KEPADATAN SYMPHYLID PADA EKOSISTEM NANAS, PISANG, JAMBU BIJI, DAN LIDAHBUAYA DI KAWASAN
TERBANGGI BESAR LAMPUNG TENGAH
(Skripsi)
Oleh
EKO ANDRIANTO
FAKULTAS PERTANIAN UNIVERSITAS LAMPUNG
ABSTRACT
SYMPHYLID ABUNDANCE IN PINEAPPLE, BANANA, GUAVA AND ALOEVERA ECOSYSTEMS IN TERBANGGI BESAR AREA,
CENTRAL LAMPUNG
By
EKO ANDRIANTO
A survey was conducted in the Great Giant Pineapple Company (PT GGP) land, Terbanggi Besar area Central Lampung, from May to June 2015 to determine the abundance of symphylids in four ecosystems in the area. Four ecosystems in the area, i.e. pineapple, banana, guava, aloevera, each size 5 – 20 ha, were observed. In each ecosystem, four sample plots, each size 0.2 – 0.8 ha, were taken for further selection of the sample points. Each sample point was selectively located at the corner (10 m by 10 m coordinate) of each sample plot. Three data were obtained from each sample point, i.e. abundance of symphylids (collected using bait trap & hand-sorting method), litter weight (raw surface necromass), and soil pH (using pH meter). Results show that the highest symphylid abundance
occurred in the guava ecosystem. The symphylid abundance in the pineapple was fair and that in the banana and aloevera ecosystems were the lowest. The
abundance of symphylids in the guava and banana ecosystems was affected by the litter weight and soil pH. Either factor affected the symphylid abundance in the pineapple ecosystem (litter weight) and that in the aloevera ecosystem (soil pH).
KEPADATAN SYMPHYLID PADA EKOSISTEM NANAS, PISANG, JAMBU BIJI, DAN LIDAHBUAYA DI KAWASAN
TERBANGGI BESAR LAMPUNG TENGAH
Oleh
EKO ANDRIANTO
Skripsi
Sebagai Salah Satu Syarat untuk Mencapai Gelar SARJANA PERTANIAN
pada
Jurusan Agroteknologi
FAKULTAS PERTANIAN UNIVERSITAS LAMPUNG
RIWAYAT HIDUP
Penulis dilahirkan di Desa Bandar Agung Kecamatan Terusan Nunyai, Lampung Tengah pada tanggal 21 Januari 1992. Penulis merupakan anak petama dari dua bersaudara pasangan Bapak Suparni dan Ibu Siti Nur Hayati.
Penulis menyelesaikan pendidikan dasar di Sekolah Dasar Negeri 3 Bandar Agung, Lampung Tengah pada tahun 2004. Kemudian melanjutkan ke jenjang sekolah menengah di SMP Negeri 3 Terusan Nunyai, Lampung Tengah dan lulus pada tahun 2007. Pendidikan menengah atas ditempuh di SMA Negeri 1 Terusan Nunyai, Lampung Tengah dan lulus pada tahun 2010. Pada tahun yang sama penulis melanjutkan ke jenjang perkuliahan dan berhasil terdaftar sebagai Mahasiswa Jurusan Agroteknologi, Fakultas Pertanian, Universitas Lampung melalui jalur Penelusuran Kemampuan Akademik dan Bakat (PKAB) dan dalam program Beasiswa BIDIK MISI 2010 angkatan 1 (pertama).
Penelitian dan Pengembangan Pengurus Besar PII Periode 2015-2017. Selain itu dalam bidang kepenulisan, Penulis tergabung dalam Forum Lingkar Pena (FLP) Bandarlampung, pernah menjadi Juara I Lomba Penulisan Buletin Jumat Tingkat Nasional Lazuardi Birru tahun 2012 dan Juara II Lomba Penulisan Artikel Pekan Cinta Tanah Air (PCTA) Kementerian Pertahanan tingkat Provinsi Lampung tahun 2014.
Di tengah aktivitas organisasi dan akademiknya, penulis tercatat pernah menjadi asisten dosen praktikum untuk beberapa mata kuliah umum dan khusus bidang proteksi tanaman. Mata kuliah tersebut meliputi Ilmu Hama Tumbuhan Umum (2012), Teknologi Benih (2012), Hama Penting Tanaman (2013), Bioekologi Hama Tumbuhan (2013, 2014), Hama Nir Serangga (2014), Pengendalian Hama Gudang dan Urban (2015), Pengendalian Terpadu Hama & Penyakit Tanaman (2013), Kewirausahan Pertanian (2012), Kewirausahaan (2014), Pengendalian Hama & Penyakit Terpadu (PHT) Tanaman Karet (2014), PHT Tanaman Sawit (2013), PHT Tanaman Tebu (2014, 2015), Klinik Tanaman (2015), Entomologi Pertanian (2014, 2015), Metodologi Penelitian (2015) dan Bahasa Indonesia (2015).
Dengan penuh rasa syukur dan ta’zim, karya ilmiah ini
didedikasikan untuk:
Keluargaku Tercinta,
Ayahanda Suparni dan Ibunda Siti Nurhayati,
Adinda Mei Irianti,
Nenek Sutini tersayang dan
Keluarga Besar Moeniran.
Seluruh Insan Akademis dan
“Adakah sama orang-orang yang mengetahui dengan orang-orang yang tidak mengetahui?.
Sesungguhnya orang yang berakallah yang dapat menerima pelajaran.”
(Qs. Az- Zumar: 9)
“Jika benar hidup ini hanyalah permainan, bukankah tugas kita hanya bermain dengan baik dan bermain hingga akhir.”
(Nasihat F.X Susilo untuk Eko Andrianto)
“If We knew what it was we’re doing, it would not be called Research, would it?” (Jika kita mengetahui apa (hasil) yang akan kita lakukan, itu tidak akan disebut riset, bukan?)
(Albert Einstein)
“Jika dilupakan itu menakutimu, maka abaikan saja, karena Tuhan tahu itu menimpa semua orang.”
(Film “The fault on our star”)
“Better to fight for something than live for nothing” (General George S. Patton Jr.)
Yesterday I was clever....So I wanted to change the world....
Today I am wise....So I am changing Myself....
SANWACANA
Puji dan syukur kehadirat Allah SWT yang telah melimpahkan rahmat dan hidayah-Nya sehingga penulis dapat menyelesaikan penulisan skripsi ini. Pada kesempatan kali ini penulis ingin mengucapkan rasa terima kasih kepada : 1. Bapak Prof. Dr. Ir. F.X. Susilo, M.Sc., pembimbing utama yang telah
memberikan banyak ilmu dan wawasan, nasihat serta semangat, bimbingan dan juga teguran pada setiap proses yang terlewati dalam penelitian hingga selesainya penulisan skripsi ini.
2. Bapak Ir. Solikhin, M. P., pembimbing kedua yang dengan sabar telah menguatkan di kala lemah, memberikan teladan sikap di kala lengah, serta bimbingan dan ilmu yang sangat berharga bagi penulis.
3. Bapak Ir. Agus. M. Hariri, M. P., pembahas atas segala curahan perhatian dan nasehat, bimbingan dan doa, serta kritik yang membangun dalam penulisan skripsi ini.
4. Bapak Prof. Dr. Ir.Wan Abbas Zakaria, M. S., Dekan Fakultas Pertanian Universitas Lampung.
5. Bapak Dr. Ir. Kuswanta F. Hidayat, M. P., Ketua Jurusan Agroteknologi. 6. Bapak Dr. Ir. Rusdi Evizal, M. S., dosen pembimbing akademik
yang luar biasa dalam melakukan penelitian ini baik di rumah, lapangan maupun di laboratorium.
8. Ayahanda dan Ibunda serta Nenek tercinta yang telah memberikan doa dan harapan, cinta dan kepercayaan dalam setiap diam dan untaian panjang nasehatnya. Serta adinda, Mei Irianti, atas segala doa serta kebanggaan yang telah diberikan dalam keseharian.
9. Bapak Muh. Basuki, beserta staf R&D PT GGP yang tidak bisa penulis sebutkan satu persatu, atas kesempatan dan kerja samanya selama penelitian . 10. Sahabat-sahabat: Mustajab, S.P., Yoseph Albert LDJP, S.P., dan Aldi I.P,
terimakasih atas bantuan dan dukungan selama ini, biarlah jalanan menjadi saksi dan Tuhan yang membalas kebaikan kalian.
11. Adinda-adinda Kelompok Studi Proteksi Tanaman (KS Protekta), terima kasih untuk setiap bantuan yang diberikan dengan rela maupun terpaksa. 12. Teman-teman agroteknologi kelas A dan agroteknologi 2010 yang telah
mengisi makna dalam ruang interaksi selama penulis berada di kampus.
Penulis menyadari bahwa skripsi ini jauh dari sempurna, oleh karena itu penulis menerima saran dan kritik yang bersifat membangun agar skripsi dapat
bermanfaat bagi semua pihak.
