PENINGKATAN PERAKARAN BIBIT MANGGIS
(Garcinia mangostana L.) MELALUI INOKULASI
Agrobacterium rhizogenes
L I Z A W A T I
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
BOGOR
PENINGKATAN PERAKARAN BIBIT MANGGIS
(Garcinia mangostana L.) MELALUI INOKULASI
Agrobacterium rhizogenes
OLEH :
L I Z A W A T I
A. 361020071
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
SURAT PERNYATAAN
Dengan ini saya menyatakan dengan sebenar-benarnya bahwa segala
pernyataan dalam disertasi saya yang berjudul :
”PENINGKATAN
PERAKARAN
BIBIT
MANGGIS
(
Garcinia
mangostana
L.) MELALUI INOKULASI
Agrobacterium
rhizogenes
”
merupakan gagasan atau hasil penelitian disertasi saya sendiri dengan
bimbingan Komisi Pembimbing, kecuali yang dengan jelas ditunjukkan
rujukannya. Disertasi ini belum pernah diajukan untuk memperoleh gelar sejenis
di Perguruan Tinggi lain.
Semua data dan informasi yang digunakan telah dinyatakan dengan
jelas dan dapat diperiksa kebenarannya.
Bogor, 2007
L I Z A W A T I
ABSTRACT
LIZAWATI. Improvement of mangosteen (Garcinia mangostana L.) seedling root system through Agrobacteriun rhizogeneses inoculation. Supervised by ROEDHY POERWANTO, IMAN RUSMANA, SOBIR, TRI MUJI ERMAYANTI.
Mangosteen is known as a slow growing plant, this is due to the root that is fragile, sensitive to the environmental condition and easily disturbed. Great care is therefore required during the transplanting of seedling, which form a long taproot with few laterals. Many writers note an apparent absence of root hairs at all stage of growth. The use of Agrobacterium rhizogenes may improve root system of mangosteen. The soil bacterium Agrobacterium rhizogenes can induce the abundant adventitious root formation at the infection site through the transfer of genetic material T-DNA, a part of the Root inducing (Ri) plasmid from bacterium to the plant genome. This research is aimed to develop a technique in improving root system of Mangosteen using Agrobacterium transformation technique in order to improve seedling growth. This research is divide into two step of research, I) improvement of mangosteen seedling root system through A. rhizogeneses inoculation at the mangosteen nursery ; and II) induction of root formation using A. rhizogenes in vitro. The materials used in this experiment were ; mangosteen fruit originated from Purwakarta and A. rhizogenes collection from Puslit Biotechnology LIPI Cibinong-Bogor. The result showed that strains inoculation of ATCC-15834, 509 , 07-20001, A4, and R-1000 increased : stem diameter, plant height, leaf number, lateral and tersier roots number better than control. Inoculation with cutting root method results in the higher live plant percentage compared with dipping root method. A. rhizogenes strain ATCC-15834 (OD600 = 1.0) was able to infect 6 week old of mangosteen seedling root.
Based on PCR product, TL-DNA from A. rhizogenes strain ATCC-15834
succeeded to be transferred into genome of mangosteen seedling root cell since the rolB gene was detected at 780 bp agarose electrophoresis. The A. rhizogenes strains ATCC-15834 could increase the root anatomy in terms of the mean xylem vascular diameter, conductivity, total xylem width, roots hair, nutrient uptake (N, P and K), IAA hormone content and was better than and control. 509, 07-20001, ATCC-158343 strains induced root formation. All explant with cotyledon that were root formation were induced were able to survive at acclimation stage, but none for cotyledoneless explants. Anatomy observation showed that 509 resulted in higher xylem diameter, total number of xylem, conductivity, and higher ration of conductivity/total root transversal area in comparation to control.
RINGKASAN
LIZAWATI. Peningkatan Perakaran Bibit Manggis (Garcinia mangostana L.)
Melalui Inokulasi Agrobacterium rhizogenes. Dibimbing oleh ROEDHY
POERWANTO, IMAN RUSMANA, SOBIR, TRI MUJI ERMAYANTI.
Pertumbuhan tanaman manggis yang lambat, disebabkan antara lain akar tumbuh dengan lambat, rapuh, jumlah akar lateral terbatas dan tidak mempunyai rambut akar, mudah rusak dan terganggu akibat lingkungan yang tidak menguntungkan. Salah satu upaya yang dapat dilakukan untuk meningkatkan sistem perakaran manggis adalah dengan pemanfaatan bakteri
Agrobacterium rhizogenes. Bakteri A. rhizogenes merupakan bakteri tanah yang mempunyai kemampuan untuk menstranfer sebagian bahan genetiknya (T-DNA) pada sel tanaman melalui pelukaan. T-DNA tersebut membawa gen-gen yang terlibat dalam proses induksi akar. Secara umum penelitian ini bertujuan untuk mendapatkan teknologi peningkatan sistem perakaran tanaman manggis melalui inokulasi A. rhizogenes sehingga mempercepat laju pertumbuhan bibit tanaman manggis. Penelitian terdiri dari dua percobaan, yaitu; I) Perbaikan sistem perakaran tanaman manggis melalui inokulasi A. rhizogenes terhadap semai
manggis, dan II) Induksi perakaran eksplan tunas manggis dengan A.
rhizogenes secara in vitro. Hasil penelitian menunjukkan bahwa inokulasi A. rhizogenes strain ATCC-15834, 07-20001, A4, dan 509 mampu meningkatkan pertumbuhan panjang akar primer, jumlah akar sekunder dan tersier serta pertumbuhan tajuk, yaitu : diameter batang, tinggi tanaman, dan jumlah daun tanaman bibit manggis. Metode inokulasi dengan cara akar dipotong lebih efektif dalam menginokulasi akar bibit manggis dibandingkan dengan akar ditusuk. Bakteri A. rhizogenes strain ATCC-15834 konsentrasi OD600=1.0 dapat
menginokulasi akar bibit manggis umur 6 minggu. Anatomi akar bibit manggis hasil inokulasi strain ATCC-15834 menghasilkan rata-rata diameter pembuluh xilem, luas serapan permukaan sayatan melintang dan total luasan xilem akar yang lebih besar serta menghasilkan pertumbuhan rambut akar yang lebih baik dibandingkan bibit yang tidak diinokulasi. Serta menghasilkan serapan hara (N dan P) daun, dan kandungan total hormon IAA yang paling tinggi. Strain ATCC-15834 dapat mentransfer daerah TL-DNAnya pada akar bibit manggis yang dibuktikan dengan terdeteksinya gen rolB 780 bp pada gel elektroforesis. Induksi perakaran eksplan tunasmanggis secara in vitro dengan A. rhizogenes mampu menginduksi terbentuknya akar adventif pada tempat infeksi setelah diinokulasi dengan strain 509, 07-20001, dan ATCC-15834 dan mampu tumbuh hingga 75 % pada tahap aklimatisasi. Hasil anatomi akar menunjukkan bahwa inokulasi
A. rhizogenes strain 509 menghasilkan rata-rata diameter pembuluh xilem, jumlah total pembuluh xilem, luas serapan permukaan sayatan melintang, total luasan sayatan melintang dan rasio (luas serapan / total luasan sayatan melintang) yang lebih besar dibandingkan bibit manggis yang tidak diinokulasi (kontrol).
© Hak cipta milik Institut Pertanian Bogor,
Tahun 2007
Hak cipta dilindungi undang-undang
1. Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa
mencantumkan atau menyebutkan sumber
a. Pengutipan hanya untuk kepentingan pendidikan,
penelitian, penulisan karya ilmiah, penyusunan laporan,
penulisan kritik atau tinjauan suatu masalah.
b. Pengutipan tidak merugikan kepentingan yang wajar
IPB
PENINGKATAN PERAKARAN BIBIT MANGGIS
(Garcinia mangostana L.) MELALUI INOKULASI
Agrobacterium rhizogenes
L I Z A W A T I
DISERTASI
Sebagai salah satu syarat untuk memperoleh gelar
Doktor pada
Program Studi Agronomi
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Judul Disertasi : Peningkatan Perakaran Bibit Manggis (Garcinia mangostana L.) melalui Inokulasi Agrobacterium rhizogenes .
Nama Mahasiswa : Lizawati
NO. MAHASISWA : A 361020071
Disetujui
Komisi Pembimbing
Prof. Dr. Ir. H. Roedhy Poerwanto, M.Sc Dr. Ir. S o b i r, MS Ketua Anggota
Dr.Ir. Iman Rusmana, MS Dr. Tri Muji Ermayanti Anggota Anggota
Diketahui
Ketua Program Studi Agronomi Dekan Sekolah Pascasarjana IPB
Dr. Ir. Satriyas Ilyas, MS Prof. Dr. Ir. Khairil Anwar Notodiputro, MS
RIWAYAT HIDUP
Penulis dilahirkan di Kabupaten Kerinci Provinsi Jambi pada tanggal 5
Desember 1970, adalah anak keempat dari pasangan Drs. H. M. Rusli dan Hj
Azizah. Penulis menyelesaikan pendidikan dasar di SD Negeri 52/IV Jambi
pada tahun 1984, SMP Negeri 8 Jambi pada tahun 1987, dan SMA Negeri I
Jambi pada tahun 1990. Penulis melanjutkan pendidikan di Universitas Jambi
Fakultas Pertanian dan lulus pada tahun 1994. Tahun 1995 penulis diangkat
menjadi staf pengajar Fakultas Pertanian Universitas Jambi. Penulis
melanjutkan pendidikan Master di Institut Pertanian Bogor pada Program Studi
Bioteknologi dan lulus tahun 2002. Pada tahun yang sama penulis mendapat
kesempatan untuk melanjutkan ke jenjang doktor pada Program Studi Agronomi
Institut Pertanian Bogor.
