HUBUNGAN KADAR UBIQUTIN C-TERMINAL
HYDROLASE – L1 DENGAN TINGKAT KEPARAHAN
CEDERA KEPALA
TESIS
ANDRE MAROLOP PANGIHUTAN SIAHAAN
NIM. 097116006
PROGRAM MAGISTER KEDOKTERAN KLINIK
DEPARTEMEN ILMU BEDAH SARAF
FAKULTAS KEDOKTERAN
UNIVERSITAS SUMATERA UTARA
HUBUNGAN KADAR UBIQUITIN TERMINAL HYDROLASE – L1 DENGAN TINGKAT KEPARAHAN CEDERA KEPALA
TESIS
Diajukan sebagai salah satu syarat untuk memperoleh Gelar Magister Kedokteran Klinik dalam Program Studi Ilmu Bedah Saraf pada Fakultas Kedokteran
Universitas Sumatera Utara
ANDRE MAROLOP PANGIHUTAN SIAHAAN NIM. 097116006
PROGRAM MAGISTER KEDOKTERAN KLINIK DEPARTEMEN ILMU BEDAH SARAF
FAKULTAS KEDOKTERAN UNIVERSITAS SUMATERA UTARA
LEMBAR PENGESAHAN
Judul Tesis : HUBUNGAN KADAR UBIQUITIN C TERMINAL
HYDROLASE L1 DENGAN TINGKAT
KEPARAHAN CEDERA KEPALA
Nama : ANDRE MAROLOP PANGIHUTAN SIAHAAN
NIM : 097116006
Program Studi : ILMU BEDAH SARAF
Menyetujui,
Pembimbing I Pembimbing II
Prof. Dr. dr. Iskandar J, SpBS (K)
NIP. 194903311977111001 NIP. 194607181977031000
Prof. dr. Adril A. Hakim, SpS, SpBS(K)
Mengetahui / Mengesahkan
Ketua Departemen Ketua Program Studi
Prof. Dr. dr. Iskandar J, SpBS (K)
NIP. 194903311977111001 NIP. 194405071977031001
HALAMAN PERNYATAAN ORISINALITAS
Tesis ini adalah hasil karya penulis sendiri
dan semua sumber yang dikutip maupun dirujuk
telah penulis nyatakan benar
Nama : Andre Marolop Pangihutan Siahaan
NIM : 097116006
HALAMAN PERNYATAAN PERSETUJ UAN PUBLIKASI KARYA
ILMIAH
UNTUK KEPENTINGAN AKADEMIS
Sebagai sivitas akademika Universitas Sumatera Utara, saya yang bertanda
tangan di bawah ini
Nama : Andre Marolop Pangihutan Siahaan
NIM : 097116006
Program Studi : Magister Kedokteran Klinik
Konsentrasi : Ilmu Bedah Saraf
Jenis Karya : Tesis
demi pengembangan ilmu pengetahuan, menyetujui untuk memberikan kepada
Universitas Sumatera Utara Hak Bebas Royalti Non-eksklusif (Non-exclusive
Royalty Free Right) atas tesis saya yang berjudul:
HUBUNGAN KADAR UBIQUITIN C-TERMINAL HYDROLASE –
L1 DENGAN TINGKAT KEPARAHAN CEDERA KEPALA
beserta perangkat yang ada (jika diperlukan). Dengan Hak Bebas Royalti
Non-eksklusif ini, Universitas Sumatera Utara berhak menyimpan, mengalih media,
mengelola dalam bentuk database, merawat, dan mempublikasikan tesis saya
tanpa meminta izin dari saya selama tetap mencantumkan nama saya sebagai
penulis dan pemilik hak cipta.
Demikian pernyataan ini saya buat dengan sebenarnya
Dibuat di : Medan
Pada tanggal : 27 Juli 2013
Yang menyatakan
ABSTRAK
LATAR BELAKANG
Salah satu permasalahan utama dalam cedera kepala adalah bagaimana menilai tingkat keparahan cedera kepala.Modalitas yang banyak digunakan adalah pemeriksaan fisik dan radiologi.Saat cedera kepala, penanda cedera saraf yang dilepaskan dari otak menuju serum mungkin memberikan tambahan diagnosis yang bermakna.
TUJUAN
Untuk mengetahui kadar Ubiquitin C-terminal Hydrolase – L1 (UCH-L1), suatu penanda kerusakan neuron, pada serum penderita cedera kepala dan untuk mengetahui hubungannya dengan karakteristik serta hasil akhir klinis.
METODE
Ini merupakan studi potong lintang yang melibatkan delapan puluh orang penderita cedera kepala, baik ringan, sedang, maupun berat. Dengan menggunakan metode ELISA, dilakukan pengukuran kadar UCH-L1 serum.
HASIL
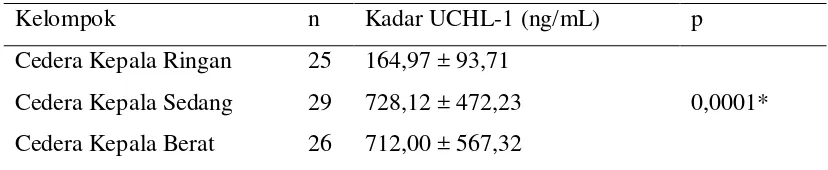
Kadar UCH-L1 serum penderita cedera kepala sedang dan berat lebih tinggi dibandingkan penderita cedera kepala ringan (p < 0,001), tetapi tidak ditemukan perbedaan bermakna antara penderita cedera kepala sedang dan cedera kepala berat. Tidak ditemukan korelasi yang signifikan antara kadar UCH-L1 serum dengan lama rawatan dan hasil akhir klinis pasien.
KESIMPULAN
Kadar UCH-L1 serum memiliki potensi yang besar sebagai tambahan modalitas diagnosis cedera kepala, tetapi tidak terbukti berhubungan dengan hasil akhir klinis pasien
ABSTRACT
BACKGROUND
One of the main problems in head injury is assessing the severity. The main modalities commonly used are physical examination and imaging; both have limitation in assessing the severity. Neuronal damage markers released in blood may provide valuable information about diagnosis the traumatic brain injury (TBI).
OBJECTIVE
To analyze the concentrations of ubiquitin C-terminal hydrolase-L1 (UCH-L1), a neuronal injury biomarker, in serum of TBI patients and their association with clinical characteristics and outcome.
METHODS
There are 80 TBI subjects, i.e. mild, moderate, and severe involved in this case-control study. Using ELISA, we studied the profile of serum UCH-L1 levels for TBI patients RESULT
Serum UCH-L1 level in moderate and severe head injury is higher than in mild head injury (p< .001), but we don’t find significant difference between moderate and severe head injury patients. There is no significant correlation found between serum UCH-L1 level and outcome.
CONCLUSION
Serum levels of UCH-L1 appear to have potential clinical utility in diagnosing TBI, but do not have correlation with outcome.
KATA PENGANTAR
Puji dan syukur penulis panjatkan kehadirat Tuhan Yang Maha Kuasa yang telah
memberikan rahmat dan karuniaNya, sehingga penulis dapat menyelesaikan tesis
ini.Penulis sangat menyadari bahwa tanpa bantuan dari semua pihak, tesis ini
tidak mungkin dapat penulis selesaikan.Oleh karena itu perkenankanlah penulis
mengucapkan terima kasih serta penghargaan yang setinggi-tingginya kepada
semua pihak yang telah membantu penulis dalam menyelesaikan tesis ini, baik
secara langsung maupun tidak langsung.
Rasa hormat, penghargaan dan ucapan terima kasih sebesar - besarnya penulis
sampaikan kepada:
1. Prof. Dr. dr. Iskandar Japardi, SpBS (K) selaku Ketua Departemen Ilmu
Bedah Saraf FK USU yang telah memberikan kesempatan pada penulis
untuk mengikuti pendidikan serta senantiasa membimbing, memberi
dorongan dan kemudahan selama penulis menjalani pendidikan.
2. Prof. dr. Abdul Gofar Sastrodiningrat, SpBS (K) selaku Ketua Program
Studi Ilmu Bedah Saraf FK USU yang telah dengan sungguh - sungguh
membantu, membimbing, dan memberi dorongan kepada penulis,
3. Pembimbing penulisan tesis ini, Prof. Dr. dr. Iskandar Japardi, SpBS (K)
dan Prof. dr. Adril A Hakim, SpS, SpBS (K) yang telah memberikan
bimbingan, bantuan serta saran - saran yang sangat berharga dalam
pelaksanaan penelitian dan penyelesaian tesis ini
4. Staf Departemen Ilmu Bedah Saraf FK USU/ RSUPHAM, Dr. dr. Rr.
Suzy Indharty, M.Kes, SpBS dan Dr. dr. Ridha Dharmajaya, SpBS yang
telah banyak memberikan bantuan kepada penulis selama penulisan tesis
ini.
5. Dr. dr. Arlinda Sari Wahyuni, M. Kes sebagai pembimbing statistik dalam
penulisan tesis ini
6. Seluruh staf Laboratorium Patologi Klinik RSUP HAM yang telah
7. Teman-teman PPDS Departemen Ilmu Bedah Saraf FK USU
8. Perawat di ruang rawat inap RA-4 Bedah Saraf dan Unit Perawatan
Intensif RSUP HAM
9. Serta seluruh pihak yang tidak mungkin saya sebutkan satu per satu.
Rasa terima kasih yang sebesar-besarnya saya haturkan kepada orang tua saya, Ir.
Sahat M. Siahaan dan Dr. dr. Sarma L. Raja, SpOG (K) atas doa, pengorbanan,
nasihat, dan dukungan kepada saya selama ini. Kepada kakak saya, dr. Hatsari M.
Siahaan, SpOG, terima kasih atas dukungannya selama ini, serta kepada sahabat
saya, Aries Akira Pulungan, saya ucapkan terima kasih sebesar-besarnya atas
segala bantuan yang telah diberikan selama pendidikan ini.
Semoga tulisan ini berguna untuk kita semua.