Bandar Lampung, September 2015 Penulis
i
2.3.4 Pengendalian Hayati Hama Symphylid ... 18
2.3.5 Pengendalian Kimiawi ... 19
2.5 Populasi Hama & Faktor yang Mempengaruhinya ... 20
2.6 Interaksi Herbvora Pemakan Akar, Akar Tanaman, dan Faktor Abiotik Tanah ... 22
ii
3.4.2.1 Penghitungan Berat Serasah ... 33
3.4.2.2 Analisis pH Tanah ... 34
3.5 Teknik Pengamatan dan Pengumpulan Data ... 34
3.5.1 Pengamatan Kepadatan Relatif Symphylid ... 34
3.5.2 Pengamatan Bahan Organik ... 34
3.5.3 Pengamatan pH Tanah ... 35
3.6 Analisis Data ... 35
IV. HASIL DAN PEMBAHASAN ………...…………... 37
4.1 Hasil Penelitian ... 37
4.1.1 Kepadatan Relatif Symphylid ... 37
4.1.2 Faktor Abiotik Ekosistem ... 38
4.1.2.1 Berat Serasah ... 38
4.1.2.2 pH Tanah ... 39
iii DAFTAR TABEL
1. Hubungan habitat dengan keberadaan symphylid. ... 5
2. Spesies symphylid yang memiliki kepentingan ekonomis. ... 9
3. Tipe pertahanan akar terhadap HPA, contoh dan mekanismenya. 24
4. Tempat pelaksanaan penelitian. ... 27
5. Waktu pelaksanaa penelitian. ... 28
6. Hubungan antara faktor abiotik dengan kepadatan symphylid pada ekosistem jambu biji. ... 41
7. Hubungan antara faktor abiotik dengan kepadatan symphylid pada ekosistem nanas. ... 42
8 Hubungan antara faktor abiotik dengan kepadatan symphylid pada ekosistem pisang. ... 43
9 Hubungan antara faktor abiotik dengan kepadatan symphylid pada ekosistem lidahbuaya. ... 45
10. Data hasil pengamatan kepadatan symphylid. ... 62
11. Data hasil pengamatan berat serasah. ... 62
iv
13. Analisis ragam kepadatan symphylid. ... 64
14. Analisis ragam berat serasah. ... 64
15. Analisis ragam pH tanah. ... 65
16. Uji lanjut BNT α=0,05. ... 65
v DAFTAR GAMBAR
1. Diagram kerangka berfikir penelitian. ... 6
2. Gejala serangan hama symphylid. ... 10
3. Salah satu spesies hama symphylid (Cephalostigmata: Scutigerellidae). ... 12
4. Symphylid dewasa. ... 13
5. Embelan-embelan kepala symphylid. ... 14
6. Tergum khas beberapa genus symphylid. ... 15
7. Fase hidup symphylid. ... 18
8 Komponen pengendalian alami yang bertaut kepadatan dan yang bebas kepadatan. ... 21
9 Pengaruh kondisi lingkungan abiotik tanah terhadap interaksi akar- HPA. ... 22
10. Lokasi penelitian. ... 28
11 Posisi titik sampel dalam plot tanaman. ... 30
12 Titik sampel yang tersusun dari subtitik sampel berbentuk trapesium. ... 31
13 Langkah pembuatan umpan jebakan (bait trap) symphylid. ... 32
14 Grafik kepadatan relatif symphylid. ... 38
15. Grafik berat serasah. ... 39
vi 17. Grafik regresi hubungan faktor abiotik dengan kepadatan symphylid
pada ekosistem jambu biji. ... 41
18. Grafik regresi hubungan faktor abiotik dengan kepadatan symphylid pada ekosistem nanas. ... 43
19. Grafik regresi hubungan faktor abiotik dengan kepadatan symphylid pada ekosistem pisang. ... 44
20. Grafik regresi hubungan pH tanah dengan kepadatan symphylid pada ekosistem lidahbuaya. ... 45
21. Pengaruh kondisi lingkungan abiotik tanah terhadap interaksi akar-Symphylid HPA. ... 49
22. Penggalian dengan sekop bait trapping. ... 67
23. Bait trap dipendam menyisakan tali. ... 67
24. Ekosistem lidahbuaya. ... 67
25. Ekosistem pisang. ... 68
26. Ekosistem nanas. ... 68
I. PENDAHULUAN
1.1 Latarbelakang dan Masalah
Symphylid merupakan salah satu golongan arthropoda yang sebagian dikenali
sebagai perombak bahan organik dan sebagian lainnya dikenal sebagai hama
(herbivora pemakan akar, HPA). Symphylid telah banyak diketahui menjadi
hama yang tidak hanya menyerang sayuran dan buah-buahan (Berry & Robinson,
1974; Morais & Silva, 2009) yang ditanam di kebun tetapi juga di rumah kaca
(Edwards, 1990; Singh, 1991). Karena itu hama ini dikenal sebagai hama rumah
kaca (Singh, 1991) namun karena seringkali juga turut terbawa tanah ia juga
dikenal sebagai parasit polifagus terbawa tanah (Alphonsine et al., 2011; Soler et
al., 2011).
Pada dasarnya arthropoda ini memiliki kepentingan ekonomis (economic
importance) yang rendah (Morais dan Silva, 2009). Namun spesies Scutigerella
immaculata (Newport) di beberapa negara justru dilaporkan menjadi hama serius
pada pertanian. Bahkan symphylid dilaporkan telah menimbulkan kerugian di
perkebunan sayuran milik UCSC (University of California at Santa Cruz) pada
2
Keberadaannya yang tersebar luas di negara-negara Amerika, Eropa dan Asia
membuat symphylid dikenali sebagai arthropoda kosmopolitan. Laporan terkait
dengan keberadaan symphylid di Indonesia (Jawa & Sumatera) sendiri telah lebih
awal dilaporkan oleh Pocock pada 1894 &1897 kemudian Jupeau pada 1957 dan
Scheller pada 1988 yang menemukan spesies baru symphylid dari Pulau Krakatau
dan Semenanjung Ujung Kulon (Joseph, 2001; Scheller, 1988; Hansen, 1903).
Laporan selanjutnya terkait keberadaan symphylid tidak hanya dalam kerangka
khazanah zoologi tetapi juga marak terkait dengan peranannya sebagai hama
tanaman. Pada tahun 1966 Beeler melaporkan symphylid merusak tanaman
jagung di Indiana (Koontz, 1968). Rao pada 1959 dan David pada 1978
melaporkan bahwa symphylid dapat mengakibatkan kematian tanaman tebu
berkisar 30–40% (Singh, 1991). Symphylid Hanseniella sp., dilaporkan telah
menyerang akar padi (Loureiro & Galvao, 1970 dalam Morais & Silva, 2009) dan
nanas (Loureiro & Fortes, 1972 dalam Morais & Silva, 2009) di Brazil tepatnya di
Minas Gerais. Kerusakan tanaman nanas juga dilaporkan di Bahia (Sanches, 1981
dalam Morais & Silva, 2009). Belakangan dilaporkan hama ini menyerang salah
satu perkebunan nanas di Indonesia tepatnya di PT Great Giant Pineapple (PT
GGP) (Rusydi et al., 2012).
Terkait dengan keberadaan dan tingkat serangannya, symphylid terhitung unik.
Symphylid justru menjadi hama pada pertanian dengan menejemen tanah yang
baik yakni perawatan tanah dengan pengolahan yang baik, kandungan bahan
organik yang tinggi, dan kompaksi tanah yang rendah (Umble et al., 2006).
3
GGP justru meningkatnya populasi dan kepadatan populasi symphylid (Ningrum
et al., 2014).
PT GGP sendiri tengah mengembangkan penelitian symphylid ke arah gulma
inang potensial dari symphylid. Sejauh ini diketahui bahwa kepadatan symphylid
dapat ditemukan di daerah perakaran gulma Lantana camara dan Paspalum
conjugatum. Pada sekitar 53,33% sampel survai kepadatan symphylid pada
gulma L. camara dapat ditemukan symphylid rata-rata 9,75 individu/gulma.
Sedangkan pada gulma P. conjugatum ditemukan pada sekitar 46,67% sampel
dengan populasi rata-rata 8,57 individu/gulma. Proyek survai ini masih dalam
pengembangan untuk mengetahui gulma mayor lain yang dimungkinkan sebagai
inang symphylid (Rusydi et al., 2012).
Selain membudidayakan tanaman nanas, PT GGP juga membudidayakan jambu
biji (Psidium guajava), pisang (Musa sp) dan lidahbuaya (Aloe vera). Jenis
tanaman yang dibudidayakan dalam konsepsi agroekosistem akan membentuk
ekosistem sendiri (Oka, 2005) sehingga pertanaman nanas, jambu biji, pisang dan
lidahbuaya dapat disebut sebagai ekosistem nanas, ekosistem jambu biji,
ekosistem pisang dan ekosistem lidahbuaya. Untuk mendapatkan informasi yang
komprehensif tentang symphylid di PT GGP maka perlu diketahui keberadaan
4
1.2 Tujuan
Adapun tujuan dari penelitian ini yakni:
1. Untuk mengetahui kepadatan symphylid pada ekosistem nanas, lidahbuaya,
pisang, dan jambu biji.
2. Mengetahui hubungan antara kepadatan symphylid dan berat serasah dan pH
tanah di ekosistem nanas, lidahbuaya, pisang, dan jambu biji.
1.3 Kerangka Pemikiran
Budidaya tanaman sehat dan kuat menjadi bagian dari program pengendalian
hama. Dalam pengelolaan hama terpadu (PHT) setiap usaha budidaya tanaman
mulai dari pemilihan varietas, pegolahan tanah, penyiapan bibit dan pembibitan,
penanaman, pemeliharaan tanaman, sampai ke penanganan pasca panen perlu
dikelola secara tepat sehingga diperoleh pertanaman sehat, kuat dan produktif
(Untung, 2006). Berkaitan dengan pengolahan tanah yang baik sebagai sub unit
budidaya tanaman sehat tersebut, hama symphylid justru kerap kali menyerang.
