DAN STRATEGI PEMANFAATANNYA SECARA
BERKELANJUTAN
ARON BATUBARA
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
BOGOR
Dengan ini saya menyatakan bahwa disertasi yang berjudul “Studi Keragaman Fenotipik dan Genetik Beberapa Sub Populasi Kambing Lokal Indonesia dan Strategi Pemanfaatannya Secara Berkelanjutan” adalah karya saya dengan arahan Komisi Pembimbing, dan belum diajukan dalam bentuk apapun kepada perguruan tinggi manapun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun yang tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir disertasi ini.
Bogor, … …Juli 2011
ARON BATUBARA. Study of Phenotypics and Genetics Diversity of Some Indonesian Local Goat Sub Populations and Its Strategies for Sustainable Use. Supervised by RONNY RACHMAN NOOR, ACHMAD FARAJALLAH and BESS TIESNAMURTI.
The genetic resources of local goats are very important because it was well adapted to the local environment with low input management systems. Local goats are very potential for breeding stock program which suitable to the Indonesia local conditions. The study was carried out in order to describe the phenotypic and genetic of 6 sub-populations of Indonesian local goats based on the analysis of the morphometrics, mitochondrial DNA, Y Chromosome and GDF9 gene. The sampling was conducted in four of the Provinces in Indonesia, namely North Sumatra Province (Samosir goat n=42 in the Samosir District, Muara goat n=34 in the North Tapanuli district, Kacang goat n=217 in the Deli Serdang District), Central Java Province (Jawarandu goat n=94 in the Blora District); Province of South Sulawesi (Marica goat n=60 in the Maros District, Makassar City, Jeneponto District) and Nusa Tenggara Timur Province (Bengala goat n=96 in the Kupang area, Sikka District, Ende District). All of them were measured for morphometric and was bleeding to collect blood as DNA sources. The blood sample was preserve with absolute ethanol. DNA extracted was analyzed by PCR-RFLP methods for D-loop region, SRY gene and GDF gene on Indonesian local goat does. If there were polymorphism then the procedure followed by the sequencing methods. The results showed that the morphometric discriminant factor variables between local goat were canon circumference, tail width and body length was 0,7 and than rump width, ear widths, wither height, skull height, skull width, body weight, chest girth, chest width and chest depth was 0,5. The colours and colour patterns of Benggala goat was dominant with black colour, Samosir goat with white colour and then the Kacang, Muara, Jawarandu and Marica goats were very high varieties of the combination almost all colours were observed. There were found 50 polymorphic sites and 19 unique haplotypes of the D-loop sequence. The average genetic diversity region of mt DNA was very high (0.014±0.002) and was significantly different between each sub-population with clustering indexs 63-99. The average distance of Y chromosome was lower (0.004±0.002) and to be founded 6 polymorphic sites with 4 unique haplotypes. The origin of local goats reveal to the maternal ancestors was including haplogroups lineage B, but the paternal origin ancestor was classified in to 4 groups; Kacang and Jawarandu one haplotype, Marica and Samosir one haplotype, Muara one haplotype and Benggala goat one haplotype. Based on the phenotypics, mitochondrion and Y chromosom analysis showed that the sub populations of Indonesian local goat as adistinct breed. Identification of the GDF9 gene promotor were polymorphic and have related with the prolific traits on the twinning does of Kacang and PE goats, but the GDF9 gene promotor were monomorphic on the Samosir and Muara goats.
ARON BATUBARA. Studi Keragaman Fenotipik dan Genetik Beberapa Sub Populasi Kambing Lokal Indonesia dan Strategi Pemanfaatannya Secara Berkelanjutan. Dibimbing oleh RONNY RACHMAN NOOR, ACHMAD FARAJALLAH, dan BESS TIESNAMURTI
Keragaman sumberdaya genetik ternak kambing merupakan modal dasar untuk meningkatkan produktivitas dan mengembangkan ternak kambing. Populasi kambing lokal bisa terancam habis antara lain disebabkan oleh program persilangan atau penggantian dengan bibit unggul eksotis. Pelestarian dan konservasi kambing lokal penting karena telah beradaptasi baik dengan lingkungan setempat, umumnya lebih bertahan hidup pada kondisi pedesaan.
Kambing lokal berpotensi besar untuk dimanfaatkan menjadi sumber pembentukan bibit unggul yang adaptif terhadap kondisi lokal di Indonesia. Data dan informasi tentang karakterisasi fenotipik dan genetik ternak kambing lokal Indonesia sampai saat ini masih sangat terbatas.
Tujuan penelitian ini adalah untuk memperoleh informasi keragaman karakteristik fenotip dan genetik 6 sub-populasi kambing lokal Indonesia yang mencakup karakteristik morfometrik dan warna dominan; keragaman genetik berdasarkan DNA mitokondria, koromosom Y dan keragaman gen GDF9.
Penelitian dirancang dengan metode survei dan analisis di laboratorium. Metode survei menggunakan purposive sampling, yaitu dengan memilih lokasi yang diduga masih terdapat populasi kambing lokal yang diamati. Penelitian lapangan untuk koleksi data fenotipik dilakukan di empat Propinsi, yaitu Propinsi Sumatera Utara (Kambing Samosir n=42 ekor di Kabupaten Samosir , Kambing Muara n=34 di Kabupaten Tapanuli Utara, Kambing Kacang n=217 di Kabupaten Deli Serdang); Propinsi Jawa Tengah (Kambing Jawarandu n=94 di Kabupaten Blora); Propinsi Sulawesi Selatan (Kambing Marica n=60: Kabupaten Maros, Kota Makassar, Kabupaten Jeneponto) dan Propinsi Nusa Tenggara Timur (Kambing Benggala n=96 di Kota Kupang, Kabupaten Kupang, Kabupaten Sikka, Kabupaten Ende). Dilakukan pengambilan data warna tubuh dominan dan warna belang kambing, penimbangan, pengukuran bagian-bagian tubuh dan pengambilan sampel darah. Analisis morfologi dengan pendekatan tehnik diskriminan dan kanonikal untuk menduga hubungan filogenik antar kambing lokal. Analisis diskriminan dilakukan pada bobot badan, bagian-bagian tubuh dan warna dominan. Kegiatan laboratorium terdiri dari ekstraksi DNA mengikuti metode Sambrook yang dimodifikasi, menganalisis DNA mitokondria, kromosom Y dan gen fekunditas (gen GDF9).
Dari 879 bp runutan ruas D-loop ditemukan 50 situs polimorfik dengan nilai keragaman 0.014±0.002 dan terdapat 19 kelompok haplotip. Asal-usul secara maternal keenam sub populasi kambing lokal Indonesia yang diteliti termasuk dalam kelompok garis keturunan (lineage) B mengelompok dengan kambing Asia Timur, Afrika Selatan dan Afrika Utara, Asia Selatan, Cina, Mongolia, Malaysia, Pakistan dan India.
Sebanyak 13 sampel DNA kambing jantan dianalisis dan diperoleh masing-masing situs nukleotida sepanjang 773 pb. Ditemukan 6 situs polimorfik dengan nilai keragaman 0.004±0.002 dan terdapat 4 kelompok haplotip unik yaitu Kambing Kacang dan Jawarandu, Marica dan Samosir, Muara serta Benggala.
Hasil identifikasi keragaman gen GDF9 ruas promotor pada induk beranak kembar bersifat polimorfik dan diduga berhubungan dengan sifat prolifik pada Kambing Kacang dan Peranakan Etawah (PE), akan tetapi bersifat monomorfik pada Kambing Samosir dan Muara.
Berdasarkan persyaratan kualitatif, kuantitatif, pemetaan genetik yang diperoleh dalam penelitian ini, maka hasil penelitian ini dapat dipertimbangkan sebagai salah satu bahan dalam penetapan dan standarisasi 6 rumpun/galur kambing lokal Indonesia yang saling berbeda di tingkat lapangan atau peternak dalam rangka pengembangan dan pelestarian sumberdaya genetik kambing lokal Indonesia secara berkelanjutan.
© Hak Cipta milik Institut Pertanian Bogor, tahun 2011
Hak Cipta dilindungi Undang-undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa
mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk
kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan
laporan, penulisan kritik, atau tinjauan suatu masalah dan pengutipan
tersebut tidak merugikan kepentingan yang wajar IPB. Dilarang
menggunakan dan memperbanyak sebagian atau seluruh Karya Tulis
DAN STRATEGI PEMANFAATANNYA SECARA
BERKELANJUTAN
ARON BATUBARA
Disertasi
Sebagai salah satu syarat untuk memperoleh gelar
Doktor pada
Program Studi Ilmu Produksi dan Teknologi Peternakan
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
BOGOR
Penguji pada Ujian Tertutup : Prof. (R) Dr. Ir. Ismeth Inounu, M.S.
Dr.Ir. Dedy Duryadi Solihin, DEA.
Penguji pada Ujian Terbuka : Dr.Ir. Riwantoro, MM.
Judul Disertasi : Studi Keragaman Fenotipik dan Genetik Beberapa Sub Populasi Kambing Lokal Indonesia dan Strategi Pemanfaatannya Secara Berkelanjutan
Nama : Aron Batubara
NRP : D161070041
Program Studi : Ilmu Produksi dan Teknologi Peternakan
Disetujui Komisi Pembimbing
Ketua
Prof.Dr.Ir. Ronny Rachman Noor, M.Rur.Sc.
Dr.Ir. Achmad Farajallah, M.Si.
Anggota Anggota
Dr.Ir. Bess Tiesnamurti, M.Sc.
Mengetahui
Ketua Program Studi/Mayor Dekan Sekolah Pascasarjana
Ilmu Produksi dan Teknologi Peternakan
Dr.Ir. Rarah R.A. Maheswari, DEA. Dr.Ir. Dahrul Syah, M.Sc.Agr.
Dengan mengucapkan puji dan syukur kepada Tuhan Yang Maha Kuasa, karena berkat karunia-Nya penulis dapat menyelesaikan penelitian dan penyusunan disertasi dengan judul “Studi Keragaman Fenotipik dan Genetik Beberapa Sub Populasi Kambing Lokal Indonesia dan Strategi Pemanfaatannya Secara Berkelanjutan”. Disertasi ini sebagai salah satu syarat untuk memperoleh gelar Doktor pada Program Studi Ilmu Produksi dan Teknologi Peternakan, Sekolah Pascasarjana Institut Pertanian Bogor.