DAFTAR ISI
DAFTAR ISI i
DAFTAR TABEL v
DAFTAR GAMBAR vi
DAFTAR SINGKATAN vii
ABSTRAK viii
ABSTRACT ix
BAB 1 PENDAHULUAN 1
A. Latar Belakang Penelitian 1
a. Epidemiologi Cedera Kepala 1
b. Mengapa Biomarker Cedera Saraf Diperlukan 3
B. Rumusan Masalah 6
C. Pertanyaan Penelitian 7
D. Tujuan Penelitian 7
a. Tujuan Umum 7
b. Tujuan Khusus 7
E. Manfaat Penelitian 8
BAB 2 TINJAUAN PUSTAKA 9
A. Cedera Kepala 9
a. Neuropatologi 9
b. Degenerasi Akson post trauma 11
i. Konsep mekanis 12
ii. Degenerasi Akson akut 12
iii. Degenerasi Wallerian 14
iv. Mekanisme molekuler degenerasi akson 17
v. Kalsium 18
vi. Kejadian lanjutan setelah influks kalsium 20
vii. Cedera mitokondria 22
viii. Agregasi 22
ix. Transport Akson 24
x. Aktivasi kinase 26
xi. Autofagi dan sistem ubiquitin protease 27
B. Sistem Ubiquitin Proteasome 27
a. Enzim Deubiquitinating 30
b. Fungsi sistem ubiquitin proteasome pada manusia 31
c. Penyakit neurodegeneratif dan SUP 33
d. Ubiquitin Carboxyl-Terminal Hydrolase (UCH-L1) 35
dan hubungannya dengan degenerasi saraf
C. Biomarker pada cedera kepala 38
BAB 3 KERANGKA TEORI, KERANGKA KONSEP, 43
HIPOTESIS PENELITIAN
A. Kerangka teori 43
B. Kerangka konsep 43
BAB 4 METODOLOGI PENELITIAN 45
A. Rancangan penelitian 45
B. Waktu dan tempat penelitian 45
C. Populasi dan sampel penelitian 45
D. Kriteria sampel penelitian 46
a. Kriteria inklusi 46
b. Kriteria eksklusi 46
E. Besar sampel penelitian 47
F. Alur penelitian 48
G. Organisasi Penelitian 49
H. Cara kerja 49
I. Batasan operasional variabel 58
J. Kontrol Kualitas 59
K. Analisis Data 59
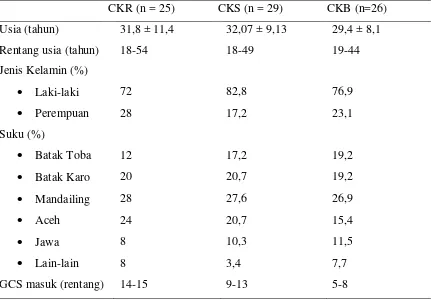
BAB 5 HASIL PENELITIAN 60
A. Kadar UCH-L1 Serum 61
a. Analisis berdasarkan jenis kelamin 61
b. Analisis berdasarkan suku 62
c. Analisis berdasarkan kelompok cedera 63
d. Analisis kadar UCH-L1 pada penderita cedera kepala 64
yang meninggal dalam tiga hari pertama
B. Korelasi kadar UCH-L1 dengan GCS saat masuk dan 66
BAB 6 PEMBAHASAN 67
BAB 7 KESIMPULAN DAN SARAN 72
A. Kesimpulan 72
B. Saran 73
DAFTAR TABEL
Tabel 1 Penanda cedera kepala 34
Tabel 2 Indikasi operasi pada Perdarahan Intrakranial 44
Tabel 3 Glasgow Outcome Scale 45
Tabel 4 Data demografi subjek penelitian 52
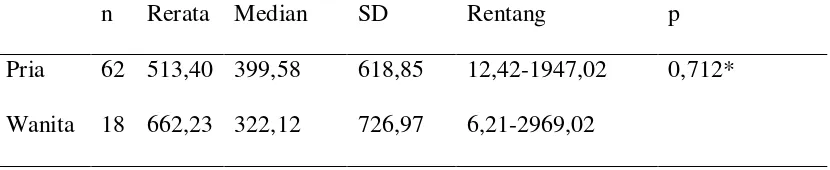
Tabel 5 Kadar UCHL-1 pria dan wanita 54
Tabel 6 Kadar UCHL-1 kelima suku terbesar yang ikut 55
dalam penelitian
Tabel 7 Kadar UCHL-1 ketiga kelompok 56
Tabel 8 Kadar UCHL-1 kelompok mortalitas tiga hari 57
pertama
Tabel 9 Korelasi kadar UCHL-1 dengan GCS saat masuk 58
DAFTAR GAMBAR
Gambar 1 Beberapa mekanisme molekul yang terlibat 14
dalam degenerasi akson
Gambar 2 Terbentuknya biomarker setelah trauma 31
dan penyebarannya pada darah
Gambar 3 Proses cedera kepala dengan protein yang 32
potensial menjadi biomarker
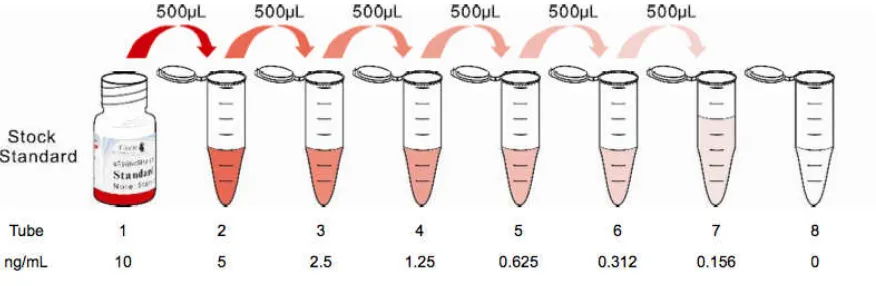
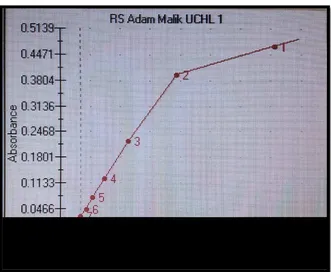
Gambar 4 Dilusi bertahap dalam pembuatan standar 55
Gambar 5 Kurva standar acuan penghitungan konsentrasi 57
UCHL-1 pada penelitian ini
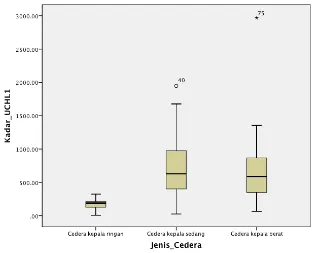
Gambar 5.2 Kadar UCHL-1 serum penderita CKR, CKS, dan 63
CKB
Gambar 5.3 Kadar UCHL-1 serum subjek penelitian yang 65
meninggal dalam tiga hari pertama dan yang
DAFTAR SINGKATAN
CKB Cedera Kepala Berat
CKR Cedera Kepala Ringan
CKS Cedera Kepala Sedang
CSF Cerebrospinal Fluid
CT Computed Tomography
CDC Centers for Disease Control and Prevention
DUB Deubiquitinating enzyme
GFAP Glial Fibrillary Acidic Protein
GCS Glasgow Coma Scale
ICP Intracranial Pressure
IL Interleukin
ISS Injury Severity Score
MAP Mean Arterial Pressure
SUP Sistem Ubiquitin Protease
ABSTRAK
LATAR BELAKANG
Salah satu permasalahan utama dalam cedera kepala adalah bagaimana menilai tingkat keparahan cedera kepala.Modalitas yang banyak digunakan adalah pemeriksaan fisik dan radiologi.Saat cedera kepala, penanda cedera saraf yang dilepaskan dari otak menuju serum mungkin memberikan tambahan diagnosis yang bermakna.
TUJUAN
Untuk mengetahui kadar Ubiquitin C-terminal Hydrolase – L1 (UCH-L1), suatu penanda kerusakan neuron, pada serum penderita cedera kepala dan untuk mengetahui hubungannya dengan karakteristik serta hasil akhir klinis.
METODE
Ini merupakan studi potong lintang yang melibatkan delapan puluh orang penderita cedera kepala, baik ringan, sedang, maupun berat. Dengan menggunakan metode ELISA, dilakukan pengukuran kadar UCH-L1 serum.
HASIL
Kadar UCH-L1 serum penderita cedera kepala sedang dan berat lebih tinggi dibandingkan penderita cedera kepala ringan (p < 0,001), tetapi tidak ditemukan perbedaan bermakna antara penderita cedera kepala sedang dan cedera kepala berat. Tidak ditemukan korelasi yang signifikan antara kadar UCH-L1 serum dengan lama rawatan dan hasil akhir klinis pasien.
KESIMPULAN
Kadar UCH-L1 serum memiliki potensi yang besar sebagai tambahan modalitas diagnosis cedera kepala, tetapi tidak terbukti berhubungan dengan hasil akhir klinis pasien
ABSTRACT
BACKGROUND
One of the main problems in head injury is assessing the severity. The main modalities commonly used are physical examination and imaging; both have limitation in assessing the severity. Neuronal damage markers released in blood may provide valuable information about diagnosis the traumatic brain injury (TBI).
OBJECTIVE
To analyze the concentrations of ubiquitin C-terminal hydrolase-L1 (UCH-L1), a neuronal injury biomarker, in serum of TBI patients and their association with clinical characteristics and outcome.
METHODS
There are 80 TBI subjects, i.e. mild, moderate, and severe involved in this case-control study. Using ELISA, we studied the profile of serum UCH-L1 levels for TBI patients RESULT
Serum UCH-L1 level in moderate and severe head injury is higher than in mild head injury (p< .001), but we don’t find significant difference between moderate and severe head injury patients. There is no significant correlation found between serum UCH-L1 level and outcome.
CONCLUSION
Serum levels of UCH-L1 appear to have potential clinical utility in diagnosing TBI, but do not have correlation with outcome.