Selama mengikuti program S3, penulis menyajikan karya ilmiah
berjudul Metode Inokulasi Agrobacterium rhizogenes pada Bibit Tanaman Manggis (Garcinia mangostana L.) pada Seminar Nasional Perhimpunan Hortikultura Indonesia pada bulan November 2006. Sebuah artikel akan
diterbitkan pada Jurnal Buletin Agronomi Vol. XXXV, No. 2, dengan judul
Pertumbuhan Bibit Tanaman Manggis (Garcinia mangostana L.) setelah Inokulasi dengan Berbagai Strain Agrobacterium rhizogenes. Karya ilmiah tersebut merupakan bagian dari program S3 penulis.
PRAKATA
Puji syukur Alhamdulillah penulis panjatkan ke hadirat Alloh SWT,
karena atas rahmat dan hidayat-Nya penelitian dan penulisan Disertasi ini
berhasil diselesaikan. Penelitian ini dibiayai oleh Program Riset Unggulan
Strategis Nasional (RUSNAS) melalui Pusat Kajian Buah-Buahan Tropika,
LPPM-IPB.
Pada kesempatan ini penulis mengucapkan rasa terimakasih yang
tulus kepada :
1. Prof.Dr. Ir. Roedhy Poerwanto, MSc, Dr. Ir. Sobir, MS; Dr. Ir. Iman Rusmana,
MSi, Dr. Ir. Tri Muji Ermayanti, selaku komisi pembimbing yang telah
memberikan kepercayaan dan bimbingan selama penelitian sampai
penyusunan Disertasi.
2. Dr. Ir. Satriyas Ilyas, MS selaku Ketua Program Studi Agronomi dan seluruh
dosen Program Studi Agronomi yang selalu memberikan dukungan.
3. Dr. Ir. Nurul Khumaida, MSc selaku penguji luar komisi pada saat ujian
tertutup yang telah banyak memberikan saran dan Dr. Ir. Darda Effendi, MS
selaku penguji luar komisi pada saat ujian prakualifikasi bersama Dr. Ir.
Maya Melati, MSc sebagai wakil Program Studi Agronomi.
4. Dr. Ir. Agus Purwito, MSc dan Dr. Ir. Ika Mariska APU selaku penguji luar
komisi pada saat ujian sidang terbuka.
5. Rektor Universitas Jambi yang telah memberikan kesempatan untuk
mengikuti program S3 di IPB.
6. Direktorat Jenderal Pendidikan Tinggi, yang telah memberikan beasiswa
BPPS.
7. Staf dosen, peneliti dan karyawan di Pusat Kajian Buah-buahan Tropika
LPPM-IPB Prof. Dr. Ir. Hj. Syafrida Manuwoto, MSc., Ir. Hj. Yayah K.
Wagiono, MEc., Dr. Ir. Sriani Sutjiprihati, MS., Dr. Ir. Rahmad Suhartanto MS.
Dr. Ir. M. Firdaus, MSi., Ir. Ivone O. Sumaraw, MS., Endang Gunawan SP.
MSi., Kusuma Darma SP. MSi., Heri Harti SP., Rena Destriani Amd., Rika
Lesmawati Amd., Naekman Naiboho SP dan M. Syafrudin atas segala
bantuan dan dukungan yang telah diberikan.
8. Bapak Sulaeman, pak Sukardi, dan bu Ade serta seluruh karyawan Kebun
9. Sulassih SP dan Sapitri atas bantuan dan kerjasamanya selama penelitian
berlangsung di Laboratorium PKBT-IPB serta Erwin dan Deritha di Puslit
Bioteknologi LIPI-Cibinong.
10. Dr. Ir. Juliarni M.Agr, Dr. Ir. Ragapadmi Purnamaningsih, M.Si, Dr. Ir. Ireng
Darwati, Ir. Dorly MSi, Ir. Muhammad Arif Nasution, MP, Ir. La Ode Safuan,
MP, Ir. Dirvamena Boer, M.Sc, Ir. Liferdi, M.Si dan Dewi Sukma, SP. M.Si
atas kebersamaan dan diskusinya selama penelitian berlansung.
11. Dr. Ir. Nurita Toruan-Matius MS dan Dr. Ir. Hajrial Aswidinnoor M.Sc, yang
telah memberikan dorongan agar penulis melanjutkan program S3.
12. Ayahanda H. M. Rusli, ibunda Hj. Azizah, ayah dan ibunda mertua H. Dja’far
Madjid dan Hj. Zubaidah serta seluruh keluarga atas doa dan dukungan
yang telah diberikan kepada penulis.
13. Suami tercinta Ir. Zainuddin, M.Si dan ananda tersayang Puja Ahmad Habibi
atas pengorbanan, ketulusan, kesabaran dan pengertian yang telah
diberikan selama ini.
14. Semua pihak yang telah membantu dan memberikan dukungan selama
pendidikan S3.
Akhirnya, diiringi doa semoga seluruh kegiatan studi ini bernilai ibadah
dihadapan Alloh SWT, baik bagi penulis maupun semua pihak yang terlibat di
dalamnya, semoga hasil-hasil penelitian ini dapat didayagunakan lebih lanjut
bagi kemaslahatan masyarakat maupun bagi kemajuan ilmu pengetahuan.
Bogor, 2007
GLOSARIUM
Adventif : Perkembangan organ seperti tunas, akar bunga; atau
emberio yang berasal dari suatu titik tumbuh yang tidak lazim.
Agrobacterium
rhizogenes : Bakteri tanah yang bersifat aerobik, beraksi negatif terhadap pewarnaan gram dan dapat menyebabkan terbentuknya akar rambut pada tanaman dikotil dan monokotil.
Aklimatisasi : Masa adaptasi planlet dari lingkungan fisik aseptik terkendali ke lingkungan tanah.
Apomiksis : Reproduksi melalui bentuk seperti biji tetapi tanpa melalui penyerbukan.
Auksin (IAA) : Zat pengatur tumbuh tanaman yang mendorong pertumbuhan tanaman. Auksin alami yang dikenal ialah indolaseticacid (IAA) terutama mempengaruhi perbesaran sel dan pertumbuhan pucuk apikal tanaman, inisiasi akar dan perkembangan akar samping, penghambatan masa tunas samping, menunda penuaan daun dan absisi. Aktifitas IAA dapat rusak karena intensitas cahaya yang tinggi.
Bakteri : Suatu mikroorganisme prokariotik dalam domain bacteria
Eksplan : Bagian tanaman yang digunakan sebagai bahan untuk inisiasi suatu kultur misalnya kultur in vitro.
Elektroforesis : Pemisahaan molekul berdasarkan muatan listriknya.
Floem : - Kulit kayu bagian dalam pada batang yang berguna untuk mendistribusikan protein dan karbohidrat, merupakan serangkaian sel yang membentuk pembuluh ayak, mempunyai sel dasar berupa sel tapis yang berdinding sel tipis.
- Sekumpulan sel jaringan pembuluh yang berdinding tebal pada akar dan berada diantara jaringan xilem dan kulit luar akar.
Gen : Unit dasar pembawa sifat keturunan yang merupakan sekuen nukleotida-nukleotida DNA penyandi produk fungsional dari RNA.
Hara : Bahan kimia anorganik (dari dalam tanah) yang diserap oleh tanaman untuk dibentuk menjadi senyawa-senyawa organik yang kompleks yang dimanfaatkan sebagai pembentuk sel, jaringan dan organ tanaman.
Infeksi : Kemasukan bibit penyakit.
Inokulasi :Pemasukan bakteri kedalam tubuh melalui luka atau melalui alat yang digoreskan pada kulit dan tidak selalu menimbulkan infeksi.
In vitro :Di dalam tabung atau di dalam botol kultur.
In vivo : Di dalam tanaman utuh yang tumbuh dirumah kaca atau dilapang.
Jumlah kopi
(turunan) : Jumlah molekul suatu plasmid yang terdapat dalam satu sel
Juvenil : Suatu periode dalam tanaman ketika pembungaan tidak terjadi dan tanaman tidak dapat dirangsang untuk berbunga dengan ZPT atau perangsang pembungaan lainnya.
Kromosom : Molekul asam nukleat yang dapat mengadakan replikasi sendiri serta membawa sejumlah gena.
Konjugasi : Pada bakteri merupakan transper DNA diantara dua sel yang bersambungan secara temporer.
Konduktivitas
xilem : Daya hantar pembulah xilem untuk mengirimkan atau mengalirkan senyawa organik kebagian pucuk tanaman.
Kofaktor : Setiap molekul atau ion non protein yang diperlukan agar suatu enzim bisa berfungsi dengan baik. Kofaktor dapat berikatan secara permanen dengan tempat aktif enzim atau bisa berikatan secara longgar dengan subtrat selama kalisis.
Meristem : Jaringan tanaman yang terdiri dari sel-sel hidup dan berdinding tipis yang mampu membelah berulang-ulang.
Nursery : Tempat untuk pemeliharaan tanaman. Struktur atau bangunan (biasanya bernaungan) tempat tanaman dirawat , diperbanyak dan diperbesar hingga siap dipasarkan.
Planlet :Tanaman lengkap hasil regenerasi kultur in vitro
Plasmid : Molekul DNA yang biasanya sirkular, yang terpisah dari kromosom tuan rumah sering ditemukan dalam bakteri dan beberapa sel jenis lain.
Primer (pemula) : Oligonukleotid untai tunggal pendek yang bila melekat melalui pasangan basa pada molekul cetakan untai tunggal, akan bertindak sebagai titik permulaan sintesis untai yang komplementer yang diarahkan oleh enzim polimerase DNA.
