IDENTIFIKASI GEN PROLAKTIN EKSON DUA DAN PRODUKSI
TELUR PADA ITIK PERSILANGAN PEKIN MOJOSARI
PUTIH (PMp) GENERASI KELIMA
IRMA
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN SUMBER
INFORMASI SERTA PELIMPAHAN HAK CIPTA
Dengan ini saya menyatakan bahwa tesis berjudul Identifikasi Gen Prolaktin Ekson Dua dan Produksi Telur pada Itik Persilangan Pekin Mojosari Putih (PMp) Generasi Kelima adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apapun kepada perguruan tinggi manapun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, 21 Agustus 2014
Irma
RINGKASAN
IRMA. Identifikasi Gen Prolaktin Ekson Dua dan Produksi Telur pada Itik Persilangan Pekin Mojosari Putih (PMp) Generasi Kelima. Dibimbing oleh CECE SUMANTRI dan TRIANA SUSANTI.
Itik persilangan Pekin Mojosari putih (PMp) merupakan hasil persilangan antara itik Pekin jantan dan itik betina Mojosari putih yang dilakukan di Balai Penelitian Ternak Ciawi. Generasi itik persilangan PMp telah mencapai generasi kelima. Pengamatan produksi dan kualitas telur serta keragaman gen prolaktin ekson dua terhadap itik persilangan PMp dilakukan dalam rangka melihat mutu ternak sebagai calon bibit unggul pedaging. Penelitian tersebut dilaksanakan di Balai Penelitian Ternak Ciawi dan Laboratorium Genetika Molekuler Ternak Fakultas Peternakan IPB sejak April 2013 sampai Januari 2014.
Materi penelitian berasal dari 168 itik milik Balai Penelitian Ternak Ciawi. Itik terdiri dari 19 ekor Pekin, 36 ekor Mojosari dan 113 ekor PMp. Sebanyak 98 itik persilangan PMp betina digunakan untuk pengamatan produksi dan kualitas telur. Ternak dipelihara pada kondisi lingkungan kandang dan pakan yang sama. Peubah kualitas terdiri dari berat telur utuh, bentuk atau indeks telur, berat putih telur, berat dan warna kuning telur, tebal dan berat kerabang, serta nilai Haugh Unit (HU). Produksi telur itik dikoleksi selama enam bulan pertama. Berat telur utuh, putih telur, kuning telur dan HU diukur dengan alat Egg AnalyzerTM. Tebal kerabang diukur dengan alat Egg Shell ThicknessTM sedangkan berat kerabang ditimbang dengan timbangan digital. Warna kuning telur dinilai dengan alat
Roche Yolk Colour Fan.
Sampel darah sebanyak 1-1.5 ml/ekor digunakan untuk analisis gen prolaktin ekson dua. Sampel darah diekstrak dengan menggunakan kit (Mini Genomic, Qiagen) sesuai dengan protokol yang tercantum. Hasil ekstraksi dianalisis kualitas dan kuantitas menggunakan spektrofotometer. Ekstraks DNA digunakan untuk proses PCR (Polymerase Chain Reaction). Primer disusun berdasarkan database sekuens prolaktin itik (Kode Akses GenBank AB158611.1) dengan bantuan softwareprimer3. Kondisi PCR yaitu suhu: pradenaturasi95°C (5 menit), denaturasi95°C (10 detik), suhu penempelan (annealing)60°C (20 detik), suhu ekstensi awal 72°C (30 detik), ekstensi akhir 72°C (5 menit) dan 35 siklus. Visualisasi hasil amplifikasi dilakukan melalui elektroforesis dengan menggunakan metode gel agarosa 1.5%. Pita-pita yang muncul dibandingkan dengan posisi marka untuk penentuan panjang basa. Sekuensing bagian forward
dilakukan pada seluruh sampel. Sekuens gen prolaktin ekson dua dilakukan oleh jasa sekuensing dengan menggunakan mesin sekuenser (ABI PRISM Genetic Analyzer) di First Base, Selangor, Malaysia.
Hasil penelitian menunjukan rataan produksi telur individu periode enam bulan yaitu 67%, bobot telur 59.09 g, indeks telur 73.8%, berat kuning telur 19.25 g, berat putih telur 33.02 g, nilai HU 101, warna kuning telur 8, berat kerabang 6.84 g dan tebal kerabang 0.4 mm. Kemurnian dan konsentrasi hasil ekstraksi yaitu 1.81 dan 263.34 µg/ml. Produk PCR berhasil diamplifikasi dengan panjang amplikon sekitar 400 pb. Terdapat satu insersi basa adenin di lokasi 2001 pb di intron dua pada keseluruhan itik yang digunakan. Gen prolaktin ekson dua dan parsial pada keseluruhan sampel hanya terdapat satu tipe genotipe dengan frekuensi alel 100%. Uji homologi menunjukan 99% kesamaan dengan sekuens gen prolaktin ekson dua dan parsial intron itik. Semua sampel itik yang digunakan dalam keadaan homozigot dan monomorfik. Kondisi homozigot dan monomorfik menjadikan gen prolaktin ekson dua tidak dapat dijadikan marka biomolekuler untuk sifat produksi dan kualitas telur itik PMp.
SUMMARY
IRMA. Identification of Prolactin Gene Exon Two and Egg Production in Pekin Mojosari Putih (PMp) Crossbred. Supervised by CECE SUMANTRI and TRIANA SUSANTI.
Pekin Mojosari Putih (PMp) is a crossbred duck between Pekin drake and Mojosari duck which conducted in Institute of Research in Animal Production (IRIAP) Ciawi. Recently, PMp generations reach out for fifth generation. Egg quality and production, and prolactin exon two observation in PMp was conducted to evaluates quality of waterfowl as superior breed candidate. The research has been performed in Institute of Research of Animal Production (IRIAP) Ciawi dan Laboratory of Animal Molecular Genetic Faculty of Animal Science, Bogor Agriculture University from April 2013 until January 2014.
Research materials comes from 168 ducks from IRIAP. Ducks consist of 19 Pekin, 36 Mojosari Putih and 113 of PMp crossbred. A total of 98 duck PMp was used for egg quality and production observation. All of ducks are keep in same backyard environment condition and feeds. Variables observation consist of egg production and quality of first laying egg. Egg quality observations involved : egg weight, egg shape or index, albumin weight, yolk weight, yolk colour, eggshell thickness, eggshell weight and Haugh Unit (HU). Egg production measured for six months periode from first laying. Egg weight, albumin weight, yolk weight and HU measured by using Egg AnalyzerTM. Eggshell thickness measured by using Eggshell ThicknessTM, eggshell weight measured by using digital scale weighting. Yolk colour determined by using 15 score Roche Yolk Colour Fan.
A total of 1-1.5 ml blood samples from individual duck was used for exon two of prolactin gene analysis. Blood samples was extracted by using Mini Genomic Kit (Qiagen). Evaluation of quality and quantity of genome extraction was performed by using spectrophotometre. Extraction quality indicated by DNA purity while quantity of extraction indicated by concentration. DNA extraction was used as DNA template for PCR (Polymerase Chain Reaction). Primer designed based on whole prolactin sequence database (GeneBank Code Access: AB158611.1) by using Primer3 software. PCR condition were: predenaturation temperature 95°C (5 min), denaturation 95°C (10 sec), annealing 60°C (20 sec), early extention 72°C (30 sec) and final extention 72°C (20 sec) with 35 cycles. Visualization of amplified PCR was performed by electrophoresis in 1.5% agarose gell. Bands position were compared to the marker to determine bases size. Sequencing for forward sequences was performed in all of samples. Sequencing was conducted in First Base, Malaysia, by using ABI PRISM Genetic Analyzer.
Descriptive analysis was conducted to egg quality and production data. Data analysis for prolactin gene exon two was conducted by alleles frequention and sequence analysis approach. Genetic mutation was identified by using Moleculer Evolutionary Genetics Analysis (4th Version) software. DNA sequences were analyze by Bioedit Sequence Alligment Editor. Homology test between prolactin exon two sequence and duck prolactin database in genebank (GeneBank Code Access: AB158611.1) by using on line Basic Local Alignment Search Tool
The result show that average egg production for six month periode was 67%, egg weight 59.09 g, egg index 73.8%, yolk weight 19.25 g, albumin weight 33.02 g, Haugh Unit 101, yolk colour 8, eggshell weight 6.84 g, and eggshell thickness 0.4 mm. DNA purity and concentration were 1.81 and 263.34 µg/ml, respectively. PCR product length approximately 400 bp. There is one adenin base insertion in 2001 bp location at intron two in all of duck samples. There only one genotype with 100% allele frequention. Homology test showed 99% identity with exon two and partial intron of prolactin gene in genebank database (GeneBank Code Access: AB158611.1). All of duck samples were homozigot and monomorphic. Prolactin gene exon two and partial intron based on this condition cannot become as biomarker for egg production and egg quality for PMp crossbred.
Keywords : Mojosari, Pekin, Pekin Mojosari crossbred, prolactin, egg production, egg quality.
© Hak Cipta Milik IPB, Tahun 2014
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah: dan pengutipan tersebut tidak merugikan IPB
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains
pada
Program Studi Ilmu Produksi dan Teknologi Peternakan
IDENTIFIKASI GEN PROLAKTIN EKSON DUA DAN
PRODUKSI TELUR PADA ITIK PERSILANGAN PEKIN
MOJOSARI PUTIH (PMp) GENERASI KELIMA
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
BOGOR
2014
Judul Tesis : Identifikasi Gen Prolaktin Ekson Dua dan Produksi Telur pada Itik Persilangan Pekin Mojosari Putih (PMp) Generasi Kelima
Nama : Irma
NIM : D151120141
Disetujui oleh Komisi Pembimbing
Prof Dr Ir Cece Sumantri, MScAgr Ketua
Dr Ir Triana Susanti, MSi Anggota
Diketahui oleh
Ketua Program Studi Ilmu Produksi dan Teknologi Peternakan
Dr Ir Salundik, MSi
Dekan Sekolah Pascasarjana
Dr Ir Dahrul Syah, MScAgr
PRAKATA
Bismillahirrahmanirrahim. Assalamu’alaikum wa rahmatullahi wa barakatuhu. Puji dan syukur penulis panjatkan kehadirat Allah SWT atas rahmat dan karuniaNya sehingga dapat menyelesaikan tesis ini sebagai tugas akhir dalam memperoleh gelar Magister Sains pada Sekolah Pascasarjana Fakultas Peternakan Institut Pertanian Bogor. Shalawat dan salam tak lupa disampaikan kepada Nabi Muhammad SAW, keluarganya, para sahabatnya, dan kepada kita semua yang senantiasa mengikuti sunnah-sunnahnya. Amin ya rabbal a’lamin.