BAB I PENDAHULUAN
A.Latar Belakang Penelitian 1. Epidemiologi Cedera Kepala
Cedera kepala merupakan salah satu permasalahan kesehatan utama di
dunia (Hyder et al., 2007; Reilly, 2007). Di Amerika Serikat, diperkirakan 1,7 juta
orang menderita cedera kepala setiap tahunnya. 275.000 orang akan dirawat di
rumah sakit dan 52.000 orang di antaranya meninggal (CDC, 2010). Hal yang
sama juga ditemui di Inggris. Setiap tahunnya ada sekitar 700.000 kunjungan ke
unit gawat darurat akibat cedera kepala (Goodacre, 2008).Dampak cedera kepala
terhadap ekonomi cukup besar.Pada tahun 2000, diperkirakan biaya yang
diperlukan dalam penatalaksanaan cedera kepala, mencakup biaya rehabilitasi dan
kerugian akibat disabilitas, cukup besar yaitu sekitar 60 miliar dollar (Finkelstein,
2006). Selain biaya ekonomi, CDC memerkirakan setidaknya lima juta penduduk
Amerika memerlukan bantuan jangka waktu panjang untuk dapat melakukan
aktivitas sehari-hari terkait efek cedera kepala yang berkepanjangan (Thurman et
al., 1999). Hal yang lebih mengkhawatirkan adalah kenyataan bahwa insiden
cedera kepala global meningkat dalam dua dekade terakhir, kemungkinan terjadi
akibat peningkatan penggunaan kendaraan bermotor di negara berkembang (Lin et
al., 2008; Bener et al., 2010).CDC melaporkan bahwa kecelakaan bermotor
merupakan penyebab cedera kepala terbesar kedua, setelah mekanisme jatuh
Sampai saat ini, pilihan terapi penderita cedera kepala pada seluruh
kelompok umur, baik pada fase akut maupun kronis, sangat terbatas (Reinert dan
Bullock, 1999; Menon, 2009).Hal ini memacu penelitian mengenai biomekanika
cedera otak, dalam hal pencegahan terjadinya cedera, seperti helm dan sabuk
pengaman (Levy et al., 2004; Nirula et al., 2004) dan pengembangan standar
keamanan yang sesuai (Zhang et al., 2004).
2. Mengapa Biomarker Cedera Kepala Diperlukan
Permasalahan utama yang ada saat ini adalah kesulitan dalam
mendiagnosis dan memprediksihasil akhir setelah cedera kepala akut berdasarkan
keadaan klinis dan radiologi (Zink, 2001).Pemeriksaan klinis dapat dipengaruhi
oleh terapi yang diberikan, intoksikasi alkohol, atau cedera multipel.Terapi
penderita cedera kepala yang disertai kebingungan, agitasi, atau kejang dapat
memengaruhi informasi yang didapat dari pemeriksaan klinis (Mirski et al.,
1995). Banyak penderita cedera kepala ringan (GCS 14-15) juga disertai
intoksikasi alkohol yang nantinya akan memengaruhi pemeriksaan neuropsikologi
(Kelly, 1995). Cedera kepala juga sering luput dari diagnosis pada penderita
trauma multipel (Buduhan dan McRitchie, 2000).
Radiologi dapat menyediakan tambahan informasi, tetapi kemampuan
teknik ini terbatas pada sensitivitasnya dan biaya.Walaupun cedera kepala ringan
dapat menyebabkan disabilitas dalam jangka waktu panjang, gambaran patologi
otak kemungkinan besar hanya dapat dideteksi oleh magnetic resonance imaging
ada, namun kemampuannya mendeteksi cedera otak difus terbatas.Single photon
emission CT dapat mendeteksi perubahan aliran darah regional, tetapi tidak dapat
mendeteksi kerusakan struktural. Pada pasien dengan keadaan kritis, ketersediaan
dan waktu yang diperlukan untuk menghasilkan imejing menjadi keterbatasan
utama MRI dan single photon emission CT Scan. Selain itu, baik MRI dan CT
tidak dapat menjadi dasar untuk memprediksi hasil akhir (Hanlon et al., 1999).
Karena itu, diperlukan modalitas diagnosis untuk mendiagnosis tingkat
keparahan cedera kepala dan memprediksi hasil akhir.Salah satu modalitas yang
dapat digunakan adalah biomarker.Dalam dekade terakhir, biomarker cedera saraf
telah menarik perhatian banyak peneliti.Dengan menganalogikan biomarker pada
infark miokard akut, beberapa tinjauan pustaka telah menekankan pentingnya
informasi mengenai luasnya kerusakan jaringan otak (Konchanek et al., 2008;
Dash et al., 2010).
Biomarker yang telah diteliti dalam skala besar sebelumnya adalah
neuron-specific enolase (Ross et al., 1996),glial protein, S100β (Pelinka et al.,
2004) dan myelin basic protein (Yamazaki et al., 1995).S100β merupakan protein
pengikat kalsium yang diekspresikan oleh astrosit dan dilepaskan oleh sel yang
sekarat.Awalnya, S100β dianggap menjadi suatu biomarker cedera kepala yang
menjanjikan.Namun, S100β juga ditemui pada oligodendrosit, microglia, neuron,
dan jaringan ekstraserebral.Protein ini disekresikan secara aktif, tidak hanya oleh
sel yang sekarat (Donato, 1999).Neuron-specific enolase terdapat pada sitoplasma
neuron dan dilaporkan muncul di serum segera setelah kecelakaan (Wang et al.,
pada selubung myelin sel-sel susunan saraf pusat dan dilaporkan akan dilepaskan
ke serum setelah cedera otak atau penyakit demyelinating (Berger et al., 2007).
Walaupun sejumlah penelitian telah menggambarkan keuntungan penggunaan
biomarker untuk menggambarkan tingkat keparahan cedera, hasil penelitian yang
ada masih bertentangan, sehingga penggunaan rutin biomarker ini sulit dilakukan.
Akibat kemajuan di penelitian di bidang proteomik, para ahli berhasil
menemukan ubiquitin C-terminal hydrolase-L1 (UCHL-1), suatu enzim yang
spesifik dan terkonsentrasi dalam sel saraf, sebagai penanda cedera otak yang
potensial (Jackson dan Thompson, 1981). UCHL-1 terlibat dalam penambahan
atau pengurangan ubiquitin dari protein yang akan dimetabolisme melalui jalur
ATP-dependent proteasome (Tongaonkar et al., 2000). Varian UCHL-1 telah
dihubungkan dengan penyakit Parkinson familial (Lincoln et al., 1999).Pada
iskemia serebri dan cedera spinal cord, telah ditemukan adanya peningkatan
agregrat protein dan penurunan aktivitas proteasome (Hu et al., 2000; Keller et al.,
2000). Data ini menggambarkan bahwa UCHL-1 memegang peranan penting
dalam membuang protein yang berlebihan, teroksidasi, atau misfolded (Gong dan
Lenik, 2007).
B.Rumusan Masalah
Dengan modalitas diagnosis yang rutin digunakan saat ini, sulit untuk
menentukan derajat kerusakan otak dan hasil akhir penderita cedera
kepala.Diperlukan suatu biomarker derajat kerusakan otak yang dapat
Ubiquitin C-terminal hydrolase-L1 (UCHL-1) merupakan protein yang
spesifik terdapat pada neuron dan banyak terdapat pada neuron.Dahulu, protein ini
digunakan sebagai penanda neuron pada histopatologi karena spesifisitasnya.
Papa et al., (2010) melaporkan bahwa kadarUCHL-1 meningkat pada
Cerebrospinal Fluid penderita cedera kepala dengan hubungan yang signifikan
antara kadar UCHL-1 dengan keparahan cedera, antara lain ada tidaknya kelainan
pada CT Scan kepala, tingkat kesadaran berdasarkan GCS, dan mortalitas dalam
enam minggu pertama. Meskipun demikian, pengukuran biomarker pada CSF
memerlukan pemasangan kateter ventrikel terlebih dahulu.Tindakan ini sendiri
bersifat invasif, tidak praktis, dan tidak selalu dapat dilakukan (Berger et al.,
2002).
C.Pertanyaan Penelitian
Berdasarkan uraian di atas, diajukan pertanyaan penelitian “Apakah kadar
Ubiquitine C- Terminal Hydrolase L-1 (UCHL-1) serumberhubungan
dengantingkat keparahan cedera kepala?”
D.Tujuan Penelitian 1. Tujuan Umum
Untuk mengetahui hubungan kadarUCHL-1 serum dengan tingkat
2. Tujuan Khusus
a. Mengetahui hubungan kadar UCHL-1 serum dengan GCS awal setelah
resusitasi
b. Mengetahui hubungan kadar UCHL-1 serum dengan mortalitas pada
tiga hari pertama
c. Mengetahui hubungan kadar UCHL-1 dengan lama rawatan pasien
E. Manfaat Penelitian
1. Dari hasil penelitian sebelumnya, ditemukan bahwa kadarUCHL-1 akan
meningkat pada CSF penderita cedera kepala berat. Pada penelitian ini,
akan didapatkan kadar UCHL-1 serum, yang jauh lebih mudah untuk
dilakukan bila dibandingkan dengan mendapatkan CSF.
2. Dengan didapatkan kadar tertentu, klinisidapat melakukan tindakan
yang tepat dan sesuai serta dapat menghindarkan klinisi dari terapi yang
berlebihan (over treated) maupun terapi yang tidak optimal (under
treated).
3. Dengan penanganan yang tepat, dari segi ekomonis akan menurunkan
biaya yang dikeluarkan oleh masyarakat khususnya dan menurunkan
BAB II
TINJAUAN PUSTAKA
A. Cedera Kepala 1. Neuropatologi
Cedera kepala dapat ditandai dengan coup dan contra coup serta dengan
shearing dan tearing akson di otak akibat akselerasi rotasional dari
kepala.Diagnosis umum sehubungan cedera kepala meliputi fraktur tengkorak,
contusion, confusion, laserasi, dan lesi fokal (Silver et al., 2005). Cedera sekunder
yang meliputi perdarahan lebih lanjut, deformitas mekanis, dan peningkatan
tekananan intrakranial dapat terjadi sebagai komplikasi sebagai cedera awal
(Bigler & Maxwel., 2011). Saat ini telah terbukti bahwa penderita cedera kepala
mengalami kehilangan sebagian volume otak sampai setidaknya satu tahun setelah
kecelakaan (Bendlin et al., 2008; Sidaros et al., 2009; Trivedi et al., 2007).