Primer spesifik : Primer yang susunan nukleotidanya tertentu dan merupakan komplemen dari cetakan DNA yang akan dianalisis
Rambut akar : Seperti tabung yang tidak bercabang, terbentuk di bagian belakang daerah pemanjangan akar, permukaan luarnya berlendir dan berfungsi memperluas permukaan serapan akar
Reaksi rantai
Polimerase (PCR) : Suatu teknik untuk perbanyakan DNA in vitro dengan cara menginkubasi dengan primer khusus, molekul DNA polimerase dan nukleotida.
Rekombinasi : Pertukaran urutan DNA antara molekul-molekul yang berbeda yang terjadi baik secara alamiah maupun sebagai hasil manipulasi DNA.
Sistim jaringan
vaskuler : Sistem yang dibentuk oleh xilem dan floem diseluruh tumbuhan, yang berfungsi sebagai sistem transpor untuk air (xilem) dan nutrien (floem)
Stele : Silinder pembuluh pusat pada akar di mana xilem dan floem berada
T-DNA : Bagian plasmid Ri yang ditransfer pada DNA tanaman.
Transformasi : Proses pengambilan DNA asing oleh suatu sel yang kemudian DNA asing tersebut dapay berintegrasi dengan DNA kromosom dari sel yang mengambilnya melalui proses rekombinasi.
Virulen :Kemampuan patogen untuk menyebabkan penyakit.
Zat pengatur
DAFTAR ISI
Halaman
DAFTAR TABEL ...
xiii
DAFTAR GAMBAR ...
xv
DAFTAR LAMPIRAN ...
xvii
PENDAHULUAN
Latar Belakang... 1Perumusan Masalah... 3
Tujuan Penelitian... 4
Hipotesis... 5
Kegunaan Penelitian... 5
Strategi Penelitian... 5
TINJAUAN PUSTAKA
Tinjauan Umum Tanaman Manggis ... 8Sistem Perakaran dan Upaya Perbaikan Akar Bibit Manggis.. 10
Perbaikan Sistem Perakaran Tanaman dengan Transformasi Agrobacterium rhizogenes ... 17
PENINGKATAN PERAKARAN BIBIT MANGGIS MELALUI
INOKULASI
Agrobacterium rhizogenes
TERHADAP SEMAI
MANGGIS
Abstrak ... 27Abstract ……….. 28
Pendahuluan ………. 29
Bahan dan Metode ……….. 31
Hasil Penelitian ………. 43
Pembahasan ... 62
Simpulan ... 69
INDUKSI AKAR EKSPLAN MANGGIS (
Garcinia mangostana
L.)
DENGAN
Agrobacterium rhizogenes
SECARA
IN VITRO
Abstrak ... 70Abstract ……….. 71
Pendahuluan ………. 72
Bahan dan Metode ……….. 73
Hasil Penelitian ………. 77
Pembahasan ………. 88
Simpulan ……… 93
PEMBAHASAN UMUM ………..
95SIMPULAN DAN SARAN UMUM ...
105DAFTAR TABEL
Halaman
1. Panjang akar primer bibit (cm) dan jumlah akar sekunder bibit
manggis umur 24 MSI ... 43
2. Jumlah akar tertier dan pertambahan diameter batang bibit manggis umur 24 MSI ... .. 44
3. Pertambahan jumlah daun dan tinggi bibit manggis umur 24 MSI ... 46
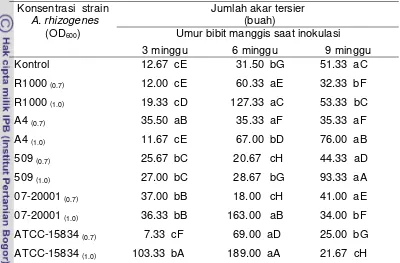
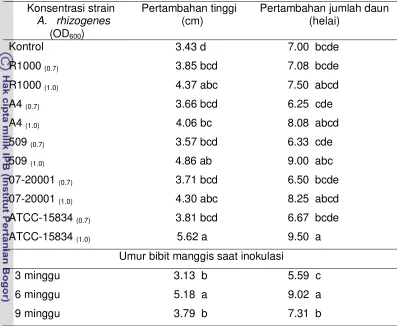
4. Panjang akar primer, jumlah akar sekunder dan panjang akar tampak bibit manggis umur 15 bulan setelah inokulasi dengan berbagai strain A. rhizogenes... .... 49
5. Jumlah akar tertier bibit manggis umur 15 BSI... 50
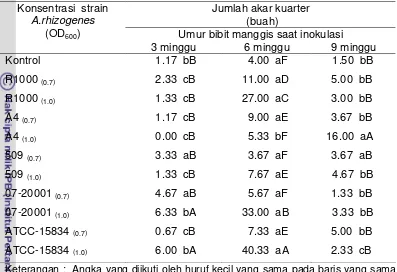
6. Jumlah akar kuarter bibit manggis umur 15 BSI... 51
7. Anatomi akar bibit manggis umur 15 bulan BSI... 52
8. Pertambahan tinggi dan jumlah daun bibit manggis 15 bulan BSI... 56
9. Pertambahan diameter batang bibit manggis umur 15 bulan BSI... 57
10. Berat kering tajuk bibit manggis umur 15 bulan BSI... 58
11. Serapan hara daun bibit manggis umur 15 bulan BSI... 60
12. Kandungan hormon IAA akar bibit manggis umur 15 bulan BSI... 61
13. Persentase bertunas dan pembentukan tunas majemuk dari 1-8 MSP ……… 77
14. Pengaruh inokulasi beberapa strain A.rhizogenes terhadap kecepatan terbentuknya akar eksplan pucuk manggis dengan biji... 79
15. Penambahan diameter batang, tinggi dan jumlah daun tanaman manggis setelah diaklimatisasi ... 81
16. Panjang akar primer, jumlah akar sekunder, dan jumlah akar tersier tanaman manggis setelah diaklimatisasi ... 82
18. Pengaruh inokulasi beberapa strain A.rhizogenes terhadap kecepatan terbentuknya akar eksplan pucuk manggis tanpa biji... 87
19. Pertumbuhan kultur setelah di inokulasi dengan berbagai strain
DAFTAR GAMBAR
Halaman
1. Alur kerja penelitian... 6
2. Penampang membujur zona pertumbuhan pada ujung akar... 10
3. Sayatatan melintang akar tumbuhan dikotil... 12
4. Peta genetik Agrobacterium rhizogenes (Jeng-Sheng 2001)……….. 17
5. Biosíntesis IAA pada tanaman dan bakteri………. 23
6. Bahan tanam ……….. 33
7. Tahapan Inokulasi………. 33
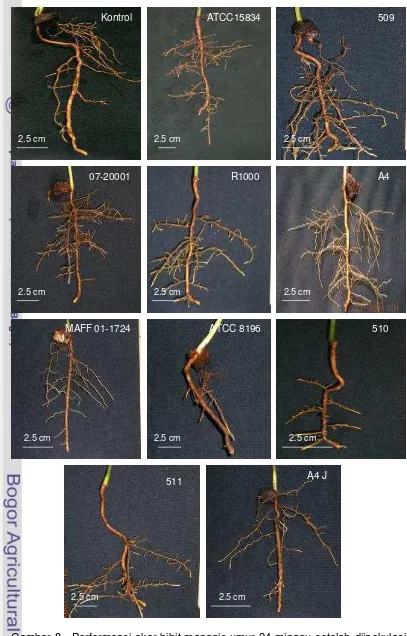
8. Performansi akar bibit manggis yang diinokulasi A. rhizogenes dengan metode akar dipotong...………. ... 45
9. Performansi bibit manggis hasil inokulasiberbagai stain A. rhizogenes dengan metode akar dipotong...…………... 47
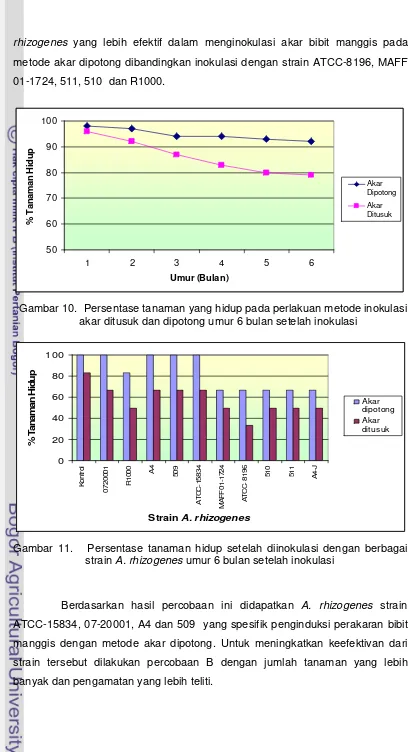
10. Persentase tanaman yang hidup pada berbagai perlakuan metode inokulasi strain A.rhizogenes... 48
11. Persentase tanaman yang hidup pada berbagai strain A.rhizogenes.. 48
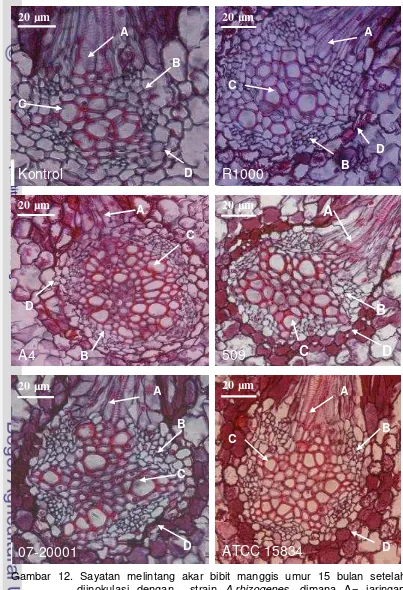
12. Sayatan melintang akar bibit manggis... 53
13. Rambut akar dari akar sekunder bibit tanaman manggis umur 15 bulan setelah inokulasi ... 54
14. Hasil Scanning Electron Microscop (SEM) rambut akar bibit manggis umur 15 bulan setelah diinokulasi dengan strain A.rhizogenes... 54
15. Performansi bibit manggis umur 15 bulan setelah inokulasi..…... 59
16. Hasil PCR akar bibit manggis yang diinokulasi dengan berbagai strain A. rhizogenes ...... 62
17. Induksi tunas manggis secara kultur in vitro... 78
18.Tunas terbanyak yang berhasil diinduksi secara kultur in vitro..... 78
19. Performansi akar planlet manggis umur 6 minggu setelah inokulasi.... 80
21. Performansi bibit manggis umur 3 bulan setelah aklimatisasi... 83
22. Sayatan melintang akar bibit manggis umur 3 bulan setelah aklimatisasi... 85
23. Primordia akar yang terbentuk 6 MSP... 86
24. Eksplan manggis tanpa biji umur 12 minggu... 88
25. Proses tranfer T-DNA Agrobacterium (de la Riva et al, 1998)... 97
DAFTAR LAMPIRAN
Halaman
1. Metode perhitungan anatomi akar bibit manggis... 115
2. Pembuatan larutan scanning electron microscop..... 116
3. Komposisi larutan dan bahan-bahan yang digunakan untuk isolasi DNA, PCR dan elektroforesis... 117
4. Penetapan kandungan nitrogen dengan metode semi-mikro kjeldahl.. 118
5. Penetapan kandungan P dan K dengan metode pengabuan... 119
6. Prosedur analisis hormon IAA... 120
7. Komposisi media MS ... 121
PENDAHULUAN
Latar Belakang
Manggis (Garcinia mangostana L) merupakan tanaman asli Asia Tenggara dan diduga berasal dari Indonesia (Yaacob & Tindall 1997). Buah
manggis dianggap sebagai salah satu jenis buah yang terbaik, rasanya lezat dan
segar sehingga menyebabkan buah ini banyak disukai dan dijuluki sebagai
“Queen of the tropical fruit”. Manggis memiliki prospek yang cerah untuk dikembangkan di Indonesia karena kesesuaian agroklimat dan ketersediaan
lahan.
