TOLERANSI HOTONG (
Setaria italica
L. Beauv)
PADA BERBAGAI CEKAMAN KEKERINGAN:
PENDEKATAN ANATOMI DAN FISIOLOGI
YATI TUASAMU
SEKOLAH PASCA SARJANA
INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI
Dengan ini saya menyatakan bahwa tesis Toleransi Hotong (Setaria italica L. Beauv) pada berbagai Cekaman Kekeringan: Pendekatan Anatomi dan Fisiologi adalah karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apapun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Bogor, Agustus 2009
ABSTRACT
YATI TUASAMU. (Tolerance Hotong (Setaria italica L. Beauv) to Drought Stress: Anatomy and Physiology responses). Under supervisor of HAMIM and TRIADIATI.
Hotong (Setaria italica L. Beauv) is one of local excellent crop in Buru Island which becomes an important food alternative. However, because this plants are normally cultivated during dry season, drought stress is one of the major factor limiting plant growth and production. The objective of the study was to examine the adverse effects of drought stress on 3 accessions of hotong (Buru 1 accession (A1), Buru 2 accession (A2), and AGH accession (A3) based on anatomical and physiological responses. Plants were grown in 6 kg pot containing soil and sand (1:1, v/v) in the greenhouse. Drought was imposed at 14 days after the plants were 6 weeks old. The parameters were examined during the drought stress were media water content (MWC), relative water content (RWC), plant growth, photosynthetic parameters including maximum efficiency of photosynthetic (Fv/Fm), photochemical quenching (qP), non-photochemical quenching (qN), and quantum yield (qY), chlorophyll content, ascorbic acid (ASA), and proline concentration. Anatomy characteristics of leaf, stem, and root were observed using paradermal and transversal section. The results showed that drought stress caused decrease of MWC and RWC which generally influenced leaf area, plant height, root length, and dry weight. Seed productiondecreased due to the stress, andthe biggest was happened on A2 compared to A1 and A3 accessions. The Fv/Fm of all plants was not change during the drought periode, but showed a slight decrease in the last periode of drought. The A1 and A2 presented the higher decrease in qP and qY in response to drought, as compared to A3. On the contrary, qN increased in all accessions during the drought periode, but the increment was higher in the A3. The chlorophyll content of the plants subjected to drought stress was relatively stable and equal to that of plant control. The ASA content of A1 and A2 increased until 8 days drought treatment and decreased after 12 days drought treatment and rewatering. Drought stress increased significant accumulation of proline in all accessions until 14 days and have positive correlation to improvement of root length especially of A3. Leaf antomical characteristics of the plants subjected to drought stress decreased significantly in primary vena, vein rib, leaf, and mesophyll thickness, but the lower reduction was showed by A3 compared to other accessions especially in leaf and mesophyll thickness. The A1 showed the higher decrease in xylem vessels diameter of leaf, as compared to A2 and A3. The xylem vessels diameter of stem and root of all plants were relatively stable and equal to the plant control.Stomata density decreased significant in A2, but in A1 and A3 it was stable. The results showed that A3 more tolerant than A1 and A2, indicated by stable of photosynthetic rate (Pn) and seed production during the drought.
RINGKASAN
YATI TUASAMU. Toleransi Hotong(Setaria italica L. Beauv) pada berbagai Cekaman Kekeringan: Pendekatan Anatomi dan Fisiologi. Dibimbing oleh HAMIM dan TRIADIATI.
Hotong (Setaria italica L. Beauv) merupakan salah satu komoditi tanaman pangan unggul lokal yang dikonsumsi masyarakat di Pulau Buru sebagai pangan alternatif penghasil karbohidrat. Tanaman ini tumbuh pada lahan kering dan toleran terhadap suhu tinggi namun tidak toleran pada periode cekaman kekeringan yang panjang. Budidaya tanaman hotong terutama di prioritaskan pada lahan-lahan kering dengan pola penanaman tadah hujan, sehingga kondisi kekeringan merupakan salah satu faktor lingkungan utama yang dapat menghambat pertumbuhan dan produksinya.
Ketersediaan air yang rendah pada fase awal pertumbuhan tanaman ini dapat mengakibatkan cekaman kekeringan (drought stress). Pada periode cekaman kekeringan yang panjang tanaman ini tidak mampu melakukan recovery, dan apabila terjadi kerusakan jaringan akan bersifat tidak dapat balik (permanen) sehingga mempengaruhi pertumbuhan dan menurunkan produksinya.
Antisipasi yang dapat dilakukan untuk mengurangi tingkat penurunan produksi akibat cekaman kekeringan adalah dengan menanam varietas unggul hotong yang toleran kekeringan. Dalam upaya pengembangan varietas unggul tersebut diperlukan sejumlah informasi mengenai mekanisme toleransi tanaman ini terhadap cekaman kekeringan, sehingga proses seleksi bisa berjalan secara efisien dan efektif.
Penelitian ini mengamati mekanisme toleransi terhadap cekaman kekeringan dengan pendekatan anatomi dan fisiologi pada tiga aksesi hotong (aksesi Buru 1 (A1), aksesi Buru 2 (A2), dan aksesi AGH (A3). Tanaman hotong ditumbuhkan dalam pot berukuran 6 kg menggunakan media tanam yang terdiri atas campuran tanah dan pasir dengan perbandingan (1:1, v/v) di dalam rumah kaca. Perlakuan cekaman kekeringan dilakukan selama 14 hari setelah tanaman berumur 6 minggu. Parameter-parameter yang diukur selama cekaman kekeringan meliputi: Kadar Air Media (KAM), Kandungan Air Relatif (KAR) daun, pertumbuhan dan produksi tanaman, parameter fotosintetis yang terdiri atas: efisiensi maksimum fotosintetis (Fv/Fm), pelepasan energi untuk reaksi fotokimia (qP), non fotokimia (qN), dan hasil kuatum fotosintesis (qY), kandungan asam askorbat (ASA), prolin, dan klorofil daun. Karakteristik anatomi daun, batang, dan akar diamati melalui pembuatan sayatan paradermal dan transversal menggunakan mikroskop cahaya.
peningkatan akumulasi asam askorbat (ASA) yang puncaknya pada 8 HSP kekeringan dan signifikan pada aksesi 3. Selain itu perlakuan cekaman kekeringan mengakibatkan peningkatan akumulasi prolin yang tertinggi pada aksesi 1 dan 2 pada akhir periode cekaman. Peningkatan akumulasi prolin selama periode cekaman berkorelasi positif terhadap peningkatan panjang akar terutama pada aksesi 3 yang merupakan mekanisme toleransi terhadap cekaman kekeringan. Peningkatan akumulasi prolin pada akhir periode kekeringan pada ketiga aksesi menunjukkan bahwa tanaman dalam keadaan stres yang sangat berat.
Dengan demikian, dapat disimpulkan bahwa aksesi 3 lebih toleran terhadap perlakuan cekaman kekeringan selama 14 hari dibanding aksesi 1 dan 2. Hal ini dapat dilihat berdasarkan kestabilan proses fotosintesis selama cekaman kekeringan yang ditunjukkan oleh nilai reaksi-reaksi fotokimia (qP) dan hasil kuantum fotosintesis (qY), mekanisme regulasi fotosintesis melalui proses non fotokimia (qN), kestabilan struktur anatomi daun terutama kerapatan stomata, tebal daun, dan lapisan mesofil. Terjadi peningkatan panjang akar dan produksi biji yang relatif tidak mengalami penurunan dibandingkan dengan tanaman kontrolnya.
©
Hak Cipta milik IPB, tahun 2009
Hak Cipta dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumber. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan yang wajar IPB
TOLERANSI HOTONG (
Setaria italica
L. Beauv)
PADA BERBAGAI CEKAMAN KEKERINGAN:
PENDEKATAN ANATOMI DAN FISIOLOGI
YATI TUASAMU
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains pada
Mayor Biologi Tumbuhan
SEKOLAH PASCA SARJANA
INSTITUT PERTANIAN BOGOR
Judul Tesis : Toleransi Hotong (Setaria italica L. Beauv) pada berbagai Cekaman Kekeringan: Pendekatan Anatomi dan Fisiologi
Nama : Yati Tuasamu
NRP : G353070011
Disetujui
Komisi Pembimbing
Dr. Ir. Hamim, M.Si. Dr. Dra. Triadiati, M.Si.
Ketua Anggota
Diketahui
Tanggal Ujian: 06 Agustus 2009 Tanggal Lulus: 25 Agustus 2009 Koordinator Mayor
Biologi Tumbuhan
Dekan Sekolah Pascasarjana
Dr. Ir. Miftahudin, M.Si.
PRAKATA
Alhamdulillah, puji dan syukur penulis panjatkan ke hadirat Allah SWT atas segala karunia-Nya sehingga karya ilmiah (tesis) ini berhasil diselesaikan. Penelitian yang dilaksanakan sejak bulan Juni 2008 sampai April 2009 di rumah kaca Kebun Percobaan Cikabayan, Laboratorium Fisiologi, Laboratorium Anatomi dan Morfologi Tumbuhan, Departemen Biologi, FMIPA, Institut Pertanian Bogor ini tentang cekaman kekeringan, dengan judul Toleransi Hotong (Setaria italica L. Beauv) pada berbagai Cekaman Kekeringan: Pendekatan Anatomi dan Fisiologi.
Terima kasih dan penghargaan yang tinggi penulis sampaikan kepada semua pihak yang telah membantu dalam pelaksanaan penelitian dan penyelesaian penulisan tesis ini yaitu;
1 Bapak Dr. Ir. Hamim, M.Si. dan Ibu Dr. Dra. Triadiati, M.Si. selaku komisi pembimbing atas waktu, kesabaran, ilmu, dan kemudahan yang diberikan selama bimbingan mulai dari tahap awal persiapan penelitian sampai akhir penyelesaian tesis ini.
2 Dr. Ir. Juliarni, M.Agr. atas waktu, kesabaran, ilmu, dan kemudahan yang diberikan selama bimbingan pada tahap awal persiapan penelitian ini.
3 Dr. Ir. Munif Ghulamahdi, M.S. selaku penguji luar komisi atas ilmu dan saran yang telah diberikan untuk kesempurnaan tesis ini.
4 Ketua Mayor, seluruh dosen, karyawan, dan pengelola Laboratorium pada lingkup Biologi Tumbuhan atas ilmu, bantuan, dan fasilitas yang diberikan selama penelitian dilakukan sampai penyelesaian tesis ini.