Penelitian ini dilakukan atas dasar bahwa kambing lokal Indonesia sebagai salah satu kekayaan plasma nuftah ternak di Indonesia perlu di eksplorasi identitas, karakteristik dan potensi genetiknya sebelum terlanjur hilang. Informasi tentang karakteristik secara fenotipik dan genetik berguna untuk menginventarisasi kekayaan sumberdaya genetik kambing Indonesia. Dengan semakin meningkatnya kebijakan pencapaian produksi daging nasional di berbagai daerah penggunaan bibit unggul eksotis terus meningkat, sehingga pemanfaatan bibit ternak lokal semakin kurang mendapat perhatian. Oleh sebab itu perlu dilakukan penelitian karakterisasi fenotipik dan keragaman genetik yang merupakan tahap awal sebelum penelitian potensi produktivitas dan upaya pembentukan bibit unggul yang berbasis lokal, serta upaya konservasi dan pemanfaatan ternak secara berkelanjutan.
Pengembangan Pertanian selaku Ketua Komisi Pembinaan Tenaga, yang telah memberikan kesempatan kepada penulis untuk mengikuti Program S3, Kepala Pusat Penelitian dan Pengembangan Peternakan dan Dr.Ir. Simon Petrus Ginting, M.Sc. selaku Kepala Loka Penelitian Kambing Potong Sei Putih, yang memberikan motivasi, dorongan dan restu kepada penulis untuk melanjutkan ke jenjang Program S3 di Sekolah Pascasarjana, Institut Pertanian Bogor.
Ucapan terimakasih yang sama penulis sampaikan kepada Dr.Ir. Rarah Ratih Adjie Maheswari, DEA, selaku Ketua Program Studi/Mayor Ilmu Produksi dan Teknologi Peternakan (IPTP), Prof.Dr.Ir. Cece Sumantri, M.Agr.Sc selaku Ketua Departemen Ilmu Produksi dan Teknologi Peternakan Fakultas Peternakan Institut Pertanian Bogor yang sekaligus sebagai penguji luar Komisi pada Ujian Kualifikasi Doktor dan Prof.Dr.Ir. Muladno, MSA sebagai penguji luar Komisi pada Ujian Kualifikasi Doktor, Rektor Institut Pertanian Bogor, Dekan Sekolah Pasca Sarjana IPB, Dekan Fakultas Peternakan IPB Bogor.
Terima kasih yang sebesar-besarnya juga disampaikan kepada Kepala Dinas Peternakan dan Perikanan Kabupaten Tapanuli Utara, Kepala Dinas Pertanian, Peternakan dan Perikanan Kabupaten Samosir, Kepala Dinas Peternakan Kabupaten Blora Jawa Tengah, Dr. Ir. Jacob Nulik, M.Sc. sebagai Kepala BPTP Nusa Tenggara Timur, Kepala BPTP Sulawesi Selatan, Ir. Matius Sariubang MS, Ir. Daniel Pasambe, Ir Bonggas Pasaribu, Ir. Rosianna Tarigan, Ir. Deborah Kana Hau, M.Si., Drh. Wasito M.Si., Imanianto, Wagimin yang telah banyak membantu selama pengambilan data dan sampel di lapangan, Wildan Muttaqin S.Si., M.Si dan Muhammad Rizal Hasan S.Si., M.Si yang telah banyak membantu selama proses analisis molekuler di Laboratorium Fungsi Hayati dan Perilaku Hewan Departemen Biologi IPB. Rekan-rekan seperjuangan Ir. Suryana M.Si., Ir. Eko Handiwinawan M.Si, drh. Bambang Ngaji Utomo M.Sc., Ben Juvarda, SPt., M.Si., yang selalu memberikan semangat dan motivasi kepada penulis.
Kepada istri saya tercinta Helena Lydia Tobing, S.Pt dan ketiga putri saya tersayang Ruth Amelia Batubara, Desy Margaretta Batubara dan Joice Deatri Batubara dan seluruh keluarga besar saya, terimakasih atas perhatian, kesabaran, pengertian, dorongan, pengorbanan dan doa yang diberikan selama ini kepada Penulis, serta kepada semua pihak yang telah membantu dan tidak dapat penulis sebutkan satu persatu.
Akhirnya Penulis berharap semoga disertasi ini memberikan sumbangsih dan bermanfaat bagi perkembangan ilmu pengetahuan dan pembangunan peternakan di Indonesia.
Bogor,……Juli 2011
Penulis dilahirkan di Desa Hutatonga, Kecamatan Sipirok (saat ini Kecamatan Arse), Kabupaten Tapanuli Selatan, Sumatera Utara pada tanggal 22 Mei 1968 sebagai anak kedelapan dari sepuluh bersaudara dari pasangan Ali Musa Batubara (alm) dan Mince Ritonga (alm). Pendidikan sarjana ditempuh pada Program Studi Produksi Ternak, Fakultas Peternakan, Universitas HKBP Nommensen Medan, lulus tahun 1992. Kesempatan untuk melanjutkan ke jenjang program magister pada Program Studi Produksi Ternak pada Departemen of Animal Production and Health, Institute of Tropical Medicine, Antwerp, Belgia tahun 1995 dan lulus tahun 1997. Pada tahun 2007 penulis melanjutkan ke jenjang program Doktor pada Program Studi/Mayor Ilmu Produksi dan Teknologi Peternakan (IPTP) dengan minat Ilmu Pemuliaan dan Genetika Ternak, Sekolah Pascasarjana Institut Pertanian Bogor, dengan beasiswa dari DIPA Badan Penelitian dan Pengembangan Pertanian, Kementerian Pertanian.
ABSTRAK……….. iii
RINGKASAN..……….. iv
DAFTAR ISI……….. xvi
DAFTAR TABEL……….………... xix
DAFTAR GAMBAR………... xx
DAFTAR LAMPIRAN……….……….. xxi
PENDAHULUAN Latar Belakang……….……….. 1
Tujuan Penelitian……….………….. 3
Manfaat Penelitian………..……... ……... 3
Kerangka Pemikiran…….………... 4
TINJAUAN PUSTAKA Sejarah Budidaya Ternak Kambing………. 6
Keragaman Genetik Ternak………... 7
Pelestarian Sumberdaya Genetik Ternak……….. 8
Sumberdaya Genetik Kambing Indonesia………..…………... 9
Sifat Kuantitatif dan Kualitatif……… 12
Penanda Genetik……….……….. 13
DNA Mitokondria……… 14
DNA Kromosom Y……….. 16
Gen yang Berhubungan dengan Sifat Prolifik…… ……….. 17
KARAKTERISTIK MORFOMETRIK DAN ANALISIS FILOGENI PADA ENAM SUB POPULASI KAMBING LOKAL INDONESIA Pendahuluan………... 20
Bahan dan Metode………. 21
Waktu dan Tempat Penelitian………. 21
Pengumpulan Sampel Kambing………. 22
Analisis Statistik………... 24
Analisis Morfometrik………. 25
Hasil dan Pembahasan Bobot Badan ………. Parameter Ukuran Tubuh………... 26 27 Plot Penyebaran Kambing menurut Ukuran Fenotipik…………... 34
Nilai Campuran Fenotipik antar Kelompok……….. 36
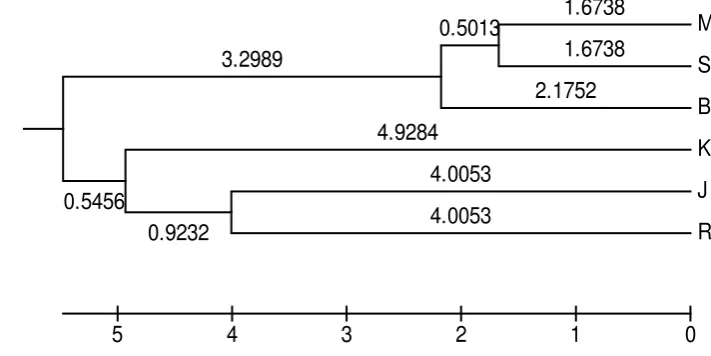
Penentuan Jarak Genetik dan Pohon Fenogram………... 37
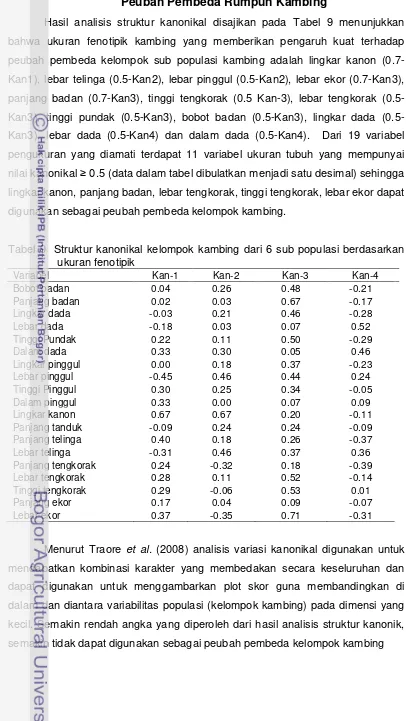
Peubah Pembeda Rumpun Kambing………... 39
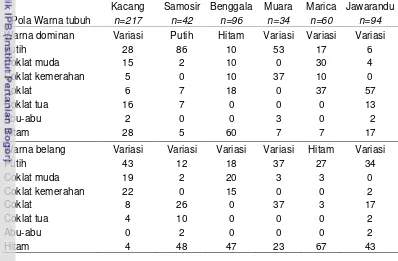
Pola Warna Tubuh………... 40
Simpulan……….…. 43
Amplifikasi DNA..………... 45
Perunutan DNA………. 46
Analisis Data………... 46
Hasil dan Pembahasan Polimorfisme Segmen daerah D-loop DNA Mitokondria……….... 47
Keragaman Runutan Nukleotida……….... 50
Jarak Genetik Kambing Penelitian dengan Kambing Lainnya 50 Simpulan……… 57
KARAKTERISASI MOLEKULER PADA ENAM SUB POPULASI KAMBING LOKAL INDONESIA BERDASARKAN ANALISIS DNA KROMOSOM Y Pendahuluan……….……… 58
Bahan dan Metode Sampel Darah Kambing Penelitian……….… 58
Ekstraksi DNA……… 58
Amplifikasi DNA ………..……….. 58
Perunutan DNA……….……. 59
Analisis Data……….…. 59
Hasil dan Pembahasan Polimorfisme DNA Kromosom Y Segmen gen SRY……….... 60
Frekuensi Nukleotida dan Jarak Genetik………... 62
Simpulan……….………... 65
IDENTIFIKASI KERAGAMAN GEN GDF9 DAN HUBUNGANNYA DENGAN SIFAT PROLIFIK PADA KAMBING LOKAL INDONESIA Pendahuluan……… 66
Bahan dan Metode……….. 67
Pengambilan Sampel Darah Kambing ……….. 67
Ekstraksi DNA………. 68
Amplifikasi Gen GDF9…....……….. 68
Genotiping PCR-RFLP dan sekuensing……….……… 69
Analisis Data………..………. 69
Hasil dan Pembahasan…………..……….……… 69
Simpulan……….……….………. 72
PEMBAHASAN UMUM……….…... 73
SIMPULAN UMUM DAN SARAN Simpulan……… 79
Saran……….. 81
DAFTAR PUSTAKA……….……….. 82
1 Rataan, simpangan baku dan kisaran bobot hidup dewasa 6 sub
populasi kambing lokal……… 26
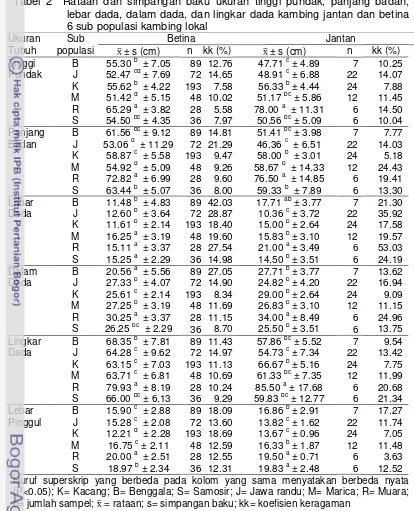
2 Rataan dan simpangan baku ukuran tinggi pundak, panjang badan, lebar dada, dalam dada, dan lingkar dada kambing jantan dan betina 6 sub populasi kambing lokal ……….