Di Indonesia tanaman manggis ditemukan di sebagian besar wilayah
dan tersebar mulai dari pulau Sumatera, Jawa, Bali, Kalimantan, Sulawesi, Nusa
Tenggara dan Kepulauan Maluku. Luasnya penyebaran tanaman manggis ini
mengakibatkan panjangnya masa panen manggis di Indonesia bila dibandingkan
dengan masa panen di negara-negara penghasil manggis lainnya. Hal ini
merupakan suatu potensi yang jika dimanfaatkan akan dapat menambah
pendapatan masyarakat.
Buah manggis selain dikonsumsi dalam negeri juga telah diekspor ke
mancanegara. Pada saat ini volume ekspor manggis secara konsisten terus
meningkat sejak tahun 1999, dan pada tahun 2005 ekspor manggis mencapai
8.471 ton dengan nilai US $ 6,3 juta sehingga menempati urutan pertama dari
seluruh ekspor buah segar nasional (Departemen Pertanian 2005).
Pembudidayaan tanaman manggis seharusnya telah lebih maju dari
tanaman buah-buahan tropika lainnya, akan tetapi tanaman manggis yang ada
sekarang masih sangat sedikit mendapat input budidaya. Tanaman manggis
yang ada sekarang sebahagian besar tanaman manggis tua yang terdapat di
hutan-hutan campuran, kebun campuran dan pekarangan rumah masyarakat
dimana pertumbuhan dan perkembangannya tergantung pada alam sehingga
produktivitas manggis belum optimal. Pembudidayaan tanaman manggis yang
relatif lambat dibanding tanaman buah tropika lain diduga berhubungan dengan
laju pertumbuhan manggis yang sangat lambat dengan masa yuwana (juvenil)
yang panjang (8–15 tahun) sehingga menimbulkan keengganan para petani,
Banyak laporan penelitian menyebutkan bahwa lambatnya
pertumbuhan manggis antara lain disebabkan oleh (a) buruknya sistem
perakaran, sehingga (b) penyerapan air dan hara lambat, (c) rendahnya laju
fotosintesis, dan (d) rendahnya laju pembelahan sel pada meristem pucuk
(Wible, Chacko & Downtown, 1992; Ramlan et al. 1992; Poerwanto, 2000). Pada tanaman manggis akar tumbuh dengan sangat lambat, rapuh, jumlah akar
lateral terbatas dan tidak mempunyai rambut akar, mudah rusak, dan terganggu
akibat lingkungan yang tidak menguntungkan, sehingga luas permukaan kontak
antara akar dan media tumbuh sempit yang menyebabkan serapan air dan hara
terbatas (Cox 1988). Rendahnya serapan hara dan air ke dalam tanaman
kurang mendukung aktifitas fisiologi tanaman dan menganggu ritme endogen
secara keseluruhan di dalam tanaman (Hidayat 2002).
Upaya penelitian untuk memperbaiki sistem perakaran bibit manggis
telah dilakukan oleh Poerwanto et al. (1998) dengan pemberian endomikoriza
Gigaspora sp. pada bibit manggis dan Hidayat et al. (1999) dengan pemberian IBA (50-150 ppm) pada biji dan akar manggis. Percobaan untuk meningkatkan
pertumbuhan bibit manggis secara in vitro juga telah dilakukan oleh Goh et al. (1990); Te-chato (1997) dan Pertamawati (2003). Pada penelitian ini dilakukan
percobaan menginokulasi bibit manggis dengan bakteri Agrobacterium
rhizogenes. Perbaikan sistem perakaran menggunakan A. rhizogenes telah banyak dilaporkan sebelumnya pada tanaman buah-buahan, seperti pada kiwi
(Rugini et al. 1991), apel (Sutter & Luza 1993; Damiano & Monticelli 1998), almond (Damiano et al. 1995), walnut (Caboni et al. 1996) dan pada beberapa tanaman tahunan berkayu, seperti pada Eucalyptus (MacRae & Staden 1993),
Larix dan pinus (McAfee et al. 1993; Li & Leung 2001).
Agrobacterium rhizogenes merupakan bakteri tanah gram-negatif yang termasuk pada kelompok Rhizobiacea, mempunyai kemampuan untuk
menstranfer sebagian bahan genetiknya (DNA) pada sel tanaman yang luka
(Nilson & Olsson 1997; Han et al. 1997). DNA yang ditransfer disebut dengan T-DNA yang memiliki gen-gen untuk mensintesis fitohormon yaitu auksin. Salah
satu fungsi auksin adalah diperlukan dalam proses pembelahan sel dan inisiasi
akar (Davies 2004).
memperbaiki sistem perakaran manggis dengan membentuk akar-akar adventif.
Akar ini dapat membantu peningkatan proses penyerapan air dan unsur hara
yang diperlukan dalam metabolisme tanaman, sehingga pertumbuhan tanaman
manggis yang telah ditransformasi dengan menggunakan A. rhizogenes akan lebih baik dibandingkan yang tidak diinokulasi.
Perumusan Masalah
Produksi bibit merupakan faktor penting pada suatu mata rantai usaha
di bidang pertanian sehingga tersedianya bibit bermutu dalam jumlah banyak
dan dalam waktu yang relatif singkat sangat diharapkan dalam menunjang
keberhasilan pengembangan budidaya dan perbaikan kualitas produksi. Kendala
dalam pengembangan usaha tanaman manggis antara lain pertumbuhannya
yang lambat yang disebabkan oleh buruknya sistem perakaran, sehingga
penyerapan air dan hara menjadi lambat (Wible et al. 1992; Poerwanto 2000). Menurut Damiano & Monticelli (1998), pada tanaman tahunan berkayu
perakaran merupakan masalah utama yang sulit dipecahkan untuk bibit yang
diperbanyak dengan perkecambahan biji maupun in vitro. Hal ini disebabkan tingkat oksidasi fenol yang tinggi dan kurangnya ko-faktor perakaran (Mariska &
Purnamaningsih 2001).
Pada penelitian ini dilakukan inokulasi bakteri A. Rhizogenes pada bibit manggis dengan tujuan untuk meningkatkan sistem perakaran tanaman
manggis. Bakteri A. rhizogenes diharapkan dapat menginfeksi tanaman manggis, sehingga menyebabkan terjadinya proliferasi akar. Akar dapat terbentuk karena
terjadi transfer sebagian fragmen DNA yaitu T-DNA dari Agrobacterium ke dalam sel tanaman. T-DNA tersebut membawa gen-gen yang terlibat dalam
proses induksi akar yaitu daerah root loci (rol) A, B, C dan D pada bagian TL (Slightom et al. 1986; Chriqui et al. 1996), sedangkan bagian TR membawa gen iaaM dan iaaH yang terlibat dalam biosintesis auksin, selain itu transformasi ini
membawa gen yang berguna untuk menyandi sintesis senyawa opin (Giri &
Narasu 2000).
Integrasi T-DNA plasmid Ri ke dalam genom sel tanaman, apabila
reaksi hipersensitif tanaman terhadap auksin sehingga menyebabkan
berkembangnya akar (Jeng-Sheng 2001; Gelvin 2003). Akar yang terbentuk
dapat terus tumbuh walaupun bakteri sudah mati. Selain itu, jaringan tersebut
dapat tumbuh secara in vitro dalam media tanpa zat pengatur tumbuh auksin yang biasa diperlukan untuk memacu pertumbuhan jaringan tanaman.