Pemahaman patologi seluler pada cedera kepala merupakan hal penting
dalam pengembangan terapi baru cedera kepala.Perubahan neurokimiawi otak
sesudah cedera dapat berupa agen “neuroprotektif” dan
“autodestruksi”.Peningkatan acetylcholine terjadi segera setelah cedera (Donat et
al., 2008; Lyeth & Hayes, 1992).Peningkatan epinefrin dan norepinefrin pada
serum terjadi seiringan dengan tingkat keparahan cedera.Pada daerah sekitar
tempat cedera, ditemui peningkatan serotonin dan dopamin (Kobori et al.,
2011).Namun sampai sekarang bagaimana peranan agen neurokimiawi ini
minggu setelah cedera terjadi peningkatan faktor neurotropic, seperti nerve
growth factor (NGF) dan fibroblast growth factor (FGF). Faktor-faktor
neurotropik ini diduga akan membantu pemulihan (Ziebell & Morganti –
Kossmann, 2010).
Autofagi merupakan proses dimana otak membuang jaringan yang mati
atau rusak agar sel sehat dapat berfungsi dengan lebih efektif. Proses ini juga
diduga akan membantu pemulihan (Clark et al., 2008). Meskipun demikian,
proses ini dapat menyebabkan nekrosis dan apoptosis yang akan memicu
degenerasi Wallerian dalam jumlah yang tidak diketahui (Zhou et al., 2012).
Degenerasi Wallerian merupakan perluasan dari cedera kepala. Cedera yang
awalnya terjadi hanya pada akson akan meluas sampai ke badan sel (Kelley et al.,
2006). Proses ini sepertinya memegang peranan, baik dalam perubahan white
matter yang banyak dijumpai pada cedera kepala, maupun dalam degenerasi dan
reorganisasi. Proses perbaikan dan degenerasi yang berlangsung sekaligus ini
menyebabkan microenvironment otak yang sangat beragam.
2. Degenerasi Axon Post Trauma
Cedera kepala akan menghasilkan cedera akson, terutama akibat regangan
(Meythaler et al., 2001). Berbeda dengan cedera fokal, cedera akson pada cedera
kepala bersifat lebih difus.Transeksi komplit jarang terjadi, tetapi regangan
menyebabkan kerusakan struktur akson, yang dapat menyebabkan disfungsi sel
Permeabilitas membran akson akan segera terganggu, bahkan setelah
cedera kepala sedang dan ini akan disertai dengan pemadatan lokal dari
neurofilamen (Pettus et al., 1994). Pada model kucing percobaan, gangguan
permeabilitas lokai ini akan muncul dalam lima menit setelah trauma dan
pemadatan ini bertahan setidaknya selama enam jam setelah kejadian (Pettus &
Povlishock, 1996). Model trauma lain yang sering digunakan adalah regangan
pada neuron in vitro. Setelah regangan, terjadi distorsi berbentuk undulasi di
sepanjang akson yang terjadi akibat kerusakan mikrotubulus. Gangguan
mikrotubulus ini akan menyebabkan gangguan transport akson dan akumulasi
protein (Tang-Schomer et al., 2010).
3. Mekanisme Degenerasi Akson
a. Konsep Mekanis
Seperti apoptosis, kebanyakan bentuk degenerasi akson merupakan proses
self-destructing seluler yang aktif dan melibatkan kaskade tertentu dengan
keterlibatan banyak faktor (Raff et al., 2002). Meskipun demikian, pada dasarnya,
apoptosis dan degenerasi akson melibatkan proses biokimia yang berbeda dan
dapat terjadi secara terpisah (Whitmore et al., 2003). Beberapa bentuk degenerasi
akson telah dijelaskan berdasarkan lokasinya pada akson dan waktu
terjadinya.Penelitian yang paling banyak dilakukan adalah mengenai degenerasi
sesudah trauma, baik degenerasi akut pada tempat trauma maupun degenerasi
b. Degenerasi Akson Akut
Istilah degenerasi akson akut mengacu pada disintegrasi akut akson dalam
beberapa jam setelah trauma susunan saraf pusat. Dalam 10-30 menit setelah
trauma, akson akan relatif stabil dengan gambaran makroskopis yang normal.
Meskipun demikian, pada tingkat molekuler, suatu sinyal sudah diaktivasi dengan
hasil akhir berupa fragmentasi akson. Proses ini diawali dengan influks kalsium
ke dalam akson dalam waktu cepat yang menyebabkan peningkatan konsentrasi
kalsium yang transien dalam 40 detik setelah cedera. Pemberian calcium channel
inhibitor pada saat ini akan menghambat peningkatan kalsium aksonal dan
degenerasi akut (Knoferle et al., 2010). Influks kalsium menyebabkan aktivasi
calpain yang mencapai nilai maksimal dalam 30 menit setelah cedera.
Perubahan pertama struktur mikroskopis dapat dinilai dalam 30 menit
pertama setelah cedera.Perubahan ini meliputi kondensasi dan perubahan
alignment neurofilamen yang diikuti fragmentasi mikrotubulus (Knoferle et al.,
2010). Pada kelainan SSP lain, pemadatan neurofilamen fokal dan proteolisis
mikrotubulus terbukti berhubungan dengan aktivasi calpain (Veerana et al., 2004).
Karena itu, aktivasi calpain pada saat awal kemungkinan besar juga berperan
dalam degenerasi akson akut. Kerusakan sitoskeleton akan menyebabkan
gangguan transport akson.
Gambaran degenerasi akson akut lain adalah aktivasi autofagi lokal.
jam pertama setelah cedera. Inhibisi autofagi dengan obat-obatan seharusnya
mengurangi degenerasi akut, namun tidak seperti setelah pemberian calcium
channel blocker, pengurangan degenerasi tidak ditemui. Ini kemungkinan
menggambarkan bahwa autofagi merupakan proses lanjutan influks kalsium
(Knoferle et al., 2010).
Meskipun pemberian obat-obatan seharusnya akan mengurangi degenerasi
akson akut, efek jangka panjang dari terapi ini masih belum jelas. Karena
degenerasi akut hanya terjadi pada sekitar 400 μm akson di sekitar tempat cedera,
keuntungan menyelamatkan bagian ini tidak bermakna.
c. Degenerasi Wallerian
Degenerasi Wallerian secara sederhana didefenisikan sebagai degenerasi
akson yang terjadi distal dari tempat cedera. Setelah trauma, bentuk bagian akson
yang tidak terkena degenerasi akut akan tetap normal dalam 24 sampai 72 jam
pertama. Kemudian, bagian distal akson akan menjalani fragmentasi progresif
yang menyerupai fragmentasi pada degenerasi akson akut (Kerschensteiner et al.,
2005) yang pada akhirnya menyebabkan removal seluruh bagian distal akson.
Degenerasi Wallerian berlangsung dengan kecepatan mulai 0,4 mm/jam pada
penelitian in vitro (Sievers et al., 2003) sampai 24 mm/jam pada saraf sciatic tikus
(Beirowski et al., 2005).Pada susunan saraf tepi, arah degenerasi Wallerian pada
akson tampaknya bergantung jenis lesi. Transeksi komplit saraf menyebabkan
fragmentasi anterograd mulai proksimal ke distal, sementara crush injuryakan
al., 2005). Meskipun makrofag dan glia sepertinya memegang peranan penting,
mekanisme degenerasi Wallerian tampaknya bersifat intrinsik (MacDonald et al.,
2006).
Mekanisme molekuler yang mendasari degenerasi Wallerian belum
dipahami sepenuhnya.Kemajuan pesat terjadi dengan ditemukannya tikus mutan
WldS (Lunn et al., 1989). Pada tikus ini, potongan akson distal dari lesi bertahan
sepuluh kali lebih lama dibandingkan akson pada binatang wild-type, dan survival
badan sel tidak terganggu (Adalbert et al., 2006).
Protein mutan WldS merupakan produk gen yang terdiri dari potongan
faktor poliubiquiti UFD2a/UBE4b dan nicotinamide monocleotide
adenytransferase -1 (NMNAT1; Mack et al., 2001).NMNAT1 adalah protein
kunci jaras nicotinamide-adenine dinucleotide +(NAD+) pada mamalia.UBE4b
adalah ubiquitin ligase tipe E4 yang dapat menambahkan rantai multiubiquitin
pada jaras ubiquitin/proteasome (Hatakeyama et al., 2001).Tempat molekuler
WldS yang penting adalah tempat pengingkatan ATP dan tempat pengikatan
NMN+
Protein WldS berada terutama pada nukleus, meskipun juga dideteksi pada
aksoplasma dan organel aksoplasma (Yahata et al., 2009).Ekspresi NMNAT1 pada NMNAT1 dan tempat pengikatan valocin-containing protein (VCP)
pada UBE4b. NMNAT1 dan UBE4b yang fungsional diperlukan untuk aktivitas
neuroproteksi dari WldS. Ini didukung oleh fakta bahwa ekspresi NMNAT1 saja
tidak cukup untuk mencegah degenerasi akson pada neuron mammalia meskipun
penurunan aktivitas NMNAT1 pada tikus WldS transgrenik menyebaban
lokal pada kompartemen akson menyebabkan efek protektif yang menyerupai
tikus transgenik WldS (Babetto et al., 2010).Data ini menunjukkan bahwa
aktivitas protektif SldS dimediasi oleh transport protein terus-menerus pada
akson. Sesuai dengan penjelasan di atas, isoform NMNAT lain meningkatkan
survival akson lokal. NMNAT2 terus menerus ditranspor dari badan sel menuju
akson dengan waktu turnover yang singkat, sekitar 4 jam. Down regulation dari
NMNAT2 atau inhibisi transportnya menuju akson menyebabkan degenerasi
akson. Sebaliknya, overekspresi akan menghambat degenerasi (Gilley &
Coleman, 2010). Efek yang sama juga terlihat pada overekspresi isoform
mitokondria NMNAT3 (Yahata et al., 2009). Target isoform NMAT untuk
meningkatkan survival akson masih belum jelas. Seluruh NMNAT mempunyai
domain katalis sintesis NAD+ (Berger et al., 2005), meskipun data yang
mendukung peranan NAD+ dalam mempertahankan survival akson tidak
konsisten.Pemberian NAD+ konsentrasi tinggi pada ekstraseluler menyebabkan
perlindungan akson yang cedera.Sebaliknya, berbagai usaha untuk meningkatkan
konsentrasi NAD+ intrasel tidak memberikan efek pada degenerasi akson (Sasaki
et al., 2009).
d. Mekanisme Molekuler Degenerasi Akson
Degenerasi akson akut, degenerasi fokal akson, dan degenerasi Wallerian
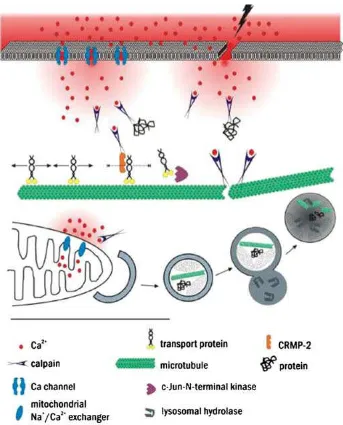
Gambar 1 Beberapa mekanisme molekul yang terlibat dalam degenerasi akson. (Lingor et al, 2012)
e. Kalsium
Beberapa proses yang menyebabkan peningkatan konsentrasi kalsium
akson pada berbagai lesi antara lain : (1) influks kalsium dari ruang ekstrasel
channel dari rongga ekstrasel, dan (3) lepasan kalsium dari depot kalsium intrasel
(Stirling & Stys, 2010).
