KEBERADAAN DAN STATUS FUNGI MIKORIZA ARBUSKULA
PADA LAHAN KELAPA SAWIT
DI DATARAN TINGGI DAN DATARAN RENDAH
SKRIPSI
Oleh :
YEPTA JAYA SAHPUTRA BARUS
PROGRAM STUDI KEHUTANAN
FAKULTAS PERTANIAN
UNIVERSITAS SUMATERA UTARA
MEDAN
KEBERADAAN DAN STATUS FUNGI MIKORIZA ARBUSKULA
PADA LAHAN KELAPA SAWIT
DI DATARAN TINGGI DAN DATARAN RENDAH
SKRIPSI
Oleh :
YEPTA JAYA SAHPUTRA BARUS
101201061/BUDIDAYA HUTAN
Skripsi sebagai salah satu syarat untuk memperoleh gelar
Sarjana Kehutanan pada Program Studi Kehutanan Fakultas Pertanian Universitas Sumatera Utara
PROGRAM STUDI KEHUTANAN
FAKULTAS PERTANIAN
UNIVERSITAS SUMATERA UTARA
MEDAN
LEMBAR PENGESAHAN
Judul Penelitian : Keberadaan dan Status Fungi Mikoriza Arbuskula pada Lahan Kelapa Sawit Dataran Tinggi dan Dataran Rendah
Nama : Yepta Jaya S Barus
Nim : 101201061
Program Studi : Kehutanan
Disetujui oleh, Komisi Pembimbing
Dr. Deni Elfiati, S.P, M.P. Dr. Delvian, S.P, M.P Ketua Anggota
Mengetahui,
ABSTRACT
Yepta Jaya Sahputra Barus: Existence and Status of Arbuscular Mycorrhizal Fungi on Highland and Lowland Oil Palm Field. Supervisied by DENI ELFIATI and DELVIAN.
The goal of this research is to know the existence and the status of Arbuskular Mycorrhiza at Oil Palm ecosystems in highland and lowland. Soil sample has been taken from Adia Nangka Village, District of Lae Parira, Region of Dairi and Batang Terap Village, District of Perbaungan, Region of Serdang Bedagai. This research use soil separating method to obtain spores and root coloring method to find out root colonization. The trapping result with host of Zea mays obtained an average spore density on the highland is 938 spores/50 g soil, while the lowland obtained 128 spores/50 g soil. The result shows on the highland obtained 14 spores types of Glomus genus and 6 spores types of Acaulospora genus with up to 37% colonization percentage. Lowland obtained 7 spores types of Glomus genus and 3 spores types of Acaulospora genus with up to 25% colonization percentage.
ABSTRAK
Yepta Jaya Sahputra Barus: Keberadaan dan Status Fungi Mikoriza Arbuskula Pada Lahan Kelapa Sawit di Dataran Tinggi dan Dataran Rendah. Dibimbing oleh DENI ELFIATI dan DELVIAN.
Penelitian ini bertujuan untuk mengetahui keberadaan dan status FMA pada lahan kelapa sawit di dataran tinggi dan dataran rendah. Contoh tanah dan akar berasal dari desa Adia Nangka, Kec Lae Parira, Kab Dairi dan desa Batang Terap, Kec. Perbaungan, Kabupaten Serdang Bedagai. Penelitian ini menggunakan metode tuang saring untuk mendapatkan spora dan metode pewarnaan untuk mengetahui kolonisasi akar. Hasil trapping dengan tanaman inang Zea mays diperoleh kepadatan spora rata-rata pada dataran tinggi adalah 938 spora/50 gr tanah, sementara pada dataran rendah diperoleh kepadatan spora rata-rata adalah 128 spora/50 gr tanah. Hasil penelitian pada dataran tinggi didapat 14 tipe spora genus Glomus dan 6 tipe spora Acaulospora dengan persentase kolonisasi mencapai 37%. Pada dataran rendah diperoleh 7 tipe spora genus Glomus dan 3 tipe spora dari genus Acaulospora dengan persentase kolonisasi mencapai 25%.
RIWAYAT HIDUP
Penulis dilahirkan di Tiga Juhar, Kec. STM Hulu, Kab. Deli Serdang pada
8 Maret 1992 dari pasangan Ajartaki Barus dan Erni Br Ginting. Penulis
merupakan anak pertama dari tiga bersaudara.
Penulis menyelesaikan pendidikan Sekolah Dasar (SD) di SD Negeri
105403 Tiga Juhar lulus pada tahun 2004 dan melanjutkan pendidikan di SMP
Negeri 1 Kec STM Hulu hingga lulus tahun 2007. Kenudian penulis melanjutkan
Pendidikan di SMA Methodist-1 Medan dan lulus pada tahun 2010.
Pada tahun 2010 penulis mengikuti Ujian Masuk Bersama Perguruan
Tinggi Negeri (UMB-PTN) dan diterima di Program Studi Kehutanan, Fakultas
Pertanian, Universitas Sumatera Utara. Penulis memilih minat Budidaya Hutan.
Di masa perkuliahan penulis aktif dalam kepengurusan maupun kepanitiaan
dibeberapa organisasi kemahasiswaan seperti HIMAS USU, KorimUSU, dan
KMKS.
Dalam menyelesaikan kegiatan akademik, pada tahun 2012 penulis
mengikuti kegiatan Praktik Pengenalan dan Pengelolaan hutan (P3H) di hutan
pendidikan Universitas Sumatera utara, Tahura Bukit Barisan di Tongkoh
Kabupaten Karo. Penulis melaksanakan Praktik Kerja Lapang (PKL) di Taman
Nasional Gunung Gede Pangrango, Jawa Barat pada tanggal 4 Februari sampai
KATA PENGANTAR
Puji dan syukur penulis ucapkan kehadirat Tuhan Yang Maha Esa, karena
rahmatNya penulis dapat menyelesaikan skripsi yang berjudul “Keberadaan dan
Status Fungi Mikoriza Arbuskula pada Lahan Kelapa Sawit Dataran Tinggi dan
Dataran Rendah ” ini dengan baik.
Pada kesempatan ini penulis ingin mengucapkan terima kasih
sebesar-besarnya kepada orang tua penulis yang selalu memberi dukungan, kasih sayang,
doa, dan seluruh tenaganya. Kepada Dr. Deni Elfiati, S.P, M.P dan Dr. Delvian, SP., M.P. selaku Komisi Pembimbing yang telah banyak membimbing,
mengarahkan dan memberikan saran kepada penulis dalam menyelesaikan
penelitian dan penulisan skripsi ini.
Penulis juga ingin mengucapkan terimakasih kepada semua rekan-rekan
mahasiswa di Program Studi Kehutanan Fakultas Pertanian USU yang tak dapat
disebutkan satu persatu disini. Saudara-saudariku di HIMAS USU, Korim USU,
KMKS, Keluarga Besar KMBI. Para sahabat yang selalu memberi semangat,
dorongan, dan pengajaran kepada penulis.
Semoga penelitian ini akan memberi manfaat dan menyumbangkan
kemajuan bagi ilmu pengetahuan, khususnya bidang kehutanan.
Penulis
DAFTAR ISI
Faktor yang Mempengaruhi Kolonisasi dan Pemebentukan Spora FMA.... 6
Sebaran dan Ekologi Fungi Mikoriza ... ... 9
Persentase Kolonisasi Akar………... 19
DAFTAR TABEL
No Halaman 1. Hasil analisis tanah lahan kelapa sawit dataran tinggi
dan dataran rendah serta kriterianya menurut Penelitian
Tanah (1983) dalam Mukhlis (2007)………..………... 18 2. Data komponen Iklim pada kedua lokasi pengambilan sampel tanah... 19 3.Persentase kolonisasi akar kelapa sawit oleh FMA
pada dataran tinggi dan dataran rendah. ... ... 19 4. Kepadatan Spora FMA pada dataran tinggi dan
dataran rendah. ………..………... 21 5. Tipe dan karakteristik spora FMA pada lahan kelapa sawit
dataran tinggi... 21 6. Tipe dan karakteristik spora FMA pada lahan kelapa sawit
DAFTAR GAMBAR
ABSTRACT
Yepta Jaya Sahputra Barus: Existence and Status of Arbuscular Mycorrhizal Fungi on Highland and Lowland Oil Palm Field. Supervisied by DENI ELFIATI and DELVIAN.
The goal of this research is to know the existence and the status of Arbuskular Mycorrhiza at Oil Palm ecosystems in highland and lowland. Soil sample has been taken from Adia Nangka Village, District of Lae Parira, Region of Dairi and Batang Terap Village, District of Perbaungan, Region of Serdang Bedagai. This research use soil separating method to obtain spores and root coloring method to find out root colonization. The trapping result with host of Zea mays obtained an average spore density on the highland is 938 spores/50 g soil, while the lowland obtained 128 spores/50 g soil. The result shows on the highland obtained 14 spores types of Glomus genus and 6 spores types of Acaulospora genus with up to 37% colonization percentage. Lowland obtained 7 spores types of Glomus genus and 3 spores types of Acaulospora genus with up to 25% colonization percentage.
ABSTRAK
Yepta Jaya Sahputra Barus: Keberadaan dan Status Fungi Mikoriza Arbuskula Pada Lahan Kelapa Sawit di Dataran Tinggi dan Dataran Rendah. Dibimbing oleh DENI ELFIATI dan DELVIAN.
