikan
kontribusi pada koloninya. Nekrofagi yaitu memakan kadaver sesamanya
(Tarumingkeng 2004).
Aktifitas Makan
Sumber makanan rayap berupa selulosa.Terdapat hubungan antara rayap
dengan mikoorganisme simbion pada saluran pencernaan rayap, yaitu protozoa
pada rayap tingkat rendah dan bakteri pada rayap tingkat tinggi. Sumber makanan
rayap pada umumnya dikelompokkan ke dalam dua tipe, yaitu sumber makanan
mentah (crude nutrient) dan sumber makanan dari kasta pekerja. Sumber makanan
mentah berupa tanaman atau pohon hidup, kayu atau tanaman yang sudah mati,
bahan makanan lain seperti humus, rumput, jamur (Nandika et al. 2003).
Aktifitas Kawin
Kopulasi rayap dapat ditandai dengan terbangnya laron (swarming) yang
dipengaruhi oleh perubahan cuaca di luar sarang. Laron akan berkumpul pada
tempat tertentu di dalam sarang yang menunjukkan atraksi tertentu dengan
mencaricahaya. Kemudian menanggalkan sayapnya untuk menemukan
pasangan.Setelah menemukanpasangannya,calon ratuakanberjalan di depan dan
calon raja mengikuti dari belakang untuk menemukan habitat yang sesuai.Setelah
3-8 hari berada pada habitat yang baruditemukan akan berkopulasi (Nandika et
al.2003).
Daya Jelajah
Rayap akanberjelajah untuk menemukan sumber makanan dengan
lingkungan yang optimal. Persentuhan fisik antar individu rayap dan bau yang
dikeluarkan melalui jejaknya merupakan sebuah mekanisme penyampaian
informasi dalam sebuah koloniterhadap sumber makanan yang baru ditemukan.
Rayap Nasutitermes dalam melakukan daya jelajah berhubungan dengan feromon
yang dikeluarkan melalui jejak dan dihasilkan oleh kelenjer sternal yang terdapat
pada abdomen(Krishna & Weesner 1969). Rayap tanah C. formosanus melakukan
daya jelajah hingga mencapai 100 m dengan jumlah individu 1-7 juta individu
perkoloni. Reticulitermes flavivesmenjelajah hingga 79 m dengan 2-5 juta
individu anggota koloni yang mengikutinya (Su 1994).
Ekologi Rayap
Rayap berperan penting dalam siklus biogeochemical (dekomposisi bahan
organik), seperti nitrogen, carbon, oksigen, fosfor, yang dapat meningkatkan
kesuburan tanah, karena mampu mengubah profil tanah, mempengaruhi tekstur
tanah dan mendistribusikan bahan organik (Khrisna & Weesner 1969).
Dengan beralih fungsinya hutan menjadi perkebunan monokultur
menyebabkan rayap ini menjadi hama penting di dunia pertanian khususnya di
bidang perkebunan(Tarumingkeng 2001).Pengembangan sektor perkebunan dan
hutan tanaman industri yang dilakukan pada lahan gambut dan lahan bekas hutan
primer merupakan salah satu penyebab terjadinya serangan rayap terhadap
tanaman perkebunan (Sudohadi 2001).
Serangan hama rayapsulit dideteksi secara dini karena merusak tanaman
mulai dari bagian akar tunggang di dalam tanahdan diketahui setelahmembentuk
ini menyerang tanaman Eucalyptus alba pada umur enam bulan dengan tingkat
kematiannya secara berturut-turut adalah 60% dan 100% di Kebun Percobaan
Darmaga dan Demplot HTI Universitas Winaya Mukti (Nandika et al. 2003).
Serangan rayap hama pada tanaman perkebunan biasanya dipengaruhi oleh
tingkat preferensinya terhadap jenis tanaman, tingkat kesehatan tanaman, dan
kondisi tempat tumbuh, serta tanaman yang tertekan karena serangan patogen,
kerusakan fisik, atau akibat kekurangan air dan unsur hara. Biasanya tanaman
eksotik lebih tinggi tingkat serangan rayap dibandingkan tanaman lokal, hal ini
diduga tanaman lokal dapat mengembangkan mekanisme resistensinya terhadap
gangguan rayap, yaitu melalui proses adaptasi dan evolusi. Tanaman yang berada
di dataran rendah lebih banyak diserang oleh rayap dibandingkan tanaman yang
berada di dataran tinggi. Hal ini berhubungan dengan distribusi rayap yang
dibatasi oleh faktor suhu, kelembaban serta ketinggian tempat (Pribadi 2009).
Aktivitas rayap disuatu daerah dipengaruhi oleh berbagai faktor seperti
tanah, tipe vegetasi, faktor iklim dan ketersediaan air. Faktor-faktor tersebut saling
berinteraksi dan saling mempengaruhi satu sama lain. Kelembaban dan suhu
merupakan faktor yang mempengaruhi aktivitas rayap. Bila kondisi lingkungan
berubah akan mempengaruhi perkembangan, aktifitas dan perilaku rayap
(Nandika et al. 2003).
