GEN cry UNTUK PENGELOLAAN HAMA PENGGEREK
BATANG PADI KUNING Scirpophaga incertulas (WALKER)
(LEPIDOPTERA: PYRALIDAE)
N. USYATI
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
DAN SUMBER INFORMASI
Dengan ini saya menyatakan bahwa disertasi “Keefektifan Padi Transgenik yang Mengandung Gen cry Untuk Pengelolaan Hama Penggerek Batang Padi Kuning Scirpophaga incertulas (Walker) (Lepidoptera: Pyralidae)” adalah hasil penelitian saya, dengan arahan dari komisi pembimbing selama mengikuti program S3 di Sekolah Pascasarjana, Institut Pertanian Bogor. Hasil penelitian ini belum pernah dipublikasi dalam bentuk apapun dan kemana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka yang ada dalam disertasi ini.
Bogor, 23 Agustus 2010
N. USYATI. The Effectiveness of Transgenic Rice Containing cry Gene to Control The Rice Yellow Stemborer Scirpophaga incertulas (Walker) (Lepidoptera: Pyralidae). Under supervision of DAMAYANTI BUCHORI, SYAFRIDA MANUWOTO, PURNAMA HIDAYAT, and INEZ HORTENSE SLAMET LOEDIN.
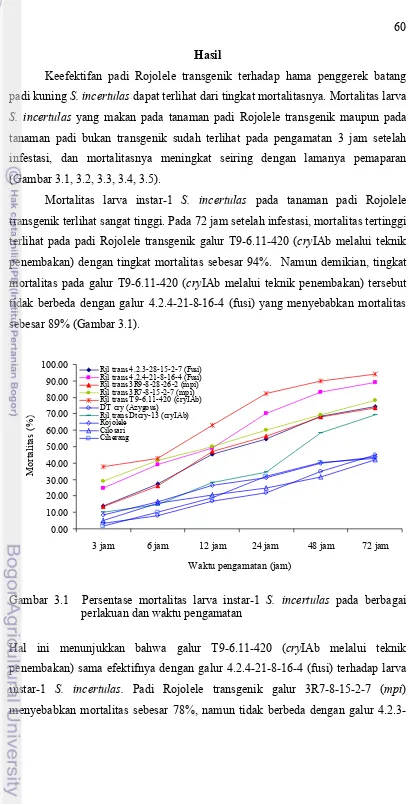
Rice stemborer is one of the major pests on rice plant in Indonesia. Transgenic rice plant that is resistant to rice stemborer is an alternative to control this pest. However, uses of transgenic crops has raised debate about their potential impact on the environment, such as on natural enemies. This situation rendez the importance doing a research assessment of transgenic rice to environment and natural enemies. The researches were conducted with the purpose: (1) to study the effectiveness of transgenic Rojolele rice to the rice yellow stemborer Scirpophaga incertulas; (2) to study larvae development and survival of insect predator Verania lineata in transgenic Rojolele rice; (3) to study nontarget impact of transgenic Rojolele rice. These studies were conducted at Indonesian Institute of Science, Cibinong-Bogor and at Indonesian Centre for Rice Research, Subang-West Java from November 2007–October 2009. The result showed that protoxin in transgenic Rojolele rice (trans Rjl) were highly effective in increasing the mortality of S. incertulas larvae and had high resistance compared to non transgenic rice varieties. There were differences of effectiveness and resistance value among protoxin in trans Rjl lines. Protoxin in trans Rjl T9-6.11-420 line was the highest (94% mortality; 0 scale) followed by 4.2.4-21-8-16-4 line (89% mortality; 1 scale), 3R7-8-15-2-7 line (78% mortality; 1 scale), 4.2.3-28-15-2-7 line (74.5% mortality; 3 scale), 3R9-8-28-26-2 line (73.5% mortality; 3 scale), and DTcry-13 line (69.5% mortality; 7 scale). DTcry (Azygous) line (45% mortality; 9 scale) was susceptible. Protoxin in trans Rjl were effective in increasing the mortality of S. incertulas larvae, but its effectiveness declined gradually along the development of larvae growth. Protoxin in trans Rjl T9-6.11-420 line effective to suppress S. incertulas infestation in the field. At the vegetative stage, protoxin in trans Rjl T9-6.11-420 line could suppress S. incertulas infestation about 3-4 fold and at the generative stage about 5-57 fold compared to non insecticide treatment. The yield of trans Rjl T9-6.11-420 line was the lowest, i.e. it was only 1/7 than Rojolele. There were differences of larvae development and survival of insect predator V. lineata among trans Rjl lines. In trans Rjl T9-6.11-420 line and DTcry-13 line, life time, developmental stage, the number eclosion of adult female, adult weight, and survival of preimaginal and adult of insect predator V. lineata were consistantly low. In trans Rjl 4.2.3-28-15-2-7 line, 3R9-8-28-26-2 line, and 3R7-8-15-2-7 line had no consistant effect on larvae development and survival of insect predator V. lineata. DTcry (Azygous) line had no effect on larvae development and survival of insect predator V. lineata. Whereas trans Rjl 4.2.4-21-8-16-4 line had one effect in decreasing of adult weight of insect predator V. lineata. Based on method in this research, effect of protoxin in trans Rjl T9-6.11-420 line on nontarget pests and natural enemies in the field can not be concluded.
N. USYATI. Keefektifan Padi Transgenik yang Mengandung Gen cry Untuk Pengelolaan Hama Penggerek Batang Padi Kuning Scirpophaga incertulas (Walker) (Lepidoptera: Pyralidae). Dibimbing oleh DAMAYANTI BUCHORI, SYAFRIDA MANUWOTO, PURNAMA HIDAYAT, dan INEZ HORTENSE SLAMET LOEDIN.
" "
Penggerek batang padi merupakan salah satu hama utama pada pertanaman padi di Indonesia. Tanaman padi transgenik tahan penggerek batang padi adalah salah satu cara alternatif untuk mengendalikan hama ini. Namun demikian tanaman transgenik ini masih diperdebatkan terutama mengenai potensi pengaruhnya terhadap lingkungan, diantaranya terhadap musuh alami.
Penelitian dengan tujuan: (1) mempelajari tingkat keefektifan padi Rojolele transgenik terhadap hama penggerek batang padi kuning Scirpophaga incertulas; (2) mempelajari perkembangan pradewasa dan kemampuan hidup predator Verania lineata pada padi Rojolele transgenik; (3) mempelajari populasi serangga nontarget pada pertanaman padi Rojolele transgenik telah dilakukan pada November 2007-Oktober 2009. Penelitian ini di lakukan di laboratorium dan rumah kaca Biologi Molekuler Tanaman, Pusat Penelitian Bioteknologi-LIPI, Cibinong-Bogor serta di Kebun Percobaan Balai Besar Penelitian Tanaman Padi Pusakanegara, Kabupaten Subang, Jawa Barat. Penelitian keefektifan padi Rojolele transgenik terhadap hama penggerek batang padi kuning S. incertulas dilakukan dalam 3 tahap, yaitu tahap in vitro, in planta, dan lapangan terbatas. Materi penelitian yang digunakan terdiri atas 6 galur padi Rojolele transgenik, yaitu galur 4.2.3-28-15-2-7 dan 4.2.4-21-8-16-4 yang mengandung fusi dua gen cry (cryIB-cryIAa), galur 3R9-8-28-26-2 dan 3R7-8-15-2-7 yang mengandung gen mpi::cryIB, galur T9-6.11-420 yang mengandung gen cryIAb melalui teknik penembakan, galur DTcry (Azygous) yaitu segregan yang mengalami proses kultur jaringan dan tidak mengandung gen cry (null), dan galur DTcry-13 yang mengandung gen cryIAb melalui Agrobacterium, serta tanaman padi bukan transgenik yang meliputi varietas Rojolele, Cilosari, dan Ciherang.
dengan bertambah lanjutnya perkembangan larva.
Protoxin dalam padi Rojolele transgenik galur T9-6.11-420 (cryIAb melalui teknik penembakan) efektif dalam menekan serangan S. incertulas baik pada stadium vegetatif (sundep) maupun pada stadium generatif (beluk). Pada stadium vegetatif, protoxin dalam padi Rojolele transgenik galur T9-6.11-420 (cryIAb melalui teknik penembakan) mampu menekan serangan S. incertulas (sundep) sebesar 3-4 kali dan pada stadium generatif sebesar 5-75 kali jika dibandingkan dengan perlakuan tanpa insektisida. Keefektifan protoxin dalam padi Rojolele transgenik galur T9-6.11-420 (cryIAb melalui teknik penembakan) sama dengan perlakuan insektisida dengan bahan aktif karbofuran dan dimehipo. Namun demikian, dari sisi produksi padi Rojolele transgenik galur T9-6.11-420 (cryIAb melalui teknik penembakan) sangat rendah, yaitu hanya 1/7 jika dibandingkan dengan varietas Rojolele.
Ada perbedaan perkembangan pradewasa dan kemampuan hidup predator V. lineata antar galur padi Rojolele transgenik yang diuji. Pada padi Rojolele transgenik galur T9-6.11-420 (cryIAb melalui teknik penembakan) dan galur DTcry-13 (cryIAb melalui Agrobacterium), lama perkembangan, keberhasilan dalam mencapai setiap stadium perkembangan, kemunculan imago betina, berat imago, dan kemampuan hidup pradewasa dan dewasa predator V. lineata konsisten rendah. Pada padi Rojolele transgenik galur 4.2.3-28-15-2-7 (fusi), 3R9-8-28-26-2 (mpi), dan 3R7-8-15-2-7 (mpi), perkembangan pradewasa dan kemampuan hidup predator V. lineata tidak konsisten. Pada galur DTcry (Azygous), perkembangan pradewasa dan kemampuan hidup predator V. lineata konsisten tidak berbeda dibandingkan dengan padi bukan transgenik varietas Rojolele. Padi Rojolele transgenik galur 4.2.4-21-8-16-4 (fusi) hanya berpengaruh dalam pengurangan berat imago predator V. lineata.