Tesis ini merupakan hasil riset mengenai identifikasi gen prolaktin ekson dua dan produksi telur pada itik persilangan Pekin Mojosari putih (PMp) generasi kelima. Riset dilaksanakan di Kompleks Itik, Balai Penelitian Ternak Ciawi dan Laboratorium Genetika Molekuler Ternak Fakultas Peternakan IPB sejak bulan April 2013 sampai Januari 2014. Sumber dana riset dan sampel itik yang digunakan berasal dari Balai Penelitian Ternak Ciawi. Penulis mengucapkan terimakasih kepada Biro Perencanaan dan Kerjasama Luar Negeri (BPKLN) Kementrian Pendidikan Nasional atas Beasiswa selama studi Pascasarjana di IPB.
Penyusunan tesis ini tidak lepas dari dukungan dan bantuan dari banyak pihak. Ucapan terimakasih kepada kedua orang tua tercinta, Bapak H. Ridhwan Shafiyan dan Ummi Siti Maryati atas doa, semangat dan dukungannya yang tiada henti. Semoga karya tulis ini berbuah pahala bagi mereka. Penulis menyampaikan terimakasih yang sebesar-besarnya kepada komisi pembimbing, Prof Dr Ir Cece Sumantri, MScAgr dan Dr Ir Triana Susanti, MSi yang telah banyak mengarahkan penulis sejak riset dimulai hingga selesai. Terimakasih juga disampaikan kepada Ibu Rahmawati Pulungan dan Bapak Indra, tim redaktur dari Jurnal Ilmu Ternak dan Veteriner (JITV) Pusat Penelitian dan Pengembangan Peternakan Bogor yang telah banyak membantu dalam proses publikasi jurnal ilmiah.
Ucapan terimakasih yang sebesar-besarnya juga disampaikan kepada rekan-rekan di Laboratorium Genetika dan Molekuler Ternak Fakultas Peternakan IPB (Erik Andreas, SPt, MSi, rekan-rekan Animal Breeding and Genetics Club: Tia Irmayanti Amalianingsih, SPt, MSi, Marlinda Indriati, SPt, MSi, Fuad Hasan Harahap, SPt, MSi, Annisa Oktavia Rini, SPt, MSi, Wike Andre Septian, SPt, Sri Wahyuni Siswanti, SPt, MSi, Pandu Permata Sari, SPt, Raden Jatu Winantoro, SPt, Komang Alit P, SPt, MSi, Isyana Khaerunnisa, SPt, Ahmad Furqon, SPt dan Selvi, SSi), staf Kompleks Itik Balitnak Ciawi (Bapak Hamdan dan H. Miftah) dan staf Laboratorium Puslitbangnak Pajajaran (Ibu Ella dan Ibu Anne). Kepada Dept. IPTP (Prof Dr Ir Muladno, MSA, Dr Ir Salundik, MSi, Dr Ir Niken Ulupi, MSi, Ibu Ade dan Mb Okta) yang telah banyak membantu selama di pascasarjana Fapet IPB. Tidak lupa pula saya ucapkan terimakasih kepada Dr Yulnawati, SKH, MSi dan Dr Ir Jakaria SPt, MSi atas diskusinya yang membangun. Semoga karya ilmiah ini dapat memberikan manfaat. Wassalamu’alaikum wa rahmatullahi wa barakatuhu.
Bogor, 21 Agustus 2014
DAFTAR ISI
Produksi Telur Periode Enam Bulan 11
Kualitas Telur Pertama Itik PMp 13
Gen Prolaktin Ekson Dua 16
Amplifikasi Gen Prolaktin Ekson Dua 16
Identifikasi Mutasi Gen Prolaktin Ekson Dua 17
Homologi Gen Prolaktin Ekson Dua 18
Komposisi Nukleotida Gen Prolaktin Ekson Dua 19 Frekuensi dan Keragaman Alel Gen Prolaktin Ekson Dua 20
5 SIMPULAN DAN SARAN 20
Simpulan 20
Saran 20
DAFTAR PUSTAKA 21
DAFTAR TABEL
1. Titik-titik mutasi pada gen prolaktin itik 7 2. Karakteristik kualitas telur pertama itik PMp generasi kelima 14 3. Identifikasi mutasi hasil riset dengan referensi 18 4. Komposisi basa penyusun gen prolaktin ekson dua 19
DAFTAR GAMBAR
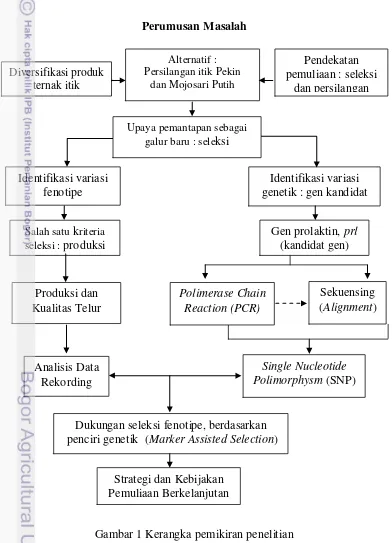
1. Bagan kerangka penelitian 2
2. Itik Mojosari putih di Balitnak 4
3. Itik Pekin di Balitnak 5
4. Itik persilangan PMp di Balitnak 6
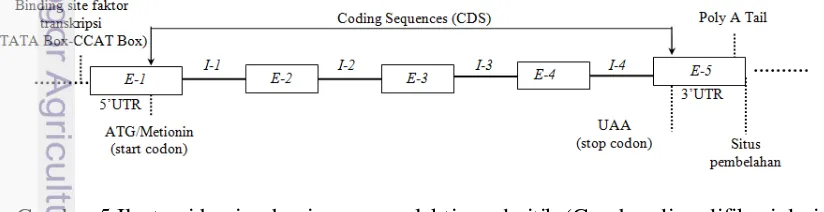
5. Ilustrasi gen prolaktin pada itik 6
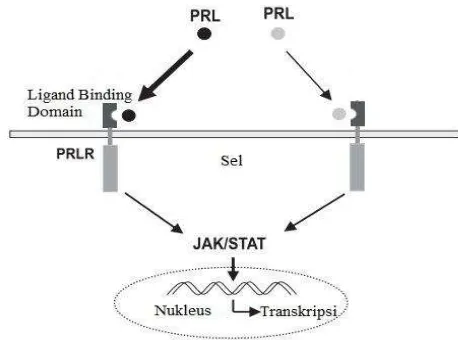
6. Jalur sinyal gen prolaktin 8
7. Dimensi pengukuran bentuk telur 10
8. Grafik produksi telur selama enam bulan 11
9. Visualisasi hasil PCR gen prolaktin ekson dua 17 10. Lokasi penempelan primer pada gen prolaktin ekson dua 17 11. Insersi basa adenin pada lokasi 2001 pb gen prolaktin ekson dua 18 12. Kromatogram insersi pada gen prolaktin ekson dua 19
DAFTAR LAMPIRAN
1. Database prolaktin itik Pekin (Kode Akses GenBank: AB158611.1) 26 2. Komposisi reagen PCR (Polimerase Chain Reaction) 29 3. Komposisi gel agarosa untuk evaluasi ekstraksi dan PCR 29 4. Komposisi pakan itik periode bertelur dalam penelitian 29 5. Komposisi kit ekstraksi (Genomic Minikit) 29 6. Informasi primer forward dan reverse gen prolaktin ekson dua 30
7. Protokol Ekstraksi DNA (Genomic MiniKit) 31
8. Informasi Sekuens Itik Mojosari Putih Jantan 32 9. Informasi Sekuens Itik Mojosari Putih Betina 33 10. Informasi Sekuens Itik Persilangan PMp Jantan 34 11. Informasi Sekuens Itik Persilangan PMp Betina 35
12. Informasi Sekuens Itik Pekin Jantan 36
13. Informasi Sekuens Itik Pekin Betina 37
14. Hasil uji BLAST sekuens sampel dengan database rujukan 38 15. Hasil sekuens gen prolaktin ekson dan parsial intron 39
1
PENDAHULUAN
Latar Belakang
Ternak itik yang dibudidayakan dan tersebar diberbagai kepulauan di nusantara adalah itik tipe petelur dengan karakteristik bertubuh kecil, ramping dan perototan rendah. Itik persilangan Pekin Mojosari putih (PMp) berpotensi menjadi galur induk itik pedaging dengan produksi telur yang tinggi. Keunggulan itik persilangan PMp yakni memiliki produksi telur yang tinggi dan mencapai 72% (Suparyantob 2005). Induk itik pedaging dengan produksi telur yang tinggi sangat diperlukan dalam mendukung peternakan unggas. Hal dikarenakan telur fertil yang dihasilkan akan ditetaskan untuk memproduksi anak itik (day old duck) sebagai komponen utama budidaya.
Upaya menyeleksi itik persilangan PMp telah dilakukan Balai Penelitian Ternak Ciawi berbasis data fenotipik yang salah satunya melalui jumlah produksi telur. Variasi individu masih terdapat pada itik persilangan PMp yang terlihat dari itik-itik persilangan PMp yang tidak bertelur (Suparyanto 2005). Keragaman ini perlu diperhatikan karena semakin banyak itik yang tidak mampu bertelur maka produksi telur akan semakin rendah. Upaya mengidentifikasi gen-gen yang berkaitan dengan variasi produksi telur telah banyak dilakukan pada ayam. Gen-gen yang berkorelasi dengan produksi telur pada ayam diantaranya prolaktin(Cui
et al. 2006), reseptor prolaktin (Zhang et al. 2012), dan janus kinase (Liu et al. 2010). Gen prolaktin merupakan kandidat gen yang secara spesifik mengontrol variasi jumlah produksi telur melalui reduksi biosintesis telur selama periode mengeram (Chen et al. 2007). Upaya untuk mendukung seleksi yang selama ini telah dilakukan pada itik persilangan PMp dan tetuanya dapat dilakukan dengan memanfaatkan gen prolaktin.