Perawatan tanah dengan pengolahan yang baik, kandungan bahan organik yang
tinggi, dan kompaksi tanah yang rendah (Umble et al., 2006) menjadikan tanah
tersebut cocok sebagai habitat hama ini.
Terkait dengan habitat yang sesuai bagi symphylid, Edwards (1958) kembali
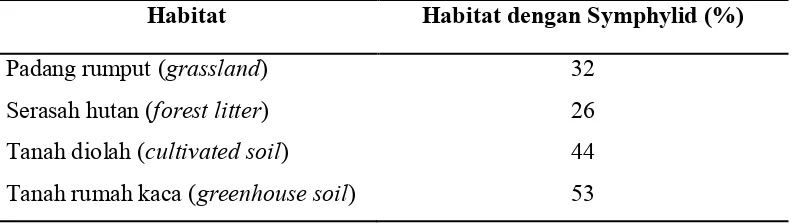
menegaskan bahwa tipe pengolahan tanah akan membuat meningkatnya
kepadatan symphylid. Dari 415 tempat yang dijadikan sampel, tipe tanah
kultivasi menunjukkan kecenderungan keberadaan symphylid yang tinggi.
5
habitat padang rumput, symphylid dapat ditemukan pada lebih dari 30 % sampel
yang dipasang. Pada habitat serasah hutan symphylid justru terhitung jarang
ditemukan (26%). Symphylid paling mudah ditemukan pada habitat tanah rumah
kaca (53 %) (Tabel 1).
Tabel 1. Hubungan habitat dengan keberadaan symphylid.
Habitat Habitat dengan Symphylid (%)
Padang rumput (grassland) 32
Serasah hutan (forest litter) 26
Tanah diolah (cultivated soil) 44 Tanah rumah kaca (greenhouse soil) 53 Sumber : Edwards (1958).
Dalam lingkup yang lebih luas habitat tanah yang diolah mengarah pada
ekosistem pertanian itu sendiri. Pada dasarnya informasi hubungan jenis
ekosistem pertanian yang terbentuk dengan kepadatan symphylid mengarah pada
hubungan hama dengan inangnya. Dalam konsep pengelolaan ekosistem
pertanian pemutusan rantai makanan pada hama (melalui usaha rotasi tanaman )
adalah hal yang penting. Hal ini terkait erat dengan fokus pengelolaan ekosistem
pertanian yakni menemukan titik lemah (weak link) pada perkembangan hama
kemudian memutusnya (Pedigo, 1989 dalam Hasibuan, 2003). Namun upaya
rotasi ini hanya efektif untuk mengendalikan hama yang memiliki kisaran inang
6
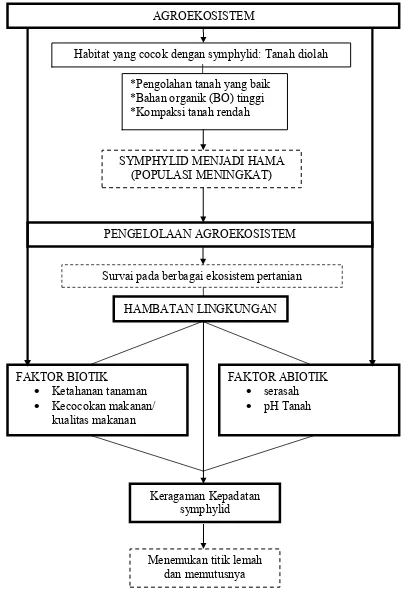
Gambar 1. Diagram kerangka berpikir penelitian. *Pengolahan tanah yang baik
*Bahan organik (BO) tinggi *Kompaksi tanah rendah
SYMPHYLID MENJADI HAMA (POPULASI MENINGKAT)
PENGELOLAAN AGROEKOSISTEM
Menemukan titik lemah dan memutusnya AGROEKOSISTEM
Habitat yang cocok dengan symphylid: Tanah diolah
HAMBATAN LINGKUNGAN
FAKTOR BIOTIK
Ketahanan tanaman Kecocokan makanan/
kualitas makanan
FAKTOR ABIOTIK serasah pH Tanah
Keragaman Kepadatan symphylid
7
Walaupun symphylid memakan tanaman dengan kisaran yang luas dan juga dapat
persisten dalam tanah gundul (bare soil) dengan memangsa organisme tanah lain
tetapi tanaman pasti memiliki variasi yang sangat beragam dalam kerentanannya
terhadap serangan symphylid (Umble et al., 2006). Symphylid sendiri memiliki
kecocokan yang beragam dengan sumber makanan yang ia dapatkan
(Michelbacher, 1938). Pada penelitian skala laboratorium, Shanks pada 1966 dan
Ramsey pada 1969 menemukan bahwa kepadatan symphylid tumbuh lebih baik
pada daun letus (Latuca sativa L.), dan akar wortel (Daucus carota L.)
dibandingkan akar kacang buncis (Phaseolus vulgaris L.), alfalfa (Medicago
sativa L.), barley (Astrebla pectinata (Lindl.) F. Muell.), rye (Secale cereale L.),
dan jagung manis (Zea mays L.) (Umble & Fisher, 2003b).
Perbedaan jenis tanaman selain memiliki ketahanan yang beragam terhadap
serangan symphylid juga merupakan faktor biotik yang akan berinteraksi dengan
faktor abiotik, dalam hal ini pH tanah dan bahan organik (berat serasah),
membentuk karakteristik tersendiri dalam ekosistem. Karakteristik ekosistem ini
kemudian dapat menjadi hambatan lingkungan yang akan mempengaruhi
beragamnya kepadatan symphylid itu sendiri (Gambar 1).
1.4Hipotesis
Berdasarkan kerangka pikir yang telah diurai di atas, disusunlah hipotesis yakni
kepadatan symphylid bervariasi menurut jenis ekosistem pertanian (ekosistem
II. TINJAUAN PUSTAKA
2.1 Symphylid sebagai Hama
Pada awalnya symphylid memiliki kepentingan ekonomis (economic important)
yang rendah (Morais dan Silva, 2009), tetapi pada abad awal ke 20 kepentingan
ekonomis dari arthopoda ini semakin banyak dirasakan dan dicatat dalam banyak
publikasi (Joseph, 2001). Beberapa publikasi yang menunjukkan kepentingan
ekonomis dari hewan ini misalnya oleh Brade-Birks pada 1929 (Joseph, 2001),
Filinger (1931), Michelbacher (1938), Edwards (1958, 1959a, 1961), Morrison
(1961), Swenson (1965), Koontz (1968), Berry & Robinson (1974), Pai &
Prabhoo (1991), Umble et al. (2006), dan Rusydi et al. (2012).
Umumnya spesies symphylid yang dikenali sebagai hama hanya terfokus pada
Scutigerella immaculata, namun sebenarnya terdapat cukup banyak spesies yang
turut menyebabkan kerusakan pada berbagai tanaman yang telah tercatat.
Beberapa spesies yang tercatat merusak tanaman dengan memakan akar mudanya
9
Tabel 2. Spesies symphylid yang memiliki kepentingan ekonomis.
Tanaman Spesies Sumber
Nanas Scutigerella immaculata
Umble et al. (2006)
S. sakimurai Sakimura (1966) dalam Soler et al. (2011)
Hanseniella unguiculata
Sakimura (1966) dalam Soler et al. (2011)
H. ivorensis Juberthie-Jupeau & Kehe (1978) dalam Soler et
al. (2011)
H. colombiana Juberthie-Jupeau (1997) dalam Soler et al.
(2011)
Hanseniella sp. Loureiro & Fortes (1972) dalam Morais & Silva
(2009)
Symphylella tennela Rohrbach & Johnson (2003)
Kubis S. immaculata Joseph (2001); Filinger (1931)
Tomat S. immaculata Joseph (2001); Filinger (1931); Michelbacher (1938)
S. lineatus Joseph (2001)
Letus S.immaculata Joseph (2001); Filinger (1931)
S. lineatus Joseph (2001)
Strawberi S.immaculata Joseph (2001); Berry & Robinson (1974) Anyelir S.immaculata Joseph, 2001; Michelbacher, 1938
Padi Hanseniella sp. Loureiro & Galvao (1970) dalam Morais & Silva (2009)
Jagung Spesies symphylid S.immaculata
Beeler (1966) dalam Koontz (1968) ; Michelbacher (1938)
Sawit H. caldaria Joseph (2001) Tebu H. unichaetosa Joseph (2001) Beet H. agilis Joseph (2001) Jambu H. unichaetosa Joseph (2001)
2.2 Gejala kerusakan
Pada dasarnya serangan symphylid mengakibatkan buruknya kerja sistem akar
sehingga pertumbuhan terganggu (Gambar 2D). Pada serangan berat, serangan
hama ini dapat mengakibatkan kematian tanaman (Ghidiu, 2005). Gejala yang
ditimbulkan oleh symphylid kerap kali membingungkan, lantaran gejala yang
ditunjukkan mirip dengan gejala tanaman yang diserang nematoda atau gejala
10
warna menjadi kuning atau merah (Gambar 2B). Selain itu pada bagian akar,
nampak akar tanaman terpotong, rambut akar sangat minim, dan massa akar yang
rendah (Gambar 2C). Hal ini disebabkan lantaran hama symphylid menyerang
tanaman dengan memakan akar dan rambut akar, bahkan juga menyerang
benih-benih sayuran yang ditanam (Ghidiu, 2005).