Transformasi genetik ini terjadi secara alami, karena adanya kontak langsung
antara bakteri dengan sel tanaman. Daerah TL-DNA dari A. rhizogenes lebih penting untuk menginduksi akar, karena rolB berperan meningkatkan pool
auksin aktif dalam tanaman dengan hidrolisis konjugat IAA inaktif dan mengatur
sensivitas sel terhadap IAA. Gen rolC berperan meningkatkan level sitokinin melalui aktivitas β-glucosidase yang mampu melepaskan sitokinin aktif dari konjugatnya. Hasil akhir dari ekspresi berbagai gen rol pada T-DNA dari Ri-plasmid adalah terbentuknya jaringan akar (Jeng-Sheng 2001; Valpuesta 2002).
Sampai sejauh ini, pemanfaatan bakteri A. rhizogenes untuk
meningkatkan sistem perakaran tanaman manggis belum pernah dilaporkan.
Oleh karena itu perlu diteliti bagaimana peranan bakteri A. rhizogenes dalam meningkatkan sistem perakaran tanaman manggis sehingga didapatkan metode
yang sesuai untuk memperbaiki sistem perakaran tanaman manggis.
Perbaikan sistem perakaran tanaman manggis diharapkan dapat
meningkatkan pertumbuhan karena perakaran yang lebih baik dapat
meningkatkan suplai hara ke tanaman, meningkatkan laju metabolisme tanaman
dan akhirnya dapat meningkatkan produksi tanaman. Dari penelitian ini
diharapkan diperoleh suatu teknologi yang dapat meningkatkan sistem
perakaran pada bibit tanaman manggis sehingga mempercepat pertumbuhan
bibit di lapangan.
Tujuan Penelitian
Tujuan Umum
Secara umum penelitian ini bertujuan untuk mendapatkan teknologi
perbaikan sistem perakaran tanaman manggis melalui inokulasi A. rhizogenes
Tujuan Khusus
1. Mendapatkan galur A. rhizogenes yang dapat menginfeksi akar manggis baik secara in vivo maupun in vitro.
2. Mempelajari sistem perakaran tanaman manggis (Garcinia mangostana L.) yang telah diinfeksi A. rhizogenes.
3. Mendapatkan bibit tanaman manggis yang telah terintegrasi T-DNA dari A. rhizogenes.
Hipotesis
1. Satu atau beberapa galur A. rhizogenes dapat menginfeksi tanaman manggis baik secara in vivo maupun in vitro.
2. T-DNA dari A. rhizogenes dapat terintegrasi pada kromosom tanaman manggis.
3. Inokulasi A. rhizogenes dapat menginduksi pembentukan rambut akar pada akar semai dan planlet manggis.
4. Infeksi A. rhizogenes memberikan pengaruh terhadap pertumbuhan,
morfologi, anatomi akar dan penyerapan hara bibit manggis.
Kegunaan Penelitian
1. Didapatkan landasan untuk langkah-langkah upaya perbaikan sistem
perakaran bibit manggis.
2. Mendapatkan bibit manggis dengan sistem perakaran yang lebih baik dengan
pertumbuhan yang lebih cepat.
3. Diperoleh suatu teknologi yang dapat meningkatkan sistem perakaran pada
tanaman manggis sehingga dapat mempercepat pertumbuhan bibit manggis
Strategi Penelitian
Disertasi ini disusun berdasarkan dua topik penelitian. Topik penelitian
pertama adalah “Peningkatan perakaran bibit manggis melalui inokulasi A.
rhizogenes terhadap semai manggis”, penelitian ini terdiri atas dua tahapan
percobaan kedua yang berjudul “Pengembangan protokol inokulasi A. rhizogenes yang efektif untuk menginduksi perakaran manggis”.
Topik penelitian kedua adalah “Induksi perakaran eksplan tunas manggis dengan A. rhizogenes secara in vitro”, penelitian ini terdiri atas dua tahapan percobaan yang saling berkaitan, yaitu Multiplikasi tunas tanaman manggis melalui kultur in vitro, yang bertujuan untuk mendapatkan media yang terbaik untuk multiplikasi tunas manggis secara in vitro. Eksplan manggis yang dihasilkan pada percobaan ini, diinduksi perakarannya dengan berbagai
galur A. rhizogenes pada percobaan kedua yang berjudul “Induksi perakaran eksplan tunas manggis dan aklimatisasi planlet manggis hasil inokulasi A. rhizogenes. Percobaan ini bertujuan untuk mendapatkan galur A. rhizogenes
yang dapat menginduksi perakaran eksplan tunas manggis secara in vitro. Bagan alur kerja penelitian yang menunjukkan keterkaitan antar
Gambar 1. Alur kerja penelitian
Agrobacterium
rhizogenes
Peningkatan
perakaran
bibit
manggis
melalui
inokulasi
A.
rhizogenes
terhadap semai manggis
Induksi perakaran eksplan tunas
manggis dengan
A. rhizogenes
secara
in vitro
Seleksi galur
A. rhizogenes
spesifik
yang menginduksi perakaran bibit
manggis
Pengembangan protokol inokulasi
A. rhizogenes
yang efektif untuk
menginduksi perakaran manggis
Mendapatkan bibit manggis dengan sistem perakaran
yang baik dengan pertumbuhan yang lebih cepat
Multiplikasi
tunas
tanaman
manggis melalui kultur
in vitro
Induksi perakaran eksplan tunas
manggis dan aklimatisasi planlet
manggis
hasil
inokulasi
A.
rhizogenes
II. TINJAUAN PUSTAKA
2.1. Tinjauan Umum Tanaman Manggis
Manggis (Garcinia mangostana L.) yang termasuk ke dalam familia Guttiferae merupakan tanaman yang berasal dari daerah Asia Tenggara
khususnya Thailand, Malaysia dan Indonesia (Nakasone & Paull 1999).
Tanaman manggis merupakan pohon besar berdaun lebar dan rimbun. Tinggi
pohon yang telah dewasa mencapai 10-25 m. Bentuk tajuk pohon bervariasi
dari bulat silindris hingga kerucut dengan penyebaran simetris ke semua arah.
Lebar tajuk merentang hingga 12 m dan semakin mengecil ke arah puncak
pohon. Diameter batang pokok pohon dewasa dapat mencapai 60 cm dengan
percabangan ke semua arah. Daunnya tunggal dan berpasangan di sisi ranting.
Bentuk daun bulat panjang dengan ukuran panjang 13-26 cm dan lebar 6-12 cm.
Helai daunnya kaku dan tebal. Daun muda yang baru tumbuh berwarna cokelat
kemerahan, kemudian sesuai dengan umur pertumbuhannya berubah menjadi
cokelat kehijauan, hijau muda, lalu hijau tua (Tirtawinata et al. 2000).
Bunga manggis terletak di ujung ranting, memiliki tangkai bunga yang
pendek dan tebal, daun kelopak empat helai tersusun dalam dua pasang dan
daun mahkota empat helai. Kedua pasang kelopak memiliki panjang 2 cm,
berwarna hijau kekuningan, berlekuk dan tumpul, sedangkan mahkotanya
berwarna hijau kekuningan dengan bagian di sekelilingnya berwarna kemerahan,
tebal, tumpul dan berdaging. Bunga manggis muncul secara menyendiri atau
berpasangan pada bagian ujung ranting di luar kanopi (Nakasone & Paull 1999).
Bunga manggis adalah dioecious (berumah dua), tetapi hanya bunga betina
yang banyak ditemui karena bunga jantan tidak berkembang sempurna (Cox
1988).
Proses pembentukan dan perkembangan buah manggis terjadi pada
laju yang konstan antara 100–160 hari dari awal pembungaan hingga
pematangan buah. Buah manggis berdiameter 4–8 cm, berbentuk bulat,
berwarna kekuningan hingga berwarna ungu kehitaman pada saat masak dan
beratnya berkisar 30-180 g. Daging buah (aril) terdiri atas 5-7 segmen berwarna
putih, rasanya manis dan hanya mengandung 1-2 biji. Menurut Prove (1998),
komponen nutrisi dalam 100 g buah manggis yang dapat dimakan adalah : 34,0
7,0 mg kalsium; 13,0 mg magnesium; 13,0 mg fosfor; 7,0 mg sodium; 45,0
potasium; 1,0 mg zat besi; 0,03 mg vitamin B1; 0,03 mg vitamin B2; dan 4,2 mg
asam arkorbat (vitamin C).
Buah manggis dapat disajikan dalam bentuk segar sebagai buah
kaleng, dibuat sirop dan sari buah. Secara tradisional buah manggis adalah
obat sariawan, wasir dan luka. Kulit buah dimanfaatkan sebagai pewarna
termasuk untuk tekstil dan air rebusannya dimanfaatkan sebagai obat diare.
Batang pohon dipakai sebagai bahan bangunan, kayu bakar dan kerajinan.
Biji manggis merupakan biji apomiktik, yaitu biji yang dihasilkan tanpa
fertilisasi, berwarna coklat, pipih, tidak berendosperm yang ditutupi
permukaannya oleh jaringan pembuluh (vascular bundles). Biji manggis bersifat poliembrioni dan nutrisi untuk perkembangan embrionya didukung oleh nuselus
atau jaringan integumen dan inti endosperm. Secara normal biji manggis selalu dalam keadaan lembab dan bila keadaan lembab tersebut berkurang maka biji
dapat mati, keadaan biji seperti ini dikenal dengan nama recalcitrant seed. Sekitar 10% dari biji yang berkecambah dapat menumbuhkan lebih dari satu
tunas dan masing-masing tunas dapat tumbuh pada posisi yang berlainan dan
masing-masing membawa perakarannya sendiri-sendiri (Nakasone & Paull
1999).
