EFISIENSI WERENG HIJAU DAN WERENG BATANG
COKELAT SEBAGAI VEKTOR VIRUS
PADA TANAMAN PADI
AMELIA FERYNA BULAN DINI
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA
Dengan ini saya menyatakan bahwa tesis berjudul Efisiensi Wereng Hijau dan Wereng Batang Cokelat sebagai Vektor Virus pada Tanaman Padi adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam daftar pustaka di bagian akhir tesis ini.
Dengan ini saya melimpahkan hak cipta karya tulis saya kepada Institut Pertanian Bogor.
Bogor, Oktober 2015
RINGKASAN
AMELIA FERYNA BULAN DINI. Efisiensi Wereng Hijau dan Wereng Batang Cokelat sebagai Vektor Virus pada Tanaman Padi. Dibimbing oleh SRI HENDRASTUTI HIDAYAT dan I WAYAN WINASA.
Penyakit virus pada padi merupakan masalah penting dalam produksi beras di Indonesia. Kehilangan hasil yang disebabkan oleh penyakit tungro dan penyakit kerdil umumnya dilaporkan sejak masa tanam. Penyakit tungro disebabkan oleh infeksi ganda virus, Rice tungro bacilliform virus (RTBV) dan Rice tungro spherical virus (RTSV); sementara penyakit kerdil disebabkan oleh Rice grassy stunt virus (RGSV) atau Rice ragged stunt virus (RRSV). Vektor yang paling efisien dalam menularkan penyakit virus padi telah diketahui, di antaranya wereng hijau, Nephotettix virescens (Distant), untuk penyakit tungro dan wereng batang cokelat, Nilaparvata lugens (Stal.), untuk penyakit kerdil.
Penyakit tungro dan kerdil sudah menyebar di Provinsi Jawa Barat yang merupakan salah satu pemasok padi terbesar secara nasional. Diagnosis penyakit tidak dapat mengandalkan hanya dari gejala saja karena gejala yang muncul beranekaragam serta mirip dengan gejala kekurangan hara dan kekeringan sehingga perlu dilakukan diagnosis secara molekuler. Penelitian ini dilakukan dengan tujuan menentukan efisiensi wereng hijau dan wereng batang cokelat dalam menularkan penyakit tungro dan kerdil ke tanaman padi. Selain itu juga dilakukan deteksi virus dengan metode polymerase chain reaction (PCR) untuk mengonfirmasi gejala penyakit tungro dan penyakit kerdil.
Penelitian meliputi tiga kegiatan, yaitu (1) pengamatan populasi wereng hijau dan wereng batang cokelat, dan insidensi penyakit tungro dan penyakit kerdil di lapangan; (2) percobaan penularan virus dengan vektor serangga di rumah kaca menggunakan padi varietas IR 64; dan (3) deteksi virus dengan metode PCR dan/atau RT-PCR dan analisis sikuen nukleotida. Pengamatan lapangan dilakukan di Desa Sukamandi Kabupaten Subang Provinsi Jawa Barat pada bulan Oktober - November 2014. Percobaan penularan virus dengan vektor serangga terdiri atas 10 kombinasi perlakuan, yaitu 1 (sumber virus tungro, serangga vektor wereng hijau); 2 (sumber virus kerdil, serangga vektor wereng batang cokelat); 3 (sumber virus tungro, serangga vektor wereng hijau dan wereng batang cokelat); 4 (sumber virus kerdil, serangga vektor wereng hijau dan wereng batang cokelat); 5 (sumber virus tungro dan kerdil, serangga vektor wereng hijau dan wereng batang cokelat); 6 (sumber virus tungro dan kerdil, serangga vektor wereng hijau); 7 (sumber virus tungro dan kerdil, serangga vektor wereng batang cokelat); 8 (tanaman sehat serangga vektor wereng hijau dan wereng batang cokelat); 9 (tanaman sehat, serangga vektor wereng hijau); 10 (tanaman sehat, serangga vektor wereng batang cokelat). Deteksi RTBV, RGSV, dan RRSV dilakukan menggunakan primer spesifik yang mengamplifikasi gen protein selubung.
hampa, kerdil rumput dan campuran kerdil hampa dan kerdil rumput. Fluktuasi populasi terjadi karena proses adaptasi setelah migrasi dari lahan sebelumnya. Wereng yang mampu beradaptasi akan bertahan dan berkembangbiak, sedangkan wereng yang tidak mampu beradaptasi pada tempat baru akan mati. Keberadaan musuh alami dan beberapa faktor lingkungan juga mempengaruhi populasi wereng. Pengamatan populasi wereng dilakukan pada masa tanam di musim penghujan, Daerah Sukamandi merupakan sentra padi dimana padi ditanam secara terus menerus tanpa adanya rotasi tanaman dan penanaman dilakukan secara tidak serempak sehingga tanaman padi selalu ada sebagai makanan wereng.
Percobaan penularan menunjukkan wereng batang cokelat lebih efisien menularkan RRSV daripada wereng hijau menularkan virus tungro, yaitu ditandai dengan periode inkubasi yang lebih singkat dan insidensi penyakit yang lebih tinggi. Periode inkubasi RRSV pada padi varietas IR 64 adalah 7 hari, dengan insidensi penyakit 100%, dan keparahan penyakit tertinggi 97.03%. Periode inkubasi virus tungro adalah 7 – 14 hari, dengan insidensi penyakit berkisar dari 76.67% sampai 100 %, dan keparahan penyakit tertinggi 94.06%. Gejala penyakit yang muncul ditentukan oleh kombinasi perlakuan. Perlakuan wereng hijau dengan sumber inokulum virus tungro menghasilkan gejala tungro, perlakuan wereng batang cokelat dengan sumber inokulum virus kerdil menghasilkan gejala kerdil hampa, perlakuan wereng hijau dan wereng batang cokelat dengan sumber inokulum virus tungro dan virus kerdil menghasilkan gejala campuran tungro dan kerdil hampa.
Deteksi virus dari sampel tanaman dari lapangan tidak berhasil mengamplifikasi virus tungro, tetapi berhasil mengamplifikasi gen protein selubung RRSV menggunakan primer RRSV-S9-F/RRSV-S9-R pada ± 445 pb, dan gen protein selubung RGSV menggunakan primer RGSV-S3-F/ RGSV-S3-R pada ± 750 pb. Hal ini menunjukkan bahwa tanaman padi di Sukamandi tidak terinfeksi oleh virus tungro, tetapi ditemukan infeksi virus kerdil rumput dan kerdil hampa. Metode deteksi dengan PCR juga berhasil mengonfirmasi infeksi virus dari sampel tanaman hasil percobaan penularan. Pita DNA berukuran ± 1400 pb berhasil diamplifikasi menggunakan primer spesifik virus tungro RTBV2L/ RTBV2R dari sampel tanaman bergejala tungro. Pita DNA spesifik RRSV juga berhasil diamplifikasi dari sampel tanaman bergejala kerdil, sedangkan pita DNA spesifik RGSV tidak teramplifikasi. Analisis sikuen nukleotida menunjukkan bahwa RRSV dan RGSV isolat Sukamandi tersebut memiliki homologi tertinggi berturut-turut dengan isolat Vietnam, Filipina dan Thailand (97.1 %); dan isolat Longan, Vietnam (95.8 %). Sikuen isolat RTBV asal Indonesia (Purwakarta) memiliki homologi tertinggi dengan isolat Chainat dari Thailand (85.7 %). Analisis sikuen gen protein selubung RTBV, RGSV, dan RRSV menunjukkan adanya homologi yang tinggi antara virus-virus padi di Asia Tenggara.
Spesifikasi spesies wereng menularkan virus padi telah dibuktikan pada penelitian ini. Wereng hijau hanya menularkan virus tungro, sedangkan wereng batang cokelat hanya menularkan virus kerdil hampa. Efisiensi wereng batang cokelat menularkan virus kerdil lebih tinggi dibandingkan wereng hijau menularkan virus tungro.
SUMMARY
growing season. Tungro disease is caused by infection of two viruses, Rice tungro bacilliform virus (RTBV) and Rice tungro spherical virus (RTSV); whereas stunt diseases is caused by Rice grassy stunt virus (RGSV) or Rice ragged stunt virus (RRSV). The most efficient vectors for viral diseases on rice has been known, i.e. green leafhopper, Nephotettix virescens (Distant), for tungro disease and brown planthopper, Nilaparvata lugens (Stal.), for stunt diseases.Tungro and stunt diseases has spread in West Java province, one of the main rice growing area. Diagnosis of diseases could not rely only on symptoms due to similar symptoms caused by nutrient deficiencies and drought. Therefore, laboratory test using molecular approach should be done to confirm virus infection. This research was aimed to evaluate the efficiency of green leafhopper (N. virenscens) and brown planthopper (N. lugens) as vectors of tungro and stunt diseases, respectively. Detection method based on polymerase chain reaction (PCR)was also carried outto confirm tungro and stunt diseases symptoms.