Penelitian ini bertujuan untuk mengetahui keberadaan dan status FMA pada lahan kelapa sawit di dataran tinggi dan dataran rendah. Contoh tanah dan akar berasal dari desa Adia Nangka, Kec Lae Parira, Kab Dairi dan desa Batang Terap, Kec. Perbaungan, Kabupaten Serdang Bedagai. Penelitian ini menggunakan metode tuang saring untuk mendapatkan spora dan metode pewarnaan untuk mengetahui kolonisasi akar. Hasil trapping dengan tanaman inang Zea mays diperoleh kepadatan spora rata-rata pada dataran tinggi adalah 938 spora/50 gr tanah, sementara pada dataran rendah diperoleh kepadatan spora rata-rata adalah 128 spora/50 gr tanah. Hasil penelitian pada dataran tinggi didapat 14 tipe spora genus Glomus dan 6 tipe spora Acaulospora dengan persentase kolonisasi mencapai 37%. Pada dataran rendah diperoleh 7 tipe spora genus Glomus dan 3 tipe spora dari genus Acaulospora dengan persentase kolonisasi mencapai 25%.
PENDAHULUAN
Latar Belakang
Organisme tanah sebagai komponen biotik di dalam tanah ada yang
bermanfaat dan ada pula keberadaannya mengganggu ataupun merugikan.
Organisme tanah yang menguntungkan yaitu yang terlibat dalam proses
dekomposisi bahan organik dan pengikat/penyedia unsur hara. Organisme tanah
yang merugikan adalah organsime yang memanfaatkan tanaman hidup, baik
sebagai sumber pangan atau sebagai inangnya, yang disebut sebagai hama atau
penyakit tanaman maupun sebagai kompetitor dalam penyerapan hara dalam
tanah. Keduanya bermanfaat pada penyediaan hara tersedia bagi tanaman serta
sebagai pemangsa parasit (Hanafiah, 2005).
Salah satu contoh dari organisme tanah yang menguntungkan adalah
mikoriza. Keberadaan mikoriza mampu memberi manfaat bagi tanah dan tanaman
di atasnya. Fungi mikoriza merupakan fungi obligat, dimana kelangsungan hidup
sporanya berasosiasi dengan akar tanaman. Asosiasi akar dengan mikoriza dibagi
menjadi dua berdasarkan pertumbuhan hifanya, yaitu ektomikoriza dan
endomikoriza. Ektomikoriza adalah asosiasi antara fungi dan akar tanaman
dimana fungi pelindung melakukan penetrasi pada lapisan-lapisan dinding sel
hanya pada bagian luarnya, sedangkan fungi endomikoriza yaitu hifa fungi
melakukan penetrasi hingga ke bagian dalam sel-sel inangnya
(Yulipriyanto, 2010).
Peranan mikoriza secara spesifik dalam membantu tanaman antara lain
dalam memperbaiki nutrisi tanaman dengan meningkatkan penyerapan posfat,
Keberadaan mikoriza sangat bermanfaat dalam penyerapan air dan unsur hara
terutama posfor (Smith dan Read, 1997). Akar bermikoriza ternyata juga
meningkatkan penyerapan seng dan sulfur dari dalam tanah lebih cepat dari pada
tanaman tidak bermikoriza.
Faktor lingkungan dan faktor biotik diketahui sangat mempengaruhi
produksi spora, derajat kolonisasinya terhadap akar tanaman, maupun banyaknya
kolonisasi mikoriza yang terjadi pada suatu lahan. Safir dan Duniway (1988)
menyatakan bahwa sebaran mikoriza dipengaruhi oleh banyak faktor, antara lain
jenis dan struktur tanah, unsur hara P dan N dalam tanah, air, pH, dan suhu tanah.
Banyak studi yang menunjukkan bahwa perkembangan FMA dipengaruhi
oleh musim atau bersifat musiman. Gay et al. (1982) mempelajari pengaruh
musim terhadap kolonisasi FMA pada Abroniaumbellata dan
Camissoniacalifornica. Hasilnya menunjukkan kolonisasi FMA berkurang
selama atau sesudah periode pembungaan, yaitu antara April – Juni untuk
Abroniaumbellata dan Mei – Nopember untuk Camissoniacalifornica.
Setiap lahan memiliki sifat yang spesifik dan ditumbuhi oleh keragaman
jenis tanaman yang berbeda, sehingga FMA lokal yang ditemukan akan berbeda
juga. Keberadaan dan pemanfaatan hasil simbiosis FMA dengan tanaman sudah
banyak dipelajari, namun penelitian yang dilakukan untuk mengetahui keberadaan
dan status FMA pada suatu lahan terkait perbedaan lingkungan masih sedikit.
Sementara semakin kita memerlukan peranan FMA untuk meningkatkan
produktivitas lahan semakin perlu kita harus mempelajari ekologi FMA dan
faktor-faktor yang mempengaruhi kolonisasi dan sporulasinya (Delvian, 2006).
mengenai keberadaan dan status FMA berkaitan dengan perbedaan ekologi tempat
tumbuh.
Tujuan Penelitian
Tujuan dari penelitian ini adalah untuk mempelajari keberadaan dan status
FMA berkaitan dengan perbedaan ketinggian tempat tumbuh. Diduga adanya
perbedaan ekologi tempat tumbuh akan mempengaruhi keberadaan dan status
FMA pada tanah.
Hipotesis
Keberadaan dan status FMA berbeda antara dataran tinggi dan dataran rendah.
Manfaat Penelitian
Hasil penelitian ini diharapkan:
1. Memberi informasi mengenai faktor-faktor yang mempengaruhi
keberadaan dan status FMA di dalam tanah.
2. Memberi informasi mengenai keberadaan dan status FMA pada berbagai
TINJAUAN PUSTAKA
Fungi Mikoriza Arbuskula
Mikoriza adalah simbiosis mutualistik, hubungan antara fungi dan akar
tanaman. Beberapa fungi membentuk mantel yang melindungi akar,
kadang-kadang berambut, berwarna keputihan. Akar-akar tanaman mengantarkan
bahan-bahan ke fungi (sebagian sebagai eksudat-oksidat), dan fungi membantu
meneruskan nutrisi-nutrisi dan air ke akar tanaman. Hifa fungi keluar dari
perakaran tanaman hingga mencapai tanah dan membantu menyerap beberapa
unsur hara tertentu untuk selanjutnya ditransmisikan ke tanaman, terutama
hara-hara yang tidak mobil seperti posfat(P), seng(Zn), tembaga(Cu), dan
molibdat(Mo) (Yulipriyanto, 2010).
FMA merupakan asosiasi simbiotik yang terbentuk antara spesies tanaman
dalam skala luas termasuk angiosperm, gymnosperm, pteridophyta, dan beberapa
bryophyte, dan skala fungi terbatas termasuk dalam ordo tunggal, Glomales.
Simbiosis terjadi dalam akar tanaman dimana fungi mengkolonisasi apoplast dan
sel korteks untuk memperoleh karbon dalam tanaman.
Kontribusi FMA pada peristiwa simbiosis sangat kompleks, tetapi aspek
utama meliputi transfer nutrient mineral, khususnya posfat dari tanah ke tanaman.
Perkembangan asosiasi yang sangat cocok ini memerlukan koordinasi molekuler
dan differensiasi selular dari kedua simbion untuk membentuk suatu sistem
dimana transfer nutrient terjadi dua arah (Delvian, 2006).
Spora FMA dalam tanah dapat saja berkecambah secara spontan jika
lingkungannya mendukung akan tetapi hifa akan sangat terbatas pertumbuhannya
maka hifa akan membentuk percabangan yang banyak dan ini menjadi penanda
dimulainya fase awal simbiosis atau dikenal dengan fase presimbiotik
(Giovannetti et al. 1993).
Tanaman yang ketergantungan akan unsur fosfat tinggi akan cenderung
berasosiasi dengan mikoriza. Cahaya dan temperatur merupakan unsur iklim yang
sangat mempengaruhi proses kolonisasi mikoriza arbuskula. Temperatur optimum
bagi perkembangan spora Gigaspora spp. adalah 34oC, sedang untuk Glomus spp.
adalah 20oC. Sedangkan faktor tanah yang berpengaruh adalah keasaman tanah
(pH) dan kandungan unsur hara terutama P dan N. Menurut Hudson (1986),
kandungan unsur hara di dalam tanah sangat mempengaruhi pertumbuhan
mikoriza arbuskula.
Peranan FMA dalam Ekosistem
Manfaat mikoriza dalam ekosistem sangat penting, yaitu berperan dalam
siklus hara, memperbaiki struktur tanah dan menyalurkan karbohidrat dari akar
tanaman ke organisme tanah yang lain (Brundrett et al. 1996). Mikoriza dapat
membebaskan P yang tidak tersedia bagi tanaman, misalnya dalam batuan fosfat,
menjadi tersedia bagi tanaman. Mikoriza mengeluarkan enzim fosfatase dan asam
asam organik, khususnya oxalat, yang dapat membantu membebaskan fosfat.