Cedera mekanis akson menyebabkan kerusakan kontinuitas membrane dan
influks kalsum ekstrasel ke dalam sitoplasma.Sesuai percobaan Ziv dan Spira
(1995) pada akson in vitro, gelombang kalsium diinduksi oleh transeksi satu
cabang dendrit yang menyebar dengan cepat (dalam satuan detik) sampai
percabangan dendrit. Kalsium intra akson akan menurun dengan cepat beberapa
menit kemudian menuju tingkat tertentu setelah ujung akson yang cedera ditutup.
Meskipun demikian, kadar kalsium intrasel bervariasi, mulai >1 mM di dekat lesi
sampai beberapa ratus mikromolar distal dari lesi (Ziv & Spira, 1995). Pada
penelitian in vitro, diketahui bahwa diperlukan kadar kalsium ekstra akson > 200
μM untuk menyebabkan peningkatan kalsium intra akson setelah axotomy.
Kalsium juga terbukti memasuki akson melalui L-type calcium channel, bukan N
type (George et al., 1995).
Ruang ekstra akson bukanlah satu-satunya sumber kalsium.Depot kalsium
intrasel juga memberikan kontribusi yang cukup besar pada peningkatan kalsium
sitoplasimik.Misalnya, pada kerusakan iskemia di akson pada kolum dorsalis,
kalsium dilepaskan melalui reseptor ryanodine oleh reticulum endoplasma
(Ouardouz et al., 2003) atau mitokondria (Nikolaeva et al., 2005).
Pada akson dengan myelin, reseptor ryanodine dapat diaktivasi melalui
L-type calcium channel. Baik reseptor ryanodine dan L-type calcium channel berada
pada axolemma.Namun, penghambatan pelepasan kalsium intra akson, misalnya
menggambarkan kemungkinan sumber kalsium lain atau adanya suatu mekanisme
lain yang tidak tergantung kalsium (Ouardouz et al., 2003). Pada proses iskemia,
kebanyakan lepasan kalsium intraaksonal dimediasi oleh pompa Na+/Ca2+ yang
dibuktikan pada percobaan dengan CGP37157, suatu penghambat pompa
Na+/Ca2+
Selain itu, kalsium intrasel yang berlebihan pada neuron dapat berasal dari
pompa pada membrane sel, seperti plasma-membrane calcium ATPase isoform 2.
Penurunan kadar ponpa ini akan memperbaiki patologi akson pada hewan
percobaan.
pada mitokondria (Nikolaeva et al., 2005).
f. Kejadian Lanjutan Setelah Influks Kalsium
Influks kalsium diikuti aktivasi calcium-dependent protease, seperti
calpains, yang akan membelah dan mendegradasi protein sitoplasma. Peningkatan
aktivitas calpains sudah terbukti pada diffuse axonal injury setelah trauma kepala,
stroke, cedera spinal cord, dan kelainan neurodegenerative (Vosler et al., 2008).
Calpains akan mendegradasi sejumlah target, seperti protein sitoskeleton, enzim,
reseptor, channel, dan faktor transkripsi melalui proses proteolisis (Saatman et al.,
2010). Calpains juga akan mendegradasi substrat yang penting untuk stabilitas
akson. Ini akan diikuti oleh pemecahan collapsing response mediator protein-2
(CRMP-2) secara proteolotik (Touma et al., 2007). Karena CRMP-2 berikatan
dengan kinesin-1 dan berpartisipasi dalam transport akson, transport akson pada
Selain calpain, ada beberapa enzim yang tergantung kalsium yang
berpartisipasi dalam degenerasi akson. Cedera akson transien akibat regangan
akan menyebabkan lepasan kalsium yang pertama kali terjadi dari depot intrasel.
Ini akan diikuti dengan peningkatan kadar kalsium intrasel dalam 48 jam. Dalam
proses ini, penghambatan calcineurin, suatu calcium-dependent phosphataseakan
menghalangi degenerasi akson sekunder (Staal et al., 2010). Calpain dan
calcineurin hanyalah dua contoh dari protein tergantung kalsium yang terlibat
dalam proses degenerasi lebih lanjut.
g. Cedera Mitokondria
Cedera mitokondria memegang kunci penting dalam lokalisasi gangguan
kontinuitas akson. Pada model cedera kepala dengan diffuse axonal injury, influks
kalsium diduga dimediasi melalui pori-pori axolemma. Namun, kerusakan akson
dan aktivasi calpain tidak terjadi secara simultan sekaligus pada seluruh
akson.Kerusakan terjadi pada titik-titik fokal disertai akumulasi mitokondria
(Kilinc et al., 2009). Akumulasi fokal mitokondria mungkin akan menyebabkan
gangguan sitoskeleton fokal dan penumpukan substrat. Pada FAD, mitokondria
dirusak oleh oksigen dan nitrogen reaktif yang kemungkinan besar berasal dari
makrofag. Proses ini sendiri akan memacu degenerasi akson lebih lanjut (Nikic et
al., 2011).
Protein amiloidogenik, seperti alpha-synuclein, tau, dan Aβ diduga
berperan menyebabkan degerasi akson pada beberapa kelainan neurodegeneratif,
yaitu melalui hambatan pada mekanisme transport akson.Agregasi protein tidak
terjadi pada seluruh jenis degerasi akson, tetapi dapat menyebabkan gangguan
akson.Overekspresi human wild-type alpha-synuclein oleh lentivirus
menyebabkan agregasi dan degenerasi akson SSP (Decressac et al., 2011).
Agregasi protein amiloidogenik tidak dapat dianggap suatu proses yang berdiri
sndiri, karena penelitian telah membuktikan peningkatan konsentrasi kalsium
akan menyebabkan agregasi alpha-synuclein pada kultur sel (Nath et al., 2011).
Karena itu, kita dapat mengeluarkan hipotesis bahwa lesi akson dengan influks
kalsium dapat menyebabkan agregasi protein amyloid. Sebaliknya, adanya
alpha-synuclein sendiri akan memengaruhi degenerasi akson akibat trauma. Tikus
transgrenik dengan over ekspresi human alpha-synuclein (Thy1-αSynWT)akan
memiliki agregat alpha-synuclein pada akson saraf sciatic. Hewan ini akan
mengalami peningkatan degenerasi Wallerian setelah axotomy saraf sciatic.
Sementara itu, degenerasi yang terjadi setelah axotomy pada tikus tanpa
ekspresi alpha synuclein (C57BL/6-Ola-hsd strain by Harlan B6),
degenerasi akson akan terjadi dalam kecepatan yang lebih lambat (Siebert
et al., 2010). Hasil penelitian ini cukup membingungkan karena
alpha-synuclein selama ini dianggap sebagai kunci pada kelainan
neurodegeneratif sel saraf pusat.Meskipun demikian, penelitian ini
mekanisme kerusakan akson yang lebih luas, termasuk lesi akibat trauma.
Proses yang terjadi pada kelainan neurodegeneratif kemungkinan besar
merupakan suatu proses yang terpisah. Mekanisme kerja yang tepat belum
dipecahkan sepenuhnya, tetapi data yang ada mengarahkan kita pada
kemungkinan adanya suatu interaksi langsung dengan sitoskeleton, seperti
neurofilamen, tau, dan tubulin (Kanazawa et al., 2008) serta interaksi
dengan protein transport, seperti dynein dan kinesin-1 (Utton et al., 2005).
i. Transport Akson
Jika kita berasumsi bahwa degenrasi akson pada trauma merupakan suatu
proses mekanis, gangguan transport akson sesudah trauma merupakan suatu hal
yang sangat mungkin terjadi. Karena akson memiliki hubungan yang kompleks
dengan inti sel, dibutuhkan suatu transport yang efektif pada tujuan, seperti sinaps
terminal atau nodes of Ranvier. Gangguan transport akson sudah terbukti terjadi
pada beberapa kelainan degeneratif, seperti Parkinson, Alzheimer’s, dan
Huntington (Morfini et al., 2009).
Transport akson dimediasi oleh dua kelompok proten utama. Kelompok
pertama adalah kinein.Kinein berperan dalam memediasi transport
anterograde.Sementara itu, kelompok kedua adalah dynein, yang berperan dalam
transport retrograd. Tikus percobaan dengan mutasi gen KIF1Bβ, gen pengkode
kinesin menunjukkan gangguan vesikel sinaps dan kelemahan otot progresif
akibat neuropati perifer. Pada manusia, mutasi KIF1Bβ ditemukan pada penderita
Gangguan transport akibat mutasi kinesin light chain-1 juga akan mengaktivasi
stress kinase, seperti c-Jun-N terminal kinase. Ini akan menyebabkan fosforilasi
yang abnormal dan agregasi tau (Falzone et al., 2009). Gangguan transport
retrograd sendiri tidak memberikan efek klinis yang relevan.Missense point
mutation pada dynein heavy-chain menyebabkan degenerasi motoneuron pada
tikus heterozygous dan pembentukan inclusion body pada binatang homozygous
(Hafezparast, 2003).Mutasi dynein juga mungkin dapat dihubungkan dengan
degenerasi akson pada penyakit motoneuron (Ravikumar et al., 2005).
