ANALISA PROFIL PROTEIN ISOLAT Escherichia coli S1
HASIL IRADIASI SINAR GAMMA
IKMALIA
103096029804
PROGRAM STUDI KIMIA
FAKULTAS SAINS DAN TEKNOLOGI
UNIVERSITAS ISLAM NEGERI SYARIF HIDAYATULLAH
JAKARTA
Motto:
“Jika engkau kesulitan dalam pekerjaanmu, jangan putus asa, jangan gelisah dan jangan ragu. Percayalah, jalan keluar akan segera datang”.
“Jalan kebahagiaan ada di depanmu. Carilah ia dalam lautan ilmu, amal shaleh dan akhlak yang mulia. Bersikaplah obyektif dalam setiap hal, niscaya engkau
akan bahagia”.
“
Ya Allah, Engkau adalah Tuhanku. Tidak ada Tuhan selain Engkau.
Engkau telah menciptakan aku dan aku adalah hamba-Mu. Aku berada
dalam janji dan akan menepati perjanjian itu semampuku. Aku berlindung
dari buruk yang aku perbuat. Aku mengakui nikmat-Mu padaku. Aku
mengakui dosa dosaku. Ampunilah aku, sesungguhnya tidak ada yang
mengampuni dosa kecuali Engkau”.
ABSTRAK
IKMALIA. Analisa Profil Protein Isolat Escherichia coli S1 Hasil Iradiasi Sinar Gamma. Dibawah bimbingan IRAWAN SUGORO dan SANDRA HERMANTO.
Penelitian ini bertujuan untuk mengetahui profil protein Escherichia coli
S1 hasil iradiasi sinar gamma sebagai bahan vaksin mastitis inaktif. Kultur sel (1013 sel/ml) diiradiasi dengan dosis 0, 50, 200, 500, 600, 700, 800, 900, dan 1000 Gy dengan laju dosis 1089,59 Gy/Jam. Kultur sel dianalisis kandungan protein intraselular dan ekstraselular dengan metoda Lowry. Profil protein dianalisis menggunakan SDS-PAGE dengan konsentrasi gel 10% dan protein standar (marker) dengan berat molekul 10 – 220 kDa. Hasil penelitian menunjukkan bahwa bakteri E. coli mampu diinaktivasi pada dosis diatas 800 Gy. Pada dosis inaktivasi menunjukan tidak ada pengaruh yang signifikan (Sign > 5%) terhadap profil protein yang dihasilkan, bila dibandingakan dengan kontrol. Hal ini menunjukan bahwa protein hasil iradiasi pada dosis inaktif dapat digunakan sebagai bahan vaksin mastitis inaktif.
ABSTRACT
IKMALIA. Analyze the protein profile of Escherichia coli S1 which irradiated by gamma rays. Advisor IRAWAN SUGORO and SANDRA HERMANTO.
The experiment has been conducted to study the protein profile of
Escherichia coli S1 which irradiated by gamma rays as inactive mastitis vaccine. Cell culture (1013 cells/ml) was irradiated with dose 0, 50, 200, 500, 600, 700, 800, 900, and 1000 Gy with dose rate of 1089,59 Gy/Jam. The cell culture was analyzed for total intracellular and extra cellular protein by Lowry method. The protein profile was analyzed by SDS-PAGE with 10% gel concentration and standard protein (marker) of molecular weight was 10 – 220 kDa. The result showed that E. coli cells could be inactivated by doses higher than 800 Gy. The profile protein of inactivated doses weren’t affected significantly than control. Based on the result, the protein of inactivated doses can be used as inactive mastitis vaccine.
KATA PENGANTAR
Assalamu’alaikum wr. wb.
Segala puji serta syukur penulis panjatkan kepada Allah SWT yang telah melimpahkan rahmat serta hidayah-Nya, sehingga penulis dapat menyelesaikan skripsi ini dengan judul “Analisa Profil Protein Isolat Escherichia coli S1 Hasil Iradiasi Sinar Gamma”.
Shalawat serta salam semoga senantiasa dilimpahkan kepada Nabi Muhammad SAW beserta segenap keluarga, para sahabat dan para pengikutnya hingga yaumul kiyamah.
Terselesaikannya skripsi ini tidak lepas dari peranan berbagai pihak yang telah ikut secara langsung maupun tidak langsung. Penulis sadar sepenuhnya, bahwa bagaimanapun usaha yang ditempuh tanpa adanya bimbingan dan bantuan dari pihak lain, penulisan skripsi ini tidak akan terselesaikan dengan baik. Maka dalam kesempatan ini, penulis ingin mengucapkan terima kasih yang sebesar-besarnya kepada:
1. Bapak DR. Syopiansyah Jaya Saputra, M.Sis, selaku dekan Fakultas Sains dan Teknologi.
2. Ibu Sri Yadial Chalid, M.Si, selaku ketua Program Studi Kimia dan dosen Penguji I.
4. Bapak S. Hermanto, M.Si, selaku dosen Pembimbing II yang telah banyak membantu dan memberikan kritik dan saran yang membangun dalam analisa hasil dan penulisan.
5. Bapak DR. Mirzan T. Razzak,M.Eng.,APU, selaku dosen Penguji II. 6. Ibunda dan Ayahanda tercinta yang kasih sayangnya sepanjang masa dan
telah memberikan dukungan moril, materil serta spiritual, serta kakanda dan adinda tersayang.
7. Ahmad Hudori yang selalu mendampingi, meluangkan waktu, dan memberi semangat serta dukungan baik moril maupun spiritual.
8. Ibu Nuniek dan bapak Dinar yang ikut membantu pada penelitian dan teman-teman seperjuangan khususnya Ericca, Rizkina, Ai, Eva dan Septi yang memberikan motivasi dan kenangan.
Tak ada gading yang tak retak dan tidak ada sesuatu apapun yang sempurna di dunia kecuali Allah SWT, sehingga penulis sangat menyadari akan masih banyak kekurangan dan kelemahan dalam penulisan skripsi ini. Saran dan kritik yang membangun demi perbaikan sangat diharapkan.
Akhirnya, penulis hanya dapat berdo’a semoga semua amal baik yang telah diberikan tersebut mendapat balasan yang berlipat ganda dari Allah SWT. Dan semoga skripsi ini dapat memberikan sumbangan pemikiran demi kemajuan dan keberhasilan bersama. Amin Ya Mujibas Sailin.
Wassalamu’alaikum wr. wb.
DAFTAR ISI
BAB II TINJAUAN PUSTAKA... 2.1Bakteri Escherichia coli... 5
2.2Protein ... 11
2.3 Antigen... 16
2.4 Faktor Virulensi Mikroba... 18
2.5 Vaksin ... 21
2.6 Radiasi dan Iradiasi ... 29
2.7 Sinar Gamma... 36
BAB III METODE PENELITIAN ...
3.1Tempat dan Waktu Pelaksanaan... 44
3.2Bahan dan Alat... 44
3.3Metoda Kerja... 45
3.4Prosedur Kerja... 46
3.4.1 Pembuatan Kurva Tumbuh Escherichia coli... 46
3.4.2 Penentuan Dosis Iradiasi Inaktivasi sel E. coli dengan Sinar Gamma.... 46
3.4.3...Peng ukuran Protein Sel E. coli Metode Lowry... 47
3.4.4...Kara kteristik Profil Protein Bakteri E. coli... 47
BAB IV HASIL DAN PEMBAHASAN ... 4.1Pertumbuhan bakteri E. coli ... 50
4.2Hasil Inaktivasi Sel E. coli dengan Iradiasi Sinar Gamma ... 51
4.3Konsentrasi Protein sel E. coli Hasil Iradiasi... 53
4.4Karakteristik Profil Protein Bakteri E. coli... 55
BAB V KESIMPULAN DAN SARAN ... 5.1Kesimpulan ... 59
5.2Saran... 59
DAFTAR PUSTAKA ... 60
DAFTAR GAMBAR
Halaman
Gambar 1. Morfologi bakteri Escherichia coli... 5
Gambar 2. Bagian yang terdapat pada bakteri gram negatif... 6
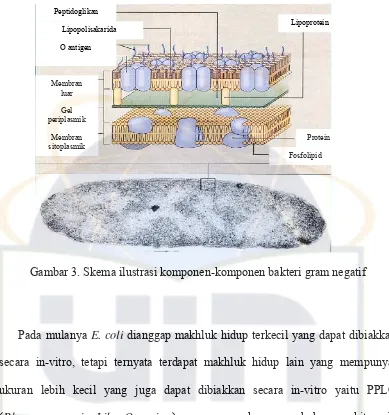
Gambar 3. Skema ilustrasi komponen-komponen bakteri gram negatif... 7
Gambar 4. Faktor-faktor yang mempengaruhi terjadinya mastitis ... 9
Gambar 5. Proses infeksi mastitis pada ambing sapi ... 11
Gambar 6. Struktur primer protein... 13
Gambar 7. Struktur sekunder protein ... 14
Gambar 8. Beberapa jenis ikatan yang terdapat pada polipeptida ... 15
Gambar 9. Struktur kuartener protein globular yang kompleks... 15
Gambar 10. Sketsa proses denaturasi protein (Winarno, 2002)... 16
Gambar 11. Mekanisme kerja vaksin (Scorvia, 2000)... 22
Gambar 12. Efek radiasi terhadap sel yang dapat dimanfaatkan untuk pembuatan bahan vaksin iradiasi ... 26
Gambar 13. Peristiwa terjadinya eksitasi dan ionisasi ... 31
Gambar 14. Formula asam amino ... 34
Gambar 15. Efek fotolistrik... 37
Gambar 16. Efek penghamburan compton... 37
Gambar 17. Pertumbuhan Bakteri E. coli dalam medium TSB yang diinkubasikan pada suhu 370C dan agitasi 120 rpm ... 50
Gambar 18. Hubungan dosis iradiasi sinar gamma terhadap jumlah sel bakteri E. coli... 52
Gambar 19. Konsentrasi protein E. coli hasil iradiasi... 54
DAFTAR TABEL
Halaman Tabel 1. Beberapa enzim ektraselular yang turut menentukan
DAFTAR LAMPIRAN
Halaman
Lampiran 1. Komposisi buffer sampel dan Komposisi larutan Lowry... 65
Lampiran 2. Skema kerja ... 66
Lampiran 3. Kurva standar protein ... 67
Lampiran 4. Kurva standar pertumbuhan bakteri E. coli S1... 68
Lampiran 5. Hasil analisis LabImage 1D 2006... 69
Lampiran 6. Hasil analisis statistik SPSS 11.5 ... 74
Lampiran 7. Gambar bakteri E. coli S1 dan Inkubator shaker... 75
Lampiran 8. Gambar LabImage 1D 2006 ... 76
Lampiran 9. Gamma Chamber 4000 A dan Bahan-bahan SDS-PAGE ... 77
BAB I
PENDAHULUAN
1.1LATAR BELAKANG
Mastitis merupakan penyakit yang mengakibatkan peradangan pada kelenjar susu (ambing), akibat infeksi oleh mikroba. Apabila sapi perah mengalami penyakit mastitis, maka produksi susu yang dihasilkan akan lebih sedikit karena terdapat penyumbatan pada ambing susu sapi, atau bahkan dapat menyebabkan kematian dan kerusakan bagian organ tubuh pada sapi. (Tetriana dan Sugoro, 2007).