Three activities was conducted during the research , i.e. (1) field observation of leaf hopper and brown planthopper populations, and tungro and stunt diseases incidence; (2) greenhouse experiment to study virus tramsmission by insect vector using IR 64 rice varieties; and (3) Detection of virus infection using PCR and/or RT-PCR method, followed by nucleotide sequence analysis. Field observation was carried out in Sukamandi Village, West java Province in October – November 2014. Virus transmission experiment consisted of 10 treatments combination, i.e. 1 (tungro virus, with green leafhopper insect vector); 2 (stunt virus with brown planthopper insect vector); 3 (tungro virus with green leafhopper and brown planthopper insect vectors); 4 (stunt virus with green leafhopper and brown planthopper insect vectors); 5 (tungro and stunt virus with green leafhopper and brown planthopper insect vectors); 6 (tungro and stunt virus with green leafhopper insect vectors); 7 (tungro and stunt virus with brown planthopper insect vectors); 8 ( healthy plants with green leafhopper and brown planthopper); 9 (healthy plants with green leafhopper); 10 (healthy plants with brown planthopper). Detection of RTBV, RGSV, and RRSV was conducted using specific primers for coat protein gene.
survive and multiply, while the population which failed to adapt in new place will die. The presence of natural enemies and several environment condition may also affect the population of brown planthoppers. The field observation was done during the rainy season. Sukamandi is paddy rice growing area, in which the paddy rice was planted continuously without crop rotation and simultaneously so that is always as brown planthopper food. Transmission experiment showed that brown planthopper transmitted RRSV more efficiently than green leafhopper transmitted rice tungro virus. Shorter incubation period (7 days), higher disease incidence and severity (100% and 97.03%, respectively) was observed for RRSV in var. IR64. Incubation period of rice tungro virus was 7 – 14 days, with disease incidence ranging from 76.67% to 100% and disease saverity 94.06%. Diseases symptoms was determined by treatment combination. Green leafhopper treatment with tungro virus caused tungro symptoms, brown planthopper treatment with stunt virus caused stunt symptoms, mix of green leafhopper and brown planthopper treatment with mix tungro and stunt virus caused mix symptoms of tungro and stunt disease . Detection of viruses from field samples was unable to amplify tungro virus, but was successfully amplified coat protein gene of RRSV and RGSV using RRSV-S9-F/ RRSV-S9-R primers and RGSV-S3-F/RGSV-S3-R primers, respectively. DNA bands of ±445 bp and ± 750 bp was amplified for RRSV dan RGSV, respectively. This result indicated that paddy rice plants in Sukamandi was not infected by tungro virus, but positively infected by RRSV and RGSV. The same detection method confirmed virus infection from plant samples collected from transmission experiment., DNA bands of ± 1400 bp was successfully amplified using spesific primers RTBV2L/ RTBV2R from plant samples showing tungro symptoms. Specific DNA bands of RRSV(450 bp) was also successfully amplified from plant showing stunt symptoms, whereas DNA bands of RGSV was not amplified. Nucleotide sequence analysis showed that RRSV and RGSV isolates from Subang has the highest homology Vietnam, Filipina and Thailand (97.1 %); and Longan, Vietnam (95.8%), respectively. The sequence of RTBV isolate from Indonesia (isolate Purwakarta) has the highest homology with Chainat isolate from Thailand (85.7 %) and the lowest with Kanyakumari isolate from India (68.9 %). This sequence analysis of coat protein gene of RTBV, RGSV, and RRSV indicated a high homology among rice viruses in Southeast Asia.
Species specification of the insect vector in transmitting rice viruses was evidenced in this study. Green leaf hopper only transmitted tungro virus, whereas brown plant hopper only transmitted ragged stunt virus. Incidence and severity of tungro disease was lower than those of stunt disease.
© Hak Cipta Milik IPB, Tahun 2015
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan mengutip tersebut tidak merugikan kepentingan IPB.
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains
pada
Program Studi Fitopatologi
EFISIENSI WERENG HIJAU DAN WERENG BATANG
COKELAT SEBAGAI VEKTOR VIRUS
PADA TANAMAN PADI
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
BOGOR
2015
PRAKATA
Alhamdulillahirrabbil’alamin, puji dan syukur penulis panjatkan kepada Allah SWT atas segala karunia-Nya sehingga penelitian ini dapat diselesaikan. Penelitian dengan judul “Efisiensi Wereng Hijau dan Wereng Batang Cokelat sebagai Vektor Virus pada Tanaman Padi” telah dilaksanakan sejak bulan Oktober 2014 sampai Mei 2015. Terima kasih penulis ucapkan kepada Prof Dr Ir Sri Hendrastuti Hidayat, MSc. dan Dr Ir I Wayan Winasa, MS selaku komisi pembimbing, Dr Bambang Nuryanto, MS. selaku pembimbing lapangan, yang telah banyak memberi dukungan, kritik serta saran kepada penulis, serta Dr Ir Kikin Hamzah Mutaqin, MSi. yang telah bersedia bertindak sebagai penguji luar komisi pada ujian tesis. Penelitian ini merupakan bagian dari Hibah Penelitian KKP3N 2013-2015 atas nama Sri Hendrastuti Hidayat yang didanai oleh Kementerian Pertanian. Ungkapan terima kasih juga disampaikan kepada kedua orang tua penulis bapak Marsolini SPd. dan ibu Nurhayana, serta seluruh keluarga, atas segala dukungan, doa dan kasih sayangnya
Penulis menyampaikan banyak terimakasih kepada pak Oco, pak Usep selaku teknisi lapang BB Padi Sukamandi dan warga desa Sukamandi yang telah banyak membantu penulis dalam menyelesaikan penelitian selama di Sukamandi. Terimakasih juga penulis ucapkan kepada Sari Nurulita SP MSi, Ni Nengah Putri Adnyani SP MP, Teguh Pamungkas SP, Kgs Angga Pratama SP, Siska Irhamnawati Pulogu SP, Fitria Yuliani SP, Novi Irawati SP, Diana Putri SSi, dan Syaiful Khoiri SP yang telah banyak membantu dan memberi masukan kepada penulis, serta seluruh anggota Laboratorium Virologi Tumbuhan IPB, Fitopatologi 2013 dan seluruh pihak yang tidak bisa disebutkan satu per satu atas segala ilmu dan kebaikan yang diberikan.
Semoga karya ilmiah ini bermanfaat.
DAFTAR ISI
DAFTAR TABEL DAFTAR GAMBAR DAFTAR LAMPIRAN
1 PENDAHULUAN 1
Latar Belakang 1
Tujuan Penelitian 2
Hipotesis Penelitian 2
Manfaat Penelitian 2
2 TINJAUAN PUSTAKA 3
Sifat Umum Tanaman Padi (Oryza sativa) 3
Karakteristik Penyakit Tungro 3
Penyakit Tungro 3
Serangga Vektor Wereng Hijau 4
Karakteristik Penyakit Kerdil 6
Penyakit Kerdil Hampa 6
Penyakit Kerdil Rumput 6
Serangga Vektor Wereng Batang Cokelat 7
3 METODE 9
Lokasi dan Waktu Penelitian 9
Pengamatan Insidensi Penyakit dan Populasi Wereng di Lapangan
9 Uji Efisiensi Penularan Wereng Hijau dan Wereng Batang
Cokelat sebagai Vektor Virus Padi
9 Perbanyakan Wereng Hijau dan Wereng Batang Cokelat 9 Perbanyakan Inokulum Virus Tungro dan Kerdil 9
Uji Penularan 10
Peubah Pengamatan 10
Deteksi Virus Penyebab Penyakit Tungro dan Kerdil 12 Deteksi RRSV dan RGSV dengan Riverse Transcription-
Polymerase Chain Reaction (RT-PCR)
12 Deteksi RTBV dengan metode Polymerase Chain Reaction
(PCR)
13
Analisis Susunan Nukleotida 14
4 HASIL DAN PEMBAHASAN 15
DAFTAR ISI (lanjutan)
Pengamatan Populasi Wereng di Lapangan 15
Pengamatan Insidensi Penyakit di Lapangan 17 Hubungan Populasi Wereng Batang Cokelat dan Insidensi
Penyakit Kerdil
19 Efisiensi Penularan Wereng Hijau dan Wereng Batang Cokelat
sebagai Vektor Virus Padi
21
Periode Inkubasi 21
Insidensi Penyakit 23
Keparahan Penyakit 23
Deteksi RRSV dan RGSV dengan metode RT-PCR dan Deteksi RTBV dengan metode PCR
24
Analisis Sekuen Nukleotida 26
5 SIMPULAN 28
DAFTAR PUSTAKA 29
LAMPIRAN 33
DAFTAR TABEL
1 Tahapan penularan virus tungro oleh wereng hijau dan kerdil oleh wereng batang cokelat
10 2 Perlakuan kombinasi virus dan serangga vektor pada uji
efisiensi penularan wereng hijau dan wereng batang cokelat sebagai vektor virus padi
11
3 Skor gejala penyakit tungro dan kerdil berdasarkan Standard Evaluation System for Rice (SESR) yang telah dimodifikasi
12 4 Periode inkubasi, insidensi penyakit dan keparahan penyakit
tungro dan kerdil pada tanaman padi yang ditularkan berturut-turut dengan wereng hijau dan wereng batang cokelat
22
5 Tingkat homologi sikuen nukliotida gen protein selubung Rice ragged stunt virus isolat Subang, Indonesia dan negara lain
26 6 Tingkat homologi sikuen nukliotida gen protein selubung Rice
grassy stunt virus isolat Indo-RG3, Indonesia dan negara lain
26 7 Tingkat homologi sikuen nukleotida gen protein selubung Rice
tungro baciliform virus isolat Purwakarta, Indonesia dan negara lain
27
DAFTAR GAMBAR
1 Gejala serangan penyakit tungro pada tanaman padi dengan gejala menguning pada daun (Dokumentasi pribadi)
4 2 Wereng hijau (Nephotettixvirescens Distant) 5 3 Gejala serangan Rice ragged stunt virus pada tanaman padi.