Gen prolaktin pada itik berhasil diidentifikasi oleh Kansaku et al. (2005). Gen tersebut terdiri dari lima daerah koding protein yang dipisahkan oleh empat intron dan mengkode 229 asam amino. Penelitian yang menghubungkan polimorfisme gen prolaktin dengan sifat produksi telur telah banyak dilakukan pada itik di Taiwan dan Cina. Titik-titik mutasi teridentifikasi pada bagian koding protein yaitu ekson dua, empat dan lima diitik lokal Cina (Wang et al. 2011). Titik-titik mutasi lainnya diidentifikasi pada bagian non-koding gen prolaktin di itik Tsaiya dan Gaoyou (Li et al. 2009; Chang et al. 2012). Informasi mengenai keragaman gen prolaktin pada unggas lokal di Indonesia sampai saat ini masih terbatas pada spesies ayam kampung. Informasi gen prolaktin pada ayam lokal bersifat polimorfik dan berkaitan dengan sifat produksi telur melalui mekanisme mengeram (Sartika 2004). Data mengenai keragaman gen prolaktin pada itik lokal sampai saat ini masih belum tersedia. Upaya membantu ketepatan seleksi secara molekuler pada itik PMp melalui pendekatan gen prolaktin dapat mempercepat program seleksi yang sedang dilaksanakan. Hal ini penting untuk dilakukan mengingat penelitian pendahuluan MAS (Marker Assisted Selection) dapat membantu kegiatan seleksi pada sifat-sifat dengan nilai heritabilitas rendah seperti pada produksi telur.
2
Penelitian ini disusun dalam rangka mengidentifikasi ada tidaknya polimorfisme gen prolaktin ekson dua pada itik persilangan PMp generasi kelima, mengamati sifat produksi selama enam bulan dan kualitas telur pertama serta menelusuri sumber keragaman tetuanya yakni itik Pekin dan Mojosari putih. Harapan dari penelitian ini yakni tersedianya bioinformasi dari ternak itik lokal Mojosari, itik persilangan PMp dan Pekin. Sistem seleksi berbasis penanda genetik atau MAS dapat bermanfaat dalam mendukung upaya seleksi yang selama ini telah dilakukan di Balai Penelitian Ternak Ciawi.
Perumusan Masalah penciri genetik (Marker Assisted Selection)
3 Tujuan Penelitian
Penelitian ini bertujuan untuk mengidentifikasi gen prolaktin ekson dua pada itik persilangan Pekin Mojosari generasi kelima, serta mengamati produksi telur periode enam bulan dan kualitas telur pertama.
Manfaat Penelitian
Hasil dari penelitian ini yaitu tersedianya bioinformasi tentang itik lokal di Indonesia. Bioinformasi tersebut bermanfaat dalam mendukung program seleksi berbasis data fenotipik yang telah dilakukan oleh Balai Penelitian Ternak Ciawi.
Ruang Lingkup Penelitian
Ruang lingkup penelitian terbatas pada pengamatan bagian gen prolaktin ekson dua dan parsial intron pada jenis itik itik persilangan PMp dan tetuanya. Peubah yang diamati meliputi aspek genetik dan fenotipik. Aspek genetik terdiri dari frekuensi alel, kejadian mutasi, analisa homologi, dan komposisi nukleotida. Pengamatan fenotipik terdiri dari produksi telur selama enam bulan dan kualitas telur pertama dari itik betina PMp generasi kelima.
2
TINJAUAN PUSTAKA
Domestikasi Itik
Itik domestik yang banyak dibudidayakan diturunkan dari itik liar wild mallard (Anas platyrhynchos). Itik liar memiliki ciri-ciri, antara lain warna bulu penutup tubuh coklat, warna kepala hijau, warna paruh dan kaki kuning, serta bulu sayap berwarna biru (Crawford 1993). Menurut Cherry dan Morris (2008), sejarah domestikasi itik terjadi di Cina sekitar 4.000-10.000 tahun yang lalu pada zaman batu baru (neolithikum). Daerah perairan merupakan tempat yang paling disukai itik sehingga dikenal sebagai unggas air (water fowl). Berdasarkan taksonomi, itik dapat diklasifikasikan berdasarkan berikut
4
(Cahyono 2011). Bangsa itik petelur yang paling banyak dibudidayakan yaitu Indian Runner dan Cherry Valley. Itik Pekin dan Aylesbury merupakan contoh itik pedaging. Contoh itik dwiguna dengan produktifitas telur dan daging yang tinggi yaitu Khaki Campbell dan Cayuga. Adapun salah satu contoh itik hias atau ornamen yaitu mandarin dan Blue Swedish (Cahyono 2011).
Itik Mojosari
Itik Mojosari ditetapkan sebagai rumpun melalui keputusan Mentri Pertanian Nomor 2837/Kpts/LB.430/8/2012 yang menyatakan bahwa itik Mojosari merupakan itik lokal Indonesia yang berasal dari persilangan itik Jawa dengan itik liar di Desa Modopuro, Kecamatan Mojosari, Kabupaten Mojokerto, Jawa Timur. Itik tersebut menyebar ke Pulau Kalimantan, Sulawesi, Jawa Tengah dan Nusa Tenggara Timur. Sifat kualitatif postur tubuh itik dewasa yaitu ramping seperti botol, warna kerabang hijau kebiruan, warna kaki dan paruh hitam, serta warna ekor coklat bagi betina dan hitam bagi jantan (Gambar 2).
Bobot badan dewasa itik Mojosari yaitu 1.6-1.7 kg dengan produksi telur 200-220 butir/tahun. Puncak produksi telur mencapai 90-95%. Bobot telur itik Mojosari sekitar 65-70 g dan mulai berproduksi pada umur 22-24 minggu. Itik betina Mojosari mampu berproduksi telur hingga waktu tiga tahun. Penetapan itik Mojosari sebagai rumpun dikukuhkan pada Keputusan Menteri Pertanian Nomor 2837/Kpts/LB.430/8/2012 sebagai itik lokal. Hal ini disebabkan karena itik Mojosari memiliki keseragaman bentuk fisik, kemampuan adaptasi yang baik pada keterbatasan lingkungan dan memiliki ciri khas yang berbeda dengan rumpun itik asli atau itik lokal lainnnya.
Persyaratan bibit induk Mojosari berdasarkan Standar Nasional Indonesia (SNI) Nomor 7558 Tahun 2009 yakni warna bulu badan secara umum berwarna coklat, paruh dan kaki berwarna hitam keabuan, bobot meri jantan dan betina umur sehari yaitu minimal 37 g dan meri jantan memiliki tonjolan kecil pada kloaka. Bibit induk meri Mojosari harus berasal dari induk yang mempunyai rataan produksi minimal 60% selama satu periode produksi, memiliki daya tetas minimal 60% dari telur fertil dan bobot telur tetas minimal yakni 58 g.
Bibit niaga itik Mojosari dara memiliki umur 4-5 bulan dan siap bertelur. Standar Nasional Indonesia (SNI) Nomor 7359 Tahun 2008 mensyaratkan bobot badan bibit niaga itik Mojosari dara yaitu 1400-1500 g. Bibit niaga (final stock)
5 itik Mojosari meri umur sehari berdasarkan SNI Nomor 7357 Tahun 2008 yaitu memiliki bobot minimum 37 g, warna bulu yakni coklat, warna paruh dan kaki yaitu hitam keabuan dengan kondisi fisik sehat. Persyaratan kualitatif lainnya yaitu kaki normal dan dapat berdiri tegak, mata bersinar dan tampak segar, aktif, tidak dehidrasi, tidak cacat fisik dan kondisi sekitar pusar dan dubur kering.
Itik Pekin
Itik Pekin merupakan itik pedaging yang berasal dari Cina dan memiliki warna bulu putih krim karena homozigot resesif (Cherry dan Morris 2008). Warna paruh dan kaki itik Pekin yakni oranye terang dengan warna mata biru tua. Bobot badan dewasa itik Pekin betina yaitu 3.6 kg sedangkan untuk jantan yaitu 4.1 Kg. Produksi telur tahunan dapat mencapai 140-220 butir dan memiliki tingkah laku maternal yang buruk (Roberts 2008). Tren bobot badan itik Pekin hingga umur 10 minggu terus mengalami kenaikan hingga mencapai bobot hidup 6 Kg (Cherry dan Morris 2008).
Tren perkembangan rasio konversi pakan dari itik Pekin dapat mencapai 2.5 (Cherry dan Morris 2008). Itik tersebut dapat berproduksi dengan baik pada zona temperatur netral yakni 7-22oC. Suhu diatas 27oC akan meningkatkan laju evaporasi dan pada suhu 40oC akan mengalami stres panas akibat hipertermia (Cherry dan Morris 2008). Telur itik Pekin berwarna putih dan memiliki kisaran bobot 71-93 g (Kokoszynski et al 2007). Komposisi telur itik Pekin disusun oleh 59% albumin, 21% oleh kuning telur dan 10% kerabang telur (Adamski et al
Itik persilangan Pekin Mojosari putih atau PMp merupakan itik persilangan antara Pekin jantan dengan itik Mojosari putih betina. Pemilihan warna Mojosari putih didasarkan atas pertimbangan tujuan pembentukan galur itik pedaging. Warna putih pada itik tetua induk Mojosari putih merupakan akibat dari gen homozigot resesif (cc) (Suparyanto et al. 2005a). Persilangan kedua itik tersebut
6
menghasilkan itik PMp yang berwarna putih seragam. Warna seragam putih dari keseluruhan PMp diduga disebabkan oleh kedua tetuanya yang sama-sama memiliki gen homozigot resesif putih (Suparyanto et al. 2005a). Itik persilangan PMp memiliki karakteristik warna paruh pink dan kaki berwarna orange. Karakteristik itik PMp disajikan pada Gambar 4.