28
3 Rataan dan simpangan baku ukuran panjang tengkorak, lebar tengkorak, dan tinggi tengkorak jantan dan betina dewasa 6 sub
populasi kambing lokal ……….……… 30
4 Rataan dan simpangan baku panjang dan lebar ekor jantan dan betina 6 sub populasi kambing lokal……….…… 31
5 Rataan dan simpangan baku panjang telinga dan lebar telinga jantan dan betina dewasa 6 sub populasi kambing lokal………. 32
6 Rataan dan simpangan baku lingkar kanon jantan dan betina 6 sub
populasi kambing lokal ………..………. 34
7 Persentase nilai kesamaan dan campuran 6 sub populasi kambing
lokal ……… 36
8 Jarak genetik berdasarkan ukuran tubuh antar 6 sub populasi
kambing ……..………..… 37
9 Struktur kanonikal kelompok kambing dari 6 sub populasi kambing lokal berdasarkan ukuran fenotipik………... 39
10 Persentase pola warna tubuh dominan dan warna belang pada 6 sub populasi kambing lokal …………. …..………..…... 40
11 Jumlah haplotip berdasarkan runutan nukleotida D-loop mtDNA setiap sub populasi kambing lokal Indonesia ……….. 47
12 Mutasi nukleotida sebagai penciri kelompok kambing lokal Indonesia dibandingkan dengan Capra hircus (GenBank no. akses
NC_005044) ………..…….. 49
13 Keragaman nukleotida D-loop mtDNA pada 6 kambing lokal
Indonesia ……….. 50
14 Jarak genetik berdasarkan runutan nukleotida pada 6 sub populasi kambing lokal Indonesia ………….……….….. 52
15 Matrik keragaman nukleotida gen SRY kromosom Y pada kambing
Halaman
1 Diagram alir kerangka pemikiran penelitian………. 5
2 Skema genom daerah D-loop kambing………. 16
3 Skema genom gen SRY kromosom Y dan kromosom X.………. 17
4 Peta lokasi pengambilan sampel dan data penelitian karakterisasi enam sub populasi kambing lokal Indonesia……….……….. 22
5 Titik pengukuran morfometrik kambing ..………..…….. 24
6 Plot penyebaran kelompok kambing berdasarkan ukuran-ukuran
fenotipik pada 6 sub populasi kambing lokal …..………. 35
7 Pohon fenogram penyebaran kelompok kambing lokal Indonesia berdasarkan ukuran fenotipik………... 38
8 Pola warna dominan dan belang pada 6 sub populasi kambing lokal … 42
9 Struktur genom mitokondria yang diapit oleh primer AF23 dan AF22. Nomor mengacu pada Capra hircus (GenBank no. akses AF 533441)..
47
10 Polimorfisme runutan nukleotida daerah D-loop DNA mitokondria pada 6 sub populasi kambing lokal mengacu pada Capra hircus (GenBank no. akses AF 533441) (tiga baris pertama dibaca secara vertikal
merupakan posisi nukleotida)………. 48
11 Dendogram 6 sub populasi kambing lokal Indonesia berdasarkan ruas
D-loop mtDNA ………….……….. 54
12 Posisi 6 sub populasi kambing lokal Indonesia dalam dendogram
kambing-kambing di dunia berdasarkan ruas D-loop ………..…………... 55
13 Median joining network dari 19 haplotip nukleotida daerah D-loop DNA
mitokondria pada 6 sub populasi kambing lokal Indonesia……... 57
14 Struktur ruas gen SRY yang diapit oleh AF126 dan AF127 pada 6 sub populasi kambing lokal Indonesia………
61
15 Polimorfisme nukleotida gen SRY pada 6 sub populasi kambing lokal
Indonesia……… 62
16 Dendogram Neigbour Joining berdasarkan runutan nukleotida gen SRY
Benggala (B1)………... 65
18 Mutasi nukleotida gen GDF9 ruas promotor pada kelompok induk prolifik (mutan) dan induk non-prolifik (wild) pada Kambing Kacang dan
Peranakan Etawah ……….. 70
19 Dendogram kambing lokal Indonesia berdasarkan runutan nukleotida
1 Pola warna dominan dan warna belang 6 sub populasi kambing lokal……….
94
2 Form daftar karakter morfologi yang diukur………...…..……… 100
3 Lokasi pengambilan data dan sampel darah kambing lokal (Kambing Kacang, Samosir, Marica, Jawarandu, Muara dan
Benggala)……….. 101
4 Nama/bangsa kambing, nomor akses dan asal sampel yang digunakan dari GenBankuntuk membentuk pohon filogeni ……….
102
5 Pensejajaran berganda nukleotida ruas D-loop DNA mitokondria pada kambing lokal……….………..
103
6 Pensejajaran berganda nukleotida gen SRY pada kambing lokal … 112
7 Pensejajaran berganda nukleotida gen GDF 9 pada kambing lokal Indonesia……….………..……….
PENDAHULUAN
Latar Belakang
Sumberdaya genetik ternak dihadapkan pada dua tantangan yang saling
bertolak belakang yaitu pemanfaatan ternak unggul eksotis untuk memenuhi
kebutuhan daging dan susu, sementara disisi lain sumberdaya genetik ternak
lokal terus berkurang. Permintaan produksi asal ternak terus meningkat di
negara-negara sedang berkembang. FAO memperkirakan kebutuhan daging
akan meningkat dua kali lipat pada tahun 2030 dibandingkan dengan kebutuhan
pada tahun 2000 dan kebutuhan susu meningkat menjadi lebih dari dua kali lipat.
Selain itu, ada kecenderungan penggunaan ternak unggul eksotis untuk
mencapai target produksi ternak secara maksimal sehingga perhatian terhadap
pengembangan ternak lokal termarjinalkan. Dampak negatif peningkatan ternak
unggul eksotis adalah penurunan populasi sumberdaya genetik ternak lokal
secara cepat terutama di negara-negara yang sedang berkembang (FAO 2000;
Cardellino & Boyazoglu 2009).
Sekitar 190 dari 7600 rumpun ternak di dunia yang telah tercatat akan
punah selama 15 tahun terakhir, dan 1500 rumpun diantaranya berada dalam
status beresiko mendekati kepunahan. Sekitar 60 rumpun terdiri atas sapi,
kambing, babi, kuda dan unggas telah punah/hilang (lost) selama 5 tahun
terakhir. Tingkat kehilangan rumpun ternak lokal paling tinggi dijumpai di
negara-negara sedang berkembang (FAO 2007). Konservasi dan pengembangan
rumpun ternak lokal sangat penting, sebab rumpun lokal dapat memanfaatkan
pakan berkualitas rendah, lebih tahan terhadap tekanan perubahan iklim dan
serangan penyakit, serta sebagai sumber gen-gen yang secara ekonomi
menguntungkan untuk peningkatan kesehatan dan performan sifat-sifat produksi
pada rumpun ternak unggul komersial (Cardellino 2006; FAO 2007; Ruane
2000).
Salah satu komoditas kekayaan plasma nutfah nasional di sub sektor
peternakan adalah ternak kambing. Kambing menyebar di berbagai daerah
dengan iklim yang berbeda dan terpisah dalam jangka waktu yang lama. Faktor
lingkungan dan perlakuan seleksi yang sangat bervariasi mengakibatkan laju
perubahan genetik yang sangat beragam (Rout et al. 2008).
Di Asia Tenggara khususnya di Indonesia, ternak kambing memegang
juga sebagai sumber daging, pupuk, pengoptimalan tenaga kerja keluarga dan
status sosial serta dibutuhkan dalam aspek budaya (Subandriyo 2008). Hampir
99% ternak ruminansia kecil di Indonesia merupakan skala usaha ternak kecil
(Soedjana 2008). Kontribusi ternak ruminansia kecil dalam usahatani sangat
berperanan penting. Ternak kambing dapat mengkonversi hijauan berkualitas
rendah menjadi protein hewani, sebagai sumber pupuk kandang serta sebagai
tabungan. Usahatani ternak kambing merupakan bagian dari sistim usahatani
secara umum di Indonesia.
Sampai saat ini, tampilan morfologi masih umum digunakan secara
praktis untuk mengkarakterisasi dan menyeleksi ternak. Penampilan morfologi ini
banyak dipengaruhi oleh faktor lingkungan luar seperti ketersediaan pakan dan
iklim. Hal ini menjadikan seleksi ternak berdasarkan morfologi membutuhkan
waktu lebih lama (Mabrouk et al. 2008; Nsoso et al. 2004; Lanari et al. 2003).
Perkembangan ilmu pengetahuan dan teknologi biologi molekuler akhir-akhir ini,
khususnya yang berhubungan dengan penggunaan penanda molekuler telah
mempercepat karakterisasi sifat-sifat yang bernilai ekonomi tinggi, daya tahan
terhadap penyakit, asal-usul dan kekerabatan suatu individu atau rumpun ternak
tertentu (Nijman et al. 2003).