tepi tanaman padi menjadi bergerigi (A), ujung tanaman padi memutar (B) (Dokumentasi pribadi), Partikel RRSV dilihat melalui mikroskop elektron (C)
6
4 Gejala penyakit kerdil rumput pada rumpun tanaman padi yang menunjukkan perubahan warna daun menjadi hijau pucat hingga kuning (A) (Dokumentasi pribadi), Hasil pemurnian RGSV 2 dengan pewarnaan uranil asetat, ukuran 100nm (B) (Hibino 1985)
7
5 Imago N. lugens bersayap panjang (Makroptera) (A), Imago N. lugens bersayap pendek (Brakhiptera) (B) (Dokumentasi Pribadi)
8
6 Skema percobaan (A), Contoh arena percobaan pengujian efisiensi wereng hijau dan wereng batang cokelat sebagai vektor virus padi dengan menggunakan ember (B)
11
7 Rata-rata populasi wereng batang cokelat pada petak pengamatan di Sukamandi pada periode pengamatan November 2014-Desember 2014
15
8 Wereng batang cokelat stadia imago makroptera (A), wereng batang cokelat stadia nimfa (B), dan imago brakhiptera (C)
9 Rata-rata insidensi penyakit kerdil pada petak pengamatan di Sukamandi pada periode pengamatan November – Desember 2014
17
10 Tanaman padi yang terserang penyakit kerdil diantara tanaman sehat. Gejala penyakit kerdil rumput pada umur tanaman 5 mst (A), gejala penyakit kerdil hampa pada umur tanaman 10 mst (B), gejala penyakit kerdil rumput dan kerdil hampa pada tanaman singgang (C)
18
11 Penyakit kerdil hampa dengan gejala permukaan daun menjadi kasar karena adanya garis-garis melintang yang terbentuk (A),tepi daun terlihat bergelombang (B), adanya sobekan-sobekan pada daun (C), ujung daun menggulung (D), terdapat pada bagian bawah helai daun (E)
19
12 Daun tanaman padi yang terserang penyakit kerdil rumput. Daun mengalami perubahan warna menguning dan perubahan ukuran daun yang menjadi lebih sempit dari daun sehat (A), terdapat bintik-bintik menyerupai karat pada bagian daun (B)
19
13 Hubungan populasi wereng batang cokelat dengan insidensi penyakit kerdil tanaman padi pada musim tanam Oktober-November 2014
21
14 Gejala menguning pada daun tanaman padi yang terinfeksi virus Tungro (A), gejala malformasi pada daun tanaman padi yang terinfeksi virus kerdil (B), gejala tanaman padi yang terinfeksi oleh virus Tungro dan virus Kerdil (C)
22
15 Visualisasi pita DNA hasil amplifikasi menggunakan pasangan primer RRSV-S9-F/RRSV-S9-R (1), RGSV-S3-F/
16 Visualisasi pita DNA hasil amplifikasi menggunakan pasangan primer RRSV-S9-F/RRSV-S9-R (1), RGSV-S3-F/
1 Perkembangan insidensi penyakit tungro (T) untuk perlakuan 1, 3, 5, 6 dan perkembangan insidensi penyakit kerdil (K) untuk perlakuan 2, 4, 5, 7 serta perlakuan 8, 9, 10 sebagai kontrol sehat tanpa sumber inokulum (S).
33
2 Perkembangan keparahan penyakit tungro yang ditunjukkan dengan tanda T dan penyakit kerdil yang ditunjukkan dengan tanda K pada setiap perlakuan, dengan tanda S untuk perlakuan 8,9,10 yang merupakan tanaman sehat dengan serangga vektor.
3 Variasi ketinggian sampel tanaman untuk percobaan efisiensi penularan virus tungro yang ditularkan oleh wereh hijau dan virus kerdil yang ditularkan oleh wereng coklat untuk masing-masing perlakuan (1-10)
34
4 Penghambatan pembentukan bulir padi pada umur 8 mst pada tanaman padi sehat (A), tanaman padi terinfeksi penyakit kerdil (B), tanaman padi terinfeksi penyakit tungro (C), dan tanaman padi terinfeksi penyakit kerdil dan tungro (D).
35
5 Visualisasi pita DNA hasil amplifikasi menggunakan pasangan primer RRSV-S9-F/RRSV-S9-R (1), RGSV-S3-F/ RGSV-S3-R (2), dan RTBV2L/RTBV2R (3). (M) penanda DNA 1 kb ladder, (A) sumber inokulum virus kerdil, (B) sumber inokulum virus tungro
35
6 Visualisasi pita DNA hasil amplifikasi menggunakan pasangan primer RGSV-S3-F/RGSV-S3-R. Sampel no. 1 – 10, berasal dari masing-masing perlakuan penularan; M, penanda DNA 1 kb ladder, K-, kontrol negatif, K+, kontrol positif.
36
7 Runutan basa nukleotida gen CP RRSV asal Subang (Sukamandi) dan beberapa isolat RRSV asal luar negeri
37 8 Runutan basa nukleotida gen CP RGSV asal Subang
(Indo-RG3) dan beberapa isolat RGSV asal luar negeri
39 9 Runutan basa nukleotida gen CP RTBV asal Purwakarta dan
beberapa isolat RTBV asal luar negeri
1
PENDAHULUAN
Latar Belakang
Indonesia merupakan negara pengonsumsi dan penghasil beras ketiga di dunia setelah Tiongkok dan India. Gangguan serangga hama yang banyak menimbulkan kerugian pada tanaman padi di Indonesia adalah wereng hijau (Nephotettix virescens Distan), dan wereng batang cokelat (Nilaparvata lugens Stal.). Selain sebagai serangga hama, kedua spesies serangga tersebut dapat berperan sebagai vektor virus, sehingga keberadaannya di pertanaman berpotensi mengakibatkan kerugian yang lebih besar. Wereng hijau merupakan vektor penyakit tungro yang disebabkan oleh Rice tungro spherical virus (RTSV)dan Rice tungro bacilliform virus (RTBV) (Rahim dan Nasrudin 2010), sedangkan wereng batang cokelat merupakan vektor penyakit kerdil hampa yang disebabkan oleh Rice ragged stunt virus (RRSV), dan penyakit kerdil rumput yang disebabkan oleh Rice grassy stunt virus (RGSV) (Cabautan et al. 2009)
Penyakit tungro tersebar hampir di seluruh wilayah Indonesia, terutama di daerah sentra produksi beras nasional seperti di Pulau Jawa, Bali, Nusa Tenggara Barat, Sulawesi, dan Kalimantan Selatan (BB Padi 2010). Di Propinsi Jawa Barat populasi wereng hijau dan insidensi penyakit tungro telah ditemukan di Kabupaten Purwakarta, Garut, Cirebon, Subang, Bogor, Bekasi (Dirjen Tanaman Pangan 1992; Yuliani 2014). Gejala utama penyakit tungro antara lain tampak pada perubahan warna daun muda menjadi kuning-oranye dimulai dari ujung daun, jumlah anakan berkurang dan tanaman menjadi kerdil, dan beberapa mengalami klorosis intervenal (Bunawan et al. 2014). Penyebaran penyakit tungro sangat bergantung pada populasi dan aktivitas serangga vektornya.
Selain penyakit tungro, penyakit kerdil juga menjadi kendala peningkatan produksi padi nasional. Penyakit kerdil hampa pada padi dilaporkan pertama kali di Indonesia pada tahun 1976 (Hibino et al.1977). Tanaman padi yang terserang penyakit kerdil hampa mengalami pengerdilan, daun menjadi berwarna gelap dengan tepi bergerigi atau ujung memutar, dan tulang daun mengalami pembengkakan atau benjolan di bagian bawah helai daun dan bagian luar permukaan pelepah daun (Cabautan at al. 2009). Penyakit kerdil rumput dilaporkan pertama kali di Indonesia pada tahun 1971 dan disebut sebagai kerdil rumput tipe I, kemudian pada tahun 2006 ditemukan penyakit kerdil rumput tipe II. Gejala penyakit kerdil rumput diantaranya tanaman menjadi sangat kerdil, anakan banyak, daun hijau pucat sampai kuning atau daun-daun sempit berwarna kuning sampai oranye, daun sempit dengan bintik-bintik karat kecil (IRRI 2002). Dilaporkan oleh Nurbaeti et al. (2010) dan Romadhon (2007) bahwa daerah endemik serangan wereng batang cokelat di Propinsi Jawa Barat meliputi Kabupaten Cirebon, Bekasi, Majalengka, Sukabumi, Tasikmalaya, Garut, Cianjur, Subang, Karawang dan Indramayu; sedangkan insidensi penyakit kerdil dilaporkan masih terbatas di Kabupaten Subang dan Cirebon (Maulana dan Abdurahman 2014).
2
diikuti dengan ledakan hama wereng hijau pada tahun terjadinya ledakan penyakit tersebut (Baehaki dan Suharto 1985).
Sementara itu penelitian Nurbaeti et al. (2010) menunjukkan hal yang berbeda pada dinamika populasi wereng batang cokelat. Saat terjadinya ledakan populasi wereng batang cokelat pada tahun 2010 yang mengakibatkan lahan pertanaman padi seluas 4874 ha mengalami kerusakan, ledakan populasi wereng tersebut juga berdampak pada munculnya insidensi penyakit kerdil di lapangan. Hal ini menunjukkan bahwa setiap serangga vektor memiliki kemampuan yang berbeda-beda dalam menularkan dan menyebarkan virus.
Diagnosis penyakit tungro dan penyakit kerdil tidak dapat mengandalkan hanya dari gejala saja karena gejala yang muncul beranekaragam serta mirip dengan gejala kekurangan hara dan kekeringan. Penggunaan teknik molekuler untuk mendeteksi penyakit tungro dan kerdil telah banyak dilaporkan, diantaranya menggunakan metode polymerase chain reaction (PCR) (Uehara-Ichiki et al. 2013). Analisis keragaman genetika virus-virus padi di Asia sudah dilaporkan oleh beberapa peneliti (Arenal et al. 2003; Cheng et al. 2013; Huang et al. 2015). Analisis keragaman virus tungro asal daerah endemis di Indonesia menunjukkan adanya indikasi kombinasi pada tingkat molekuler antara isolat-isolat RTSV dan RTBV dari daerah geografi yang berbeda (Praptana 2013). Sementara itu analisis keragaman virus kerdil masih sangat terbatas.
Tujuan Penelitian
1. Mengamati insidensi penyakit kerdil dan mengetahui populasi wereng batang cokelat di pertanaman padi di Sukamandi, Kabupaten Subang, Propinsi Jawa Barat.
2. Mempelajari hubungan insidensi penyakit dengan populasi wereng.
3. Menentukan efisiensi wereng hijau dan wereng batang cokelat dalam menularkan penyakit tungro dan kerdil ke tanaman padi.
4. Mengetahui kekerabatan isolat virus tungro, virus kerdil hampa dan virus kerdil rumput asal Indonesia dengan isolat virus asal negara lain.