Karakteristik pertumbuhan itik PMp sejak menetas hingga umur 90 hari menunjukan perkembangan yang cepat. Lebar dan dalam dada mencapai puncaknya pada umur 22-25 minggu. Lebar dan dalam dada mencapai 89 mm dan 74 mm pada umur 90 hari. Pertambahan bobot badan itik persilangan PMp mencapai puncaknya pada umur sembilan minggu. Itik persilangan PMp generasi pertama memiliki rasio konversi pakan diatas 3.5 dengan produksi telur tahunan mencapai 72%. Warna kerabang telur itik PMp yaitu biru kehijauan dengan berat 66.84 g. Bentuk telur mengarah ke oval dengan indeks 76%. Meri itik persilangan PMp yang ditetaskan menghasilkan bobot tetas 39.6 g (Suparyantoet al. 2005c).
Prolaktin
Prolaktin merupakan hormon polipeptida rantai tunggal yang disintesis di kelenjar pituari anterior. Hormon prolaktin pada itik tersebut disusun dari 229 asam amino yang disandikan oleh gen prolaktin (Kansaku et al. 2005). Gen prolaktin pada itik terdiri dari lima ekson dan empat intron (Gambar 5).
Gambar 4 Itik persilangan Pekin Mojosari Putih (PMp) di Balitnak Ciawi
7 Keterangan
Ekson 1 : 243 - 270 pb Ekson 4 : 3753-3932 pb Intron 2 : 1956-2358 pb Ekson 2 : 1774-1955 pb Ekson 5 : 5844-6035 pb Intron 3 : 2467-3752 pb Ekson 3 : 2359-2466 pb Intron 1 : 271-1773 pb Intron 4 : 3933-5843 pb
Penelitian polimorfisme gen prolaktin itik intron satu berasosiasi dengan berat telur (Li et al. 2009) sedangkan ekson lima menunjukan hubungan dengan produksi telur tahunan (Wang et al. 2011). Analisis haplotipe menunjukan bahwa masing-masing mutasi berkaitan dengan produksi telur dan sifat reproduksi (Chang et al. 2012). Mutasi gen prolaktin itik banyak terjadi di intron (Tabel 1).
Gen prolaktin pada ayam telah ditemukan di lokasi kromosom dua sedangkan letaknya pada kromosom itik belum teridentifikasi. Menurut Forsyth (2002), prolaktin memerankan fungsi pleitropi biologis yang sangat luas pada makhluk hidup. Fungsi-fungsi tersebut diantaranya yaitu penyeimbang elektrolit dan air pada ikan, pertumbuhan dan perkembangan sel, metabolisme dan regulasi endokrin, tingkah laku, reproduksi, regulasi sistem imun, penghancuran sel, regulasi sitoskeleton dan penyelamatan sel. Jalur sinyal prolaktin dalam memerankan fungsi biologinya banyak melibatkan gen-gen lain seperti reseptor prolaktin (prlr), jak dan stat (Radhakrishnan et al. 2012).
Salah satu gen utama yang menunjang kerja prolaktin yaitu reseptor prolaktin (plrr). Prolaktin reseptor pada itik termasuk kedalam famili sitokin kelas I dan tersusun dari 2.439 pb yang mengkode 832 asam amino (Wang et al.
2009a). Reseptor prolaktin terdiri dari domain ekstraseluler sebagai situs penempelan ligan, domain transmembran dan domain intraseluler (Radhakrishnan
et al. 2012). Salah satu fungsi biologisnya yaitu sebagai situs penempelan polipeptida prolaktin. Gen lain yang banyak terlibat dalam mekanisme kerja prolaktin yaitu jak (janus kinase) dan stat (signal transducer and activators of transcription). Gen jak berfungsi sebagai sebagai transmiter sinyal dari luar sel ke nukleus sedangkan stat berfungsi sebagai faktor transkripsi (Gambar 6).
Tabel 1 Titik titik mutasi pada gen prolaktin itik Intron C213Ta A1842Ga T2231Ca INDEL3724e T3777Cd INDEL3939e C5961Ta
T233Cc T3785Gd C3941Td,e T6052Aa
8
Jalur sinyal prolaktin diinisiasi oleh penempelan prolaktin dengan reseptornya (Binart et al. 2000). Ikatan prolaktin dengan reseptornya akan mengaktivasi jak. Reseptor prolaktin akan menginduksi sinyal melalui kinase yang berinteraksi dengan ekor sitoplasmik. Gen jak mengikat sinyal transduksi dan mengaktivasi faktor transkripsi protein stat (Dasilva et al. 1996). Protein stat akan terdisosiasi dengan reseptornya, dan translokasi menuju nukleus untuk mengikat target gen bagian promotor prolaktin. Penempelan pada bagian regulatori prolaktin akan mengatur mekanisme aktif dan inaktif gen prolaktin (Radhakrishnan et al. 2012).
Kualitas dan Produksi Telur
Kualitas telur diartikan sebagai kondisi eksterior-interior dari telur. Parameter yang sering dijadikan indikator kualitas eksterior telur yakni bentuk telur, kondisi kerabang, warna kerabang, tebal dan berat kerabang, luas permukaan kerabang, kekuatan deformasi kerabang, densitas kerabang dan jumlah pori-pori kerabang. Indeks telur merupakan persentase perbandingan lebar dan panjang telur. Luar permukaan telur dihitung rumus Paganelli yaitu Lp=4.835 x Berat Telur (BT)0.662. Warna kerabang dan kuning telur merupakan ukuran kualitatif yang nilainya berupa kisaran (Kokoszynski et al. 2007).
Penampilan kualitas interior telur terdiri dari berat dan tinggi putih telur, berat dan tinggi kuning telur, indeks dan diameter kuning telur, nilai Haugh Unit (HU), warna kuning telur, proporsi dan densitas kuning telur, proporsi dan densitas putih telur, pH kuning dan putih telur. Haugh Unit merupakan korelasi antara bobot telur dengan tinggi putih telur. Nilai HU dihitung dengan formula
Raymond Haugh yaitu, HU = 100 log (H + 7.57-1.7 W0.37) (Monira et al. 2003). Penyimpanan telur dalam waktu yang lama dapat menyebabkan menurunnya nilai HU karena terjadinya pengurangan bobot telur akibat penguapan (Haryono 2000). Warna kuning telur merupakan nilai kualitatif dari kisaran warna kuning telur antara sangat pucat hingga sangat kuning mendekati warna oranye. Bobot sebutir telur merupakan penjumlahan dari komponen-komponen didalamnya. Sebutir
9 telur itik tersusun rata-rata dari 9% kerabang, 59% putih telur dan 32% kuning telur (Adamski et al. 2005).
Sifat produksi telur pada itik memiliki nilai heritabilitas yang rendah dengan kisaran 0.15-0.22 (Cheng et al. 1995; Hu et al. 2004). Berat telur itik memiliki nilai heritabilitas yang lebih tinggi yaitu 0.35 dan nilai repitabilitas 0.68 (Suparyanto et al. 2004c). Produksi telur pada itik secara genetik berkorelasi negatif dengan berat telur dan bobot badan (Cheng et al. 1995). Karakteristik kualitas telur itik seperti tebal kerabang, indeks telur, berat kuning telur, dan warna kuning telur memiliki nilai heritabilitas sedang berturut-turut yaitu 0.26, 0.317, 0.29, dan 0.28 (Tai et al. 1985; Hu dan Tai 1993).
3 METODE
Lokasi dan Waktu
Penelitian ini telah dilaksanakan di Kompleks Itik, Balai Penelitian Ternak Ciawi dan Laboratorium Genetika Molekuler Ternak Fakultas Peternakan IPB. Pengamatan produksi telur dilakukan sejak bulan April-Desember 2013. Analisis molekuler dilakukan pada bulan Oktober 2013 sampai Januari 2014.
Materi
Sampel Darah
Sampel darah yang digunakan sebanyak 1-1.5 ml/ekor yang dikoleksi dari 168 itik yang terdiri dari tiga populasi itik yang terdiri dari: 19 ekor itik Pekin, 36 ekor itik Mojosari dan 113 ekor itik PMp. Sebanyak 98 itik PMp betina digunakan untuk pengamatan fenotipik. Ternak dipelihara pada kondisi lingkungan kandang dan pakan yang sama. Pakan yang diberikan yaitu ransum komersial dengan kandungan protein kasar 18% dengan jumlah pemberian sekitar 250-300 g/ekor/hari. Kandang individu yang digunakan berukuran p x l x t yaitu 45 cm x 45 cm x 45 cm.
Prosedur
Koleksi Darah dan Ekstraksi Total Genom
Pengambilan sampel darah dilakukan pada bagian pembuluh darah sayap. Sebanyak 1.0-1.5 ml sampel diambil menggunakan syringe. Darah dimasukkan ke dalam tabung berisi antikoagulan edta (ethylenediaminetetraacetic). Sampel kemudian disimpan dalam refrigerator dengan suhu 4oC hingga siap untuk dilakukan ekstraksi. Ekstraksi darah menggunakan metode kit (Mini Genomic Kit, Qiagen) sesuai dengan protokol yang tercantum.
Uji Kualitas dan Kuantitas Total Genom
10
melalui elektroforesis dengan menggunakan metode gel agarosa yang dibuat pada konsentrasi 0.8%.
Amplifikasi Gen Prolaktin Ekson Dua dan Parsial Intron
Perbanyakan ruas gen prolaktin ekson dua dan parsial intron dilakukan dengan metode PCR (Polymerase Chain Reaction). Campuran reagen dimasukkan ke dalam mesin thermocycler dengan kondisi suhu pradenaturasi 95°C (5 menit), denaturasi 95°C (10 detik), suhu penempelan (annealing) 60°C (20 detik), suhu ekstensi awal 72°C (30 detik), dan ekstensi akhir 72°C (5 menit). Siklus yang digunakan sebanyak 35 kali. Visualisasi hasil amplifikasi dilakukan melalui elektroforesis dengan menggunakan metode gel agarosa 1.5%. Pita-pita yang muncul dibandingkan dengan posisi marka untuk penentuan ukuran pasang basa. Primer untuk PCR dibuat dengan bantuan software Primer3 berdasarkan database prolaktin itik di GenBank (Kode Akses GenBank: AB158611.1). Sekuens primer
forward (F) yaitu 5’- CCT GAC TGT TTA CAT CCA CAGC-3’ sedangkan sekuens primer reverse (R.) yaitu 3’- GGG ACT ATC ACT GC CTG TGC -5’. Sekuensing Gen Prolaktin Ekson Dua
Sekuensing gen prolaktin ekson dua dan parsial intron dilakukan pada bagian forward di seluruh itik Pekin, Mojosari dan persilangan PMp dengan jumlah 168 sampel. Sekuens gen prolaktin ekson dua dan parsial intron dilakukan melalui jasa perusahaan sekuensing dengan menggunakan mesin sekuenser (ABI PRISM Genetic Analyzer) di First Base, Selangor, Malaysia.