Peran ini sangat penting mengingat sebagian besar tanah-tanah di
Indonesia bersifat asam, dimana fosfat diikat oleh Al dan Fe. Pada tanah-tanah
kapur, fosfat diikat oleh Ca sehingga tidak tersedia bagi tanaman. Disamping
membebaskan fosfat yang tidak tersedia, hifa mikoriza juga mengkonservasi
unsur hara agar tidak hilang dari ekosistem. Manfaat mikoriza secara langsung
membentuk tubuh buah yang mudah dikenali. Tubuh buah dari fungi
ektomikoriza ini dapat dijadikan sebagai bahan pangan (Scleroderma
sinnamariense yang bersimbiosis dengan melinjo), bahan obat, untuk keindahan
(tubuh buah fungi ektomikoriza beraneka bentuk, ukuran dan warna).
Keanekaragaman fungi juga dapat dijadikan indikator kualitas lingkungan
(Brundrett et al. 1996).
Faktor yang Mempengaruhi Kolonisasi dan Pembentukan Spora FMA
Keberadaan dan kolonisasi dipengaruhi oleh beberapa faktor lingkungan,
faktor-faktor tersebut antara lain:
1.Cahaya dan Fotoperiodesitas
Intensitas cahaya dan panjang hari yang lama akan memperbaiki
kolonisasi dan produksi spora pada Pueraria javanica, jagung dan lain-lain
(Graham et al. 1987). Meningkatnya kolonisasi FMA adalah akibat meningkatnya
proses fotosintesis yang berakibat pada meningkatnya konsentrasi karbohindrat di
dalam akar atau meningkatnya senyawa-senyawa eksudat. Untuk
memaksimumkan produksi inokulum FMA perlu memaksimumkan fotosintesis
inang dan cahaya.
Adanya naungan yang berlebihan terutama untuk tanaman yang senang
cahaya dapat mengurangi kolonisasi akar dan produksi spora, selain itu respon
tanaman terhadap fungi mikoriza akan berkurang. Hal ini disebabkan adanya
hambatan pertumbuhan dan perkembangan internal hifa dalam akar yang
2. Suhu
Suhu berpengaruh terhadap kolonisasi yakni pada perkembangan spora,
penetrasi hifa pada sel akar dan perkembangan pada korteks akar, selain itu suhu
juga berpengaruh pada ketahanan dan simbiosis. Semakin tinggi suhu semakin
besar terbentuknya kolonisasi dan meningkatnya produksi spora. Schenk dan
Schroder (1974) menyatakan bahwa suhu terbaik untuk perkembangan arbuskula
yakni pada suhu 30oC tetapi untuk koloni miselia terbaik berada pada suhu 28–
34oC, sedangkan perkembangan bagi vesikula pada suhu 35oC.
3. Kandungan air tanah
Kandungan air tanah dapat berpengaruh baik secara langsung atau tidak
langsung terhadap kolonisasi dan pertumbuhan fungi mikoriza. Pengaruh secara
langsung tanaman bermikoriza dapat memperbaiki dan meningkatkan kapasitas
serapan air. Sedangkan pengaruh tidak langsung karena adanya miselia eksternal
menyebabkan fungi mikoriza efektif dalam mengagregasi butir-butir tanah,
kemampuan tanah menyerap air meningkat. Penjenuhan air tanah yang lama
berpotensi mengurangi pertumbuhan dan kolonisasi fungi mikoriza karena kondisi
yang anaerob. Daniels dan Trappe (1980) menggunakan Glomus epigaeum
dikecambahkan pada lempung berdebu pada berbagai kandungan air. Glomus
epigaeum ternyata berkecambah paling baik pada kandungan air di antara
kapasitas lapang dan kandungan air jenuh.
4. Kemasaman Tanah
Fungi mikoriza pada umumnya lebih tahan terhadap perubahan pH tanah.
Meskipun demikian adaptasi masing-masing spesies fungi mikoriza terhadap pH
perkembangan dan peran mikoriza terhadap pertumbuhan tanaman
(Maas dan Nieman, 1978).
Perkembangan fungi mikoriza pada pH optimum berbeda-beda tergantung
pada adaptasi fungi mikoriza terhadap lingkungan. Aktivitas enzim yang
berperan dalam perkecambahan spora fungi mikoriza dapat dipengaruhi oleh pH.
Misalnya Glomus mosseae biasanya pada tanah alkali dapat berkecambah dengan
baik pada air atau pada soil extract agar pada pH 6-9. Spora Gigaspora
coralloidea dan Gigaspora heterogama dari jenis yang lebih tahan asam dapat
berkecambah dengan baik pada pH 4-6. Glomus epigaeum perkecambahannya
lebih baik pada pH 6-8.
5.Bahan organik
Bahan organik merupakan salah satu komponen dalam tanah yang penting
disamping air dan udara. Jumlah spora FMA berhubungan erat dengan kandungan
bahan organik dalam tanah. Jumlah maksimum spora ditemukan pada tanah-tanah
yang mengandung bahan organik 1-2% sedangkan pada tanah-tanah berbahan
organik kurang dari 0,5% kandungan spora sangat rendah (Pujiyanto, 2001).
6. Logam berat dan unsur lain
Adanya logam berat dalam larutan tanah dapat mempengaruhi
perkembangan mikoriza. Beberapa spesies mikoriza arbuskula diketahui mampu
beradaptasi dengan tanah yang tercemar seng (Zn), tetapi sebagian besar spesies
mikoriza peka terhadap kandungan Zn yang tinggi. Pada beberapa penelitian lain
diketahui pula strain-strain fungi mikoriza tertentu toleran terhadap kandungan
Sebaran dan Ekologi Fungi Mikoriza
Sebaran dan ekologi mikoriza arbuskula terdapat pada hampir pada semua
jenis tanaman. Mikoriza berasosiasi pada akar tanaman angiosperma,
pterydophyta, bryophyta dan beberapa Gymnospermae. Hanya terdapat beberapa
saja tumbuhan yang tidak bermikoriza terutama tumbuhan yang hanya
membentuk Ektomikoriza misalnya Pinnaceae (Imas et al, 1989).
Meyer (1973) dalam Setiadi (1989) menambahkan mikoriza arbuskula ini
mempunyai penyebaran yang luas, meliputi hutan hujan rapat, padang pasir, semi
gurun dan jarang ditemukan dalam hutan temperate areal yang amat basah
(didominasi oleh Ektomikoriza). Perbedaan lokasi, ekosistem, dan rizosfer
ternyata menunjukan keanekaragaman spesies dan populasi fungi mikoriza,
misalnya yang didominasi oleh fraksi lempung berdebu merupakan tanah yang
baik bagi perkembangan Glomus (Baon dan Widiastuti, 1997), begitu juga dengan
tanah mangrove yang bercirikan tanah berlumpur dan cenderung liat hanya
Glomus sp. yang dapat hidup, sedangkan tanah yang berpasir genus Acaulospora
dan Gigaspora ditemukan dalam jumlah yang tinggi.
Kelapa Sawit Sebagai Inang FMA
Tanaman kelapa sawit (Elaeis guineensis Jack) diketahui berasal dari
kawasan Afrika utara, tepatnya di Nigeria (Fauzi, et al, 2002). Menurut
Sastrowardoyo (2004) tanaman kelapa sawit dapat diklasifikasikan dalam:
Kingdom: Spermatophyta
Subdevisi: Angiospermae
Kelas: Monocotyledonae
Familli: Palmaceae
Genus: Elaeis
Species: Elaeis guineensis jack
Tanaman kelapa sawit tumbuh optimal pada ketinggian tempat 0-100
mdpl. Pertumbuhan dan produksi terbaik kelapa sawit diperoleh pada lahan
dengan ketinggian 0-100 mdpl. Kelapa sawit dapat tumbuh pada berbagai jenis
tanah, tetapi pertumbuhan optimal akan tercapai jika jenis tanahnya sesuai dengan
syarat tumbuh kelapa sawit. Sifat fisika dan kimia tanah yang harus dipenuhi
untuk pertumbuhan kelapa sawit secara optimal diantaranya harus memiliki
ketebalan tanah lebih dari 57 cm dan tidak berbatu agar perkembangan akar tidak
terganggu, tekstur ringan dan terbaik (memiliki pasir 20%-60%, debu 10%-40%,
dan liat 20%-50%), drainase baik dan permukaan air tanah cukup dalam, dan
kemasaman (pH) tanah 4,0-6,0 dan pH optimal 5,0-5,5.
Curah hujan optimal rata-rata tahunan untuk kelapa sawit berkisar
2000-2500 mm pertahun. Distribusi hujan idealnya merata sepanjang tahun tanpa bulan
kering yang berkepanjangan. Lama penyinaran matahari yang dibutuhkan kelapa
sawit minimum 1600 jam/tahun dan optimum sekitar 6-7 jam/hari. Kelembaban
udara yang optimum untuk pertumbuhan kelapa sawit sekitar 80%
(Sastrowardoyo, 2004).
Tanaman kelapa sawit memerlukan temperatur udara yang optimal sekitar
24-280C untuk tumbuh dengan baik. Namun, tanaman masih dapat tumbuh pada
suhu terendah 180C dan tertinggi 32 0C. Tanaman kelapa sawit cenderung tahan
angin, namun sebaiknya kecepatan angin rata-rata tidak melebihi 40 km/jam.