Hipotesis Penelitian
Ada hubungan antara insidensi dan keparahan penyakit yang disebabkan oleh virus dengan populasi wereng hijau dan wereng batang cokelat di pertanaman padi. Masing-masing jenis serangga vektor bersifat spesifik dan memiliki efisiensi penularan yang tinggi, yaitu wereng hijau hanya menularkan penyakit tungro dan wereng batang cokelat hanya menularkan penyakit kerdil.
Manfaat Penelitian
2 TINJAUAN PUSTAKA
Sifat Umum Tanaman Padi (Oryza sativa)
Tanaman padi (Oryza sativa L.) merupakan salah satu tanaman pangan utama di dunia, khususnya Indonesia. Tanaman padi menghasilkan beras dengan kandungan karbohidrat 80%, protein 7-8%, lemak 3% dan serat 3% (Ahuja et al. 2008). Pertumbuhan tanaman padi dapat dibagi dalam tiga fase yaitu fase vegetatif, fase reproduktif, dan fase pematangan. Fase vegetatif merupakan fase dimana berlangsung pertumbuhan organ-organ vegetatif seperti pertambahan jumlah anakan, tinggi tanaman, jumlah, bobot, dan luas daun. Fase reproduktif ditandai dengan memanjangnya beberapa ruas teratas batang tanaman, berkurangnya jumlah anakan, munculnya daun bendera, bunting dan pembungaan. Fase terakhir yaitu fase pematangan sekitar 30 hari. Pada fase ini gabah mulai terisi, malai mulai merunduk, gabah mulai menguning, kemudian gabah terisi penuh dan berwarna kuning, daun bagian atas mengering (Makarim dan Suhartatik 2009).
Morfologi tanaman sangat berpengaruh terhadap produktivitas tanaman padi tersebut. Varietas padi yang saat ini banyak ditanam di Indonesia diantaranya ialah varietas Mekongga dan IR64. Karakteristik varietas Mekongga antara lain tanaman berumur 116–125 hari, bentuk tanaman tegak, tinggi tanaman sekitar 91 sampai 106 cm, anakan produktif berjumlah 13 sampai 16 batang. Tekstur nasi yang dihasilkan pulen, kadar amilosa pada bulir padi sebesar 23 %, rata-rata hasil produksi padi 6.0 t/ha, varietas ini tahan terhadap wereng batang cokelat biotipe 2 dan 3, sedangkan untuk penyakit agak tahan terhadap hawar daun bakteri strain IV (Puslittan 2007). Karakteristik varietas IR64 menurut Daradjat et al. (2001) antara lain adalah tanaman berumur sedang (100−125 HSS), postur tanaman pendek sampai sedang sekitar 95 sampai 115 cm, bentuk tanaman tegak, posisi daun tegak, jumlah anakan sedang antara 20 sampai 25 anakan/rumpun, dengan
anakan produktif 15−16 anakan/rumpun, panjang malai sedang, responsif terhadap pemupukan, tahan rebah, daya hasil 5.0 sampai 6.0 t/ha, tahan hama dan penyakit utama, mutu giling baik, dan rasa nasi enak. Beberapa penelitian menunjukan bahwa varietas IR64 merupakan varietas dengan tingkat ketahanan yang moderat terhadap serangan wereng.
Karakteristik Penyakit Tungro
Penyakit Tungro
4
Infeksi penyakit tungro pada tanaman padi dapat terjadi sejak tanaman di persemaian dan kerugian dapat berupa kehilangan hasil yang besarannya tergantung pada stadia tanaman saat terjadi infeksi, tingkat ketahanan varietas dan pola perkembangan serangan. Tungro merupakan penyakit virus yang biasanya menyerang tanaman padi pada fase pertumbuhan vegetatif. Gejala utama pada tanaman padi yang terinfeksi virus tungro adalah perubahan warna daun menjadi kuning-oranye, kerdil, dan penurunan jumlah anakan (Hibino et al. 1978).Selain itu menurut IRRI (2002) gejala tungro mulai dari ujung daun yang lebih tua. Daun menguning berkurang bila daun yang lebih tua terinfeksi. Pelepah dan helaian daun memendek. Daun muda sering berlurik atau strip berwarna hijau pucat sampai putih dengan panjang berbeda sejajar dengan tulang daun.
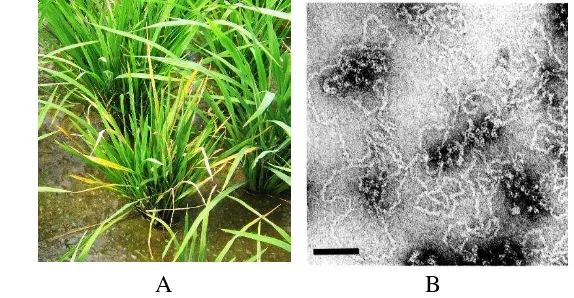
Gambar 1 Gejala serangan penyakit tungro pada tanaman padi dengan gejala menguning pada daun (Dokumentasi pribadi)
Penyakit tungro disebabkan oleh infeksi 2 virus, yaitu Rice tungro bacilliform tungrovirus (RTBV) dan Rice tungro spherical wakaivirus (RTSV). Kedua virus tersebut ditularkan terutama oleh wereng hijau Nephotettix virescens Distant (Hemiptera: Cicadelidae) (Hibino dan Cabunagan 1986).Apabila tanaman padi hanya terinfeksi oleh RTBV saja maka gejala yang ditimbulkan adalah gejala khas tungro ringan sedangkan apabila tanaman padi hanya terinfeksi oleh RTSV maka tanaman hampir tidak menunjukkan gejala atau kerdil sangat ringan (Hibino 1983). Dalam hal ini RTBV berfungsi sebagai penentu gejala sedangkan RTSV adalah virus helper (Hibino 1996). Berdasarkan laporan dari Jones et al. (1991) diketahui bahwa RTBV termasuk kedalam genus Badnavirus. Genom RTBV ini memiliki DNA beruntai ganda dengan bentuk partikel bacilliform berdiameter 30-35 nm dan panjangnya kira-kira 100-300 nm. Rice tungro spherical virus (RTSV) termasuk kedalam genus Waikavirus. Beberapa sifat molekul virus yang menunjukkan bahwa dan RTSV adalah virus dengan asam nukleat berupa RNA (Hull 2002).
Serangga Vektor Wereng Hijau
5
wilayah non endemik 52% (Supriyadi et al 2008). Penelitian Rahim dan Nasrudin (2010) menunjukkan efektivitas wereng hijau menularkan virus tungro dari sumber inokulum pada umur inkubasi yang berbeda berhasil tertular sebesar 96%. Penyakit ini ditularkan dari satu tanaman ke tanaman lain secara semi persisten yaitu, periode makan akuisisi yang dibutuhkan untuk menularkan kembali penyakit tungro berkisar antara 5-30 menit, tidak ada periode laten, dengan waktu makan inokulasi 7-30 menit dan periode retensi hanya 6 hari.
Wereng hijau dewasa berwarna hijau kekuningan dengan bercak hitam pada ujung sayapnya dan kadang bercak hitam pada bagian tengah sayap serta ujung kepala meruncing, memiliki sepasang sayap dimana sepasang sayap belakang tebal berwarna hijau dengan bercak hitam dan sepasang sayap belakang tipis seperti selaput. Bagian tubuhnya meruncing ke arah belakang dan memiliki satu deret duri pada bagian tibia tungkai belakang (Suwarno et al. 2010).
Wereng hijau betina meletakkan telurnya pada pelepah padi muda dan telur diletakkan secara berkelompok, biasanya 8-16 butir. Umur serangga dewasa 20-30 hari dan telur menetas dalam waktu 6-7 hari. Nimfa mengalami lima instar dan lamanya stadium nimfa 16-18 hari. Nimfa menghisap cairan tanaman dari jaringan xilem pada helai daun dan pelepah daun. Stadium nimfa dapat juga berperan sebagai vektor penyakit virus (Baehaki dan Widiarta 2009).
Gambar 2 Imago wereng hijau (Nephotettix virescens Distant) (Pathak dan Khan 1994)
6
Karakteristik Penyakit Kerdil
Penyakit Kerdil Hampa
Penyakit kerdil hampa pada padi ditemukan pertama kali di Indonesia tahun 1976, selanjutnya ditemukan di Taiwan pada tahun 1978, dan kemudian menyebar di beberapa negara di Asia. Penyakit ini disebabkan oleh Rice ragged stunt virus (RRSV). Tanaman padi yang terserang kerdil hampa mengalami pengkerdilan, daun menjadi berwarna gelap dengan tepi bergerigi atau ujung memutar, dan vena mengalami pembengkakan atau benjolan di bagian bawah helai daun dan dagian luar permukaan pelepah daun. Benjolan ini dihasilkan dari hiperplasia dan hipertropi pada jaringan floem (Cabautan et al. 2009).
Gambar 3 Gejala serangan Rice ragged stunt virus pada tanaman padi. tepi tanaman padi menjadi bergerigi (A), ujung tanaman padi memutar (B) (Dokumentasi pribadi), Partikel RRSV dilihat melalui mikroskop elektron (C) (Chen et al.1997)
Rice ragged stunt virus (RRSV) termasuk dalam Famili Reoviridae, Genus Oryzavirus. Virus kerdil hampa ini mempunyai partikel yang berbentuk polihedral dengan ukuran antara diameter 50-70 nm. Hibino et al. (1978) melaporkan bahwa penyakit kerdil hampa disebabkan oleh virus yang berbentuk bulat dengan ukuran 60 nm. Penularan RRSV melalui wereng batang cokelat memerlukan periode makan akuisisi minimal selama 24 jam, dengan periode laten 2 jam, periode makan inokulasi 6 jam serta periode retensi selama hidup serangga vektor (IRRI 2002). Wereng batang cokelat dapat menularkan penyakit virus kerdil hampa sampai akhir masa hidupnya, tetapi tidak dapat menularkan kepada keturunannya lewat telur (Ling et al. 1978).