5 IPB yang telah memberikan kesempatan bagi penulis untuk memperoleh dana BPPS dalam rangka penyelesaian studi dan penelitian pada Program Magister Sains. 6 Yayasan Dana Beasiswa Maluku (YDBM) dan Yayasan Tahija di Jakarta yang telah
memberikan bantuan dana dalam pelaksanaan penelitian ini.
7 Papa dan mama tercinta, Ed dan adik-adikku tersayang serta seluruh keluarga atas nasehat, kepercayaan, doa, dan semangat yang telah diberikan sehingga penulis dapat menyelesaikan penelitian dan penulisan tesis ini tepat waktu.
8 Rekan-rekan Pascasajana Biologi FMIPA IPB tahun 2007 atas kerjasama dan kebersamaannya selama menyelesaikan studi.
Semoga Allah senantiasa membalas kebaikan semuanya dengan pahala yang berlipat ganda, amin. Penulis berharap semoga karya ilmiah ini bermanfaat bagi pihak yang terkait terutama dalam upaya pemuliaan dan pengembangan budidaya tanaman hotong khususnya di Pulau Buru maupun di Indonesia.
Bogor, Agustus 2009
TOLERANSI HOTONG (
Setaria italica
L. Beauv)
PADA BERBAGAI CEKAMAN KEKERINGAN:
PENDEKATAN ANATOMI DAN FISIOLOGI
YATI TUASAMU
SEKOLAH PASCA SARJANA
INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI
Dengan ini saya menyatakan bahwa tesis Toleransi Hotong (Setaria italica L. Beauv) pada berbagai Cekaman Kekeringan: Pendekatan Anatomi dan Fisiologi adalah karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apapun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Bogor, Agustus 2009
ABSTRACT
YATI TUASAMU. (Tolerance Hotong (Setaria italica L. Beauv) to Drought Stress: Anatomy and Physiology responses). Under supervisor of HAMIM and TRIADIATI.
Hotong (Setaria italica L. Beauv) is one of local excellent crop in Buru Island which becomes an important food alternative. However, because this plants are normally cultivated during dry season, drought stress is one of the major factor limiting plant growth and production. The objective of the study was to examine the adverse effects of drought stress on 3 accessions of hotong (Buru 1 accession (A1), Buru 2 accession (A2), and AGH accession (A3) based on anatomical and physiological responses. Plants were grown in 6 kg pot containing soil and sand (1:1, v/v) in the greenhouse. Drought was imposed at 14 days after the plants were 6 weeks old. The parameters were examined during the drought stress were media water content (MWC), relative water content (RWC), plant growth, photosynthetic parameters including maximum efficiency of photosynthetic (Fv/Fm), photochemical quenching (qP), non-photochemical quenching (qN), and quantum yield (qY), chlorophyll content, ascorbic acid (ASA), and proline concentration. Anatomy characteristics of leaf, stem, and root were observed using paradermal and transversal section. The results showed that drought stress caused decrease of MWC and RWC which generally influenced leaf area, plant height, root length, and dry weight. Seed productiondecreased due to the stress, andthe biggest was happened on A2 compared to A1 and A3 accessions. The Fv/Fm of all plants was not change during the drought periode, but showed a slight decrease in the last periode of drought. The A1 and A2 presented the higher decrease in qP and qY in response to drought, as compared to A3. On the contrary, qN increased in all accessions during the drought periode, but the increment was higher in the A3. The chlorophyll content of the plants subjected to drought stress was relatively stable and equal to that of plant control. The ASA content of A1 and A2 increased until 8 days drought treatment and decreased after 12 days drought treatment and rewatering. Drought stress increased significant accumulation of proline in all accessions until 14 days and have positive correlation to improvement of root length especially of A3. Leaf antomical characteristics of the plants subjected to drought stress decreased significantly in primary vena, vein rib, leaf, and mesophyll thickness, but the lower reduction was showed by A3 compared to other accessions especially in leaf and mesophyll thickness. The A1 showed the higher decrease in xylem vessels diameter of leaf, as compared to A2 and A3. The xylem vessels diameter of stem and root of all plants were relatively stable and equal to the plant control.Stomata density decreased significant in A2, but in A1 and A3 it was stable. The results showed that A3 more tolerant than A1 and A2, indicated by stable of photosynthetic rate (Pn) and seed production during the drought.
RINGKASAN
YATI TUASAMU. Toleransi Hotong(Setaria italica L. Beauv) pada berbagai Cekaman Kekeringan: Pendekatan Anatomi dan Fisiologi. Dibimbing oleh HAMIM dan TRIADIATI.
Hotong (Setaria italica L. Beauv) merupakan salah satu komoditi tanaman pangan unggul lokal yang dikonsumsi masyarakat di Pulau Buru sebagai pangan alternatif penghasil karbohidrat. Tanaman ini tumbuh pada lahan kering dan toleran terhadap suhu tinggi namun tidak toleran pada periode cekaman kekeringan yang panjang. Budidaya tanaman hotong terutama di prioritaskan pada lahan-lahan kering dengan pola penanaman tadah hujan, sehingga kondisi kekeringan merupakan salah satu faktor lingkungan utama yang dapat menghambat pertumbuhan dan produksinya.
Ketersediaan air yang rendah pada fase awal pertumbuhan tanaman ini dapat mengakibatkan cekaman kekeringan (drought stress). Pada periode cekaman kekeringan yang panjang tanaman ini tidak mampu melakukan recovery, dan apabila terjadi kerusakan jaringan akan bersifat tidak dapat balik (permanen) sehingga mempengaruhi pertumbuhan dan menurunkan produksinya.
Antisipasi yang dapat dilakukan untuk mengurangi tingkat penurunan produksi akibat cekaman kekeringan adalah dengan menanam varietas unggul hotong yang toleran kekeringan. Dalam upaya pengembangan varietas unggul tersebut diperlukan sejumlah informasi mengenai mekanisme toleransi tanaman ini terhadap cekaman kekeringan, sehingga proses seleksi bisa berjalan secara efisien dan efektif.
Penelitian ini mengamati mekanisme toleransi terhadap cekaman kekeringan dengan pendekatan anatomi dan fisiologi pada tiga aksesi hotong (aksesi Buru 1 (A1), aksesi Buru 2 (A2), dan aksesi AGH (A3). Tanaman hotong ditumbuhkan dalam pot berukuran 6 kg menggunakan media tanam yang terdiri atas campuran tanah dan pasir dengan perbandingan (1:1, v/v) di dalam rumah kaca. Perlakuan cekaman kekeringan dilakukan selama 14 hari setelah tanaman berumur 6 minggu. Parameter-parameter yang diukur selama cekaman kekeringan meliputi: Kadar Air Media (KAM), Kandungan Air Relatif (KAR) daun, pertumbuhan dan produksi tanaman, parameter fotosintetis yang terdiri atas: efisiensi maksimum fotosintetis (Fv/Fm), pelepasan energi untuk reaksi fotokimia (qP), non fotokimia (qN), dan hasil kuatum fotosintesis (qY), kandungan asam askorbat (ASA), prolin, dan klorofil daun. Karakteristik anatomi daun, batang, dan akar diamati melalui pembuatan sayatan paradermal dan transversal menggunakan mikroskop cahaya.
peningkatan akumulasi asam askorbat (ASA) yang puncaknya pada 8 HSP kekeringan dan signifikan pada aksesi 3. Selain itu perlakuan cekaman kekeringan mengakibatkan peningkatan akumulasi prolin yang tertinggi pada aksesi 1 dan 2 pada akhir periode cekaman. Peningkatan akumulasi prolin selama periode cekaman berkorelasi positif terhadap peningkatan panjang akar terutama pada aksesi 3 yang merupakan mekanisme toleransi terhadap cekaman kekeringan. Peningkatan akumulasi prolin pada akhir periode kekeringan pada ketiga aksesi menunjukkan bahwa tanaman dalam keadaan stres yang sangat berat.
Dengan demikian, dapat disimpulkan bahwa aksesi 3 lebih toleran terhadap perlakuan cekaman kekeringan selama 14 hari dibanding aksesi 1 dan 2. Hal ini dapat dilihat berdasarkan kestabilan proses fotosintesis selama cekaman kekeringan yang ditunjukkan oleh nilai reaksi-reaksi fotokimia (qP) dan hasil kuantum fotosintesis (qY), mekanisme regulasi fotosintesis melalui proses non fotokimia (qN), kestabilan struktur anatomi daun terutama kerapatan stomata, tebal daun, dan lapisan mesofil. Terjadi peningkatan panjang akar dan produksi biji yang relatif tidak mengalami penurunan dibandingkan dengan tanaman kontrolnya.
©
Hak Cipta milik IPB, tahun 2009
Hak Cipta dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumber. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan yang wajar IPB
TOLERANSI HOTONG (
Setaria italica
L. Beauv)
PADA BERBAGAI CEKAMAN KEKERINGAN:
PENDEKATAN ANATOMI DAN FISIOLOGI
YATI TUASAMU
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains pada
Mayor Biologi Tumbuhan
SEKOLAH PASCA SARJANA
INSTITUT PERTANIAN BOGOR
Judul Tesis : Toleransi Hotong (Setaria italica L. Beauv) pada berbagai Cekaman Kekeringan: Pendekatan Anatomi dan Fisiologi
Nama : Yati Tuasamu
NRP : G353070011
Disetujui
Komisi Pembimbing
Dr. Ir. Hamim, M.Si. Dr. Dra. Triadiati, M.Si.
Ketua Anggota
Diketahui
Tanggal Ujian: 06 Agustus 2009 Tanggal Lulus: 25 Agustus 2009 Koordinator Mayor
Biologi Tumbuhan
Dekan Sekolah Pascasarjana
Dr. Ir. Miftahudin, M.Si.
PRAKATA
Alhamdulillah, puji dan syukur penulis panjatkan ke hadirat Allah SWT atas segala karunia-Nya sehingga karya ilmiah (tesis) ini berhasil diselesaikan. Penelitian yang dilaksanakan sejak bulan Juni 2008 sampai April 2009 di rumah kaca Kebun Percobaan Cikabayan, Laboratorium Fisiologi, Laboratorium Anatomi dan Morfologi Tumbuhan, Departemen Biologi, FMIPA, Institut Pertanian Bogor ini tentang cekaman kekeringan, dengan judul Toleransi Hotong (Setaria italica L. Beauv) pada berbagai Cekaman Kekeringan: Pendekatan Anatomi dan Fisiologi.