Peubah yang Diamati
Peubah yang diamati terdiri dari produksi dan kualitas telur. Produksi telur itik dikoleksi selama enam bulan pertama. Peubah kualitas meliputi berat telur utuh, bentuk atau indeks telur, berat putih telur, berat dan warna kuning telur, tebal dan berat kerabang, serta nilai HU. Berat telur utuh, putih telur, kuning telur dan HU diukur dengan alat Egg AnalyzerTM. Tebal kerabang diukur dengan alat
Egg Shell ThicknessTM sedangkan beratnya ditimbang dengan timbangan digital. Warna kuning telur dinilai dengan alat Roche Yolk Colour Fan. Indeks telur dihitung dengan rumus : Indeks = [lebar telur : panjang telur] x 100%.
11 Analisis Data
Analisis Sekuens Gen Prolaktin Ekson Dua
Analisis gen prolaktin ekson dua dan partial intron menggunakan program
Moleculer Evolutionary Genetics Analysis (Tamura et al. 2007). Hasil sekuens dianalisis dengan program Bioedit Sequence Alignment Editor (Hall 1999). Kesamaan dengan gen prolaktin itik pada database GenBank dianalisis dengan metode Basic Local Alignment Search Tool (BLAST) yang tersedia online di
https://blast.ncbi.nlm.nih.gov.
4 HASIL DAN PEMBAHASAN
Produksi dan Kualitas Telur
Produksi Telur Periode Enam Bulan
Berdasarkan data pengamatan produksi telur selama enam bulan, diperoleh rataan persentasi produksi telur itik persilangan PMp sebesar 67%. Hasil ini lebih tinggi dibandingkan produksi telur tahunan itik persilangan lainnya yaitu itik persilangan Pekin Alabio yang menghasilkan produksi telur tahunan yang mencapai 63% dan itik persilangan Alabio Pekin memiliki produksi telur tahunan sebesar 62% (Susanti et al. 2012). Itik persilangan Mojosari Alabio pada waktu periode pengamatan yang sama menghasilkan produksi telur yang lebih rendah yaitu 66% (Ketaren et al. 1999). Itik persilangan PMp generasi pertama memiliki produksi telur tahunan yang sangat tinggi yaitu 72% (Suparyanto 2005b). Produksi telur bulanan itik persilangan PMp generasi kelima disajikan pada Gambar 8.
12
Produksi telur mengalami peningkatan pada bulan kedua saat umur itik mencapai enam bulan. Penurunan terjadi pada bulan ketiga yaitu pada umur itik mencapai tujuh bulan sebagai tanda itik-itik akan memasuki fase ranggas bulu. Pada umumnya itik mengalami rontok bulu pada bagian dada dan pantat pada umur tujuh bulan (Supriyadi 2009). Menurut Susanti (2012), mekanisme ranggas bulu pada itik selalu diawali dengan berhentinya produksi telur yang diikuti kejadian bulu meranggas dan kemudian itik berproduksi telur kembali. Terdapatnya itik PMp yang mengalami ranggas bulu menyebabkan itik berhenti berproduksi. Kejadian ranggas bulu pada itik persilangan Pekin Alabio (PA) dan itik persilangan Alabio Pekin (AP) sangat nyata mempengaruhi produksi telur. Itik persilangan PA dan AP yang tidak mengalami rontok bulu dapat mencapai produksi telur tahunan yang lebih tinggi yaitu 73% dan 86% dibandingkan dengan itik yang sama namun mengalami kejadian rontok bulu yang hanya mencapai 63% dan 62% (Susanti et al. 2012).
Fenomena ranggas bulu pada unggas merupakan peristiwa alamiah dalam rangka penggantian bulu-bulu lama oleh folikel baru yang mendorong bulu lama lepas. Banyak faktor yang mempengaruhi peristiwa ranggas bulu pada itik. Menurut Susanti (2012) ranggas bulu dapat disebabkan oleh faktor genetik, lingkungan, dan stres. Kadar hormon prolaktin yang tinggi dalam darah dapat menjadi salah satu pemicu yang menyebabkan rontok bulu (Hafez 2000) dan regresi ovarium (Ramesh et al. 2004). Faktor lain yang mempengaruhi produksi telur yakni ketersediaan pakan. Produksi telur itik persilangan Mojosari Alabio (MA) pada umur 22-42 minggu mampu mencapai 83% pada kondisi pakan ad libitum sedangkan pembatasan pakan 15% dan 30% menghasilkan produksi telur yang lebih rendah yaitu 64% dan 46% (Ketaren et al. 2002). Pakan yang diberikan untuk itik PMp periode layer merupakan ransum petelur komersial dengan kandungan protein kasar yaitu 18%. Standar dari SNI pakan itik petelur (01-3910-2006) merekomendasikan ransum dengan kandungan protein kasar minimum 15% agar produksi telur berjalan baik.
Variasi genetik antar individu itik persilangan PMp memiliki koefisien keragaman untuk produksi telur tergolong tinggi yaitu 34%. Keragaman menunjukan bahwa produksi telur antar individu itik persilangan PMp masih relatif heterogen. Produksi telur yang tinggi dengan keragaman yang rendah merupakan faktor penting dalam menyeleksi calon induk itik betina. Hal ini dikarenakan betina tersebut akan dijadikan induk untuk menghasilkan telur yang akan ditetaskan sebagai sumber bibit niaga. Penetapan kriteria bibit induk (parent stock) mensyaratkan bahwa calon induk tersebut harus berproduksi telur tinggi, cepat dewasa kelamin, viabilitas tinggi, fertilitas tinggi, daya tetas tinggi dan umur induk minimal enam bulan (SNI No7558 Tahun 2009). Menurut Kurnianto (2010), keragaman dikatakan sudah kecil apabila nilainya kurang dari 5% dan dikategorikan besar apabila nilainya lebih dari 15%.
13 dan perkawinan itik PMp jantan dan betina generasi sebelumnya. Perkawinan
interse biasanya dilakukan dalam upaya pemantapan galur. Galur tersebut dikatakan telah stabil apabila telah memiliki nilai keragaman yang kurang dari 5% (Susanti 2012). Seleksi individu dapat saja dilakukan terhadap sifat produksi telur di itik PMp. Namun karena sifat yang akan diseleksi berkaitan dengan jenis kelamin (sex limited) maka kurang akurat untuk diterapkan.
Rataan produksi telur itik persilangan PMp betina selama periode enam bulan yakni 112 butir. Produksi telur tertinggi dicapai hingga 163 butir (97%) dan produksi terendah yaitu 17 butir (10%). Pengamatan produksi telur pada itik hasil persilangan Mojosari lainnya pada periode pengamatan yang lebih singkat (tiga bulan) menghasilkan jumlah telur yang lebih sedikit. Itik persilangan Mojosari Alabio (MA), persilangan Mojosari dengan Mojosari (MM), persilangan Alabio dengan Alabio (AA) dan persilangan Alabio Mojosari (AM) hanya menghasilkan produksi telur berturut-turut yaitu 74, 66, 66 dan 61 butir (Prasetyo dan Susanti 2000). Produksi telur pada itik yang merupakan sifat kuantitatif dilaporkan memiliki nilai heritabilitas yang rendah yaitu 0.15-0.22 (Cheng et al. 1995; Hu et al. 2004). Rendahnya nilai heritabilitas dari sifat produksi telur itik tersebut menunjukan kecilnya proporsi kontribusi genetik terhadap fenotipe yang diakibatkan tingginya pengaruh lingkungan.
Beberapa penelitian pengaruh genetik terhadap produksi telur pada itik dilaporkan spesifik dengan pendekatan gen kandidat tertentu. Wang et al. (2011) melaporkan bahwa produksi telur pada itik salah satunya dipengaruhi oleh mutasi pada gen prolaktin. Genotipe mutant akibat mutasi gen prolaktin di wilayah koding protein pada itik lokal Cina menghasilkan produksi telur tahunan yang lebih tinggi yaitu 317 butir. Namun mutasi gen prolaktin itik di wilayah non-koding dilaporkan tidak mempengaruhi produksi telur pada itik Gaoyou. Itik Gaoyou bergenotipe mutant pada bagian intron satu menghasilkan rataan produksi telur 82 butir selama 100 hari periode pengamatan (Li et al. 2009). Laporan Chang et al. (2012) menyatakan bahwa rataan produksi telur itik Tsaiya selama 40 minggu periode yang mengalami mutasi pada wilayah non koding (intron satu dan empat) yaitu 149 butir.
Menurut Cheng et al. (1995) seleksi terhadap produksi telur pada itik memiliki korelasi genetik negatif dengan bobot badan. Seleksi yang tinggi terhadap bobot badan pada itik Pekin menghasilkan itik yang memiliki pertumbuhan sangat cepat dengan produksi telur rendah. Seleksi bobot badan pada itik Mojosari betina terkendala karena sifatnya sebagai itik petelur yang memiliki pertumbuhan yang lambat dibandingkan itik Pekin. Tingginya produktivitas bertelur itik persilangan PMp betina diwariskan dari tetua Mojosari yang merupakan itik petelur. Tetua induk Mojosari putih dapat menghasilkan produksi telur tahunan yang mampu mencapai 238 butir (Prasetyo dan Susanti 2000). Tingginya produksi telur itik PMp berpotensi untuk dijadikan sebagai bibit. Berdasarkan Standar Nasional Indonesia No.7558 Tahun 2009, kriteria calon bibit induk yang baik memiliki rata-rata produksi telur minimal 60% sehingga itik PMp memenuhi kriteria.