Mastitis juga merupakan hal yang komplek dan tidak ditemukan suatu solusi yang sederhana untuk mengontrolnya. Beberapa aspek telah banyak dikaji dan di dokumentasikan dalam literatur ilmiah. Ada yang kontrovesial dan ada opini yang didasarkan pada fakta. Ada 3 faktor penyebab yang mempermudah terjadinya mastitis, yaitu kondisi sapi sebagai inang, kondisi lingkungan yang buruk dan mikroorganisme sebagai agen penyebab penyakit. Salah satu mikroorganisme penyebab mastitis adalah Escherischia coli, akibatnya produksi susu sapi menurun (Sugoro, 2004).
karena kurangnya informasi dan harga yang relatif mahal dan penggunaan vaksin komersil tidak cocok dengan strain lokal. Salah satu produk vaksin yang beredar di pasaran adalah J5 Bacterin dan Mastiguard untuk bakteri Coliform dan
Endovac bovi untuk bakteri Gram negatif. Vaksin-vaksin tersebut ini telah banyak digunakan oleh para peternak di Amerika Serikat, Selandia Baru dan Australia serta dapat menurunkan kejadian mastitis sampai dengan 60%(Ruegg, 2001).
Penelitian ini didasarkan pada pembuatan vaksin dengan cara menginaktivasi sel bakteri. Vaksin dapat diperoleh dengan cara konvensional, baik secara kimia maupun pemanasan. Alternatif lainnya adalah dengan menggunakan iradiasi sinar gamma untuk menginaktivasi sel bakteri. Keuntungan metode iradiasi, yaitu dapat mengaktifkan seluruh fase sistem imun, meningkatkan respon imun terhadap seluruh antigen (proses inaktivasi dapat menyebabkan perubahan antigenitas), durasi imunitas lebih panjang, biaya lebih murah, lebih cepat menimbulkan respon imunitas dan mudah dibawa ke lapangan. Tetapi vaksin jenis ini memiliki beberapa kelemahan di mana vaksin ini kurang baik apabila digunakan pada daerah tropis dan pada penderita penyakit immunodeficiency serta adanya kemungkinan terjadi mutasi balik yang menyebabkan daya virulensi menjadi tinggi (Tetriana dan Sugoro, 2007).
Inaktivasi bakteri dengan cara iradiasi sinar gamma akan mengubah konfigurasi molekuler dari sel bakteri, salah satunya adalah molekul protein yang diasumsikan bahwa konfigurasi 3 dimensinya berubah menjadi terbuka dan siap melakukan suatu reaksi (Darussalam, 1996). Namun demikian, untuk mendukung hal tersebut perlu dilakukan analisis profil protein sebelum dan sesudah diiradiasi. Analisa tersebut dapat dilakukan melalui elektroforesis SDS-PAGE (Sodium dodecyl sulphate polyacrylamide gel electrophoresis), sehingga dapat diketahui apakah protein bakteri tersebut mengalami perubahan yang signifikan atau tidak setelah proses iradiasi.
1.2 RUMUSAN MASALAH
Dari uraian latar belakang di atas, maka masalah penelitian yang dapat dirumuskan adalah bagaimana kecenderungan profil protein isolat E. coli S1 hasil iradiasi sinar gamma sebagai bahan vaksin mastitis inaktif?
1.3 TUJUAN PENELITIAN
Penelitian ini bertujuan untuk:
1. Mengetahui dosis iradiasi sinar gamma yang dapat menginaktivasi sel bakteri E. coli.
1.4 HIPOTESIS
Hipotesis yang digunakan dalam penelitian ini sebagai berikut:
H0 = Ada pengaruh yang signifikan antara dosis iradiasi terhadap profil
protein yang dihasilkan.
H1 = Tidak ada pengaruh yang signifikan antara dosis iradiasi terhadap
profil protein yang dihasilkan.
1.5 MANFAAT PENELITIAN
BAB II
TINJAUAN PUSTAKA
2.1Bakteri Escherischia coli
Klasifikasi Ilmiah Filum : Proteobacteria
Kelas : Gamma Proteobacteria Ordo : Enterobacteriales Familia : Enterobacteriaceae Genus : Escherichia
Spesies : E. coli
Gambar.1. Morfologi Bakteri E. coli (Feng, et. al, 2002)
(terdiri atas fosfolipid, protein dan lipopolisakarida). Sedangkan di dalam sitoplasma terdapat beberapa substansi, yaitu:
1. Ribosom, merupakan partikel kecil yang tersebar dalam sitoplasma terdiri dari 35% protein dan 65% asam ribonukleat (RNA), berfungsi sebagai tempat sintesis protein.
2. Granul, merupakan tempat penyimpanan makanan berupa polisakarida, lipida, polifosfat, metafosfat, sulfur, dan poli beta hidroksi asam butirat (PBH).
3. Bahan Nukleus, merupakan untaian ganda DNA mencapai 1,2 mm, tetapi terlipat menjadi diameter 2 nm, berfungsi sebagai pembawa informasi genetik.
(a) Flagel (b) Pili
Gambar 2. Bagian permukaan membran luar yang terdapat pada bakteri gram negatif
Pada E. coli terdapat tiga jenis antigen, yaitu O-antigen yang merupakan inti lipopolisakarida dan unit-unit polisakarida, K-antigen yang merupakan kapsul, dan H-antigen yang merupakan flagel (Lehtolainen, 2004).
Gambar 3. Skema ilustrasi komponen-komponen bakteri gram negatif
Pada mulanya E. coli dianggap makhluk hidup terkecil yang dapat dibiakkan secara in-vitro, tetapi ternyata terdapat makhluk hidup lain yang mempunyai ukuran lebih kecil yang juga dapat dibiakkan secara in-vitro yaitu PPLO (Pleuropneumonia Like Organism) yang merupakan penyebab penyakit pada binatang maupun manusia, PPLO ini mempunyai ukuran diameter 0,1-0,25 mikron berarti jauh lebih kecil daripada bakteri apalagi sel-sel manusia (Juwono dan Zulfa, 2000).
Terdapat 3 (tiga) jenis bakteri E. coli, yaitu (1) Enteric - termasuk jenis yang paling umum. Tanda klinis utama yang ditimbulkan pada inangnya adalah diare hebat. Apabila Pedet (anak sapi) terserang bakteri ini, maka akan cepat menjadi lemas dan mengalami dehidrasi. Biasanya diawali dulu dengan demam yang kemudian dengan cepat kembali normal, atau mendekati normal, dan dapat
menyebabkan kematian. (2) Enterotoxigenic - Disebabkan oleh bakteri E. coli dari jenis K-99. Infeksi dari strain ini berakibat fatal. Racun menyebabkan cairan yang dipompa ke dalam usus sedemikian banyak sehingga pedet (anak sapi) biasanya mati bahkan sebelum gejala diare (mencret) muncul. Diare seperti ini adalah salah satu diare yang dapat muncul pada umur pedet (anak sapi) di bawah 3 hari. (3)
Septicemic - Jenis ini bekerja mirip bakteri Salmonella. Metodanya adalah dengan menginfeksi aliran darah dan masuk ke dalam jaringan tubuh sehingga menyebabkan infeksi global. Luka dan jejak dari infeksi bakteri jenis ini biasanya tidak tampak secara jelas. Ini merupakan jenis E. coli yang ganas, seringkali menyebabkan kematian tanpa gejala klinis diare terlebih dahulu. Pedet (anak sapi) yang tidak mendapat atau dihentikan pemberian kolostrum, biasanya mati karena jenis septisemik ini(Manglayang, 2006).
Infeksi Bakteri
Bakteri E. coli menginfeksi pada masa laktasi hari ke-60. Ada 3 (tiga) faktor penyebab yang mempermudah
terjadinya mastitis, yaitu kondisi sapi sebagai inang, kondisi lingkungan yang buruk dan mikroorganisme sebagai agen
penyebab penyakit (Gambar 4).
Lingkungan Sapi
Gambar 4. Faktor-faktor yang mempengaruhi terjadinya mastitis.
Kondisi sapi yang sakit karena infeksi alat reproduksi (radang rahim), infeksi saluran pencernaan (mencret), radang kuku, dan penyakit kulit (kutil, eksim, dan cacar) akan mempermudah terinfeksi mastitis. Selain itu bentuk ambing, pakan, umur dan stadium laktasi pun dapat mempengaruhi, ambing yang bergantung sangat rendah akan mudah kontak dengan lantai kandang sehingga beresiko terserang mastitis. Kuantitas dan kualitas pakan yang tidak memadai (kurang gizi) menyebabkan hewan menjadi kurus, kelemahan dan terganggunya metabolisme. Makin tua sapi maka semakin peka, di mana mekanisme penutupan lubang puting susu semakin menurun dan proses penyembuhan semakin lama. Stadium laktasi, terutama pada minggu pertama dan minggu terakhir beresiko pula terinfeksi mastitis (Fuad, 2006).
Luka atau lecet pada ambing atau puting susu yang diakibatkan oleh lantai kandang yang kasar, kuku panjang atau
tajam, sikat yang keras, memerah susu dengan cara kasar, memerah dengan cara menarik puting dapat menyebabkan
mastitis di mana luka akan mendatangkan lalat (Tetriana dan Sugoro, 2007).
Kondisi lingkungan yang mempermudah kejadian mastitis adalah kondisi kandang ternak yang basah dan kotor dan peternak/pemerah/pekerja tidak memelihara tubuh seperti kuku yang panjang, memakai pakaian kotor dan lain sebagainya (Sugiwaka, 2005).