Penanda molekuler yang populer saat ini, antara lain DNA-mitokondria
dan DNA mikrosatelit. Penanda DNA-mitokondria menggambarkan karakteristik
yang diturunkan melalui garis induk (maternal) (Fan-Bin 2007). Penanda DNA
mikrosatelit menggambarkan karakter rekombinasi (Wimmers et al. 2000; Tadelle
2003; Zhang et al. 2002). Selain itu penanda molekuler DNA kromosom Y
menggambarkan karakteristik yang diwariskan melalui pejantan (paternal)
(Verkaar et al. 2003).
Sejalan dengan perkembangan yang pesat lalu lintas ternak antar
kawasan maka lama kelamaan terjadilah proses adaptasi terhadap
agroekosistem yang spesifik sesuai dengan lingkungan dan manajemen
pemeliharaan yang ada ditempat tertentu. Dengan demikian terjadi evolusi yang
membuka kemungkinan munculnya rumpun/galur/ras kambing yang baru. Balai
Penelitian Ternak Ciawi sejak tahun 1995 sudah memulai mengkarakterisasi
Kambing Kacang, Peranakan Etawah, Kosta dan Gembrong pada tahun 1997
(Setiadi et al. 1995; 1997). Diperkirakan masih banyak lagi rumpun kambing lokal
Indonesia lainnya yang belum dikarakterisasi dan sebagian mungkin sudah
Perwujudan tujuan pelestarian plasma nutfah dan spesifikasi ternak
menurut bangsa dapat dimulai dengan sebutan khusus menurut wilayah,
misalnya Kambing Marica yang ada di Sulawesi Selatan, Kambing Benggala di
Nusa Tenggara, Kambing Lakor di Maluku, Kambing Samosir dan Kambing
Muara di Sumatera Utara, Kambing Jawarandu di Jawa Tengah dan jenis
kambing lainnya di daerah Indonesia. Penelitian tentang karakterisasi kambing
lokal Indonesia perlu dilakukan untuk mendapatkan data dan informasi potensi
sumberdaya genetik kambing lokal Indonesia.
Tujuan Penelitian
1. Menginventarisasi karakteristik fenotip secara kualitatif dan kuantitatif ciri-ciri
morfologi pada kambing lokal Indonesia.
2. Mengkarakterisasi keragaman genetik daerah D-loop DNA Mitokondria pada
kambing lokal Indonesia untuk mengetahui asal usul menurut garis keturunan
maternal.
3. Mengkarakterisasi keragaman gen SRY ruas DNA kromosom Y pada
kambing lokal Indonesia untuk mengetahui asal usul menurut garis keturunan
paternal.
4. Mengidentifikasi keragaman gen GDF9 yang berhubungan dengan sifat
prolifik pada kambing lokal Indonesia.
Manfaat Penelitian
Hasil penelitian ini diharapkan dapat memberikan sumbangan pemikiran yang
bermanfaat untuk
1. Mengembangkan kambing lokal Indonesia dengan memperhatikan
keragaman populasi yang ada.
2. Menyediakan informasi yang dapat digunakan Pemerintah Daerah khususnya
Dinas Peternakan sebagai pedoman dalam menyusun perencanaan
pembangunan peternakan, khususnya pengembangan kambing lokal
Indonesia dalam rangka pelestarian plasma nutfah kambing lokal Indonesia
yang bertujuan untuk pengembangan dan pemanfaatannya secara
3. Menyediakan informasi tentang pemanfaatan ciri-ciri fisik kambing lokal
Indonesia yang dapat digunakan sebagai sumber keragaman genetik untuk
pembentukan galur bibit kambing unggul.
Kerangka Pemikiran
Keragaman sumberdaya genetik merupakan modal dasar untuk
meningkatkan produktivitas kambing. Data dan informasi tentang karakter
fenotipik dan genetik ternak kambing lokal Indonesia sampai saat ini masih
sangat terbatas. Penelitian ini perlu dilakukan untuk mendapatkan data fenotifik
dan genotip beberapa kambing lokal Indonesia.
Penelitian dirancang dengan metode survei dan laboratorium. Metode
survei menggunakan purposive sampling, yaitu dengan memilih lokasi yang
diduga masih bisa ditemukan populasi kambing lokal terutama daerah yang
paling padat populasinya. Lokasi dipilih berdasarkan informasi dari instansi
terkait di Propinsi kemudian menuju Kabupaten dan selanjutnya menuju
Kecamatan dan Desa. Sampel kambing diusahakan diambil dari lokasi yang
berjauhan. Pada lokasi terpilih dilakukan pengambilan sampel darah,
penimbangan, pengamatan warna dan pola warna kambing serta ukuran-ukuran
tubuh diukur dengan pita, jangka dan tongkat ukur. Kegiatan laboratorium terdiri
atas ekstraksi DNA dari darah mengikuti metode Sambrook yang dimodifikasi,
menganalisis DNA mitokondria, DNA kromosom Y dan gen GDF9.
Diagram alir kerangka pemikiran penelitian Studi keragaman fenotipik dan
genetik beberapa sub populasi kambing lokal Indonesia dan strategi
Gambar 1 Diagram alir kerangka pemikiran penelitian KAMBING LOKAL INDONESIA
POTENSI
• Sebagai sumberdaya genetik ternak daerah/ nasional • Kambing penghasil daging, susu, pupuk
• Berkontribusi signifikan terhadap pendapatan peternak dan status sosial
PERMASALAHAN
• Data dan informasi karakter morfometrik kambing lokal masih terbatas
• Data dan informasi pemetaan genetik tentang pengelompokan dan asal usul secara maternal dan paternal masih terbatas
• Data dan informasi potensi keragaman gen fungsional yang berhubungan dengan sifat prolifik pada kambing lokal masih terbatas.
ANALISIS MORFOMETRIK Data kualitatif (warna dan pola warna tubuh) dan kuantitatif (ukuran tubuh) ANALISIS DNA MITKONDRIA Keragaman genetik DNA mitokondria untuk mengiden tifikasi hubungan asal usul kambing lokal secara maternal ANALISIS DNA KROMOSOM Y Keragaman genetik DNA kromosom Y untuk mengiden tifikasi hubungan asal-usul secara paternal ANALISIS KERAGAMAN GEN GDF9 Identifikasi keragaman gen GDF9 pada kambing lokal
Status pemetaan keragaman fenotipik dan identitas genetik enam sub populasi kambing lokal Indonesia
TINJAUAN PUSTAKA
Sejarah Budidaya Ternak Kambing
Ternak kambing (Capra hircus) sering diartikan sebagai ternak yang
dapat membantu memecahkan masalah kemiskinan di kalangan peternak,
karena kemampuannya dalam memanfaatkan hijauan dalam jumlah terbatas
seperti pada lingkungan yang kritis dan kering/lahan marjinal (MacHugh &
Bradley 2001). Kambing merupakan hewan pertama yang didomestikasi, diduga
berasal dari Kambing liar Capra aegargus. Pada awalnya sekitar 10 000-11 000
tahun yang silam di daerah Kawasan Timur Tengah manusia zaman Neolithic
mulai memelihara kambing dalam jumlah kecil untuk mendapatkan susu, daging
dan kotorannya sebagai bahan bakar, juga sebagai bahan untuk pakaian dan
bangunan yang terbuat dari bulu, tulang, kulit dan urat daging (MacHugh et al.
2001; Zeder et al. 2000).
Saat ini lebih dari 300 rumpun ternak kambing yang hidup di berbagai
iklim dan ketinggian, mulai dari dataran tinggi sampai ke daerah dataran rendah.
Ahli arkeologi melaporkan dua tempat yang berbeda sebagai asal dari pertama
kali proses domestikasi kambing dilakukan, yaitu; Lembah Sungai Eupharate di
Nevali Cori, Turki (11 000 B.C.) dan di Pegunungan Zagros di Garj Dareh, Iran
(10 000 B.C.). Kemungkinan situs yang lain adalah Indus Basin, di daerah
Mehgarh, Pakistan (9 000 B.C.) dan kemungkinan di Pusat Anatolia dan bagian
utara Levant. Situs arkeologi yang lain yang penting menunjukkan adanya
proses domestikasi kambing di Cayonu, Turki (8 500 - 8 000 B.C.), Tell Abu
Hureyra, Syria (8 000 – 7 400 B.C.), Jerico, Israel (7 500 B.C.) dan Ain Ghazal,
Jordan (7 600 – 7 500 B.C.) (Hirst 2008).
Kambing yang dipelihara (Capra aegagrus hircus) berasal dari 3
kelompok kambing liar yang telah dijinakkan, yaitu Bezoar goat atau kambing liar
Eropa (Capra aegagrus), kambing liar India (Capra aegagrus blithy), dan Makhor
goat atau Kambing Makhor di pegunungan Himalaya (Capra falconeri).
Sebagian besar kambing yang diternakkan di Asia berasal dari keturunan
Bezoar, termasuk Kambing Gunung Sumatra (Caprinae sumatraensis) atau
disebut juga Kambing Gurun (Maddox & Cockett 2007).
Kambing biasanya dibedakan berdasarkan letak geografis, karakteristik
morfologi, dan performan produksi. Kambing berdasarkan ukuran tubuh
sedang dan tipe kecil. Berdasarkan performan produksi kambing dibedakan atas
kambing tipe perah, tipe pedaging dan tipe dwi guna (dual purpose). Saat ini
usaha ternak kambing juga sangat berperan mendukung kebutuhan akan ternak
Qurban bagi yang beragama Islam yang merupakan agama mayoritas di
Indonesia dan juga pada aliran kepercayaan seperti agama Parmalim di Pulau
Samosir dan daerah sekitar Danau Toba. Saat ini, usaha ternak secara
komersial sudah berkembang di beberapa daerah di Indonesia untuk
memproduksi susu kambing, dimana kualitas susu kambing mempunyai
beberapa keunggulan jika dibandingkan dengan jenis susu ternak lainnya.
Usaha ternak kambing perah berperan sekaligus menghasilkan ternak kambing
potong.
Keragaman Genetik Ternak
Keragaman genetik terjadi tidak hanya antar rumpun tetapi juga di dalam
satu rumpun yang sama, antar populasi maupun di dalam populasi. Pada
spesies ternak domestik suatu identifikasi tingkat keragaman, terutama pada
lokus-lokus yang mempunyai sifat bernilai penting mempunyai keterkaitan
dengan seleksi dalam program pemuliaan (Handiwirawan & Subandriyo 2004;
Abdullah 2008). Salah satu alat bantu yang dapat digunakan untuk mendeteksi
keragaman populasi adalah DNA mitokondria dan DNA mikro satelit (Muladno
2006; Yuwono 2006) dan DNA kromosom Y segmen gen SRY.