Gambar 2. Gejala serangan hama symphylid. (A). Lahan pertanaman nanas sehat, (B). Tanaman nanas terserang symphylid, (C). Akar tanaman nanas yang terserang symphylid, (D). Tanaman terung terserang symphylid dan (E). Tanaman terung sehat. [Sumber: (A–B). Rusydi et al., 2012; (D–E) Umble et al., 2006].
(A)
(D)
(C) (B)
11
2.3 Karakteristik Symphylid
2.3.1 Taksonomi Hama Symphylid
Symphylid digolongkan ke dalam kelas Symphyla. Kelas Symphyla sendiri
merupakan bagian dari filum Arthropoda subfilum Atelocerata (Myriapoda).
Kelas ini hanya memiliki 1 (satu) ordo yakni Cephalostigmata Verhoeff, 1934
dan terdiri dari sekitar 160 spesies symphylid (Umble & Fisher, 2003a; Ghidiu,
2005; Burden, 2008).
Selain hanya memiliki satu ordo, kelas ini juga hanya terdiri dari dua famili yakni
Scutigerellidae dan Scolopendrellidae (Scheller, 1961). Mayoritas publikasi
terfokus pada dua genus yang kerap kali merusak yakni Scutigerella dan
Hanseniella (Camacho, 2009). Semua spesies hama dari kedua genus tersebut
tergolong ke dalam famili Scutigerellidae (Gambar 3). Secara sederhana
Australian Faunal Directory (2008) menyusun klasifikasi hama ini seperti
berikut:
Kingdom : Animalia
Filum : Arthropoda
Subfilum : Myriapoda
Kelas : Symphyla
Ordo : Cephalostigmata
Superfamili : Scutigerelloidea
12
Gambar 3. Salah satu spesies hama symphylid (Cephalostigmata: Scutigerellidae).
2.3.2 Morfologi Eksternal Symphylid
Secara morfologi symphylid menyerupai kelabang yang juga tinggal di dalam
tanah (Carr, 2003) karenanya dahulu Michelbacher (1938) menyebutnya sebagai
“garden centipede”. Namun berbeda dengan kelabang symphylid berwarna putih,
dan berukuran kecil dengan panjang tubuh sekitar 3–7 mm. Ia hanya memiliki
11–12 pasang kaki pada saat dewasa (Gambar 4) (Carr, 2003; Edwards, 1990;
Berry & Robinson, 1974).
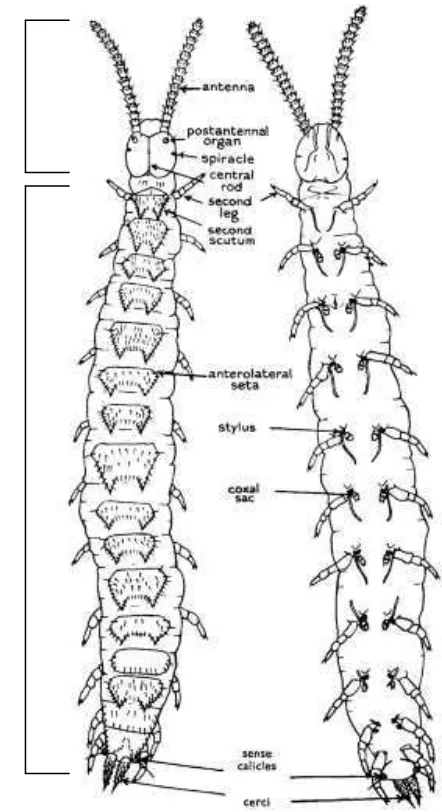
Tubuh symphylid terdiri atas dua tagmata yakni kepala dan tubuh (trunk)
(Gambar 4). Kepala memiliki beberapa embelan seperti antena dan alat mulut.
Tubuh symphylid bersegmen dan memiliki embelan berupa tungkai, stilus, sense
13
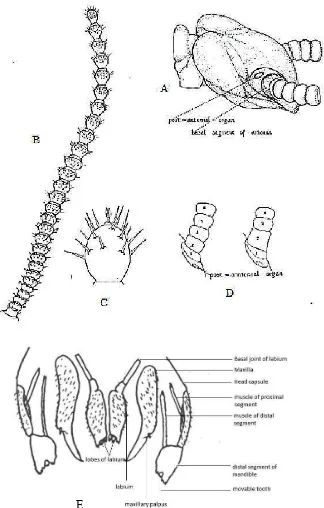
Gambar 4. Symphylid dewasa (Famili Scolopendrellidae). A. tampak dorsal, B. tampak ventral. (Sumber : Edwards, 1959b).
Kepala symphylid berbentuk seperti hati dan terdapat antena serta embelan alat
mulut. Tidak terdapat mata pada kepala symphylid tetapi terdapat organ
posantenal organ pada basal setiap antenna (Gambar 5A ). Terdapat satu pasang
stigma (bercak) di bawah organ posantenal. Antenna yang dimiliki panjang dan
bersambung dengan organ indera pada ujungnya (Gambar 5B–C) dan dapat terdiri
lebih dari 60 segmen antenna (Edwards, 1990). Symphylid memiliki alat mulut
menggigit-mengunyah atau bertipe mandibuta (Edwards, 1990; Filinger, 1931). Kepala
14
Struktur alat mulut symphylid terdiri dari labium, hipofaring, maksila dan
mandibel (Szucsich et al., 2011; Filinger, 1931) (Gambar 5E).
Gambar 5. Embelan-embelan pada kepala symphylid. (A–D). Bagian–bagian antena. (E). Bagian-bagian alat mulut. [Sumber : (A–D) Michelbacher, 1938; (E) Filinger, 1931)].
15
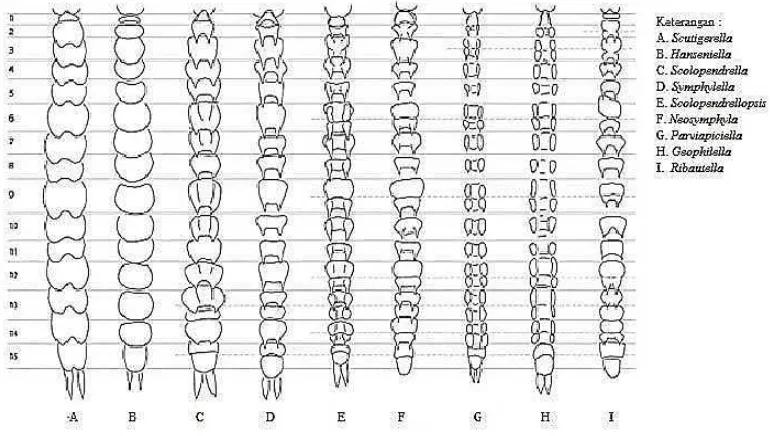
Badan (trunk) symphylid dewasa terdiri dari 12 segmen yang terlihat jelas dan
telson posterior. Pada permukaan dorsalnya terdapat 15 hingga 22 dorsal skuta
dengan ukuran dan pola yang berbeda pada setiap genus (Gambar 6). Tubuhnya
terdiri dari 15 segmen tubuh dan terdapat cerci seperti spin di bagian posteriornya
(Edwards, 1990).
Gambar 6. Tergum khas beberapa genus symphylid (Sumber: Camacho, 2009).
2.3.3 Siklus Hidup
Siklus hidup symphylid dimulai dari fase telur kemudian pradewasa (nimfa) dan
fase dewasa. Fase telur symphylid berakhir setelah menetas pada kisaran 7–20
hari. Setelah menetas symphylid memasuki fase pra dewasa (nimfa instar 1)
kemudian nimfa terus mengalami molting (ganti kulit) dengan setiap kali molting
membutuhkan waktu yang beragam. Setidaknya symphylid membutuhkan 6 kali
16
molting bahkan tercatat dapat mencapai 50 kali (Michelbacher, 1938). Umumnya
symphylid membutuhkan waktu sekitar 90 hari untuk menjadi dewasa, hanya saja
waktu ini dapat dipengaruhi oleh temperatur lingkungan (Capinera, 2001).
1.3.3.1Telur
Secara morfologi telur symphylid terlihat unik, selain berukuran sangat kecil
dengan diameter 0,5 mm (Filinger, 1931) telur symphylid seperti mutiara putih
dan bulat dengan punggung berbentuk heksagonal (Gambar 7A). Warna telur
yang putih seperti mutiara tersebut berangsur berubah menjadi gelap sebelum
akhirnya menetas (Edwards, 1990).
Umumnya telur diletakkan secara berkelompok seolah menjadi satu paket. Setiap
paket dapat terdiri dari 9–25 butir telur. Paket telur ini selalu dilindungi oleh
induk betinanya. Karena apabila ditinggalkan telur rentan terserang jamur dan
membuat telur tidak menetas. Pada titik ini menunjukkan bahwa symphylid
dewasa juga memakan jamur (Edwards, 1990; Joseph, 2001).
Telur memiliki masa inkubasi yang beragam bergantung pada suhu lingkungan.
Masa inkubasi telur pada suhu berkisar antara 50° – 70°F sekitar 25 – 40 hari,
tetapi penetasan muncul pada kurang lebih 12 hari saat temperatur mencapai 77°F
17
2.3.3.2 Pradewasa
Fase pradewasa dimulai setelah telur symphylid menetas dan memasuki fase
nimfa instar pertama. Instar pertama muncul dari telur dengan enam (tujuh)
pasang kaki dan enam segmen antennal, tubuh mereka ditutupi dengan bulu–bulu
halus (Gambar 7A). Pada fase ini panjangnya sekitar 0,75 mm (Filinger, 1931).