Dengan metode yang digunakan pada penelitian ini belum bisa diambil kesimpulan mengenai pengaruh protoxin dalam padi Rojolele transgenik galur T9-6.11-420 (cryIAb melalui teknik penembakan) terhadap serangga hama nontarget dan musuh alami di lapangan.
Kata kunci: keefektifan, padi transgenik, S. incertulas, hama nontarget, musuh alami
© Hak cipta milik Institut Pertanian Bogor, tahun 2010
Hak cipta dilindungi Undang Undang
1. Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumber
a. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik atau tinjauan suatu masalah
b. Pengutipan tidak merugikan kepentingan yang wajar IPB
GEN cry UNTUK PENGELOLAAN HAMA PENGGEREK
BATANG PADI KUNING Scirpophaga incertulas (WALKER)
(LEPIDOPTERA: PYRALIDAE)
N. USYATI
Disertasi
Sebagai salah satu syarat untuk memperoleh gelar Doktor
pada
Program Studi Entomologi-Fitopatologi
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Penguji Luar Komisi pada Ujian Tertutup: Dr.Ir. Hermanu Triwidodo, M.Sc (Staf Pengajar Departemen Proteksi Tanaman,
Fakultas Pertanian, IPB)
Dr.Ir. Sutrisno
(Staf Peneliti BB Biogen, Badan Litbang Pertanian, Kementerian Pertanian)
Penguji Luar Komisi pada Ujian Terbuka: Dr. Ir. I Wayan Winasa, M.Si (Staf Pengajar Departemen Proteksi Tanaman,
Fakultas Pertanian, IPB)
Prof. Dr. Ir. Baehaki SE, APU (Staf Peneliti BB Padi,
Gen cry Untuk Pengelolaan Hama Penggerek Batang Padi Kuning Scirpophaga incertulas
(Walker) (Lepidoptera: Pyralidae)
Nama Mahasiswa : N. Usyati
NIM : A461060071
Disetujui
Komisi Pembimbing
Dr.Ir. Damayanti Buchori, M.Sc Prof. Dr. Ir. Syafrida Manuwoto, M.Sc Ketua Anggota
Dr.Ir. Purnama Hidayat, M.Sc Dr.Ir. Inez Hortense Slamet Loedin, M.Sc Anggota Anggota
Diketahui
Ketua Program Studi Dekan Sekolah Pascasarjana Entomologi-Fitopatologi
Dr. Ir. Pudjianto, M.S Prof.Dr.Ir. Khairil A. Notodiputro, M.S
Puji dan syukur penulis panjatkan kepada Allah SWT karena dengan rakhmat dan hidayah-Nya maka disertasi yang berjudul “Keefektifan Padi Transgenik yang Mengandung Gen cry Untuk Pengelolaan Hama Penggerek Batang Padi Kuning Scirpophaga incertulas (Walker) (Lepidoptera: Pyralidae)” dapat terselesaikan.
Penelitian dan penulisan disertasi dapat terlaksana atas bimbingan dan arahan komisi pembimbing yaitu Dr. Ir. Damayanti Buchori, M.Sc sebagai Ketua, Prof. Dr. Ir. Syafrida Manuwoto, M.Sc, Dr. Ir. Purnama Hidayat, M.Sc, dan Dr. Ir. Inez Hortense Slamet Loedin, M.Sc masing-masing selaku Anggota Komisi Pembimbing, untuk semua bimbingan dan arahan tersebut penulis mengucapkan terima kasih.
Penulis juga mengucapkan terima kasih kepada Kepala Badan Litbang Pertanian, Ketua Komisi Pembinaan Tenaga Badan Litbang Pertanian, Kepala Puslitbangtan, Kepala Balai Besar Penelitian Tanaman Padi, Ketua Kelompok Peneliti Entomologi-Fitopatologi BB Padi, dan atasan langsung Ibu Ir. H. Hendarsih Suharto, M.Sc yang telah menugaskan dan memberi kesempatan kepada penulis untuk melanjutkan pendidikan, serta Pemimpin dan staf Bendahara Badan Litbang Pertanian yang telah membantu mempermudah penyaluran dana pendidikan penulis.
Kepada Kepala Puslit Bioteknologi LIPI dan Kepala Bidang Biologi Molekuler Puslit Bioteknologi LIPI yang telah memberi izin dan kesempatan penulis untuk melakukan penelitian dan mengerjakan sebagian dari proyek penelitian yang dibiayai dana DIPA Puslit Bioteknologi LIPI penulis juga mengucapkan terima kasih.
APU (BB Padi), penulis mengucapkan terima kasih atas kesediaannya untuk menjadi penguji pada ujian terbuka.
Kepada Ketua Program Studi Entomologi-Fitopatologi dan semua staf dosen IPB penulis mengucapkan terima kasih atas ilmu yang telah diberikan, kepada staf administrasi dan teknisi laboratorium Departemen Proteksi Faperta IPB penulis mengucapkan terima kasih atas bantuan dalam kelancaran studi di IPB. Penulis juga mengucapkan terima kasih kepada Dr. Ir. Ali Nurmansyah, M.Si atas bantuan dalam analisis data.
Ucapan terima kasih penulis sampaikan kepada Kepala Kebun Percobaan Pusakanegara Atito Dirdjoseputro, SP atas penyediaan lahan penelitian dan kepada Kepala Koperasi BB Padi Ir. Dede Kusdiaman penulis mengucapkan terima kasih atas segala bantuannya.
Kepada Ketua Dewan Redaksi dan Redaksi Pelaksana Jurnal Penelitian Pertanian Tanaman Pangan yaitu Prof Dr. A. Karim Makarim, Drs. Hermanto, dan Edi Hikmat, SE, penulis mengucapkan terima kasih atas bantuannya dalam publikasi karya ilmiah.
Ucapan terima kasih juga penulis sampaikan kepada semua teman-teman peneliti (Dr. Satya Nugraha, Dr. Amy Estiati, Syamsidah Rachmawati, M.Si, Enung Sri Mulyaningsih, M.Si, Agus Rachmat, M.Si, Yuli Sulistyowati, M.Si, Fatimah Azzahra, M.Si, dan Sri Indrayani, S.Si) dan teknisi (Budi Satrio, M. Taufik Hidayat, Pak Tohar, dan Pak Adang) di Laboratorium dan Rumah Kaca Biologi Molekuler LIPI yang telah membantu dalam pelaksanaan penelitian di laboratorium dan rumah kaca, dan teman-teman peneliti dan teknisi Kelti Entomologi-Fitopatologi BB Padi terutama kepada Nia Kurniawati SP, Undang Suryana, Oco Rumasa, Oo, dan Toha atas bantuan dalam kelancaran penelitian di lapangan dan pemeliharaan serangga uji.
Ucapan terima kasih juga penulis sampaikan kepada teman-teman Program Studi Entomologi-Fitopatologi IPB terutama kepada Rahmini, M.Si dan Dra. Ifa Manzila, M.Si atas dukungannya kepada penulis.
penulis dapat menyelesaikan pendidikan.
Akhirnya penulis berharap mudah-mudahan tulisan ini dapat bermanfaat bagi kita semua.
Bogor, 23 Agustus 2010
Penulis dilahirkan di Cirebon pada tanggal 07 Agustus 1968 dari ayah H. Hasyim dan ibu Hj. Aisyah (Alm). Penulis merupakan putri pertama dari tujuh bersaudara.