Keragaman genetik dalam populasi merupakan modal dasar aplikasi
teknologi pemuliaan dalam pemanfaatan hewan. Keragaman genetik populasi
yang digambarkan dalam keragaman penampilan hewan adalah refleksi
informasi genetik yang dimilikinya. Perbedaan penampilan disebabkan selama
proses domestikasi tipe atau rumpun-rumpun hewan terpisah secara genetik
karena adanya proses adaptasi dengan masing-masing lingkungan lokal dan
kebutuhan komunitas lokal sehingga dihasilkan rumpun yang berbeda (Muladno
2006). Adanya kemampuan adaptasi hewan disebabkan hewan memiliki
kemampuan menghasilkan lebih dari satu alternatif bentuk morfologi, status
fisiologi, dan atau tingkah laku sebagai reaksi terhadap lingkungan (Noor 2008).
Lebih dari 12 000 tahun yang lalu terdapat 12 spesies ternak telah
didomestikasikan dan berevolusi sehingga menjadi rumpun (breed) yang secara
genetik unik dan berbeda, beradaptasi terhadap lingkungan dan komunitas
spesies yang telah terdomestikasi, bersama dengan lebih dari 80 spesies
kerabat liarnya yang merupakan sumberdaya genetik ternak di bumi ini yang
berperan penting untuk pangan dan produksi pertanian. Berbagai rumpun ternak
yang telah berkembang dalam berbagai sistem dan lingkungan yang ada saat ini
telah menghasilkan berbagai kombinasi gen yang unik. Gen-gen ini tidak hanya
menentukan kualitas sifat produksi dari masing-masing rumpun, tetapi juga
terhadap kemampuan adaptasinya pada perubahan kondisi lingkungan lokal
termasuk makanan, ketersediaan air, iklim dan hama penyakit (FAO 2001).
Berbagai macam kebutuhan manusia sehari-hari dipenuhi dari spesies
ternak, dalam bentuk pangan maupun kebutuhan lainnya. Namun hanya
sebagian kecil dari total keragaman genetik ternak dan kerabat liarnya, yakni
sekitar 40 spesies yang memenuhi sebagian besar proporsi dari produksi ternak
global. Keragaman ternak di dalam genetik ternak dan beberapa kerabat lainnya
telah menjadi sumber keragaman dari rumpun dan populasi ternak. Keragaman
genetik ini penting dalam pembentukan ternak modern dan akan terus
berkelanjutan di masa mendatang (Subandriyo & Setiadi 2003). Sumberdaya
genetik ternak sedikitnya memiliki empat manfaat, yaitu (1) keberlanjutan dan
peningkatan produksi pangan; (2) memaksimumkan produktivitas lahan dan
sumberdaya pertanian; (3) pencapaian pertanian berkelanjutan untuk
memberikan keuntungan masa kini dan generasi rumpun ternak yang akan
datang; (4) pemenuhan keanekaragaman baik yang telah maupun yang belum
diketahui manfaatnya bagi kehidupan sosial masyarakat. Ketersediaan
keanekaragaman genetik ternak, termasuk kambing akan mempengaruhi
keberhasilan strategi pemuliaan untuk masa yang akan datang (FAO 2007).
Pelestarian Sumberdaya Genetik Ternak
Keragaman genetik ini penting dalam pembentukan rumpun dan populasi
ternak modern dan akan terus berlanjut untuk masa mendatang. Punahnya
keragaman plasma nutfah ternak tidak akan dapat diganti meskipun dengan
kemajuan bioteknologi, paling tidak sampai saat ini. Negara-negara sedang
berkembang pada umumnya berada pada iklim dengan perubahan temperatur
yang ekstrim antara musim panas dan hujan. Pada kondisi seperti ini akan
terbentuk rumpun ternak yang beradaptasi. Walaupun produktivitasnya rendah
apabila dibandingkan dengan dengan rumpun yang terdapat di daerah temperate
penyakit; tahan terhadap fluktuasi ketersediaan dan mutu pakan dan air; tahan
terhadap perubahan temperatur, kelembaban dan pengaruh iklim ekstrim
lainnya. Rumpun ternak ini juga beradaptasi terhadap pemeliharaan yang
kurang baik sehingga memiliki nilai yang sangat berharga untuk mengantisipasi
berbagai perubahan alam dan lingkungan diwaktu yang akan datang (FAO
2007).
Dengan demikian, pelestarian terhadap sumberdaya genetik ternak lokal
sebagai bagian dari komponen keanekaragaman hayati adalah penting untuk
memenuhi kebutuhan pangan, pertanian dan perkembangan sosial masyarakat
di masa yang akan datang. Ada beberapa alasan untuk ini, antara lain: (1) lebih
dari 60 persen dari rumpun-rumpun hewan ternak di dunia berada di
negara-negara sedang berkembang, (2) konservasi rumpun ternak lokal tidak menarik
bagi petani, (3) secara umum tidak ada program monitoring yang sistematis dan
tidak tersedianya informasi deskriptif dasar sebagian besar sumberdaya genetik
hewan ternak, serta (4) sedikit sekali rumpun-rumpun hewan ternak asli yang
telah digunakan dan dikembangkan secara aktif (FAO 2001; 2007).
Pelestarian sumberdaya genetik ternak pada dasarnya dapat dilakukan
salah satu atau gabungan dari: (1) mempertahankan populasi ternak hidup baik
dalam bentuk in-situ maupun ex-situ pada satu tempat tertentu, (2) penyimpanan
beku (cryogenic), dan (3) penyimpanan dalam bentuk DNA. Dalam beberapa
hal, mempertahankan populasi merupakan metode yang lebih praktis.
Pelestarian pada ternak hidup mempunyai beberapa keuntungan antara lain;
rumpun-rumpun ternak yang dilestarikan secara bertahap dapat merespon
terhadap perubahan pengaruh eksternal dan memungkinkan dilakukan evaluasi
kinerjanya (FAO 2007).
Sumberdaya Genetik Kambing Indonesia
Sumberdaya ternak kambing di Indonesia saat ini terdiri dari tiga
kelompok, yakni: (1) ternak asli, (2) ternak impor, dan (3) ternak yang telah
beradaptasi dalam jangka waktu lama sehingga membentuk karakteristik
tersendiri (ternak lokal). Pentingnya nilai konservasi pada kelompok hewan
ternak ini, beberapa rumpun ternak ini perlu dijadikan target konservasi sekaligus
pemanfaatannya (Utoyo 2002). Rumpun ternak kambing di Indonesia dan
rumpun kambing lainnya adalah merupakan hasil domestikasi sekitar 10000
terus melalui Khyber Pass, kemudian menyebar melalui pulau Sumatra,
Kalimantan, Sulawesi, Jawa sampai Indonesia bagian Timur.
Kambing Kacang merupakan rumpun kambing asli Indonesia, bentuk
badannya kecil dengan tinggi pundak sekitar 50-60 cm serta prolifik. Introduksi
rumpun kambing impor Benggala dari India dimulai oleh orang-orang Arab dan
kambing-kambing tersebut didatangkan melalui pelabuhan pantai utara Pulau
Jawa. Mulai pada tahun 1911-1931 didatangkan rumpun-rumpun Kambing
Kashmir, Angora (Montgomey), Benggala dan Etawah untuk stasiun ternak
kambing atau stasiun peternakan di Keresidenan Kedu, Solo, Yogyakarta,
Banyumas, Pekalongan, Pangalengan, Padang Mangatas, Wlingi (Blitar),
Sumba, dan Sumbawa. Disamping dari India pada tahun 1928 pernah pula
diimpor dari Negeri Belanda yaitu “Hollandse Edelgeiten” (Kambing Belanda
Murni). Rumpun kambing dari India selanjutnya disilangkan dengan rumpun
kambing lokal Indonesia dengan cara digaduhkan atau menempatkan pejantan
Etawah murni atau persilangan dengan proporsi darah Etawah yang cukup tinggi
di desa-desa yang akan dikembangkan peternakan kambingnya. Hasil
persilangan tersebut dikenal dengan nama Peranakan Etawah, yang proporsi
darah Etawahnya sangat beragam.
Selain itu juga terdapat rumpun kambing lain yang berkembang di daerah
tertentu yang merupakan kambing lokal tradisional diantaranya Kambing
Gembrong (di Bali), Kambing Kosta (di Banten), Kambing Bligon, Kambing
Jawarandu (di Jawa Tengah dan Daerah Istimewa Yogyakarta) dan beberapa
jenis kambing lokal lainnya yang belum diidentifikasi secara ilmiah. Setelah
zaman kemerdekaan diimpor atau diintroduksikan beberapa rumpun kambing,
baik dalam bentuk hidup atau mani beku. Rumpun kambing yang pernah
dintroduksikan antara lain Kambing Saanen dan Kambing Anglo Nubian. Bahkan
akhir-akhir ini telah diintroduksikan pula Kambing Boer dari Australia yang
dipersilangkan dengan Kambing Kacang atau Peranakan Etawah dalam bentuk
pejantan hidup atau mani beku (Subandriyo 2004).
Terjadinya persilangan antara kambing impor dengan kambing asli
Indonesia (Kacang) serta adanya aklitimasi dan isolasi selama puluhan bahkan
ratusan tahun di suatu lokasi tertentu dapat menyebabkan terbentuknya
kelompok kambing lokal atau sub populasi dengan komposisi genetik yang unik
pula. Terbentuknya galur/kelompok kambing bisa juga disebabkan terisolasinya
adanya penghanyutan genetik (random genetic drift) seperti dilaporkan Freeland
(2005).
Kambing Muara dijumpai di daerah Kecamatan Muara, Kabupaten
Tapanuli Utara. Berdasarkan pada penampilannya kambing ini nampak gagah,
tubuh kompak dan sebaran warna bervariasi antar warna bulu coklat kemerahan,
putih dan ada juga yang berwarna bulu hitam. Bobot Kambing Muara ini lebih
besar dari pada Kambing Kacang dan diduga mempunyai potensi sebagai ternak
prolifik. Kambing Benggala menurut cerita dari peternak diduga merupakan hasil
persilangan Kambing Black Bengal dengan kambing lokal yang diduga dibawa
pendatang/pedagang dari India, Bangladesh dan Arab ke daerah sekitar Pulau
Timor dan Pulau Flores di Propinsi Nusa Tenggara Timur sebelum zaman
penjajahan Hindia Belanda. Selang waktu yang sudah ratusan tahun persilangan
kambing tersebut mengalami proses adaptasi dengan lingkungan setempat
(Batubara et al. 2007). Kambing Benggala secara umum lebih besar dari
Kambing Kacang, umumnya didominasi warna hitam dan sedikit berwarna
kecoklatan.