Terima kasih dan penghargaan yang tinggi penulis sampaikan kepada semua pihak yang telah membantu dalam pelaksanaan penelitian dan penyelesaian penulisan tesis ini yaitu;
1 Bapak Dr. Ir. Hamim, M.Si. dan Ibu Dr. Dra. Triadiati, M.Si. selaku komisi pembimbing atas waktu, kesabaran, ilmu, dan kemudahan yang diberikan selama bimbingan mulai dari tahap awal persiapan penelitian sampai akhir penyelesaian tesis ini.
2 Dr. Ir. Juliarni, M.Agr. atas waktu, kesabaran, ilmu, dan kemudahan yang diberikan selama bimbingan pada tahap awal persiapan penelitian ini.
3 Dr. Ir. Munif Ghulamahdi, M.S. selaku penguji luar komisi atas ilmu dan saran yang telah diberikan untuk kesempurnaan tesis ini.
4 Ketua Mayor, seluruh dosen, karyawan, dan pengelola Laboratorium pada lingkup Biologi Tumbuhan atas ilmu, bantuan, dan fasilitas yang diberikan selama penelitian dilakukan sampai penyelesaian tesis ini.
5 IPB yang telah memberikan kesempatan bagi penulis untuk memperoleh dana BPPS dalam rangka penyelesaian studi dan penelitian pada Program Magister Sains. 6 Yayasan Dana Beasiswa Maluku (YDBM) dan Yayasan Tahija di Jakarta yang telah
memberikan bantuan dana dalam pelaksanaan penelitian ini.
7 Papa dan mama tercinta, Ed dan adik-adikku tersayang serta seluruh keluarga atas nasehat, kepercayaan, doa, dan semangat yang telah diberikan sehingga penulis dapat menyelesaikan penelitian dan penulisan tesis ini tepat waktu.
8 Rekan-rekan Pascasajana Biologi FMIPA IPB tahun 2007 atas kerjasama dan kebersamaannya selama menyelesaikan studi.
Semoga Allah senantiasa membalas kebaikan semuanya dengan pahala yang berlipat ganda, amin. Penulis berharap semoga karya ilmiah ini bermanfaat bagi pihak yang terkait terutama dalam upaya pemuliaan dan pengembangan budidaya tanaman hotong khususnya di Pulau Buru maupun di Indonesia.
Bogor, Agustus 2009
RIWAYAT HIDUP
Penulis dilahirkan di Ambon pada tanggal 09 Juli 1976 sebagai anak sulung dari tujuh bersaudara dari ayah Kadir Tuasamu dan ibu Hadija Maruapey.
Tahun 1994 penulis lulus SMA Negeri 7 Ambon dan pada tahun yang sama penulis diterima sebagai mahasiswa pada Jurusan Peternakan Universitas Pattimura Ambon melalui jalur PSSB. Penulis menyelesaikan studi S1 pada tahun 2001 dengan beasiswa Peningkatan Prestasi Akademik (PPA) dari Yayasan Supersemar.
Penulis menikah pada tahun 2000 dengan Serka. A. Tuharea dan telah dikaruniai dua orang putra dan putri yaitu: Jafar Umar Thalib Tuharea (10 tahun) dan Humaira Sukma Ayu Tuharea (6 tahun).
Tahun 2005 penulis diangkat sebagai tenaga edukatif pada lingkup Kopertis Wilayah XII Ambon, dpk di Universitas Iqra Buru, Kabupaten Buru, Provinsi Maluku pada Jurusan Budidaya Pertanian, Fakultas Pertanian sampai sekarang. Penulis juga mengabdi sebagai guru bantu Biologi dan Muatan Lokal Peternakan pada Madrasah Aliyah Yayasan Daarul Arqam, Kabupaten Buru, Provinsi Maluku sejak tahun 2006 sampai sekarang.
DAFTAR ISI
Halaman DAFTAR TABEL ………... xiii DAFTAR GAMBAR ………... xiv DAFTAR LAMPIRAN ………... xv PENDAHULUAN
Latar Belakang ... 1 Tujuan Penelitian ... 5 Manfaat Penelitian ... 5 Hipotesis ... 5 TINJAUAN PUSTAKA
Gambaran Umum Tanaman Hotong ... 6 Fungsi Air bagi Tanaman ... 6 Cekaman Kekeringan bagi Tanaman ... 7 Pengaruh Cekaman Kekeringan terhadap Fotosintesis Tanaman ... 9 Akumulasi Asam Askorbat (ASA) dan Prolin sebagai Respon
Terhadap Kekeringan ... 10 BAHAN DAN METODE
Waktu dan Tempat Penelitian ... 14 Bahan dan Alat ... 14 Rancangan Percobaan ... 15 Pelaksanaan Penelitian ... 15 Persiapan bahan tanaman dan media tanam ... 15 Penanaman dan pemeliharaan ... 16 Pemberian perlakuan cekaman kekeringan ... 16 Pengamatan ... 16 Pengukuran parameter fotosintesis ... 18 Analisis asam askorbat (ASA) ... 18 Analisis prolin ... 19 Analisis kandungan klorofil daun ... 19 Pembuatan sayatan paradermal ... 20 Pembuatan sayatan transversal ... 21 HASIL DAN PEMBAHASAN
Kadar Air Media (KAM) dan Kandungan Air Relatif (KAR) Daun ... 22 Respon Umum Tanaman ... 25 Parameter Anatomi Daun, Batang, dan Akar ... 30 Kualitatif Anatomi Daun, Batang, dan Akar ... 34 Parameter Fotosintesis ... 36 Akumulasi Asam Askorbat (ASA), Prolin, dan Kandungan
KESIMPULAN DAN SARAN
DAFTAR TABEL
Halaman
1 Pengaruh cekaman kekeringan terhadap luas daun, tinggi tajuk, dan panjang akar ketiga aksesi hotong pada 14 hari setelah perlakuan
cekaman kekeringan ... 26 2 Pengaruh cekaman kekeringan terhadap bobot kering tajuk dan akar
dan bobot kering biji ketiga aksesi hotong ... 27 3 Pengaruh cekaman kekeringan terhadap kerapatan dan indeks stomata
pada lapisan epidermis daun bagian atas dan bawah pada 14 hari setelah
perlakuan cekaman kekeringan ... 31 4 Pengaruh cekaman kekeringan terhadap struktur anatomi daun, batang,
dan akar ketiga aksesi hotong pada 14 hari setelah perlakuan cekaman
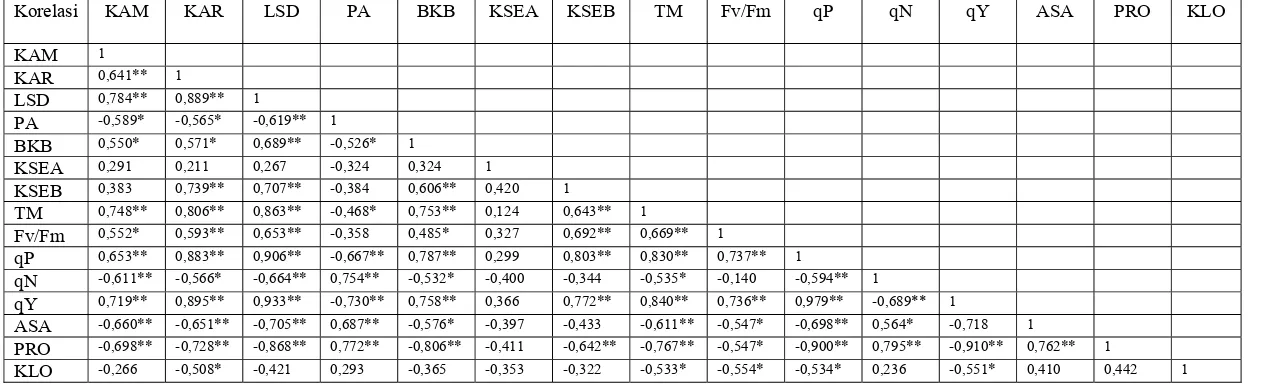
kekeringan ... 32 5 Korelasi antar parameter ketiga aksesi hotong pada 14 hari setelah
DAFTAR GAMBAR
Halaman
1 Mekanisme pembentukan senyawa reactive oxygen species (ROS) ... 11 2 Mekanisme penyelamatan terhadap cekaman oksidatif oleh asam
askorbat (ASA) ... 12 3 Kadar Air Media (KAM) (%) dan Kandungan Air Relatif (KAR)
daun (%) pada 0-14 hari setelah perlakuan cekaman kekeringan dan
recovery pada taraf uji 5% (DMRT) ... 22 4 Sayatan transversal daun, batang, dan akar dan sayatan paradermal daun
ketiga aksesi hotong kontrol dan perlakuan pada 14 hari setelah perlakuan
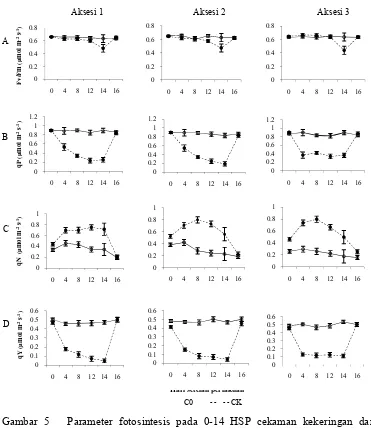
cekaman kekeringan ... 35 5 Parameter fotosintesis pada 0-14 hari setelah perlakuan cekaman
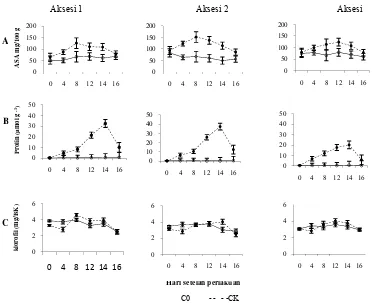
Kekeringan dan recovery pada taraf uji 5% (DMRT) ... 38 6 Akumulasi asam askorbat (ASA), prolin, dan kandungan klorofil daun
pada 0-14 hari setelah perlakuan cekaman kekeringan dan recovery
pada taraf uji 5% (DMRT) ... 42 7 Morfologi aksesi Buru 1 dan Buru 2 ... 55 8 Morfologi ketiga aksesi hotong kontrol dan perlakuan pada 8 hari
setelah perlakuan cekaman kekeringan ... 56 9 Morfologi ketiga aksesi hotong kontrol dan perlakuan pada 14 hari
setelah perlakuan cekaman kekeringan ... 56 10 Morfologi akar ketiga aksesi hotong kontrol dan perlakuan pada 14 hari
DAFTAR LAMPIRAN
Halaman 1 Korelasi antar parameter ketiga aksesi hotong pada 14 hari setelah
perlakuan cekaman kekeringan ... 53 2 Grafik korelasi parameter penentu sifat toleransi ketiga aksesi hotong
pada 8 dan 14 hari setelah perlakuan cekaman kekeringan ... 54 3 Letak geografis, morfologi, dan deskripsi aksesi Buru 1 dan Buru 2 ... 55 4 Morfologi ketiga aksesi hotong pada 8 dan 14 hari setelah perlakuan
cekaman kekeringan ... 56 5 Morfologi akar ketiga aksesi hotong pada 14 hari setelah perlakuan
PENDAHULUAN
Latar Belakang
Dalam rangka mendukung program diversifikasi pangan Nasional yang sedang digalakkan saat ini, maka pengembangan tanaman pangan alternatif sumber karbohidrat bagi masyarakat perlu dilakukan. Usaha pengembangan tanaman pangan alternatif tersebut harus tetap memperhatikan berbagai keunggulan komparatif wilayah yang sasaran utamanya adalah pengembangan dan peningkatan produksi komoditas unggul daerah (unggul lokal).