Halaman
DAFTAR TABEL ………... xvi
DAFTAR GAMBAR ... xviii
DAFTAR LAMPIRAN ... xix
BAB I PENDAHULUAN ………... 1
Latar Belakang ...………... 1
Perumusan Masalah ……...………... 4
Tujuan Penelitian ………... 6
Manfaat Penelitian ………... 6
Ruang Lingkup Penelitian ………... 7
Kerangka Pemikiran ………... 14
Daftar Pustaka ... 16
BAB II TINJAUAN PUSTAKA ………... 19
Ketahanan Tanaman ... 19
Strategi Pengendalian Serangga dengan Rekayasa Genetika ... 21
Gen cry, Penyandi δ-Endotoksin dari Bacillus thuringiensis ... 23
Proteinase Inhibitor ... 25
Maize Proteinase Inhibitor (mpi) dan Regulasinya... 25
Penggerek Batang Padi ... 26
Biologi Penggerek Batang Padi Kuning S. incertulas ... 27
Predator V. lineata ... 28
Interaksi Tritropik ... 29
Pengaruh Tanaman Transgenik Tahan Hama Terhadap Musuh Alami ... 35
Tanaman Transgenik Komersial ... 46
MENGANDUNG GEN cry UNTUK PENGELOLAAN HAMA PENGGEREK BATANG PADI KUNING Scirpophaga incertulas (WALKER) (LEPIDOPTERA:
PYRALIDAE) PADA TAHAP IN VITRO ... 53
Abstrak ... 53
Abstract ... 54
Pendahuluan ... 54
Bahan dan Metode ... 56
Hasil ... 60
Pembahasan ... 64
Kesimpulan ... 70
Daftar Pustaka ... 71
BAB IV KEEFEKTIFAN PADI TRANSGENIK YANG MENGANDUNG GEN cry UNTUK PENGELOLAAN HAMA PENGGEREK BATANG PADI KUNING Scirpophaga incertulas (WALKER) (LEPIDOPTERA: PYRALIDAE) PADA TAHAP IN PLANTA ... 75
Abstrak ... 75
Abstract ... 76
Pendahuluan ... 76
Bahan dan Metode ... 78
Hasil ... 82
Pembahasan ... 86
Kesimpulan ... 90
Daftar Pustaka ... 90
BAB V KEEFEKTIFAN PADI TRANSGENIK YANG MENGANDUNG GEN cry UNTUK PENGELOLAAN HAMA PENGGEREK BATANG PADI KUNING Scirpophaga incertulas (WALKER) (LEPIDOPTERA: PYRALIDAE) PADA TAHAP LAPANGAN TERBATAS... 93
Abstrak ... 93
Abstract ... 94
Pendahuluan ... 94
Pembahasan ... 103
Kesimpulan ... 105
Daftar Pustaka ... 106
BAB VI PERKEMBANGAN PRADEWASA DAN KEMAM-PUAN HIDUP PREDATOR Verania lineata THURNBERG (COLEOPTERA: COCCINELLIDAE) PADA PADI TRANSGENIK ... 109
Abstrak ... 109
Abstract ... 110
Pendahuluan ... 110
Bahan dan Metode ... 112
Hasil ... 116
Pembahasan ... 124
Kesimpulan ... 127
Daftar Pustaka ... 127
BAB VII POPULASI SERANGGA NONTARGET PADA PERTANAMAN PADI TRANSGENIK ... 130
Abstrak ... 130
Abstract ... 131
Pendahuluan ... 131
Bahan dan Metode ... 133
Hasil ... 135
Pembahasan ... 145
Kesimpulan ... 147
Daftar Pustaka ... 147
BAB VIII PEMBAHASAN UMUM ... 150
Daftar Pustaka ... 154
BAB IX KESIMPULAN DAN SARAN ... 156
Kesimpulan ... 156
Halaman 2.1 Pengaruh tanaman transgenik tahan hama terhadap musuh
alami ... 40 2.2 Pengaruh tanaman transgenik tahan hama terhadap musuh
alami ... 41 2.3 Pengaruh tanaman transgenik tahan hama terhadap musuh
alami ... 42 2.4 Lanjutan pengaruh tanaman transgenik tahan hama terhadap
musuh alami ... 43 2.5 Lanjutan pengaruh tanaman transgenik tahan hama terhadap
musuh alami ... 44 2.6 Lanjutan pengaruh tanaman transgenik tahan hama terhadap
musuh alami ... 45 2.7 Luas area global tanaman transgenik pada tahun 2009 ... 47 4.1 Rata-rata intensitas serangan S. incertulas pada 2 minggu
setelah infestasi dan nilai ketahanan tanaman padi pada
berbagai perlakuan ... 83 4.2 Rata-rata intensitas serangan S. incertulas pada 4 minggu
setelah infestasi dan nilai ketahanan tanaman padi pada
berbagai perlakuan ... 84 4.3 Rata-rata bobot pupa S. incertulas pada 4 minggu setelah
infestasi pada berbagai perlakuan ... 85 4.4 Jumlah salinan gen (copy number) pada berbagai perlakuan .... 88 6.1 Rata-rata lama perkembangan predator V. lineata pada
berbagai perlakuan dan stadia perkembangan ... 118 6.2 Persentase individu predator V. lineata yang gagal mencapai
perkembangan pada berbagai perlakuan dan stadia
perkembangan ... 119 6.3 Persentase individu predator V. lineata yang berhasil
mencapai perkembangan pada berbagai perlakuan dan stadia
perkembangan ... 120 7.1 Rata-rata jumlah individu hama nontarget pada berbagai
varietas padi dan waktu pengamatan ... 137 7.2 Rata-rata jumlah individu musuh alami pada berbagai varietas
Halaman 1.1 Diagram latar belakang kerangka pemikiran penelitian ... 14 1.2 Diagram kerangka pemikiran penelitian ... 15 3.1 Persentase mortalitas larva instar-1 S. incertulas pada
berbagai perlakuan dan waktu pengamatan ... 60 3.2 Persentase mortalitas larva instar-2 S. incertulas pada
berbagai perlakuan dan waktu pengamatan ... 61 3.3 Persentase mortalitas larva instar-3 S. incertulas pada
berbagai perlakuan dan waktu pengamatan ... 62 3.4 Persentase mortalitas larva instar-4 S. incertulas pada
berbagai perlakuan dan waktu pengamatan ... 63 3.5 Persentase mortalitas larva instar-5 S. incertulas pada
berbagai perlakuan dan waktu pengamatan ... 64 3.6 Persentase mortalitas larva instar 1-5 S. incertulas pada
berbagai perlakuan dan waktu pengamatan ... 69 3.7 Gejala larva S. incertulas yang makan pada tanaman padi
transgenik dan tanaman padi bukan transgenik ... 70 4.1 Pupa S. incertulas pada tanaman padi bukan transgenik ... 86 5.1 Rata-rata intensitas serangan S. incertulas pada tanaman padi
transgenik dan bukan transgenik pada stadium vegetatif ... 100 5.2 Rata-rata intensitas serangan S. incertulas pada tanaman padi
transgenik dan bukan transgenik pada stadium generatif ... 101 5.3 Rata-rata hasil panen pada tanaman padi transgenik dan bukan
transgenik ... 103 6.1 Persentase imago predator V. lineata yang muncul pada
berbagai perlakuan ... 121 6.2 Rata-rata berat imago predator V. lineata (total jantan dan
betina) yang muncul pada berbagai perlakuan ... 122 6.3 Rata-rata berat imago predator V. lineata jantan dan betina
yang muncul pada berbagai perlakuan ... 123 6.4 Persentase kemampuan hidup predator V. lineata pada
berbagai perlakuan dan stadia perkembangan ... 124 7.1 Populasi serangga hama nontarget pada berbagai varietas padi
dan waktu pengamatan ... 142 7.3 Populasi kelompok musuh alami pada berbagai perlakuan dan
Halaman 1 Deskripsi galur padi transgenik ... 159 2 Hasil uji PCR galur padi transgenik ... 160 3 Persentase mortalitas larva instar-1 S. incertulas pada berbagai
perlakuan dan waktu pengamatan ... 161 4 Persentase mortalitas larva instar-2 S. incertulas pada berbagai
perlakuan dan waktu pengamatan ... 162 5 Persentase mortalitas larva instar-3 S. incertulas pada berbagai
perlakuan dan waktu pengamatan ... 163 6 Persentase mortalitas larva instar-4 S. incertulas pada berbagai
perlakuan dan waktu pengamatan ... 164 7 Persentase mortalitas larva instar-5 S. incertulas pada berbagai
PENDAHULUAN
Latar Belakang
Penggerek batang padi sampai saat ini masih merupakan kendala utama peningkatan produksi padi di Indonesia. Pada tahun 2006, luas daerah yang terserang penggerek batang padi mencapai 112 950 ha atau 21.75% dari total luas serangan OPT seluas 519 200 ha (DIRJENTAN PANGAN 2007).
Untuk mengatasi masalah hama penggerek batang padi ini, beberapa teknik pengendalian telah diterapkan antara lain penangkapan ngengat dengan lampu perangkap (Rauf et al. 1992), pengumpulan kelompok telur, penggunaan karbofuran (Oka 1992), teknik pengendalian secara kultur teknis yaitu dengan mengatur waktu tanam dan pola tanam (Hendarsih 1994), dan teknik penangkapan massal dengan menggunakan feromon seks (Hendarsih & Beevor 2001), tetapi hasilnya masih belum memuaskan.
Varietas tahan hama sebagai salah satu komponen pengendalian hama terpadu (PHT) merupakan salah satu teknik pengendalian yang murah, mudah diterapkan, dan tidak mencemari lingkungan. Namun sampai saat ini belum ada varietas padi yang mempunyai ketahanan pada tingkat yang memadai untuk penggerek batang padi (Rao & Padhi 1988).
ketahanan kimiawi dari tingkat ketahanan toleran sampai moderat, tetapi tidak ada yang mempunyai ketahanan kimiawi pada tingkat ketahanan yang tinggi. Ketahanan morfologi untuk penggerek batang padi telah lama diketahui, tetapi tidak ada varietas yang dilepas berdasarkan sifat ketahanan morfologi ini dikembangkan. Upaya lain dengan metode pemuliaan secara konvensional dengan memanfaatkan zat antibiosis sangat sulit dilakukan karena di alam sumber gen ketahanan untuk penggerek batang padi belum ditemukan pada padi atau kerabat liarnya (Rao & Padhi 1988).
Rekayasa genetika tanaman merupakan alternatif yang diharapkan dapat membantu menjawab tantangan ini. Dengan rekayasa genetika, gen tahan untuk penggerek batang padi yang berasal dari spesies lain dapat diintroduksikan pada tanaman padi dengan menggunakan teknologi transfer gen.
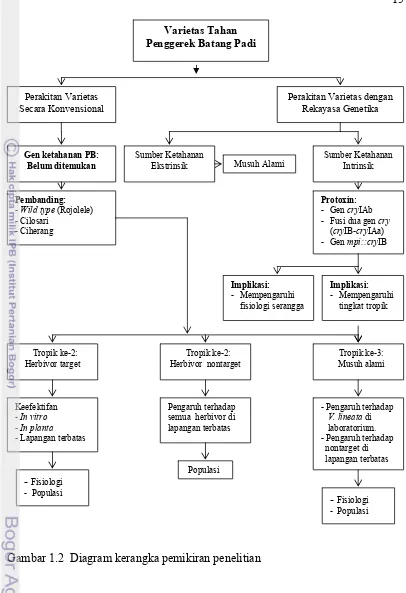
Teknik transformasi genetika yang bertujuan untuk ketahanan terhadap hama telah dikembangkan pada varietas padi Indonesia. Salah satu gen ketahanan yang telah berhasil disisipkan ke varietas Rojolele (Javanica) adalah gen cryIAb yang berasal dari bakteri Bacillus thuringiensis penyandi sumber ketahanan terhadap hama dari ordo lepidoptera melalui teknik penembakan (Slamet-Loedin et al. 1998). Berdasarkan pengujian sampai dengan generasi kelima, terbukti bahwa gen cryIAb tetap diwariskan pada setiap generasinya dan ditemukan beberapa galur potensial tahan penggerek batang padi (Satoto 2003). Sementara itu dari hasil pengujian di lapangan terbatas pada musim tanam tahun 2003 dan 2004 telah diperoleh 5 galur padi transgenik cv. Rojolele mengandung gen cryIAb tahan penggerek batang padi kuning S. incertulas.
penggerek batang padi kuning S. incertulas dan bergaris Chilo suppressalis (Walker) (Lepidoptera: (Pyralidae) (Breitler et al. 2000).