Gerakannya lambat dan posterior yang membengkak membuat instar pertama kali
sepintas lebih seperti Collembola daripada symphylid dewasa. Instar pertama
dalam beberapa hari kemudian akan berganti kulit ke instar kedua. Pada instar ini
symphylid telah menyerupai symphylid dewasa (Gambar 7C) (Michelbacher,
1938).
2.3.3.3 Dewasa
Fase dewasa symphylid terjadi ketika symphylid memasuki instar ke 7 dengan 6
kali proses ganti kulit. Setiap kali dari enam kali pergantian kulit akan
menghasilkan sepasang tungkai dan beberapa segmen tubuh juga segmen antenna.
Total waktu dari telur hingga dewasa yang matang secara seksual (instar ke tujuh)
adalah sekitar lima bulan pada 50°F, menurun menjadi sekitar tiga bulan pada
70°F dan kurang dari dua bulan pada 77°F. Oleh karena itu, dimungkinkan untuk
memiliki dua generasi lengkap setahun (Berry, 1972 dalam Umble et al., 2006).
Menariknya, tidak seperti serangga dewasa, yang tidak berganti kulit, symphylid
18
Gambar 7. Fase hidup symphylid. (A). Fase telur, (B). Fase Pradewasa, dan (C). Fase Dewasa.
2.4 Pengendalian Hama Symphylid
2.4.1 Pengendalian Hayati Hama Symphylid
Perkembangan informasi terkait pengendalian hayati symphylid diawali dengan
penemukan predator symphylid oleh Menge tahun 1851. Ia mencatat bahwa
centipede sejati telah mempredasi “garden centipede”. Kemudian Fillinger,
Wymore, Savor, dan Waterhouse berturut-turut pada 1828 & 1931, 1931, 1958,
dan 1969 melaporkan observasi yang serupa pada total 7 spesies centipede
(Koontz, 1968; Filinger, 1931; Waterhouse, 1969).
Selanjutnya perkembangan eksplorasi musuh alami symphylid terus berkembang.
Wymore pada 1931 melaporkan tungau Gasamid memangsa symphylid.
Illingsworth pada 1927–1928 menyatakan bahwa dua spesies dari kumbang telah
diketahui memangsa symphylid. Savos pada 1958 melaporkan bahwa millipedes
dan Collembola memangsa symphylid mati dan telurnya (Koontz, 1968).
Michelbacher (1938) menerangkan bahwa studi terkait musuh alami dari “garden
centipede” telah terabaikan. Khususnya para parasit sejati dan penyakit dari
symphylid itu sendiri. Hingga Getzin pada 1963 menemukan Entompophthora
19
coronata (Constantin) Kevorkian membunuh Symphylan dalam kaleng-kaleng
tanah. Getzin dan Shanks pada 1964 melaporkan bahwa kultur jamur
Entompophthora coronata (Constantin) Kevorkian dan Metarhizium anisopliae
(Metschnikoff) Sorokin yang diintroduksikan ke dalam kaleng–kaleng tanah yang
sebelumnya diinfestasi symphylid dan juga pada lahan uji coba dapat bertahan
hidup dan menginfeksi symphylid dalam satu atau dua bulan (Koontz, 1968).
Setelah itu Swenson (1965) berhasil menemukan bahwa nematoda patogenik
DD-136 dapat menginfeksi S. immaculata.
Penggunaan agensia hayati berupa predator symphylid, jamur patogen, atau
nematoda (Soler et al., 2011) dapat dijadikan teknik pengendalian hayati hama ini.
Namun demikian memang masih sedikit informasi yang diketahui terkait
kemampuan agensia hayati tersebut dalam mengendalikan kepadatan symphylid
(Carr, 2003).
2.4.2 Pengendalian Kimiawi
Berbagai senyawa kimia telah dicoba untuk menengendalikan symphylid dalam
100 tahun terakhir (Howitt & Bullock, 1955 dalam Umble et al., 2006). Sejauh
ini, fumigan dan pestisida organofosfat menjadi jenis pestisida yang dianggap
efektif (Umble et al., 2006). Morrison (1961) menyebutkan pestisida fumigan
yang potensial misalnya Vorlex dan Dichloropropane-dichloropropene mixture .
Rusydi et al. (2012) merekomendasikan menggunakan pestisida berbahan aktif
20
2.5. Populasi Hama dan Faktor yang Mempengaruhinya
Naik turunnya populasi organisme dalam hal ini hama ditentukan oleh dua
kekuatan ekosistem yaitu kemampuan hayati atau potensi biotik dan hambatan
lingkungan (Untung, 2006). Potensi biotik merupakan kemampuan organisme
untuk berkembangbiak dalam kondisi yang normal. Potensi biotik suatu jenis
organisme di alam selalu akan dipengaruhi oleh faktor biotik (misalnya, makanan
dan pasangan) dan faktor abiotik atau lingkungan fisik (misalnya, temperatur,
kelembaban dan intensitas cahaya). Berbagai faktor biotik dan abiotik di
ekosistem yang cenderung menurunkan fertilitas dan kelangsungan hidup
individu-individu dalam populasi organisme dikenal dengan istilah hambatan
lingkungan (Untung, 2006) atau ketahanan lingkungan (environmental resistance)
(Sembel, 2012).
Terdapat beberapa teori yang muncul untuk menjelaskan faktor apakah yang
menentukan kepadatan rata-rata suatu spesies dalam ekosistem, dalam hal ini
symphylid. Pertama, teori biologis, yang menyatakan bahwa di alam terdapat
faktor-faktor yang bertaut kepadatan (density dependent) yang juga menghalangi
kenaikan populasi dan menentukan kepadatan rata-rata pada banyak spesies
populasi (Oka, 2005). Faktor-faktor yang dimaksud adalah musuh alami dari
symphylid. Secara teoritis, musuh alami tersebut merupakan faktor yang juga
memiliki hubungan timbal balik dan bertaut kepadatan dengan populasi hama
(Untung, 2006). Artinya, populasi musuh alami akan meningkat ketika populasi
21
Teori kedua yakni teori iklim, yang berpendapat bahwa faktor fisik atau abiotik
sebagai faktor yang mengendalikan kepadatan suatu populasi. Faktor ini
merupakan pengendali yang tidak bertaut kepadatan (Oka, 2005). Artinya, daya
penekanannya tidak dipengaruhi oleh kepadatan symphylid.
Teori selanjutnya yakni teori komprehensif, teori ini yang paling banyak
digunakan. Bahwa semua faktor baik yang bertaut kepadatan maupun bebas
kepadatan mempengaruhi kepadatan populasi. Faktor biotik maupun abiotik yang
kompleks yang berbeda dalam ruang dan waktu membuat perubahan atau
perbedaan populasi (Oka, 2005). Secara skematik pengendalian alami hama
dalam teori ini dapat digambarkan sebagai berikut (Gambar 8):
Gambar 8. Komponen pengendalian alami yang bertaut kepadatan dan yang bebas kepadatan. (Sumber : dimodifikasi dari Untung, 2006)
Secara teoritis terdapat indikasi yang kuat pengaruh parameter abiotik pada
keberadaan suatu hama pemakan akar (HPA) (Johnson et al., 2011; Barnett &
Johnson, 2013). Umumnya, faktor abiotik yang Symphylid sebagai HPA meliputi
struktur dan tekstur tanah, kelembaban tanah, suhu tanah, CO2 di tanah, pH tanah,
22
Kondisi Abiotik
2.Tidak Langsung
serta pemupukan (dengan pupuk kandang) (Barnett & Johnson, 2013; Edwards,
1961; Michelbacher, 1938).
2.6 Interaksi Herbivora Pemakan Akar, Akar Tanaman dan Faktor Abiotik Tanah
Interaksi herbivora pemakan akar (symphylid), akar tanaman dan faktor abiotik
tanah dapat dijelaskan terkait 3 hal (Gambar 9.A). Pertama, lingkungan abiotik
tanah dapat secara langsung mempengaruhi kemelimpahan dan distribusi HPA.
Edwards (1961) menemukan bahwa temperatur dan kelembaban mempengaruhi
distribusi vertikal symphylid. Kedua, faktor abiotik dapat mempengaruhi secara
tidak langsung symphylid. Hal ini disebabkan oleh perubahan fisiologi dan
metabolisme akar mengikuti tekanan faktor abiotik (Gao et al., 2007 dalam Erb &
Lu, 2013) yang akan mengubah kualitas makanan dan kapasitas pertahanan akar.
Terakhir, faktor abiotik juga dapat mempengaruhi lingkungan biotik dimana
tanaman dan HPA bertemu (Fierer & Jackson, 2006 dalam Erb & Lu, 2013),
termasuk, misalnya, musuh alami yang dapat mengurangi serangan symphylid.
23
Lebih lanjut, Erb & Lu (2013) menjelaskan bahwa kondisi abiotik dapat
mempengaruhi interaksi tanaman dan lingkungan secara kuantitatif (Gambar 9B).
Secara umum kondisi yang ekstrim akan mengurangi HPA (symphylid) dan
kesehatan tanaman, termasuk organisme yang spesifik hidup beradaptasi di bawah
kondisi tersebut. Pada banyak kasus, kondisi abiotik berfluktuasi pada level
dalam rentang kompensasi fisiologi dari keduanya (tanaman dan symphylid).
Dengan demikian perubahan faktor abiotik mempengaruhi dengan kuat hasil
interaksi tanaman dengan symphylid.