Hotong atau yang sering dikenal sebagai ”foxtailmillet” (Setaria italica L. Beauv) termasuk salah satu jenis tanaman pangan unggul lokal yang dikonsumsi oleh masyarakat Pulau Buru sebagai pangan alternatif penghasil karbohidrat dan masih dianggap langka. Biji hotong selain dikonsumsi untuk memproduksi aneka jenis makanan olahan seperti bubur, nasi, wajik dan aneka penganan lain, juga memiliki nilai ekonomis penting antara lain sebagai bahan baku pembuatan mi instan, minuman berenergi, dan makanan balita. Kandungan nutrisi biji hotong cukup tinggi yaitu mengandung protein sebesar 11,18%, lemak 2,36%, karbohidrat 73,36%, air sebesar 11,78%, dan abu sebesar 1,32%. Energi yang dihasilkan dalam 100 g biji adalah 359 kalori. Dilihat dari kandungan gizinya yang cukup tinggi, maka tanaman hotong dapat dijadikan komoditi alternatif dalam program diversifikasi pangan alternatif penghasil karbohidrat (Andrawulan 2003). Untuk itu perlu dibudidayakan secara luas serta di jaga kelestariannya sebagai komoditi unggul lokal.
data dan informasi hasil penelitian tentang pemuliaan dan pengembangan budidaya tanaman hotong masih sangat sedikit (Diptan 2003).
Pulau Buru termasuk daerah rawan kekeringan dengan jumlah curah hujan tahunan relatif rendah yaitu berkisar antara 1000 - 1400 mm. Curah hujan rata-rata pada bulan lembab (Januari–Maret) 262 mm/bln, sedangkan bulan kering (April– Desember) kurang dari 100 mm/bln (Diptan 2003). Curah hujan rata-rata bagi pertumbuhan tanaman hotong berkisar antara 200 - 300 mm/bln atau lebih sedikit dari curah hujan musiman (Dekany 1999). Rendahnya curah hujan di daerah tersebut dapat menyebabkan terjadinya defisit air bagi tanaman hotong sehingga mempengaruhi pertumbuhan dan produksinya.
Ketersediaan air yang rendah pada masa awal pertumbuhan tanaman
dapat mengakibatkan cekaman kekeringan (drought stress). Kekeringan
merupakan faktor lingkungan utama yang dapat menghambat pertumbuhan tanaman dan menurunkan produksi bergantung pada besarnya tingkat cekaman yang dialami dan fase pertumbuhan tanaman ketika mendapat cekaman kekeringan (Levitt 1980). Pada periode cekaman kekeringan yang panjang akan mempengaruhi seluruh proses metabolismeme di dalam sel dan mengakibatkan penurunan produksi tanaman (Bohnert et al. 1995).
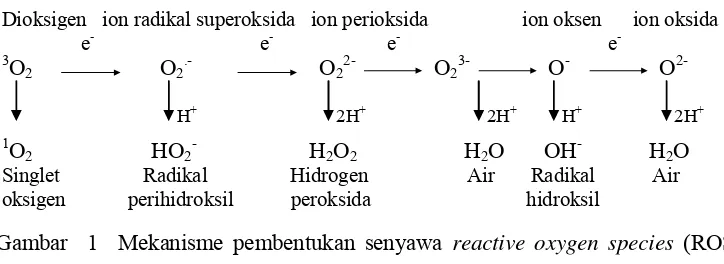
Hamim (2004) menyatakan bahwa pada tahap awal, kekeringan menyebabkan berkurangnya pembukaan stomata untuk meminimalisir kehilangan air di bawah kondisi cahaya berlebihan. Peristiwa ini mengakibatkan terjadinya penurunan konsentrasi CO2 intrasel, sehingga tanaman mengalami overreduksi pada transfer elektron fotosintesis (Zlatev & Yordanov 2004). Hal ini dapat mengakibatkan terbentuknya reactive oxygen species (ROS) yang diawali dengan pengikatan elektron pada transfer elektron fotosintesis oleh oksigen (Asada 2006). Senyawa ROS ini dapat menyebabkan cekaman oksidatif bagi tanaman yang dapat menimbulkan kerusakan jaringan dan ultrastruktur sel (Wise & Naylor 1987).
dilaporkan meningkat pada beberapa spesies tanaman saat mengalami cekaman kekeringan. Selain itu tanaman juga mengakumulasi senyawa prolin yang berfungsi untuk pengaturan derajat osmotik sel (osmotic adjustment). Akumulasi prolin dapat menurunkan potensial osmotik sehingga menurunkan potensial air dalam sel tanpa membatasi fungsi enzim dan menjaga turgor sel (Hamim 1995). Sintesis senyawa-senyawa ini mengindikasi toleransi tanaman terhadap cekaman kekeringan. Cekaman kekeringan juga dilaporkan dapat memacu tanaman untuk beradaptasi secara morfologi dan anatomi (Radwan 2007).
Hotong umumnya tumbuh pada daerah kering dan toleran terhadap suhu tinggi seperti jagung dan sorgum, akan tetapi tidak toleran terhadap periode cekaman kekeringan yang panjang sebaik sorgum (Leder & Monda 2004). Tanaman hotong yang mengalami cekaman kekeringan panjang pada fase pertumbuhannya sukar mengalami recovery setelah dilakukan pengairan kembali (rewatering) (Karyudi & Fletcher 2003). Antisipasi yang dapat dilakukan untuk mengurangi tingkat penurunan produksi akibat cekaman kekeringan adalah dengan menanam varietas unggul tanaman hotong yang toleran kekeringan. Dalam upaya pengembangan varietas unggul tersebut diperlukan sejumlah informasi mengenai mekanisme toleransi tanaman tersebut terhadap cekaman kekeringan, sehingga proses seleksi bisa berjalan secara efisien dan efektif.
Karyudi dan Fletcher (1999) telah melakukan penelitian tentang kapasitas osmoregulasi 11 aksesi S. italica dan 14 aksesi Panicum muliaceum terhadap cekaman kekeringan. Berdasarkan penelitian tersebut dilaporkan bahwa: (1) S. italica mampu tumbuh pada kondisi cekaman kekeringan, tetapi tidak toleran pada periode kekeringan yang panjang dibanding P. muliaceum, (2) tanaman ini tidak
mampu melakukan recovery setelah mengalami cekaman kekeringan berat
solut di dalam jaringan daun, (5) terjadi peningkatan prolin dan kalsium tetapi tidak berkontribusi terhadap akumulasi solut dan perkembangan luas daun.
Kemampuan penetrasi akar untuk mencapai zona yang lebih dalam pada beberapa spesies millet merupakan suatu mekanisme penghindaran terhadap cekaman kekeringan untuk meningkatkan kemampuan penyerapan air. Pada kondisi cekaman kekeringan, kemampuan penetrasi akar pada S. italica kurang efektif dibanding spesies millet yang lain (pearl millet, barnyar millet, finger millet, dan job’stears) dan terjadi penurunan laju fotosintesis akibat perlakuan cekaman kekeringan (Zegada-Lizaruzu & Iijima 2004).
Veeranagamallaiah et al. (2007) melaporkan bahwa peningkatan
akumulasi prolin pada S. italica kultivar Prasad dan Lepakshi merupakan indikasi toleransi terhadap kondisi cekaman. Prolin berperan sebagai senyawa osmoregulator dan osmoprotektan ketika tanaman menghadapi cekaman abiotik termasuk cekaman kekeringan yang secara umum respon tersebut bersifat multigenetik.
Tujuan Penelitian
1 Mempelajari mekanisme toleransi tiga aksesi hotong terhadap cekaman
kekeringan dengan pendekatan aspek anatomi (stabilitas struktur anatomi daun, batang, dan akar) dan fisiologi (parameter fotosintesis, akumulasi asam askorbat, prolin, dan kandungan klorofil daun) yang dihubungkan dengan pertumbuhan dan produksinya.
2 Membandingkan mekanisme toleransi tiga aksesi hotong dari aspek anatomi dan fisiologi yang dihubungkan dengan produksinya sebagai indikator toleran terhadap cekaman kekeringan.
Manfaat Penelitian
Hasil penelitian ini diharapkan dapat menjadi bahan informasi dan data awal bagi pihak terkait dalam upaya pemuliaan dan budidaya tanaman hotong.
Hipotesis Penelitian
1 Perbedaan aksesi akan memberikan respon yang berbeda dalam hal
pertumbuhan dan produksi.
2 Aksesi yang toleran akan memberikan respon anatomi dan fisiologi yang
TINJAUAN PUSTAKA
Gambaran Umum Tanaman Hotong
Hotong (Setaria italica L. Beauv) atau yang di kenal dengan foxtail millet adalah tanaman liar yang di domestikasi. Tanaman ini di domestikasi dari tipe liarnya yaitu green millet (Setaria viridis) sekitar 5000 tahun yang lalu di bagian selatan padang rumput Sahara dan di bagian barat sungai Nil. Distribusinya berbeda secara geografis dan umumnya memiliki persamaan morfologi, hanya memiliki perbedaan warna bulu pada malai dan kulit biji (Lampiran 3) serta memiliki perbedaan genetik walaupun masih dalam satu spesies (Dekany 1999).
Hotong biasa tumbuh pada lahan kering dengan suhu optimum berkisar antara 24-29oC. Tanaman ini memerlukan intensitas cahaya matahari penuh. Besar kecilnya curah hujan sangat berpengaruh terhadap ketersediaan air tanah dan kelembaban di sekitar pertanaman. Jumlah curah hujan yang menunjang pertumbuhannya berkisar antara 200-300 mm/bln. Ketinggian tempat berpengaruh langsung terhadap umur panen. Ketinggian tempat yang ideal bagi pertumbuhan hotong adalah 0-450 m dpl. Tanaman ini dapat tumbuh pada berbagai jenis tanah, namun secara umum lebih menghendaki tanah yang gembur dan sedikit berpasir serta mengandung bahan organik yang cukup untuk menunjang pertumbuhannya (Karyudi & Fletcher 2003).