Pada umumnya mastitis disebabkan oleh bakteri seperti Streptococcus agalactiae, Staphylococcus aureus, Streptococcus dysgalactiae, Streptococcus.
Escherichia coli, Klebsiella pneumoniae, Klebsiella oxytoca, dan Enterobacter aerogenes.
Kerugian yang diakibatkan mastitis adalah berkurangnya produksi susu hingga 25-30% atau berhenti sama sekali, kualitas susu menjadi turun sehingga tidak dapat dijual atau dikonsumsi, biaya perawatan meningkat, dan sapi dapat diafkir lebih awal (Fuad, 2006).
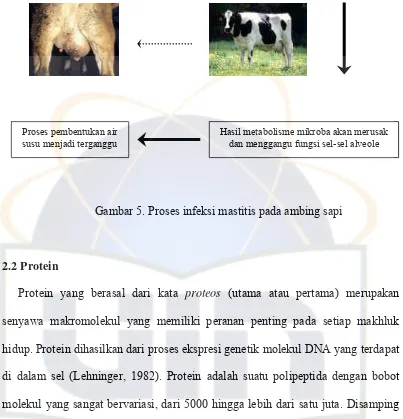
Mastitis terjadi karena masuknya bakteri melalui saluran puting dan berkembang biak dengan cara pembelahan di dalam susu yang diproduksi jaringan sehingga menimbulkan radang (Gambar 5). Mikroorganisme akan menembus saluran puting dengan berbagai cara diantaranya pemerahan, di mana mikroorganisme masuk melalui saluran puting atau hasil perpindahan secara fisik dari ujung puting yang ditekan akibat pergerakan sapi. Selama pemerahan dengan mesin, mikroorganisme mungkin terdorong masuk atau melalui saluran puting masuk ke dalam puting. Penularan dari ambing yang terinfeksi mastitis ke ambing sehat dapat terjadi melalui kain lap ambing yang digunakan untuk seluruh ternak, tangan pemerah yang kotor, urutan pemerahan yang salah dan peralatan pemerahan yang kotor (Tetriana dan Sugoro, 2007).
Mikroba masuk ke dalam ambing melalui lubang puting
Gambar 5. Proses infeksi mastitis pada ambing sapi
2.2Protein
Protein yang berasal dari kata proteos (utama atau pertama) merupakan senyawa makromolekul yang memiliki peranan penting pada setiap makhluk hidup. Protein dihasilkan dari proses ekspresi genetik molekul DNA yang terdapat di dalam sel (Lehninger, 1982). Protein adalah suatu polipeptida dengan bobot molekul yang sangat bervariasi, dari 5000 hingga lebih dari satu juta. Disamping berat molekul yang berbeda-beda, protein mempunyai sifat yang berbeda-beda pula (Poedjiadi, 1994), dengan fungsi yang spesifik ditentukan oleh gen yang sesuai (Lehninger, 1982).
Pada sel E. coli protein terbagi atas 2 (dua) bagian, yaitu protein intraselular dan protein Ekstraselular. Protein intraselular adalah protein yang terdapat dalam membran sel, ribosom, dan bahan nukleus. Sedangkan protein ekstraselular adalah protein yang terdapat dalam dinding sel, membran luar, dan flagel (Yatim, 1999).
Proses pembentukan air susu menjadi terganggu
Berdasarkan fungsi dan aktifitas biologisnya protein dikelompokkan menjadi beberapa golongan, antara lain enzim yang merupakan biokatalisator dari hampir semua reaksi yang terjadi pada makhluk hidup. Protein lain berfungsi sebagai protein transport seperti hemoglobin yang mengikat dan membawa oksigen menuju jaringan peri-peri dan lipoprotein yang membawa lipid dari hati ke organ lain, sementara protein lainnya berfungsi sebagai protein nutrien seperti ovalbumin dan kasein, serta protein pelindung seperti immunoglobulin yang berperan dalam sistem kekebalan tubuh. Di samping itu terdapat pula protein yang berperan sebagai protein pengatur seperti hormon insulin yang berperan dalam mengatur keseimbangan kadar glukosa dalam darah (Lehninger, 1982).
Struktur protein
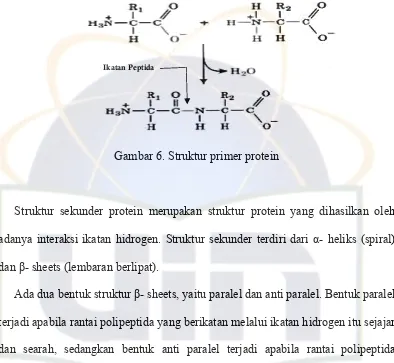
Protein tanpa memandang fungsi dan aktivitas biologisnya bangun oleh susunan dasar yang sama yaitu 20 asam amino yang molekulnya sendiri tidak memiliki aktivitas biologis. Masing-masing asam amino dalam suatu protein terintegrasi melalui ikatan peptida yang tersusun secara kovalen membentuk struktur yang beragam bergantung dari proses pembentukan dan fungsi dari protein tersebut (Hermanto, 2003).
struktur primer protein juga menunjukkan ikatan peptida yang urutannya diketahui.
Gambar 6. Struktur primer protein
Struktur sekunder protein merupakan struktur protein yang dihasilkan oleh adanya interaksi ikatan hidrogen. Struktur sekunder terdiri dari α- heliks (spiral) dan β- sheets (lembaran berlipat).
Ada dua bentuk struktur β- sheets, yaitu paralel dan anti paralel. Bentuk paralel terjadi apabila rantai polipeptida yang berikatan melalui ikatan hidrogen itu sejajar dan searah, sedangkan bentuk anti paralel terjadi apabila rantai polipeptida berikatan dalam posisi sejajar tetapi berlawanan arah.
α- heliks β- sheets (paralel) β- sheets (anti paralel)
Gambar 7. Struktur sekunder protein
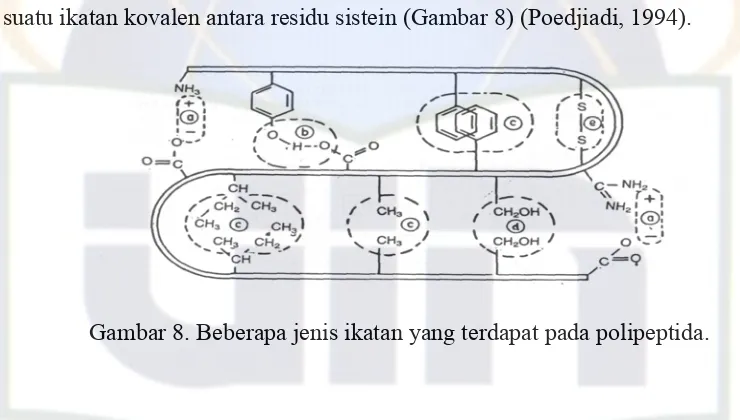
Struktur tersier menunjukkan kecenderungan polipeptida membentuk lipatan atau gulungan, dan dengan demikian membentuk struktur yang lebih kompleks. Struktur ini dimantapkan oleh adanya beberapa ikatan antara gugus R pada molekul asam amino yang membentuk protein. Beberapa jenis ikatan tersebut misalnya (a) ikatan elektrostatik, (b) ikatan hidrogen, (c) interaksi hidropob antara rantai samping non polar, (d) interaksi dipol-dipol dan (e) ikatan disulfida yaitu suatu ikatan kovalen antara residu sistein (Gambar 8) (Poedjiadi, 1994).
Gambar 8. Beberapa jenis ikatan yang terdapat pada polipeptida.
Gambar 9. Struktur kuartener protein globular yang kompleks.
Stabilitas Protein
Stabilitas struktur protein yang terbentuk karena adanya interaksi intramolekuler dan interaksi intermolekuler sangat dipengaruhi oleh kondisi lingkungan di sekitarnya. Kondisi eksternal seperti perubahan pH yang ekstrim atau pengaruh panas dapat menyebabkan struktur tiga dimensi protein rusak dan kehilangan aktivitas biologisnya. Oleh karenanya protein memerlukan kondisi tertentu yang memungkinkan senyawa tersebut dapat menjalankan aktivitas biologisnya secara optimal (Hermanto, 2003).
Gambar 10. Sketsa porses denaturasi protein (Winarno, 2002)
2.3Antigen
Antigen ialah suatu substansi yang bila memasuki inang vertebrata, menimbulkan respons kekebalan yang membawa kepada terbentuknya kekebalan dapatan. Respon kekebalan ini mengakibatkan pembentukan antibodi spesifik yang beredar di dalam aliran darah (imunitas humoral) atau merangsang peningkatan jumlah sel-sel reaktif khusus yang disebut limfosit (imunitas yang diperantarai sel atau cell-mediated immunity) atau keduanya. Limfosit ini telah memperoleh kemampuan yang lebih tinggi untuk menghancurkan sel-sel lain. Baik antibodi maupun limfosit khusus bereaksi dengan antigen yang digunakan sebagai bahan untuk membentuk kekebalan. Dengan cara ini kekebalan dapatan memungkinkan tubuh dapat menghancurkan atau menetralkan mikroorganisme penyerang ataupun toksinnya. Ini merupakan jalur utama pertahanan internal tubuh terhadap mikroba patogenik (Pelchzar dan Chan, 1988).
Sifat-sifat antigen
respons kekebalan. Hanya ada dua kelompok senyawa, secara alamiah yang jelas bersifat imunogenik, artinya mempunyai kemampuan untuk merangsang respons kekebalan. Senyawa yang dimaksud ialah protein dan polisakarida. Protein pada umumnya lebih efektif dalam merangsang pembentukan antibodi dibandingkan dengan polisakarida. Namun, polisakarida kompoleks berukuran besar, seperti polisakarida pada kapsul pneumokokus, merupakan antigen yang baik karena menimbulkan reaksi kekebalan yang kuat. Oligosakarida, lipid, dan asam-asam nukleat tidak merangsang pembentukan antibodi bila berdiri sendiri, tetapi dapat melakukannya bila bergabung dengan protein, yang disebut hapten (Pelchzar dan Chan, 1988).