Curah hujan merupakan salah satu faktor pemicu perkembangan eksternal
yang merangsang keluarnya kasta reproduktif dari sarang. Laron biasanya keluar
pada saat musim hujan. Bila curah hujan terlalu tinggi dapat menurunkan daya
jelajah rayap yang berpengaruh langsung terhadap koloni rayap, terutama bila
sarangnya berada diatas permukaan tanah. Koloni Neotermes tectonae bersarang
di dalam kayu dan terlindungi secaralangsung dari pengaruh curah hujan. Di hutan
jati rayap ini memiliki kisaran suhu optimum 22‐26°C dengan ketinggian 0- 700 m
dpl (Nandika et al. 2003). Rayap Macrotermes spmenyukai kelembaban 75
sampai 90% dan pada saat musim panas melakukan daya jelajah membentuk
tabung kembara menuju habitat dengan suhu yang lebih rendah. Cryptotermes sp
merupakan rayap kayu kering dan dengan suhu optimum 15-38°C dapat
melakukan daya jelajah tanpa memerlukan air atau kelembaban tinggi(Khrisna &
tinggi dibandingkan rayap Reticulitermes flavipes. Bila suhu sangat tinggi rayap
akan berada di bawah permukaan tanah atau masuk dalam sarangnya atau tetap
beradadi permukaan tanah bila terdapat naungan agar mendapatkan suhu
optimum. Di daerah semi gurun dengan penutupan vegetasi yang rendah, rayap
Psammotermes sering ditemukan di bawah batu atau naungan karena dapat
menciptakan suhu dan kelembaban yang lebih baik. Jenis tanaman penutup tanah
juga mempengaruhi suhu tanah. Di lapangan dengan tanaman sereal dapat
memberikan sedikit perlindungan dibandingkan jenis tanaman lain atau semak
(Lee & Wood 1971).
Pendugaan Ukuran Populasi
Menurut Krebs (1978) dua kriteria mendasar yang mempengaruhi
pemilihan metode pendugaan ukuran populasi adalah kerapatan dan mobilitas dari
individu-individu penyusun populasi yang akan diteliti. Pengukuran kepadatan
populasi dapat dilakukan dengandua cara,yaitu (a) kepadatan absolut; jumlah
organisme per unit areal atau volume (b) kerapatan relatif; kerapatan satu populasi
relatif terhadap populasi lainnya (Krebs 1989). Penggunaan metode untuk
memperoleh informasi mengenai ukuran populasi biasanya berhubungan dengan
sifat mobilitas organisme yang diteliti. Untuk organisme yang bergerak umumnya
menggunakan teknik tanda tangkap dan transek garis. Sedangkan untuk organisme
yang tidak bergerak menggunakan metode kuadrat, seperti pada tumbuhan
(Tarumingkeng 2001).
Umumnya ukuran populasi kolonirayap tingkat rendah terdiri atas
beberapa ratus atau beberapa ribu ekor.Rayap Kalotermes flavicollis
menghasilkan 15-20 ekor rayap untuk tahun pertama, namun beberapa tahun
kemudian populasi koloninya menjadi bertambah hingga 600-1.000 ekor
(Richards& Davies 1996).Di Afrikarayap Macrotermes spplebih besar ukuran
populasi koloninya yang terdiri atas beberapa juta ekor(Lee & Wood 1971).
Teknik Tanda Tangkap
Menurut Rismayadi (1999)teknik tanda tangkap pertama kali digunakan
(1996) juga mempelajari ukuran populasi dan daya jelajah rayap tanah dengan
metode teknik tanda tangkap.Asumsi-asumsi yang mendasari semua analisis
teknik tanda tangkap adalah: (1) individu bertanda tidak dipengaruhi oleh
penandaan dan tanda yang digunakan tidak hilang selama periode pengamatan, (2)
individu bertanda bercampursecaraacak dalam populasi, (3) penarikan contoh
dilakukan secaraacak (berdasarkan asumsi ini maka individu dalam populasi dari
kelompok umur dan dari jenis kelamin berbeda akan memiliki peluang tertangkap
berdasarkan perbandingan yang ada dalam populasi dan semua individu
mempunyai peluang yang sama untuk tertangkap dalam habitatnya), (4)
pengambilan contoh dilakukan dalam waktu tertentu (Southwood 1975, Krebs
1978& 1989).
Metode Schnabel menjadi dasar dari teknik yang digunakan oleh Su
(1994) dan Surnnuwat et al.(1996) untuk mempelajari ukuran populasi dan daya
jelajah rayap tanah. Namun dalam penelitian ini untuk pendugaan ukuran populasi
koloni rayap digunakan metode triple mark reupture technique, yaitu:
N = (∑Mi.ni)/[(∑mi)+1]
SE = N/{[1/(∑mi)+1)]}+{(2/((∑mi)+1)2+[(6/(∑mi)+1)3]}1/2 dimana:
N = Ukuran populasi
SE = Simpangan baku
ni = Jumlah keseluruhan rayap yang tertangkap pada penangkapan ke-i
mi = Jumlah rayap bertanda yang tertangkap pada penangkapan ke-i
Mi = Jumlah total rayap bertanda sampai penangkapan ke-i
Teknik Penandaan
Menurut Southwood (1975) bahan penanda yang digunakan oleh para
peneliti terhadap beberapa studi populasi adalah: cat dan larutan bahan pewarna,
penandaan internal dengan penyuntikan, bahan pewarna fluorescent, label,
mutilasi (pemotongan) dan penandaan melalui pengumpanan yang mengandung
bahan pewarna.
Pada penelitian pendugaan ukuran populasi koloni dan daya jelajah rayap
al.1984). Kemudian Su et al. (1991) menggunakan beberapa bahan pewarna
neutral red dan nile blue A, ternyata hasilnya sangat efektif digunakan sebagai
pewarna rayap. Setelah itu bahan pewarna ini telah banyak digunakan untuk studi
populasi rayap. Su (1994) pernah menggunakan bahan pewarna ini untuk
menandai Reticulitermes flavipes dan C.formosanus, juga pernah di pakai untuk
menandai C. gestroi. Hal yang sama juga pernah dilakukan oleh Harahap et al.
(2005) dalam mengamati perilaku agonistik pada rayap R. flavipes dan R.
nirginicus; menggunakan bahan pewarna nile blue A (0,05%) dan neutral red
(0,25%), ternyata sangat efektif digunakan sebagai bahan penandaan rayap.