Tanaman kelapa sawit memiliki respon yang sangat baik terhadap kondisi
lingkungan dan perlakuan yang diberikan. Kondisi iklim dan tanah merupakan
faktor utama yang menentukan produktivitasnya, disamping pemberian faktor lain
seperti pemberian organisme seperti mikoriza (Lubis, 1992).
Berdasarkan penelitian yang dilakukan Widiastuti dan Goenadi (2000)
ditemukan bahwa pada perakaran kelapa sawit dijumpai beberapa organ FMA
seperti hifa internal, hifa eksternal, arbuskula, dan vesicular. Organ yang paling
jarang dijumpai adalah arbuskula. Hal ini menunjukkan bahwa FMA dapat
mengkolonisasi akar kelapa sawit dan selanjutnya membentuk organ-organ secara
METODE PENELITIAN
Waktu dan Tempat penelitian
Penelitian ini dilaksanakan pada bulan Januari sampai Juli 2014. Kegiatan
penelitian terdiri dari beberapa tahapan kegiatan yang diawali dengan
pengambilan sampel (eksplorasi) lapangan yang dilakukan di dataran tinggi lahan
kelapa sawit di desa Adia Nangka, Kec. Lae Parira, Kabupaten Dairi dan dataran
rendah di lahan kelapa sawit desa Batang Terap, Kec. Perbaungan, Kabupaten
Serdang Bedagai. Pembuatan kultur trapping di Rumah Kaca Fakultas Pertanian,
Universitas Sumatera Utara. Analisis tanah di Laboratorium Riset dan Teknologi
Fakultas Pertanian, dan pengamatan kolonisasi akar dan identifikasi spora FMA di
Laboratorium Biologi Tanah, Program Studi Agrekoteknologi, Fakultas Pertanian,
Universitas Sumatera Utara.
Bahan dan Alat
Bahan yang digunakan dalam penelitian ini adalah contoh tanah dan akar
kelapa sawit (Elaeis guineensis jack) . Pasir sungai sebagi campuran media tanam,
terrabuster guna merangsang pembentukan spora, hyponex merah sebagai sumber
hara tanaman, dan jagung (Zea mays) sebagai inang pada perlakuan
pemerangkapan. Untuk ekstraksi dan identifikasi spora mikoriza digunakan bahan
berupa larutan glukosa 60%, dan larutan Melzer’s sebagai bahan pewarna spora.
Larutan trypan blue untuk bahan proses pewarnaan akar (staining). Larutan KOH
10% untuk mengeluarkan cairan sitoplasma dalam akar, sehingga akar pucat dan
sebagai pengawet. Larutan HCl 2% untuk mempermudah masuknya trypan blue
Alat yang digunakan dalam untuk pengambilan contoh tanah dan akar
tanaman antara lain GPS, meteran, tali plastik, cangkul, kantong plastik, spidol,
dan kertas label. Alat untuk pengamatan di laboratorium adalah saringan 200 mm,
710 μm, 250 μm, dan 53 μm, tabung sentrifuse, cawan petri, pipet tetes,
mikroskop binokuler, mikroskop stereo, kaca preparat, dan kaca penutup. Alat
yang digunakan untuk pemerangkapan di rumah kaca berupa pot (aqua cup), dan
sprayer.
Prosedur Penelitian
1. Pengambilan Contoh Tanah dan Akar
Pengambilan contoh tanah dan akar tanaman dilakukan pada lima titik
dalam satu petak ukur. Petak ukur berukuran 20 x 20 m sebanyak 5 petak dalam
satu lahan. Contoh tanah diambil pada daerah rizosfir atau pada kedalaman 0
sampai 20 cm. Berat tanah yang diambil dari setiap titik dalam satu petak adalah
sebanyak 500 gram secara komposit. Sedangkan contoh akar tanaman diambil
dengan cara memotong akar-akar halus dari anakan dan tumbuhan bawah pada
petak contoh.
2. Analisis Tanah
Sebelum melakukan penelitian, terlebih dahulu dilakukan analisa awal
terhadap kondisi tanah meliputi pH tanah, C-organik, dan P-tersedia untuk
mengetahui sifat tanah.
3. Pembuatan Kultur Trapping.
Teknik trapping yang digunakan mengikuti metoda Brundrett et al.
(1994) dengan menggunakan pot kultur terbuka. Media tanam yang digunakan
dalam pot kultur adalah pot kultur diisi dengan pasir sungai sampai sepertiga
volume pot, kemudian dimasukkan contoh tanah dan terakhir ditutup dengan pasir
sungai sehingga media tanam tersusun atas pasir sungai-contoh tanah-pasir
sungai. Selanjutnya bibit jagung (Zea mays) ditaruh pada lubang tanam yang
sudah diisi dengan pasir sungai, tanah kemudian ditutupi lagi dengan pasir sungai.
Dari setiap contoh tanah dibuat 5 pot kultur. Disamping itu diberikan
penambahan terrabuster guna merangsang pembentukan spora yang lebih baik.
Perlakuan terrabuster diberikan dengan konsentrasi 0,4% (1:250) sebanyak 20 ml
tiap pot. Frekuensi pemberian terrabuster adalah 3 x 1 minggu selama satu bulan
pertama dan 1 x 1 minggu selama 1 bulan kedua. Penambahan terrabuster ini
diharapkan berpengaruh terhadap pembentukan spora fungi mikoriza.
Setelah kultur berumur 8 minggu kegiatan penyiraman dihentikan dengan
tujuan menkondisikan kultur pada keadaan stress kekeringan. Proses pengeringan
ini berlangsung secara perlahan sehingga dapat merangsang pembentukan spora
lebih banyak. Periode pengeringan ini akan berlangsung selama lebih kurang 2
minggu
Pemeliharaan kultur meliputi kegiatan penyiraman, pemberian hara dan
pengendalian hama secara manual. Larutan hara yang digunakan adalah Hyponex
merah (25-5-20) dengan konsentrasi 1 g/l. Pemberian larutan hara dilakuan setiap
minggu sebanyak 20 ml tiap pot kultur.
Pemanenan dilakukan setelah pembentukan spora-spora baru diasumsikan
sudah cukup baik setelah dilakukan stressing selama 2 minggu terhadap tanaman
jumlah spora per 50 g media tanam dan jenis spora. Selanjutnya spora-spora yang
diperoleh dari kultur ini akan diidentifikasi jenisnya.
4. Pengamatan Contoh Tanah dan Akar
a. Ekstraksi Spora
Teknik yang digunakan dalam mengekstraksi spora FMA adalah teknik
tuang – saring dari Pacioni (1992) dan akan dilanjutkan dengan teknik sentrifugasi
dari Brundrett et al. (1996). Prosedur kerja teknik tuang – saring ini, pertama
adalah mencampurkan tanah sampel sebanyak 50 g dengan 200–300 ml air dan
diaduk sampai butiran-butiran tanah hancur. Selanjutnya disaring dalam satu set
saringan dengan ukuran 200 mm, 710 μm, 250 μm, dan 53 μm secara berurutan
dari atas ke bawah. Dari saringan bagian atas disemprot dengan air kran untuk
memudahkan bahan saringan lolos. Kemudian saringan paling atas dilepas dan
saringan kedua kembali disemprot dengan air kran. Setelah saringan kedua
dilepas sejumlah tanah sisa yang tertinggal pada saringan terbawah dipindahkan
ke dalam tabung sentrifuse.
Ekstraksi spora teknik tuang – saring ini kemudian diikuti dengan teknik
sentrifugasi dari Brundrett et al. (1996). Hasil saringan dalam tabung sentrifuse
ditambahkan dengan glukosa 60% yang diletakkan pada bagian bawah dari
larutan tanah dengan menggunakan pipet. Tabung sentrifuse ditutup rapat dan
disentrifuse dengan kecepatan 2500 rpm selama 3 menit. Selanjutnya larutan
supernatan tersebut dituang ke dalam saringan 53 µm, dicuci dengan air mengalir
(air kran) untuk menghilangkan glukosa. Endapan yang tersisa dalam saringan di
binokuler untuk penghitungan kepadatan spora dan pembuatan preparat guna
identifikasi spora FMA yang ada.
Pembuatan preparat spora menggunakan bahan pewarna Melzer’s.
Spora-spora FMA yang diperoleh dari ekstraksi setelah dihitung jumlah diletakkan
dalam larutan Melzer’s. Selanjutnya spora-spora tersebut dipecahkan secara
hati-hati dengan cara menekan kaca penutup preparat menggunakan ujung lidi.
Perubahan warna spora dalam larutan Melzer’s adalah salah satu indikator untuk
menentukan tipe spora yang ada.
b. Kolonisasi FMA pada Akar Tanaman Sampel
Pengamatan kolonisasi FMA pada akar tanaman contoh dilakukan melalui
teknik pewarnaan akar (staining). Metoda yang digunakan untuk pembersihan
dan pewarnaan akar sampel adalah metoda dari Kormanik dan McGraw (1982).
Langkah pertama adalah memilih akar-akar halus dengan diameter 0,5-2,0 mm
(Rajapakse dan Miller Jr., 1992) segar dan dicuci dengan air mengalir hingga
bersih.
Akar sampel dimasukkan ke dalam larutan KOH 10% dan dibiarkan
selama lebih kurang 24 jam sehingga akar akan berwarna putih atau pucat.