Kualitas Telur Itik Persilangan PMp
14
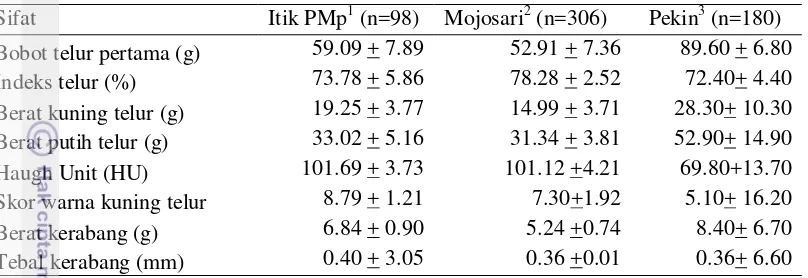
Mojosari putih dan bobot telur itik Pekin. Karakteristik kualitas telur pertama itik persilangan PMp generasi kelima disajikan dalam Tabel 2.
Bobot telur itik persilangan PMp generasi kelima berdasarkan Tabel 2 diatas didapatkan rataan yaitu 59.09 g. Sebanyak 72.77% bobot telur yang dihasilkan berada pada kisaran 51.2-66.98 g. Koefisien keragaman bobot telur itik persilangan PMp tergolong sedang yaitu 13.35%. Hal tersebut menunjukan bobot telur relatif beragam dari ukuran telur itik PMp terkecil (51 g) hingga ukuran telur itik PMp terbesar (67 g). Pengelompokan bobot telur penting untuk dilakukan agar kualitas anak itik atau DOD (day old duck) yang ditetaskan seragam. Keseragaman DOD sangat memudahkan dalam manajemen budidaya seperti pada pemberian pakan. Upaya penyeragaman bobot telur saat penetasan dapat dilakukan dengan melakukan kegiatan pengelompokan.
Suparyanto (2005b) melaporkan bahwa seleksi terhadap bobot telur itik persilangan PMp generasi pertama memiliki korelasi positif terhadap bobot anak itik yang ditetaskan. Oleh karena itu untuk menghasilkan bobot tetas yang tinggi, Setioko (1998) menganjurkan telur itik yang akan ditetaskan sebaiknya berbobot pada kisaran 65-75 g. Pedoman Pembibitan Itik Nomor 237/Kpts/1PD.430/6/2005 menganjurkan bobot telur untuk ditetaskan sebaiknya tidak kurang dari 60 g. Sekitar 43.66% telur-telur itik PMp memiliki bobot di atas 60 g. Telur itik PMp dengan bobot awal 65.36 g akan menghasilkan bobot DOD itik PMp dengan berat 38.5 g atau mengalami 61.5% derajat kehilangan bobot telur (Suparyanto et al. 2004c). SNI bibit itik Mojosari Nomor 7357 Tahun 2008 dan Alabio Nomor 7557 Tahun 2009 menstandarkan bobot DOD final stock yaitu 37 g.
Bobot telur pada itik secara genetik berkorelasi negatif dengan produksi telur yang dihasilkan (Cheng et al. 1995). Hal ini berarti semakin banyak telur yang diproduksi maka akan mengarah pada bentuk telur yang semakin mengecil. Seleksi terhadap bobot badan itik Pekin berdampak pada produksi telur yang rendah dengan ukuran telur yang besar. Telur itik Mojosari yang berproduksi tinggi memiliki ukuran telur yang lebih kecil dibandingkan telur itik Pekin yang diperuntukan sebagai itik pedaging. Berdasarkan penelitian Kokoszynski et al. (2007), telur itik Pekin memiliki kisaran bobot yaitu 71.7-86.7 g dengan dimensi panjang dan lebar telur berkisar 61.9-65.1 mm dan 45-48 mm.
15 telur itik Gaoyou genotipe mutant memiliki berat telur yang lebih tinggi 77 g dibandingkan itik dengan genotipe non mutant dengan berat telur 74 g. Penelitian lainnya bahwa bobot telur itik dipengaruhi oleh mutasi gen prolaktin disampaikan oleh Chang et al. (2012). Bobot telur itik Tsaiya yang mengalami mutasi lebih tinggi yaitu 67 g dibandingkan itik Tsaiya yang tidak mengalami mutasi yaitu 64 g. Bobot telur pada itik memiliki kisaran potensi genetik untuk diwariskan (heritabilitas) sedang yaitu 0.30 (Cheng et al. 1995).
Bobot telur itik persilangan PMp generasi kelima yang hampir mencapai rata-rata 60 g meningkat dibandingkan bobot telur tetua induknya Mojosari putih. Kontribusi peningkatan bobot telur itik persilangan diwariskan dari tetuanya yaitu itik Pekin. Namun bobot telur itik persilangan PMp generasi kelima mengalami penurunan dibandingkan dengan bobot telur itik PMp generasi pertama yang mencapai berat 66.84 g (Suparyanto 2005b). Purba et al. (2000) melaporkan bahwa telur itik Mojosari pada populasi seleksi generasi awal yaitu 51 g. Peningkatan bobot telur itik persilangan PMp dapat disebabkan pengaruh heterosis dari efek persilangan dua tipe itik yang berbeda.
Dampak heterosis yakni meningkatkan kombinasi gen-gen heterozigot dari dua tipe itik berbeda sangat nampak pada itik PMp generasi awal. Nilai heterosis paling baik diperoleh pada generasi pertama (Noor 2010). Upaya seleksi yang dilakukan pada itik-itik bergenotipe PMp pada generasi pertama hingga kelima untuk perkawinan interse dalam rangka pemantapan galur meningkatkan frekuensi gen-gen target seleksi ke arah homozigot dan seragam. Kriteria-kriteria seleksi digunakan oleh Balai Penelitian Ternak Ciawi untuk mendapatkan kondisi optimum dan ideal dari itik persilangan PMp diantaranya yaitu keseragaman genotipe PMp yang mengarah pada sifat pertengahan itik Pekin dan Mojosari putih. Itik PMp memiliki karakteristik bobot badan lebih rendah dari Pekin namun lebih tinggi dari Mojosari putih, perilaku yang tenang, cepat dewasa kelamin, dan berproduksi telur tinggi.
Ciri lain dari itik persilangan PMp generasi kelima yaitu memiliki rataan nilai indeks telur 73.78%. Nilai tersebut mengarah pada bentuk telur yang oval normal (mendekati 75%). Sebanyak 86.85% bentuk telur berada pada kisaran 67-79%. Tinggi rendahnya indeks telur menunjukan tingkat kelonjongan telur. Semakin tinggi angka indeks telur akan mengarah pada bentuk telur yang bulat dan semakin rendah angka indeks telur akan mengarah pada bentuk telur yang lonjong. Nilai koefisien keragaman untuk indeks telur rendah yaitu 5.86%. Rendahnya nilai keragaman nilai indeks telur menunjukan bahwa telur-telur itik persilangan PMp memiliki bentuk yang hampir seragam atau homogen baik telur yang berkuran kecil, sedang, maupun besar. Penelitian Mahi (2012) dan Hermawan (2000) yang mengamati dampak bentuk telur pada puyuh dan ayam terhadap bobot tetas dan jenis kelamin menunjukan hasil yang tidak signifikan.
16
itik merupakan potensi genetik dengan nilai heritabilitas moderat yang berkisar 0.2-0.4 (Tai et al. 1985; Hu dan Tai 1993). Sifat kualitas telur itik (berat putih telur, berat kuning telur, nilau HU, berat dan tebal kerabang) memiliki korelasi yang positif dengan berat telur (Suparyanto et al. 2004c; Cheng et al. 1995). Hal ini berarti peningkatan bobot komponen penyusun telur secara positif berdampak meningkatkan bobot telur utuh.
Selain faktor genetik, kualitas telur juga dipengaruhi oleh lingkungan yang berupa nutrisi dari pakan yang diberikan. Asupan kalsium yang lebih tinggi dibutuhkan untuk itik tipe petelur dibandingkan itik pedaging (NRC 1994). Hal ini dikarenakan kalsium sangat dibutuhkan dalam proses pembentukan kalsium karbonat pada kerabang itik (Shen dan Chen 2003). Pakan petelur yang diberikan untuk itik PMp yang digunakan dalam riset mengandung asupan kalsium sekitar 4%. Pakan itik petelur yang baik sesuai standar dari SNI 01-3910-2006 mengandung 3-4% kalsium. Faktor lain yang mempengaruhi kualitas telur adalah faktor penyimpanan. Monira et al. (2003) melaporkan bahwa waktu penyimpanan telur sejak hari pertama ditelurkan hingga hari ke-21 sangat signifikan menurunkan bobot telur, tinggi albumin dan nilai HU telur ayam.
Pengamatan terhadap komponen penyusun telur lainnya dilakukan untuk melihat kualitas telur itik persilangan PMp. Nilai Haugh Unit (HU) adalah peubah yang menunjukan kualitas putih telur yang dicirikan dengan tinggi albumin dengan berat telur secara keseluruhan (Monira et al. 2003). Pengamatan terhadap tebal dan berat kerabang penting dilakukan dalam proses inkubasi selama proses penetasan. Tebal kerabang yang tipis akan menyebabkan telur mudah retak sedangkan kerabang yang terlalu tebal akan menyulitkan proses pelubangan cangkang (pipping). Rataan nilai HU telur itik PMp sangat tinggi yaitu 101 dengan grade telur AA (sangat baik). Nilai HU yang tinggi menunjukan tingkat kekentalan albumin dari telur itik persilangan PMp yang masih sangat segar pada saat dilakukan pengukuran.
Gen Prolaktin Ekson Dua
Amplifikasi Fragmen Gen Prolaktin Ekson Dua
17
Panjang amplikon prolaktin ekson dua dan parsial intron berdasarkan posisi marker yaitu berukuran sekitar 400 pb. Ukuran tersebut sesuai dengan panjang region yang diapit oleh primer yaitu 1694-2093 pb yang mencakup basa-basa pada primer forward dan reverse, ekson dua, intron satu dan dua (Gambar 10).