Penyakit Kerdil Rumput
Penyakit kerdil rumput dilaporkan pertama kali di temukan pada tahun 1970 di Taiwan (Chen dan Chiu 1982). Pada tahun 1982-1983 penyakit ini dilaporkan ditemukan di Filipina (Hibino et al. 1985). Du et al. (2005) melaporkan bahwa pada tahun 2000 negara Vietnam mengalami kerugian ekonomi yang cukup besar akibat penyakit kerdil rumput, kerdil hampa dan akibat langsung serangan wereng batang cokelat. Gejala pada tanaman yang terserang penyakit kerdil rumput ini
7
terlihat bahwa tanaman menjadi sangat kerdil, anakan banyak, daun hijau pucat sampai kuning atau daun-daun sempit berwarna kuning sampai oranye, daun sempit dengan bintik-bintik karat kecil (IRRI 2002).
A B
Gambar 4 Gejala penyakit kerdil rumput pada rumpun tanaman padi yang menunjukkan perubahan warna daun menjadi hijau pucat hingga kuning (A) (Dokumentasi pribadi), Hasil pemurnian RGSV 2 dengan pewarnaan uranil asetat, ukuran 100nm (B) (Hibino 1985)
Rice grassy stunt virus (RGSV) termasuk Famili Bunyaviridae, Genus Tenuivirus (Hull 2002). Organisasi genom RGSV terdiri atas 6 segmen RNA yang seluruhnya adalah molekul single stranded RNA yang ambisense (Miranda et al. 2000). Partikel RGSV berbentuk pleomorphic, dapat terlihat seperti filamen tipis ataupun filamen yang melingkar dan seringkali membentuk konfigurasi spiral (Toriyama et al. 1998). N. lugens mendapatkan virus dengan makan pada tanaman padi sakit (periode makan akuisisi) selama 5 sampai 10 menit. Jumlah virus pada tubuh N. lugens akan semakin meningkat jika periode makan akuisisi semakin lama. Periode laten rata-rata 10 hari dan langsung dapat menularkan virus. Masa retensi yang dibutuhkan RGSV di tubuh N. lugens selama siklus hidup vektornya atau 24 hari (Reissig et al. 1986). Cabauatan et al. (2009) melaporkan bahwa penularan RGSV bersifat transtadial yaitu virus tetap berada dalam tubuh vektor selama siklus hidupnya. RGSV dapat ditularkan oleh nimfa maupun imago N. lugens.
RRSV dan RGSV ditularkan oleh wereng batang cokelat secara persisten propagatif (Chen et al. 1997). Hull (2002) menjelaskan bahwa virus yang ditularkan secara persisten propagatif memperbanyak diri dalam vektornya terlebih dahulu sebelum ditularkan pada tanaman inang dan waktu yang dibutuhkan disebut sebagai periode laten.
Serangga Vektor Wereng Cokelat
8
batang tanaman (Mueller 1974). Panjang tubuh serangga dewasa 2 – 4.4 mm. Serangga dewasa mempunyai 2 bentuk, yaitu bersayap pendek (brakhiptera) dan bersayap panjang (makroptera). Serangga makroptera mempunyai kemampuan untuk terbang, sehingga dapat bermigrasi cukup jauh. Wereng batang cokelat adalah serangga monofag, inangnya terbatas pada padi dan padi liar (Oryza parennis dan O. spontanea) (BPTP 2010).
Gambar 5 Imago N. lugens bersayap panjang (Makroptera) (A), Imago N. lugens bersayap pendek (Brakhiptera) (B) (Dokumentasi Pribadi)
Wereng batang cokelat berkembangbiak secara seksual, masa pra peneluran 3-4 hari untuk brakiptera (bersayap pendek) dan 3-8 hari untuk makroptera (bersayap panjang). Satu ekor betina mampu meletakkan telur 100-500 butir. Telur diletakkan berkelompok, satu kelompok telur terdiri dari 3-21 butir. Telur menetas setelah 9 hari di daerah tropis. Telur biasanya diletakkan pada jaringan pangkal pelepah daun, tetapi jika populasinya tinggi telur diletakkan di ujung pelepah daun dan tulang daun. Nimfa mengalami lima instar, dan rata-rata waktu yang diperlukan untuk menyelesaikan periode nimfa adalah 12-13 hari (Badan litbangtan 2010). Wereng batang cokelat menyukai lingkungan dengan kelembapan tinggi (70-80%), suhu siang hari optimum (28-30 oC), intensitas cahaya matahari rendah, pemupukan N tinggi, tanaman rimbun, lahan basah, angin lemah (BPTP 2010).
3 METODE
Lokasi dan Waktu Penelitian
Penelitian dilakukan di Balai Besar Penelitian Padi (BB Padi) Sukamandi, Jawa Barat mulai November hingga Desember 2014, dan di Laboratorium Virologi Tumbuhan, Departemen Proteksi Tanaman, Fakultas Pertanian, Institut Pertanian Bogor, mulai Februari hingga Mei 2015.
Pengamatan Insidensi Penyakit Kerdil dan Populasi Wereng Batang Cokelat di Lapangan
Pengamatan dilakukan di lahan penanaman padi milik BB Padi di Desa Sukamandi, Kecamatan Patok Beusi, Kabupaten Subang, Provinsi Jawa Barat. Petak pengamatan berjumlah 3 plot, masing-masing berukuran 300 m2, jarak tanam 25 × 25 cm. Untuk menghitung populasi wereng setiap plot pengamatan terdiri atas 100 tanaman yang ditentukan menggunakan metode pengambilan sampel diagonal . Insidensi penyakit dihitung berdasarkan rasio jumlah tanaman yang menunjukkan gejala dengan jumlah seluruh tanaman yang diamati dikali 100%, dan diamati mulai umur tanaman 1 minggu setelah tanam (MST) sampai 8 MST dengan interval 7 hari.
Gejala penyakit kerdil yang diamati mencakup gejala kerdil rumput atau kerdil hampa, yaitu penyempitan daun, warna daun berubah menjadi hijau pucat, adanya daun yang menggulung, penurunan tinggi tanaman, jumlah anakan yang lebih banyak, adanya pembengkakan pada pangkal daun. Selain gejala penyakit diamati pula populasi wereng batang cokelat. Pengamatan serangga dimulai sejak awal pindah tanam bibit padi sampai fase generatif awal dengan interval 7 hari. Populasi wereng batang cokelat dihitung secara langsung dari pangkal batang tanaman padi dalam unit contoh.
Uji Efisiensi Penularan Wereng Hijau dan Wereng Batang Cokelat sebagai Vektor Virus Padi
Perbanyakan wereng hijau dan wereng batang cokelat
Populasi wereng hijau dan wereng batang cokelat yang digunakan untuk percobaan di rumah kaca berasal dari populasi lapangan yang diambil di daerah Subang. Sebanyak 20 ekor imago masing-masing wereng dari lapangan dimasukkan ke dalam sungkup kasa terpisah berisi tanaman padi varietas IR64 berumur 10 hari setelah semai (HSS) sebagai pakan dan media peneluran. Tanaman padi tersebut dikeluarkan dari dalam sungkup setelah 1 hari, kemudian dipindahkan ke dalam kurungan yang berbeda di rumah kaca untuk mendapatkan imago wereng yang bebas virus. Penggantian pakan dilakukan secara terus-menerus hingga terbentuk populasi wereng dewasa.
Perbanyakan inokulum virus tungro dan kerdil
10
inokulum virus. Propagasi virus dilakukan di rumah kaca pada tanaman padi varietas IR 64 dengan metode penularan tabung reaksi dengan serangga vektor (Cabautan et al. 1994). Perbanyakan inokulum virus tungro dilakukan dengan menginfestasikan imago serangga wereng hijau dan untuk virus kerdil dilakukan dengan menginfestasikan nimfa instar 3-4 serangga wereng batang cokelat dengan periode makan akuisisi, periode laten, dan periode makan inokulasi yang berbeda (Tabel 1). Periode makan inokulasi dilakukan didalam tabung, setiap tabung terdapat satu tanaman sehat dengan 2 ekor vektor viruliferous. Setelah periode makan inokulasi serangga dimatikan dengan perlakuan insektisida berbahan aktif buthylphenylmethyl carbomate (BPMC). Tanaman padi yang sudah terinfeksi virus tungro atau virus kerdil dari hasil penularan dipelihara untuk digunakan sebagai sumber inokulum.
Tabel 1 Tahapan penularan virus tungro oleh wereng hijau dan virus kerdil oleh wereng batang cokelat
Tahapan penularan Virus tungro Virus kerdil
Periode makan akuisisi (hari) 1 2
Periode laten (hari) tidak dilakukan 4
Periode makan inokulasi (hari) 1 1
Uji penularan
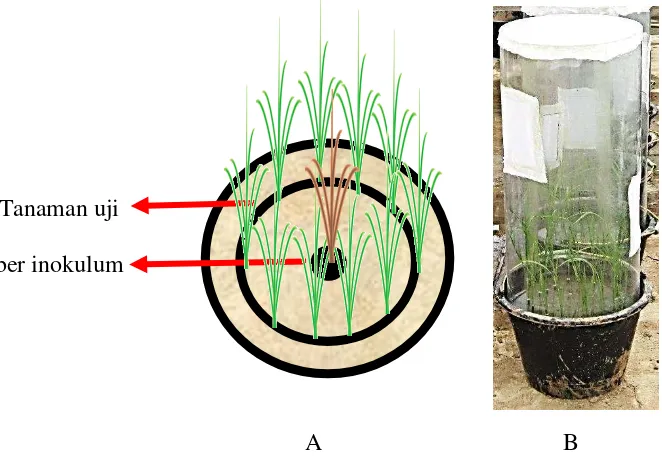
Sebanyak 10 bibit padi varietas IR64 berumur 10 HSS dipindahkan ke dalam ember percobaan berdiameter 30 cm yang berisi tanah dengan kondisi menyerupai sawah. Bibit padi ditanam dengan pola melingkar (Gambar 6), pada bagian tengah dari lingkaran tersebut ditanam bibit padi yang terinfeksi virus sebagai sumber penyakit. Masing-masing ember percobaan disungkup dengan kurungan yang kedap serangga. Sebanyak 10 imago wereng hijau dan/atau wereng batang cokelat hasil perbanyakkan diinfestasikan ke dalam setiap ember percobaan sesuai dengan perlakuan (Tabel 2). Serangga wereng tersebut dibiarkan berada pada ember percobaan selama 7 hari, kemudian dimusnahkan dengan perlakuan insektisida. Peubah Pengamatan
Percobaan terdiri atas 10 perlakuan, masing-masing diulang sebanyak 3 kali. Peubah pengamatan meliputi periode inkubasi, insidensi penyakit, dan keparahan penyakit. Pengamatan dilakukan selama satu bulan sejak infestasi serangga ke dalam ember percobaan, dengan interval pengamatan 7 hari. Periode inkubasi ditentukan pada saat pertama gejala muncul. Insidensi penyakit dan keparahan penyakit dihitung dengan rumus :
�� =� � %
11
A B
Tanaman uji Sumber inokulum
Dengan KP, keparahan penyakit; n, jumlah tanaman yang menunjukkan nilai skor tertentu; v, nilai skor untuk setiap kategori gejala penyakit; N, jumlah sampel yang diamati; Z, skor tertinggi kategori gejala penyakit. Keparahan penyakit dievaluasi berdasarkan sistem skor sesuai dengan Standard Evaluation System for Rice (SES) yang dimodifikasi (IRRI 2002) (Tabel 3).