Pengendalian Rayap
Pengendalian rayap dengan teknik pengumpanan lebih menguntungkan
karena tanah tidak terkontaminasi oleh bahan kimia dan meringankan pekerjaan
dari perlakuan yang intensif.Melalui penempatan umpan pertama tidak beracun
pada koloni rayap dandengan mengulangi penempatan umpan beracun yang slow
action, akan lebih efektif untuk menyebarkan racun terhadap individu lain dalam
sebuah koloni. Agar pengendalian cara ini lebih efektif maka umpan harus lebih
menarik dari makanan di sekitarnya. Penambahan gula, madu, jamur pelapuk,
asam amino, sumber nitrogen, bahkan feromon dapat meningkatkan laju
komsumsi rayap (Pearce 1997). Pengendalian rayap dengan menggunakan
chlorprenapyr pada berbagai konsentrasi melalui pengumpanan mampu
menghasilkan mortalitas yang berkorelasi positif dengan waktu aplikasi dan
konsentrasi perlakuan(Sudohadi Y 2001).
Menurut Scheffrahn dan Su (1994) kelebihan dari metode pengumpanan
yaitu ramah lingkungan, mudah diterima oleh masyarakat karena sedikit
menggunakan bahan kimia yang telah dikemas dalam bentuk yang efektif, dan di
sukai rayap dengan kerja racun yang slow action, sehingga dapat mengeliminasi
koloni rayap. Menurut Su et al. (1994) penggunaan hexaflumuron dengan dosis
0,5% yang dilarutkan pada gulungan kertas tissue (Whatman No. 1) dan
diumpankan pada rayap, maka semua rayap yang mencerna umpan tersebut tidak
metabolisme dan menghambat proses ganti kulit, kemudian setelah beberapa hari
rayap akan mati.
Hingga saat ini pengendalian rayap masih tergantung pada insektisida
sintetik yang berdampak negatif terhadap lingkungan (Kartika et al. 2007),
keracunan bagi penggunadan lainnya. Oleh karena itu perlu alternatif
pengendalian cara lain yang ramah lingkungan. Salah satunya dengan
memanfaatkan bioinsektisida (Soetopo & Indrayani 2009). Ada enam
kelompokmikroorganisme yang bermanfaat sebagai bioinsektisida, yaitu
cendawan, bakteri, nematoda, virus protozoa, dan riketsia (Santoso 1993).
Cendawan Metarhizium brunneum Petch
CendawanM. brunneummerupakan salah satu jenis bioinsektisida yang
dapat menimbulkan penyakit terhadap serangga dandapat ditemukan hampir di
berbagai tempat, seperti tanah gambut di USA dan Oregon (CABI 2001).
Desyanti (2007) mengisolasikan cendawan M. brunneum dari tanah. Untuk
pertumbuhannya memerlukan suhu optimum 22-27°C (Roddam & Rath 1997),
dengan tingkat keasaman (pH) antara 3,3-8,5, sedangkan pH optimal 6,5 (Domsch
& Gams 1980). Konidia akan berkecambah pada kelembaban 90% dan dapat
tumbuh baik pada media PDA, jagung dan beras (Ginting 2008).
Butt et al. (2001) mengklasifikasikan cendawan kepada dua divisi yaitu
Myxomycota dan Eumycota.Lebih rinci Roy et al.(2006) mengklasifikasikan
cendawan entomopatogen Metarhizium sp sebagai berikut:
devisi: Ascomycota
kelas: Sordariomycetes
ordo: Hypocreales
famili: Clavicipitaceae
genus: Metarhizium
spesies:Metarhizium brunneum Petch
CendawanM. anisopliaemempunyai koloni berwarna putih dan dengan
bertambahnya umurwarnanya berubah menjadi gelap, sedangkan strain M.
brunneum pada awal pertumbuhannya bewarna putih hingga kekuningan,
µm, konidiofor tersusun tegak, berlapis, dan bercabang yang dipenuhi oleh
konidia. Konidia bersel satu berwarna hialin, berbentuk bulat silinder dengan
ukuran 9,94 x 3,96 mμ (Strack 2003, Ginting 2008).
Keragaman intraspesiespada cendawan entomopatogen umumnya terlihat
pada perbedaan virulensinya (Hajek& Leger 1994). Faktor yang dapat
mempengaruhi perbedaan keragaman tersebut adalah sumber isolat, inang dan
daerah geografis asal isolat (Varela & Morales 1995, Baretta et al. 1998).
Umumnya strain cendawan entomopatogen yang diisolasi dari inang yang sama
akan lebih virulen untuk diaplikasikan terhadap inang tersebut, dibandingkan dari
inang yang berbeda. Seperti cendawanB. bassiana yang koleksi dari kadaver
rayap apabila kembali diinfeksikan terhadap individu rayap lain dari spesies yang
sama maka toxisitasnya lebih efektif (Boucias& Pendland 1998).
Pemanfaatan agens hayati cendawanM. brunneum di areal pertanaman
belum pernah dilakukan. Penggunaan cendawan ini baru dilakukan Desyanti
(2007) dalam studi pengendalian rayap tanah Coptotermes spp di laboratorium
dan hasilnya efektif sebagai agens hayati dengan tingkat patogenisitasnya lebih
tinggi dan dapat membunuh rayap Coptotermes spp dengan LC50 terendah
dibandingkan spesies M. anisopliae, B. bassiana, Fusarium oxysporum
danAspergillus flavus (Desyanti 2007). Keefektifan dalam penggunaan cendawan
tersebut didukung oleh daya kecambah dan kerapatan konidia yang dihasilkan.
Menurut Desyanti (2007) daya kecambah dari cendawan M. brunneum dan M.
anisopliaesetelah diinkubasiselama 12-24 jammasing-masing 97,20% dan 85
sampai 90%.Kerapatan konidia yang dihasilkan oleh cendawan M. brunneumlebih
tinggi (223,66 x 107/cawan Petri) dibandingkan cendawan M. anisopliae(6,18 x
107/cawan Petri).