Fungsi Air bagi Tanaman
Karena pentingnya air pada berbagai proses di dalam tanaman, maka ketidakstabilan jumlah curah hujan di daerah tropik merupakan penyebab utama terjadinya variasi produksi tanaman dari tahun ke tahun atau dari musim ke musim. Ketidakstabilan curah hujan ini menyebabkan ketidaktentuan keadaan air tanah dan suplai hara bagi tanaman (Levitt 1980).
Cekaman Kekeringan bagi Tanaman
Tanaman mengalami cekaman kekeringan ketika terjadi ketidakseimbangan antara penyerapan air oleh akar dan kehilangan air akibat tingginya laju transpirasi yang ditandai dengan proses pelayuan. Cekaman kekeringan (drought stress) adalah faktor lingkungan yang menyebabkan tidak/kurang tersedianya air secara cukup bagi tanaman. Hal itu dapat disebabkan oleh kekurangan suplai air terutama di daerah perakaran dan adanya kebutuhan air yang berlebihan oleh daun karena laju transpirasi melebihi laju absorpsi air oleh akar tanaman, walaupun keadaan air tanah mencukupi (Bohnert et al. 1995). Kekurangan suplai air di daerah perakaran banyak dialami oleh tanaman terutama yang ditanam pada lahan-lahan kering di daerah tropis (Hamim 1995).
meningkatkan toleransinya melalui perubahan kimia sel dengan mensintesis senyawa terlarut (fisiologi) yang berfungsi sebagai osmoregulator seperti prolin dan gula (Yamada et al. 2005).
Penurunan kandungan air pada tanaman akan menyebabkan penurunan tekanan turgor pada dinding sel, sehingga sel mengalami pengkerutan. Perkembangan sel bergantung pada tekanan turgor sehingga penurunan tekanan turgor dapat menghambat perkembangan sel yang perlahan-lahan akan menghambat pertumbuhan dan perkembangan luas daun (Passiuora 1994).
Menurut Taiz dan Zeiger (2002), ketika tanaman mengalami kekurangan air akibat cekaman kekeringan, maka akan menyebabkan penurunan asimilasi karbon dan pembentukan energi untuk proses fotosintesis. Fotosintat sebagian besar didistribusikan ke akar untuk perkembangan perakaran agar mampu mencapai zona yang lebih lembab. Dalam hal ini distribusi karbohidrat terlarut melebihi kecepatan asimilasi sehingga secara simultan menurunkan pertumbuhan tanaman yang selanjutnya mengakibatkan penurunan produksi (Levitt 1980).
Pengaruh Cekaman Kekeringan terhadap Fotosintesis Tanaman
Cekaman kekeringan menyebabkan penurunan potensial air tanaman yang sangat besar pengaruhnya terhadap proses fisiologis dan metabolisme yang terjadi di dalam tanaman. Penurunan potensial air tanaman dapat menyebabkan terjadinya penurunan laju fotosintesis. Hal ini terjadi karena adanya hambatan yang ditimbulkan oleh penutupan stomata (stomatal limitation) maupun hambatan akibat penurunan proses biokimia di dalam tanaman (non-stomatal limitation) (Kalefetoglu & Ekmekci 2005).
Penurunan laju fotosintesis berhubungan dengan kombinasi beberapa proses antara lain: (1) penutupan stomata secara hidroaktif mengurangi suplai CO2 ke dalam daun, (2) dehidrasi kutikula, dinding epidermis dan membran sel, sehingga mengurangi permeabilitasnya terhadap CO2, (3) bertambahnya tahanan mesofil terhadap pertukaran gas menyebabkan penurunan efisiensi sistem fotosintesis yang berkaitan dengan proses-proses biokimia dan aktivitas enzim dalam sitoplasma (Grassi & Magnani 2005).
Pengaturan konduktansi stomata berkaitan dengan sinyal hidrolik (hydrolic signaling) dan sinyal kimia (chemical signaling). Ketika tanaman mengalami cekaman kekeringan, akan terjadi perubahan potensial air pada tanaman. Pada keadaan ini terjadi penurunan gradien potensial air antara akar dan tanah, sehingga laju penyerapan air oleh akar tanaman menurun. Gradien potensial air ini akan menimbulkan sinyal hidrolik terhadap konduktansi stomata sebagai respon tanaman terhadap cekaman kekeringan sehingga stomata menutup (Comstock 2002).
akan di hambat oleh ABA, sehingga banyak ion K+ yang keluar dari sel penjaga. Kondisi ini akan menurunkan potensial osmotik sel penjaga sehingga stomata menutup (Roberts & Snowman 2000). Proses ini dikenal dengan istilah long-distance chemical signaling (Comstock 2002).
Kondisi kekeringan yang terus berlanjut akan menyebabkan penurunan proses metabolik di dalam tanaman salah satunya yang berhubungan dengan proses transfer elektron fotosintesis. Penurunan proses metabolik disebabkan oleh berkurangnya difusi CO2 ke dalam kloroplas mesofil yang selanjutnya mengarah ke penurunan kandungan ribulosa-1,5-bisphosphat (RuBP) pada proses fotosintesis (Chaves 1991). Yang et al. (2004) menambahkan bahwa intesitas cahaya tinggi dan peningkatan suhu pada Phaseolus radiatus dan S. italica menurunkan afinitas enzim Rubisco dan konsentrasi CO2 di dalam kloroplas mesofil.
Cekaman kekeringan menginduksi terjadinya fotoinhibisi yang selanjutnya akan menurunkan kandungan protein D1 pada PSII (Pastenes et al. 2004). PSII sebagai sistem penangkap cahaya (light harvesting system) memiliki kompleks protein subunit yang berikatan dengan klorofil yang dikenal dengan protein D1 dan D2. Protein ini memiliki 2 fungsi esensial yaitu: (1) menangkap cahaya pada proses fotosintesis dan (2) melepas energi tereksitasi apabila terjadi kelebihan energi. Oleh karena itu, PSII akan merespon sinyal eksternal dari lingkungan. Proses ini berkaitan dengan perubahan gradien pH yang berfungsi sebagai kontrol balik terhadap kelebihan transfer elektron fotosintesis (Horton et al. 1996) yang dikenal dengan non-phtochemical quenching (qN). Proses perubahan nilai qN melibatkan siklus xantophyll dan protein PsbS pada PSII, sebagai mekanisme pertahanan terhadap fotoinhibisi (Taiz & Zeiger 2002).
Akumulasi Asam Askorbat (ASA) dan Prolin sebagai Respon terhadap
Kekeringan
[image:37.612.135.497.313.445.2]
tanaman akibat terbentuknya senyawa oksidatif (Blokhina et al. 2003). Proses ini menginduksi peningkatan akumulasi asam askorbat (ASA). ASA merupakan salah satu antioksidan biologi pada tanaman yang berperan sebagai agen reduksi yang dapat menetralisir reactive oxygen species (ROS) yang meliputi molekul-molekul seperti: superoksida (O2-), singlet oksigen (-O2), radikal hidroksil (.OH), dan hidrogen peroksida (H2O2) (Shgerri et al. 2000). Radikal oksigen merupakan molekul yang sangat reaktif, karena memiliki elektron yang tidak berpasangan dan dapat mengakibatkan kerusakan organel seperti kloroplas, mitokondria, dan membran plasma (Asada 2006). Reaksi pembentukan ROS dapat dilihat pada Gambar 1:
Gambar 1 Mekanisme pembentukan senyawa reactive oxygen species (ROS) (Apel & Hirt 2004).
Peluang terjadinya pembentukan ROS pada proses fotosintesis antara lain: (1) terjadi fotooksidasi pada pusat reaksi yaitu reaksi transfer elektron ke oksigen atau senyawa lain, salah satunya adalah molekul klorofil selama eksitasi energi sehingga terbentuk molekul klorofil triplet (3Chl) yang secara langsung bisa bereaksi dengan molekul oksigen dan meningkatkan transfer energi dari bentuk triplet ke bentuk singlet (1O2) (Mckersie dan Leshem 1994), (2) terjadi oksidasi dengan mentransfer 4 elektron singlet dari H2O membentuk triplet atau ground
state oksigen pada PSII, (3) terjadi reduksi oksigen melalui reaksi Mehler pada PSI dan atau deviasi elektron terhadap aliran elektron siklik pada PSII. Reduksi oksigen ini terjadi pada transfer elektron feredoksin yang terjadi ketika NADP+
Dioksigen ion radikal superoksida ion perioksida ion oksen ion oksida e
e- e- e- 3
O2 O2.- O22- O23- O- O2-
H+ 2H+ 2H+ H+ 2H+ 1O
2 HO2- H2O2 H2O OH- H2O Singlet Radikal Hidrogen Air Radikal Air oksigen perihidroksil peroksida hidroksil
terbatas, dan salah satunya disebabkan oleh berkurangnya penggunaan NADPH untuk fiksasi karbon pada siklus Calvin (Ghannoum et al. 2003).
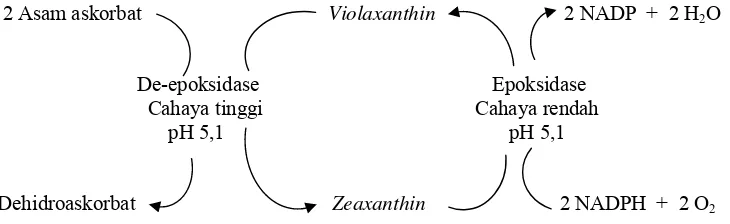
[image:38.612.139.504.333.445.2]Keterlibatan ASA dalam mekanisme penyelamatan terhadap ROS adalah melalui siklus xantophyll. ASA dapat secara langsung menetralisir ROS dan dapat bertindak sebagai ko-faktor enzim violaxanthin de-epoxidase (Fedoroff 2006) untuk mengubah violaxanthin menjadi zeaxanthin. Proses ini dilibatkan dalam perlindungan pelepasan penyerapan cahaya (energy dissipation) dalam bentuk panas (heat) dan bisa diukur sebagai non photochemical quenching dari Chlorophyll flourescence (Asada 2006). Dengan demikian akumulasi ASA merupakan suatu mekanisme penyelamatan tanaman terhadap cekaman oksidatif. Keterlibatan ASA dalam siklus xantophyll dapat di lihat pada Gambar 2:
Gambar 2 Mekanisme penyelamatan terhadap cekaman oksidatif oleh asam askorbat (ASA) (Apel & Hirt 2004).