Selain itu terkait interaksi HPA dengan akar tanaman, penelitian satu dekade
terakhir oleh para ahli telah berhasil mengungkap bahwa serangan HPA dapat
menginduksi pertahanan akar. Tipe pertahaan yang terbentuk dapat digolongkan
ke dalam beberapa tipikal yakni tipe langsung, tipe tidak langsung, tipe toleransi
dan tipe pengelakan. Toleransi akar terhadap HPA ditunjukkan dengan
pertumbuhan yang lebih atau tumbuh kembalinya jaringan yang hilang (rusak)
atau menolak dengan cara memproduksi metabolit sekunder yang beracun
terhadap HPA atau menarik musuh alami HPA tersebut (Rasmann et al., 2011).
24
Tabel 3. Tipe pertahanan akar terhadap HPA, contoh dan mekanismenya.
Tipe Pertahanan Contoh Mekanisme
Toleransi Tumbuh kembali Realokasi sumber daya dari jaringan lain Kompensasi
berlebih
Peningkatan metabolisme utama tanaman setelah dirusak HPA
Langsung Pertahanan kimiawi Pengeluaran metabolit sekunder dalam konsentrasi tinggi pada area yang terluka Pertahanan
struktural
Pengerasan jaringan akar
Nutrisional N (C : N ratio)
Tidak langsung Interaksi tritrofik Rekrutmen NEP oleh emisi volatil dari area yang terluka
Menyimpan nutrisi yang kecil pada jaringan terserang
Sumber : Rasmann et al. (2011).
Pada interaksi tritrofik (tiga trofik: tumbuhan, hama, dan musuh alami), peranan
biota tanah lain (musuh alami symphylid) yang mungkin terjadi dan membentuk
interaksi pihak ketiga (tripartite) terhadap akar maupun symphylid yang juga
dipengaruhi oleh faktor abiotik. Pola pengaruh ini dikenal dengan istilah efek
tripartit (tripartite effects) yang terjadi secara langsung maupun tidak langsung.
Pengaruh secara langsung terhadap symphylid dilakukan mikroorganisme sebagai
patogen. Sedangkan secara tidak langsung, mikroorganisme dapat mempengaruhi
pertahanan tanaman dengan berperan sebagai mutualis bermanfaat (Zamioudis &
Pieterse, 2012) atau sebagai antagonis (Millet et al., 2010). Misalnya Glomus
mosseae diketahui meningkatkan resistensi tanaman Taraxacum officinale dan
Glomus etunicatum meningkatkan resistensi tanaman Glycine max terhadap
serangan hama pemakan akar (Gange et al., 1994; Borowicz, 1997). Ketahanan
25
mode kegiatan memakan dan derajat spesialisasi dari herbivora (Koricheva et al.,
2009).
Secara umum, ketika HPA dalam hal ini symphylid menyerang, tanaman mulai
memproduksi senyawa organik volatil yang menyebar melalui tanah dan dapat
digunakan oleh nematoda entomopatogenik (NEP) untuk menemukan lokasi
mangsanya (Rasmann et al., 2005; Ali et al., 2010; Turlings et al., 2012; Hiltpold
et al., 2013). Nematoda yang umum dikenali sebagai NEP tergolong ke dalam
Famili Heterorhabditidae dan Steinernematidae (Purnomo, 2010). Sejauh ini
telah dilaporkan spesies nematoda Heterorahabditis marealatus, Steinernema
feltiae, S.carpocapsae terbukti dapat menimbulkan mortalitas pada symphylid
hingga 75% (Brown et al., 2001). Terkait dengan jenis senyawa volatil yang
dapat mengundang NEP, Hiltpold et al. (2010) menemukan bahwa S. fertiae
tertarik dengan senyawa (E)-β-caryophyllene (EβC) yang dikeluarkan oleh akar
tanaman jagung yang terserang HPA. Kemampuan pencarian mangsa dari NEP
sendiri umumnya dipengaruhi arsitektur akar tanaman (Demarta et al., 2014).
Beberapa jenis NEP dapat ditemukan keberadaannya di pertanaman pisang
(Mwaitulo et al., 2011).
Namun demikian pola interaksi tritrofik yang melibatkan akar dan NEP sebagai
agensia hayati HPA (symphylid) tetap dipengaruhi oleh faktor abiotik. Menurut
Erb & Lu (2013) setidaknya pengaruhnya dapat dijelaskan dalam 3 cara, yakni :
1. Induksibilitas dari sinyal volatil dapat dipengaruhi oleh ketersediaan
nutrisi (Ibrahim et al., 2008) dan status air (Gouinguene & Turling, 2002
26
2. Difusi volatil melalui tanah dipengarahui oleh kelembaban tanah,
kelembaban yang terlalu tinggi atau terlalu rendah tidak cocok dalam
proses difusi ini (Hiltpold & Turlings, 2008).
3. Aktivitas pergerakan NEP dipengaruhi oleh struktur tanah (Schroeder &
Beavers, 1987 dalam Erb & Lu, 2013) dan kelembaban (Grant & Villani,
III. BAHAN DAN METODE
3.1 Tempat dan Waktu Penelitian
Penelitian ini dilaksanakan di dua tempat sesuai dengan tahapan penelitian yang telah disinggung di atas. Tahap pertama dilaksanakan di PT Great Giant
Pineapple, Kec. Terbanggi Besar, Kab. Lampung Tengah, Lampung (Gambar 10). Sedangkan tahap kedua dilakukan di laboratorium Jurusan Agroteknologi,
Fakultas Pertanian, Universitas Lampung, Kota Bandarlampung, Lampung (Tabel 4).
Tabel 4. Tempat pelaksanaan penelitian.
Tahap Penelitian
Kegiatan Tempat Pelaksanaan
Pertama Survai ekosistem nanas Plantation Group (PG)1 (Lok. 030 C)
Survai ekosistem lidahbuaya PG 1 (Lokasi 036) Survai ekosistem pisang PG 1 (Lokasi 055) Survai ekosistem jambu biji PG (Lokasi 042A)
Kedua Penghitungan berat serasah Laboratorium Hama
28
Gambar 10. Lokasi penelitian.
Kegiatan penelitian ini terdiri dari 2 tahap berlangsung pada bulan Mei–Juni 2015. Tahap pertama merupakan tahap survai di lapangan yang meliputi kegiatan
pemasangan bait trap symphylid dan pengamatan (ekstraksi symphylid) serta pengambilan sampel tanah. Tahap kedua merupakan tahap lanjutan yang meliputi kegiatan penghitungan berat serasah, dan pH tanah (Tabel 5).
Tabel 5. Waktu pelaksanaan penelitian.
Tahap
Penelitian Kegiatan
Waktu Pelaksanaan
Mei Juni
1 2 3 4 1 2 3 4
Pertama Pemasangan 12
Pemasangan+ Ekstraksi 15 Pemasangan+ Ekstraksi 19 Pemasangan+ Ekstraksi 21
Ekstraksi 25
Kedua Penghitungan berat serasah 6
29
3.2 Bahan dan Alat
Bahan-bahan yang digunakan pada penelitian ini digolongkan berdasarkan tahapan penelitian. Bahan yang digunakan pada tahap pertama meliputi tanah, daun pepaya dan alkohol. Sedangkan bahan pada tahap kedua merupakan bahan analisis pH tanah. Bahan yang digunakan pada analisis pH tanah meliputi air destilata dan sampel tanah.
Alat yang digunakan pada penelitian ini terdiri dari dua kelompok alat, juga berdasarkan tahapan penelitian. Kelompok alat pertama merupakan alat survai symphylid meliputi alat sekop bait trapping symphylid, nampan plastik, sendok, jaring, golok atau pisau, tali rafia, kertas label dan alat tulis. Sedangkan
kelompok alat kedua merupakan alat-alat analisis tanah. Alat analisis tanah sendiri dikelompokkan kembali berdasarkan kegiatan analisis. Pada penghitungan berat serasah digunakan ayakan tanah dengan ukuran besar (besek, wadah nasi dengan lubang-lubang persegi disekeliling sisinya) dan ayakan berukuran 1mm, timbangan digital Scaltec SPO 61, sendok, palu mortar, plastik, nampan, spidol permanen. Analisis pH tanah dilakukan dengan menggunakan alat meliputi cup
plastik, dan pH meter Jenway 3520.
3.3Metode Penelitian
30
pertanaman nanas, pisang, jambu biji,dan lidahbuaya. Setiap ekosistem terletak pada lokasi berbeda (Gambar 10), masing-masing seluas 5 sampai dengan 10 ha.
Gambar 11. Posisi titik sampel dalam plot tanaman.
Dari setiap lokasi ekosistem dipilih empat plot yang berdekatan (Plot 1 sampai dengan Plot 4) yang berbentuk persegi panjang dan berukuran 0,2 sampai dengan 0,8 ha. Dari setiap plot itu diambil satu titik sampel yang letaknya di sudut diagonal plot (Gambar 11) dan dari setiap titik sampel ini ditentukan lima subtitik sampel pada posisi trapesium (Gambar 12). Pada setiap titik sampel dilakukan pengambilan tiga data secara bersamaan, yaitu (1) kepadatan symphylid, (2) berat serasah, dan (3) pH tanah. Setiap kali pengambilan data itu dilakukan di setiap titik sampel di empat ekosistem sekaligus pada hari dan tanggal yang sama (Tabel 5). Ukuran 5 bait trap dipilih untuk setiap plot karena merupakan ukuran contoh minimal yang dapat digunakan dalam pendugaan sederhana keberadaan
31
Gambar 12. Titik sampel yang tersusun dari subtitik sampel berbentuk trapesium.