Cendawanini mempunyai kemampuan untuk menempel dan menembus
kutikula inang dan tumbuh ke bagian internal tubuh inang (hemocoel) sebagai
sumber nutrisi untuk pertumbuhan yang mengakibatkan kematian inang.
Kemudian cendawan tersebut juga dapat menghancurkan jaringan lain dengan
melepaskan toksin dan akan mempengaruhi perkembangan inang secara normal
Metarhizium spp terhadap serangga yaitu:destruxin, cytohalasins, swainsonine
(Desyanti 2007,Boucias& Pendland 1998).
Diharapkandengan memanfaatkan potensi sumber daya hayati lokal yang
berpotensi di alam Indonesia, seperti pemanfaatan cendawan M. brunneumsebagai
agens hayati yang berpotensi akan dapat mengurangipenggunaan pestisida sintetik
yang selama ini banyak menimbulkan dampak negatif terhadap lingkungan
(Desyanti 2007).
Jarak Pagar (Jatropha curcas L.)
Tahun 1912 tanaman jarak pagar telah diperkenalkan di Indonesia oleh
bangsa Jepang (Hambali et al. 2007). Tanaman ini dapat tumbuh pada tanah kritis
unsur hara, beriklim panas danpada ketinggian tempat 0-1.000 m diatas
permukaan laut (dpl). Namun lebih optimal pertumbuhannya pada tanah yang
berstruktur ringan atau tanah lempung berpasir pH(5,0-6,5), suhu (18-30°C), dan
curah hujan (300-1.200 mm) per tahun (Hariyadi 2005). Pada lahan yang kurang
subur populasitanaman jarak pagar 1.666 per hektar (Prihandana & Hendroko
2006). Sedangkan lahan yang subur daya tampungnya2.500 tanamanper hektar
dengan jarak tanam setiap tanaman 2 m. Tanaman jarak pagar dapat menghasilkan
3,5-4,5 kg biji/pohon/tahun . Bila rendemen minyak dari bijitanaman jarak pagar
sebesar 30% maka setiap hektar lahan dapat diperoleh 1,5-3 ton minyak/ha/tahun.
Produksi akan stabil setelah tanaman berumur lebih dari 5 tahun.Kemungkinan
jarak pagar ini dapat bertahan hidup lebih dari 20 tahun apabilaperawatannya
efektif(Hambali et al. 2007).
Di KIJP Pakuwon budidaya tanaman jarak pagar mengalami beberapa
kendala, salah satu diantaranyaadalah serangan hama rayap M. gilvus yang
merusak sistem perakaran tanaman dan mengakibatkan pengambilan unsur
haradari dalam tanah oleh tanaman menjadi terhambat.Semakin lama serangan
hama ini semakin meningkat hingga meluas pada tanaman lainnya.Data kuantitatif
serangan rayap terhadap tanaman jarak pagar umur tegakan 5 tahun
mencapai16,33%. Oleh karenanya serangan rayap hama ini perlu segera
BAB III
POPULASI KOLONI RAYAP Macrotermes gilvus Hagen
(Isoptera: Termitidae) DI KIJP PAKUWON SUKABUMI JAWA
BARAT
[Colony size population ofMacrotermes gilvus
Hagen(Isoptera:Termitidae) in KIJP Pakuwon Sukabumi West
Java]
Abstrak
Tujuan penelitian ini adalah untuk menduga ukuran populasi koloni rayap M.
gilvus di KIJP Pakuwon dengan menggunakan tekniktriple markrecapture. M. gilvus dikoleksi dari perkebunan jarak pagar (J. curcas) yang ditentukan dalam tiga blok (I, II, III).Koleksi rayap diwarnai denganneutral red0,25% dan nile
blueA 0,05% di laboratorium.Di blok1 dengan luas areal 15.210 m2ditemukan 8
koloni (150.388 individu), di blok 2 dengan luas areal 5.700 m2hanya ditemukan
satu koloni (59.459 individu), dan di blok 3 dengan luas areal 27.000 m2 15
koloni(149.459 individu). Total populasi M. gilvusyang ditemukan terdiri atas 24
koloni dengan 359.066 individu yang tersebar dalam luas areal 47.910 m2.
Kata kunci: Populasi, Macrotermes gilvus, Jatropha curcas, KIJP Pakuwon
Abstract
The aim of this study was to predict the population size of M. gilvus colonyat KIJP Pakuwon using Triple Mark Recapture Technique. M. gilvus was collected from castor plantation whichwas divided in to three blocks (I,II, III). Later M.
gilvus were cleaned, counted and maintained in a Petri dish fed with filter papers
that have been colored by neutral red 0,25% and nile blue A 0,05%. Eight colonies were found on block 1 and the area width is 15.210 m2 (150.388 individuals), only one colony on block 2 with the area width is 5.700 m2 (59.459 individuals), and 15 colonies on block 3 with area width is 27.000 m2 (149.459 individuals). The total population of M. gilvus was 24 colonies counsisted of 359.066 individuals with total area width 47.910 m2.
Pendahuluan
Populasi merupakan sekelompok individu suatu organismedan termasuk
dalam satu spesies yang dapat melangsungkan interaksi genetik dengan spesies
yang sama pada waktu tertentu di suatu wilayah atau ruang tertentu
(Tarumingkeng 1992). Karakteristik suatu populasi yaitu kepadatan (densitas),
laju kelahiran (natalitas), laju kematian (mortalitas), sebaran (distribusi) umur,
potensi biotik, sifat genetik, perilaku dan pemencaran (dispersal) (Tarumingkeng
1992). Ukuran populasi sebuah koloni rayap sangat bervariasi dan dipengaruhi
oleh habitat, mikroklimat (kelembaban, suhu, curah hujan), jenis tanah dan umur
ratu (Khrishna & Weesner 1969).