Tanaman manggis dapat tumbuh baik pada ketinggian 460-610 m dpl
di atas permukaan laut. Verheij (1992) menyatakan bahwa di daerah tropis
tanaman manggis masih dapat tumbuh pada ketinggian tempat lebih dari 1000
meter di atas permukaan laut, tetapi semakin tinggi tempat tumbuh akan
semakin lambat pertumbuhannya dan semakin lama awal pembungaannya.
Tanah yang disukai tanaman manggis adalah jenis tanah gembur
yang kaya kandungan bahan organik dengan drainase yang baik. Tanaman
manggis tumbuh baik pada tanah lempung berpasir dengan kandungan bahan
organik yang tinggi, di samping itu untuk pertumbuhan yang optimum tanah
harus subur, air tanahnya harus dangkal dengan ke dalaman 2-3 meter dari
permukaan tanah dan dijaga agar tanah tidak sampai kering. Derajat keasaman
tanah yang baik untuk tanaman manggis antara 5-7, tetapi tanaman toleran
terhadap pH tanah yang rendah.
Untuk kesuksesan penanaman, manggis membutuhkan curah hujan
1500-2500 mm per tahun, untuk menstimulir pembungaan tanaman manggis
membutuhkan curah hujan lebih dari 100 mm per bulan dengan musim kering
yang pendek. Menurut Tirtawinata et al. (2000) bahwa pada masa awal pertumbuhan, manggis menyukai naungan, akan tetapi menjelang dewasa, sinar
matahari penuh dapat mempercepat masa awal produksinya.
Nakasone & Paull (1999) mengemukakan bahwa udara yang lembab
dengan suhu udara berkisar 25-35oC sangat menunjang pertumbuhan tanaman
manggis. Pada suhu di bawah 20oC pertumbuhannya terhambat dan suhu di
bawah 5oC dan di atas 38oC menyebabkan kematian tanaman manggis.
Kelembaban udara optimal untuk tanaman manggis ialah sekitar 80% (Verheij
1992).
2.2. Sistem Perakaran dan Upaya Perbaikan Akar Bibit Manggis
2.2.1. Pertumbuhan dan Perkembangan Akar
Organ yang pertama terbentuk pada kebanyakan tanaman adalah
akar. Akar yang tumbuh langsung dari benih (radikel) berkembang menjadi
akar primer atau disebut akar tunggang (tap root) pada tanaman dikotil. Pertumbuhan lebih lanjut dari akar primer tergantung pada aktivitas dari
meristem apikalnya. Pembelahan sel berlansung sangat aktif pada bagian
meristem akar ini. Bagian meristem akar ini dilindungi oleh tudung akar (root cap). Peranan tudung akar penting sekali dalam proses pemanjangan akar pada saat akar melakukan penetrasi ke dalam tanah. Tudung akar juga
menghasilkan sejenis bubur polisakarida yang disebut musigel (mucigel) yang berfungsi sebagai pelumas untuk mempermudah penetrasi akar ke dalam tanah
(Lakitan 1995).
Sel-sel muda yang terbentuk pada meristem kemudian berkembang
menjadi sel-sel epidermis, korteks, endodermis, perisikel, xilem, dan floem. Di
balik tudung akar (di depan meristem) terdapat suatu zona yang terdiri beberapa
sel yang tidak aktif membelah diri. Zona ini disebut quinscent center. Zona ini berfungsi sebagai pengganti jika tudung akar atau meristem mengalami
kerusakan. Zona pemanjangan (elongation zone) akar berkisar antara 0.5-1.5 cm pada bagian ujung akar. Laju pemanjangan akar dapat mencapai 2 cm/hari
(Gambar 2). Akar primer memanjang lebih cepat dibandingkan dengan akar
dengan akar tersier. Laju pemanjangan akar juga dipengaruhi oleh faktor
internal dan berbagai faktor lingkungan. Faktor internal yang mempengaruhi laju
tersebut adalah pasokan fotosintat (umumnya dalam bentuk sukrosa) dari daun.
Faktor lingkungan yang berpengaruh antara lain suhu tanah dan kandungan air
tanah (Campbell et al. 2000).
Selain tumbuh memanjang, akar juga tumbuh secara radial. Akar
tanaman gimnosperma dan tanaman dikotil mempunyai kambiun vaskuler yang
terletak pada posisi di antara pembuluh floem dan xilem. Kambiun berperan
dalam penambahan diameter akar (pertumbuhan radial), terutama karena
kambiun ini berperan dalam pembentukan sel-sel xilem (ke arah internal) dan
sel-sel floem (ke arah eksternal). Tanaman monokotil tidak memiliki kambiun
vaskuler. Pertumbuhan radial pada akar tanaman monokotil hanya disebabkan
oleh pembesaran sel-sel nonmeristematik. Dengan demikian, pertumbuhan
radial pada akar tanaman monokotil sangat terbatas.
Gambar 2. Penampang membujur zona pertumbuhan pada ujung akar (Sumber : Campbell et al. 2000)
Akar primer selanjutnya akan membentuk cabang yang disebut
sebagai akar sekunder. Akar sekunder umumnya tumbuh secara lateral
(horizontal) oleh sebab itu sering pula disebut sebagai akar lateral. Akar
sekunder ini terbentuk beberapa milimeter atau beberapa sentimeter dari ujung
Epidermis
Rambut akar
Stele
Kortek
Tudung
akar Meristem
apikal
Zona
pemanjangan
Zona
pembelahan Zona diferensiasi
Meristem primer
akar primer. Pertumbuhan akar sekunder dimulai pada sel-sel perisikel calon
akar sekunder ini sangat aktif membelah diri dan tumbuh menembus lapisan
sel-sel korteks dan epidermis akar primer. Sel-sel-sel perisikel calon akar sekunder ini
diduga menghasilkan enzim hidrolitik yang berperan mengurai bahan-bahan
penyusun dinding sel korteks dan epidermis yang dilalui dalam proses
pertumbuhannya (Lloret & Casero 2000). Melalui proses yang sama, akar-akar
tertier akan tumbuh dari sel-sel perisikel akar sekunder (Lakitan 1995).
2.2.2. Rambut Akar
Absorpsi air dan zat-zat terlarut oleh tumbuhan berlansung melalui
sistem perakaran. Sebagian besar absorbsi terjadi pada daerah rambut akar
yang terletak beberapa milimeter di atas ujung akar. Rambut akar adalah sel
epidermis berbentuk tabung memanjang mempunyai vakuola lebar dan biasanya
berdinding tipis, hanya beberapa tumbuhan rambut tersebut bercabang. Rambut
akar panjangnya 80–1500 µm dengan diameter antara 5–20 µm dan dapat
mencapai 200 lembar/mm2 (Hidayat 1995).
Rambut akar mulai dibentuk di luar daerah meristematik bagian akar
muda yang epidermisnya masih dapat memanjang. Rambut akar biasanya
pertama kali tampak sebagai gelembung kecil di dekat ujung apikal sel
epidermis. Jika sel epidermis terus memanjang setelah terlihat adanya
gelembung, rambut akar ditemukan agak jauh dari ujung apikal sel epidermis
yang menjelang dewasa. Rambut akar memanjang di ujungnya yang dindingnya
tipis, lunak dan lebih lembut.
Pada beberapa tanaman hanya sel epidermis akar tertentu yang
disebut trikoblas yang dapat menghasilkan rambut akar, yakni berupa sel-sel
kecil hasil pembelahan sel epidermis yang tidak sama. Cutter & Feldman (1970)
dalam Fahn (1995) mempelajari trikoblas pada Hydrocharis, selama perkembangan trikoblas, nukleus dan nukleolus bertambah volumenya.
Trikoblas berisi lebih banyak nukleohiston, protein total, RNA dan DNA inti.
Trikoblas tidak berbagi, dan nukleusnya makin menjadi poliploid makin jauh dari
ujung akar. Hal tersebut merupakan akibat pengunduran proses pendewasaan
dari rambut akar yang berkembang. Terlambatnya pendewasaan ini mungkin
Rambut akar biasanya hanya hidup dalam waktu yang singkat,
umumnya hanya beberapa hari. Dengan kematian rambut akar dan jika sel tidak
mengelupas, dinding sel epidermis menjadi bergabus dan lignin. Pada
beberapa tumbuhan, rambut akar ditemukan tetap ada pada tumbuhan. Dinding
dari rambut akar seperti itu menebal dan kehilangan kemampuan mengambil air
dari tanah.
2.2.3. Struktur Internal Akar
Epidermis adalah jaringan pelindung dan terdiri atas satu lapisan sel
yang tersusun padat. Di bawah epidermis terdapat daerah yang relatif tebal
disebut korteks. Korteks terutama tersusun dari sel-sel yang tidak terspesialisasi
secara struktural, sel parenkima, dengan ruang antar sel yang luas. Lapisan
terdalam korteks terdiri atas sebaris sel disebut endodermis. Dalam keadaan
primer dinding semua sel endodermis itu tipis kecuali penebalan seperti pita
pada sisi-sisi radial dan melintang sel terdebut. Penebalan ini dikenal sebagai
jalur Caspary atau pita Caspary (Fahn 1995). Pada akar primer jalur caspary
merupakan batas dalam dari ruang bebas dan tidak permeabel terhadap air dan
zat-zat terlarut, sehingga air dan ion-ion terlarut dipaksa melewati protoplas sel
untuk mencapai jaringan pembuluh (Harran & Tjondronegoro 1992).
Gambar 3. Sayatan melintang akar tumbuhan dikotil (Sumber : Campbell et al. 2000)
Xilem
Perisikel Floem kambium
Endodermis
Rambut akar Kortek
Bagian tengah akar dinamakan silinder pembuluh. Silinder ini terdiri
atas jaringan penyalur air (xilem) dan jaringan penyalur makanan (floem).