Pada genom itik, sekuens gen prolaktin ekson dua membentang pada region 1774-1955 pb dengan panjang yaitu 181 pb (Kode Akses GenBank AB158611.1). Jenis itik Pekin digunakan oleh Kansaku et al. (2005) untuk proses kloning gen prolaktin utuh pada itik pedaging tersebut. Pada proses amplifikasi gen prolaktin ekson dua dan parsial intron, primer mengapit gen prolaktin dengan rentang 1694-2093 pb sehingga sekuens fragmen prolaktin dalam penelitian ini merupakan gabungan dari sebagian intron satu, ekson dua dan intron dua. Proporsi basa ekson dua pada keseluruhan gen prolaktin itik yang berukuran 6.332 yaitu 2.85%. Adapun proporsi bagian ekson dua dengan keseluruhan wilayah ekson gen prolaktin itik yang berukuran 685 pb yaitu 26.42%.
Identifikasi Mutasi Gen Prolaktin Ekson Dua
Hasil sekuens gen prolaktin ekson dua dan parsial intron pada semua jenis itik jantan dan betina baik itik Pekin, Mojosari putih, dan persilangan PMp
Gambar 10 Lokasi primer dan titik mutasi target (primer forward, primer reverse (bergaris bawah), ekson dua (huruf kapital), intron satu dan dua (huruf kecil), Titik
mutasi target yaitu basa adenin pada lokasi ekson dua gen prolaktin itik ditunjukan dengan garis bawah) (Kode Akses GenBank AB158611.1).
18
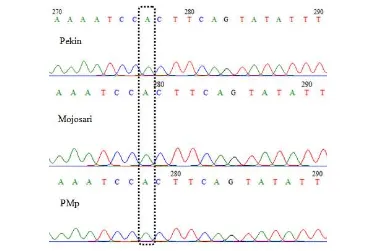
diperoleh hanya satu macam genotipe dan alel. Pada lokasi 2001 pb di intron dua, ditemukan satu tipe insersi atau penyisipan satu basa adenin pada keseluruhan sampel itik yang digunakan dalam penelitian (Gambar 11). Terdapat jenis itik lain dalam GenBank yang memiliki kejadian insersi yaitu itik Linwu (Kode Akses GenBank: JQ677091.1)
Pada penelitian ini, tipe insersi yang terjadi pada tetua itik Pekin di Balitnak Ciawi tidak terdapat pada itik Pekin yang digunakan sebagai referensi pada kode akses GenBank AB158611.1. Mutasi target A-1842-G pada lokasi ekson dua tidak ditemukan baik pada itik Pekin, Mojosari putih dan itik persilangan PMp dalam penelitian. Mutasi yang ditemukan pada penelitian tidak mengubah susunan asam amino karena terletak di wilayah non-koding. Titik mutasi juga tidak terletak dalam situs-situs splicing baik pada donor, akseptor maupun situs percabangan (intron brach site). Walaupun tidak secara langsung berdampak kepada susunan amino, mutasi intron sangat berdampak signifikan pada proses regulasi gen (Chorev dan Liran 2012). Beberapa titik mutasi yang terjadi pada wilayah ekson dua dan parsial intron pengapit intron satu dan dua pada gen prolaktin itik disajikan pada Tabel 3.
Homologi Gen Prolaktin Ekson Dua
Hasil uji kesamaan sekuens fragmen gen prolaktin ekson dua dan parsial intron dengan gen prolaktin itik Pekin acuan pada GenBank (Kode Akses GenBank: AB158611.1) menunjukan kesamaan yang tinggi yakni 99%. Perbedaan terletak pada adanya insersi basa pada lokasi 2001 pb di intron dua (Gambar 12). Itik lain yaitu itik Linwu dan Jingxi memiliki kesamaan 100% dengan itik-itik yang digunakan dalam penelitian yaitu memiliki tipe insersi. Itik
Tabel 3 Identifikasi mutasi basa tunggal pada gen prolaktin itik lokal Cina, Taiwan dan dan hasil penelitian
19 tersebut merupakan tipe itik dwiguna penghasil daging dan telur dari Tiongkok. Itik Linwu (Kode Akses GenBank: JQ677091.1). Itik tersebut sama-sama memiliki basa adenin di posisi 2001 gen prolaktin. Menurut Wheeler dan Baghwat (2007), metode BLAST mencari kesamaan nukleotida atau protein suatu sekuens dan membandingkannya dengan database di GenBank untuk dilihat signifikansi kesamaannya secara statistik
Komposisi Nukleotida Gen Prolaktin Ekson Dua
Analisa komposisi nukleotida dilakukan untuk membandingkan komposisi sekuens sampel dengan sekuens dari database yang dirujuk. Komposisi nukleotida juga berguna dalam melihat konformasi dan struktur DNA (Ohyama 2005). Analisa komposisi nukleotida fragmen gen prolaktin ekson dua dan parsial intron disajikan pada Tabel 4.
Komposisi nukleotida menunjukan bahwa persentase basa adenin pada itik-itik yang digunakan dalam penelitian lebih tinggi dibandingkan database itik-itik Pekin dalam GenBank (Kode Akses: AB158611.1). Hal ini menjadi klarifikasi adanya penambahan satu basa adenin pada insersi di intron dua. Hal tersebut berdampak juga pada tingginya persentase pasangan basa A+T. Verkuil et al.
Tabel 4 Komposisi nukleotida sekuens hasil PCR (sekitar 400 pb)
Sampel A T G C A+T C+G
Pekin (Kode Akses GenBank: AB158611.1)
28.97 29.91 18.38 22.74 58.88 41.12
Pekin (Penelitian) 29.19 29.81 18.32 22.67 59.01 40.99 Mojosari (Penelitian) 29.19 29.81 18.32 22.67 59.01 40.99 PMp (Penelitian) 29.19 29.81 18.32 22.67 59.01 40.99
20
(2008) menyatakan bahwa terdapat korelasi positif antara persentase jumlah G+C dengan temperatur leleh (melting temperature). Semakin tinggi persentase G+C akan diikuti dengan semakin tingginya temperatur leleh. Hal ini dimungkinkan karena ikatan tiga hidrogen antara G+C lebih kuat dibandingkan ikatan dua hidrogen antara adenin dan timin (Muladno 2010).
Frekuensi dan Keragaman Alel Gen Prolaktin Ekson Dua
Hasil analisis frekuensi alel fragmen gen prolaktin ekson dua dan parsial pada itik Pekin, Mojosari dan persilangan PMp hanya ditemukan satu macam genotipe dengan frekuensi alel 100%. Suatu alel dikatakan polimorfik apabila memiliki frekuensi sama dengan atau kurang dari 0.99 (Hartl 2000). Pada penelitian ini, alel yang diperoleh bersifat monomorfik. Sifat monomorfik pada penelitian ini tidak dapat dijadikan marka molekuler untuk sifat-sifat kuantitatif telur selama enam bukan dan kualitas telur pada itik persilangan PMp.
Tipe alel ini tidak ditemukan pada itik lainnya bahkan pada jenis itik yang sama yakni Pekin dalam database. Program seleksi itik PMp telah dilakukan oleh Balai Penelitian Ternak sejak generasi kesatu hingga generasi kelima. Terdapatnya tipe insersi pada tetua Pekin jantan dan Mojosari betina diwariskan kepada itik hasil persilangan PMp. Proses perkawinan antar sesama PMp dalam generasi yang sama (perkawinan interse) untuk menghasilkan PMp generasi selanjutnya telah menyebabkan alel homozigot tersebut terus menerus diwariskan dari satu generasi ke generasi berikutnya.
5 SIMPULAN DAN SARAN
Simpulan
Gen prolaktin ekson dua pada itik persilangan PMp generasi kelima bersifat monomorfik dan homozigot sehingga tidak dapat digunakan sebagai marka untuk produksi dan kualitas telur.
Saran
Pengamatan gen prolaktin perlu dilakukan lebih spesifik pada bagian regulator (promotor) dan reseptor prolaktin (plr-r). Penambahan jumlah sampel dan spesies itik lain dari luar Balai Penelitian Ternak dapat dilakukan untuk melihat keragaman gen prolaktin pada wilayah yang lebih luas pada itik lokal lainnya. Penelitian ekspresi gen prolaktin pada fase sebelum, sedang dan setelah masa bertelur dapat dilakukan untuk melihat lebih jauh mekanisme gen prolaktin tersebut pada sifat produksi telur pada itik-itik lokal di Indonesia.
21
DAFTAR PUSTAKA
Adamski M, Bernacki Z, Kuzniacka J. 2005. Changes in biological value of duck eggs defined by egg quality. Folia Biol 53: 107-114.
Binart N, Ormandy CJ, Kelly PA. 2000. Mammary gland development and the prolactin receptor. Adv Exp Med Biol 480: 85-92.
Bu G, Wang CY, Cai G, Leung FC, Xu M, Wang H, Huang G, Li J, Wang Y. 2013. Molecular characterization of prolactin receptor (cPRLR) gene in chickens: Gene structure, tissue expression, promoter analysis, and its interaction with chicken prolactin (cPRL) and prolactin-like protein (cPRL-L. Mol Cell Endocrinol 370: 149–162.
Cahyono B. 2011. Pembibitan Itik. Jakarta (ID): Penebar Swadaya.
Chang MT, Cheng YS, Huang MC. 2012. Association of prolactin haplotypes with reproductive traits in Tsaiya ducks. Ani Repro Sci 135 (1): 91-96. Chen CF, Shiue YL, Yen CJ, Tang PC, Chang HC, Lee YP. 2007. Laying traits
and underlying transcripts, expressed in the hypothalamus and pituitary gland, that were associated with egg production variability in chickens.
Theriogenol 68: 1305-1315.
Cheng YS, Rouvier R, Poivey JP, Tai C. 1995. Genetic parameters of body weight, egg production and shell quality traits in the Brown Tsaiya laying duck. Genet Select Evol 27: 459–472.
Cherry P, Morris TR. 2008. Domestic Duck Production. London (GB): CABI Chorev M, Liran C. 2012. The Function of Introns. Frontiers in Genetics.