Selain ASA, tanaman juga mengakumulasi prolin untuk menjaga keseimbangan osmotik sel saat mengalami cekaman kekeringan. Prolin diakumulasi oleh banyak tanaman budidaya sebagai respon terhadap cekaman kekeringan yang dapat diamati pada daun yang masih melekat maupun yang telah gugur (Robert & Verslues 1999). Prolin juga ditanspor ke bagian ujung akar terutama pada zona pemanjangan akar untuk merangsang pertumbuhan akar sebagai respon awal ketika terjadi defisit air (Voetberg & Sharp 1991). Akumulasi prolin pada tanaman yang terpapar kondisi kekeringan sangat penting karena senyawa ini berfungsi sebagai osmoregulator, agen pelindung bagi enzim-enzim
2 Asam askorbat Violaxanthin 2 NADP + 2 H2O
De-epoksidase Epoksidase Cahaya tinggi Cahaya rendah pH 5,1 pH 5,1
sitoplasma dan membran juga sebagai bahan simpanan untuk pertumbuhan setelah tanaman mengalami cekaman (Yamada et al. 2005).
BAHAN DAN METODE
Waktu dan Tempat Penelitian
Penelitian dilaksanakan di rumah kaca Kebun Percobaan Cikabayan, kemudian dilanjutkan di Laboratorium Fisiologi Tumbuhan, Laboratorium Anatomi dan Morfologi Tumbuhan, Departemen Biologi, FMIPA IPB. Penelitian ini telah dilaksanakan sejak bulan Juni 2008 sampai April 2009.
Bahan dan Alat
Bahan tanaman yang digunakan adalah tiga aksesi hotong (A1, A2, dan A3), aksesi 1 dan 2 merupakan aksesi lokal yang diperoleh dari Dinas Pertanian Kabupaten Buru propinsi Maluku dan aksesi 3 merupakan aksesi hasil pemuliaan dari Departemen Agronomi dan Hortikultura IPB. Media tanam merupakan campuran tanah dan pasir (1:1 v/v) masing-masing 3 kg. Pupuk dasar terdiri atas pupuk kandang sebanyak 10% dari bobot media dan Urea:SP-36: KCl sebanyak 200:150:200 kg/Ha (Diptan 2003).
Bahan kimia yang digunakan adalah untuk analisis asam askorbat (ASA), prolin, dan klorofil terdiri atas; nitrogen cair, asam askorbat (4,0 mg/l-1), asam metaphosphorik 5%, dicholorophenol-indophenol (DCIP) 0,8 g L-1, ninhidrin, asam asetat glasial, asam fosfat 6 M, asam sulfosalisilik 3%, toluen, prolin murni, aseton 80%, kertas saring (Wathman no 1). Sedangkan untuk preparasi anatomi daun, batang, dan akar tediri atas: alkohol (pa, 95, 70, 60, 50, & 30%), HNO3 40%, larutan pemutih (bahan aktif), safranin 2%, fast green 0.5%, gliserin 30%, larutan albumin gliserin 30%, larutan FAA, larutan giford, larutan seri n-butanol III – VII, paraplas, entelan, dan aquades.
pipet volumetrik, pipet tetes, pipet 1 ml, 5 ml dan 10 ml, gelas piala, tabung reaksi, batang pengaduk, bulb, lemari asam, tabung film, gunting, pisau mikrotom, kater, silet, kuas, mikroskop cahaya, cawan petri, gelas objek, cover gelas, pinset, kamera, kertas label, mikrotom putar, botol sampel, luxmeter, bunsen, hotplate, cetakan blok parafin, kotak preparat, dan sebagainya.
Rancangan Percobaan
Penelitian dirancang dan dilaksanakan dengan metode rancangan faktorial dalam rancangan acak kelompok dengan 3 ulangan. Sedangkan faktor yang diamati terdiri atas 2 faktor. Faktor pertama adalah aksesi terdiri atas 3 taraf yaitu; aksesi Buru 1 (A1), aksesi Buru 2 (A2), dan aksesi AGH (A3). Faktor kedua adalah perlakuan terdiri atas 2 taraf yaitu; disiram setiap hari sesuai volume air pada kondisi kapasitas lapang (C0) dan cekaman kekeringan dengan penundaan penyiraman selama 14 hari (CK). Dengan demikian terdapat 18 unit uji dari 6 kombinasi perlakuan dengan 3 kali ulangan. Total tanaman yang disiapkan adalah 126 tanaman. Data yang di peroleh dianalisis dengan sidik ragam RAK dengan uji jarak berganda Duncan (DMRT) sebagai uji lanjut pada tarap kepercayaan 95% (α = 0,05) dengan SPSS versi 13.
Pelaksanaan Penelitian
Persiapan bahan tanaman dan media tanam
Penanaman dan pemeliharaan
Benih hotong ditanam langsung pada pot yang telah disiapkan, sebanyak 4 biji/pot untuk setiap perlakuan. Setelah umur satu minggu, tanaman dijarangkan menjadi dua tanaman/pot. Pupuk dasar yang diberikan terdiri atas pupuk kandang sebanyak 10% dari bobot media tanam dan Urea:SP-36:KCl dengan perbandingan 200:150:200 kg/Ha (Diptan 2003).
Penyemprotan untuk pencegahan hama dan penyakit tanaman dilakukan dengan menggunakan insektisida sesuai dengan tingkat dan jenis serangan hama. Selain itu dilakukan penyiangan tanaman pengganggu secara berkala. Penyiraman dilakukan setiap hari secara merata sesuai volume air pada kondisi kapasitas lapang sampai tanaman tumbuh sempurna.
Pemberian perlakuan cekaman kekeringan
Setiap aksesi mendapatkan dua perlakuan, yaitu perlakuan disiram setiap hari (C0), dan tanpa disiram selama 14 hari (CK). Sejak penanaman sampai tanaman berumur 6 minggu, air diberikan sesuai volume air pada kondisi kapasitas lapang. Setelah itu dilakukan penundaan penyiraman selama 14 hari sampai tanaman menunjukkan gejala layu berat. Keadaan layu berat ditandai dengan tanaman tetap layu apabila dipindahkan keruang lembab selama satu malam.
Pengamatan
14 hari setelah perlakuan (HSP) cekaman kekeringan dan 2 hari setelah pengairan kembali (rewatering). Pengukuran luas daun, tinggi tajuk, panjang akar, bobot kering tajuk dan akar, serta struktur anatomi daun, batang, dan akar melalui pembuatan preparasi sayatan paradermal dan transversal dilakukan pada 14 HSP cekaman kekeringan. Bobot kering biji diukur setelah tanaman berproduksi.
Penentuan KAM dilakukan secara gravimetri dengan mengambil sampel tanah ± 100/g (di bagian atas, tengah, dan bawah). Sampel tanah ditimbang untuk memperoleh bobot basah (BB), kemudian dioven pada suhu 80oC selama 48 jam untuk mendapatkan bobot kering (BK). KAM diperoleh dengan persamaan sebagai berikut:
BB - BK
KAM (%) = x 100% BB
KAR daun ditetapkan mengikuti metode (Barr & Weatherley 1962). Sebanyak 10 bulatan sampel daun diambil menggunakan cork borer berdiameter 1 cm. Selanjutnya sampel daun ditimbang untuk memperoleh bobot segar (BS), kemudian dilakukan hidrasi selama 24 jam di dalam botol film untuk memperoleh bobot jenuh (BJ) yang sebelumnya dikeringkan terlebih dahulu menggunakan kertas tissue. Sampel daun selanjutnya dioven pada suhu 80oC selama 48 jam untuk mendapatkan bobot kering (BK) yang sebelumnya didinginkan terlebih dahulu di dalam desikator. KAR daun diperoleh dengan persamaan sebagai berikut:
BS - BK
KAR daun (%) = x 100% BJ - BK
Luas daun (LD) dihitung dengan mengukur panjang daun (P) yang diukur mulai dari pangkal daun sampai ujung daun terpanjang dan lebar daun (L) yang diukur pada bagian helai daun yang terlebar mengukuti metode Guritno dan Sitompul (1995) dengan persamaan sebagai berikut:
LD (cm2) = P x L x k
k adalah konstanta kalibrasi (0,74).
Tinggi tajuk diukur mulai dari permukaan tanah sampai pada titik tumbuh. Panjang akar diukur mulai dari pangkal akar sampai ujung akar terpanjang, sedangkan bobot kering akar dan tajuk diukur secara terpisah dengan cara dioven pada suhu 80oC selama 48 jam. Bobot kering biji diukur setelah tanaman berproduksi. Biji yang diperoleh dijemur selama 2 minggu di dalam rumah kaca, kemudian ditimbang untuk mendapatkan bobot kering biji.
Pengukuran parameter fotosintesis
Parameter fotosintesis diukur dengan menggunakan alat Photosynthetic Chlorophyll Flourescence Analyzer (Qubit Systems tipe ATC 1). Sampel daun yang diukur adalah daun muda yang berkembang sempurna (daun ketiga). Daun tanaman diletakkan pada kuvet kemudian diadaptasi gelap dengan cara ditutup dengan kain hitam selama 20 menit. Selanjutnya diberi cahaya saturasi (cahaya jenuh). Pada saat di beri cahaya jenuh, fluorescence akan meningkat dari nilai ground state (Fo) ke nilai maksimum (Fm). Dari sini diperoleh nilai efisiensi maksimum fotosintesis (Fv/Fm) dengan persamaan Fv/Fm = (Fm-Fo)/Fm. Selang 2 menit dinyalakan Actinic light, pada kondisi ini flourescence yield meningkat pada kondisi nilai steady state (Ft). Pada interval 20 detik Saturating flash dinyalakan, maka akan meningkatkan flourescence yield pada kondisi maksimum (Fm’) (nilainya akan lebih rendah dari kondisi pada saat adaptasi gelap/tanpa actinic light on). Maka diperoleh photochemical quenching (qP) = (Fm’ – Ft)/(Fm’ – Fo), non-photochemical quenching (qN) = (Fm – Fm’)/(Fm – F0) dan quatum yield (qY) dari transfer elektron pada PSII (qY) = (Fm’ – Ft)/Fm’.