Namun seperti halnya hasil pemuliaan konvensional, ketahanan tanaman transgenik dapat dipatahkan. Berbagai strategi dikembangkan untuk memperlambat laju pematahan ketahanan oleh serangga. Cohen (2000) menyarankan untuk hanya menanam tanaman transgenik yang mengandung “high dose” toxin, menanam tanaman non transgenik diantara tanaman transgenik (refugia), atau menanam tanaman yang mengandung lebih dari satu gen ketahanan yang mempunyai “binding site” yang berbeda di dalam pencernaan larva serangga. Alcantra et al. (2000) melaporkan beberapa toxin cry mempunyai “binding site” yang berbeda di dalam pencernaan larva dua spesies penggerek batang padi, C. suppressalis dan S. incertulas. Selanjutnya mereka menyarankan untuk mengkombinasikan penggunaan toxin cry yang berbeda binding site ini untuk memperoleh tanaman transgenik yang memiliki ketahanan yang lama. Misalnya kombinasi gen cryIAb atau cryIAc dengan gen cryIC, cry2A, atau cry9c. Penemuan ini sangat menguntungkan dalam mengembangkan strategi untuk memperlambat laju pematahan ketahanan oleh serangga, karena kemungkinan mutasi dua receptor yang berbeda pada serangga lebih kecil.
Untuk mendapatkan padi tahan penggerek batang padi yang memiliki ketahanan yang panjang (tidak mudah patah), dua pendekatan telah dilakukan oleh Kelompok Penelitian Padi, Puslit Bioteknologi LIPI yaitu (a) transformasi dua gen cry (cryIB-cryIAa) yang mempunyai binding site berbeda dalam sistem pencernaan larva serangga, dan (b) transformasi gen cryIB dibawah kendali promoter terinduksi pelukaan yaitu promoter dari gen maize proteinase inhibitor (mpi). Dari hasil penelitian pada tahun 2003 dan 2004 pada generasi pertama dan kedua, telah diperoleh 2 galur padi transgenik cv. Rojolele mengandung fusi dua gen cry (cryIB-cryIAa)dan 4 galur padi transgenik cv. Rojolele mengandung gen mpi::cryIB potensial tahan penggerek batang padi kuning S. incertulas yang bersegregasi mengikuti hukum Mendel (3:1) (Rahmawati 2004).
dapat mengeliminasi gen penyeleksi antibiotik, meskipun antibiotik yang digunakan masuk dalam kategori aman (Rachmat 2006).
Dengan didapatkannya padi transgenik tahan penggerek batang padi, bukan berarti langsung padi tersebut dapat dilepas ke pasar. Sebelum suatu produk rekayasa genetika dilepas maka sebelumnya, sesuai kesepakatan pada protokol keamanan hayati global (Protocol Cartagena) perlu dilakukan analisis resiko untuk keamanan lingkungan dan keamanan pangan. Di Indonesia hal tersebut diatur oleh Peraturan Pemerintah Republik Indonesia Nomor 21 Tahun 2005 (Tentang Keamanan Hayati Produk Rekayasa Genetik) dan Surat Keputusan Bersama (SKB) 4 Menteri Tahun 1999 (Menteri Pertanian, Menteri Kehutanan dan Perkebunan, Menteri Kesehatan, dan Menteri Negara Pangan dan Hortikultura) (Tentang Keamanan Hayati dan Keamanan Pangan Produk Pertanian Hasil Rekayasa Genetik). Untuk itu ditingkat Nasional dibentuk Komisi Keamanan Hayati dan Pangan (KKHP) yang secara teknis dibantu oleh Tim Teknis Keamanan Hayati dan Keamanan Pangan (TTKHP). Hal ini dilakukan untuk meminimalisasi dampak negatif yang mungkin ditimbulkan oleh produk transgenik terhadap keamanan lingkungan dan kesehatan manusia. Aspek yang perlu dipelajari dari pengujian keamanan lingkungan diantaranya adalah menganalisis pengaruh padi transgenik terhadap serangga nontarget.
Perumusan Masalah
Penggerek batang padi sampai saat ini masih merupakan kendala utama peningkatan produksi padi di Indonesia. Untuk mengatasi masalah hama penggerek batang padi ini, beberapa teknik pengendalian telah diterapkan antara lain pengendalian secara fisik, mekanis, kimiawi, kultur teknis yaitu dengan mengatur waktu tanam dan pola tanam, serta teknik penangkapan massal dengan menggunakan feromon seks, tetapi hasilnya masih belum memuaskan.
Upaya untuk mendapatkan varietas padi tahan hama penggerek batang padi telah lama dilakukan dengan metode pemuliaan secara konvensional dengan memanfaatkan pemahaman mekanisme ketahanan genetik tanaman terhadap serangga hama. Mekanisme tersebut adalah mekanisme ketahanan genetik nonpreference atau antixenosis, baik antixenosis kimiawi maupun antixenosis morfologi. Dengan cara ini telah teridentifikasi galur-galur padi yang mempunyai ketahanan kimiawi dari tingkat ketahanan toleran sampai moderat, tetapi tidak ada yang mempunyai ketahanan kimiawi pada tingkat ketahanan yang tinggi. Ketahanan morfologi untuk penggerek batang padi telah lama diketahui, tetapi tidak ada varietas yang dilepas berdasarkan sifat ketahanan morfologi ini dikembangkan. Upaya lain dengan memanfaatkan zat antibiosis sangat sulit dilakukan karena di alam sumber gen ketahanan untuk penggerek batang padi belum ditemukan pada padi atau kerabat liarnya.
Alternatif lain untuk mendapatkan varietas padi tahan hama penggerek batang padi adalah dengan dilakukannya introduksi gen tahan penggerek batang padi yang berasal dari spesies lain menggunakan teknologi transfer gen melalui rekayasa genetika. Teknologi DNA rekombinan merupakan upaya untuk dapat melakukan transfer gen antar spesies. Gen ketahanan yang digunakan dalam penelitian ini adalah gen yang dapat memfasilitasi produksi bahan pestisida alami yaitu gen cry yang berasal dari bakteri B. thuringiensis.
Hipotesis
Hipotesis nol (H0) dalam penelitian ini adalah:
1. Tidak ada perbedaan keefektifan padi Rojolele transgenik terhadap hama penggerek batang padi kuning S. incertulas.
2. Tidak ada perbedaan perkembangan pradewasa dan kemampuan hidup predator Verania lineata Th (Coleoptera: Coccinellidae) pada padi Rojolele transgenik.
3. Tidak ada perbedaan populasi serangga nontarget pada pertanaman padi Rojolele transgenik.
Hipotesis satu (H1) dalam penelitian ini adalah:
1. Ada perbedaan keefektifan padi Rojolele transgenik terhadap hama penggerek batang padi kuning S. incertulas.
2. Ada perbedaan perkembangan pradewasa dan kemampuan hidup predator V. lineata pada padi Rojolele transgenik.
3. Ada perbedaan populasi serangga nontarget pada pertanaman padi Rojolele transgenik.
Tujuan Penelitian
Tujuan penelitian ini adalah:
1. Mempelajari tingkat keefektifan padi Rojolele transgenik terhadap hama penggerek batang padi kuning S. incertulas.
2. Mempelajari perkembangan pradewasa dan kemampuan hidup predator V. lineata pada padi Rojolele transgenik.
3. Mempelajari populasi serangga nontarget pada pertanaman padi Rojolele transgenik.
Manfaat Penelitian
hama penggerek batang padi kuning S. incertulas dan begitu pula dapat diperoleh informasi mengenai pengaruh tanaman transgenik terhadap lingkungan hidup termasuk organisme bukan sasaran.
Ruang Lingkup Penelitian Penggerek Batang Padi
Keterangan:
Pengendalian Hama
Pengendalian hama terpadu (PHT) adalah teknik pengendalian yang dianjurkan untuk mengatasi masalah hama di lahan pertanian. Dalam penerapannya di lapangan, PHT tidak tergantung hanya pada satu jenis teknik pengendalian hama, tetapi semua teknik pengendalian harus dimanfaatkan dan dipadukan agar dapat menekan populasi hama berada di bawah Ambang Ekonomi. Teknik-teknik pengendalian hama tersebut termasuk pengendalian secara fisik, pengendalian mekanik, pengendalian secara budidaya tanaman, pengendalian hayati, pengendalian kimiawi, penggunaan varietas tanaman tahan hama, dan teknik-teknik pengendalian hama lainnya, termasuk peraturan (Untung 2006).
Pengendalian fisik merupakan usaha mengubah faktor lingkungan fisik sehingga dapat menimbulkan mortalitas dan mengurangi populasi hama. Mortalitas hama disebabkan karena faktor fisik seperti suhu, kelembaban, dan suara diluar batas toleransi serangga hama sasaran. Pengendalian mekanik bertujuan mematikan atau memindahkan hama secara langsung baik dengan tangan atau dengan bantuan alat dan bahan lain. Cara pengendalian fisik dan mekanik ini tidak menimbulkan pengaruh negatif pada lingkungan.
Pengendalian hama secara budidaya atau bercocok tanam bertujuan mengelola lingkungan budidaya tanaman sedemikian rupa sehingga menjadi kurang cocok bagi kehidupan dan perkembangan hama serta dapat mengurangi laju peningkatan populasi hama dan kerusakan tanaman. Teknik pengendalian ini tidak mengakibatkan pencemaran lingkungan dan relatif mudah dikerjakan oleh petani baik secara perorangan maupun secara berkelompok.
terhadap serangga atau organisma bukan sasaran, karena musuh alami biasanya adalah khas inang.
Pengendalian kimiawi adalah teknik pengendalian hama dengan menggunakan bahan kimiawi seperti insektisida. Pengendalian kimiawi tidak aman bagi lingkungan karena memiliki dampak samping terhadap lingkungan seperti hama menjadi resisten, resurjensi atau ledakan hama sekunder, pengaruh negatif terhadap organisma nontarget, dan residu pestisida (Metcalf 1982; Sutanto 2002).