Antigen dapat berupa substansi yang dapat larut seperti toksin bakteri atau protein serum (bagian zat alir dari darah yang terkoagulasi). Antigen dapat pula bersifat partikulat, seperti sel bakteri atau virion. Antigen partikulat biasanya lebih ampuh daripada antigen yang dapat larut. Antigen alami salah satunya adalah antigen bakteri yang diekskresikan sebagai eksotoksin dan enzim atau komponen struktural sel (Pelchzar dan Chan, 1988).
2.4 Faktor Virulensi Mikroba
Pada kebanyakan kasus, sifat-sifat yang menyebabkan virulensi suatu mikroorganisme patogenik itu tidak jelas atau belum diketahui. Namun, telah diketahui bahwa beberapa bakteri mengekskresikan substansi, sedangkan yang lain mempunyai struktur khusus yang turut menyumbang kepada virulensinya. Beberapa faktor mikrobial tersebut antara lain:
Beberapa mikroorganisme menghasilkan zat beracun yang dikenal sebagai toksin. Kemampuan suatu mikroorganisme untuk menghasilkan suatu toksin yang mempunyai efek buruk terhadap inang dan keampuhan toksin tersebut merupakan faktor penting di dalam kemampuan organisme tersebut untuk menyebabkan penyakit. Toksin yang dihasilkan mikroorganisme mungkin diekskresikan ke medium di sekitarnya (eksotoksin) atau di simpan di dalam selnya (endotoksin) sebagai bagian dari sel tersebut (Pelchzar dan Chan, 1988).
Eksotoksin.
Eksotoksin dapat berdifusi dan diekskresikan dari sel mikroba yang menghasilkannya ke dalam medium biakan atau ke dalam sistem peredaran dan jaringan inang. Eksotoksin adalah protein. Toksisitasnya akan hilang bila dipanaskan atau diberi perlakuan dengan zat kimia. Fenol, formaldehid, β– propiolakton, dan berbagai asam dapat memodifikasi eksotoksin secara kimiawi sehingga toksisitasnya lenyap; dalam hal demikian maka disebut
toksoid. Toksin dan toksoid mempunyai kemampuan untuk merangsang pembentukan antitoksin, yaitu substansi yang menetralkan toksisitas toksin di dalam tubuh inang. Kemampuan ini penting untuk melindungi inang yang rentan terhadap penyakit-penyakit yang disebabkan oleh toksin bacterial (Pelchzar dan Chan, 1988).
Endotoksin.
menghasilkan endotoksin yang dilepaskan hanya bila selnya hancur. Adanya substansi beracun di dalam populasi bakteri semacam itu disebabkan karena terlisisnya beberapa dari sel itu. Endotoksin bakteri gram negatif terletak pada dinding sel dan merupakan substansi kompleks yang mengandung fosfolipid dan karbohidrat (lipopolisakarida). Dibandingkan dengan eksotoksin, endotoksin itu (1) relatif stabil terhadap panas (2) tidak membentuk toksoid, dan (3) kurang toksik (Pelchzar dan Chan, 1988).
2. Enzim ekstraselular.
Virulensi beberapa mikroorganisme sebagian disebabkan oleh produksi enzim ekstraseluler (lihat tabel 1). Kendati tidak ada satu pun enzim ekstraselular tunggal yang telah diperlihatkan tanpa meragukan sebagai satu-satunya faktor penyebab virulensi, tidak disangsikan lagi enzim-enzim semacam itu berperan di dalam proses patogenik (Pelchzar dan Chan, 1988). Tabel 1. Beberapa enzim ekstraselular yang turut menentukan virulensi mikroba.
Leukosidin
(suatu serat jaringan)
Membunuh leukosit Staphylococcus aureus
Sumber: Pelchzar dan Chan, 1988.
3. Kapsul
Virulensi bakteri dalam banyak hal dipengaruhi oleh ada atau tidak adanya kapsul. Bila patogen kehilangan kapsulnya, misalnya oleh mutasi, maka mereka kehilangan kemampuannya sebagai penyebab penyakit. Nampaknya bertambahnya virulensi pada galur berkapsul disebabkan oleh kemampuan polisakaride kapsul untuk mencegah fagositosis atau penelanan oleh fagosit inang. Kemampuan ini mungkin disebabkan oleh sifat-sifat permukaan kapsul yang mencegah fagosit sehingga tidak membentuk kontak yang cukup erat dengan bakteri yang menelannya (Pelchzar dan Chan, 1988).
4. Pili
Banyak bakteri nonpagotenik memiliki pili, sama halnya seperti banyak bakteri pagotenik memiliki kapsul. Organel-organel ini dapat meningkatkan virulensi beberapa patogen. Telah dilaporkan bahwa dimilikinya pili membantu organisme melekat dengan lebih baik pada permukaan sel inang dan jaringan inang (Pelchzar dan Chan, 1988).
2.5 Vaksin
mematikan atau menatenuasi (menghentikan transkripsi DNA pada suatu kodon, sehingga mRNA yang terbentuk lebh pendek dari biasanya), sehingga tidak akan menimbulkan penyakit dan dapat merangsang pembentukan kekebalan/antibodi bila di inokulasikan (Sugoro, 2004), atau hasil-hasil pemurniannya seperti protein, peptida, partikel serupa virus, dan sebagainya (Pelchzar dan Chan, 1988).
Bila unsur asing ini menyerang tubuh, maka sistem kekebalan akan mengaktifkan sel-sel tertentu untuk menghancurkan unsur asing tersebut. Jika tubuh diserang ulang bakteri atau virus di masa datang, sel ingatan akan diaktifkan dan menjawab lebih cepat dan lebih kuat untuk menghancurkan bakteri atau virus. Vaksin juga bisa membantu sistem kekebalan tubuh melawan sel-sel degeneratif seperti kanker (Medical Research News, 2006).
Pada gambar 11 menunjukkan mekanisme kerja vaksin, di mana ketika vaksinasi berlangsung, vaksin yang berasal dari virus, bakteri atau organisme yang telah mati maupun yang sudah dalam bentuk aman, disuntikkan ke dalam sistem (kiri). Vaksin merangsang sistem kekebalan tubuh untuk memproduksi
Gambar 11. Mekanisme kerja vaksin (Scorvia, 2000)
Vaksin Vaksin Antibodi Antibodi Organisme
antibodi terhadap suatu organisme (tengah). Kapanpun tubuh terserang oleh kuman ini setelah vaksinasi, antibodi pada sistem kekebalan tubuh menyerang dan menghentikan infeksi (kanan).
Aplikasi Teknik Nuklir Dalam Bidang Vaksin
Infeksi merupakan masalah besar dalam kesehatan dan telah menghabiskan dana yang sangat besar. Hilangnya harapan hidup atau produktivitas akibat penyakit infeksi bukan sekedar masalah kesehatan semata, tetapi juga menyangkut permasalahan sosial dan ekonomi. Infeksi ini dapat menyerang manusia maupun hewan sebagai inang atau vektor (Tetriana dan Sugoro, 2007).
Infeksi ataupun penyakit akibat infeksi pada manusia telah menyebabkan kematian sebesar 13 juta orang di seluruh dunia setiap tahun, terutama di negara-negara berkembang seperti Indonesia (IAEA, 2000). Empat puluh tiga persen kematian di negara berkembang disebabkan oleh penyakit infeksi sedangkan di negara maju hanya sebesar satu persen. Kematian yang besar tersebut dapat dicegah jika dilakukan diagnosa yang cepat dan tepat serta didukung oleh penanganan yang efektif dan efisien (Machi, S, 2002). Pada hewan, penyakit infeksi telah menurunkan tingkat produksi dan kualitas hasil ternak. Selain itu, terinfeksi suatu penyakit menyebabkan hewan di karantina atau dibunuh (Tetriana dan Sugoro, 2007).
terjadi melalui kulit, membran mukosa, transfusi darah, ataupun kontaminasi dari makanan dan minuman. Oleh karena itu diperlukan penanganan yang tepat dalam mengendalikan penyakit infeksi. Tujuan utama dari penanganan adalah menghilangkan atau membunuh organisme patogen penyebab infeksi. Beberapa penanganan yang umumnya dilakukan adalah mencegah kontaminasi dari lingkungan, memutuskan siklus hidup dari organisme patogen, memutuskan hubungan antara inang dan vektor, menghancurkan reservoir host dan mencegah perkembangan organisme patogen melalui pengobatan dan vaksinasi (Machi, S, 2002).
Salah satu alternatif yang dapat digunakan dalam penangan penyakit infeksi ini adalah dengan menggunakan teknik nuklir. Berbagai penyakit yang bersumber dari virus, bakteri, protozoa dan cacing telah banyak yang memanfaatkan teknik nuklir dalam proses pembuatan bahan vaksinnya (Tabel 2). Vaksin dapat merangsang sistem imun pada inang untuk melawan infeksi organisme patogen (Tetriana dan Sugoro, 2007; Ramamoorthy,S., et. al., 2006).
Tabel 2. Vaksin iradiasi yang telah dihasilkan
Penyakit Jenis vaksin Target vaksinasi
TBC
Young (1981), dalam percobaannya menyatakan bahwa iradiasi dapat mengubah agen penyakit patogen menjadi non patogen yang mampu menstimulasi sistem kekebalan dalam tubuh. Smith (1992), juga menyatakan bahwa teknik nuklir/ iradiasi dapat melemahkan agen penyakit tanpa menghilangkan daya imunogeniknya dan mampu meningkatkan daya kekebalan pada hewan yang dicobakan.
Berdasarkan bahan dasarnya, vaksin dibagi menjadi empat tipe yaitu (1) vaksin dengan bahan dasar organisme patogen yang dimatikan atau inaktif; (2) vaksin dengan parasit yang dilemahkan atau daya virulensinya rendah; (3) vaksin dengan subunit protein hasil purifikasi, rekombinasi atau proses kimia; dan (4) vaksin asam nukleat baik DNA maupun RNA (Nature, 2005).
Suatu materi hidup seperti sel, apabila terkena sinar gamma akan mengalami kerusakan secara langsung atau tidak langsung. Efek langsung adalah terjadinya pemutusan ikatan senyawa-senyawa penyusun sel. Efek tidak langsung terjadi karena materi sel terbanyak adalah air yang apabila terkena sinar gamma akan mengalami hidrolisis dan menghasilkan radikal bebas. Radikal bebas inilah yang akan menyebabkan kerusakan materi sel seperti molekul enzim, DNA, RNA, dan yang lainnya. Mengingat bahwa 80% komposisi organisme hidup terdiri dari air, maka hampir setiap efek radiasi terhadap sistem biologi sebagian besar diawali
Vaksin inaktif Vaksin aktif
Vaksin aktif
Gambar 12. Efek radiasi terhadap sel yang dapat dimanfaatkan untuk pembuatan bahan vaksin iradiasi.