Gambar 6 Skema percobaan penularan virus dengan serangga vektor (A), Contoh arena percobaan pengujian efisiensi wereng hijau dan wereng batang cokelat sebagai vektor virus padi menggunakan ember (B)
Tabel 2 Perlakuan kombinasi virus dan serangga vektor pada uji efisiensi penularan wereng hijau dan wereng batang cokelat sebagai vektor virus padi
No Sumber virus Serangga vektor
1. Tungro Wereng hijau
2. Kerdil Wereng batang cokelat
3. Tungro Wereng hijau dan wereng batang cokelat
4. Kerdil Wereng hijau dan wereng batang cokelat
5. Tungro dan kerdil Wereng hijau dan wereng batang cokelat 6. Tungro dan kerdil Wereng hijau
7. Tungro dan kerdil Wereng batang cokelat
8. Tanaman sehat (tanpa inokulum) Wereng hijau dan wereng batang cokelat 9. Tanaman sehat (tanpa inokulum) Wereng hijau
12
Tabel 3 Skor gejala penyakit tungro dan kerdil berdasarkan Standard Evaluation System for Rice (SESR) yang dimodifikasi
Skor Deskripsi gejala penyakit
Tungro Kerdil
0 Tidak ada gejala serangan Tidak ada gejala serangan 3 Tinggi tanaman lebih
pendek1-Terjadi penurunan tinggi tanaman 11-30%, jumlah anakan banyak dan kecil, daun berwarna hijau pucat atau beberapa daun menggulung atau pembengkakan pangkal daun terdapat perubahan warna menjadi hijau pucat daun menjadi kasar, menggulung atau pembengkakan pada pangkal daun.
Deteksi Virus Penyebab Penyakit Tungro dan Kerdil
Deteksi dilakukan pada sampel tanaman dari lapangan dan pada beberapa sampel tanaman hasil uji penularan.
Deteksi RRSV dan RGSV dengan metode Reverse Transcription-Polymerase Chain Reaction (RT-PCR). Tahapan deteksi terdiri atas ekstraksi RNA total dengan metode Cetyl Trimethyl Ammonium Bromide (CTAB) (Doyle dan Doyle 1990), sintesis cDNA, amplifikasi DNA target, dan visualisasi DNA.
13
disentrifugasi dengan kecepatan 8 000 rpm selama 5 menit. Cairan dibuang, lalu pelet RNA dikeringanginkan, selanjutnya ditambahkan 50 µL bufer TE 1x (10 mM Tris-HCl pH 8.0 mM EDTA.
Sintesis cDNA. RNA total digunakan sebagai templet dalam reaksi transkripsi balik (reverse transcription/RT) untuk menghasilkan cDNA. Reaksi RT (10 µL) terdiri atas 2 µL bufer RT, 1 µL dNTP 10 mM, 1 µL DTT 50 mM, 0.50 µL RNAse Inhibitor (Thermo Scientific, US), 0.50 µL M-MuLV (Thermo Scientific, US), 2 µL H2O bebas nuklease, 1 µL Oligo d(T) 10 mM, dan 2 µL templet RNA. Pertama-tama RNA total dan Oligo d(T) diinkubasi dalam penangas air selama 5 menit pada suhu 60 oC. Bahan pereaksi lainnya kemudian ditambahkan kedalam tabung mikro, selanjutnya tabung mikro disentrifugasi selama 1 menit dengan kecepatan 1 000 rpm. Tabung mikro dinkubasi pada suhu 42 oC selama 60 menit ,
selanjutnya diinkubasi pada suhu 70 oC selama 5 menit .
Amplifikasi DNA target. Setelah diperoleh cDNA dilanjutkan dengan amplifikasi DNA target. Setiap reaksi amplifikasi (25µL) terdiri atas 1 µL cDNA, 1 µL primer F 10 µM, 1 µL primer R 10 µM, 12.5 µL GTG Master mix, dan 2H2O sebanyak 9.5 µL. Primer yang digunakan adalah primer spesifik RGSV dan RRSV, yaitu primer RGSV-S3-F (5’-AGAATTTTTATGTCACTTAG-3’) dan RGSV
-S3-R (5’-TATCCAGATTCCAG GTGC-3’) dengan target amplikon ±738 pb untuk RGSV; RRSV-S9-F (5’-ATGAAGCGCTCGGAACCAA-3’) dan RRSV-S9-R (5’ -GCGTCTAGTCCCGTATGG-3’) dengan target amplikon ±445 pb untuk RRSV. Amplikasi cDNA dimulai dengan tahapan pradenaturasi pada suhu 94 oC selama 5 menit sebanyak 1 siklus, dilanjutkan dengan amplifikasi sebanyak 35 siklus dengan tahapan denaturasi pada suhu 94 oC selama 1 menit, aneling selama 1 menit pada suhu 50 oC untuk RGSV atau 56 oC untuk RRSV, dan pemanjangan utas baru pada suhu 72 oC selama 1 menit , dan diakhiri dengan 1 siklus pemanjangan akhir pada suhu 72 oC selama 7 menit.
Visualisasi DNA. Visualisasi produk amplifikasi dilakukan dengan elektroforesis pada 1% gel agarosa (0.5xTris-Borate EDTA /TBE). Elektroforesis dilakukan pada 50 volt selama 50 menit, selanjutnya gel agarosa diinkubasi pada zat pewarna yang berisi Etidium Bromida (1%) selama 15 menit, lalu dicuci dengan H2O selama 10 menit. Pengambilan gambar dilakukan menggunakan UV transiluminator dan didokementasikan dengan kamera digital.
Deteksi RTBV dengan metode Polymerase Chain Reaction (PCR). Deteksi RTBV secara molekuler dengan metode PCR terdiri atas 3 tahapan, yaitu ekstraksi DNA total, amplifikasi DNA target, visualisasi hasil amplifikasi.
14
disentrifugasi dengan kecepatan 13 000 rpm selama 10 menit. Supernatan diambil dan ditempatkan kedalam tabung mikro yang baru, dan ditambahkan dengan Sodium Asetat sebanyak 1/10 dari total supernatan yang didapat, kemudian ditambahkan Isopropanol sebanyak 2/3 dari jumlah total supernatan. Selanjutnya tabung dibolak-balik kemudian disentrifugasi selama 5 menit dengan kecepatan 13 000 rpm. Supernatan dibuang dan pelet DNA diambil, kemudian ditambahkan dengan Etanol 70% sebanyak 600 µL, selanjutnya disentrifugasi dengan kecepatan 8 000 rpm selama 5 menit. Etanol 70% dibuang, lalu pelet DNA dikeringkan dengan meletakkan tabung dalam posisi terbalik. Setelah pelet DNA kering, ditambahkan 50 µL bufer TE 1x (10 mM Tris-HCl pH 8.0 mM EDTA).
Amplifikasi DNA. Setiap reaksi amplifikasi (25µL) terdiri atas 1 µL DNA, 1 µL primer F 10 µM, 1 µL primer R 10 µM, 12.5 µL GTG Master mix, dan 2H2O sebanyak 9.5 µL. Primer yang digunakan adalah primer spesifik dengan target protein selubung RTBV, yaitu primer ForwardRTBV2L (5’-GCTGAGGTG CTTACATAGGTT-3’) dan primer Reverse RTBV2R (5-GGTCTTGGATGGAT GCTAGA-3’) (Hikmahayati 2010) dengan target amplikon ±1400 pb. Amplikasi DNA dimulai dengan tahapan pradenaturasi pada suhu 94 oC selama 5 menit sebanyak 1 siklus, denaturasi pada suhu 94 oC selama 1 menit, aneling selama 1 menit di suhu 55 oC dan ekstensi pada suhu 72 oC selama 1 menit sebanyak 34 siklus. Selanjutnya sebanyak 1 siklus untuk ekstensi akhir pada suhu 72 oC selama 7 menit. Hasil amplifikasi divisualisasi seperti diuraikan sebelumnya.
Analisis Susunan Nukleotida
4 HASIL DAN PEMBAHASAN
Populasi Wereng dan Insidensi Penyakit di Lapangan
Pengamatan Populasi Wereng di Lapangan
Berdasarkan hasil tangkapan menggunakan jaring serangga untuk mengamati populasi wereng hijau diketahui bahwa pada lahan pengamatan di Sukamandi tidak didapatkan serangga wereng hijau. Baehaki dan Suharto (1985) menyatakan bahwa populasi wereng hijau dipengaruhi oleh keadaan ekologi, dengan faktor iklim akan mempengaruhi distribusi, pertumbuhan, perilaku, mortalitas serta migrasi wereng hijau tersebut. Wereng hijau lebih sering ditemukan di daerah dataran sedang sampai tinggi dengan kisaran ketinggian 500 m – 995 m dpl (Yuliani 2014). Provinsi Jawa Barat, Jawa Tengah, Yogyakarta, Sulawesi Tenggara, Sulawesi Barat, Bali dan Nusa Tenggara Barat merupakan daerah endemis wereng hijau di Indonesia (Praptana et al. 2013). Desa Sukamandi di Kabupaten Subang termasuk dalam daerah dataran rendah dengan ketinggian 10 m dpl (Romadhon 2007). Kondisi lokasi pengamatan tersebut tampaknya tidak sesuai dengan kondisi iklim yang mendukung perkembangan wereng hijau.