Analisis asam askorbat (ASA)
daun segar sebanyak 5 g diekstraksi dalam 10 ml asam metaphosphorik 5%. Hasil ekstraksi difiltrasi dengan kertas saring Wathman no 1. Filtrat yang di peroleh dititrasi dengan dicholorophenol-indophenol (DCIP) 0,8 g L-1. Sebelum dititrasi, dilakukan standarisasi pewarna (larutan DCIP) dengan larutan ASA murni, yaitu 1
ml larutan yang mengandung asam askorbat (4,0 mg L-1) dan 9 ml asam
metaphosphorik 5% dititrasi dengan dicholorophenol-indophenol (DCIP) 0,8 g L-1. Titrasi dihentikan ketika filtrat tepat berwarna merah muda. Kandungan ASA dihitung berdasarkan persamaan sebagai berikut:
1 Untuk standarisasi larutan ASA ( 4 mg ASA murni = 1 ml pewarna): ASA (mg) 4 mg ASA murni
=
1ml DCIP DCIP yang dititrasi (ml)
2 Untuk mengetahui kandungan ASA daun tanaman (ASA / 100 g jaringan daun):
Total volume ekstrak (ml) 100 = (mg ASA) x x
Volume yang dititrasi (ASA) g sampel
Analisis prolin
kemudian diabsorbansi pada λ 520 nm dengan spektrofotometer. Sebagai blangko digunakan toluen. Konsentrasi prolin ditentukan dari kurva standar dan dihitung berdasarkan persamaan sebagai berikut:
(µg prolin / ml x ml toluene) / 115,5 µg / µmol (g sampel) / 5
= µ mol prolin / g bobot segar sampel.
Anlaisis kandungan klorofil
Analisis kandungan klorofil dilakukan mengikuti metode Arnon menggunakan spektrofotometer (Arnon 1949). Sebanyak 1 g sampel daun segar tanpa tulang daun digerus dengan aseton 80%. Supernatan disaring dengan kertas saring dan diencerkan sampai volume 50 ml. Selanjutnya diambil 2,5 ml larutan dan diencerkan lagi sampai volume 25 ml. Kemudian di absorbansi dengan spektrofometer pada λ 663 nm (klorofil a) dan λ 645 nm (klorofil b). Kandungan klorofil a, b, dan total dihitung berdasarkan persamaan sebagai berikut:
Kla = 0,0127 . D663 – 0,00269 . D645 Klb = 0,0229 . D645 – 0,00468 . D663
50 100
Kltot = 20,2 . D645 + 8,02 . D663 x x x 0,5 (mg/g BK) 1000 50
Pembuatan sayatan paradermal
yang telah diberi media gliserin 30%. Selanjutnya ditutup dengan cover gelas dan diamati pada mikroskop cahaya. Kerapatan dan indeks stomata pada lapisan epidermis daun bagian atas dan bawah dihitung berdasarkan persamaan (Wilmer 1983) sebagai berikut:
Jumlah stomata Kerapatan stomata =
Luas daun (mm2)
Luas daun diperoleh dari luas bidang pandang lensa objektif dengan perbesaran 10 x 40 dengan persamaan πr2 (π = 3,14 & r = 0,22). Luas daun = 0,152 mm2.
Jumlah stomata
Indeks stomata = x 100% Jumlah stomata + Jumlah sel epidermis
Pembuatan sayatan transversal
HASIL DAN PEMBAHASAN
Kadar Air Media (KAM) dan Kandungan Air Relatif (KAR) Daun
Dalam percobaan pendahuluan yang dilakukan untuk menentukan periode waktu yang tepat bagi perlakuan cekaman kekeringan pada ketiga aksesi hotong diperoleh hasil bahwa perlakuan cekaman kekeringan dengan penundaan penyiraman selama 14 hari merupakan waktu yang cukup tepat saat kadar air media diperkirakan terletak diantara kapasitas lapang dan titik layu permanen. Keadaan ini ditandai setelah diberi penyiraman kembali (rewatering), tanaman perlakuan kembali segar seperti tanaman kontrol. Hal ini menunjukkan bahwa ketiga aksesi hanya mengalami layu sementara dan memiliki kemampuan untuk tumbuh kembali setelah mendapat cekaman kekeringan. Dengan demikian pada 14 HSP cekaman kekeringan tanaman belum mencapai titik layu permanen, walaupun kadar air media rata-rata pada ketiga aksesi di bawah 10%.
Gambar 3 Kadar air media (KAM) (%) (A) dan kandungan air relatif (KAR) daun (%) (B) pada 0-14 HSP cekaman kekeringan dan recovery pada taraf uji 5% (DMRT). Tanda panah menunjukkan titik rewatering.
Perlakuan cekaman kekeringan dengan menunda waktu penyiraman selama 14 hari pada ketiga aksesi hotong mengakibatkan penurunan KAM (Gambar 3A). Nilai rata-rata KAM pada tanaman kontrol sebesar 12,8%,
Aksesi 1 Aksesi 2 Aksesi 3
Hari setelah perlakuan
C0 ‐ ‐ ‐ ‐CK
A B 0 5 10 15 20
0 4 8 12 14 16
KA M (% ) 0 5 10 15 20
0 4 8 12 14 16
0 5 10 15 20
0 4 8 12 14 16
0 25 50 75 100
0 4 8 12 14 16
K AR ( % ) 0 25 50 75 100
0 4 8 12 14 16
0 20 40 60 80 100
sedangkan pada tanaman perlakuan sebesar 7,8%, sehingga terjadi penurunan kadar air media sebesar 39,1%. Selain itu secara fisual tampak pula perbedaan antara media pada kontrol dan perlakuan cekaman kekeringan. Media pada perlakuan tampak kering di bagian permukaan sampai ke bagian yang lebih dalam.
Penurunan KAM terjadi secara linear pada 14 HSP kekeringan. Penurunan KAM terbesar terjadi pada aksesi 1 sebesar 44,3% diikuti aksesi 3 sebesar 39,4% dan terendah pada aksesi 2 sebesar 28,6%. Penurunan KAM pada aksesi 1 berbeda nyata dengan tanaman kontrol mulai terjadi pada 8-14 HSP, sedangkan pada aksesi 2 dan aksesi 3 menunjukkan berbeda nyata dengan tanaman kontrol hanya pada 14 HSP kekeringan. Hal ini menunjukkan bahwa kemungkinan laju evaporasi lebih tinggi pada aksesi 1 dibanding pada aksesi 2 dan 3. Secara umum pada 14 HSP cekaman kekeringan nilai KAM rata-rata pada ketiga aksesi di bawah 10%. Fenomena ini kemungkinan disebabkan oleh rendahnya daya ikat air oleh komponen media sehingga terjadi kehilangan air yang cukup tinggi melalui proses evaporasi pada ketiga aksesi selama periode cekaman. Penurunan ini kemungkinan juga disebabkan oleh peningkatan laju absorpsi air oleh akar tanaman yang berkaitan dengan aktivitas metabolisme dan laju transpirasi pada tanaman (Levitt 1980).
Penurunan KAM dapat mengakibatkan terjadinya ketidakseimbangan aliran air pada tanaman, sehingga terjadi defisit air dan gangguan fungsi fisiologis di dalam sel tanaman. Laju aliran air dalam tanah bergantung pada 2 faktor yaitu besarnya gradien tekanan air tanah dan konduktivitas hidrolik tanah. Aliran air ini terkait dengan potensial air, potensial osmotik, dan gradien tekanan. Ketika kandungan air tanah menurun, konduktivitas hidrolik tanah menurun drastis. Penurunan ini terjadi karena adanya penguapan (Taiz & Zeiger 2002).
media kurang dari kisaran tersebut maka media harus diairi karena air tersedia bagi kebutuhan tanaman tidak terpenuhi (Hanafiah 2007).
Secara umum KAM berkorelasi positif dengan KAR daun (Lampiran 1) pada ketiga aksesi terutama pada 14 HSP kekeringan. Dengan demikian penurunan KAM akibat perlakuan kekeringan menyebabkan penurunan KAR daun secara nyata pada ketiga aksesi (Gambar 3B) dengan pola yang hampir sama dengan penurunan KAM (Gambar 3A). Penurunan KAR daun dari ketiga aksesi secara berturut-turut adalah: aksesi 2 sebesar 44,6%, aksesi 1 sebesar 43,7%, dan aksesi 3 sebesar 40,9%. Penurunan KAR daun secara tajam ini mungkin disebabkan oleh laju transpirasi yang jauh lebih besar dari pada laju penyerapan air oleh akar. Penurunan KAR daun juga diikuti oleh penurunan turgor daun yang ditandai dengan proses pelayuan daun sejak 8-14 HSP cekaman kekeringan (Lampiran 4). Proses pelayuan ini meyebabkan terjadinya penggulungan daun pada ketiga aksesi yang merupakan salah satu mekanisme tanaman menghindari cekaman kekeringan (hidronasti). Penggulungan daun terjadi karena proses pengkerutan sel bulliform yang berfungsi untuk melindungi jaringan dibawahnya agar tidak mengalami kerusakan akibat kehilangan air yang lebih besar (Fahn 1990).
KAR daun pada 14 HSP kekeringan secara umum berkorelasi positif dengan beberapa parameter lainnya seperti luas daun, kerapatan stomata, tebal mesofil, Fv/Fm, qP, qY, dan bobot kering biji pada ketiga aksesi (Lampiran 1). Penurunan KAR daun ini mungkin menyebabkan penurunan konduktansi stomata dan bertambahnya tahanan mesofil terhadap difusi CO2 ke dalam kloroplas mesofil, sehingga mempengaruhi perubahan laju fotosintesis dan proses metabolisme dasar lainnya (Kramer 1995). Hal ini berkontribusi terhadap penurunan produksi pada ketiga aksesi yang mendapat perlakuan cekaman kekeringan.
kontrol. Peningkatan KAR daun diperlukan untuk perbaikan tanaman dari kerusakan akibat perlakuan cekaman kekeringan. Fusiana (1997) dan Violita (2007) telah melakukan penelitian pada tanaman padi gogo, kedelai, dan jagung bahwa pemberian air kembali pada tanaman yang mendapat perlakuan cekaman kekeringan dapat meningkatkan KAM dan KAR daun hingga mencapai nilai yang sama pada level kontrol. Air sebagai komponen utama tanaman dibutuhkan untuk berbagai proses metabolisme tanaman untuk pertumbuhan dan perkembangan tanaman termasuk transportasi air dan hara. Pada kondisi ini tanaman telah mampu menjaga keseimbangan gradien potensial osmotik antara media-akar dan tajuk (Taiz & Zeiger 2002).