Akibat kerusakan DNA
Sel mati Mutasi Transformasi
Akibat kerusakan DNA
Memperbaiki DNA
oleh peristiwa pengaktifan (radiolisis) molekul air (Hall, E.J., 1994 ; Darussalam 1996).
Vaksin yang menggunakan iradiasi dibagi menjadi dua macam, yaitu vaksin aktif dan vaksin inaktif. Vaksin aktif adalah vaksin dengan bahan dasar organisme hidup yang telah dilemahkan dengan proses iradiasi, sedangkan vaksin inaktif adalah vaksin dengan bahan dasar organisme mati hasil iradiasi. Vaksin inaktif sendiri dibagi menjadi dua, yaitu vaksin inaktif rekombinan dan non rekombinan. Vaksin inaktif rekombinan diperoleh dengan cara melemahkan organisme terlebih dahulu melalui teknik rekombinan setelah itu diinaktivasi dengan iradiasi. Vaksin inaktif non rekombinan adalah pemakaian iradiasi untuk inaktivasi organisme patogen secara langsung (Nature, 2005).
Vaksin aktif yang telah dilemahkan biasanya digunakan untuk parasit yang bersifat intraselular yang berasal dari protozoa dan cacing. Beberapa penelitian vaksin yang saat ini dikembangkan baik pada manusia maupun hewan menggunakan teknik iradiasi untuk melemahkan organisme patogen, seperti protozoa dan cacing (Jenkins, 2002).
mutasi balik yang menyebabkan daya virulensi menjadi tinggi (Tetriana dan Sugoro, 2007).
Hasil percobaan terdahulu menunjukkan bahwa booster yang diberikan akan bermanfaat apabila diberikan pada saat tingkat produksi/ titer antibodi menjelang puncaknya, sehingga akan meningkatkan daya kekebalan pada hewan yang bersangkutan. Di samping itu pertambahan bobot badan hewan tidak terganggu karena parasit penantang yang diberikan tidak bisa berkembang dan tidak aktif lagi (Arifin, 2006).
Vaksin inaktif contohnya Leishmania, yaitu penyakit Kala-azar yang ditimbulkan oleh protozoa. Keuntungan vaksin ini adalah memberikan imunitas humoral yang tinggi bila booster diberikan, tidak menyebabkan mutasi atau reversion, dapat digunakan untuk pasien immuno-defisiensi, cocok digunakan untuk daerah tropis tetapi vaksin jenis ini membutuhkan biaya yang lebih tinggi karena membutuhkan booster (Gaffar, A., 2006).
Vaksin inaktif rekombinan contohnya untuk penyakit yang disebabkan bakteri
2.6Radiasi dan Iradiasi
Radiasi merupakan proses yang kejadiannya berlangsung tanpa unsur kesengajaan atau tanpa adanya perlakuan khusus, misalnya: bentuk mutasi pada tanaman dapat terjadi secara alamiah (spontan) akibat radiasi sinar kosmik di alam. Sedangkan iradiasi merupakan proses yang kejadiannya berlangsung karena adanya perlakuan khusus terhadap sesuatu obyek yang dilakukan secara disengaja (misalnya untuk tujuan melakukan suatu pengamatan atau penelitian), contoh: bahan makanan yang telah diiradiasi (the irradiated food) dengan sinar gamma dapat menjadi awet dan tidak cepat membusuk ataupun rusak (Darussalam, 1996).
Beberapa macam radiasi dapat bersifat letal (mematikan) terhadap sel-sel mikroba dan juga sel-sel organisme lain. Radiasi macam ini meliputi bagian dari spektrum elektromagnetik (radiasi ultraviolet, gamma, dan sinar X) dan sinar-sinar katode (elektron berkecepatan tinggi).
Proses perubahan materi akibat radiasi sangatlah kompleks, tetapi untuk penyederhanaannya dapat digambarkan dalam empat tahapan:
1. Tahap Fisik Awal
Tahap awal ini berlansung hanya kira-kira 10-16 detik, energi terdeposit di dalam sel dan menyebabkan ionisasi. Di air reaksinya dapat dinyatakan sebagai: H2O → H2O+ + e-. (H2O+ adalah ion positif dan e- ion negatif).
Gambar 13. Peristiwa terjadinya ionisasi dan eksitasi
2. Tahap Kimia – Fisik
Tahap ini berlangsung kira-kira 10-6 detik. Ion-ion berinteraksi dengan molekul air lainnya yang menghasilkan beberapa produk baru. Sebagai contoh, ion positif terdisosiasi :
H2O+→ H+ + OH
-Ion negatif, yaitu elektron yang terikat pada molekul air netral yang selanjutnya terdisosiasi
H2O+ + e-→ H2O
H2O-→ H + OH-
Sehingga produk dari reaksinya adalah H+ , OH- ,H dan OH. Dua ion pertama yang ada dalam sebagian besar air, tidak mengambil bagian dalam reaksi berikutnya. Dua produk lainnya, H dan OH disebut radikal bebas, yaitu mereka yang mempunyai elektron yang tidak berpasangan dan secara kimia sangat reaktif. Hasil reaksi lainnya adalah hidrogen peroksida H2O2, yang
merupakan oksidan yang sangat kuat dan terbentuk dengan reaksi: OH + OH
3. Tahap Kimia
Tahap kimia berlangsung hanya beberapa detik, dimana hasil reaksi berinteraksi dengan molekul-molekul organik yang penting dalam sel. Radikal bebas dan oksidan dapat menyerang molekul komplek yang membentuk kromosom. Misalnya, sebagai contoh radikal tersebut dapat mengikatkan dirinya ke molekul atau meyebabkan ikatan rantai panjang menjadi putus. Perubahan tingkat molekular secara kimia dapat diakibatkan oleh:
a) Kerja langsung radiasi (direct action), dimana perubahan atau kerusakan terjadi pada molekul-molekul biologi yang menyerap lansung energi radiasi.
b) Kerja tidak lansung radiasi (indierct action), dimana perubahan atau kerusakan pada molekul-molekul terjadi akibat pengaruh senyawa radikal-radikal bebas dan pengoksida. Sedangkan senyawa radikal-radikal-radikal-radikal bebas dan pengoksida merupakan hasil peristiwa radiolisis molekul air dalam plasma akibat radiasi (Darussalam, 1996).
4. Tahap Biologi
1. Efek fisiologis radiasi, bersifat sementara dan dapat pulih kembali (recovery). Contoh: penghambatan pertumbuhan dan gangguan sifat permeabilitas selaput (dinding) sel.
2. Efek genetik radiasi, menghasilkan bentuk mutasi pada keturunan organisme dengan sifat-sifat yang berbeda dari induknya. Kejadian mutasi menyusul kerusakan pada molekul DNA dan kromosom.
3. Efek somatik radiasi, menghasilkan bentuk sel-sel somatik abnormal sebagai hasil pembelahan sel secara mitosis.
Terdapat 2 (dua) macam efek somatik radiasi:
a. Jenis stokastik, yaitu merupakan efek tertunda. Contoh: terjadinya kanker pada darah (leukimia), tulang dan paru-paru. b. Jenis stokastik, di mana terdapat hubungan antara dosis paparan
radiasi dan efek radiasi. Contoh: luka bakar pada kulit (erythema), katarak mata, penurunan jumlah sel-sel gonad (kemandulan).
O
CH N H2 R
O H
Efek Radiasi terhadap Molekul Penting
a) Protein
Protein terdiri dari beberapa asam amino yang secara karakteristik di tandai oleh adanya gugus -NH2, gugus -COOH dan rantai samping R.
Gambar 14. Formula asam amino
Gugus -NH2 dan rantai samping –R asam amino yang paling radiosensitif di
antara tiga kelompok di atas. Kerusakan fungsi protein sebagian besar disebabkan oleh timbulnya perubahan pada rantai samping (-R) yang kritis. Diasumsikan bahwa radiasi dapat mempengaruhi konfigurasi 3 dimensi molekul protein sehingga menjadi terbuka dan siap melakukan suatu reaksi. (Darussalam, 1996).
b) Kromosom
Radiasi dapat menyebabkan perubahan atau kerusakan struktur kromosom (aberasi). Perubahan atau kerusakan itu dapat berupa antara lain: pelengketan, translokasi, inversi, defisiensi, dan lain sebagainya (Darussalam, 1996).
c) Molekul Asam Deoksiribonukleat (DNA)
Setiap Asam nukleat (nucleic acid) merupakan makromolekul yang berperan sebagai pembawa informasi genetik. Kerusakan secara biokimiawi yang terjadi pada molekul DNA dapat mengarah kepada kejadian mutasi baik pada sel-sel germa maupun pada sel-sel somatik. Bentuk mutasi pada sel-sel germa akan tampak pada keturunan berikutnya, sementara bentuk mutasi pada sel-sel somatik terlihat pada bentuk abnormal pada sel-sel anakan hasil pembelahan mitosis (Darussalam, 1996).
d) Enzim
Radiasi dapat menyebabkan kerusakan biokimiawi pada struktur enzim. Kerusakan enzim dapat mengakibatkan penurunan pada fungsi enzim, yaitu terjadinya penurunan daya katalisasi enzim dalam proses reaksi kimia tertentu (Darussalam, 1996).
e) Lemak
Radiasi dapat menimbulkan perubahan dan kerusakan pada bagian ikatan rangkap asam lemak (Darussalam, 1996).
f) Karbohidrat
Radiasi dapat menimbulkan penguraian dan kerusakan (degradasi) pada rantai karbohidrat (Darussalam, 1996).
Efek Radiasi terhadap Pembelahan Sel
kromosom pada stadia anafase ataupun telofase. Semua ini dapat mengarah kepada tertundanya atau terhentinya proses pembelahan sel. Jika proses pembelahan sel dihambat atau dihalangi secara terus-menerus dapat menimbulkan kematian sel atau jaringan (Darussalam, 1996).
2.7 Sinar Gamma
Sinar gamma (seringkali dinotasikan dengan huruf Yunani gamma, γ) adalah sebuah bentuk energi dari radiasi elektromagnetik tinggi yang diproduksi oleh transisi energi karena percepatan elektron atau radioaktifitas atau proses nuklir atau subatomik yang lainnya seperti penghancuran elektron-positron.