Ukuran populasi koloni merupakan bagian penting dari sebuah koloni
rayap (Riklefs 1990). Dasar kriteria menggunakan metode pendugaan ukuran
populasi adalah kerapatan dan mobilitas individu penyusun populasi (Krebs
1978). Untuk organisme bergerak, umumnya digunakan teknik tanda tangkap,
metode kuadrat atau transek garis, sedangkan organisme yang tidak bergerak
seperti tumbuhan digunakan metode kuadrat (Tarumingkeng 2001).
Meskipun ukuran populasi rayap tanah sulit dipelajari karena hampir
semua aktivitasnya di bawah permukaan tanah, namun penelitian di bidang
tersebut telah berkembang menggunakan berbagai macan metode baru. Penelitian
tentang pendugaan ukuran populasi koloni rayap C. curvignathus dan M.
inspiratusdapat dilakukan menggunakan metode triple mark reapture
tehnique(teknik tanda tangkap) (Tarumingkeng 1992).
Agar pengendalianrayap dapat berlangsung lebih efektif dan efisien perlu
diketahui ukuran populasi koloninya (Nandika et al. 2003). Rayap hama M. gilvus
pada tanaman jarak pagar perlu dikendalikan sedini mungkin agar kerusakan yang
ditimbulkan tidak mencapai ambang ekonomi.
Bahan dan Metode Tempat dan Waktu
Penelitian ini dilakukan di Laboratorium Taksonomi Serangga,
P P P d 9 t p y k s p G I y B d L
Pertanian I
Pakuwon Su
Persiapan S Loka
dalam tiga b
9 stasiun (bl
terdiri dari 1
pipa PVC Ф
yang ditutup
kemungkina
setiap blok
pemasangan
Gambar 1 P × P d p Identifikasi Seran yang dikole Barat. Cont dimasukkan Laboratorium a Institut Pert ukabumi Jaw Stasiun Peng asi percobaa
blok (I,II, da
lok II), dan
10 potong ka
Ф 5 inci. Kem
p dengan bah
an gangguan
k. Berikut
nnya di KIJP
Perakitan Sta ×10 cm) dib PVC berisi dari bahan permukaan s
i Rayap ngga yang d
ksi dari Keb
toh beberap
ke dalam
m Taksono
tanian Bogo
wa Barat, seja
gamatan da an adalah l
n III) yang m
45 stasiun (
ayu pinus (1
mudian pipa
han plastik b
n dari luar.
ini
bagian-P bagian-Pakuwon (G
asiun Penga bungkus den
kayu pinus
plastik (wa
stasiun peng
digunakan d
bun Induk J
pa individu
m botol ber
omi Serang
or, dan di K
ak Nopembe
an Pemasan lahan KIJP
masing- mas
(blok III). S
cm × 2 cm×
a PVC dimas
berwarna ge
Jarakantara
[image:35.612.132.500.340.483.2]bagian dari
Gambar 1 da
amatan: (a) p ngan kertas yang terbun ater straine amatan. dalam penel Jarak pagar
u rayap (ka
risi alkohol
gga Depart b
Kebun Indu
er 2009 samp
ngannya Pakuwon.
sing terdiri d
Stasiun peng
×10cm) yang
sukkan dalam
elap, agar tid
astasiun pen
i stasiun p
an 2).
potongan kay kardus bek ngkus kardu
r) sebagai
itian ini ada
(KIJP) Pak
asta pekerja
l 70%. Ide
temen Prot
uk Jarak Pag
pai Juli 2010
Percobaan
dari 23 stasiu
gamatan yan
g dimasukka
m lubang ga
dak tembus c
ngamatan 20
pengamatan
yu pinus (1 kas, (b) poto us bekas, dan penutup pa
alah spesies
kuwon Sukab
a dan kast
entifikasi d
teksi Tanam
gar (KIJP)
0.
ditentukan
un (blok I),
g dipasang
n ke dalam
alian 15 cm
cahaya dan
0 m dalam
dan cara
cm × 2 cm ongan pipa n (c) tutup ada bagian
s M. gilvus
bumi Jawa
a prajurit)
dilakukandi
Karak pengen Gamb techni beriku Tahap dari ta (What
dan ni
rayap Rayap dilepa pelepa dikum berwa Tahap kteristik morf nalan rayap
ar 2 (a) sta
Percobaan que(Marini ut: p Pertama Kayu ump anahserta di
tman No 1)
ile blue A 0
selama 3 ha
p yang telah
skan ke stas
asan rayap b
mpulkan. Ra rnadihitung p Kedua fologi spesie (Tarumingk siun pengam P ini me
& Roberto 1
pan yang ter
ihitung jum yang telah 0,05% (Hara ari, sehingga h berwarna siun pengam bertanda.Kay ayap yang kembali.
es rayap ini
keng 1971).
matan, dan (b
Pelaksanaan enggunakan 1998). Adap rserang rayap mlahnya. Pew direndam da
hapet al. 20
a diperoleh r
tersebut dih
matan tempa
yu umpan d
tertangkap a diketahui be b) pemasang n Penelitian metode
pun tahapan k
p dikumpulk warnaannya alam bahan 005). Kertas rayap warna hitung kemb
at rayap dita
dari setiap s
p baik yan
erdasarkan k
gannya di KI
triple m
kerja dari m
kandan dipi
menggunak
pewarna ne
tissue dium
biru dan me
bali jumlahn angkap. Satu stasiun peng ngberwarna kunci identif JP Pakuwon mark recap
metode ini seb
sahkan raya
kan kertas t
eutral red 0
mpankan terh
erah (Gamba
nya dan kem
u minggu se
p s t T s r N S d N S n m M G Raya prosedur pad semula ditan tahap pertam Tahap Keti Pena seperti pros
rayap M. gil
N = (∑Mi.n
SE = N/{[1/
dimana:
N = Uk
SE = Sim
ni = Jum
mi = Jum
Mi = Jum
Gambar 3 R d a
ap yang ter
da tahap per
ngkap. Sem
ma dengan ta
iga
andaan, pele
edur tahap
lvusdigunaka
ni)/[(∑mi)+1
(∑mi)+1)]+
kuran popula
mpangan Ba
mlah keselur
mlah rayap b
mlah total ra
Rayap M. gi
dan (b) nile
rtangkap pad
rtamadan k
minggu setela
ahap berikutn
epasan dan p
pertama da
an metode B
]
{(2/((∑mi)+
asi.