Antara jaringan pembuluh (xilem dan floem) dan endomermis terdapat lapisan
sel parenkima yang tak terpsesialisasi (perisikel) yang berasal dari kumpulan sel
meristimatik yang sama seperti xilem dan floem. Perisikel yang tetap
mempertahankan sifat meristematiknya membentuk akar-akar lateral. Xilem
terdiri atas sel-sel penyalur (trakeid) dan anggota pembuluh maupun serat dan
parenkima. Trakeid dewasa merupakan sel tunggal yang memanjang. Di dalam
dinding yang menebal terdapat bagian-bagian tipis (noktah) yang dapat
menyalurkan air dengan mudah. Anggota pembuluh juga terdiri atas sel-sel
tunggal dengan dinding yang serupa dengan trakeid tetapi penuh
berlubang-lubang ujungnya (Gambar 3). Sel-sel tersebut biasanya lebih pendek
dibandingkan trakeid dan tersusun dalam baris vertikal. Sebaris anggota
pembuluh dinamakan trakea. Serat merupakan sel lancip memanjang yang
berdinding tebal terutama berfungsi dalam memperkuat jaringan. Parenkima
merupakan semacam jaringan pengisi dan berfungsi dalam penyimpanan
makanan (Mauseth 1988).
Floem terdiri atas pembuluh tapis, sel pengiring, serat dan parenkima.
Anggota pembuluh tapis merupakan sel hidup, tersusun dalam barisan vertikal,
yang dikenal sebagai jaringan pembuluh tapis dan berfungsi dalam translokasi
zat-zat organik. Sel-sel pengiring merupakan sel seasal dengan anggota
pembuluh tapis dan tetap berhubungan rapat sesamanya.
2.2.4. Sistem Perakaran Tanaman Manggis
Tanaman manggis biasa diperbanyak dengan menggunakan biji, waktu
yang dibutuhkan untuk perkecambahan antara 10–45 hari. Perkecambahan
dimulai dengan pembengkakan pada benih. Akar pertama muncul dari satu
bagian pembengkakan (ujung), sedangkan tunas akan tumbuh dari bagian
pembengkakan yang lain. Selanjutnya sistem perakaran berkembang dari
bagian dasar tunas dan sistem perakaran yang pertama terbentuk berhenti
berfungsi (Verheij 1992).
Satu bulan setelah biji berkecambah, sistem perakaran tanaman
manggis masih sangat jarang. Bijinya tetap melekat pada pangkal tunas sampai
masing-masing masih memperlihatkan perakarannya. Pada umur 2-4 bulan
terjadi peningkatan akar sekunder, sedangkan pertumbuhan akar tersier dimulai
pada umur 3 bulan. Akar sekunder maupun tersier tebal, dengan permukaan
halus dan tidak berakar rambut pada semua stadia tumbuh (Rukayah &
Zabedah 1992).
Pertumbuhan tanaman manggis yang lambat berkaitan erat dengan
sistem perakarannya. Tanaman manggis mempunyai akar tunggang yang
panjang dan kuat tetapi percabangan akarnya sangat sedikit, juga tidak memiliki
bulu-bulu akar. Uniknya di antara seluruh spesies Garcinia, hanya Garcinia mangostana saja yang mempunyai perakaran lemah, sedangkan jenis lainnya memiliki perakaran kuat dan lebat. Hasil pemeriksaan sitologi terhadap tanaman
manggis memperlihatkan bahwa tanaman ini mempunyai kromosom poliploid
2n=96 yang sifatnya sangat lemah, laju pembelahan selnya rendah demikian
pula pembesaran selnya lambat, sedangkan spesies Garcinia lainnya yaitu G. Hombroniana dan G. Malaccencis, masing-masingnya memiliki jumlah kromosom, yaitu 2n=48 dan 2n=46 (Verheij 1992).
Menurut Cox (1988) pohon manggis dengan tinggi 3.8 m dan lebar
tajuk 2.5 m mempunyai sebaran akar terbanyak pada kedalaman 5-30 cm dan
akar terpanjang tidak lebih dari 1 m dari pangkal batang. Selain itu Gonzales &
Anoos (1952) dalam Pertamawati (1994) mengatakan bahwa pada setiap
tanaman manggis yang tingginya lebih dari 1 m, rata-rata mempunyai 5,6 akar
primer yang lurus dan panjang, tetapi hanya 1 atau 2 dari akar primer tersebut
yang dapat berkembang baik. Hidayat (2002) melaporkan juga bahwa, semakin
tua tanaman manggis persentase akar tersier (diameter < 2 mm = feeder root) semakin rendah. Sebaliknya persentase akar primer dan akar sekunder
semakin tinggi dengan semakin tuanya umur tanaman manggis. Akar tersier
merupakan akar penyerap air dan hara mineral, sedangkan akar primer dan akar
sekunder berperan sebagai organ penyangga batang dan penyimpan cadangan
karbohidrat. Rendahnya persentase akar tersier pada tanaman manggis
menyebabkan serapan air dan hara rendah.
2.2.3. Upaya Perbaikan Akar Bibit Manggis
Dewasa ini pemerintah sedang menggalakkan komoditas nonmigas,
Negara. Upaya peningkatan ekspor komoditas pertanian memerlukan dukungan
penyediaan bibit untuk memenuhi kebutuhan yang semakin meningkat.
Penyediaan bibit yang berkualitas merupakan salah satu faktor yang
menentukan keberhasilan dalam pengembangan pertanian di masa mendatang.
Beberapa penelitian telah dilakukan untuk memperbaiki sistem
perakaran bibit manggis baik secara konvensional (in vivo) maupun secara in vitro. Pemberian mikorhiza dapat memperbaiki pertumbuhan dan perakaran bibit manggis. Poerwanto et al. (1998) melaporkan bahwa pemberian mikorhiza dapat meningkatkan pertumbuhan bibit manggis umur 4 minggu. Peningkatan
pertumbuhan bibit terbaik diperlihatkan oleh pemberian endomikorhiza
Gigaspora sp dengan meningkatkan panjang akar primer, panjang total akar dan luas daun serta berat kering akar, batang dan daun. Sementara itu, di Malaysia
dilaporkan bahwa mikorhiza jenis Scutellospora calospora dan Glamus mosseae
mampu meningkatkan panjang dan percabangan akar, meningkatkan
pertumbuhan bibit manggis dan mempersingkat waktu di pembibitan dari 24
bulan menjadi 18 bulan (Masri et al. 1998).
Penelitian Hidayat et al. (1999) diketahui bahwa pemberian 50-150 ppm IBA terhadap biji dan akar manggis meningkatkan pertambahan panjang
akar, diameter batang, bobot total tanaman, kandungan dan serapan hara daun
manggis. Pemberian trikontanol (0.075-0.150 ppm) meningkatkan panjang akar,
luas daun, bobot tanaman serta serapan hara daun bibit yang berumur 7 bulan.
Upaya perbaikan sistem perakaran manggis juga dilakukan dengan
mengiduksi perakaran manggis secara in vitro. Goh et al. (1994), berhasil menginduksi perakaran manggis dengan menggunakan IBA yang ditanam
dalam vermikulit. Hasil penelitian Te-chato & Lim (1999) perakaran manggis
lebih cepat terbentuk, lebih panjang dan kualitasnya lebih baik pada perlakuan
perendaman 4,4 mM IBA selama 15 menit dalam gelap dan dikulturkan pada
media WP yang ditambah 34,5 µM phloroglucinol. Perlakuan 10-20 ppm IBA
diinkubasi dalam gelap selama 14 hari memberikan persentase perakaran yang
baik (Triatminingsih et al. 2001).
Pertumbuhan dan perkembangan akar manggis yang dikulturkan
kultur sehingga eksplan yang mempunyai klorofil dapat melakukan proses
fotosintesis dengan optimal. Kultur seperti ini dikatakan bahwa eksplan tumbuh
dalam keadaan fotoautotrofik (perbanyakan mikro dengan media bebas gula).
Ermayanti et al. (1999) menyatakan bahwa planlet manggis yang ditanam dalam media bebas gula, menggunakan substrat vermikulit dan dengan pengaturan
CO2 menghasilkan persen perakaran dan panjang akar yang lebih tinggi
dibandingkan kontrol. Pertamawati (2003) juga mendapatkan bahwa, planlet
manggis yang dikulturkan secara in vitro dalam keadaan fotoautotrofik nyata lebih baik pertumbuhannya, akar planlet lebih panjang dan daunnya lebih luas
dibandingkan dengan keadaan miksotrofik (medium tumbuh mengandung gula).
2.3. Perbaikan Sistem Perakaran Tanaman dengan Transformasi Agrobacterium rhizogenes
2.3.1. Agrobacterium rhizogenes
Agrobacterium adalah bakteri tanah termasuk famili Rhizobiaceae bersifat aerobik, bereaksi negatip terhadap pewarnaan gram, dapat tumbuh
secara saprofit atau parasit dan menyebabkan penyakit tumor (grown gall) dan akar rambut (hairy root) pada tanaman dikotil dan monokotil. Sel-selnya yang normal berukuran 0,6–1,0 µm dan memiliki 1-6 flagela (Schaad 1988). Menurut
Winans (1992) genus ini terdiri dari dua spesies yang bersifat patogen yaitu
Agrobacterium tumefaciens dan Agrobacterium rhizogenes keduanya menginfeksi berbagai tanaman pada bagian yang luka. Infeksi sel bakteri pada
bagian yang luka menyebabkan sel tanaman berproliferasi membentuk tumor
dan akar. Infeksi oleh A. tumefaciens menyebabkan tumor yang tidak terorganisasi dan infeksi oleh A. rhizogenes menyebabkan proliferasi akar.