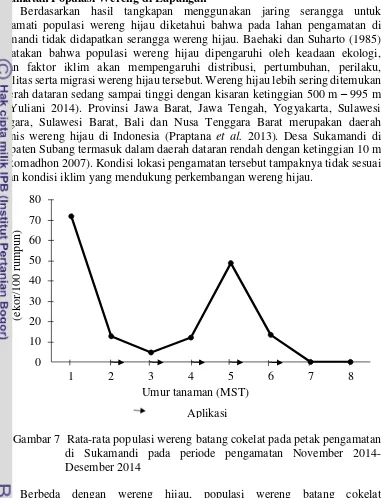
Gambar 7 Rata-rata populasi wereng batang cokelat pada petak pengamatan di Sukamandi pada periode pengamatan November 2014-Desember 2014
Berbeda dengan wereng hijau, populasi wereng batang cokelat menunjukkan adanya fluktuasi populasi pada setiap minggu pengamatan (Gambar 7). Rata-rata populasi wereng batang cokelat tertinggi mencapai 71.7 ekor per 100 tanaman pada 1 MST, sedangkan populasi terendah pada 7 dan 8 MST yaitu 0 ekor per 100 tanaman. Fluktuasi populasi ini dapat terjadi karena proses adaptasi wereng batang cokelat setelah migrasi dari lahan sebelumnya. Wereng yang mampu beradaptasi akan bertahan dan berkembangbiak, sedangkan wereng yang tidak
16
mampu beradaptasi pada tempat baru akan mati. Keberadaan musuh alami juga mempengaruhi populasi wereng batang cokelat. Saat pengamatan ditemukan laba-laba yang merupakan predator nimfa maupun imago wereng batang cokelat. Selain ditemukan populasi wereng batang cokelat tipe makroptera, selama pengamatan juga ditemukan wereng batang cokelat stadia nimfa sejak 3 MST dan populasi imago tipe brakhiptera pada 5 MST (Gambar 8). Wereng makroptera mempunyai kemampuan terbang yang tinggi dan berperan melakukan migrasi dalam jumlah banyak dari satu tempat ke tempat lainnya untuk membentuk populasi baru (Baehaki 2011); sedangkan wereng batang cokelat brakhiptera umumnya bertubuh lebih besar, mempunyai tungkai dan ovipositor lebih panjang dan berperan lebih banyak dalam hal reproduksi (Baehaki dan Widiarta 2009).
Gambar 8 Wereng batang cokelat stadia imago makroptera (A), stadia nimfa (B), dan imago brakhiptera (C)
Lahan padi di Sukamandi, Subang dikelola secara konvensional dan aplikasi insektisida diberikan secara terjadwal, yaitu pada hari kedua setiap minggu sejak minggu kedua setelah tanam. Insektisida diaplikasikan untuk hama sasaran yang berbeda-beda sesuai dengan populasi hama tertinggi di lapangan. Hal ini menunjukan bahwa aplikasi insektisida pada setiap minggunya mempengaruhi populasi hama pada minggu berikutnya. Setiap aplikasi insektisida dilakukan terjadi penurunan populasi hama target dan juga hama non target. Aplikasi insektisida tidak mampu membunuh telur wereng yang diletakkan di dalam jaringan batang padi, dan telur tetap dapat menetas sehingga terjadi fluktuasi populasi wereng batang cokelat.
Stadia tanaman juga mempengaruhi keberadaan wereng di lapangan. Populasi tertinggi wereng batang cokelat terjadi pada saat tanaman berumur 1 MST. Hal ini didukung laporan dari Ikeda dan Vaughan (2004) yang menyatakan bahwa stadia tanaman yang rentan terhadap serangan wereng batang cokelat adalah dari pembibitan sampai fase matang susu.
Hasil pengamatan populasi wereng batang cokelat yang dilakukan pada kurun waktu November sampai Desember 2014 di daerah Sukamandi, Subang tidak menunjukkan adanya populasi yang tinggi. Di Indonesia pernah terjadi ledakan hama wereng cokelat pada tahun 1982 di Sumatera Utara, tahun 1998 di Kabupaten
17
Subang, tahun 2005 di Kabupaten Cirebon, tahun 2010 di Kabupaten Subang, Karawang, Indramayu (Baehaki 2011; BPTP 2010).
Pengamatan Insidensi Penyakit di Lapangan
Selama pengamatan pada November – Desember 2014, pertanaman padi di Desa Sukamandi tidak terserang penyakit tungro. Insidensi penyakit kerdil mulai terlihat pada 2 MST dan mengalami peningkatan sampai dengan 7 MST (Gambar 9). Insidensi penyakit kerdil tergolong rendah, kemungkinan disebabkan populasi wereng batang cokelat sebagai vektornya di lapangan mengalami fluktuasi. Hal tersebut dapat terjadi karena adanya faktor lingkungan yang mempengaruhi populasi serangga vektor (Susanti et al. 2009), serta pola tanam yang mempengaruhi luasan penyakit (Baehaki 2011). Pola tanam padi di Desa Sukamandi termasuk monokultur dengan waktu tanam tidak serempak.
Gambar 9 Rata-rata insidensi penyakit kerdil pada petak pengamatan di Sukamandi pada periode pengamatan November – Desember 2014
Gejala penyakit yang ditemukan pada saat pengamatan terdiri atas gejala kerdil hampa, gejala kerdil rumput dan gejala campuran kerdil hampa dan kerdil rumput. Gejala tanaman kerdil sulit dikenali pada awal pertumbuhan tanaman. Pada pengamatan 1 MST sampai 4 MST tinggi tanaman sehat dengan tanaman yang terserang penyakit belum terlihat dengan jelas perbedaannya. Perbedaan tinggi tanaman sehat dan sakit mulai tampak jelas pada umur tanaman 5 MST.
Penghambatan tinggi tanaman tampak lebih jelas pada penyakit kerdil rumput dibandingkan dengan pada penyakit kerdil hampa (Gambar 10A dan B). Pada pengamatan 5 MST pengurangan tinggi tanaman yang terserang penyakit kerdil rumput sangat jelas dengan anakan yang sangat banyak, sedangkan penyakit kerdil hampa reduksi tanaman terlihat jelas setelah 10 MST. Selain itu juga ditemukan adanya infeksi ganda pada tanaman padi di lapangan, infeksi ganda penyakit kerdil hampa dan kerdil rumput yang ditemukan pada tanaman singgang. Gejala infeksi ganda kerdil rumput dan kerdil hampa menunjukkan adanya bagian daun yang terpelintir, dan bagian daun lainnya berwarna kuning dan terdapat
18
karat pada ujung daun serta daun menjadi lebih sempit (Gambar 10C). Du et al. (2005) melaporkan tanaman yang terinfeksi RRSV dan RGSV menunjukkan adanya pengkerdilan yang lebih parah dari infeksi tunggal. Biasanya gejala RRSV muncul pertama dengan anakan berwarna hijau gelap, lebih pendek, lebih tipis dan tegak dengan anakan kecil-kecil kemudian diikuti dengan munculnya gejala RGSV dimana daun muda menjadi lebih kecokelatan dan kadang terdapat daun tua yang menunjukkan adanya daun menguning. Tanaman singgang dapat terinfeksi virus, sehingga pembersihan lahan dari tanaman singgang perlu dilakukan untuk menghilangkan sumber inokulum dan menghindari penularan virus yang lebih luas pada musim tanam berikutnya (BB Padi 2010).
Gambar 10 Tanaman padi yang terserang penyakit kerdil diantara tanaman sehat. Gejala penyakit kerdil hampa pada umur tanaman 10 MST (A), gejala penyakit kerdil rumput pada umur tanaman 5 MST (B), gejala penyakit kerdil rumput dan kerdil hampa pada tanaman singgang (C) Terdapat perbedaan gejala yang jelas antara tanaman yang terserang penyakit kerdil hampa dan kerdil rumput. Tanaman yang terserang virus kerdil hampa memiliki gejala yang lebih bervariasi dibanding gejala serangan virus kerdil rumput pada tanaman padi (Gambar 11). Daun tetap berwarna hijau pucat, pada bagian ujung daun menggulung, pada beberapa tanaman gejala yang muncul pada daun terdapat sobekan-sobekan dan kerutan sehingga permukaan daun tampak kasar serta ada pembengkakan pada bagian bawah helai daun.
Tanaman padi yang ditemukan memiliki gejala kerdil hampa menunjukkan gejala yang khas, yaitu pengkerdilan, daun menjadi berwarna gelap dengan tepi bergerigi atau ujung memutar, dan tulang daun mengalami pembengkakan atau benjolan di bagian bawah helai daun dan bagian luar permukaan pelepah daun. Benjolan ini dihasilkan dari hiperplasia dan hipertropi pada jaringan floem (Cabautan et al. 2009).
19
Gambar 11 Penyakit kerdil hampa dengan gejala permukaan daun menjadi kasar karena adanya garis-garis melintang yang terbentuk (A), tepi daun terlihat bergelombang (B), adanya sobekan-sobekan pada daun (C), ujung daun menggulung (D), terdapat pembengkakan pada bagian pangkal helai daun (E)
Tanaman padi yang terserang virus kerdil rumput memiliki daun yang berwarna kuning, pada bagian daun terdapat bintik-bintik menyerupai karat serta daun menjadi sempit dan lebih kecil dibanding dengan daun tanaman sehat, selain itu tanaman yang terserang virus kerdil rumput akan memiliki anakan yang banyak sehingga menyerupai rumput (Gambar 12).