Tanaman tahan hama merupakan teknik pengendalian yang efektif, murah, dan kurang berbahaya bagi lingkungan. Landasan yang digunakan dalam pengendalian ini adalah pemahaman tentang sifat ketahanan yang dimiliki oleh tanaman yaitu ketahanan ekologi dan ketahanan genetik (Kogan 1982). Ketahanan ekologi merupakan sifat ketahanan tanaman yang tidak dikendalikan oleh faktor genetik tetapi sepenuhnya oleh faktor lingkungan yang memungkinkan munculnya penampakan sifat ketahanan tanaman terhadap hama tertentu. Karena sifatnya yang tidak tetap, ahli pemulia tanaman tidak mengakui sifat ini sebagai sifat ketahanan tanaman yang sesungguhnya. Sifat ketahanan ini biasanya merupakan sifat sementara dan dapat terjadi pada tanaman yang sebenarnya peka terhadap serangan hama tertentu. Ketahanan genetik merupakan sifat ketahanan tanaman yang dikendalikan oleh faktor genetik. Menurut Painter (1951) dalam Untung (2006) terdapat 3 mekanisme ketahanan genetik tanaman terhadap serangga hama yaitu (1) ketidaksukaan atau nonpreference, (2) antibiosis, dan (3) toleran.
oleh adanya sifat-sifat struktur atau morfologi tanaman yang dapat menghalangi terjadinya proses makan dan peletakan telur yang normal.
Antibiosis adalah semua pengaruh fisiologi pada serangga yang merugikan, bersifat sementara atau tetap, sebagai akibat kegiatan serangga memakan dan mencerna jaringan atau cairan tanaman tertentu. Gejala penyimpangan fisiologi terlihat apabila suatu serangga dipindahkan dari tanaman tidak memiliki sifat antibiosis ke tanaman yang memiliki sifat tersebut. Penyimpangan fisiologi tersebut berkisar mulai dari penyimpangan yang sedikit sampai penyimpangan terberat yaitu terjadinya kematian serangga. Mekanisme antibiosis sampai saat ini merupakan mekanisme ketahanan tanaman yang paling penting dan banyak dicari dan dimanfaatkan oleh ahli pemulia tanaman sebagai sumber gen untuk digunakan guna memperoleh varietas baru yang tahan hama. Banyak contoh zat antibiosis yang dijumpai pada tanaman yang telah dimanfaatkan untuk mendapatkan varietas tahan hama seperti: (a) kandungan gosipol pada kapas untuk memperoleh ketahanan kapas terhadap hama H. armigera, (b) kandungan asparagin pada padi untuk ketahanan padi terhadap hama wereng coklat padi, (c) kandungan 2,4-hydroxy-7-methoxy-2H-1,4-benzoxazin-3(4H)-one (DIMBOA) pada jagung untuk memperoleh ketahanan terhadap penggerek batang jagung Ostrinia.
Mekanisme ketahanan toleran terjadi karena adanya kemampuan tanaman tertentu untuk sembuh dari luka yang diderita karena serangan hama atau mampu tumbuh lebih cepat sehingga serangan hama kurang mempengaruhi hasil dibandingkan dengan tanaman lain yang lebih peka.
mempengaruhi pertumbuhan dan perkembangan larva, serta mengurangi lama hidup dan fekunditas serangga dewasa. Sementara mekanisme toleran tidak jelas pengaruhnya terhadap serangga.
Penggunaan varietas tahan terbukti mampu mengurangi tingkat serangan hama sehingga hasil panen dapat meningkat. Sebagian besar varietas tahan hama yang digunakan di Indonesia saat ini masih merupakan hasil teknologi pemuliaan tanaman secara konvensional. Seiring dengan perkembangan dan kemajuan teknologi akhir-akhir ini, penerapan bioteknologi modern dalam bidang pertanian dapat menghasilkan varietas tahan hama hasil rekayasa genetika (tanaman transgenik).
Tanaman transgenik dan tanaman bukan transgenik mempunyai mekanisme ketahanan intrinsik dan ketahanan ekstrinsik (Pilcher et al. 2005). Sumber ketahanan intrinsik pada tanaman bukan transgenik pada umumnya berasal dari karakteristik biokimia dan karakteristik biofisik suatu tanaman yang mempengaruhi perilaku atau metabolisme serangga (Kogan 1982). Sumber ketahanan ekstrinsik adalah musuh alami (Pilcher et al. 2005).
Sumber ketahanan intrinsik pada tanaman transgenik, contohnya padi transgenik Bt, adalah protoxin yang berasal dari gen cryIAb, fusi dua gen cry (cryIB-cryIAa), dan gen mpi::cryIB yang mempengaruhi fisiologi serangga. Dengan demikian mekanisme ketahanan tanaman transgenik ini termasuk mekanisme ketahanan antibiosis. Menurut Panda dan Khush (1995) pengaruh antibiosis mengakibatkan berkurangnya ukuran atau berat serangga, mengurangi proses metabolisme, meningkatkan kegelisahan, kematian larva dan pradewasa. Secara tidak langsung antibiosis mengakibatkan meningkatnya eksposur serangga terhadap musuh alaminya.
Tanaman padi transgenik Bt mempunyai pertahanan intrinsik untuk melawan hama penggerek batang padi kuning S. incertulas dan mempunyai pengaruh negatif langsung yaitu toksik terhadap herbivor ini. Seberapa besar toksiknya Bt toxin pada padi transgenik terhadap hama penggerek batang padi kuning S. incertulas dapat diketahui melalui pengujian keefektifan pada tahap in vitro, in planta, dan lapangan terbatas.
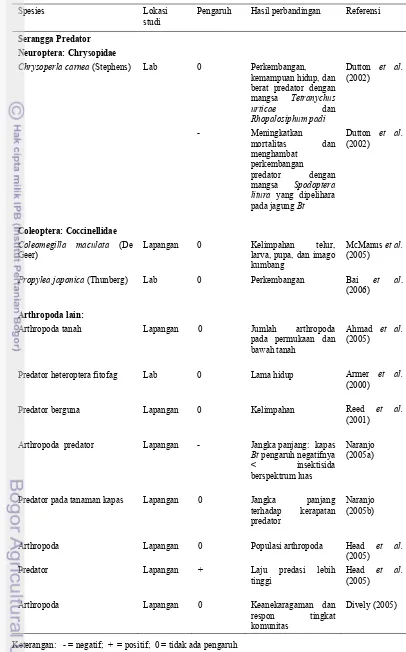
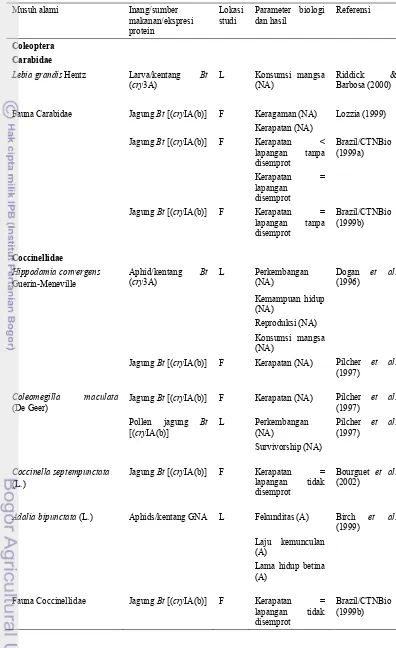
Musuh alami seperti parasitoid dan predator sebagai sumber ketahanan ekstrinsik, adalah regulator populasi serangga hama. Kemampuan hidup musuh alami tergantung pada suplai serangga inang, artinya berkurangnya jumlah inang karena makan pada tanaman transgenik akan mempengaruhi kerapatan populasi musuh alami sebagai pengendali hama (O’Callaghan et al. 2005). Menurut Losey et al. (2004) tanaman transgenik mempengaruhi musuh alami melalui tiga cara yaitu: (1) langsung makan pada jaringan tanaman transgenik seperti pollen, akar; (2) makan pada inang yang makan pada tanaman transgenik; dan (3) melalui pengurangan populasi inang.
Cara pertama menurut Losey et al. (2004) digunakan dalam penelitian ini untuk melihat pengaruh tanaman transgenik terhadap musuh alami yang langsung makan pada jaringan tanaman transgenik. Serangga model yang digunakan adalah kumbang V. lineata (Coleoptera: Coccinellidae). Kumbang ini digunakan karena jumlah spesiesnya yang besar dan distribusinya yang kosmopolitan (Ferry et al. 2007). Selain itu, kumbang dewasa dan larva V. lineata (Coleoptera: Coccinellidae) makan pollen, berbagai serangga kecil, tungau, nektar bunga, dan embun madu (Kalshoven 1981). Pollen tanaman dan nektar dari bunga digunakan sebagai makanan alternatif, sehingga walaupun makanan utamanya jarang, kumbang ini masih dapat bertahan hidup. Di dalam pertanian, kumbang Coleoptera: Coccinellidae ini penting karena sebagai spesies indikator kunci yang digunakan untuk memonitor perubahan ekologi atau lingkungan, termasuk biodiversitas (Ferry et al. 2007).
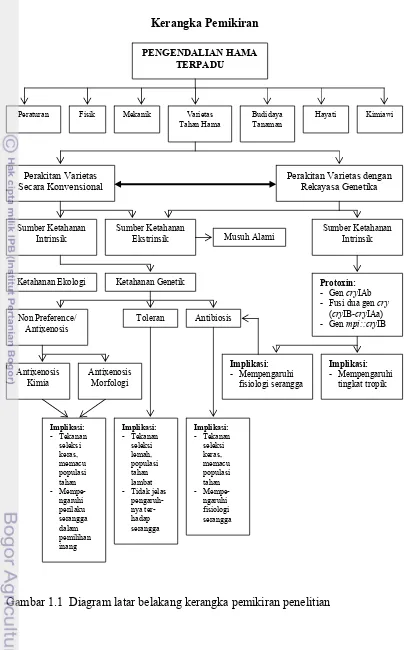
Kerangka Pemikiran
PENGENDALIAN HAMA TERPADU
Fisik Mekanik Varietas
Tahan Hama
Varietas Tahan
Daftar Pustaka
Alam MF et al. 1998. Production of transgenic deepwater rice plants expressing a synthetic Bacillus thuringiensis cryIAb gene with enhanced resistance to yellow stem borer. Plant Science 135:25-30.
Alcantra EP, R Aguda R, DH Dean, MB Cohen. 2000. Investigation of Bacillus thuringiensis δ-endotoxin binding to midgut receptors of rice stem borer. International Rice Genetics Symposium. IRRI. 221 p.