aku.
ruhan rayap
bertanda yan
ayap bertand
ilvus yang te
e blue A 0,05
da tahap pe
kembali dilep
ah pelepasan
nya (kedua d
penangkapan
an kedua. Pe
Begon (Marin
+1)2+[(6/(∑m
yang tertang
ng tertangkap
da sampai pe
elah diwarnai 5%.
ertama, diw
pas ke stasiu
n diamati ke
dan ketiga) s
n rayap untu
endugaan uk
ni & Roberto
mi)+1)3]}1/2
gkap pada pe
p pada penan
enangkapan k
i dengan (a) b
warnai kemb
un pengama
embali. Inte
selama 10 ha
uk tahap tig
kuran popu
o 1998) yaitu
enangkapan
ngkapan
ke-ke-i
) neutral red
bali seperti atan tempat rval waktu ari. ga diulangi lasi koloni u: ke-i. -i.
1. Id dan III Tabel Blo I II II mayor Gamb 2. U
M..gilv
ukuran sebesa terdiri dan 86 a dentifikasi R Semua con
I di KIJP Pa
1 Spesies ra
ok Juml
pen I
I II
Bahan ide
r (Gambar 4)
ar 4 Rayap (c) kas
Ukuran Popu Hasil peng
vus di KIJP
n populasi k
ar 25% (A) d
dari satu ko
6,87% kolon Rayap ntoh rayap y
akuwon adal
ayap dari sta
lah stasiun ngamatan 23 9 45 entifikasi ada ).
p Macroterm
sta prajurit m
ulasi Koloni gamatan me
Pakuwon d
koloni kecil.
dan 75% kol
oloni besar (A
ni kecil(D,E,F
Hasil dan
yang dijumpa
lah Macroter
asiun pengam
alah rayap k
mes gilvus (a mayor
i Rayap Ma enunjukkan
igolongkan
Persentase u
loni kecil (B
A) (Gambar
F,G,H,I,J,K, b
n Pembaha
ai di stasiun
rmes gilvus
matan di KIJ
Spes
Macrote Macrote Macrote
kasta pekerj
a) kasta peke
acrotermes g
bahwa uku
kepada ukur
ukuran popu
B,D,E,F,G,H
r 6). Blok III
,L,M,N dan san n pengamatan Hagen (Tab JP Pakuwon. sies rayap ermes gilvus ermes gilvus ermes gilvus
ja, kasta pra
erja (b) kast
gilvus Hagen
uran popula
ranpopulasi
ulasi koloni b
H) (Gambar 5
I 13,13% ko
O) (Gambar c
n pada blok
el 1). . s s s ajurit minor
ta prajurit m
n
asi koloni r
koloni besar
besar pada b
5). Blok II h
loni besar (A
[image:38.612.91.476.52.780.2]Tabel 2Ukuran populasi koloni rayap Macrotermes gilvus Hagen di KIJP Pakuwon.
No Blok Jumlah Koloni Ukuran Populasi (N) Luas areal (m2)
I 8 150.388 15.210
II 1 59.219 5.700
III 15 149.459 27.000
Jumlah 24 359.066 47.910
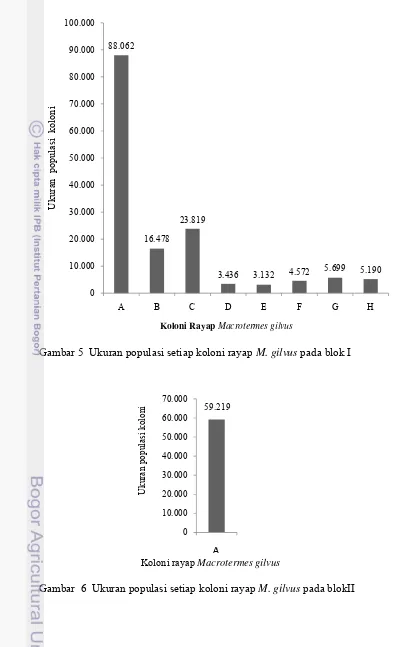
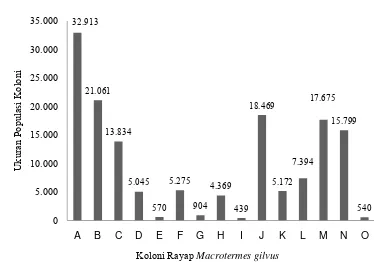
Ukuran populasi koloni tiap blok berbeda. Persentase koloni pada blok I,
II, dan III termasuk koloni ukuran kecil 73,17%, sedangkan 20,83% termasuk
koloni ukuran besar (Tabel 2).Karakteristik koloni rayap M. gilvus di KIJP
Pakuwon banyak didominasioleh kasta pekerja dan hanya dalam jumlah kecil
terdapat kasta prajurit. Hal inimenunjukkan bahwa kasta pekerja berperan penting
terhadappembangunan sebuah koloni terutamakoloni yang baru terbentuk.