Crawford RD. 1993. Origin and history of poultry species. In: Crawford RD. 1993. Poult Breed Genet. Amsterdam (NL): Elseviers.
Cui JX, Du HL, Liang Y, Deng XM, Li N, Zhang XQ. 2006. Association of polymorphisms in the promoter region of chicken prolactin with egg production. Poult Sci 85: 26-31.
Dasilva L, Rui H, Erwin RA, Howard OM, Kirken RA, Malabarba MG, Hacket RH, Larner AC, Farrar WL. 1996. Prolactin recruits STAT1, STAT3, and STAT5 independent of conserved receptor tyrosine TYR402, TYR479, TYR512, TYR580. Mol Cell Endocrinol 117: 131-140.
[DPKH] Direktorat Peternakan dan Kesehatan Hewan. 2012. Surat Keputusan Penetapan Rumpun Itik Mojosari Nomor 2837/Kpts/LB.430/8/2012. Jakarta (ID): Direktorat Jenderal Peternakan Kementrian Pertanian Republik Indonesia.
[DPKH] Direktorat Peternakan dan Kesehatan Hewan. 2005. Pedoman Pembibitan Itik yang Baik Nomor 237/Kpts/1PD.430/6/2005. Jakarta (ID): Direktorat Jenderal Peternakan Kementrian Pertanian Republik Indonesia. Forsyth IA, Wallis M. 2002. Growth hormone and prolactin : moleculer and
functional function. J Mammary Gland Neoplas 7: 291-312.
Hafez ESE. 2000. Reproduction in Farm Animal. 6th Ed. Philadelphia (US): Lea & Febiger.
Hall TA. 1999. BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucl Acids SympSer 41:95-98. Hartl DL. 2000. A Primer of Population Genetics 3rd ed. Massachusetts (GB):
22
Haryono. 2000. Langkah-langkah uji kualitas telur konsumsi ayam ras. Temu Teknis Fungsional Non Peneliti: Balai Penelitian Ternak Ciawi.
Hermawan A. 2000. Pengaruh bobot dan bentuk telur terhadap jenis kelamin pada ayam kampung pada saat menetas. [Skripsi]. Bogor (ID): Institut Pertanian Bogor.
Hu YH, Poivey JP, Rouvier R, Liu SC, Ta, C. 2004. Heritabilities and genetic correlations of laying performances in Muscovy selected in Taiwan. British Poultr Sci 45: 180–185.
Hu YH, Tai C. 1993. Estimation of genetic parameters of yolk color in I-Lan Tsaiya. J Taiwan Livestock Res 26: 131–138.
Indriati M, Sumantri C, Susanti T. 2014. Analysis of diversity prolactin gene exon 4 in Pekin, White Mojosari and Pekin Mojosari Crossbred. Media Peternakan (Jurnal dalam proses review).
Kansaku N, Ohkubo T, Okabayashi H, Guémené D, Kuhnlein U, Zadworny D, Shimada K. 2005. Cloning of duck PRL cDNA and genomic DNA. Gener Compar Endocrinol: 39–47.
Ketaren PP, Prasetyo LH, Murtisari T. 1999. Karakter produksi telur pada itik silang Mojosari x Alabio. Seminar Nasional Peternakan dan Veteriner. Ketaren PP, Prasetyo LH. 2002. Pengaruh pemberian pakan terbatas terhadap
produksi telur itik silang Mojosari x Alabio (MA): 1. Masa bertelur fase Pengembangan dan Penjaminan Mutu Pendidikan Universitas Diponegoro. Liang Y, Cui J, Yang G, Leung FC, Zhang X, 2006. Polymorphisms of 5-flanking
region of chicken prolactin gene. Domest Anim Endocrinol 30: 1-16.
Li HF, Zhu WQ, Chen KW, Zhang TJ, Song WT. 2009. Association of polymorphisms in the intron 1 of duck prolactin with egg performance. Tuk J Vet Anim Sci 33(3): 193-19.
Mahi M, Achmanu, Muharlim. 2012. Pengaruh bentuk telur dan bobot telur terhadap jenis kelamin, bobot tetas dan lama tetas burung puyuh (coturnix-coturnix japonica). Bagian Produksi Ternak Fakultas Peternakan, Universitas Brawijaya.
Monira KN, Shalahuddin, Miah G. 2003. Effect of breed and holding period on egg quality characteristics of chicken. Inter J Poult Sci 2(4): 261–263. Muladno. 2010. Teknologi Rekayasa Genetika. Bogor (ID): IPB Pr.
Noor RR. 2010. Genetika Ternak. Edisi VI. Jakarta (ID): Penebar Swadaya. [NRC] National Research Council. 1994b. Nutrient requirement of ducks. Di Mojosari I. Awal pertumbuhan dan awal bertelur. JITV 2 (3): 152-156. Prasetyo LH, Susanti T. 2000. Persilangan timbal balik antara itik Alabio dan
23 Purba M, Prasetyo LH, Susanti T. 2006. Kualitas telur itik alabio dan
mojosari pada generasi pertama populasi seleksi. Seminar Nasional Teknologi Peternakan dan Veteriner: Balai Penelitian Ternak. Bogor. Radhakrisna et al. 2012. A pathway map of prolactin signaling. J Cell Commun
Sign 6 : 169-173.
Ramesh R, Kuenzel WJ, Proudman JA. 2004. Increased proliferative activity and programmed cellular death in the turkey hen pituitary gland following interruption of incubation behavior. Reg Artie Biol Repro 64: 611-618. Roberts V. 2008. British Poultry Standard. Oxford (GB): Blackwell Pub. Samosir DJ. 1993. Ilmu Ternak Itik. Jakarta (ID): Jakarta.
Sartika T, Duryadi D, Mansjoer SS, Saefuddin A, Martojo H. 2004. Gen promotor prolaktin sebagai penanda pembantu seleksi untuk mengontrol sifat mengeram pada ayam Kampung. JITV 9(4): 239-245.
Setioko AR. 1998. Penetasan telur itik di Indonesia. Wartazoa 7(2): 40-46.
Shen TF, Chen WL. 2003. The role of magnesium and calcium in eggshell formation in tsaiya ducks and leghorn hens. Asian-Aust. J Anim Sci 16(2) : 290-296
[SNI] Standar Nasional Indonesia. 2009. Bibit Niaga Itik Alabio Meri No.7557-2009. Jakarta (ID): Badan Standardisasi Nasional.
[SNI] Standar Nasional Indonesia. 2009. Bibit Induk Itik Mojosari Meri No.7558-2009. Jakarta (ID): Badan Standardisasi Nasional.
[SNI] Standar Nasional Indonesia. 2008. Bibit Niaga Dara Itik Mojosari No.7359-2008. Jakarta (ID): Badan Standardisasi Nasional.
[SNI] Standar Nasional Indonesia. 2008. Bibit Niaga Itik Mojosari Meri No.7357-2008. Jakarta (ID): Badan Standardisasi Nasional.
[SNI] Standar Nasional Indonesia. 2006. Pakan Itik Petelur No.01-3910. Jakarta (ID): Badan Standardisasi Nasional.
Song WT, Li HF, Zhu WQ, Wang Q, Han W, Xu WJ. 2010. Analysis of single nucleotide polymorphism on prolactin gene intron of ducks. Jiangsu J Agr Sci: 2
Suparyanto A, Setioko AR, Prasetyo LH, Susanti T. 2005a. Ekspresi gen homosigot resesif (c/c) pada performans telur pertama itik Mojosari. JITV
10(1): 6-11.
Suparyanto A. 2005b. Peningkatan Produktivitas Daging Itik Mandalung melalui Pembentukan Galur Induk [Disertasi]. Bogor (ID): Institut Pertanian Bogor. Suparyanto A, Setioko AR, Prasetyo LH. 2004c. Evaluasi telur tetas hasil IB antara itik mojosari putih dengan pejantan Pekin. Seminar Nasional Teknologi Peternakan dan Veteriner.
Suparyanto A, Martojo H, Hardjosworo PS, Prasetyo LH. 2004d. Kurva pertumbuhan morfologi itik betina hasil silang antara Pekin dengan Mojosari Putih. JITV 9(2): 87-97.
Supriyadi. 2009. Panduan Lengkap Itik. Jakarta (ID): Penebar Swadaya.
Susanti T, Noor RR, Hardjosworo PS, Prasetyo LH. 2012. Keterkaitan kejadian dan lamanya rontok bulu terhadap produksi telur itik hasil persilangan Pekin dengan Alabio. JITV 17(2): 112-119.
24
Tai C, Tai LJJ, Huang HH. 1985. Estimation of genetic parameters for egg quality characters in laying ducks. J Chin Soc Anim Sci 14: 105–110.
Tamura K, Dudley J, Nei M, Kumar S. 2007. MEGA4: Molecular evolutionary genetics analysis (MEGA) software version 4.0. Mol Biol Evol 24:1596-1599.
Verkuil EP, Belkum A, Hays JP. 2008. Principles and Technical Aspect of PCR Amplification. Doordrecht (NL): Springer.
Wang J, Hou SS, Huang W, Yang XG, Zhu XY, Liu XL. 2009a. Molecular cloning of prolactin receptor of the Pekin duck. Poult Sci 88: 1016-1022. Wang SK, Wu X, Wang H, Li A, Jia HY, Wang GY. 2009b. Correlation analysis
between nucleotide polymorphism of the prolactin exon and broodiness in ducks. J Shanghai Jiaotong Univ - Agr Sci. 27(4): 331-334 .
Wang C, Liang Z, Yu W, Feng Y, Peng X, Gong Y, Li S. 2011. Polymorphism of the prolactin gene and its association with egg production traits in native Chinese ducks. S Afr J Anim Sci 41:63-70.
Wheeler D and Baghwat M. 2007. BLAST QuickStart Example-Driven Web-Based BLAST Tutorial in Comparative and Genomic. Di dalam: Bergman NH. 2007. Compar Genom. New Jersey (US): Totowa. p149
Zhang L, Li DY, Liu YP, Wang Y, Zhao XL, Zhu Q. 2012. Genetic effect of the prolactin receptor gene on egg production traits in chickens. Genet Mol Res
25