Menurut FAO (2000) rumpun adalah bagian kelompok tertentu
(subspecific group) dari ternak domestik dengan karakteristik eksternal yang
dikenal dengan penilaian visual atau kelompok yang dipisahkan oleh geografi
dan budaya secara fenotipik. Rumpun berkembang menurut perbedaan geografi
dan budaya untuk memenuhi kebutuhan yang serupa dan telah diterima sebagai
identitas yang terpisah.
Berdasarkan adaptasi terhadap kondisi lokal rumpun dibedakan atas
rumpun lokal dan rumpun introduksi. Rumpun lokal dapat dibedakan lagi atas
rumpun asli (indigenous breed, native breed) adalah ternak yang berdasarkan
sejarah terbukti berasal dari negara tersebut dan rumpun tradisional (rumpun
lokal) adalah ternak yang sejarahnya tidak terbukti berasal dari negara tersebut
tetapi selama 30-50 tahun telah diternakkan di negara tersebut, terbukti
mempunyai catatan silsilah selama lima generasi. Rumpun introduksi (rumpun
asing, exotic, alocthonous) yang tidak berasal dari suatu negara atau tidak
secara kontinu diternakkan di suatu negara lebih dari 50 tahun (Sapi, kuda) dan
30 tahun untuk ternak lainnya (FAO 2007).
Penetapan dan pengakuan rumpun/galur ternak di Indonesia dilakukan
oleh Pemerintah melalui Kementerian Pertanian. Pemerintah menyusun tatacara
pelepasan rumpun/galur ternak. Istilah “penetapan” adalah sebagai bentuk
pengakuan dari negara terhadap rumpun ternak yang telah ada dan
dibudidayakan secara turun temurun oleh peternak dan menjadi milik masyarakat
(rumpun lokal atau rumpun asli). Istilah “pengakuan” adalah suatu bentuk
pengakuan negara terhadap rumpun dan/atau galur ternak hasil pemuliaan/
introduksi/rekayasa genetik (Puslitbangnak 2007).
Sifat Kuantitatif dan Kualitatif
Penampilan individu yang nampak dari luar disebut sebagai fenotipik,
yang dapat dibedakan menjadi sifat kuantitatif dan kualitatif (Hardjosubroto
2001). Mabrouk et al. (2008) mengemukakan bahwa karakter kuantitatif adalah
ciri-ciri dari mahluk hidup yang dapat diukur, dihitung atau diskor, misalnya
ukuran-ukuran tubuh. Karakter ini ditentukan oleh banyak pasang gen
(poligenik) dan sangat dipengaruhi oleh lingkungan.
Beberapa sifat kuantitatif pada ternak mempunyai hubungan satu sama
lain, hubungan ini secara statistik disebut dengan korelasi. Sifat-sifat yang
berkorelasi menjadi penting karena seleksi terhadap satu sifat akan
menyebabkan kemajuan atau kemunduran bagi sifat lain yang berkorelasi
dengan sifat tersebut (Nsoso et al. 2004).
Berlawanan dengan karakter kuantitatif, karakter kualitatif adalah karakter
yang pada umumnya dijelaskan dengan kata-kata atau gambar. Sifat ini sedikit
sekali atau bahkan tidak ada hubungannya dengan kemampuan produksi,
namun sifat ini mungkin penting sebagai penciri bagi rumpun atau tipe ternak
tertentu, misalnya warna dan pola warna tubuh. Sifat ini diatur oleh satu atau
beberapa pasang gen saja, dan sedikit sekali dipengaruhi oleh lingkungan (Noor
2008).
Sponenberg (2004) mengemukakan bahwa tipe dasar tatawarna bulu
dapat dibedakan menurut: pertama, warna yang meliputi seluruh permukaan
tubuh sehingga membentuk warna seragam atau homogen atau warna tunggal;
dan kedua, heterogen atau campuran. Warna heterogen ini memiliki dua tipe
yang berbeda, yaitu: (a) komposit, apabila pada tubuh ditemukan bidang-bidang
warna yang berbeda (spotted); dan (b) campuran, apabila bulu-bulu dari warna
yang berbeda tampak secara bergantian satu dengan lainnya. Warna pada
kambing umumnya diklasifikasikan kedalam warna tunggal (unicoloured) dan
Penanda Genetik
Penanda adalah karakter yang dapat diwariskan dan berasosiasi dengan
genotip tertentu dan digunakan untuk mengkarakterisasi genotip. Potensi
penggunaan penanda sebagai alat untuk melakukan karakterisasi genetik telah
dikenal sejak puluhan tahun yang lalu. Penanda ini dikategorikan atas penanda
morfologi, sitologi, dan yang terbaru adalah penanda molekuler (Simianer 2006).
a. Penanda Morfologi
Penanda morfologi (fenotipik) merupakan penanda yang telah banyak
digunakan baik dalam program genetika dasar maupun dalam program praktis
pemuliaan, karena penanda ini paling mudah untuk diamati dan dibedakan.
Pengukuran parameter tubuh biasa digunakan untuk menduga asal usul
rumpun ternak. Ukuran-ukuran tubuh sangat berguna untuk menentukan
asal-usul dan hubungan filogenetik antar spesies, rumpun dan tipe ternak yang
berbeda. Beberapa penelitian telah dilakukan menggunakan ukuran-ukuran
tubuh untuk membedakan kelompok Kambing Tswana (Nsoso et al. 2004).
Mabrouk et al. (2008) juga telah melakukan penelitian menggunakan beberapa
ukuran tubuh (bobot badan, panjang badan, tinggi pundak, dalam dada, lingkar
dada, tinggi pinggul, lingkar pinggul, dalam pinggul, panjang ekor, lebar ekor dan
tebal ekor) sebagai peubah pembeda kelompok pada beberapa kelompok
kambing lokal di Tunisia.
Dossa et al. (2007) mengemukakan bahwa kesamaan fenotipik dapat
menunjukkan identitas genetik, walau terdapat beberapa batasan, antara lain:
fenotipik yang identik dapat disebabkan olel alel-alel yang berbeda atau oleh
gen-gen pada lokus yang berbeda. Dalam hal tertentu, mungkin terdapat
perbedaan dalam daya ekspresi (derajat manifestasi pada satu individu) atau
oleh gen dominan (frekuensi satu sifat diekspresikan relatif terhadap sejumlah
pembawa gen tertentu yang diketahui dalam satu populasi). Kemiripan fenotipik
dapat juga disebabkan oleh fenokopi, yakni kemiripan satu fenotip yang
diakibatkan satu genotip tertentu oleh aksi lingkungan pada genotip lainnya.
Namun demikian, penanda ini memiliki kelemahan karena dipengaruhi oleh
lingkungan, memperlihatkan sifat menurun dominan/resesif dan banyak yang
b. Penanda Molekuler
Menurut Cardellino dan Boyazoglu (2009) aplikasi penanda molekuler
yang paling penting adalah untuk pembuatan peta genetik, yang dapat
digunakan untuk memeriksa lokasi suatu gen yang bertanggung jawab terhadap
suatu sifat yang sederhana, misalnya resistensi terhadap penyakit atau sifat
kuantitatif yang komplek pada kromosom. Penanda molekuler ini ada pada
tingkat DNA, maka penanda ini bebas dari pengaruh-pengaruh epistasis,
lingkungan dan fenotip sehingga dapat menyediakan informasi genetik yang
defenitif untuk digunakan dalam mempelajari keragaman genetik, mendeteksi
gen-gen major dan mempelajari sifat-sifat genetik yang komplek.
Tehnik ini sangat membantu pemulia dalam melakukan studi genetik
dengan ketepatan yang tinggi. Untuk mendapatkan informasi genetik dapat
dilakukan dengan menggunakan penanda molekuler, seperti isozim, RFLP
(restriction fragment length polymorphism), RAPD (random amplified polimorphic
DNA), AFLP (amplified fragment length polymorphism) dan lain-lainnya.
Penanda molekuler terbaru yang relatif mudah diamati adalah DNA mikrosatelit.
DNA Mitokondria
Organisme eukariot termasuk ternak domestik, sumber DNA dapat
diperoleh oleh organel-organel sitoplasmik antara lain DNA mitokondria. DNA
mitokondria memiliki karakteristik sebagai molekul DNA yang diturunkan secara
utuh tanpa adanya rekombinasi, memiliki molekul dengan ukuran kecil/pendek
yang susunannya berbeda dengan DNA inti (Lewin 2000) dan memiliki variasi
basa nukleotida yang lebih tinggi dibandingkan dengan DNA inti. Tingginya
variasi basa nukleotida disebabkan DNA mitokondria memiliki laju perubahan
5-10 kali lebih tinggi dibandingkan DNA inti (Muladno 2006; Fan-Bin 2007). DNA
mitokondria terutama daerah D-loop, sangat baik digunakan untuk analisis
keragaman hewan, baik di dalam spesies maupun antar spesies (Muladno 2006).
Setiap sel mengandung satu hingga ratusan DNA mitokondria. DNA
mitokondria merupakan DNA utas ganda yang berbentuk sirkuler (Freeland
2005), mengandung sejumlah gen penting untuk respirasi dan pembentukan
energi sel tubuh dan fungsi lainnya, sehingga relatif lebih mudah untuk
mengisolasi nukleotidanya dari genom (MacHugh & Bradley 2001). Genom
mitokondria hewan berukuran relatif kecil dan terdapat dalam jumlah banyak,
DNA mitokondria (mtDNA) mempunyai beberapa kelebihan yang
menjadikannya banyak digunakan untuk mengidentifikasi keanekaragaman
genetik dan dinamika populasi. Beberapa kelebihan tersebut adalah (1) memiliki
ukuran yang kompak dan relatif kecil (16 000 – 20 000 pasang basa), tidak
sekomplek DNA inti sehingga dapat dipelajari sebagai satu kesatuan yang utuh;
(2) berevolusi lebih cepat dibandingkan dengan jelas perbedaan antara populasi
dan hubungan kekerabatannya; (3) hanya sel telur yang menyumbangkan
material mitokondria sehingga mitokondria DNA hanya diturunkan dari induk
betina; dan (4) bagian-bagian dari genom mitokondria berevolusi dengan laju
yang berbeda, sehingga dapat berguna untuk studi sistematika dan penelusuran
kesamaan asal-usul. DNA mitokondria telah banyak digunakan sebagai
penanda molekul untuk studi genetika populasi, penelusuran asal-usul dan
pelacakan beberapa penyakit degeneratif, penuaan serta kanker (Wandia 2001).