Di KIJP Pakuwon jugaterdapat koloni rayap M. gilvusyang mempunyai
kasta prajurit hampir seperempat dari jumlah kasta pekerja. Hal inididuga karena
banyaksemut di sekitar stasiun pengamatan sebagai musuh utama rayap.Ratu
dalam koloni tersebutmenghasilkan lebih banyak kasta prajurit untuk melindungi
anggota koloninya dari serangan semut. Pada koloni yang tidak terdapat sarang
semut jumlah kasta prajurit minor sangat sedikitbahkantidak terdapat kasta
Gambar 5 Ukuran populasi setiap koloni rayap M. gilvus pada blok I
Gambar 6 Ukuran populasi setiap koloni rayap M. gilvus pada blokII
88.062
16.478
23.819
3.436 3.132 4.572 5.699 5.190
0 10.000 20.000 30.000 40.000 50.000 60.000 70.000 80.000 90.000 100.000
A B C D E F G H
Uk
uran po
p
ulasi
ko
lon
i
Koloni Rayap Macrotermes gilvus
59.219
0 10.000 20.000 30.000 40.000 50.000 60.000 70.000
1
Ukura
n popul
asi koloni
Koloni rayap Macrotermes gilvus
Gambar 7 Ukuran populasi setiap koloni rayap M. gilvus pada blokIII
Ukuran populasi koloni rayap diduga tergantung pada umur ratu (umur
koloni), semakin tua umur ratu maka kapasitas untuk meletakkan telur akan lebih
tinggi demikian juga sebaliknya semakin muda umur ratu, kemampuan untuk
meletakkan telursemakin rendah. Menurut Faulet et al. (2006)ukuran
populasisebuah kolonirayap dengan jumlah individu kurang dari 20.223 individu
(50% kasta pekerja dan 10% kasta prajurit) termasuk dalam koloni ukuran kecil,
sedangkan lebih besar dari ukuran tersebut dikelompokkan ke dalam koloni
ukuran besar. Ukuran populasi koloni dipengaruhi oleh umur ratu dan lingkungan
sertaadanya gangguan organisme lain seperti semut (Khrisna &Weesner 1969).
Lee at al. (2007) menambahkan bahwaapabila lingkungan tidak mendukung maka
ukuran populasi koloni rayap tidak akan meningkat bahkan semakin
rendah.Berbagai faktor lingkungan seperti tanah, tipe vegetasi, iklim, dan
ketersediaan air, sangat mempengaruhi ukuran populasi koloni rayap.
Lingkungan KIJP Pakuwon dengan ciri tingkat kelembaban relatif 76
sampai 88%, suhu permukaan22-26°C, sirkulasi udararelatif lebih baik,
32.913
21.061
13.834
5.045
570 5.275
904 4.369
439 18.469
5.172 7.394
17.675
15.799
540
0 5.000 10.000 15.000 20.000 25.000 30.000 35.000
A B C D E F G H I J K L M N O
Uk
ur
an Pop
ula
si Ko
lon
i
kandungan bahan organik banyak sangat menunjang perkembangan populasi
koloni rayap
Simpulan
1. Jenis rayap yang dijumpai di KIJP Pakuwon adalah Macrotermes gilvus.
2. Ukuran populasi koloni rayap M. gilvusdi KIJP Pakuwon sebesar 359.066
individu dari 24 koloni.
3. Umumnya lebih banyak terdapat koloni ukuran kecil dibandingkan koloni
ukuran besar.
Daftar Pustaka
Faulet BM, Niamke S, Gonnety JT, Kouame LP. 2006. Purification and biochemical properties of a new thermostable xalanase from symbiotic
fungusTermitomyces sp. African Journal of Biotechnology 5(3): 273-282.
Harahap IS, Benson EP, Zungoli PA, Adler PH, Hill HS. 2005. Inter-and
intra-colony agonistic behavior of nativesubterranean termites, Reticulitermes
flavipes and Reticulitermes virginicus (Isoptera: Rhinotermitidae). Sociobiology 46: 305-316.
Khrishna K, Weesner FM. 1969. Biology of Termites.Volume II. New York: Academic Press.
Krebs J. 1978. Ecology the Experimental Analysis of Distribution and Abundance.Second Edition.Harper and Row Publisher. New York.
Lee CY, Vongkaluang , Lenz M. 2007. Challenges to subterranean termite tunnel
branches for efficient food search and resource transportation. BioSystems
90:802-807.
Marini M, Roberto F 1998. A Population survey of the Italian subterranean
termite Reticulitermes lucifuguslucifugus Rossi in Bacnacavallo (Rovenna,
Italy) using the Triple Mark Recupture Techninque (TMR). Zoological
sciencie 15:963-969
Nandika D, Rismayadi Y, Diba F. 2003. Rayap, biologi dan pengendaliannya Surakarta.Muhammadiyah University Press.
Tarumingkeng R. 1971. Biologi dan Pengenalan Rayap Perusak Kayu di Indonesia. Laporan Lembaga Penelitian Hasil Hutan. No. 138. Bogor.
Tarumingkeng R. 1992. Insektisida: sifat, mekanisme kerja dan dampak penggunaannya. Ukrida Press. Jakarta.
Tarumingkeng. R. 2001.Biologi dan Perilaku Rayap.http://tumoutou.net/
BAB IV
DAYA JELAJAH RAYAP Macrotermes gilvus Hagen (Isoptera:
Termitidae) DI KIJP PAKUWON SUKABUMI JAWA BARAT
[The foraging range of Macrotermes gilvus Hagen (Isoptera:
Termitidae) in KIJP Pakuwon Sukabumi West Java]
Abstrak
Daya jelajah maksimum rayap Macrotermes gilvus Hagen dipengaruhi oleh faktor
lingkungan terutama sumber makanan yang tersedia pada habitatnya. Penelitian
ini bertujuan untuk mempelajari daya jelajah maksimum rayap M. gilvus di KIJP
Pakuwon. RayapM. gilvusditangkapdari stasiun pengamatan, dipisahkan dari
tanah dan diwarnai denganneutral reddan nile blue.Rayap berwarna kemudian
kembali dilepaskan pada stasiun pengamatan dan seminggu kemudian diamati.