Induksi tumor dan akar yang terjadi disebabkan oleh terjadinya transfer
sebagian fragmen DNA atau yang disebut dengan T-DNA dari Agrobacterium
ditransfer ke dalam sel tanaman. T-DNA memiliki gen-gen untuk mensintesis
auksin dan sitokinin, sehingga apabila berinfeksi dengan DNA tanaman
mengakibatkan terjadinya over produksi fitohormon tersebut di dalam sel
tanaman (Klee et al. 1987; Winans 1992).
T-DNA ini terdapat pada plasmid besar yang disebut Ti-plasmid (tumor inducing) pada Agrobacterium tumefaciens dan Ri (root inducing) pada
terdapat pada A. rhizogenes umumnya berukuran sekitar 140–235 kbp dan fragmen DNA yang ditransfer ke dalam sel tanaman sekitar 14-42 kb yang
merupakan daerah TR dan TL-DNA. Plasmid Ri yang terdapat pada A.
rhizogenes mempunyai satu atau dua macam T-DNA yaitu left T-DNA (TL-DNA) dan right T-DNA (TR-DNA). Berbeda dengan TL-DNA, TR-DNA mempunyai
persamaan dengan T-DNA A. tumefaciens. TR-DNA mengandung gen-gen
iaaM dan iaaH yang berfungsi dalam biosintesis auksin dan membawa gen-gen
untuk menyandi sintesis senyawa opin (Giri & Narasu 2000). Sedangkan
TL-DNA mengandung gen-gen rol (root loci) yaitu : rolA, rolB, rolC dan rolD (Slightom et al. 1986). Hasil pengujian White et al. (1985) menunjukkan bahwa protein produk dari berbagai gen rol berperan dalam meningkatkan sensitivitas sel tanaman terhadap auksin. Selain itu produk gen rol juga diduga mendorong tanaman inang untuk mensintesis auksin.
Ri-plasmid dapat dibedakan berdasarkan tipe opin yang dihasilkan
yaitu agropin, manopin atau kukumopin (van der Salm et al., 1996). Senyawa-senyawa tersebut merupakan sumber karbon dan nitrogen bagi Agrobacterium
yang tidak dihasilkan oleh tanaman normal. Bakteri dengan tipe agropin
mempunyai kedua bagian DNA, sedangkan bakteri dengan tipe manopin hanya
memiliki TL-DNA.
A. rhizogenes galur agropin yang mengandung TL dan TR-DNA
mengandung lokus rol yang bertanggung jawab terhadap pembentukan penyakit
akar rambut. TL-DNA ini homolog dengan T-DNA tunggal dari galur A. rhizogenes yang lain. TR-DNA membawa gen agropin (ags), mannopin (mas2’ dan mas1’) dan auksin (iaaM dan iaaH) yang terlibat dalam biosintesis auksin yang juga ditemukan pada T-DNA strain A. tumefaciens (Jeng-Sheng, 2001) (Gambar 4). A.rhizogenes tipe agropin mempunyai kisaran inang yang lebih
luas dibandingkan dengan A. rhizogenes tipe lainnya. Umumnya galur
Agrobacterium yang digunakan dalam proses transformasi merupakan galur dengan inang luas dengan demikian strain bakteri tersebut patogenik terhadap
Gambar 4. Peta genetik Agrobacterium rhizogenes (Sumber : Jeng-Sheng 2001).
2. 3.2. Interaksi Agrobacterium dengan Tanaman
Proses transfer T-DNA dari Ti atau Ri plasmid ke dalam genom inti
tanaman terjadi akibat adanya interaksi antara A. tumefaciens atau A. rhizogenes dengan sel tanaman yang luka. Sel tanaman yang luka menghasilkan senyawa gula, asam amino atau senyawa fenol (Winans, 1992).
Dengan adanya isyarat tersebut maka Agrobacterium bergerak aktif menuju sel
tanaman yang luka. Untuk memperkuat interaksi ini Agrobacterium
mengeluarkan β-1,2-glukan. Beberapa gen dalam kromosom Agrobacterium
diketahui merupakan penyandi enzim yang berperan dalam sintesis berbagai
senyawa glukan, yaitu chvA, chvB dan pscA (exoC) serta gen cel yang berfungsi mensintesa selulosa fibril (Zambryski et al. 1989).
Terbentuknya senyawa fenol seperti asetosiringon (AS) dan
hidroksiasetosiringon (OH-AS) dari sel tanaman yang luka menyebabkan
protein sensor (bertugas mendeteksi adanya senyawa fenol) yang disandikan
oleh gen virA. Produk dari virA menginduksi fosforilasi dari produk virG yang kemudian mengaktifkan ekpresi dari berbagai gen vir lainnya (Powell et al. 1989; Zambryski et al. 1989; Winans 1992; Gelvin 2003).
Gen-gen lain yang aktif diantaranya adalah gen virD1 dan virD2.
Produk kedua gen ini merupakan endonuklease yang dapat menimbulkan nick
(terputusnya rantai DNA) pada basa ketiga dan keempat pada LB dan RB
sehingga dapat terjadi proses sintesis utas tunggal (ss)T-DNA. Jika virD1 dan virD2 berada dalam kondisi terbatas, digunakan juga produk virC untuk
meningkatkan sintesis (ss)T-DNA (Gelvin, 2003). Selanjutnya (ss) T-DNA
ditransfer dengan cara membentuk T-kompleks dengan produk virE2 (virE2
-T-DNA). VirE2 mengikat ssT-DNA pada situs yang tidak spesifik, berfungsi untuk
menjaga kestabilan ssT-DNA dari kerusakan akibat nuklease (Jeng-Sheng,
2001; Gelvin, 2003).
Menurut Winans (1992) produk dari virD2 akan tetap terikat pada ujung
5 dari T-kompleks dan diduga berfungsi sebagai protein pemandu dalam
pelepasan T-kompleks dari Agrobacterium ke sel tanaman, sedangkan virD1
tidak lagi berikatan dengan T-kompleks karena tidak diperlukan dalam proses
transfer T-DNA. Produk dari virB menyebabkan terbentuknya pori atau lubang yang dilewati oleh T-DNA pada proses transfer T-kompleks dari bakteri ke
tanaman.
2.3.3. Integrasi dan Ekspresi pada Genom Tanaman
Secara umum proses infeksi A. rhizogenes mirip dibandingkan dengan
A. tumefaciens. Proses transfer ssT-DNA bersifat polar, yaitu bergerak dari batas kanan ke batas kiri dimulai dari ujung 5’ ke ujung 3’ (5’–3’) mirip dengan
proses konjugasi. Satu atau beberapa kopi ssT-DNA dapat terintegrasi ke
dalam genom tanaman pada satu situs yang sama atau terpisah-pisah pada
situs yang berbeda (Armitage et al. 1987). Masuknya suatu gen asing dari suatu organisme ke sel lain (tanaman) mengalami beberapa tahap yaitu pengikatan
dan pengambilan bahan genetik pada tanaman dan penurunan sifat genetik
pada tanaman yang diregenerasikan.
Tanaman yang telah mengalami integrasi T-DNA selanjutnya
produksi fitohormon. Fitohormon tersebut diperlukan tanaman untuk
meningkatkan sintesis protein dan memacu pertumbuhan akar primordia yang
meningkatkan pertumbuhan akar. Gen-gen yang terdapat pada T-DNA
dibedakan menurut peranannya yaitu gen-gen yang mempengaruhi
keseimbangan fitohormon dalam sel tanaman inang (oncogene) atau gen yang
mempengaruhi produksi opin (opin sintetase). Pada TR-DNA gen yang berfungsi
untuk pembentuk auksin adalah iaaM (tms 1), dan iaaH (tms 2), dan juga
membawa gen untuk menyandi sintesis senyawa opin. Gen iaaM menyandi
pembentukan triptofan 2-monooksigenase yang mengkatalisis perubahan
L-triptofan menjadi senyawa indole-3-asetamida. Sedangkan iaaH menyandi
enzim amino hidrolase yang mengubah indole-3-asetamida menjadi asam
indole-3-asetat (IAA) (van der Salm et al. 1996; Jeng-Sheng 2001).
Daerah TL-DNA berukuran 20 kb, mengandung empat root loci (rol
A-D) yang terlibat dalam induksi akar, yakni rolA, rolB, rolC dan rolD yang berhubungan dengan open reading frames ORFs 10, 11, 12, dan 15 (Chriqui
1996). Jeng-Sheng (2001) menjelaskan bahwa gen rolB (ORF11) mengkodeβ
-glucosidase yang menghidrolisis indol β-glucosida yang diduga berperan meningkatkan pool auksin aktif dalam tanaman dengan hidrolisis konjugat IAA inaktif, mengatur sensivitas sel terhadap IAA dan mendorong pembentukan
meristem. Gen rolC (ORF12) diduga berperan meningkatkan level sitokinin melalui aktivitas β-glucosidase yang mampu melepaskan sitokinin aktif dari konjugatnya. Gen rolA (ORF10) terlibat lansung dalam induksi akar sedangkan
rolD (ORF15) berperan menginduksi pembungaan, namun fungsi biokimia dari kedua gen rol ini belum diketahui (Jeng-Sheng 2001; Valpuesta 2002). Rugini et al. (1992) telah membuktikan bahwa kemampuan perakaran, jumlah akar dan massa akar meningkat ketika gen rolABC terekspresi pada tanaman kiwi.
2.3.4. Perbaikan Sistem Perakaran dengan Agrobacterium rhizogenes
Perakaran masih menjadi masalah utama yang sulit dipecahkan pada
tanaman tahunan berkayu yang diperbanyak dengan metode perkecambahan
biji maupun kultur in vitro. Percobaan untuk memperbaiki sistem perakaran selama ini lebih banyak dilakukan dengan penambahan zat pengatur tumbuh
dan bahan kimia lainnya. Hasil yang diperoleh memperlihatkan bahwa awal