3.4Pelaksanaan Penelitian
Pelaksanaan penelitian ini terbagi menjadi dua tahap penelitian. Tahap pertama merupakan tahapan survai di lapang yang terdiri dari kegiatan pembuatan dan pemasangan bait trap symphylid, kegiatan ekstraksi symphylid, dan pengambilan sampel tanah. Tahap kedua merupakan tahapan analisis tanah yang terdiri dari kegiatan penghitungan berat serasah (pengayakan sampel tanah), dan analisis pH tanah.
3.4.1 SurvaiSymphylid di Lapangan
3.4.1.1Pembuatan dan Pemasangan Bait Trap Symphylid
32
dicampur dengan tanah sebanyak 700 g kemudian dimasukkan ke dalam jaring dan diikat dengan tali rafia (Gambar 13).
Pemasangan umpan dilakukan dengan dipendam di dalam galian tanah sedalam 20–30 cm yang sebelumnya digali menggunakan skop bait trapping. Setelah umpan dipendam 4 hari, umpan kemudian diambil dan dipindahkan secara hati-hati ke dalam nampan untuk ekstraksi symphylid.
33
3.4.1.2Ekstraksi Symphylid
Metode ekstraksi symphylid yang digunakan yakni hand sorting method atau metode sortir langsung dengan tangan. Ekstraksi langsung ini dilakukan dengan meletakkan tanah umpan yang diambil dari titik sampel ke dalam nampan berwarna cerah dan kemudian secara perlahan dan hati-hati diamati dan dihitung kepadatan symphylid yang terperangkap dengan bantuan sendok. Setelah itu memasukkan symphylid ke dalam botol berisi alkohol.
3.4.2 Analisis Tanah di Laboratorium
3.4.2.1 Penghitungan Berat Serasah
34
3.4.3.2 Analisis pH Tanah
Pengukuran pH tanah dilakukan dengan menggunakan pH meter Jenway 3520. Analisis pH tanah ini menggunakan 5g sampel tanah masing-masing ulangan yang dimasukkan ke dalam cup plastik kemudian dicampur dengan air destilata
sebanyak 12,5 ml. Kemudian diaduk hingga beberapa kali kemudian setelah 30 menit kemudian diaduk kembali dan dimasukkan elektroda pada alat pH meter kemudian ditunggu hingga angka pada pH meter berhenti bergerak (Thom & Utomo, 1991).
3.5 Teknik Pengamatan dan Pengumpulan Data
3.5.1 Pengamatan Kepadatan Relatif Symphylid
Pengamatan dilakukan pada variabel ini meliputi jumlah symphylid yang berhasil tertangkap dengan menggunakan bait trap yang dipasang pada pertanaman nanas, lidahbuaya, pisang dan jambu biji. Pengamatan ini dilakukan pada setiap hari ke 4 setelah pemasangan bait trap pada setiap ulangan. Kepadatan symphylid yang dihitung mencakup symphylid dari berbagai stadia (telur, pradewasa atau dewasa) dan kondisi (mati atau hidup).
3.5.2 Pengamatan Bahan Organik
35
dikompositkan kemudian diambil sekitar ½ kg dan kemudian dipisahkan tanah dari serasah bahan organik dengan menyaring tanah dengan saringan berukuran 1mm. Serasah yang berhasil dipisahkan kemudian ditimbang bobotnya pada setiap ulangan.
3.5.3 Pengamatan pH Tanah
Pengamatan pH tanah dilakukan dengan menganalisis sampel tanah yang telah dipisahkan dengan serasah. Analisis pH tanah dilakukan dengan menggunakan alat pH meter Jenway 3520 dengan prosedur yang telah diurai sebelumnya.
3.6. Analisis Data
Data variabel pengamatan pada penelitian ini dianalisis secara statistik dengan perangkat Microsoft Excel. Beberapa variabel pengamatan yang meliputi data kepadatan relatif symphylid, berat serasah, dan pH tanah diuji dengan analisis ragam (ANARA) dengan taraf nyata α = 0,05. Kemudian apabila hasil ANARA signifikan maka dilanjutkan dengan uji lanjut BNT dengan taraf nyata 0,05.
Selanjutnya untuk mengetahui hubungan masing-masing faktor abiotik dengan kepadatan symphylid pada berbagai ekosistem yang diteliti dilakukan analisis regresi dan korelasi. Analisis regresi linier dilakukan dengan terlebih dahulu melakukan ANARA regresi pada taraf nyata α = 0,05. Setelah itu dihitung persamaan regresi dan koefisien determinasi (regresi) R2. Nilai R2
36
V. KESIMPULAN DAN SARAN
5.1 Kesimpulan
Berdasarkan hasil penelitian ini dapat disimpulkan bahwa:
1. Kepadatan symphylid tertinggi terdapat pada ekosistem jambu biji, diikuti ekosistem nanas, pisang dan lidahbuaya.
2. Kepadatan symphylid dipengaruhi oleh faktor abiotik berupa berat serasah dan/atau pH tanah.
3. Pada ekosistem nanas kepadatan symphylid dipengaruhi oleh berat serasah. 4. Pada ekosistem lidahbuaya kepadatan symphylid dipengaruhi oleh pH tanah. 5. Pada ekosistem jambu biji dan pisang kepadatan symphylid dipengaruhi oleh
berat serasah dan pH tanah.
5.2 Saran
Adapun saran yang dapat diajukan yakni
1. Meningkatkan jumlah sampel pada penelitian selanutnya untuk mengetahui kepadatan symphylid secara lebih akurat
53
3. Perlu dilakukan penelitian lanjutan menggali manfaat-manfaat tanaman lidahbuaya sebagai tanaman perotasi atau tanaman sisipan.
4. Menginventarisasi keanekaragaman symphylid pada berbegai ekosistem. 5. Melakukan uji daya makan symphylid terhadap akar berbagai tanaman,
PUSTAKA ACUAN
Ali, J.G., H.T. Alborn, & L.L. Stelinski. 2010. Subterranean herbivore-induced volatiles released by citrus root upon feeding by Diaprepes abbreviatus recruit entomophatogenic nematodes. Journal of Chemical Ecology. 36 : 361–368.
Alphonsine, P.A.M., P. Fourmier, B. Dole, J.C. Govidin, P. Quénéhervé, & A. Soler. 2011. A bait and trap method for sampling symphylid
population in pineapple. In: Proceedings of 7th International Pineapple Symposium. Abdullah, H., D.P. Bartholomew, & M.N. Latifah. (Eds). ISHS Acta Horticurturae. 902. p 357–362.
Australian Faunal Directory (AFD). 2008. Species Scutigerella immaculata (Newport, 1845): garden symphylid. Australian Biological Resources Study. Australian Government. Departement of the Environment. http: //www. environment.gov.au/biodiversity/abrs/online resources/fauna/afd/ taxa/Scutigerella_immaculata. Diakses pada 8 Oktober 2015.
Barnett, K & S.N. Johnson. 2013. Living in the soil matrix : abiotic factors affecting root herbivores. In: Advances in Insect Physiology- Behavior and Physiology of Root Herbivores Volume 45. Johnson, S.N., I. Hiltpold, & T.J. Turling. (Eds). Academic Press. London. p 1–52.
Berry, R.E. & R.R. Robinson. 1974. Biology and control of the garden symphylans. In : Extention Circular 845. Oregon State University. Extention Service. Corvallis. p 1–9.
Borowicz, V.A. 1997. A fungal root symbiont modifies plant resistance to an insect herbivore. Oecologia. 112: 534–542.
55
Burden, B. 2008. Symphylans (Class Symphyla). In : Encyclopedia of entomology 2nd edition. Capinera, J.L (Ed). Springer. Heidelberg. p 3667–3668.
Camacho, M.D. 2009. Phylogeny of the Symphyla (Myriapoda). (Disertation). Department of Biology, Chemistry and Pharmacy of Freie Universitat Berlin. Madrid.
Capinera, J.L. 2001. Handbook of Vegetable Pests. Academic Press. San Diego.
Carr, P. 2003. Garden symphylid (Scutigerella immaculata). Departement of Primary Industries VegCheque extention Program. Knoxfield.
Coloma, A.G., M. Reina, C.E. Diaz, & B.M. Fraga. 2010. Natural product-based biopesticide for insect control. In: Comprehensive Natural Products II : Chemistry & Biology. Vol 3. Development & modification. Mander, L. & H-W.(Ben) Liu (Eds. in Cheif). Elsevier Ltd. London.p 237–268.
Demarta, L., B.E. Hibbard, M.O. Bohn, I. Hiltpold. 2014. The role of root architecture in foraging behavior of entomopathogenic nematodes. Journal of Invertebrate Pathology. 20: 155–162.
Edwards, C.A. 1990. Symphyla. In : Soil biology guide. Dindal, D.L. (Ed). A Wiley-Interscience Publication. John Wiley & Sons. New York.
.1958. The ecology of symphyla Part I. Population. Entomologia Experimentalis et Applicata. 1 : 308–319.
. 1959a. The ecology of symphyla Part II. Seasonal soil migrations. Entomologia Experimentalis et Applicata. 2 : 257–267.
. 1959b. Keys to the genera of the symphyla. Journal of Linnean Society of London (Zoology). 44 : 164–169.
. 1961. The ecology of symphyla Part III. Factors controlling soil distributions. Entomologia Experimentalis et Applicata. 4 : 239–256.