Rata-rata daya jelajah maksimum rayap M. gilvussejauh 140,5 m
Kata kunci : Macrotermes gilvus, daya jelajah,KIJP Pakuwon
Abstract
The maximum foraging range of M. gilvusinfluenced by environmental factors especially the availability of food resourcesin their habitat. The aim of this reseacrh was to study the maximum foraging range of M.gilvus in KIJP Pakuwon. The reseacrh was carried out by capturing M. gilvus from observation stations separated from the soil and marked with neutral red and nile blue. Marked termites then released back to the station and observed a week later. Average maximum foraging range of M. gilvus extent to140,5 m.
Pendahuluan
Wilayah jelajah didefinisikan sebagai daerah yang dikunjungi oleh
organisme secara tetap kerena tersedia sumber makanan dan dapat berfungsi
sebagai tempat berlindung, beristirahat, dan bereproduksi (Moen 1973, Alikodra
1990).Beberapa spesies rayap memiliki daya jelajah lebih luas untuk mendapatkan
sumber makanan. Perilaku persentuhan fisik dan bau yang dikeluarkan melalui
jejak dari individu rayap untuk disampaikan kepada anggota koloninya merupakan
suatu mekanisme penyampaian informasi terhadap sumber makanan yang baru
ditemukan (Pearce 1997).
Mempelajari daya jelajah rayap tanah mengalami kesulitan karena hampir
semua aktivitasnya di bawah permukaan tanah (Su et al. 1994).Pendugaan daya
jelajah rayap dapat dilakukan dengan menggali galeri rayap(King & Spink 1969)
atau menggunakan penanda radioisotop (Spragg & Paton 1980). Sornnuwat et al.
(1996) menyatakan bahwakedua cara tersebut kurang efektif digunakan untuk
mempelajari daya jelajah rayap. Metode tanda-lepas-tangkap (
mark-release-recapture method) telah banyak digunakan untuk melakukan penelitian terhadap
daya jelajah rayap (Su et al. 1994, Su et al. 1991, Sornnuwat et al. 1996).
Beberapa bahan penanda sangat efektif digunakan untuk penelitian daya jelajah
rayapC. formosanus seperti neutral red dan nile blue A (Su et al. 1991).
Bahan dan Metode Waktu dan Tempat
Penelitian ini dilaksanakan sejak Nopember 2009 sampai Juli 2010 di
Kebun Induk Jarak Pagar (KIJP) Pakuwon, Sukabumi, Jawa Barat.
Perakitan Stasiun Pengamatan dan Pemasangannya
Cara kerja sama seperti yang telah diuraikan pada BAB III
Pelaksanaan Penelitian
Seluruh stasiun pengamatan yang terdapat pada blok II diberi nomor 1
dua stasiun pengamatan sebagai tempat pelepasan awal. Rayap yang terdapat pada
stasiun pengamatan terpilih (3 dan 9) dikoleksisecara terpisah dan diwarnai
melalui pengumpanan menggunakan kertas saring yang telah direndam ke dalam
neutral red (0,25 %) dan nile blue A (0,05%) dalam Petridishsecara terpisah
(Harahap et al. 2005) selama tiga hari (Tabel 3). Setelah berwarna merah dan biru,
rayap dilepas kembali di stasiun pengamatan tempat semula. Pengamatan terhadap
daya jelajah maksimum rayap M. gilvusdilakukan mulai dari blok II(stasiun
pengamatan tempat pelepasan rayap bertanda) hingga pada blok I dan blok III
(stasiun pengamatan tempat ditemukan kembali rayap bertanda) menggunakan
alat ukur meteran pita 50 m (Gambar 12).
Tabel 3 Stasiun pengamatan terpilih untuk menduga daya jelajah rayap M. gilvus.
Nomor stasiun Rayap tertangkap Tanda Lepas
Merah Biru Merah Biru
3 556 556 482
9 495 495 411
Hasil dan Pembahasan
Rayap M. gilvus melakukan daya jelajah untuk mendapatkan sumber
makanan dan sangat ditentukan oleh lingkungan yang optimal. Rata-rata jarak
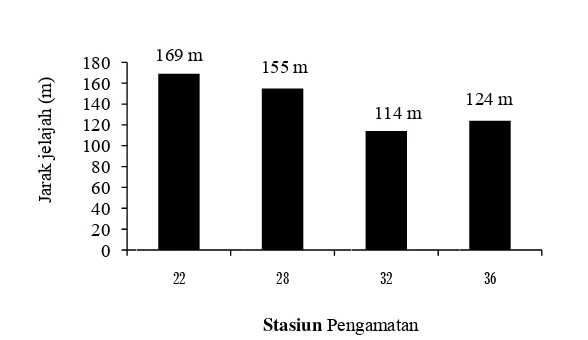
linier daya jelajah rayap M. gilvus pada blok I sejauh 140,5 m (324,25 individu)
dari luas areal 15.210 m2 (Gambar 8), dan blok III sejauh 140 m (311,26 individu)
dari luas areal 27.000 m2 (Gambar 9). Cara pengukuran daya jelajah rayap M.
gilvus pada (Gambar 13). Sedangkan hasil pengamatan daya jelajah setiap koloni
dari blok (I dan III) dapat di lihat pada Gambar (4 dan 5).
Rayap M. gilvusmelakukan daya jelajah lebih jauh pada blok I diduga
pengaruh dari beberapa faktor lingkungan terutama makanan yang tersedia sangat
terbatas (Gambar 10). Di blok Ipertumbuhan tanaman jarak pagar kerdil dengan
kerapatan tajuk tananam lebih rendah dan luas kanopinya lebih sempit. Tidak
terdapat serasah dari sisa tanaman seperti daun dan cabang yang patah pada
habitat tersebut. Sanitasi gulma dilakukan secara berkelanjutan di sekitar pangkal