Larutan KOH kemudian dibuang dan akar contoh dicuci pada air mengalir selama
5-10 menit. Selanjutnya akar contoh direndam dalam larutan HCl 2% dan
dibiarkan selama satu malam. Larutan HCl 2% kemudian dibuang dengan
mengalirkannya secara perlahan-lahan. Selanjutnya akar sampel direndam dalam
larutan Trypan blue 0,05%. Kemudian larutan Trypan blue dibuang dan diganti
dengan larutan lacto glycerol untuk proses destaining (pengurangan warna).
Penghitungan persentase kolonisasi akar menggunakan metoda panjang
akar terkolonisasi (Giovannetti dan Mosse, 1980). Secara acak diambil
potong-potongan akar yang telah diwarnai dengan panjang ± 1 cm sebanyak 10 potong-potongan
akar dan disusun pada kaca preparat, untuk setiap tanaman sampel dibuat dua
preparat akar. Potongan-potongan akar pada kaca preparat diamati untuk setiap
bidang pandang. Bidang pandang yang menunjukkan tanda-tanda kolonisasi
(terdapat hifa dan atau arbuskula dan atau vesikula) diberi tanda positif (+),
sedangkan yang tidak terdapat tanda-tanda kolonisasi diberi tanda negatif (-).
Derajat/persentase kolonisasi akar dihitung dengan menggunakan rumus:
% ∑∑ %
Variabel Pengamatan
Variabel pengamatan dibedakan menjadi dua kelompok, yaitu variabel
lingkungan dan variabel mikoriza. Variabel lingkungan meliputi (1) kesuburan
tanah (pH, C-organik, P-tersedia), dan (2) komponen iklim (suhu dan curah hujan
rata-rata bulanan). Komponen iklim yang diasumsikan mempunyai pengaruh
paling besar terhadap perubahan musim untuk daerah tropis adalah curah hujan.
Variabel mikoriza yang akan diamati meliputi (1) persentase kolonisasi akar pada
tanaman inang, (2) kepadatan spora atau jumlah spora FMA tiap gram tanah dan
(3) jenis spora FMA yang ditemukan.
Berdasarkan data-data yang diperoleh dari variabel pengamatan dilakukan
analisis untuk melihat hubungan antara variabel lingkungan dengan variabel
mikoriza. Analisis ini dilakukan untuk menjelaskan bagaimana perbedaan
keberadaan dan status FMA dengan adanya perubahan kondisi-kondisi
HASIL DAN PEMBAHASAN
Hasil
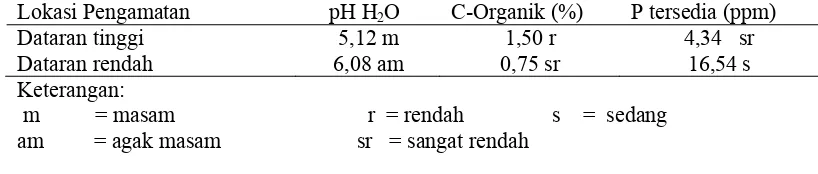
1. Analisis Sifat Kimia Tanah
Sampel tanah yang diambil dari lapangan berasal dari kedalaman 0 – 20
cm. Berdasarkan hasil analisis yang dilakukan ditemukan perbedaan sifat kimia
tanah diantara kedua lokasi penelitian. Persentase C Organik pada dataran tinggi
lebih besar yaitu 1,50 % dibanding dengan kandungan C Organik pada dataran
rendah yang hanya mencapai 0,75%. Lahan dataran rendah memiliki pH H2O
lebih tinggi yaitu 6,50 dibandingkan dengan pH H2O pada lahan dataran tinggi
yaitu 5,12. Kandungan P tersedia pada dataran rendah lebih tinggi yaitu 16,54
ppm dibandingkan dengan P tersedia pada dataran tinggi yang hanya mencapai
4,34 ppm. Hasil analisis sampel tanah yang diamati dapat dlihat pada tabel 1.
Tabel 1. Hasil analisis tanah lahan kelapa sawit dataran tinggi dan dataran rendah serta kriterianya menurut Penelitian Tanah (1983) dalam Mukhlis (2007)
Lokasi Pengamatan pH H2O C-Organik (%) P tersedia (ppm)
Komponen iklim yang diamati dan dijadikan parameter dari lokasi
penelitian adalah data curah hujan dan suhu rata-rata bulanan. Berdasarkan data
iklim yang diperoleh dari Stasiun Klimatologi kelas I Sampali Medan, diketahui
bahwa curah hujan rata-rata pada dataran tinggi dari bulan desember 2013 sampai
januari 2014 adalah 153 mm sementara curah hujan rata-rata dari bulan desember
dataran tinggi pada bulan desember 2013 sampai januari 2014 adalah 20,10C
sementara suhu rata-rata dataran rendah pada bulan desember 2013 sampai januari
2014 adalah 26,20C. Data iklim di kedua lokasi dapat dilihat pada tabel 2.
Tabel 2. Data komponen Iklim pada kedua lokasi pengambilan sampel tanah
Lokasi Pengamatan Curah hujan rata-rata (mm) Suhu rata-rata (0C)
Dataran tinggi 153 mm 20,1
Dataran rendah 207 mm 26,2
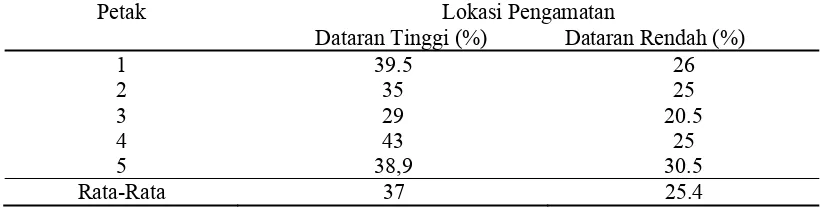
3. Persentase kolonisasi akar
Dilihat dari rataan persentase kolonisasi akar kelapa sawit pada kedua
tempat didapatkan persentase kolonisasi tertiggi adalah pada akar dari dataran
tinggi yang rata-rata kolonisasinya mencapai 37%, sedangkan persentase terendah
adalah kolonisasi akar yang berasal dari dataran rendah yang rata-rata
kolonisasinya hanya mencapai 25,4%. Rata-rata persentase kolonisasi FMA pada
akar kelapa sawit (Elaeis guineensis jack) di dua tempat dapat dilihat pada table 3
berikut.
Tabel 3. Persentase kolonisasi akar kelapa sawit oleh FMA pada dataran tinggi dan dataran rendah.
Struktur yang dibentuk FMA dengan mengkolonisasi akar kelapa sawit
(Elaeis guineensis jack) di kedua tempat yang diamati adalah struktur vesikula
Gambar 1. Akar tanpa kolonisasi FMA
Gambar 2. Vesikula FMA pada akar
Gambar 3. Hifa FMA pada akar
4. Kepadatan Spora Hasil Pemerangkapan di Rumah Kaca
Hasil ekstraksi spora dari 50 gram tanah yang dilakukan menunjukkan
bahwa rata-rata kepadatan spora dalam tanah lahan kelapa sawit dataran tinggi
lebih tinggi dari pada tanah yang berasal dari lahan kelapa sawit dataran rendah.
Rata-rata kepadatan spora yang didapatkan dari dataran tinggi adalah 938
spora/50. Hasil ekstraksi tanah lahan kelapa sawit dataran rendah, rata-rata
kepadatan sporanya hanya mencapai 128 spora/50 gram tanah. Rata-rata
Tabel 4. Kepadatan Spora FMA pada dataran tinggi dan dataran rendah.
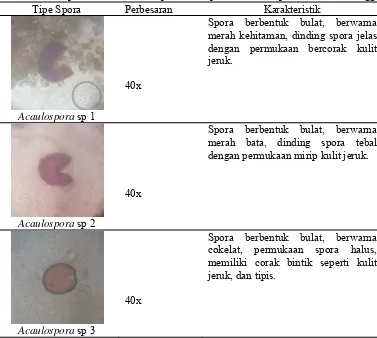
5. Tipe dan Karakteristik Spora
Hasil ekstraksi dan identifikasi terhadap spora FMA dari tanah lahan
kelapa sawit dataran tinggi ditemukan 2 genus spora FMA yaitu Acaulospora
yang terdiri dari 6 tipe spora dan dan Glomus yang ditemukan terdiri dari 14 tipe
spora. Tipe dan karakteristik spora FMA dapat dilihat pada tabel 5.
Tabel 5. Tipe dan karakteristik spora FMA pada lahan kelapa sawit dataran tinggi. Tipe Spora Perbesaran Karakteristik
Acaulospora sp 1
40x
Spora berbentuk bulat, berwarna merah kehitaman, dinding spora jelas dengan permukaan bercorak kulit merah bata, dinding spora tebal dengan permukaan mirip kulit jeruk.
Acaulospora sp 3
40x
Acaulospora sp 4
40x
Spora berbentuk bulat, berwarna hitam, dinding spora tebal dan dengan permukaan seperti kulit jeruk.
Acaulospora sp 5
40x
Spora berbentuk bulat, berwarna merah bata, dinding spora tebal dengan permukaan seperti kulit jeruk.
Acaulospora sp 6
40x
Spora berbentuk bulat, berwarna kuning keemasan, dinding spora tipis dengan permukaan seperti kulit jeruk.
Glomus sp 1
40x
Spora berbentuk bulat, berwarna merah bata, dinding spora tebal dan dengan permukaan kasar.