Sinar gamma merupakan gelombang elektromagnetik yang bergerak dengan kecepatan sangat tinggi, hampir menyamai kecepatan cahaya. Arahnya tidak dipengaruhi medan magnet dan mempunyai daya ionisasi kecil serta daya tembus yang tinggi. Dalam hal ionisasi, radiasi gamma berinteraksi dengan bahan melalui tiga proses utama, yaitu:
1. Efek fotolistrik
Gambar 15. Efek fotolistrik
2. Efek penghamburan Compton
Pada efek Compton, foton dengan energi hv berinteraksi dengan elektron terluar dari atom, selanjutnya foton dengan energi hv dihamburkan dan elektron tersebut dilepaskan dari ikatannya dengan atom dan bergerak dengan energi kinetik tertentu (Yudi, 2008).
Gambar 16. Efek penghamburan Compton
3. Efek produksi pasangan
Proses produksi pasangan hanya terjadi bila foton datang / 1,02 MeV. Apabila foton semacam ini mengenai inti atom berat, foton tersebut akan lenyap dan sebagai gantinya timbul sepasang elektron-positron. Positron adalah partikel yang massanya sama dengan elektron dan bermuatan listrik positif yang
RADIASI
besarnya juga sama dengan muatan elektron. Proses ini memenuhi hukum kekekalan energi:
hv = (2m 0C2) + (K+) + (K-)
Keterangan: K+ = energi kinetik positron
K- = energi kinetik elektron
Oleh karena itu proses ini hanya bisa berlangsung bilamana energi foton yang datang minimal. 2m0C2 (= 1,20 MeV), m0 adalah massa diam elektron dan c
adalah kecepatan cahaya. Berkaitan dengan uraian ini maka nilai atau besaran absorpsi linier akan bergantung pada energi foton yang datang disamping bergantung pada jenis media/materi/zat yang dilaluinya atau bergantung pada nomor atom (Z) media/materi yang dilaluinya (Yudi, 2008).
Sinar gamma mempunyai panjang gelombang yang lebih pendek daripada sinar X sehingga mempunyai energi yang lebih tinggi. Pada umumnya sinar gamma yang digunakan untuk radiasi adalah hasil peluruhan inti atom Cobalt-60. Cobalt-60 adalah sejenis metal yang mempunyai karakteristik hampir sama dengan besi/nikel. Cobalt-60 memancarkan dua sinar gamma dengan energi masing-masing sebesar 1,17 MeV dan 1,33 MeV yang mempunyai waktu paruh 5,27 tahun (Wayudi P., et. al., 2005; GIDAE, 1992 )
Dosimeter Standar
Dosimeter standar adalah alat untuk mengukur dosis iradiasi sinar gamma dosis tinggi, nilai dosis dapat ditentukan secara langsung dengan dosimeter standar, atau diukur dengan dosimeter lain yang sebelumnya telah dikalibrasi terhadap dosimeter standar. Ada dua jenis dosimeter standar yang hingga kini sering dimanfaatkan untuk dosimetri gamma dosis tinggi, yaitu dosimeter Fricke dan Ceri-cero (McLaughlin, 1989; IAEA, 1977).
1. Dosimeter Fricke
Dosimeter Fricke merupakan salah satu jenis pengukur dosis serap yang dipakai sebagai dosimeter acuan karena absorbsinya yang tinggi dan mempunyai hubungan yang linier terhadap dosis serap (ICRU). Dosimeter ini dibuat dari bahan kimia Fe(NH4)2(SO4)2.6H2O (ferro ammonium sulfat), NaCl
(natrium klorida) dan H2SO4 (asam sulfat) (Scrad, 1971).
tersebut disinari dihitung menggunakan persamaan SCHESTED (Bjerbakke, 1970).
Keunggulan dari dosimeter Fricke ini antara lain adalah laju dosis dari sumber yang diukur tidak melebihi 2 x 107 Gy/det dan temperatur tidak menyimpang selama proses iradiasi, sehingga laju dosis sumber tidak berpengaruh terhadap hasil pengukuran. International Commission on Radiation Units and Measurements (ICRU) juga menganjurkan penggunaan dosimeter Fricke untuk pengendalian mutu faktor kalibrasi alat ukur radiasi standar nasional, yaitu laju dosis serap yang ditentukan dengan dosimeter Fricke digunakan untuk mengevaluasi faktor kalibrasi alat ukur radiasi standar nasional yang diperoleh sebelumnya (Thamrin, 1997).
2. Dosimeter Ceri-Cero
Dosimeter Ceri-cero juga dapat dipakai sebagai dosimeter standar dalam pengukuran radiasi gamma dosis tinggi. Dosimeter ceri-cero sulfat untuk mengukur dosis tinggi dengan dengan jangkauan 10-1000 KGy sudah umum digunakan dalam proses radiasi. Larutan ceri sulfat dibuat menggunakan reagen Ce(SO4)2.4H2O, H2SO4 dan H2O2 30% yang dilarutkan dalam pelarut
perbedaan jumlah ion cero pada larutan yang diradiasi dengan larutan yang tidak diradiasi. Perubahan kerapatan optik pada dosimeter ceri-cero yang menerima paparan radiasi diukur menggunakan spektrofotometer pada panjang gelombang 320 nm (Thamrin, 1997).
Untuk keperluan pengukuran dosis secara rutin, kedua jenis dosimeter tersebut tidak tersedia dalam bentuk siap pakai, jadi pemakai harus meracik sendiri dosimeter sesuai dengan petunjuk yang telah dibakukan (McLaughlin, 1989; Scrad, 1971; Bjerbakke, 1970). Ada beberapa faktor yang membuat dosimeter tersebut kurang praktis apabila dipakai untuk keperluan rutin, misalnya Dosimeter Fricke, sangat peka terhadap pengotor organik dan cahaya (Spink, 1976). Untuk mendapatkan dosimeter yang baik diperlukan syarat-syarat yang ketat seperti kemurnian pelarut dan kebersihan peralatan. Pengukuran dosis pada dosimeter ceri-cero ini biasanya menggunakan spektrofotometer yang merupakan masalah tersendiri di lapangan karena berukuran besar dan tidak mungkin dibawa ke mana-mana. Selain itu, dalam proses pengukuran juga harus melakukan pengenceran berulang-ulang dan makan waktu yang relatif lama (Thamrin, 1997).
2.8Elektroforesis
molekul yang terseparasi pada gel dapat dideteksi dengan pewarnaan atau autoradiografi, ataupun dilakukan kuantifikasi dengan densitometer.
Menurut Yuwono (2005), elektroforesis adalah suatu teknik pemisahan molekul selular berdasarkan atas ukurannya, dengan menggunakan medan listrik yang dialirkan pada suatu medium yang mengandung sampel yang akan dipisahkan. Kecepatan gerak molekul tergantung pada nisbah (rasio) muatan terhadap massanya, serta tergantung pula pada bentuk molekulnya (Yuwono, 2005).
Kegunaan elektroforesis antara lain:
1. Menentukan berat molekul (estimasi). Penetapan BM secara lebih teliti dapat dilakukan dengan ultrasentrifuge, meskipun dengan elektroforesis cukup memenuhi syarat.
2. Dapat mendeteksi terjadinya pemalsuan bahan.
3. Dapat mendeteksi terjadinya kerusakan bahan seperti protein dalam pengolahan dan penyimpanan.
4. Untuk memisahkan spesies molekul yang berbeda secara kualitatif maupun kuantitatif, yang selanjutnya masing-masing spesies dapat dianalisis.
5. Menetapkan titik isoelektrik protein.
oligometrik, selain itu untuk menetapkan berat molekul dan jumlah rantai polipeptida sebagai subunit atau monomer.
Pada mekanisme SDS PAGE, protein bereaksi dengan SDS yang merupakan deterjen anionik membentuk kompleks yang bermuatan negatif. Protein akan terdenaturasi dan terlarut membentuk kompleks berikatan dengan SDS, berbentuk elips atau batang, dan berukuran sebanding dengan berat molekul protein. Protein dalam bentuk kompleks yang bermuatan negatif ini terpisahkan berdasarkan muatan dan ukurannya secara elektroforesis di dalam matriks gel poliakrilamid. Berat molekul protein dapat diukur dengan menggunakan protein standar yang telah diketahui berat molekulnya (Ummubalqis, 2000).
BAB III
METODA PENELITIAN
3.1Tempat dan Waktu Pelaksanaan
Penelitian ini dilaksanakan di Laboratorium Nutrisi, Kesehatan dan Reproduksi Ternak, Pusat Aplikasi Teknologi Isotop dan Radiasi (PATIR), Badan Tenaga Nuklir Nasional (BATAN), Pasar Jum’at Jakarta Selatan, mulai dari bulan Agustus 2007 sampai dengan bulan September 2007.
3.2Bahan dan Alat
Bahan:
Isolat bakteri E. coli S1 yang didapat dari Laboratorium Nutrisi, Kesehatan dan Reproduksi Ternak, Pusat Apalikasi Tenaga Isotop dan Radiasi (PATIR), Badan Tenaga Nuklir Nasional (BATAN), Pasar Jum’at Jakarta Selatan.
Alat:
Alat – alat yang digunakan dalam penelitian ini adalah: autoklaf (Hitachi), sonikator (Branson 2210), gelas beker, penangas air, bunsen, cawan petri, tabung Eppendorf , laminar flow, mikropipet, tabung reaksi, timbangan analitik, vortex, erlenmeyer, shaker inkubator, kuvet, gamma chamber 4000 A, spektrofotometer
vissble (Genesys), sentrifuge (Hitachi CR21G II 10000 rpm), mikro sentrifuge (Hitachi), vial gelas, tip kecil dan tip besar, Mini-Protein Gel Electrophoresis (Atto).
3.3Metoda Kerja
3.4Prosedur Kerja
3.4.1 Pembuatan kurva tumbuh Escherichia coli
Kultur bakteri berumur 1 hari pada medium NA diinokulasikan sebanyak 3 ose ke dalam 30 ml medium NB dan diinkubasi selama 24 jam pada suhu 37oC dan agitasi 120 rpm sebagai kultur inokulum. Setelah itu kultur diukur absorbannya dengan spektrofotometer vissible pada panjang gelombang 660 nm. Kultur diukur pada menit ke-0, 30, 60, 90, 150, 210, 270, 330, dan 390. Hasil pengukuran dibuat kurva dengan axis (waktu) dan ordinat (absorbansi).