Gangguan transport akson pada akhirnya akan menyebabkan gangguan,
mulai dari distrofi akson sampai degenerasi akson. Gangguan ini potensial
menjadi target terapi pada trauma dan kelainan neurodegenerative.Namun, proses
yang terjadi sangat kompleks dan tidak dapat ditangani hanya dengan satu
intervensi spesifik.
j. Aktivasi Kinase
Kinase berperan dalam eksekusi destruksi akson. JNK dikenal sebagai
protein kinase yang dipicu stress karena aktivitasya meningkat pada stress seluler
akibat lingkungan, seperti osmotic stress, redox stress, atau irradiation (Weston
dan Davis, 2007). Begitu diaktivasi, kinase akan mempropagasi sinyal yang
memacu apoptosis sel. Cedera juga dapat mengaktivasi JNK pada akson dan
menyebabkan gangguan transport akson. Activated pospho-JNK terdapat dalam
jumlah banyak pada traktur kortikospinal tikus dengan cedera spinal cord.
nantinya akan memperbaiki fungsional tikus (Yoshimura et al., 2011). Sebaliknya,
mutasi kinesin-light-chain 1 terjadi akibat aktivasi JNK, bersamaan dengan tau
protein yang mengalami proses hiperfosforilasi pada akson yang mengalami
proses distrofi (Falzone et al., 2009).
k. Autofagi dan sistem ubiquitin-proteasome
Degradasi protein atau organela terjadi dengan berbagai jalur
degradasi.Salah satu di antaranya adalah autofagi.Setelah kerusakan akson
mekanis, dijumpai peningkatan autofagosom pada lesi yang bergantung influks
kalsium (Koch et al., 2010). Autofagi juga diinduksi oleh neurit simpatis yang
mengalami degenerasi, seperti setelah aksotomi.
Sistem ubiquitin-proteason juga terbukti berhubungan dengan degenerasi
akson. Pada binatang percobaan dengan cedera nervus optikus, inhibisi sistem
ubiquitin-proteasome akan menghambat degenerasi Wallerian. Fragmenasi
mikrotubuli yang terjadi segera setelah trauma dapat dikurangi dengan pemberian
proteasome inhibitor MG132 (Zhai et al., 2003).
B. Sistem Ubiquitin Proteasome (SUP)
Sistem Ubiquitin Proteasome (SUP) merupakan jalur seluler utama pada
eukariotik yang mengontrol pergantian protein melalui proteasome 26 S. Banyak
proses dasar sel, seperti siklus sel, transduksi sinyal, apoptosis, dan pengontrolan
kualitas protein diatur oleh SUP (Cardozo dan Pagano, 2004). Induksi proses ini
penandaan protein yang menjadi target dengan molekul kecil, yang dikenal
dengan nama ubiquitin. Ubiquitinakan berikatan secara kovalen dengan residu
lisin pada protein yang menjadi target. Proses ini dikenal dengan nama
ubiquinisasi.
Ubiquinisasi merupakan proses bertahap yang memerlukan aktivitas tiga
kelas enzim, yaitu ubiquitin-activating enzyme (E1), ubiquitin-conjugating
enzyme (E2), dan ubiquitin-protein ligase (E3) (Pickart, 2004).
Ubiquitin-activating enzyme bekerja dengan bergantung ATP dengan mengikat active site
enzim ini (residu cysteine) pada residu C-terminal glycine pada ubiquitin, yang
pada akhirnya akan mengaktifkan ubiquitin. Ubiquitin yang sudah aktif kemudian
akan ditransfer pada E2, yang relatif sedikit terdapat pada eukariot, melalui suatu
reaksi esterifikasi. Akhirnya, ubiquitin ditempelkan pada residu lysine pada
protein target dengan ikatan isopeptida. Langkah ini memerlukan aktifitas salah
satu dari ratusan sistem E3.E3 bertindak sebagai pengenal substrat dan mampu
berinteraksi baik dengan E2 maupun dengan substrat protein. (Ardley dan
Robinson, 2005)
Ada dua jenis domain pada Ubiquitine – protein ligase (E3). Suatu E3
hanya akan memiliki salah satu di antara keduanya. Kategori pertama memiliki
domain zinc-binding RING (Really Interesting New Gene) yang memacu
ubiquitinisasi dengan berikatan pada E2 dan substrat (6).Kategori kedua memiliki
domain HECT (Homologous to E6-AP Carboxyl-terminus) yang berikatan
langsung dengan ubiquitin, kemudian mentransfer ubiquitin tersebut pada
selesai dengan sempurna, meninggalkan protein yang menjadi target hanya
berikatan dengan satu ubiquitin. Namun, ini akan sering berlanjut melalui
pengikatan molekul ubiquitin bebas lainnya pada residu lysine spesifik pada
ubiquitin terakhir, membentuk suatu rantai ubiquitin. Pada cara ini, protein target
akan berikatan dengan banyak ubiquitin dan beberapa protein memerlukan kerja
ubiquitin-chain elongation-factor (E4) supaya proses ubiquitinisasi dapat berjalan
dengan efisien (Koegi et al., 1999).
Rantai ubiquitin bekerja sebagai penanda proses yang secara fungsional
berbeda, tidak hanya terbatas pada degradasi proteasom. Awalnya, protein dengan
banyak ubiquitin dianggap hanya memiliki target berupa proteasom 26S. Namun,
saat ini diketahui bahwa proses poliubiquitinisasi dapat terjadi dalam dua bentuk,
yaitu homopolymeric dan heteropolymeric. Pada bentuk homopolymeric, molekul
ubiquitin berikatan satu sama lain menggunakan donor residu lysine yang sama.
Rantai homopolymeric ubiquitin akan memanjang melalui ikatan beberapa residu
lysine yang terdapat pada molekul ubiquitin, yaitu Lys48, Lys63, Lys6, Lys11, Lys29,
Lys27 and Lys33. Setiap jenis ikatan ini akan menyebabkan kaskade sinyal yang
spesifik. Sebagai contoh, penambahan rantai yang berikatan dengan Lys48 pada
protein akan menyebabkan munculnya sinyal degradasi, sementara Lys63 akan
menyebabkan sinyal perbaikan DNA, endositosis, atau transduksi sinyal. Rantai
polyubiquitin heteropolymeric ditandai dengan ikatan molekul ubiquitin dengan
lebih dari satu jenis pengikat (Koegi et al., 1999).Fungsi jenis ini belum diteliti
1. Enzim Deubiquitinating (DUB)
Ubiquitinisasi merupakan suatu proses yang reversibel. Terdapat
sekelompok enzim yang dapat melepaskan ikatan ubiquitin dengan protein yang
sudah mengalami ubiquitinisasi.Kelompok enzim ini dikenal dengan enzim
deubiquitinating (DUB). DUB merupakan protease yang dapat menghidrolisis
ikatan isopeptida antara ubiquitin dengan substratnya melalui suatu proses yang
bergantung ATP (Yao dan Cohen, 2002). Hampir mirip dengan ubiquitinisasi,
proses deubiquitinisasi merupakan suatu proses yang sangat dikontrol dengan
ketat dan berperan dalam regulasi siklus sel (Song dan Rape, 2008), ekspresi gen
(Daniel dan Grant, 2007), degradasi protein bergantung lysosome dan proteasome
(Schmidt, 2005), DNA repair (Kennedy dan D’Andrea, 2005), aktivasi kinase
(Komada, 2008), dan proses seluler lainnya. DUB memegang peranan pada jalur
ubiquitin. Pertama, DUB akan berperan sebagai antagonis dalam proses
ubiquitinisasi dengan melepaskan ubiquitin dari protein yang sudah mengalami
ubiquitinisasi pada suatu sel (Nijman, 2005). Kedua, DUB berperan dalam
aktivasi proprotein ubiquitin. Pada dasarnya, ubiquitin selalu diekspresikan
sebagai proprotein yang berikatan pada protein ribosomal atau diekspresikan
sebagai rantai poliubiquitin linear yang harus menjalani proses hidrolisis agar
membentuk molekul ubiquitin sederhana (Baker, 1987). Ketiga, DUB akan
mendaur ulang ubiquitin yang sudah yang sudah memiliki ikatan ester dengan
monomeric bebas dari rantai poliubiquitin yang tidak berikatan dalam sel
(Piotrowski, 1997). Kelompok ini juga mencakup poliubiquitin bebas yang
dihasilkan oleh enzim konjugasi yang sudah dilepaskan dari protein lain yang
sudah menjalani deubiquitinisasi).
Pada sel eukariot, DUB dibagi lagi menjadi lima kelompok berdasarkan
struktur dan fungsinya, yaitu ubiquitin-specific protease (USP), ubiquitin
C-terminal hydrolase (UCH), Otubain protease (OTU), Machado-Joseph disease
protease (MUD), dan suatu kelompok metalloprotease (Nijman et al., 2005).
2. Fungsi Sistem Ubiquitin Proteasome pada Neuron
SUP sudah dianggap menjadi protein kunci dalam proses modifikasi dan
degradasi yang penting dalam perkembangan, pemeliharaan, dan remodeling
koneksi sinaps pada otak (Kuo et al., 2006). Karena proses seluler dan fisiologi
yang melibatkan ubiquitin sangat banyak, tidak mengejutkan ada banyak fungsi
saraf yang sangat bergantung pada SUP yang baik. Pada sistem saraf Drosophila,
SUP terlibat dalam pruning dendrit.Pruning merupakan suatu proses yang terjadi
saat metamorphosis Drosophila. Saat metamorphosis, sekelompok neuron sensori
larva tumbuh kembali membentuk suatu jaringan dendrit yang baru, Pada tahun
2006, sekelompok peneliti dari UCSF menunjukkan bahwa proses pruning
berhubungan dengan aktivitas enzin E2/E3 yang spesifik (Kuo et al., 2006).