Glomus sp 2
40x
Spora berbentuk bulat, berwarna hitam, tidak memiliki corak. Dinding spora tebal dan memiliki Hyfal attchment.
Glomus sp 3
40x
Glomus sp 4
40x
Spora berbentuk bulat lonjong, berwarna merah gelap, dinding spora tebal dan dengan permukaan halus.
Glomus sp 5
40x
Spora berbentuk bulat, berwarna merah gelap, dinding spora tebal dengan permukaan halus. merah bata, dinding spora tebal dengan permukaan halus.
Glomus sp 8
40x
Spora berbentuk bulat lonjong, berwarna merah bata terang, dinding spora tebal dengan permukaan berbintik.
Glomus sp 9
40x
Glomus sp 10 dan memiliki Hyfal attchment.
Glomus sp 11
40x
Spora berbentuk bulat, berwarna cokelat, dinding spora tipis dengan permukaan berbintik dan memiliki berwarna merah tua, dinding spora tebal dengan permukaan bertekstur.
Glomus sp 13
40x
Spora berbentuk bulat, berwarna hitam, tidak memiliki corak. Dinding spora tebal dengan permukaan kasar
Glomus sp 14
40x
Spora berbentuk bulat telur, berwarna kuning keemasan tua, dinding spora tidak tebal dengan permukaan berbintik dan memiliki Hyfal attchment.
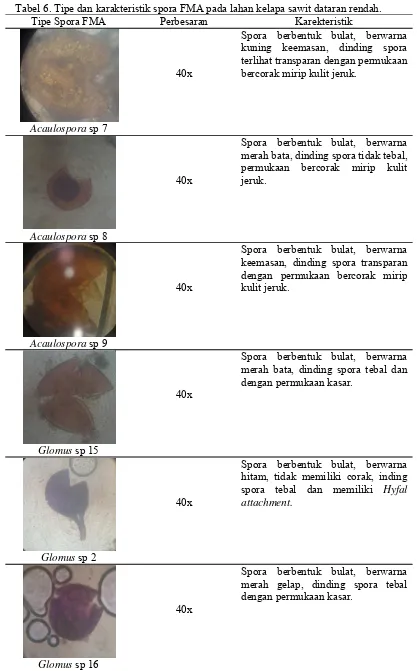
Hasil ekstraksi tanah lahan kelapa sawit dataran rendah terdapat 2 genus
yaitu Acaulospora yang terdiri atas 3 tipe spora dan Glomus yang terdiri dari 7
Tabel 6. Tipe dan karakteristik spora FMA pada lahan kelapa sawit dataran rendah. Tipe Spora FMA Perbesaran Karekteristik
Acaulospora sp 7 merah bata, dinding spora tidak tebal, permukaan bercorak mirip kulit merah bata, dinding spora tebal dan dengan permukaan kasar.
Glomus sp 2
40x
Spora berbentuk bulat, berwarna hitam, tidak memiliki corak, inding spora tebal dan memiliki Hyfal attachment.
Glomus sp 16
40x
Glomus sp 17
40x
Spora berbentuk bulat, berwarna cokelat, dinding spora tebal dengan permukaan kasar.
Glomus sp 18
40x
Spora berbentuk bulat, berwarna kuning keputihan, dinding spora tipis transparan dengan permukaan bercorak.
Glomus sp 13
40x
Spora berbentuk bulat, berwarna hitam, dinding spora tebal dan tidak begitu jelas, permukaan kasar.
Glomus sp 19
40x
Spora berbentuk bulat, berwarna merah tua, dinding spora tidak tebal dengan permukaan berbintik putih.
Pembahasan
Kolonisasi FMA pada akar dicirikan oleh adanya asosiasi antara FMA
dengan akar yang membentuk struktur-struktur yang dihasilkan oleh FMA seperti
hifa, vesikula, arbuskula maupun spora. Hifa adalah salah satu struktur dari FMA
berbentuk benang-benang halus yang berfungsi sebagai penyerap unsur hara dari
luar. Vesikula menurut Abbott dan Robson (1982), berbentuk globosa dan berasal
dari menggelembungnya hifa internal dari FMA. Arbuskula adalah unit kolonisasi
dinding sel serta membentuk sistem percabangan hifa yang kompleks
(Hudson, 1986). Adanya satu atau lebih struktur FMA tersebut, maka dapat
dikatakan terjadi kolonisasi oleh FMA.
Hasil pengamatan di laboratorium terhadap akar kelapa sawit yang
diperoleh dari dataran tinggi dan dataran rendah menunjukkan bahwa tanaman
kelapa sawit di kedua lokasi ini mampu berasosiasi dengan FMA. Namun,
persentase kolonisasi yang dijumpai relatif beragam dari setiap lokasi
pengambilan sampel.
Persentase kolonisasi akar yang terkolonisasi FMA dari kedua tempat
(Tabel 3) menunjukkan perbedaan persentase kolonisi akar yang terjadi pada akar
kelapa sawit di kedua tempat. Rataan kolonisasi akar menunjukkan bahwa akar
kelapa sawit dataran tinggi memiliki persentase kolonisasi lebih tinggi yaitu 37%
dibandingkan dengan akar kelapa sawit dari dataran rendah yaitu 25,4%.
Berdasarkan kriteria persentase akar menurut Setiadi (1992) maka persentase
kolonisasi FMA 37% pada akar kelapa sawit dataran tinggi termasuk dalam
kategori sedang, sementara untuk akar kelapa sawit dataran rendah dengan persen
kolonisasi sebesar 25,4% termasuk dalam kategori rendah.
Intensitas kolonisasi FMA dipengaruhi oleh berbagai macam faktor, baik
faktor lingkungan maupun faktor mikoriza itu sendiri. Penyebaran FMA tidak
pernah merata untuk kondisi lapangan tertentu. Hal ini kemungkinan sebagian
besar disebabkan oleh kesuburan tanah yang berbeda-beda untuk setiap lokasi
pengambilan sampel.
Berdasarkan hasil analisa di laboratorium terhadap sifat kimia tanah
sampel akar. Sifat kimia tanah diketahui sangat mempengaruhi kemampuan FMA
berasosiasi dengan tanaman. Tanah lahan kelapa sawit dataran tinggi memiliki
pH 5,14 artinya lebih rendah dari pH tanah lahan kelapa sawit dataran rendah
yang tingkat kemasaman (pH) tanahnya mencapai 6,08. Menurut Setiadi (1989),
perkembangan FMA yang optimal terjadi pada pH 3,9-5,9. Namun,
perkecambahan masih memungkinkan untuk terjadi dalam kisaran pH yang masih
memungkinkan untuk pertumbuhan. Hal ini sesuai dengan hasil temuan di
lapangan bahwa persentase kolonisasi FMA lebih tinggi ditemukan pada lahan di
dataran tinggi yang memiliki kisaran pH yang lebih optimum dalam
perkembangan FMA.
Hasil analisa laboratorium menunjukkan bahwa pada tanah lahan kelapa
sawit dataran tinggi mengandung C organik 1,50% yang termasuk dalam kriteria
rendah dan tanah lahan kelapa sawit dataran rendah mengandung C organik
0,75% yang termasuk dalam kriteria sangat rendah. FMA sangat dominan berada
pada lapisan bahan organik tanah, oleh karena itu rendahnya kandungan bahan
organik pada tanah akan mempengaruhi keberadaan dan perkembanagan FMA.
Harley et al (1976) menyatakan bahwa 95% FMA aktif terdapat pada lapisan
humus dan kayu-kayu yang busuk.
Kandungan P dalam tanah diketahui dapat mempengaruhi variasi
kolonisasi FMA pada akar tanaman. Hasil analisa tanah terhadap P tersedia dalam
tanah menunjukkan bahwa pada tanah lahan kelapa sawit dataran tinggi P
tersedia yang ditemukan adalah 4,34 ppm yang termasuk dalam kriteria rendah,
berbeda dengan kandungan P tersedia yang ditemukan pada tanah lahan kelapa
yang mengandung unsur P yang banyak sering dihubungkan dengan menurunnya
kolonisasi FMA. Pembentukan simbiosis FMA mencapai maksimum jika kadar P
dalam tanah tidak lebih besar dari 50 mg kg-1 (50 ppm) (Ishii, 2004). Menurut
Setiadi et al (1992) konsentrasi P yang tinggi di dalam tanah menghambat
kolonisasi FMA.
Selain faktor sifat tanah, faktor lingkungan seperti curah hujan dan suhu
dapat pula mempengaruhi variasi kolonisasi FMA pada akar. Berdasarkan data
yang diperoleh dari BMKG Kelas 1 Sampali Medan, diperoleh data curah hujan
dan suhu rata-rata bulanan dari kedua lokasi pengambilan sampel yang disajikan
dalam tabel 2. Setiadi (1989) menyatakan bahwa suhu optimum bagi pertumbuhan
cendawan pembentuk mikoriza beragam menurut jenis dan strain. Pertumbuhan
yang baik antara 20oC-30oC. Artinya, kedua lokasi pengambilan sampel memiliki
suhu yang optimum untuk FMA dapat mengkolonisasi akar tanaman.