Respon Umum Tanaman
[image:52.612.127.507.369.449.2]
Penurunan luas daun pada ketiga aksesi penting untuk meminimalisir kehilangan air melalui transpirasi selama periode cekaman agar potensial air sel tetap terjaga, namun berpengaruh negatif terhadap laju fotosintesis. Selama cekaman kekeringan, laju pertumbuhan dan perkembangan daun muda terhambat, terjadi pengkerutan sel dan proses senesensi yang diikuti pengguguran daun tua, sehingga menyebabkan reduksi area fotosintesis, yang selanjutnya akan berpengaruh terhadap penurunan produksi tanaman. (Salisbury & Ross 1992). Keadaan ini disebabkan oleh kurangannya suplai air yang dibutuhkan untuk proses metabolisme dan fisiologi (pemelaran & pemanjangan sel) selama fase pertumbuhan tanaman.
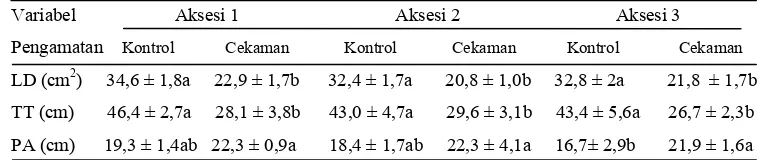
Tabel 1 Pengaruh cekaman kekeringan terhadap luas daun, tinggi tajuk, dan panjang akar ketiga aksesi hotong pada 14 hari setelah perlakuan cekaman kekeringan
Variabel Aksesi 1 Aksesi 2 Aksesi 3
Pengamatan Kontrol Cekaman Kontrol Cekaman Kontrol Cekaman
LD (cm2) 34,6 ± 1,8a 22,9 ± 1,7b 32,4 ± 1,7a 20,8 ± 1,0b 32,8 ± 2a 21,8 ± 1,7b
TT (cm) 46,4 ± 2,7a 28,1 ± 3,8b 43,0 ± 4,7a 29,6 ± 3,1b 43,4 ± 5,6a 26,7 ± 2,3b
PA (cm) 19,3 ± 1,4ab 22,3 ± 0,9a 18,4 ± 1,7ab 22,3 ± 4,1a 16,7± 2,9b 21,9 ± 1,6a
LD: luas daun, TT: tinggi tajuk, PA: panjang akar. Angka yang diikuti oleh huruf yang sama pada masing-masing baris tidak berbeda nyata antara tanaman kontrol dan perlakuan pada taraf uji 5% (DMRT).
[image:53.612.129.506.321.421.2]
ketiga aksesi (Lampiran 1). Dengan demikian perlakuan cekaman kekeringan menginduksi peningkatan akumulasi prolin yang diduga mempengaruhi peningkatan panjang akar pada ketiga aksesi dengan adanya sinyal kimia yang dikendalikan oleh asam absisat (ABA) (Comstock 2002). Peningkatan panjang akar berbeda nyata antara tanaman kontrol dan perlakuan pada aksesi 3, sedangkan pada aksesi 1 dan 2 relatif sama dengan tanaman kontrolnya. Peningkatan panjang akar tertinggi terjadi pada aksesi 3 sebesar 24% dan terendah pada aksesi 1 sebesar 13,5%.
Tabel 2 Pengaruh cekaman kekeringan terhadap bobot kering tajuk dan akar dan bobot kering biji ketiga aksesi hotong
Variabel Aksesi 1 Aksesi 2 Aksesi 3
Pengamatan Kontrol Cekaman Kontrol Cekaman Kontrol Cekaman
………..… (g) ………
BKT/pot 16,6 ± 4,3a 8,3 ± 1,9bc 12,5 ± 1,8ab 9,2 ± 0,9bc 8,2 ± 1,8bc 4,5 ± 1,5c
BKA/pot 3,1 ± 0,4a 2,0 ± 0,4ab 1,8 ± 0,3b 1,6 ± 0,4bc 1,1 ± 0,3cd 0,7 ± 0,2d
BKB/pot 9,3 ± 1,5ab 6,3 ± 2,3bc 11,0 ± 1,7a 5,1 ± 0,4c 9,1 ± 1,2ab 7,6 ± 0,4bc
BKT: bobot kering tajuk, BKA: bobot kering akar, BKB: bobot kering biji. Angka yang diikuti oleh huruf yang sama pada masing-masing baris tidak berbeda nyata antara tanaman kontrol dan perlakuan pada taraf uji 5% (DMRT).
Perbedaan peningkatan panjang akar pada ketiga aksesi ini menunjukkan bahwa aksesi 3 memiliki kemampuan adaptasi yang cukup baik terhadap cekaman kekeringan dengan nilai R2 sebesar 0,71 (Lampiran 2C) dibanding aksesi 1 dan 2. Aksesi 3 mampu memanfaatkan senyawa prolin yang terakumulasi selama periode cekaman untuk menunjang pertumbuhan akarnya guna meningkatkan penyerapan air pada zona yang lebih lembab. Akumulasi prolin di daun akan ditranspor ke akar sebagai sumber energi utama untuk merangsang pertumbuhan ujung akar. Proses ini dapat terlihat dari laju peningkatan konsentrasi dan deposit prolin pada zona pemanjangan akar primer tanaman jagung pada kondisi potensial air rendah (Voetberg & Sharp 1991).
mempengaruhi bobot kering akar pada tanaman perlakuan yang lebih rendah dibanding kontrol. Walaupun tidak dilakukan pengukuran pertumbuhan akar lateral secara kuantitatif, hal ini dapat diamati pada morfologi akar ketiga aksesi yang mendapat perlakuan kekeringan pada akhir periode cekaman kekeringan (Lampiran 5).
Penghambatan pertumbuhan tajuk dan peningkatan panjang akar selama cekaman kekeringan ini mempengaruhi keseimbangan antara pertumbuhan akar dan tajuk. Menurut Violita (2007), ketidakseimbangan pertumbuhan akar dibanding tajuk selama cekaman kekeringan mempengaruhi rasio akar terhadap tajuk yang disebabkan oleh faktor internal dan eksternal. Ketidakseimbangan pertumbuhan ini terkait dengan distribusi fotosintat yang dibutuhkan organ tanaman untuk menunjang pertumbuhan dan perkembangannya (Srivastava 2002). Tajuk akan tumbuh hingga terbatasnya penyerapan air oleh akar tanaman untuk pertumbuhan selanjutnya, sebaliknya akar akan tumbuh hingga permintaan fotosintat dari tajuk sama dengan suplai fotosintat ke tajuk. Peningkatan panjang akar pada ketiga aksesi akibat cekaman kekeringan ini merupakan respon terhadap kondisi kekeringan yang terkait dengan kemampuan akar untuk memperoleh air tanah pada zona yang lebih dalam (Karyudi & Fletcher 2003).
Akar pada tanaman yang terpapar kondisi kekeringan berfungsi sebagai sensor utama terhadap cekaman kekeringan. Perubahan kadar air tanah akan menyebabkan perubahan metabolisme akar seperti penurunan produksi sitokinin, peningkatan produksi ABA, dan gangguan metabolisme nitrogen yang akan mengirim sinyal kimia ke tajuk. Sinyal ini diduga menginduksi akumulasi senyawa-senyawa pertahanan di daun sehingga terjadi perubahan-perubahan pada tanaman seperti penurunan pertumbuhan, konduktansi stomata, dan laju fotosintesis tanpa memperhatikan status air daun, namum disisi lain menginduksi pertumbuhan akar pada beberapa species tanaman budidaya (Kramer 1995).
tajuk dan akar maupun bobot kering biji (Tabel 2). Tanaman hotong yang mendapat perlakuan cekaman kekeringan memiliki bobot kering tajuk dan akar lebih rendah dibanding tanaman kontrol. Penurunan bobot kering tajuk dan akar terbesar terjadi pada aksesi 1 dan yang terendah pada aksesi 2. Penurunan ini berbeda nyata dengan tanaman kontrol pada aksesi 1, sedangkan pada aksesi 2 dan 3 relatif sama dengan tanaman kontrolnya. Hal ini menunjukkan bahwa distribusi dan akumulasi fotosintat untuk menunjang pertumbuhan tajuk dan akar pada aksesi 2 dan 3 selama periode cekaman relatif seimbang, sehingga dapat mempertahankan bobot kering tanaman dengan nilai yang relatif sama dengan tanaman kontrolnya. Perbedaan respon ini kemungkinan juga di pengaruhi oleh perbedaan genetik antar ketiga aksesi.
Bobot kering merupakan parameter pertumbuhan yang baik untuk mengamati pengaruh cekaman kekeringan. Bobot kering merupakan akumulasi fotosintat tanaman selama pertumbuhannya (Levitt 1980). Penurunan bobot kering ini mungkin terkait dengan penurunan laju fotosintesis pada ketiga aksesi selama periode cekaman kekeringan. Hal ini terjadi karena ketidakseimbangan transpor hara dan distribusi fotosintat ke seluruh bagian tanaman akibat penurunan KAM sehingga mempengaruhi proses transpor hara dan distribusi fotosintat.
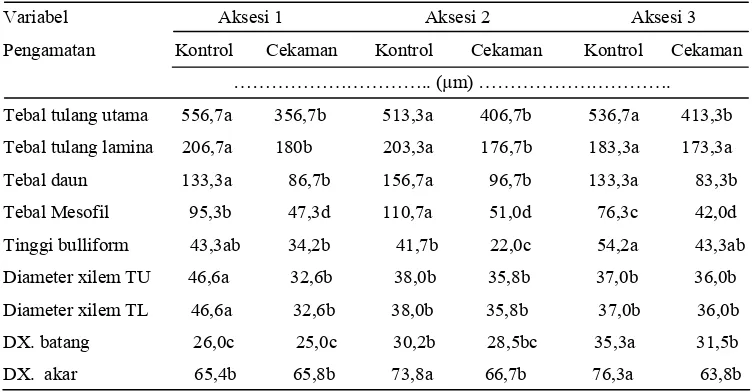
aksesi 1 dan 3. Selain itu aksesi 1 dan 3 cenderung mempertahankan stabilitas struktur anatominya terutama pada daun (tebal daun & mesofil) dibanding aksesi 2 (Tabel 4). Perubahan anatomi daun yang di amati pada 14 HSP kekeringan terutama pada lapisan mesofil berkorelasi positif (0,86) terhadap penurunan luas daun pada ketiga aksesi (Lampiran 1). Perubahan ini menyebabkan reduksi area
permukaan daun sehingga menghambat difusi CO2 kedalam daun yang
bergantung pada ukuran ruang sel mesofil (Wise & Naylor 1987). Penurunan stabilitas