Banyak penelitian menunjukkan bahwa UPS akan terlibat dalam
terbukti meningkatkan jumlah percabangan sinaps pada neuromuscular
junction.Ukuran sinaps juga diatur oleh SUP (van Roessel, 2004).
Modifikasi efikasi sinaps juga disertai dengan perubahan komposisi
protein pada sinaps (Mallnow, 2003).Perubahan komposisi protein ini dapat
terjadi dengan mengikutkan protein yang baru disintesis atau melalui degradasi
sebagian komponen protein terebut. Proses sintesis dan degradasi ini sangat diatur
oleh SUP.
Lebih lanjut lagi, aktivitas yang bergantung degradasi protein diperlukan
untuk plastisitas saraf pada hippocampus mammalia. Inhibisi proteasome dapat
memacu long term potentiation (LTP) dan long term depression (LTD). Keduanya
membentuk plastisitas sinaps pada hippocampus yang terlibat dalam pada proses
belajar dan memori. Inhibisi proteasome terbukti menghambat depresi sinaps
yang bergantung NMDA dan mGluR (Deng dan Lei, 2007).
3. Penyakit Neurodegeneratif dan SUP
SUP berperan pada banyak kelainan sistem saraf, terutama kelainan
neurodegenerative, seperti Alzheimer’s (Gong et al., 2006), Parkinson’s (Upadhya
dan Hedge, 2005), Amylotrophic Lateral Sclerosis (ALS), dan Huntington’s
(Rubinsztein, 2006). Seluruh penyakit ini sekarang dianggap sebagai
“proteinopati” sistem saraf, yang ditandai dengan akumulasi misfolded protein
yang menumpuk dan tidak dapat didegradasi. Tumpukan misfolded protein ini
dapat berupa plaque dan tangle (Alzheimer’s), Lewy bodies (Parkinson’s), dan
sudah menjalani proses ubiquitinisasi ditemukan pada tumpukan protein ini. Ini
menggambarkan suatu kemungkinan bahwa protein ini sudah ditandai untuk
proses degradasi, tetapi proses degradasi itu sendiri belum berjalan dengan efisien.
Pada penderita Dementia dengan Lewy bodies dan penderita Parkinson, hanya
ditemui satu sampai tiga rantai ubiquitin, sementara proses pengenalan dan
degradasi yang sempurna terjadi bila ikatan ubiquitin berjumlah lebih dari empat
(Anderson et al., 2006). Aktivitas proteasome menurun pada penderita
Alzheimer’s dan Parkinson’s. Karena itu, disfungsi SUP kemungkinan terjadi
karena peningkatan jumlah misfolded dan agregat protein serta ketidakmampuan
untuk mengenali dan mendegradasinya (Dahlmann, 2007).
Bagian penting pemahaman disfungsi SUP pada proses degenerasi saraf
adalah identifikasi gen yang terlibat, baik secara familial maupun sporadik. Ada
sepuluh lokus gen yang sudah diidentifikasi pada penderita Parkinson’s. Dua di
antaranya merupakan bagian dari SUP.Pertama, gen PARK2 mengkode parkin
(E2-dependent E3 ligase) (Shimura, 2000). Mutasi PARK2 menyebabkan
terjadinya Parkinson’s pada usia muda dengan hilangnya neuron dopaminergik
pada substansia Nigra tanpa adanya Lewy Bodies (van Coelln, 2004). Gen kedua
adalah PARK5 yang mengkode UCH-L1, sejenis DUB yang memiliki fungsi
hydrolase dan ligase (Osaka et al., 2003). Mutasi PARK5 berhubungan dengan
Parkinson’s, baik familial maupun sporadik pada manusia. Karena UCH-L1
dianggap membantu mempertahankan kadarubiquitin monomer pada neuron,
4. Ubiquitin Carboxyl-Terminal Hydrolase (UCH-L1) dan Hubungannya dengan
Degenerasi Saraf
UCH-L1, juga dikenal dengan PGP9.5 merupakan ikatan 223 asam amino
yang memegang peranan penting pada SUP. Ini termasuk ke dalam kelompok
DUB. Ada lima jenis kelompok ini, yaitu UCH-L1 – UCH-L5. UCH-L1 hanya
selektif ditemui pada neuron dalam jumlah banyak, sekitar 1-2% di antara seluruh
soluble protein otak (Wilkinson, 1989).Awalnya, UCH-L1 dianggap hanya
berperan sebagai DUB.Namun, beberapa penelitian terakhir menunjukkan bahwa
protein ini juga memegang peranan besar pada SUP. UCH-L1 diketahui akan
menciptakan ubiquitin bebas monomeric, baik dari bentuk prekursor (ubiquitin
polypeptide) maupun dari ubiquitin yang bergabung dengan protein ribosomal
(Larsen et al., 1998). Pada penelitian in vitro, UCH-L1 juga terbukti memiliki
aktifitas ubiquitin-ligase (Liu et al., 2002). Selain itu, UCH-L1 bersama dengan
ubiquitin dapat menghambat degradasi lisosomal, sehingga dapat
mempertahankan kadarubiquitin monomeric pada neuron (Osaka et al., 2003).
Banyak penelitian membuktikan hubungan antara UCH-L1 dengan
kelainan neurodegeneratif, seperti Alzheimer’s dan Parkinson’s.Ini diawali
dengan penemuan mutasi gen UCH-L1 pada dua saudara di keluarga Jerman yang
menderita Parkinson terkait autosomal dominan (Leroy et al., 1998).Isoleusin
pada posisi 93 diganti dengan methionine.Produk gen yang dihasilkan dinamai
UCH-L1 193M. Hanya satu dari kedua alel UCH-L1 yang berubah pada mutasi
Namun, orang tua kedua anak tersebut tidak menderita Parkinson. Ini
menggambarkan kemungkinan mutasi 193M mungkin merupakan suatu
polimorfisme yang sangat jarang terjadi yang tidak berkaitan dengan suatu
kelainan atau mutasi pada orang tua kedua anak itu bersifar inkomplit (Liu et al.,
2002). Pada penelitian in vitro, mutasi ini menyebabkan penurunan fungsi katalis
sampai 50%. Hilangnya fungsi UCH-L1 akan mengurangi ketersediaan ubiquitin
bebas dan menyebabkan gangguan degradasi protein melalui SUP. Lebih lagi,
tikus transgrenik dengan mutasi 193M mengalami penurunan neuron
dopaminergik pada substansia nigra yang signifikan (Setsuie et al., 2007).
Seperti yang disebutkan sebelumnya, UCH-L1 dipercaya memiliki
aktiivtas ubiquitin ligase. Pada percobaan invitro, aktivitas ligase ini bergantung
pada proses dimerisasi dan berperan dalam ubiquitinisasi a-synuclein,
kemungkinan dengan ikatan rantai K63. A-synuclein merupakan protein yang
banyak terdapat pada ujung pre-sinaps dan terlibat dalam pelepasan
neurotransmiter (Kahle et al., 2000). Protein ini merupakan komponen utama dari
Lewy bodies yang banyak ditemui pada penderita Parkinson. Pada tikus
percobaan, UCH-L1 berada dalam vesikel pada sinaps dan terlibat dalam
presipitasi a-synuclein. Ubiquitinisasi a-synuclein dengan UCH-L1 dipercaya
akan menghambat degradasi proteasome, sehingga menyebabkan penumpukan
dan agregasi dalam neuron dan Lewy bodies (Liu et al., 2002). Pada keadaan
fisiologis, a-synuclein berbentuk monomeric, namun dalam konsentrasi tinggi,
a-synuclein dapat berbentuk polimer, menjadi bentuk filamen yang dapat
Lewy bodies. Selain itu, ada polimorfisme UCH-L1 lain yang tampaknya
mengurangi kemampuan ligase. Mutasi ini ditemui pada populasi tertentu di Asia
Timur, berupa perubahan serine 18 menjadi serine.Mutasi ini dinamai UCH-L1
S18Y. Mutasi S18Y dari UCH-L1 menyebabkan penurunan kadara-synuclein
yang mengalami ubiquitinisasi dan berhubungan dengan penurunan resiko
terjadinya Parkinson’s.
UCH-L1 juga terlibat dalam plastisitas sinaps pada tikus percobaan. Tikus
transgenic percobaan ini dinamai gracile axonal dystrophy (gad), berupa tikus
yang sudah mengalami delesi ekson 7 dan 8 pada gen Uch-l1. Ekspresi UCH-L1
pada susunan saraf tikus-tikus ini tidak dapat dideteksi. Tikus gad mengalami
paresis motorik akibat degenerasi aksonal dari neuron spinal cord dan traktus
spinoserebelar. Tikus gad ini juga menunjukkan penurunan kadarubiquitin
monomeric (Gong et al., 2006).
C. Biomarker pada Cedera Kepala
Aplikasi penggunaan biomarker pada cedera kepala akan sangat
memberikan manfaat sebagai tambahan alat diagnosis berbagai macam cedera
kepala. Sebagai contoh, pelatih pada sepak bola dapat menggunakan biomarker
sebagai penentu keputusan yang objektif untuk penghentian permainan saat terjadi
sport concussion. Biomarker yang tervalidasi akan merevolusi penatalaksanaan
dan diagnosis cedera kepala, bahkan sekaligus dapat membantu menilai prognosis
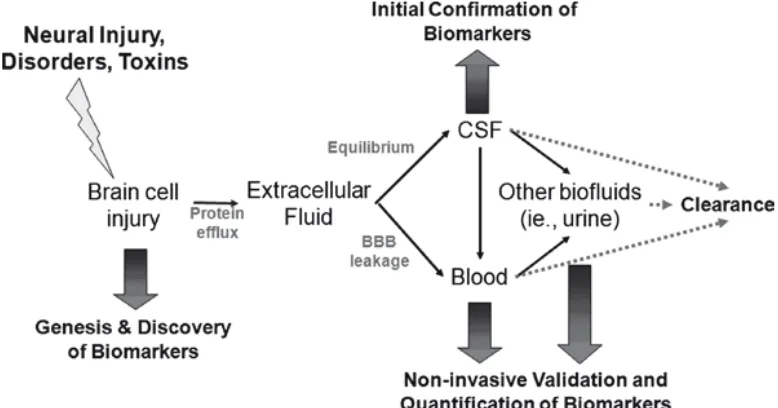
Gambar 2 menggambarkan jalur mulai terbentuknya biomarker cedera
kepala pada jaringan otak sampai deteksinya pada CSF dan darah.Selama cedera
otak, protein neural terlepas ke lingkungan ekstrasel, dan kemudian
CSF.Biomarker ini muncul dalam konsentrasi tinggi pada CSF. Protein-protein ini
kemudian akan mencapai aliran darah melalui sawar darah otak yang terganggu
atau melalui filtrasi CSF. Karena volume CSF manusia adalah sekitar 30-40 kali
lebih sedikit dibandingkan volume darah (CSF 125-150 mL, darah 4-5 L),
konsentrasi biomarker akan jauh lebih tinggi pada CSF dibandingkan darah.