Gambar 12 Daun tanaman padi yang terserang penyakit kerdil rumput. Daun mengalami perubahan warna menguning dan perubahan ukuran daun yang menjadi lebih sempit dari daun sehat (A), terdapat bintik-bintik menyerupai karat pada bagian daun (B)
Hubungan populasi wereng batang cokelat dan insidensi penyakit kerdil
Peningkatan insidensi penyakit di lapangan juga dipengaruhi oleh keberadaan wereng di lapangan. Imago makroptera wereng batang cokelat memiliki peran penting dalam menyebarkan virus kerdil dimana imago wereng ini berperan
A B C D E
20
dalam migrasi jarak jauh dan tidak diketahui apakah wereng sudah atau belum membawa virus saat melakukan migrasi. Wereng berkembangbiak dengan cepat karena memiliki sifat yang mudah beradaptasi dengan lingkungan. Imago makroptera wereng batang coklat imigran berperan sebagai generasi nol lalu muncul nimfa sebagai generasi pertama dari wereng imigran. Dalam satu musim tanam wereng batang cokelat dapat menghasilkan populasi yang tinggi dari generasi nol sampai generasi ke 3 dengan cepat karena wereng batang cokelat merupakan salah satu hama yang berkembangbiak secara r-strategik (Baehaki 2011).
Pertambahan populasi wereng mengakibatkan semakin tinggi terjadinya insidensi penyakit kerdil di lapangan, karena selain imago makroptera, nimfa juga memegang peranan penting dalam menyebar dan menularkan virus kerdil pada tanaman padi dalam satu lahan. Hal tersebut dikarenakan nimfa merupakan serangga yang aktif menghisap cairan tanaman padi karena membutuhkan nutrisi yang cukup untuk perkembangannya sampai menjadi imago. Ketika nimfa makan pada tanaman yang terinfeksi oleh virus kerdil maka nimfa tersebut dapat menularkan virus seperti imago makroptera ataupun imago brakhiptera. Baehaki dan Widiarta (2005) melaporkan bahwa pemencaran wereng batang cokelat jarak pendek dalam pertanaman dilakukan oleh nimfa, brakhiptera ataupun makroptera. Pemencaran jarak pendek antar pertanaman dan pemencaran jarak jauh dilakukan oleh makroptera. Menurut Ghosh dan John (1980), nimfa wereng batang cokelat lebih efisien daripada imago dalam menularkan virus kerdil hampa. Persentase penularan nimfa wereng batang cokelat instar pertama, instar kedua, instar ketiga, instar keempat dan instar kelima berturut-turut adalah 19.3%, 25.8%, 29%, 27.6%, dan 21.4%; namun tidak diketahui kemampuan nimfa dalam menularkan penyakit kerdil rumput.
Meskipun di lapangan tidak terjadi ledakan populasi wereng batang cokelat namun populasi wereng batang cokelat tersebut mengalami fluktuasi dan dapat menularkan virus kerdil.. Pengamatan populasi wereng dilakukan pada masa tanam di musim penghujan (Gambar 13). Susanti et al. (2009) melaporkan bahwa curah hujan yang tinggi menyebabkan kelembapan udara meningkat sehingga menyebabkan populasi wereng batang cokelat juga meningkat. Daerah Sukamandi merupakan sentra padi dimana padi ditanam secara terus menerus tanpa adanya rotasi tanaman dan penanaman dilakukan secara tidak serempak sehingga tanaman padi selalu ada sebagai makanan wereng.
21
Gambar 13 Hubungan populasi wereng batang cokelat dengan insidensi penyakit kerdil tanaman padi pada pengamatan November - Desember 2014
Efisiensi Penularan Wereng Hijau dan Wereng Batang Cokelat sebagai Vektor Virus Padi
Periode Inkubasi
Hasil penelitian menunjukkan bahwa penyakit kerdil memiliki periode inkubasi yang relatif lebih singkat dibandingkan dengan penyakit tungro (Tabel 4). Gejala penyakit kerdil umumnya mulai muncul 7 hari setelah inokulasi (perlakuan 2, 4, 5, dan 7), sedangkan gejala penyakit tungro muncul antara 7 hari (perlakuan 3) sampai 14 hari (perlakuan 5). Kemampuan serangga vektor membawa virus dengan konsentrasi yang tinggi dan menularkannya ke tanaman dapat menyebabkan munculnya gejala penyakit lebih cepat. Selain itu, virulensi virus yang tinggi dapat menyebabkan proses infeksi yang lebih cepat (Maule et al. 2007).
-2
22
Tabel 4 Periode inkubasi, insidensi penyakit dan keparahan penyakit tungro dan kerdil pada tanaman padi yang ditularkan berturut-turut dengan wereng hijau dan wereng batang cokelat
Perlakuana PI (hari) IP (%) KP (%)
Tungro Kerdil Tungro Kerdil Tungro Kerdil
1 (WH+T) 10.33 0 100 0 89.62 0
2 (WBC+K) 0 7 0 100 0 82.96
3 (WH+WBC+T) 7 0 100 0 94.06 0
4 (WH+WBC+K) 0 7 0 100 0 97.03
5 (WH+WBC+T+K) 14 7 76.67 100 65.58 81.47
6 (WH+T+K) 10 0 100 0 91.84 0
7 (WBC+T+K) 0 7 0 100 0 83.70
8 (WH+WBC) 0 0 0 0 0 0
9 (WH) 0 0 0 0 0 0
10 (WC) 0 0 0 0 0 0
aKeterangan : T, tungro; K, kerdil; WH, wereng hijau; WBC, wereng batang cokelat
Gambar 14 Gejala menguning pada daun tanaman padi yang terinfeksi virus tungro (A), gejala malformasi pada daun tanaman padi yang terinfeksi virus kerdil (B), gejala tanaman padi yang terinfeksi oleh virus Tungro dan virus Kerdil (C)
Virus kerdil bersifat persisten propagatif didalam tubuh wereng batang cokelat (Chen et al. 1997). Setelah wereng batang cokelat mengakuisisi virus kerdil, akan terjadi propagasi virus di dalam tubuh wereng sehingga konsentrasi virus akan bertambah. Berbeda dengan sifat virus tungro, yaitu non propagatif didalam tubuh wereng hijau. Tingginya konsentrasi virus didalam tubuh serangga vektor mengakibatkan lebih banyak pula virus yang ditularkan pada tanaman dan selanjutnya dapat mempercepat munculnya gejala (Reissig et al. 1986).
23
Insidensi Penyakit
Insidensi penyakit kerdil mencapai 100% pada semua perlakuan dengan sumber inokulum virus kerdil dan vektor wereng batang cokelat (perlakuan 2, 4, 5, dan 7); sedangkan insidensi penyakit tungro berkisar antara 76.67 % sampai 100% pada perlakuan dengan sumber inokulum virus tungro dan vektor wereng hijau (perlakuan 1, 3, 5, dan 6) (Tabel 4). Data pengamatan insidensi penyakit dan periode inkubasi menunjukkan bahwa virus kerdil memiliki kemampuan yang cenderung lebih cepat menimbulkan penyakit di arena percobaan dibandingkan dengan virus tungro.
Dari hasil penularan virus tungro dengan serangga vektor wereng hijau menunjukkan bahwa insidensi penyakit tungro diketahui berbeda-beda dalam perkembangan setiap minggunya setelah inokulasi. Berbeda dengan insidensi penyakit tungro, insidensi penyakit kerdil menunjukkan perkembangan yang lebih cepat. Terlihat bahwa pada minggu ke-tiga setelah inokulasi semua perlakuan menunjukkan persentase 100% (Lampiran 1).
Insidensi penyakit dipengaruhi oleh perbedaan antara kemampuan vektor menularkan virus secara semi persisten dan persisten ke tanaman (Foreres dan Moreno 2009). Virus tungro ditularkan secara semi persisten non sirkulatif sedangkan penyakit kerdil ditularkan ketanaman secara persisten propagatif., Meskipun populasi vektornya sama dalam setiap perlakuan tetapi kemampuan vektor dalam menularkan virus berbeda-beda.
Keparahan Penyakit
Demikian pula halnya dengan keparahan penyakit, secara umum infeksi virus kerdil menyebabkan keparahan penyakit yang lebih berat dibandingkan dengan virus tungro (Tabel 4). Keparahan penyakit tungro yang tertinggi terjadi pada perlakuan 3 (97.03%), sedangkan yang terendah terjadi pada perlakuan 5 (65.58%) yaitu perlakuan dengan sumber penyakit terdiri atas 2 virus dan serangga vektor terdiri atas 2 spesies wereng. Keadaan yang sama juga terjadi pada penyakit kerdil, yaitu keparahan penyakit terendah terjadi pada perlakuan 5 (81.47%), sedangkan yang tertinggi pada perlakuan 4 (97.03%). Berdasarkan hasil pengamatan, perkembangan keparahan penyakit pada perlakuan 3 dan 4 menunjukkan tingkat keparahan tertinggi untuk masing-masing virus tungro dan virus kerdil (Lampiran 2). Hal ini terjadi karena serangga non vektor dalam perlakuan tersebut membantu meningkatkan kerentanan tanaman terhadap infeksi penyakit dengan cara menghisap cairan di tanaman.
Tingkat keparahan penyakit tersebut ditentukan antara lain oleh virulensi virus dan respons tanaman inang. Manzila et al. (2013) menyatakan bahwa kejadian dan keparahan penyakit merupakan hasil interaksi antara beberapa faktor, di antaranya isolat/galur virus, varietas padi, dan kondisi lingkungan di pertanaman termasuk keberadaan serangga vektor. Selain itu Widiarta et al. (2004) melaporkan bahwa suatu varietas padi dapat memberikan respons ketahanan yang sama terhadap infeksi galur virus tertentu walaupun memiliki latar belakang gen ketahanan yang berbeda. Dilaporkan oleh Asrori et al. (2014) bahwa varietas IR 64 merupakan varietas yang moderat tahan terhadap penyakit tungro, tetapi belum diketahui responsnya terhadap penyakit kerdil.