3.4.2 Penentuan Dosis Iradiasi Inaktivasi sel E. coli dengan Sinar Gamma
Kultur pada fase mid log disentrifuge 10000 rpm dan dibilas dengan aquades steril sebanyak 2 kali. Pelet yang diperoleh diencerkan hingga jumlah sel diperoleh 1013 sel/ml dan ditempatkan di dalam vial gelas sebanyak 10 ml. Selanjutnya diiradiasi sinar gamma dengan dosis 0Gy, 50Gy, 200Gy, 500Gy, 600Gy, 700Gy, 800Gy, 900Gy, dan 1000Gy di Gamma Chamber 4000 A dengan laju dosis 1089,59 Gy/jam. Selanjutnya kultur hasil iradiasi diencerkan hingga jumlah sel diperoleh 1011 sel/ml dan ditempatkan di dalam vial gelas sebanyak 10 ml. Kultur hasil iradiasi kemudian dihitung jumlah selnya dengan metode droptest
3.4.3 Pengukuran Protein Sel E. coli Metode Lowry
Kultur hasil iradiasi diukur kandungan protein ekstraselular dan intraselular. Untuk mengetahui kandungan protein ekstraselular langsung menggunakan kultur hasil iradiasi, sedangkan untuk protein intraselular dipecah terlebih dahulu dengan melarutkan kultur hasil iradiasi ke dalam aseton (1 : 1) dan disonikasi selama 15 menit. Kemudian 1 ml sampel (ekstra dan intraselular) ditambahkan 5 ml larutan Lowry I (Lampiran 1) dan dibiarkan selama 10 menit. Setelah itu, ditambahkan 0,5 ml larutan Lowry II (Lampiran 1) dan dibiarkan selama 30 menit. Setelah itu, dibaca dengan spektrofotometer visible pada panjang gelombang 700 nm, dan dibandingkan dengan standar BSA.
3.4.4 Karakteristik Profil Protein Bakteri E. coli
Pada penelitian ini menggunakan metode elektroforesis 1 dimensi SDS-PAGE dengan sistem buffer Laemmli. Konsentrasi gel poliakrilamida yang digunakan adalah 10%.
a. Preparasi sampel
b. Preparasi gel elektroforesis - Separating gel (10%)
30% Acrylamide solution 6 ml ditambahkan separating gel buffer (1,5 M Tris-HCl, pH 8,8) sebanyak 4,5 ml, kemudian aquabides 7,5 ml dan 10%
Ammonium persulfate 0,08 ml serta TEMED 0,01 ml. - Stacking gel (45%)
30% Acrylamide solution 0,9 ml ditambahkan stacking gel buffer (0,5 M Tris-HCl, pH 6,8) sebanyak 1,5 ml, kemudian aquabides 3,6 ml dan 10%
Ammonium persulfate 0,02 ml serta TEMED 0,01 ml. c. Pembuatan kolom gel
Setelah separating gel dibuat kemudian dimasukkan sedikit demi sedikit ke dalam alat elektroforesis dengan mikropipet, lalu ditambahkan aquabides untuk meratakan separating gel tersebut. Setelah separating gel membeku dimasukkan stacking gel sedikit demi sedikit, lalu pasang sisir pembentuk kolom biarkan hingga stacking gel membeku lalu diangkat sisirnya. Kemudian dipasang hasil gel tersebut pada perangkat elektroforesis.
d. Loading sampel
Larutan buffer dimasukkan ke dalam tangki elektroforesis. Kemudian sampel sebanyak 15 µl dimasukkan ke dalam kolom gel dengan hati-hati lalu dielektroforesis selama + 100 menit pada 200 Volt, 40 mA.
e. Pewarnaan gel
f. Pencucian gel
Gel dicuci dengan larutan destain solution coomassie R-250, selama + 1 hari. Selanjutnya hasil pencucian discan dan dianalisis menggunakan LabImage 1D 2006 dan SPSS 11.5.
BAB IV
HASIL DAN PEMBAHASAN
Gambar 17 menunjukkan kurva pertumbuhan Isolat bakteri E. coli yang ditumbuhkan dalam media NB. Berdasarkan gambar tersebut bahwa kurva tumbuh tidak mengalami fase adaptasi (lag phase), tetapi langsung memasuki fase eksponensial (fase log). Dalam fase eksponensial terdapat suatu pertambahan semua komponen selular yang lain seperti DNA, RNA, dan protein secara teratur, kemudian setelah menit ke-330, pertumbuhan bakteri memasuki fase statis (stationary phase).
Gambar 17. Pertumbuhan Bakteri E. coli dalam medium TSB yang diinkubasikan pada suhu 370C dan agitasi 120 rpm
Menurut Pelchzar dan Chan (1988), bahwa kurva pertumbuhan bakteri terdiri dari 3 fase, yaitu periode awal yang tampaknya tanpa pertumbuhan (fase adaptasi), diikuti oleh suatu periode pertumbuhan yang cepat (fase log), kemudian
mendatar (fase seimbang atau stationary phase) dan akhirnya diikuti oleh suatu penurunan populasi sel hidup (fase kematian).
Kurva tumbuh ini digunakan untuk menentukan fase mid log, yaitu fase pertumbuhan dimana terjadi kecepatan pembelahan sel tertinggi. Fase ini terjadi pada menit ke-210, dengan kecepatan pertumbuhan berdasarkan nilai absorbansi adalah 0,09/menit. Fase mid log digunakan karena sel-sel dalam kondisi aktif melakukan metabolisme. Pada fase tersebut terjadi pembelahan yang cepat sehingga dinding selnya tipis dan efek radiasi dapat terjadi secara maksimal (Tetriana dan Sugoro, 2007). Menurut Alatas (2005), bahwa sel yang paling sensitif adalah sel dengan tingkat proliferasi yang tinggi (aktif melakukan pembelahan) dan tingkat diferensiasi yang rendah. Sedangkan sel yang resisten atau tidak mudah rusak akibat pengaruh radiasi yaitu sel dengan tingkat diferensiasi yang tinggi dan tidak aktif melakukan pembelahan.
4.2Hasil Inaktivasi sel E. colidengan Iradiasi Sinar Gamma
0 200 400 600 800 1000 1200
Dosis (Gy)
Gambar 18. Hubungan dosis iradiasi sinar gamma terhadap jumlah sel bakteri E. coli
Tabel 3. Jumlah sel E. coli hasil iradiasi sinar gamma.
Dosis (Gy) Jumlah sel/ml % Viabilitas
0 2,1 x 1013 100
Kondisi inaktif dapat terjadi karena terganggunya metabolisme sel yang menyebabkan sel bakteri tidak mampu bereplikasi atau hilangnya kemampuan membelah diri. Efek radiasi terhadap molekul-molekul penting, sel ataupun jaringan telah menimbulkan berbagai macam perubahan, gangguan ataupun kerusakan pada sistem biologi, seperti molekul protein, kromosom, molekul DNA, molekul enzim, lemak dan karbohidrat (Darusalam, 1996).
Berdasarkan penelitian yang dilakukan Trampuz, et al. (2006), disebutkan bahwa pada fase stasioner bakteri E. coli dapat diinaktivasi hingga dosis 4000 Gy (laju dosis 9,35 Gy/menit). Pada dosis tersebut sel bakteri E. coli telah mengalami kerusakan yang cukup parah, sehingga sel tidak dapat melakukan metabolisme, dan mengalami kematian. Dengan demikian, dosis tersebut dapat digunakan untuk bahan vaksin mastitis.
Menurut Alatas (2005), kerusakan yang terjadi pada DNA dan kromosom dapat menyebabkan sel tetap hidup atau mati. Bila tingkat kerusakan yang dialami sel tidak terlalu parah dan proses perbaikan berlangsung dengan baik dan tepat, maka sel bisa kembali normal seperti sebelum terpajan radiasi. Bila proses perbaikan berlangsung tetapi tidak tepat maka akan dihasilkan sel yang tetap dapat hidup tetapi telah mengalami perubahan. Artinya sel tersebut tidak lagi seperti sel semula, tetapi sudah menjadi sel yang baru atau abnormal tetapi hidup. Selain itu, bila tingkat kerusakan yang dialami sel sangat parah dan bila proses perbaikan tidak berlangsung dengan baik maka sel akan mati.
4.3Konsentrasi Protein Sel E. coli Hasil Iradiasi
0
Ekstaselular 104 103 126 134 135 113 87.9 91.8 113
Intraselular 154 178 182 202 190 177 165 169 170
Total 257 281 308 336 325 290 253 261 283 0 50 200 500 600 700 800 900 1000
Gambar 19. Konsentrasi protein E. coli hasil iradiasi
Pada dosis 0 Gy (kontrol). Jumlah protein intraselular, yaitu 154 mg/ml dan protein ekstraselular, yaitu 104 mg/ml. Kandungan protein total cenderung meningkat dibandingkan kontrol, tetapi pada dosis 800 Gy mengalami penurunan, yaitu 253 mg/ml. Perubahan konsentrasi protein total tertinggi terjadi pada dosis 500 Gy, yaitu 336 mg/ml kemudian meningkat lagi pada dosis iradiasi 900Gy dan 1000 Gy, yaitu 261 mg/ml dan 283 mg/ml.
iradiasi. Konsentrasi masing-masing protein dapat meningkat melalui sintesis atau melalui pelepasan selular yang nyata akibat kerusakan jaringan sel.
Wahyudi (2005), menambahkan bahwa adanya kemungkinan kecepatan peningkatan aktifitas pembelahan sel terjadi pada dosis tertentu, tidak selalu mengikuti interval peningkatan dosisnya. Seperti halnya sel kanker yang disebabkan oleh radiasi atau radikal bebas, sehingga protein bersifat karsinogenik. Harliansyah (2005), juga menyatakan jika radikal bebas menyerang asam-asam nukleat, akan menimbulkan gangguan terhadap molekul DNA yang berakibat terbentuknya mutasi basa-basa nitrogen serta berakhir dengan pembentukan karsinogenesis.