DNA mitokondria telah dikarakterisasi dengan lebih baik pada sebagian besar
ternak dan telah digunakan untuk studi evolusi (Freeland 2005).
Tingkat evolusi dari suatu gen atau bagian DNA yang berbeda
merupakan faktor penting yang menentukan penggunaan penanda DNA dalam
studi sistematika dan biogeografi. Umumnya, gen-gen yang terkonservasi
dengan baik (berevolusi lambat) dapat dijadikan dasar penelusuran asal-usul
atau filogeni. Sebaliknya, gen-gen yang tidak terkonservasi dengan baik
(berevolusi cepat) dapat digunakan untuk perbandingan galur-galur baru
(Chen et al. 2005).
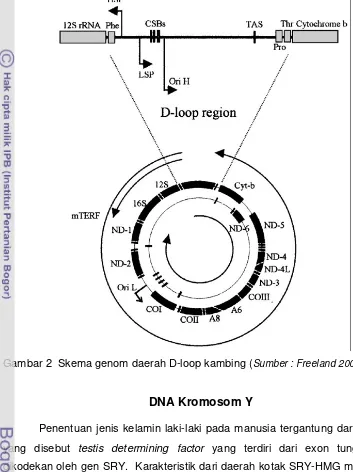
DNA mitokondria hewan secara umum memiliki jumlah dan jenis gen
yang sama yaitu 13 daerah yang mengkode protein (URF1, URF2, URF3, URF4,
URF5, URF6, URF6L, URF4L, Cytochrome Oxidase unit I, Cytochrome Oxidase
unit II, Cytochrome Oxidase unit III, Cytochrome –b dan ATPase 6); 2 gen
pengkode rRNA yaitu 12S rRNA dan 16S rRNA; 22 gen pengkode tRNA
(Freeland 2005). Perkembangan sekarang ini ke-8 URF adalah diidentifikasi
menjadi gen-gen 7 sub unit NADH-dehidrogenase (ND 1-6 dan ND 4L) dan sisa
ATPase 8 (Lewin 2000). Daerah bukan pengkode, hanya terdiri atas daerah
kontrol (control region) yang memegang peranan penting dalam proses
transkripsi dan replikasi genom mitokondria. Pada mamalia, daerah bukan
pengkode meliputi daerah bukan pengkode utama yang merupakan tempat awal
replikasi H strand (OH). Daerah bukan penyandi utama terletak pada wilayah
pengkode segmen minor yaitu tempat awal replikasi L strand (OL) yang terletak
[image:36.595.111.464.154.626.2]pada gugus gen tRNA antara gen CO I dan ND 2.
Gambar 2 Skema genom daerah D-loop kambing (Sumber : Freeland 2005)
DNA Kromosom Y
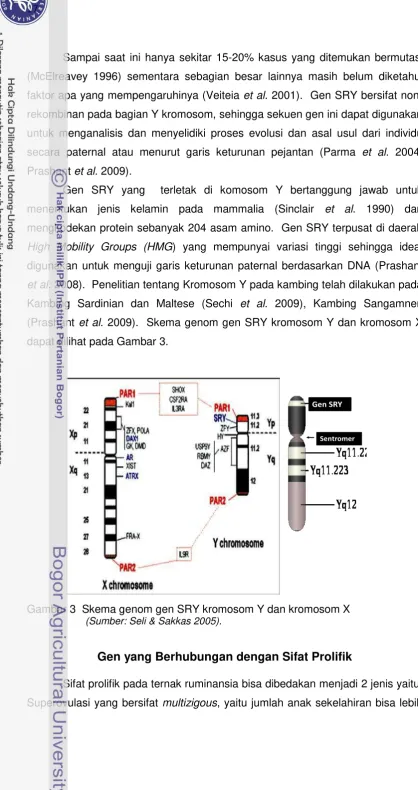
Penentuan jenis kelamin laki-laki pada manusia tergantung dari aktivitas
yang disebut testis determining factor yang terdiri dari exon tunggal dan
dikodekan oleh gen SRY. Karakteristik dari daerah kotak SRY-HMG merupakan
target ideal untuk pengembangan dari uji penentuan jenis kelamin berdasarkan
DNA (Prashant et al. 2008). Mutasi pada gen SRY berperan menentukan XY
Gonadal Dysgenesis (XYGD). Fenotip XYGD yang telah dilaporkan antara lain
pada manusia (Cohen dan Shaw 1965), kuda (Power 1986), sapi (Kawakura et
Sampai saat ini hanya sekitar 15-20% kasus yang ditemukan bermutasi
(McElreavey 1996) sementara sebagian besar lainnya masih belum diketahui
faktor apa yang mempengaruhinya (Veiteia et al. 2001). Gen SRY bersifat
non-rekombinan pada bagian Y kromosom, sehingga sekuen gen ini dapat digunakan
untuk menganalisis dan menyelidiki proses evolusi dan asal usul dari individu
secara paternal atau menurut garis keturunan pejantan (Parma et al. 2004;
Prashant et al. 2009).
Gen SRY yang terletak di komosom Y bertanggung jawab untuk
menentukan jenis kelamin pada mammalia (Sinclair et al. 1990) dan
mengkodekan protein sebanyak 204 asam amino. Gen SRY terpusat di daerah
High Mobility Groups (HMG) yang mempunyai variasi tinggi sehingga ideal
digunakan untuk menguji garis keturunan paternal berdasarkan DNA (Prashant
et al. 2008). Penelitian tentang Kromosom Y pada kambing telah dilakukan pada
Kambing Sardinian dan Maltese (Sechi et al. 2009), Kambing Sangamneri
(Prashant et al. 2009). Skema genom gen SRY kromosom Y dan kromosom X
[image:37.595.58.477.34.825.2]dapat dilihat pada Gambar 3.
Gambar 3 Skema genom gen SRY kromosom Y dan kromosom X
(Sumber: Seli & Sakkas 2005).
Gen yang Berhubungan dengan Sifat Prolifik
Sifat prolifik pada ternak ruminansia bisa dibedakan menjadi 2 jenis yaitu:
Superovulasi yang bersifat multizigous, yaitu jumlah anak sekelahiran bisa lebih
Gen SRY
dari 2 bahkan bisa sampai beranak lima ekor dalam satu periode kelahiran,
biasanya dijumpai pada ternak ruminansa kecil seperti domba dan kambing, dan
yang bersifat monozigous, yaitu pada ternak ini ada kemungkinan satu sel telur
berkembang dengan cara membelah menjadi dua, biasanya ini dijumpai pada
ternak ruminansia besar. Folikel merupakan titik awal yang perlu disoroti
sebagai salah satu ”pabrik” penghasil sel telur dan penghasil hormon conseptus
(kebuntingan) maupun mammogenic (kelenjar susu), bahkan secara genetik
dapat digunakan sebagai alat seleksi untuk memperoleh ternak unggul
berdasarkan jumlah anak yang dilahirkan. Selama ini dibidang fisiologi
reproduksi telah terjadi inefisiensi pemanfaatan folikel pada induk betina. Kedua
ovarium betina mengandung 40 000 – 300 000 folikel bahkan lebih tergantung
pada jenis ternak. Pada ruminasia (sapi,domba,kambing) berkisar 50 000 – 150
000 folikel (Hafez 1993), namun selama hidupnya ternyata hanya beberapa
folikel yang berovulasi, sehingga ratusan ribu sisanya tidak termanfaatkan.
Folikel baik sebelum maupun sesudah ovulasi menjadi corpus luteum merupakan
organ transitor yang memiliki peran regulator di dalam proses reproduksi,
terutama sebagai sumber penghasil sel telur maupun hormon conseptus
endogen (Gemmell 1995).
Kematian anak dari tipe kelahiran 3 atau lebih sangat tinggi (Inounu et al.
1993), sehingga potensi keuntungan dari betina dengan jumlah anak banyak ini
tidak terlihat. Betina-betina yang merawat anak banyak membutuhkan tingkat
nutrisi yang sangat baik pada fase sebelum dan sesudah kelahiran dan juga
memerlukan perawatan serta perhatian yang lebih banyak, jika persentase anak
yang hidup ingin dicapai. Tehnik laparoskopi dapat dilakukan pada ternak
kambing betina untuk mengamati produksi sel telur (laju ovulasi) dengan cara
menghitung jumlah corpus luteum (CL) yang dihasilkan. Batas atas keragaan
reproduksi adalah jumlah sel telur yang dihasilkan oleh seekor induk per satu
siklus birahi, yang diamati dengan cara menghitung jumlah corpus luteum (badan
kuning) dari kedua indung telur pada hari ke 3-10 setelah birahi. Faktor
lingkungan merupakan faktor pendukung apakah batas atas tersebut dapat
dicapai. Laju ovulasi adalah rataan jumlah sel telur yang dihasilkan oleh seekor
induk setiap siklus birahi. Di dunia ini ada beberapa rumpun kambing yang
sangat prolifik, ditandai dengan laju ovulasi dan jumlah anak sekelahiran tinggi
yang bisa melahirkan anak 3-5 ekor anak per kelahiran. Karakterisasi genetik
ekor tipis, Merino, Thoka, Lacaune, Cambridge (Davis et al. 2002); Awassi dan
Assap (Gootwine et al. 2008); Han ekor pendek (Chu et al. 2006); Rumpun
Aragonesa (Rovo 2008), sedangkan untuk jenis ternak kambing baru dilaporkan
pada Kambing Jining Grey (Chu et al. 2007). Meskipun demikian, kedua jenis
ternak tersebut seringkali ditemukan mempunyai jumlah anak per kelahiran yang
lebih dari satu (Odubute et al. 1992). Lan (2007) melaporkan bahwa laju ovulasi
dan litter size pada Domba Booroola dari Merino Australia disebabkan alel
FecBB dari major gene yang disebut FecB. Ditemukan pula bahwa gen utama
tersebut berada pada ovine chromosome 6 yang juga merupakan posisi dari gen
yang menyandikan salah satu anggota growth transforming factor-β, yaitu bone morphogenetik protein receptor 1B (BMPR1B). Hanrahan et al. (2004) kemudian
menambahkan bahwa ada juga jenis gen lain yang berasosiasi dengan laju
ovulasi dan litter size, yaitu Oocyte- growth derived factors GDF9 dan bone
morphogenetik protein 15 pada Domba Cambridge dan Berclare. Dalam
perkembangan berikutnya, ternyata ketiga jenis gen tersebut diketahui beraksi
sebagai gen-gen utama yang mengatur tingkat laju ovulasi pada kisaran taksa
mamalia yang luas, mulai dari manusia, sapi, tikus, dan kucing (Davis 2004;
2005) yang kemudian diikuti dengan penamaan gen yang bersinonim. Jadi
dalam keluarga transforming growth factor-β yang berasosiasi dengan sifat-sifat prolifik adalah bone morphogenetik protein receptor type 1B (EU 581862
BMPR1B, activin-like kinase 6, atau FecB) pada ovine chromosome 6, growth
differentiation factor 9 (= GDF9, Oocyte-derived growth factors, atau FecG) pada
ovine chromosome 5, dan bone morphogenetik protein 15 (BMP15, atau FecX)
pada ovine chromosome X. Selain ketiga gen fekunditas yang populer pada
domba, gen prolactin receptor juga diduga berasosiasi dengan sifat prolifik pada
Kambing Jining Grey (Zhang et al. 2007) dan pada manusia dan sapi.