Curah hujan yang berkaitan dengan status air tanah juga merupakan faktor
yang mempengaruhi kolonisasi FMA pada akar. Curah hujan rata-rata di dataran
tinggi dari bulan desember 2013 sampai januari 2014 lebih rendah yaitu 153 mm
dibandingkan dengan curah hujan rata-rata dari bulan desember 2013 sampai
januari 2014 di dataran rendah yang mencapai 207 mm. Delvian (2003)
melakukan pengamatan terhadap fluktuasi kolonisasi FMA pada akar beberapa
jenis tanaman dalam 5 periode curah hujan yang berbeda, hasilnya terjadi variasi
kolonisasi FMA ditemukan baik dalam perbedaan tanaman inang maupun
perbedaan curah hujan. Beberapa jenis tanaman menunjukkan respon yang
berbeda dari tanaman lain terhadap satu kondisi curah hujan tertentu. Namun
rendah. Oehl et al (2009) menyatakan bahwa tidak semua FMA aktif pada periode
waktu yang sama. Sebagian FMA jumlahnya melimpah pada musim hujan,
sebagian lainnya pada waktu musim kemarau, dan sebagian lainnya pada
sepanjang tahun.
Selain faktor-faktor di luar FMA, menurut Gunawan (1993), persentase
kolonisasi pada akar dan produksi spora oleh FMA dipengaruhi oleh spesies FMA
itu sendiri, lingkungan dan tanaman inangnya, sehingga baik jumlah spora
maupun persentase kolonisasi akar tidak dipengaruhi oleh satu faktor saja,
melainkan akumulasi dari berbagai faktor yang dapat mempengaruhinya.
Struktur FMA yang dijumpai pada akar kelapa sawit dari dataran tinggi
dan dataran rendah yang telah diwarnai dan diamati adalah hifa (Gambar 3) dan
vesikula (Gambar 2). Struktur FMA berupa arbuskula tidak dijumpai pada akar di
lahan perkebunan kelapa sawit di dataran tinggi dan dataran rendah. Hal ini tidak
berbeda jauh dengan hasil penelitian aplikasi FMA terhadap bibit kelapa sawit
yang dilakukan Widiastuti dan Goenadi (2000) yang menemukan bahwa pada
perakaran kelapa sawit dijumpai organ FMA seperti hifa internal, hifa eksternal,
arbuskula, dan vesikula namun struktur arbuskula sangat jarang dijumpai.
Sieverding (1991) menyatakan bahwa vesikula pada umumnya dibentuk
setelah arbuskula. Struktur arbuskula akan sukar untuk dijumpai karena
keberadaan arbuskula pada akar relatif sangat singkat, yaitu berkisar 1-3 hari saja.
Kemungkinan lain, struktur arbuskula tidak dapat ditemukan karena akar yang
diambil terlalu muda sehingga struktur arbuskula belum terbentuk atau akar sudah
tidak ditemukan juga karena sampel akar yang diambil belum dapat mewakili
keseluruhan akar inang yang terkolonisasi oleh FMA.
Selain pengamatan terhadap kolonisai FMA pada akar dilakukan juga
penghitungan kepadatan spora hasil pemerangkapan di rumah kaca. Hasil
pengamatan di laboratorium terhadap spora dari tanah hasil pemerangkapan di
rumah kaca menunjukkan bahwa kepadatan spora bervariasi terhadap lokasi yang
dijadikan sumber tanah pemerangkapan (Tabel 4). Hasil penghitungan kepadatan
spora menunjukkan kepadatan spora hasil pemerangkapan sangat tinggi.
Pada tanah asal lahan kelapa sawit dataran rendah rata-rata kepadatan
spora yang diperoleh adalah 128 spora/50 gram tanah, sementara pada tanah lahan
kelapa sawit dataran tinggi kepadatan spora adalah 938 spora/50 gram tanah.
Kepadatan spora yang bervariasi ini tidak berbeda jauh dengan hasil penelitian
yang dilakukan oleh Puspitasari (2005) yang melakukan pemerangkapan FMA
dari tanah berbagai salinitas di hutan pantai Ujung Genteng Sukabumi dengan
Shorgum sp sebagai tanaman inang. Pada penelitian ini diperoleh variasi
kepadatan spora yang sangat beragam antara lain dibawah 10 spora/50 gram tanah
sampai kepada kerapatan diatas 700 spora/50 gram tanah.
Kultur pemerangkapan pada dasarnya digunakan untuk menstimulasi
sporulasi atau meningkatkan jumlah propagul FMA yang ada di dalam tanah yang
diambil dari lapangan. Hal itu perlu dilakukan, mengingat tidak semua FMA aktif
pada periode waktu yang sama. Sebagian FMA jumlahnya melimpah pada musim
hujan, sebagian lainnya pada sepanjang tahun (Oehl et al, 2009). Tingginya
pemerangkapan yang diberikan berpengaruh positif terhadap perkecambahan dan
perkembangan FMA.
Penyebaran genus FMA dari pengamatan terhadap hasil pemerangkapan
mempunyai variasi sebaran genus yang sama di kedua tempat pengambilan
sampel tanah. Berdasarkan hasil yang diperoleh diketahui bahwa tipe spora yang
ditemukan adalah genus Glomus dan Acaulospora. Pada tanah lahan kelapa sawit
dataran tinggi ditemukan 6 tipe spora dari genus Acaulospora dan 14 tipe spora
dari genus Glomus (Tabel 5). Sementara pada lahan kelapa sawit dataran rendah
ditemukan 3 tipe spora genus Acaulospora dan 7 tipe spora dari genus Glomus
(Tabel 6). Sebaran genus FMA yang dijumpai pada lahan kelapa sawit kedua
tempat tidak berbeda jauh dengan hasil temuan Sibarani (2011) yang
mengidentifikasi tipe spora FMA pada lahan kelapa sawit ekosistem lahan gambut
desa Telaga Suka, Kecamatan Panai Tengah Kabupaten Labuhan Batu yang juga
hanya menemukan 2 genus FMA yaitu Acaulospora dan Glomus.
Genus Acaulospora yang ditemukan dari kedua lokasi pengamatan
umumnya berbentuk bulat berwarna kemerahan, cokelat, putih, sampai pada
warna hitam. Spora jelas menampakkan dinding sel yang berjumlah lebih dari satu
lapis, dan memiliki bintik-bintik atau permukaan mirip kulit jeruk. Berdasarkan
hasil pengamatan terhadap tipe spora Acaulospora dari kedua lokasi pengamatan
tidak ditemukan tipe spora yang sama-sama dapat dijumpai di kedua tempat.
Masing-masing lokasi menunjukkan tipe spora Acaulospora yang berbeda.
Menurut Hall (1984) spora Acoulospora merupakan spora tunggal di
dalam sporokarp, spora melekat secara lateral pada hifa yang ujungnya
globos, subglobos, ellips atau fusiform melebar. Spora Acaulospora yang
ditemukan memiliki bentuk bulat lonjong dan memiliki dinding spora yang relatif
tebal. Proses perkembangan Acaulospora berawal dari ujung hifa (subtending
hyphae) yang membesar seperti spora yang disebut hyphal terminus. Di antara
hyphal terminus dan subtending hypae akan muncul bulatan kecil yang semakin
lama semakin membesar dan terbentuk spora.
Spora Glomus yang ditemukan dari kedua tempat pada umumnya
berbentuk bulat sempurna, bulat telur sampai bulat lonjong dengan ukuran yang
bervariasi. Spora FMA yang dijumpai berwarna kemerahan, cokelat, kuning,
putih, sampai kehitaman dengan dinding tebal sampai transparan, dan permukaan
halus sampai kasar. Beberapa tipe spora Glomus ditemukan memiliki tangkai
spora (Hyfal attcment). Berdasarkan hasil identifikasi terhadap tipe spora Glomus
dari kedua tempat, ditemukan bahwa pada lahan kelapa sawit dataran tinggi dan
lahan kelapa sawit dataran rendah terdapat dua tipe spora yang sama-sama dapat
dijumpai di kedua tempat. Tipe spora yang dapat sama-sama dijumpai di kedua
tempat adalah tipe spora Glomus sp 2 yang memiliki ciri bentuk bulat, berwarna
hitam, tidak memiliki corak, dinding spora tebal dan memiliki tangkai spora
(Hyfal attcment), dan yang kedua adalah tipe spora Glomus sp 13 dengan spora
berbentuk bulat, berwarna hitam, dinding spora tebal yang tidak begitu jelas, dan
permukaan kasar.
Proses perkembangan spora genus Glomus sp adalah dari ujung hifa yang
membesar sampai ukuran maksimal dan terbentuk spora. Karena sporanya berasal
dari perkembangan hifa maka disebut chlamydospora, kadang hifa
Pada saat dewasa spora dipisahkan dari hifa pelekat oleh sebuah sekat, spora
bentuk globos, subglobos, ovoid ataupun obovoid (Puspitasari, 2012). Menurut
Smith dan Read (1997) spora genus Glomus dapat ditemukan dalam bentuk
tunggal atau agregat lepas, sporokarp tidak seperti pada Sclerocystis dan
sporokarp terdiri dari spora dengan dinding lateral yang saling melekat satu sama
lainnya
Sebaran genus Acaulospora dan Glomus dalam penelitian ini belum bisa
diidentifikasi secara akurat tentang penyebaran dan nama spesiesnya, karena dari
seluruh jumlah spora yang ditemukan hanya sedikit yang dapat diidentifikasi.