Gambar 2. Terbentuknya biomarker setelah trauma dan penyebarannya pada darah (Kobeissy et al., 2008)
Sampai saat ini, kebanyakan penelitian biomarker cedera kepala
berfokus pada profil protein.Namun, genom manusia diperkirakan
mengandung 23.000 gen. Separuh di antaranya ada dalam jumlah sangat
sedikit.Akibatnya, mendata seluruh proteasome yang ada menjadi sangat
proses cedera kepala.
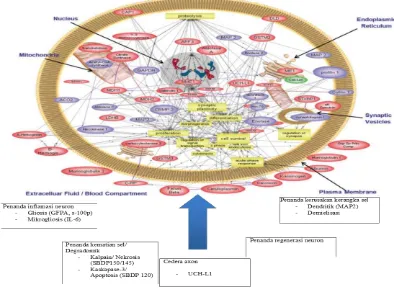
Sebuah review tahun 2008 oleh Kobeissy et al mencoba mendata
seluruh proteasome yang kemungkinan besar terlibat dalam cedera kepala
manusia (Gambar3 )
Gambar 3 Proses cedera kepala dengan protein yang potensial menjadi biomarker
(Kobeissy et al, 2008)
Penanda yang ada dapat dikelompokkan ke dalam beberapa
penanda inflamasi, metabolit, neurotransmitter, metabolit lipid, dan
biomarker secondary insult (tabel 1)
Tabel 1 Penanda cedera kepala (Dash et al., 2010)
Biomarker Kegunaan Diagnosis Sumber sampel Keterangan
α II-Spectrin BDPs Meningkat pada cedera kepala, berhubungan dengan ukuran lesi dan keparahan cedera
CSF, jaringan otak, serum Banyak ditemui pada otak, meningkat pada kematian sel
α –Synuclean C-tau Meningkat pada cedera kepala, peningkatan C-tau berhubungan dengan perburukan hasil akhir dan peningkatan TIK
Jaringan otak, CSF, serum Spesifik pada SSP, memerlukan validasi lagi pada manusia, alat diagnosis yang jelek pada CKR
3’,5’cAM/2’,3’cAMP Kadar pada CSF berkorelasi dengan derajat koma
CSF Second messenger
Ceruloplasmin/Cu Menurun pada peningkatan TIK Serum
CK-BB Gangguan blod brain barrier
mempermudah penetrasi serum, tidak berhubungan dengan volume kontusio
CSF, serum Waktu paruh pendek,
dieliminasi dengan cepat
CRP & SAA Marker cedera secara umum Serum Diinduksi dengan
cepat. Mengindikasikan politrauma
CRMP-2 Menurun pada penderita mesial
temporal lobe epilepsy. Berhubungan dengan Alzheimer’s
Jaringan otak Mungkin dapat
digunakan sebagai marker epilepsy post trauma
FABP Meningkat pada cedera kepala
ringan
Serum/plasma, jaringna otak Akurasi tinggi untuk cedera kepala ringan F2-Isoprostane Meningkat pada penyakit
multipel
CSF, serum Penanda kerusakan
oksidatif
4-HNE Meningkat pada jaringan otak
penderita cedera kepala
Jaringan otak Marker peroksidasi
lemak
5-HIAA Meningkat pada CSF, indikator
trauma
CSF Marker gangguan
neurotransmisi
GFAP Prediktif untuk peningkatan ICP,
penurunan MAP, penurunan CPP, dan GOS
CSF, serum Spesifik untuk SSP.
Mungkin kurang sensitif pada cedera kepala ringan
HVA Meningkat pada CSF CSF Marker gangguan
neurotransmisi
ICAM Meningkat pada gangguan sawar
darah otak
CSF, serum Penanda disfungsi
neurovaskuler
IL-1b Peningkatannya berhubungan
dengan hasil akhir yang buruk
CSF Muncul saat terjadi
jaringan parut astroglia
IL-6 Hasil yang meragukan.
Peningkatan IL-6 berhubungan dengan hasil akhir yang membaik
CSF, serum, dialisat intrakranial
Penanda cedera organ multipel
IL-8 Meningkat pada CSF penderita
cedera kepala berat
CSF, serum Penanda inflamasi
mortalitas
Biomarker Kegunaan Diagnosis Sumber Sampel Keterangan
IL-12p70 Meningkat pada CSF penderita
CKB
CSF Marker inflamasi
Laktat Berhubungan dengan keparahan
cedera
CSF Marker gangguan
metabolism otak
Magnesium Penurunan Mg dalam lima hari
pertama berhubungan dengan keparahan cedera
Serum
MBP Peningkatan MBP berhubungan
dengan hasil akhir yang lebih jelek pada anak-anak
CSF, serum Penanda cedera white
matter
MCP-1 Peningkatan pada otak tikus
dalam empat jam pertama setelah trauma
Jaringan otak Belum diuji pada
manusia
MIP-1a Meningkat pada CSF penderita
CKB
CSF Penanda invasi sel
inflamasi
Phospho-neurofilament
Meningkat pada penderita cedera kepala
Serum Perlu divalidasi
manusia
NSE Gangguan neuropsikologi pada
lesi intrakranial
CSF, Serum, jaringan otak Penanda small cell lung cancer, neuroendocrine bladder tumor, stroke, dan neuroblastoma
NE Meningkat pada penderita dalam
keadaan koma dan penderita politrauma
Serum Penanda gangguan
neurotransmisi
S100b Meingkat pada penderita cedera
kepala ringan
CSF, Serum Tidak spesifik untuk
cedera kepala. Dapat digunakan sebagai penanda gangguan Sawar darah otak
TGF-b Meningkat pada cedera kepala,
tetap meningkat selama 3 minggu
CSF Pertumbuhan dan
diferensiasi sel, angiogenesis, fungsi imun, apoptosis
TNF-a Meningkat pada penderita cedera
kepala berat.
CSF, serum Penanda inflamasi
UCH-L1 Meningkat pada CSF,
berhubungan dengan mortalitas, komplikasi, dan hasil akhir dalam 6 bulan pertama
CSF Marker yang banyak
ditemukan pada otak
BAB III
KERANGKA TEORI, KERANGKA KONSEP, HIPOTESIS PENELITIAN
A. Kerangka Teori
B. Kerangka Konsep
Variabel Bebas : • UCH – LI
Serum
Variabel Terikat: • GCS awal sesudah
resusitasi • Lama Rawatan • Mortalitas tiga hari
C. Hipotesis
1. Hipotesis mayor
Kadar UCH-L1serum berhubungan dengan tingkat keparahan
cedera kepala.
2. Hipotesis minor
a. Kadar UCH-L1 serum berhubungan dengan GCS awal setelah
resusitasi.
b. Kadar UCH-L1 serum berhubungan dengan mortalitas pada tiga
hari pertama.
c. Kadar UCH-L1 serum berhubungan dengan lama rawatan
BAB IV
METODE PENELITIAN
A. Rancangan Penelitian
Penelitian ini merupakan penelitian analitik dengan pendekatan
potong lintang (cross sectional).
B. Waktu dan Tempat Penelitian
Waktu penelitian mulai Juni 2012 sampai Maret 2013.Tempat
peneltian adalah Departemen Ilmu Bedah Saraf RSUP H Adam Malik
Medan.
C. Populasi dan Sampel Penelitian
1. Populasi Target
Populasi yang dilakukan generalisasi/inferensialnya yaitu seluruh
penderita cedera kepala ringan, sedang, dan berat.
2. Populasi Terjangkau
Kumpulan dari satuan/unit yang dilakukan pengambilan sampel
penelitian, yaitu penderita cedera kepala ringan, sedang, dan berat
yang dirawat di RSUP H. Adam Malik Medan.
3. Sampel Penelitian
Bagian dari populasi terjangkau yang diambil untuk dilakukan
berat yang dirawat di RSUP H. Adam Malik Medan.Sampel
penelitian diambil dengan cara consecutive sampling dengan
penetapan kriteria inklusi dan kriteria eksklusi.
D. Kriteria Sampel Penelitian
1. Kriteria Inklusi
a. Seluruh penderita cedera kepala ringan, sedang, dan berat
b. Usia penderita 18-50 tahun
c. Onset kejadian di bawah 48 jam
2. Kriteria eksklusi
a. Penderita cedera multipel
b. Penderita dengan indikasi operasi, baik saat masuk atau
selama follow up
c. Dengan riwayat dementia, psikosis, atau kelainan susunan
saraf pusat aktif
d. Penderita hamil
e. Trauma tajam kepala
E. Besar Sampel Penelitian
Besar sampel dihitung berdasarkan rumus yang berdasarkan
SampleSize Determination in Health Studies, a practical manual version
Zα : 1,96 (α = 5%)
Berdasarkan rumus di atas, didapat bahwa besar sampel satu
kelompok minimal 21 orang.
(
)
1. Penanganan trauma kepala yang komprehensif 2. Pencatatan ulang identitas (GCS masuk, suku, usia) 3. Pengambilan serum untuk pemeriksaan UCHL-1 4. Pencatatan lama rawatan
Pemeriksaan UCHL-1 serum kumulatif