Berbagai analisis terhadap pola-pola mutasi dari gen-gen fekunditas dan
asosiasinya dengan laju ovulasi, litter size dan berbagai efisiensi reproduksi
lainnya banyak menarik perhatian para pemulia ternak (Davis 2005). Akibatnya
bisa dipastikan bahwa pola-pola mutasi nukleotida pada gen-gen fekunditas
diatas memunculkan berbagai penanda molekular yang diasosiasikan dengan
KARAKTERISASI MORFOMETRIK DAN ANALISIS
FILOGENI PADA ENAM SUB POPULASI KAMBING LOKAL
INDONESIA
Pendahuluan
Berdasarkan Statistik Tahun 2010 jumlah populasi ternak kambing di
Indonesia sebanyak 16 841 149 ekor, paling tinggi di provinsi Jawa Tengah
(3 491 073 ekor), Jawa Timur (2 822 534 ekor), Jawa Barat (1 825 748 ekor),
Lampung (1 206 000 ekor), Banten (854 522 ekor), NAD (886 468 ekor),
Sumatera Utara (621 492 ekor), NTT (556 190 ekor) dan Sulawesi Selatan
(442 297 ekor). Hampir 99% ternak ruminansia kecil di Indonesia merupakan
skala usaha ternak kecil (Soedjana 2008). Sekitar 95% penduduk Indonesia
adalah Muslim, ruminansia kecil mempunyai peranan penting pada kegiatan
keagamaan terutama perayaan Idul Adha. Ternak kambing dapat mengkonversi
hijauan berkualitas rendah menjadi protein hewani, sebagai sumber pupuk
kandang serta sebagai tabungan.
Parameter fenotipik merupakan metode yang paling mudah digunakan
untuk mengidentifikasi karakterisitik ternak ruminansia (Alade et al. 2008; Khan
et al. 2006; Dossa et al. 2007; Jimmy et al. 2010). Perbedaan penampilan
disebabkan selama domestikasi tipe-tipe atau rumpun-rumpun hewan terpisah
fenotipik secara genetik karena adanya proses adaptasi (ekpresi gen) dengan
lingkungan lokal dan kebutuhan komunitas lokal sehingga dihasilkan rumpun
yang berbeda. Adanya kemampuan adaptasi hewan disebabkan hewan memiliki
kemampuan menghasilkan lebih dari satu alternatif bentuk morfologi, status
fisiologi dan atau tingkah laku sebagai reaksi atau upaya adaptasi terhadap
perubahan lingkungan berupa pengaturan ekspresi gen dan perubahan bentuk
fenotip (Riva et al. 2004; Mansjoer et al . 2007; Noor 2008; Karna et al. 2001).
Mendukung upaya pelestarian dan pemanfaatan ternak kambing lokal
secara berkelanjutan maka perlu diketahui karakteristik fenotipik dan potensi
produksi ternak yang ada di Indonesia. Tujuan penelitian ini adalah untuk
mendapatkan data dan informasi tentang karakteristik morfometrik dan jarak
Bahan dan Metode
Penelitian ini menggunakan beberapa sub populasi kambing lokal
Indonesia yang disebut Kambing Samosir, Kambing Muara, Kambing Marica,
Kambing Jawarandu, Kambing Benggala dan Kambing Kacang. Penentuan
sampel kambing dengan metode purposive sampling, yaitu pertama menentukan
Kabupaten daerah sentra produksi di setiap Propinsi, baru kemudian ditentukan
Kecamatan dan Kelompok Desa. Setiap sub populasi diambil sekitar 34 - 217
ekor kambing sebagai sampel sesuai dengan ketersediaan populasi ternak yang
bisa ditemui di lapangan dan diusahakan diambil dari desa yang jauh
kekerabatan/keturunan sampel kambing dengan sampel pada lokasi desa
pengambilan lainnya.
Peralatan penelitian yang digunakan yaitu tongkat ukur ketelitian 0.1 cm,
pita ukur ketelitian 0.1 cm, jangka sorong stainless steel buatan Jerman,
timbangan gantung (shelter) dengan ukuran kg dengan tingkat ketelitian 50
gram, kamera digital Nikon F-9. 8 mega pixel, dan tali rapiah pengikat kambing.
Waktu dan Tempat Penelitian
Penelitian lapangan untuk koleksi data fenotipik dilakukan pada bulan
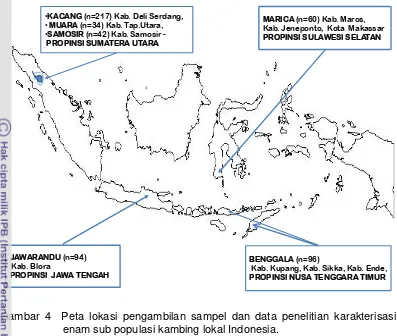
Maret 2009 sampai Maret 2011 di empat Propinsi yaitu:
1. Propinsi Sumatera Utara : Kabupaten Samosir, Kabupaten Tapanuli Utara,
Kabupaten Deli Serdang.
2. Propinsi Jawa Tengah; Kabupaten Blora.
3. Propinsi Sulawesi Selatan: Kabupaten Maros, Kota Makassar, Kabupaten
Jeneponto.
4. Propinsi Nusa Tenggara Timur : Kota Kupang, Kabupaten Kupang,
Gambar 4 Peta lokasi pengambilan sampel dan data penelitian karakterisasi enam sub populasi kambing lokal Indonesia.
Pengumpulan Sampel Kambing
Teknik pengambilan sampel ternak kambing dilakukan secara acak pada
543 ekor kambing, yaitu 96 ekor Kambing Benggala (betina=89, jantan=7), 94
ekor Kambing Jawarandu (betina=72, jantan=22), 60 ekor Kambing Marica
(betina=48, jantan=12), 217 ekor Kambing Kacang (betina=193, jantan=24), 34
ekor Kambing Muara (betina=28, jantan=6) dan 42 ekor Kambing Samosir
(betina=36, jantan=6).
Pengumpulan data fenotipik dilakukan bersamaan dengan pengambilan
sampel darah. Koleksi data dimulai dengan pencatatan jenis kelamin dan umur
kambing serta nama pemiliknya. Umur kambing penelitian ditentukan
berdasarkan minimal sudah terdapat 1 pasang gigi seri yang permanen.
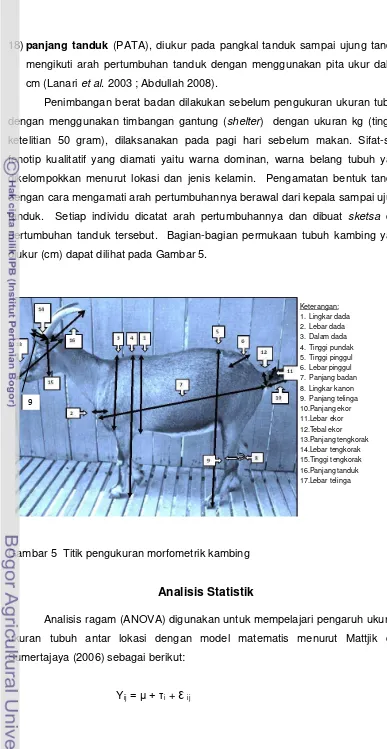
Parameter fenotipik yang digunakan dalam analisis data meliputi :
1) lingkar dada (LIDA), diukur melingkar tepat di belakang scapula, dengan
menggunakan pita ukur dalam cm;
JAWARANDU (n=94) Kab. Blora
PROPINSI JAWA TENGAH
•KACANG (n=217) Kab. Deli Serdang,
•MUARA (n=34) Kab. Tap.Utara,
•SAMOSIR (n=42) Kab. Samosir -PROPINSI SUMATERA UTARA
MARICA (n=60) Kab. Maros, Kab. Jeneponto, Kota Makassar PROPINSI SULAWESI SELATAN
BENGGALA (n=96)
2) lebar dada (LEDA), diukur antara tuberitas humeri sinister dan dexter,
dengan menggunakan tongkat ukur dalam cm;
3) dalam dada (DADA), diukur dari bagian tertinggi pundak sampai dasar dada,
dengan menggunakan tongkat ukur dalam cm;
4) tinggi pundak (TIPU), diukur dari bagian tertinggi pundak melalui belakang
scapula tegak lurus ke tanah, dengan menggunakan tongkat ukur dalam cm;
5) tinggi pinggul (TIPI), diukur dari bagian tertinggi pinggul secara tegak lurus
ke tanah, dengan menggunakan tongkat ukur dalam cm;
6) lebar pinggul (LEPI), diukur dengan jarak lebar antara kedua sendi pinggul
dengan menggunakan tongkat ukur dalam cm;
7) panjang badan (PABA), diukur dari tuber ischii sampai tuberitas humeri,
dengan menggunakan tongkat ukur dalam cm;
8) lingkar kanon (LIKA), diukur tepat melingkar pada bagian tulang canon kaki
belakang sebelah kiri dengan menggunakan pita ukur dalam cm;
9) lebar kanon (LEKA) diukur jarak antar tulang kering lutut dengan tulang
kanon; dengan menggunakan pita ukur dalam cm
10) panjang ekor (PAEK), diukur pada pangkal sampai ujung ekor, dengan
menggunakan pita ukur dalam cm
11) lebar ekor (LEEK), diukur lebar ekor pada bagian paha ekor, dengan
menggunakan jangka sorong dalam cm;
12) tebal ekor (TEEK), diukur tebal pada bagian pangkal ekor, dengan
menggunakan jangka sorong dalam cm;
13) panjang telinga (PATEL), diukur pada pangkal telinga sampai ujung telinga;
dengan menggunakan pita ukur dalam cm
14) <