Kondisi ini dikarenakan banyak ditemukan spora-spora yang rusak dan kotor
belum terpisah dengan tanah. Proses identifikasi spora juga terkendala oleh
terbatasnya peralatan di laboratorium dalam proses identifikasi sehingga
KESIMPULAN DAN SARAN
Kesimpulan
1. Sebaran keberadaan FMA lebih tinggi pada lahan kelapa sawit dataran tinggi
dibandingkan dengan lahan kelapa sawit dataran rendah.
2. Rataan kolonisasi akar kelapa sawit di dataran tinggi lebih tinggi
dibandingkan dengan rataan kolonisasi akar kelapa sawit di dataran rendah
Saran
Hasil penelitian ini hanya mendapatkan data keanekaragaman FMA dan
statusnya pada tanaman inang kelapa sawit pada satu musim pengamatan saja,
maka perlu dilakukan penelitian lanjutan untuk mengetahui potensi FMA yang
DAFTAR PUSTAKA
Abbott, L.K. dan Robson, A.D. 1991. The Effect of Mycorrizha on Plant Growth. CRC Press, Inc., Boca Raton. Florida.
Bagyaraj, D. J. 1984. Biological interaksi With VA Michorrizal fungi; Powell CL dan Bagyraj DJ. (Eds). Vesicular-arbuscular Micorriza. CRC Press. Inc. Boca Raton. Florida.
Baon, J. B. dan Widiastuti A 1997. Spore Production of VA Mycorrhizal Fungus, Acaulospora tuberculata, As Affected By Host Species Dan Nutrients Source. Procedings of International Conference on Mycorrhizal in Sustainable Tropical Agriculture dan Forest Ecosystems. Bogor.
Brundrett, M., N. Bougher, B. Dell, T. Grave dan N. Malajezuk. 1996. Working with Mycorrihiza in Forestry dan Agriculture. Australian Centre for InternationalAgricultural Research (ACIAR). Carbera.
Daniels, B. A. H. dan Trappe JM 1980 Factors affecting spora germination of the VAM fungus, Glomus epigaeus. Mycology.
Delvian, 2003. Studi Keanekaragaman dan Potensi Pemanfaatan Cendawan Mikoriza Arbuskula (CMA) di Hutan Pantai. Disertasi Program Pasca Sarjana IPB. Bogor.
Delvian. 2006. Peranan Ekologi dan Agronomi Cendawan mikoriza. USU Repositoty. Medan.
Delvian. 2006. Dinamaika Sporulasi Cendawan Mikoriza Arbuskula. USU Repository. Medan.
Fauzi, Y, et al. 2002. Kelapa Sawit: Budidaya Pemanfaatan Hasil dan Limbah, Analisis Usaha dan pemasaran. Penebar Swadaya. Jakarta.
Gay Pe, Grubb P. J. dan Hudson H. J. 1982. Why are someplants more mycorrizal than others An ecological enquiri. Di dalam: Read DJ, Lewis DH, Fitter AH, dan Alexdaner IJ(Eds). Mycorrizas in ecosystems. C. A. B. International.
Graham, W. H. A. Van der As, AJ. Van der Plants-Nitterik RA. Samson dan J.A.Stalpers.1987. CBS Course of Mycology.Centraalbureau voor Schimmelcultures,Baam-Delft.
Giovanneti, M., Avio L, Sbrana C, Citernesi AS dan Logi C. 1993. Differential hypha morphogenesis in arbucular mycorrhizal fungi during pre-infetion stages. New Phytol.
Gunawan AW. 1993. Mikoriza Arbuskula. Pusat Antar Universitas Ilmu Hayat. Institut Pertanian Bogor. Bogor.
Hall IR. 1984. Taxonomy of VA mycorrhizal fungi. In: VA mycorrhizal (ed.) C.L. Powell dan D.J. Bagyaraj. CRC. Press. Boca Raton Florida USA.
Hanafiah, et al. 2003. Biologi Tanah. Ekologi dan Makrobiologi tanah. PT. Raja Grafindo Persada. Jakarta.
Harley JL. 1969. The Biology of Mycorrhiza. Second Edition. Leonard Hill. London.
Hudson, H. J. 1986. Fungal Biology. Departemen Kehutanan. Jakarta.
Imas. T. R. S. Hadioetomo, AW, Gunawan. dan Setiadi Y. 1989. Mikrobiologi Tanah. Jilid II. Departemen Pendidikan dan Kebudayaan. Direktorat jendral Tinggi Pusat Antar Universitas Bioteknologi IPB. Bogor.
Indriyanto. 2006. Ekologi Hutan. Penerbit Bumi Aksara. Jakarta.
Ishii, T. 2004. Vasicular Arbuscular (VA) Mycorrizae. http://www.bio.kpu.ac.jp/pomlab /VANinf.html. (diakses pada Juni 2014)
Janouskova, M; Pavlikova D; Vosatka M. 2006. Potensial contribution of arbuscular mycorrhiza to cadmium immobili sation in soil. Chemosphere.
Kormanik P. P. dan McGraw AC. 1982. Quantification of VA mycorrhizae in plant root. Di Dalam : N.C.Schenk (Ed.) Methods dan principles of mycorrhizae research. The American Phytop.
Lubis, A. U. 1992. Kelapa Sawit (Elaeis guineensis Jacq) di Indonesia. Pusat Penelitian Perkebunan Mariha. Bdanar Kuala.
Maas, E. V. dan R. H. Nieman. 1978. Physiology of plant tolerance to salinity. Dalam G. A. Jung (Ed). Crop tolerance to suboptimal ldan conditions. ASA Spec. Pub.
Mosse, B. 1981. Vesicular-Arbuscular Mycorrhizal. Research of Tropical Agroculture. Research Bulletin Hawaii Institute of Tropical Agriculture dan Human Resources. Hawaii.
Oehl, F., Sieverding E, Inechen K, Mader P, Wiemken A, Boller T. 2009. Districh sporulation synamics of mycorrhizal fungal communities from different agroecosystem in long-term microcosms. Agric Ecosyst Environm 134
Pujiyanto. 2001. Pemanfatan Jasad Mikro, Jamur Mikoriza dan Bakteri dalam Sistem Pertanian Berkelanjutan di Indonesia: Tinjauan Dari Perspektif Falsafah Sains. Makalah Falsafah Sains Program Pasca Sarjana Institut Pertanian Bogor. Bogor.
Puspitasari, D., Kristanti I P., Anton M. 2012. Eksplorasi Vesicular Arbuscular Mycorrhiza (VAM) Indigenous pada Lahan Jagung di Desa Torjun, Sampang Madura. Jurnal Sain dan Seni ITS Vol 1.
Puspitasari, R. T. 2005. Keanekaragaman Cendawan Mikoriza Arbuskula di hutan pantai Ujung Genteng, Sukabumi-Jawa Barat. Tesis Program pasca Sarjana IPB. Bogor.
Safir, G. R. dan J. M. Duniway. 1988. “Evaluation of plant response to colonizayion by vesicular-arbuscular mychorrhizal fungi”. Environmental variables. The American Phytopathological. USA.
Sastrowardoyo, S. 2004. Budidaya Kelapa Sawit. Agromedia pustaka. Jakarta
Schenck, N. C. dan Schroder VN 1974. Temperature response of endogone micorrhiza on soybean roots. Mycologia.
Setiadi, Y. 1989. Pemanfaatan Mikroorganisme dalam Kehutanan. Departemen Pendidikan dan Kebudayaan. Direktorat jenderal Pendidikan Tinggi Pusat Antar Universitas Bioteknologi IPB. Bogor.
Setiadi Y. 1992. Mikoriza dan Pertumbuhan Tanaman. Departemen Pendidikan dan Kebudayaan, Direktorat Jenderal Pendidikan Tinggi Pusat Antar Universitas IPB. Bogor.
Setiadi, Y. 2001. Peranan mikoriza arbuskula dalam reboisasi lahan kritis di Indonesia. Makalah seminar penggunaan CMA dalam sistem pertanian organik dan rehabilitas lahan. Bandung.
Sibarani, Danrianus S. 2011. Keanekaragaman Fungi Mikoriza Arbuskula di Padad Tegakan Karet dan Tegakan Sawit di Ekosistem Lahan Gambut. Skripsi Program Studi Kehutanan USU. Medan.
Sieverding, E. 1991. Vesicular Arbuscular Mycorrizha Management in Tropical Agrosystem. Deutche Gessellsschaft fur Tecnosche Zusmmenourheit (GTZ) Gmbh. Federal Republic Germany.
Widiastuti dan D. H. Goenadi. 2000. Prospek Penggunaan Cendawan Mikoriza Arbuskula Untuk Pembangunan Perkebunan Kelapa Sawit. Posiding seminar Nasional mikoriza 1. Pusat penelitidanan Pengembangan hutan dan Konservasi Alam, Badan Pengembangan Kehutanan dan Perkebunan. Departemen Kehutanan dan Perkebunan.
Yayasan Ekosistem Leuser. 2013. Petunjuk Praktis Budidaya Kelapa Sawit Ramah Lingkungan Untuk Petani Kecil. Universitas Syah Kuala. Aceh.
Lampiran 1. Pengambilan Sampel Tanah dan Akar
Lampiran 3. Pengamatan di Laboratorium