Bandong JP, JA Litsinger. 2005. Rice crop stage susceptibility to the rice yellow stemborer Scirpophaga incertulas (Walker) (Lepidoptera: Pyralidae). Inter Jour Pest Manag 51(1):37-43.
Breitler JC et al. 2000. Expression of a Bacillus thuringiensis cry1B synthetic gene protects Mediterranean rice against the striped stem borer. Plant Cell Reports 19:1195-1202.
Cohen MB. 2000. Bt rice: practical steps to sustainable use. International Rice Research Notes 25(2):4-10.
[DIRJENTAN PANGAN] Direktorat Jenderal Tanaman Pangan.2007. Informasi Perkembangan Serangan OPT Padi Tahun 2006, Tahun 2005 dan Rerata 5 Tahun (2000-2004). Jakarta: Direktorat Jenderal Tanaman Pangan, Direktorat Perlindungan Tanaman.
Ferry N, EA Mulligan, MEN Majerus, AMR Gatehouse. 2007. Bitrophic and tritrophic effects of Bt Cry3A transgenic potato on beneficial, non target, beetles. Transgenic Res 16:795-812.
Ghareyazie et al. 1997. Enhanced resistance to two stem borer in an aromatic rice containing a synthetic cryIA(b) gene. Moleculer Breeding 3:401-414.
Hendarsih S. 1994. Perbandingan efektifitas pengendalian dengan cara kultur teknis, mekanis, dan kimiawi pada penggerek batang padi putih Scirpophaga innotata Wlk. Di dalam Baehaki SE, Sastrosiswojo, Hendarsih S, Tatang S, editor. Prosiding Simposium Penerapan PHT; Sukamandi, 3-4 September 1992. hlm 50-54.
Kalshoven LGE. 1981. The Pests of Crops in Indonesia. Van der Laan PA, penerjemah. Jakarta: PT Ichtiar Baru-van Hoeve. Terjemahan dari: De Plagen van de cultuurgeweassen in Indonesia.
Kogan M. 1982. Plant resistance in pest management. In: Metcalf RL, WH Luckmann, editor. Introduction to Insect Pest Management. Second Edition. New York: John Wiley & Sons. pp 93-134.
Losey JE, JJ Obrycki, RA Hufbauer. 2004. Biosafety considerations for transgenic insecticidal plants: non-target predators and parasitoids. Encyclopedia of Plant and Crop Science. New York: Marcel Dekker, Inc. pp 156-159.
Metcalf RL. 1982. Insecticides in pest management. In: Metcalf RL, WH Luckmann, editor. Introduction to Insect Pest Management. Second Edition. New York: John Wiley & Sons. pp 217-277.
Nayak P et al. 1997. Transgenic elite indica rice plants expressing CryIAc δ -endotoxin of Bacillus thuringiensis are resistant against yellow stem borer (Scirpophaga incertulas). Proc Natl Acad SciUSA 94:2111-2116.
O’Callaghan M, TR Glare, EPJ Burgess, LA Malone. 2005. Effects of plants genetically modified for insect resistance on nontarget organisms. Annu Rev Entomol 50:271-292.
Oka IN. 1992. Program nasional pelatihan dan pengembangan pengendalian hama sebagai salah satu usaha mengembangkan tenaga manusia dalam menuju pertanian tangguh. Makalah Kongres PEI IV; Yogyakarta, 28-30 Januari 1992.
Panda N, GS Khush. 1995. Host Plant Resistance to Insects. CAB International Intl Rice Res Inst. Philippines: Los Banos. 431 p.
Pilcher CD, ME Rice, JJ Obrycki. 2005. Impact of transgenic Bacillus thuringiensis corn and crop phenology on five nontarget arthropods. Environ Entomol 34(5):1302-1316.
Rachmat A. 2006. Konstruksi vektor ekspresi gen untuk mengeliminasi gen penyeleksi antibiotik pada tanaman padi (Oryza sativa L.) transgenik [tesis]. Bogor: Program Pascasarjana, Institut Pertanian Bogor.
Rao PSP, G Padhi. 1988. Improved sources of plant resistance to yellow stem borer (YSB) Scirpophaga incertulas Walker in rice. Intl Rice Res Newsl 13:5.
Rauf A, IW Winasa, R Anwar, A Tarigan, J Lestari. 1992. Kajian beberapa teknik pengendalian penggerek padi putih, Scirpophaga innotata (Wlk)(Lepidoptera: Pyralidae). Seminar Hasil Penelitian Pendukung PHT; Cisarua, 7-8 September 1992. Bogor: Kerjasama Program Nasional Pengendalian Hama Terpadu – Bappenas dengan Fakultas Pertanian – Institut Pertanian Bogor. 17 hlm.
Satoto. 2003. Kestabilan, pola pewarisan, dan keefektifan gen gna dan cry1Ab terhadap wereng batang coklat dan penggerek batang kuning pada padi rojolele transgenik [disertasi]. Bogor: Program Pascasarjana, Institut Pertanian Bogor.
Slamet-Loedin IH et al. 1998. Production of fertile transgenic aromatic Indonesian javanica rice co-expressing the snowdrop lectin and cryIAb anti-insect proteins. Proceedings of the 4th Asia Pacific Conference on Agricultural Biotechnology. pp 206-208.
Sutanto R. 2002. Pertanian Organik. Yogyakarta: Penerbit Kanisius. 218 hal.
Untung K. 2006. Pengantar Pengelolaan Hama Terpadu. Edisi Kedua. Yogyakarta: Gadjah Mada University Press. 348 hal.
TINJAUAN PUSTAKA
Ketahanan Tanaman
Ketahanan tanaman adalah sifat-sifat tanaman yang dapat diturunkan dan dapat mempengaruhi tingkat kerusakan oleh serangga. Dalam praktek pertanian, ketahanan tanaman berarti kemampuan tanaman untuk berproduksi lebih baik dibandingkan tanaman lain dengan tingkat populasi hama yang sama dan lingkungan yang sama (Panda & Khush 1995).
Berdasarkan susunan dan sifat gen, ketahanan genetik dapat dibedakan menjadi: (1) monogenik, sifat tahan diatur oleh satu gen dominan atau resesif, (2) oligogenik, sifat tahan diatur oleh beberapa gen yang saling menguatkan satu sama lain, dan (3) polygenik, sifat tahan diatur oleh banyak gen yang saling menambah dan masing-masing gen memberikan reaksi yang berbeda-beda terhadap biotipe hama sehingga mengakibatkan timbulnya ketahanan yang luas.
Ketahanan genetik juga dapat dibedakan menjadi beberapa tipe: (1) ketahanan vertikal, ketahanan hanya terhadap satu biotipe hama, dan biasanya bersifat sangat tahan tetapi mudah patah oleh munculnya biotipe baru, (2) ketahanan horizontal atau ketahanan umum, ketahanan terhadap banyak biotipe hama dengan derajat ketahanan ”agak tahan”, dan (3) ketahanan ganda, memiliki sifat tahan terhadap beberapa jenis hama .
Ketahanan tanaman terhadap serangga dilandasi pemahaman interaksi tanaman dengan serangga. Dengan berkembangnya ilmu pengetahuan dan teknologi, hubungan interaksi antara serangga dengan tanaman dapat diketahui lebih luas dan lebih baik. Tanaman secara alami mempunyai sistem pertahanan untuk mempertahankan dirinya dari serangan serangga tertentu. Sistem pertahanan tanaman tersebut diantaranya: (a) avoidance, penghindaran dalam ruang dan waktu, (b) pertahanan fisik, misalnya adanya rambut-rambut pada permukaan daun yang disebut trichome dan glandular trichome yang sangat potensial untuk memerangkap serangga, duri, daun yang licin atau mengkilat, dan adanya lapisan lilin berfungsi agar serangga sulit untuk mengkolonisasi, dan (c) pertahanan kimia, tanaman mengandung metabolit sekunder (senyawa sekunder) seperti phenol, steroid, dan terpenoid, pada kadar tertentu tahan terhadap serangan serangga tertentu. Senyawa sekunder ini dapat bersifat racun baik secara langsung atau setelah dihidrolisis di dalam sistem pencernaan serangga (Speight et al. 1999).
akibat adanya serangan hama, merupakan salah satu contoh bahwa serangga membangun strategi untuk mematahkan sistem pertahanan tanaman. Tanaman dan serangga kejar mengejar dalam mengembangkan strategi untuk mempertahankan dirinya dan mematahkan sistem pertahanan musuh (Tamayo et al. 2000).
Strategi Pengendalian Serangga dengan Rekayasa Genetika
Teknologi rekayasa genetika adalah salah satu teknologi alternatif yang dapat dimanfaatkan, diantaranya untuk meningkatkan ketahanan tanaman terhadap serangga tertentu dengan meningkatkan ekspresi gen ketahanan yang sudah ada atau dengan memasukkan gen baru yang secara alami tidak ditemukan pada tanaman tersebut maupun kerabat liarnya. Kemajuan yang pesat dibidang biologi molekuler telah memacu penemuan gen-gen ketahanan baru dari berbagai sumber (tanaman tingkat tinggi, hewan, dan mikroba) dan dimanfaatkan secara luas, sehingga tanaman target mempunyai kemampuan internal untuk melawan serangga tertentu. Gen baru tersebut dapat diekspresikan secara terus menerus (constitutive) atau setelah adanya sinyal serangan hama (inducible) pada bagian tertentu dari tanaman (akar, daun, batang, bunga, biji, dan lain sebagainya), pada fase pertumbuhan tertentu (vegetatif, generatif), dan pada konsentrasi tertentu dibawah kendali ”promoter” yang dikenali oleh mesin transkripsi tanaman inang. Sebelum dimasukkan ke dalam tanaman inang, biasanya konstruksi gen tersebut terlebih dahulu dirakit di dalam sebuah ”vektor” (plasmid) yaitu dengan menempelkan gen bermanfaat tersebut ke promoter. Vektor tersebut juga mengandung gen penanda dan penyeleksi. Gen penanda berfungsi untuk memberitahukan keberhasilan transformasi, sedangkan gen penyeleksi berfungsi untuk memudahkan seleksi sel yang tertransformasi (Rahmawati 2004).
tersebut, kedalam genom tanaman dikotil dan menyebabkan tumor (Old & Primrose 1994). Namun, A. tumefaciens yang digunakan dalam rekayasa genetika adalah bakteri yang sudah dihilangkan gen penyebab terbentuknya tumor (disarmed).