Erb, M. & J. Lu. 2013. Soil abiotic factors influence interaction between belowground herbivores and plant roots. Journal of Experimental Botany. 64 (5): 1295–1303.
Filinger, G.A. 1931. The Garden symphylid, Scutigerella immaculata Newport. Ohio Agricultural Experimen Station Bulletin. 486:1–33.
Ghidiu, G.M. 2005. Garden Symphylans. Fact Sheet 234 NJ Agricultural
56
Gange, A.C., V.K. Brown, & G.S. Sinclair. 1994. Reduction of black vine weevil larval growth by vesicular-arbuscular mycorrhizal infection.
Entomologia Experimentalis et Applicata. 70: 115–119.
Grant, J.A & M.G. Villani. 2003. Soil moisture effect on entomopathogenic nematodes. Population Ecology. 32 (1) : 80–87.
Gregorich, E.G., L.W. Turchenek, M.R. Carter, & D.A. Angers. 2001. Soil and Environmental Science Dictionary. CRC Press. Boca Raton.
Hansen, H.J. 1903. The genera and species of the order Symphyla. Quarterly Journal of Microscopical Science. 47 : 1–101.
Hasibuan, R. 2003. Pengendalian Hama Terpadu. Penerbit Universitas Lampung. Bandarlampung.
Hiltpold, I., E. Bernklau, L.B. Bjostad, N. Alvarez, N.E.M. Struttmann, J.G. Lundgren, & B.E. Hibbard. 2013. Nature, evolution and
characterisation of rhizosperic chemical exudates affecting root
herbivores. In: Advances in Insect Physiology- Behavior and Physiology of Root Herbivores Volume 45. Johnson, S.N., I. Hiltpold, & T.J. Turling. (Eds). Academic Press. London. p 97–157.
Hiltpold, I., S. Toepfer, U. Kuhlmann, & T.C.J. Turling. 2010. How maize root volatiles effect the efficacy of entomopathogenic nematodes in
controlling the western corn rootworm?. Chemoecology. 20 : 155-162.
Hiltpold, I., & T.C.J. Turlings. 2008. Belowground chemical signaling in maize: when simplicity rhymes with efficiency. Journal of Chemical Ecology. 34: 628–635.
Ibrahim, M.A., A. Stewart-Jones, J. Pulkkinen, G.M. Poppy, & J.K. Holopainen, 2008. The influence of different nutrient levels on insect-induced plant volatile in Bt and conventional oilseed rape plants. Plant Biology. 10: 97–107.
Johnson, S.N., A.T. Barton, K.E. Clark, P.J. Gregory, L.S. McMenemy, & R.D. Hancock. 2011. Elevated atmospheric carbon dioxide impairs the performance of root feeding vine weevils by modifying root growth and secondary metabolites. Global Change Biology. 17 : 688–695.
Joseph, S.K. 2001. Some Aspect of The Ecology and Biology of Symphyla. (Thesis). Departement of Zoology, N.S.S. Reserach Centre. N.S.S. College. Changanacherry.
57
symphylan, Scutigerella immaculata (Newport). (PhD Disertation). Oregon State University. Corvallis.
Koricheva, J., A.C. Gange, & T. Jones. 2009. Effect of mycorrhizal fungi on insect herbivore: A Meta-Analysis. Ecology.90 (8) : 2088–2097.
Michelbacher, A.E. 1938. The biology of the garden centipede Scutigerella immaculata. Hilgardia 11:55–148.
Millet, Y.A., C.H. Danna, N.K. Clay, W. Songnuan, M.D. Simon, D. Werck-Reicchart, & F.M. Ausubel. 2010. Innate imune responses activated in arabidopsis roots by microbe-associated molecular patterns. The Plant Cell. 22: 973–990.
Morais, J.W. de & E.P. da Silva. 2009. Occurence of Symphyla (Myriapoda) in region of the Upper Solimones River, Amazonas, Brazil. Pesquisa Agropecuária Brasileira. 44 (8): 981–983.
Morrison, H.E. 1961. Subsoiling aids symphylid control. Oregon State University. Oregon Vegetable Digest. X (4): 1–3.
Mwaitulo, S., S. Haukeland, M.-G. Saethre, A. Laudisoit, & A.P. Maerere. 2011. First report of entomopathogenic nematodes from Tanzania and their virulence against larvae and adults of the banana weevil Cosmopolites sordidus (Coleoptera: Curculionidae). International Journal of Tropical Insect Science. 31(3) : 154–161.
Ningrum, Z., B.T. Rahardjo, & H. Tarno. 2014. Kepadatan kepadatan symphylid pada berbagai kompos di pertanaman nanas (Ananas comosus L., Merr) PT Great Giant Pineapple. Jurnal Hama dan Penyakit Tumbuhan. 2 (3): 1–8.
Oka, I.N. 2005. Pengendalian Hama Terpadu dan Implemantasinya di Indonesia. Gadjah Mada University Press. Yogyakarta.
Pai, C.G.A., & N.R. Prabhoo. 1991. Effect of forest fire on symphylid population in the Ponmudi Hill of Western Ghats. In: Advance in Management and Conservation of Soil Fauna. Veeresh, G.K., D. Rajagopal, & C.A. Viraktamath. (Eds). Oxford & IBH Publsihing Co.Pvt.Ltd. New Delhi. p I.27–I.39
Purnomo, H. 2010. Pengantar Pengendalian Hayati. Penerbit ANDI. Yogyakarta.
58
Rasmann, S., T.L. Bauerie, K. Poveda, & R. Vannette. 2011. Predicting root defence against herbivores during succession. Functional Ecology. 25 : 368–379.
Rohrbach, K.G. & M.W. Johnson. 2003. Pest, diseases and weeds. In: The Pineapple: Botany, Production and Uses. D.P. Bartholomew, R.E. Paull, & K.G. Rohrbach. (Eds). CABI Publishing. New York.
Rusydi, N.E., M. Basuki, & Purwito. 2012. Symphylids control in pineapple field in Indonesia. In: Pineapple News. Newsletter of Pineapple Works Group. International Society for Horticulture Science. Issue 19. p 39–41.
Scheller, U. 1961. Studies on the symphylid fauna of the Hawaiian island. Proceedings, Hawaiian Entomological Society.17 (3): 443–456.
. 1988. Two new species of Symphyla from the Krakatau islands and the Ujung Kulon peninsula (Myriapoda: Symphyla: Scolopendrellidae, Scutigerellidae). Philosophical Transactions of the Royal Society of London Biological Sciences. 322: 401–411.
Sembel, D.T. 2012. Dasar-dasar Perlindungan Tanaman. Penerbit ANDI. Yogyakarta.
Seydel, C. 1999. Researcher seek organic solution to symphylan problem. The Center for Agroecology & Sustainable Food Systems (CASFS). The Cultivar. 17 (2): 4–5.
Singh, J. 1991. Progress in soil zoology in India. In: Advance in management and conservation of soil fauna. Veeresh, G.K., D. Rajagopal, & C.A. Viraktamath. (Eds). Oxford & IBH Publsihing Co.Pvt.Ltd. New Delhi. p I.27–I.39
Soler, A., J.M. Gaude, P.A.M. Alphonsine, F. Vinatier, B. Dole, J.C. Govidin, P. Fournier, & P. Queneherve. 2011. Development and evaluation of new method for sampling and monitoring the symphylid population in pineapple. Pest Management Science. 67: 1169–1177.
Susilo, F.X. 2013. Buku Ajar. Aplikasi Statistika untuk Analisis Data Riset Proteksi Tanaman. Anugrah Utama Raharja (AURA). Bandarlampung.
Swenson, K.G. 1965. Infection of garden symphylan, Scutigerella immaculata, with The DD-136 Nematode. Journal of Invetebrate Pathology. 8:133– 134.
59
Thom, W.O, & M. Utomo. 1991. Manajemen Laboratorium dan Metode Analisis Tanah dan Tanaman. Penerbit Universitas Lampung. Bandarlampung.
Turling, T.C.J., I. Hiltpold, & S. Rasmann. 2012. The importance of root-produces volatile as foraging cues for entomopathogenic nematodes. Plant Soil. 358 : 51–60.
Umble, J., R.Dufour, G. Fisher, J. Fisher, J. Leap, & M.V. Horn. 2006.
Symphylans: Soil pest management options. ATTRA. National Center for Appropriate Technology (NCAT). Butte.
Umble, J. R., & J.R . Fisher. 2003a. Influence of below-ground feeding by garden symphylans (Cephalostigmata: Scutigerellidae) on plant health.
Environmental Entomology. 32 (5): 1251–1261.
. 2003b. Suitability of selected crop and soil for garden symphylan population (Symphyla, Scutigerellidae: Scutigerella immaculata Newport). Applied Soil Ecology 24 : 151–163.
Untung, K. 2006. Pengantar Pengelolaan Hama Terpadu Edisi ke 2 (revisi). Gadjah Mada University Press. Yogyakarta.
Waterhouse, J.S. 1969. An evaluation of a new predaceous centipede Lamyctes sp. on the garden symphylan Scutigerella immaculata. The Canadian Entomologist. 101 : 1081–1083.
Wuyts, N., G. Lognay, L. Sagi, D. De Waele. & R. Swennen. 2005. Secondary metabolites in roots and implications for nematodes resistance in banana (Musa spp.). In: Banana Root System: Toward Better Understanding for Its Productive Management. Proceeding of International Symposium. Turner, D.W., & F. E. Rosales (Eds). International Network for The Improvement of Banana & Plantain. Montpellier. p 238–246.