4.4Karakteristik Profil Protein Bakteri E. coli
Gambar 20. Profil Protein bakteri E. coli S1 hasil iradiasi sinar gamma (elektroforesis SDS-PAGE). M (marker/standar protein), 1 (0 Gy sebagai kontrol), 2 (50 Gy), 3 (200 Gy), 4 (500 Gy), 5 (600 Gy), 6 (700 Gy), 7 (800 Gy), 8 (900Gy), dan 9 (1000 Gy).
Intensitas protein yang berbeda, kemungkinan disebabkan karena jumlah protein yang dielektroforesis relatif rendah. Hal ini sesuai dengan hasil penentuan kadar protein yang telah ditentukan sebelumnya.
Hasil analisa anova satu arah (analysis of variance) menunjukkan nilai signifikansi sebesar 0,996 atau 99,6% yang berarti lebih besar dari 5% data menunjukkan tidak adanya perbedaan yang signifikan (H0 ditolak). Hal ini
memperkuat hasil konsentrasi protein. Dengan demikian iradiasi sinar gamma sebagai metode untuk inaktivasi berpotensi besar dalam pembuatan vaksin, karena komposisi protein relatif tetap walaupun konsentrasinya bervariasi dengan bertambahnya dosis iradiasi.
Protein yang biasanya mengalami gangguan dan kerusakan akibat denaturasi, yaitu pada struktur sekunder dan tersier protein. Hal ini diketahui sejak denaturasi
tidak cukup kuat untuk memutuskan ikatan peptida, dimana struktur primer protein tetap sama setelah proses denaturasi (Ophart, C.E., 2003). Dalam penelitian Vuckovic, M., et. al (2005), menunjukkan bahwa pada dosis iradiasi 1,5 kGy profil protein tidak mengalami perubahan yang signifikan.
Menurut Winarno (2002), protein yang terdenaturasi mengalami dua kemungkinan, yaitu pembukaan rantai peptida dan pemecahan protein menjadi unit yang lebih kecil. Terjadinya kedua jenis denaturasi ini tergantung pada keadaan molekul, yang pertama terjadi pada rantai polipeptida intramolekuler, sedangkan yang kedua terjadi pada bagian-bagian molekul yang bergabung dalam bentuk protein oligomer. Ikatan-ikatan yang dipengaruhi oleh proses denaturasi ini adalah: ikatan hidrogen, ikatan hidrofobik misalnya pada leusin, valin, fenilalanin, triptofan yang saling berlekatan membentuk suatu micelle dan tidak larut dalam air, ikatan ionik antara gugus bermuatan positif dan negatif, dan ikatan intramolekular seperti yang terdapat pada gugus disulfida dalam sistin (Winarno, 2002).
Selain terjadinya denaturasi protein, iradiasi juga mampu mendegradasi protein. Degradasi protein dapat menyebabkan protein tersebut kehilangan fungsi dan aktivitas biologisnya, degradasi struktur dapat berasal dari hilangnya gugus samping seperti deaminasi asam amino atau dari rearrangement kimia dan modifikasi biomolekul. Dosis 0 – 1000 Gy tidak cukup untuk memutuskan ikatan peptida (Syaifudin, 2005).
BAB V
KESIMPULAN DAN SARAN
5.1Kesimpulan
Dari hasil penelitian yang telah dilakukan dapat disimpulkan bahwa:
1. Dosis iradiasi untuk menginaktivasi E. coli S1 adalah 800 Gy -1000 Gy dengan laju dosis 1089,59 Gy/Jam.
2. Iradiasi sinar gamma terhadap bakteri E. coli S1 hingga dosis 1000 Gy mampu menghasilkan sel E. coli S1 inaktif dengan profil protein total yang tidak mengalami perubahan yang signifikan (Sign > 5%). Dengan nilai signifikan 0,996.
5.2Saran
DAFTAR PUSTAKA
Alatas, Zubaidah, M.Sc. 2005. Efek Paparan Radiasi Pada Manusia. Pusat Penelitian dan Pengembangan Keselamatan Radiasi dan Biomedika Nuklir. Badan Tenaga Nuklir Nasional
Arifin, M. 2006. Pengaruh Vaksin Radiasi Fascioliosis terhadap Ternak Ruminansia. Jurnal Gakuryoku Vol. II. Bogor
Benneth, C, et. Al,. 2002. Comparison of gamma-irradiated and triazol-treated RNA viruses using the joint biological agent identification and diagnostic, Idaho Technology Inc. Salt Lake City. UT
Bjerbakke, E. 1970. Manual on Radiation Dosimetry. Marcel Dekker Inc. pp. 323. New York
Darussalam, M. 1996. Radiasi dan Radioisotop Prinsip Kegunaannya Dalam Biologi, Kedokteran, dan Pertanian. Bandung: TARSITO
Feng P, et. al,. (2002-09-01). Enumeration of Escherichia coli and the Coliform
Bacteria Bacteriological Analytical Manual (8th ed.). FDA/Center for
Food Safety & Applied Nutrition. Retrieved on 2007-01-25.
Fuad, Asep A. 2006. Manajemen Kesehatan Pemerahan. BPPT Sapi Perah Bumikasih
Gaffar, A. 2006. Parasitology : Blood and Tissue, Download from:
http://www.med.sc.edu:85/parasitology/blood-proto.htm, University of
South California,
Government of India Department of Atomic Energy (GIDAE). 1992. Radiation Safety Aspects of High Intensity Irradiators. Bhabha Atomic Research Center Bombay 400 085
Hall, E.J. 1994. Radiobiology for the radiobiologist, Lippincott Williams and Walkin. Philadelphia
Hames. B.D. 1998. Gel Electrophoresis of Proteins. Oxford University Press. New York Tokyo
Harliansyah. 2005. Mengunyah Halia Menyah Penyakit. Jabatan Biokimia, Fakulti Perubatan, Universiti Kebangsaan Malaysia, 43600 UKM Bangi, Selangor Darul Ehsan, Malaysia. pp 92-97
Hermanto, S. 2003. Spesifitas dan Sensitifitas Antibodi Anti eRF3 Ragi Saccharomyces cerevisiae. ITB
Hilmy, N. 2001. The Application of Nuclear Science and Technology for Human Welfare. Proc.of The Public. Informa. Seminar Jointly Organized by IAEA and BATAN. Jakarta
IAEA. 1977. Manual of Food Irradiation Dosimetry (Technical Report Series 178). IAEA, Vienna
IAEA. 2000. Combating infection in developing countries. Vienna Austria
Jenkins, M.C. 2002. Advances and prospects for subunit vaccines againsts protozoa of veterinary importance. Veterinary Parasitology 101, Elsevier, pp 291-310
Juwono dan Zulfa J. Ahmad. 2000. Biologi Sel. Buku Kedokteran EGC. Jakarta
Kogan, GD, et. al.1996. Structural and immunochemical Characterization of Type VIII group B Streptococcus Capsular Polysacharide. J. Bio. Chem.271: 8786-8790
Leammli, U.K. 1970. Nature 227: 680-685
Lehtolainen, Tanja. 2004. Escherchia coli Mastitis Bacterial factors and host response. Department of Clinical Veterinary Sciences Faculty of Veterinary Medicine University of Helsinki. Finland
Machi,S. 2002. Nuclear techniques serve mankind. Japan Atomic Industrial Forum (JAIF),Inc.
Manglayang Farm Online. 2006. Diare Pada Sapi Pedet. Artikel. Ditemukan di
http://manglayang.blogsome.com/2006/04/06/kct-diare-pada-sapi-pedet/trackback/.
Medical Research News. 2006. Using gamma radiation preserves T-cell
responses in bacterial vaccine. http://www.news-medical.net/?id=19078. University of California, San Diego (UCSD) School of Medicine. Ditemukan pada tanggal 26 juli 2006, di http://www.ucsd.edu
McLaughlin, W.L., et. al. 1989. Dosimetry for Radiation Processing. Taylor & Francis, London
Muslim LW. 1995. Mikrobiologi Lingkungan. Universitas Hasanuddin. Jakarta
Nature publishing group. 2005. Nature reviews/immunology
Ophart, C. E. 2003. Virtual Chembook. Elmhurst College.
Poedjiadi A. 1994. Dasar-dasar Biokimia. Jakarta: UI-Press
Pelchar, Michael J., Jr., dan Chan E.C.S. 1988. Dasar-Dasar Mikrobiologi. Jilid 1 dan jilid 2. Jakarta: UI-Press
Ramamoorthy,S, et. al. 2006. Vaccination with gamma-irradiated Neospora caninum tachyzoites protects mice against acute challenge with N.caninum, J. Eukaryot. Microbiol. 53(2). pp. 151-156
Sanakkayala, N, et. al. 2005. Induction of antigen-specific Th1-type Immune Responses by Gamma-Irradiated recombinant Brucella abortus RB51., Clinical and diagnostic laboratory immunology, American Society for Microbiology
Schroeder, J.W. 1997. Mastitis Control Program : Bovine Mastitis and Milking Management. AS-129. North Dakota State University
Scorvia, Imelda. 2000. How Vaccines Work. Ditemukan di
http://www.mayoclinic.com/invoke.cfm?id=ID00023
Scrad – AAMP. 1971. Phys. Med. Biol. 16, pp. 379 – 396
Sugiri, Nawangsari. 1992. Biologi Sel. Departemen Pendidikan dan Kebudayaan Direktorat Jendral Pendidikan Tinggi Pusat antar Universitas Ilmu Hayat Institut Pertanian Bogor. Bogor
Sugiwaka, Teruo. 2005. Japan International Cooperation Agency
Sugoro, I. 2004. Pengontrolan Penyakit Mastitis dan Manajemen Pemerahan Susu. Artikel PATIR BATAN
Sumarsih, I. 2003. Diktat Kuliah Mikrobiologi Dasar. Jurusan Ilmu Tanah, Fakultas Pertanian UPN Veteran. Yogyakarta
Smith, N.C. 1992. Concepts and strategies for anti-parasite immunoprophylaxis and therapy, Int. J. For Parasite 22. 1047
Spink, J.W.T. dan Woods, R.J. 1976. An Introduction to Radiation Chemistry. John Willey and Sons Inc. New York
Syaifudin, M. 2005. Indikator Biokimia Sel terhadap Radiasi Pengion. Buletin ALARA. Vol. 6. No. 3. pp. 125-131
Tetriana, D dan Sugoro, I. 2007. Aplikasi Teknik Nuklir Dalam Bidang Vaksin. Jurnal Alara Vol. I. PTKMR – BATAN, PATIR – BATAN Pasar Jum’at. Jakarta