Penguasaan teknologi rekayasa genetika yang lebih baik dan diketahuinya sifat biologi serangga, sistem penyerangan dan mekanisme pertahanan dirinya, memungkinkan pengembangan strategi pengendalian serangga hama. Secara umum strategi pengendalian serangga hama dapat dibagi atas tiga cara yaitu: pertama, dengan melaparkan serangga akan molekul penting seperti asam amino dan gula sederhana sehingga pertumbuhannya terhambat dan menyebabkan kematian. Telah diketahui bahwa banyak enzim yang berperan dalam proses pencernaan serangga seperti proteinase dan amylase, masing-masing adalah enzim yang berperan dalam pemecahan protein dan karbohidrat menjadi asam amino dan gula sederhana. Penghambatan kerja enzim ini, misalnya dengan adanya kandungan proteinase inhibitor (PI) pada tanaman menyebabkan serangga kekurangan asam amino yang dapat diserap oleh tubuhnya (Irie et al. 1996).
Kedua, dengan meracuni serangga. Salah satu contoh yang paling populer adalah dengan mengintroduksi gen cry dari B. thuringiensis yang menyandikan kristal protein Bt yang bersifat racun setelah dimakan dan dihidrolisis di dalam sistem pencernaan serangga sasaran (Metz et al. 1995; Wu et al. 1997; Maqbool et al. 1998).
Ketiga, dengan menghambat pembentukan molekul penyusun kerangka tubuh serangga. Misalnya, dengan mengekspresikan chitinase, yaitu pengurai chitin (molekul utama penyusun kerangka tubuh serangga) pada tanaman (Hilder et al. 1995).
Gen cry, Penyandi δ-Endotoksin dari Bacillus thuringiensis
Bacillus thuringiensis adalah bakteri yang menghasilkan suatu kristal protein yang bersifat racun jika terhidrolisis dalam sistem pencernaan serangga (Dekeyser et al. 1990). Kristal protein Bt yang bersifat insektisida sering disebut dengan δ-endotoxin. Kristal ini di alam merupakan protoxin yang jika larut dalam sistem pencernaan serangga karena proses proteolisis akan diubah menjadi polipeptida yang lebih pendek (27-149 kilo Dalton) serta mempunyai sifat insektisida. Toxin aktif ini berinteraksi dengan sel-sel epitel midgut serangga. Toxin Bt menyebabkan lubang-lubang kecil pada membran sel epidermis sistem pencernaan serangga, sehingga mengganggu keseimbangan osmotik sel tersebut. Sel yang terganggu tekanan osmosisnya menjadi bengkak dan pecah, sehingga serangga mati (Hofte & Whiteley 1989; Bahagiawati 2005; Manyangarirwa et al. 2006).
Pengetahuan tentang mekanisme kerja endotoxin Bt penting untuk menentukan proses utama yang bertanggung jawab terhadap kespesifikan dari suatu kristal protein. Faktor utama yang menentukan kisaran inang kristal protein adalah perbedaan pH di midgut larva yang mempengaruhi proses kelarutan (solubilization) dan pengubahan kristal yang tidak aktif menjadi aktif, serta keberadaan lokasi penempelan (binding site) yang spesifik dari protoxin di dalam sistem pencernaan serangga (Lereclus et al. 1993; Bahagiawati 2005; Manyangarirwa et al. 2006).
(5) cryV yang dapat menyerang lepidoptera dan coleoptera (Bahagiawati 2005). Berdasarkan urutan basanya gen cryI terbagi atas enam klas yaitu cryIAa, cryIAb, cryIAc, cryIB, cryIC, dan cryID.
Proteinase Inhibitor
Proteinase inhibitor biasanya ada pada benih dan umbi tanaman dimana mereka disintesis dan disimpan. Ekspresi gen proteinase inhibitor tanaman terinduksi pada jaringan vegetatif dalam merespon pelukaan, suatu respon tanaman terkait pada peningkatan ketahanan terhadap serangga. Proteinase inhibitor mempunyai peranan dalam pertahanan tanaman melawan serangga herbivor melalui penghambatan enzim proteolytic serangga, keefektifannya tergantung pada afinitas dan atau spesifikasi inhibitor terutama proteinase midgut dari serangga target (Tamayo et al. 2000).
Menurut Laskowski et al. (1988) dalam Tamayo et al. (2000), kompatibilitas antara proteinase inhibitor tanaman dan proteinase serangga menentukan tingkat aktivitas penghambatan terhadap proteinase spesifik. Adaptasi hama terhadap proteinase inhibitor tanaman inang mungkin hasil dari tekanan seleksi yang bekerja pada suatu populasi serangga ketika mereka menghadapi proteinase inhibitor tanaman inangnya. Agar dapat bertahan hidup, tanaman harus mengembangkan protein penghambatnya untuk secara efektif menghambat proteinase serangga. Oleh karena itu hama dan tanaman mengembangkan bentuk baru dari enzim dan inhibitor untuk menetralkan setiap mekanisme pertahanan lain. Ekspresi proteinase inhibitor tanaman pada tanaman transgenik bertujuan untuk mengurangi laju pertumbuhan larva dari beberapa spesies serangga. Tetapi, pengaruh perlindungannya bersifat sementara (Tamayo et al. 2000).
Maize Proteinase Inhibitor (mpi) dan Regulasinya
Fenomena ini diperjelas dengan hasil analisis gel RNA dari jaringan luka yang menunjukkan pada perlakuan pelukaan yang berurutan menghasilkan transkripsi mpi dengan tingkat tertinggi. Promoter gen ini mengatur ekspresi gen yang tinggi pada saat terjadi pelukaan.
Penggerek Batang Padi
Di Indonesia telah ditemukan 6 spesies penggerek batang padi dari ordo lepidoptera, yang terdiri dari 5 spesies famili pyralidae dan 1 spesies famili noctuidae. Ke-enam spesies penggerek batang padi tersebut adalah penggerek batang padi kuning S. incertulas (PBPK), penggerek batang padi bergaris C. suppressalis, penggerek batang padi merah jambu Sesamia inferens (Walker), penggerek batang padi putih Scirpophaga innotata (Walker) (PBPP), penggerek batang padi kepala hitam Chilo polychrysus Meyrick, dan penggerek batang padi berkilat Chilo auricilius Dudgeon (Hattori & Siwi 1986). Jenis-jenis penggerek batang padi tersebut memiliki sifat atau ciri yang berbeda dalam penyebaran dan bioekologi, namun hampir sama dalam cara menyerang atau menggerek tanaman serta kerusakan yang ditimbulkannya.
Penggerek batang padi dapat menyerang pada semua stadium pertumbuhan tanaman padi (Alam et al. 1998). Pada stadium vegetatif serangan menyebabkan kematian anakan (tiller) muda yang disebut sundep (deadhearts), dan pada stadium generatif serangan menyebabkan malai muncul putih dan hampa yang disebut beluk (whiteheads) (Bandong & Litsinger 2005). Kehilangan hasil akibat serangan penggerek batang padi pada stadium vegetatif tidak terlalu besar karena tanaman masih dapat mengkompensasi dengan membentuk anakan baru. Berdasarkan simulasi pada stadium vegetatif, tanaman masih sanggup mengkompensasi akibat kerusakan oleh penggerek sampai 30% (Rubia & De Vries 1990). Kehilangan hasil yang disebabkan setiap persen gejala beluk berkisar 1-3% (Pathak & Khan 1994).
Selatan, jenis penggerek batang padi yang dominan adalah PBPP diikuti oleh penggerek batang padi bergaris, penggerek batang padi merah jambu, penggerek batang padi kepala hitam, dan penggerek batang padi berkilat (Baco & Said 1998). Penggerek batang padi kepala hitam dan penggerek batang padi bergaris tersebar luas di Sulawesi, Jawa, Bali, Sumatera, dan dalam populasi yang rendah di Kalimantan.
Di Pantai Utara Jawa Barat terjadi perubahan dominasi spesies yang sangat dinamis. Setelah lama PBPP digeser oleh PBPK, pada tahun 1989-1990 kembali muncul ledakan populasi PBPP (Manwan et al. 1990). Namun, sejak tahun 1995 secara bertahap PBPP tersebut digeser kembali oleh PBPK, sehingga pada akhir dekade 90-an kecuali di Indramayu, PBPK sangat dominan (Hendarsih et al. 2002).
Perubahan dominasi penggerek batang padi dari PBPP ke PBPK dipengaruhi oleh adanya pembangunan jaringan irigasi dari Jatiluhur dan dam yang lainnya yang menyebabkan 300 000 ha sawah berpengairan teknis. Kondisi ini memungkinkan perubahan pola tanam dari satu kali menjadi dua kali dalam setahun (Sosromarsono 1990). Selain itu varietas berumur pendek dan responsif terhadap pemupukan menggantikan varietas lokal, serta intensifikasi tanaman dengan masukan pemupukan dan aplikasi insektisida diduga penyebab perubahan dominasi spesies penggerek batang padi. Perubahan kultur teknis besar pengaruhnya terhadap dominasi dan serangan penggerek batang padi. Di Jepang, C. suppressalis berkurang setelah ada perubahan varietas, cara tanam, dan cara panen (Kiritani 1988).
Biologi Penggerek Batang Padi Kuning S. incertulas
Mereka membuat lubang pada pelepah daun, atau menyebar dengan menggantung pada benang sutra. Larva lainnya mungkin menyebar dengan mengapung diatas air pada helaian daun yang dipotong dari tanaman. Perkembangan larva berkisar 3-6 minggu dengan panjang tubuh 25 mm. Larva berwarna putih kekuningan hingga hijau. Periode pupa adalah 8-14 hari. Total siklus hidup penggerek batang padi kuning mencapai 5-9 minggu. Ketika hendak menjadi pupa, larva turun ke ruas batang yang paling bawah, biasanya di bawah air atau sejajar dengan tanah. Pupa berwarna kekuningan dan dibentuk di dalam kokon yang berwarna putih. Ngengat dapat muncul menembus air sedalam 12.5 cm, tidak makan dan hidup hanya 5-7 hari. Serangga dewasa, khususnya betina tertarik cahaya. Serangga betina berwarna kuning krem dengan bintik hitam yang jelas pada sayap depan, tumpukan rambut yang luas, dan sisik pada ujung abdomen. Panjang tubuh pada posisi istirahat adalah 17 mm. Jantan berwarna abu-abu kecoklatan dengan panjang tubuh hanya 14 mm dan mempunyai bintik pada sayap depan yang tidak jelas. Jumlah perkembangan generasi di lahan sawah pada musim hujan berkisar 5-8 generasi tergantung pada lamanya waktu tanam dan waktu panen. Di Indonesia, penggerek batang padi kuning S. incertulas tidak mempunyai periode istirahat pada musim kemarau. Oleh karena itu kemampuan hidup penggerek batang padi kuning S. incertulas tergantung pada tidak adanya tanaman inang alternatif pada pertanaman padi (Kalshoven 1981).
Predator V. lineata
Interaksi Tritropik
Interaksi tritropik merupakan interaksi paling sederhana, yaitu interaksi antara tanaman-herbivor-musuh alami, dalam jaringan makanan. Dalam interaksi tritropik ini, tanaman merupakan tingkat tropik pertama, herbivor merupakan tingkat tropik kedua, dan musuh alami merupakan tingkat tropik ketiga (Vet & Dicke 1992). Pada tingkat tropik tersebut terdapat pertarungan kekuatan antara tingkat yang dibawah dengan tingkat yang diatasnya. Tingkat tropik yang lebih bawah berevolusi untuk mengurangi serangan, sedangkan tingkat tropik yang lebih tinggi berevolusi untuk meningkatkan serangan (Price et al. 1980).
Tanaman menyediakan makanan (nektar, pollen) dan tempat berlindung untuk musuh alami herbivor, dengan cara demikian tanaman mendapatkan pelindung yang permanen. Tanaman melepas senyawa volatil yang diinduksi oleh pelukaan dan herbivor yang dapat menarik predator atau parasitoid, ini merupakan sifat pertahanan tidak langsung dari tanaman. Pelepasan senyawa volatil organik dapat juga berfungsi sebagai pertahanan langsung dengan menolak oviposisi herbivor dan mungkin sebagai perantara interaksi antara tanaman dengan tanaman (Kessler & Baldwin 2002).
Kerentanan serangga herbivor untuk diserang oleh predator dan parasitoid sering dimediasi oleh interaksi dengan tanaman inang dimana herbivor makan. Herbivor spesialis mengatasi pertahanan tanaman dengan mengambil toxin kimia yang dihasilkan oleh tanaman di dalam tubuhnya sebagai pertahanan melawan musuh alaminya (Moraes & Mescher 2004). Misalnya larva Manduca sexta yang makan pada tanaman Nicotiana tabacum menahan nicotin pada hemolim-nya yang mempengaruhi penampilan dan kemampuan hidup endoparasit Cotesia congregata (Kessler & Baldwin 2002).
sementara kimia yang berasal dari herbivor meningkat dengan berkurangnya jarak dari korban (Vet & Dicke 1992).
Informasi kimia dapat berasal dari herbivor, dari makanannya, dari organisma yang berasosiasi dengan herbivor, atau dari interaksi antar sumber ini. Produk dari herbivor yang potensial memberikan isyarat kimia untuk penyerangnya adalah feses, kutikula, eksuvia, sekresi dari mandibula dan accessory gland, feromon, embun madu, sisik tubuh atau hemolim. Produk dari makanan herbivor seperti bunga atau daun dapat memberikan informasi kimia spesifik. Organisma yang berasosiasi dengan herbivor seperti mikroba mungkin juga sebagai sumber penting dari informasi kimia (Vet & Dicke 1992).
Menurut Vet dan Dicke (1992) stimulus yang berasal dari herbivor pada umumnya lebih dapat dipercaya sebagai sumber informasi. Tetapi, penggunaan stimulus yang berasal dari herbivor sering dibatasi oleh rendahnya kemampuan deteksi, khususnya pada jarak jauh, sebaliknya stimulus yang berasal dari tanaman biasanya lebih tersedia, karena biomas tanaman relatif lebih besar, tetapi kurang dapat dipercaya sebagai prediktor keberadaan herbivor. Dapat dipercayanya sinyal dari tanaman tergantung pada kemampuan untuk memprediksi dari tanaman yang terserang pada ruang dan waktu. Jadi musuh alami dituntut untuk menggabungkan karakteristik yang menguntungkan dari informasi yang berasal dari kedua tropik tersebut.
A. Tipe semiokimia yang digunakan pada interaksi tritropik
Semiokimia adalah kimia yang diperlukan di dalam interaksi kimiawi antar organisma (Vet & Dicke 1992; Price 1997). Semiokimia pada dasarnya ada dua tipe yaitu feromon dan allelokimia. Feromon adalah semiokimia intraspesifik yang menjadi perantara interaksi serangga ke serangga. Hal ini lebih jauh diklasifikasikan kedalam tipe yang berbeda tergantung pada fungsinya seperti sex pheromone, alarm pheromone, trail marking pheromone, territory marking
allomon. Synomon adalah allelokimia yang menghasilkan respon yang menguntungkan bagi keduanya yaitu untuk penerima dan yang mengeluarkan emisi, sementara kairomon menghasilkan respon yang menguntungkan hanya untuk penerima dan apneumon adalah allelokimia yang dilepas oleh substansi tidak hidup, yang menghasilkan respon yang menguntungkan dan digunakan untuk melawan organisma lain pada habitat yang sama (Price 1997; Ahmad et al. 2004).
Allelokimia yang menghasilkan interaksi herbivor-tanaman dan yang menarik musuh alami herbivor mungkin dipandang dalam konteks interaksi tingkat pertama-ketiga atau dalam interaksi tingkat kedua-ketiga. Pada interaksi tingkat pertama-ketiga (antara tumbuhan dan musuh alami) allelokimia berfungsi sebagai synomon dan pada interaksi tingkat kedua-ketiga (antara herbivor-musuh alami) allelokimia berfungsi sebagai kairomon (Vet & Dicke 1992).
B. Semiokimia yang dihasilkan tanaman dan interaksi tritropik
tubuh herbivor bertindak sebagai kairomon antara serangga dan musuh alami, khususnya parasitoid. Ulat jagung, Heliothis zea, mengambil trichosane dari jagung yang dilewatkan tanpa berubah ke dalam telurnya. Parasitoidnya yaitu Trichogramma evanescens menggunakan kimia ini sebagai kairomon dan untuk menemukan telur inangnya. Biokimia yang dihasilkan oleh tanaman juga mempengaruhi interaksi tritropik secara tidak langsung dengan merubah ukuran, vigour, laju pertumbuhan, dan laju kemampuan hidup dari herbivor. Ukuran serangga dikurangi oleh metabolit sekunder tanaman yang menyebabkan meningkatkan atau mengurangi keefektifan predator, karena predator menyukai ukuran yang spesifik dari mangsa untuk dikonsumsi (Ahmad et al. 2004).
Ada kasus dimana semiokimia yang dihasilkan oleh tanaman bertindak melawan organisma tingkat tropik ketiga. Kadang-kadang asosiasi volatil tanaman mungkin menutupi atraktan untuk musuh alami. Contoh, bau tubuh dari Larch dan Larch sawfly, Pristiphora erichsonii, adalah penarik parasitoid hingga mencapai 86% parasitisasi pada Larch murni, tetapi jika pohon lain ditumbuhkan pada asosiasi ini, parasitisasi berkurang hingga 12% karena penutupan pengaruh volatil yang dihasilkan oleh pohon yang berasosiasi. Pada kasus lain pertahanan alomon mungkin menjadi membahayakan untuk tanaman, ketika mereka juga menolak musuh alami, tetapi beberapa herbivor menjadi lebih spesialis terhadap kimia yang sama (Ahmad et al. 2004).
kandungan nitrogen tinggi dibandingkan dengan tanaman yang rendah nitrogen. Tanaman yang rendah kualitas nutrisinya berpengaruh buruk terhadap pertumbuhan larva dan perkembangan serangga herbivor, menyajikan suatu paradoks bagi pengamat karena serangga perlu makan yang banyak untuk mencapai ganti kulit menuju dewasa, hal ini penyebab lebih banyak kerusakan pada tanaman inang dibandingkan dengan mekanisme ketahanan lain seperti antibiosis. Tetapi, perkembangan larva yang panjang yang berasosiasi dengan kualitas tanaman inang yang rendah mungkin meningkatkan kesempatan eksposur pada predator dan patogen. Ketika kumbang chrysomelid Galerucella lineola dipelihara pada dua spesies tanaman inang, satu cocok untuk perkembangan larva (Salix viminalis) dan satu tidak cocok (Salix dasyclados), pertumbuhan dan kemampuan hidup larva lebih tinggi pada Salix viminalis tetapi laju pemangsaan setiap harinya lebih tinggi pada S. dasyclados.
C. Semiokimia yang dihasilkan herbivor dan interaksi tritropik
Herbivor secara nyata terkait dalam interaksi tritropik, dimana disatu sisi berinteraksi dengan tanaman dan satu sisi lain dengan musuh alaminya, maka, dalam dua cara mereka harus mengatasi pertahanan intrinsik tanaman dan serangan musuh alami. Jadi herbivor menghasilkan semiokimia lain untuk mengatasi pertahanan tanaman atau untuk menolak musuh alami. Semiokimia juga memainkan peranan penting dalam interaksi tritropik. Contoh dari synomon yang menjadi perantara interaksi herbivor-predator adalah larva kupu-kupu Lycaenid dan aphid yang menghasilkan synomon (embun madu yang kaya gula) yang menarik semut untuk perlindungan melawan musuh alami. Suatu asosiasi mutualisme berkembang antara semut dan aphid, semut melindungi aphid dan larva kupu-kupu dari musuh alami lain. Sebaliknya aphid menyediakan makanan untuk semut dalam bentuk embun madu.