AKTIVITAS EKST
TERHADAP
Sta
STAPH
S
INS
STRAK KASAR ALKALOID DAUN PEP
Staphylococcus aureus
DAN EKSPRESI G
APHYLOCOCCAL ENTEROTOXIN
A
LITA HANDAYANI
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
BOGOR
2013
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA
Dengan ini saya menyatakan bahwa tesis berjudul Aktivitas Ekstrak Kasar Alkaloid Daun Pepaya terhadap Staphylococcus aureus dan Ekspresi Gen Staphylococcal EnterotoxinA adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, Desember 2013
RINGKASAN
LITA HANDAYANI. Aktivitas Ekstrak Kasar Alkaloid Daun Pepaya terhadap Staphylococcus aureus dan Ekspresi Gen Staphylococcal Enterotoxin A. Dibimbing oleh HARSI DEWANTARI KUSUMANINGRUM dan DIDAH NUR FARIDAH.
Pepaya merupakan tanaman yang banyak ditanam di Indonesia. Daun pepaya digunakan oleh masyarakat sebagai sayur-mayur, bahan pengempuk daging, dan sebagai bahan obat-obatan tradisional. Ekstrak daun pepaya dilaporkan memiliki aktivitas antimikroba terhadap bakteri Gram positif maupun Gram negatif. Aktivitas antimikroba tersebut berhubungan dengan komponen fitokimia yang dimiliki oleh daun pepaya. Tanin dan terpenoid dapat mengganggu membran sel bakteri, sedangkan alkaloid dapat menyisip pada utas ganda DNA dan mengambat sintesis DNA. Oleh karena itu, selain sebagai pengempuk daging, daun pepaya dapat menjadi alternatif pengawet alami di dalam pangan.
Staphylococcus aureus merupakan patogen penyebab intoksikasi melalui produksi staphylococcal enterotoxins (SEs) pada pangan. Staphylococcal enterotoxinA (SEA) merupakan salah satu SEs yang paling sering menyebabkan kasus keracunan pangan akibat S. aureus. SEA disintesis dari gen sea dan diekspresikan pada pertengahan fase eksponensial pertumbuhan. Selain menghambat pertumbuhan bakteri, terdapat potensi bahwa ekstrak daun pepaya dapat menghambat produksi SEA melalui aktivitas alkaloid. Penelitian ini bertujuan untuk menguji pengaruh penambahan ekstrak kasar alkaloid daun pepaya dalam berbagai konsentrasi terhadap ekspresi gen penyandi SEA.
Ekstrak kasar alkaloid diekstraksi dari daun pepaya kering dengan ultrasonic-assisted extraction. S. aureus diisolasi dari pangan siap saji dan susu sapi mentah menggunakan metode Bacteriological Analytical Manual (BAM) yang dimodifikasi. Isolat dengan koloni tipikal S. aureus pada media Baird-Parker Agar (BPA) dan Mannitol Salt Agar (MSA) serta positif memproduksi koagulase diidentifikasi dengan uji API Staph serta dengan PCR gen penyandi 16S rRNA, diikuti dengan sekuensing. Sekuen yang diperoleh dianalisis menggunakan program BLAST. Konsentrasi Hambat Minimum (KHM) ekstrak kasar alkaloid ditentukan menggunakan metode pengenceran makro. Selanjutnya, isolat S. aureus dipaparkan dengan ekstrak kasar alkaloid pada konsentrasi 0, 1, dan 2 kali KHM. Setelah itu, ekspresi gen sea dianalisis dengan quantitative reverse transcriptionPCR (qRT-PCR).
metode perbandingan CT. Ekstrak kasar alkaloid daun pepaya memiliki aktivitas antibakterial terhadap S. aureus, yang tidak hanya menghambat pertumbuhan bakteri tetapi juga menghambat pembentukan toksin dengan menekan ekspresi gensea.
SUMMARY
LITA HANDAYANI. Activity of Crude Alkaloid Extract of Papaya Leaves toward Staphylococcus aureus and Staphylococcal Enterotoxin A Gene Expression. Supervised by HARSI DEWANTARI KUSUMANINGRUM and DIDAH NUR FARIDAH.
Papaya plant is widely grown in Indonesia. Papaya leaves are used by people as vegetable, meat tenderizer and traditional herbs. Extracts of papaya leaves have been reported to have antimicrobial activity against both Gram positive and Gram negative bacteria. This antimicrobial activity was related to phytochemical content of papaya leaves. Tannins and terpenoids can make cell membrane disruption, whereas alkaloids have been shown to intercalate into DNA and inhibit DNA synthesis. Therefore, besides as meat tenderizer, papaya leaves can be an alternative natural preservatives in food.
Staphylococcus aureus is known pathogen causing intoxication by producing staphylococcal enterotoxins (SEs) in food. Staphylococcal enterotoxin A (SEA) is one of SEs that is commonly implicated in staphylococcal food poisoning. SEA is synthesized from sea gene and expressed from the mid-exponential phase of growth. In addition to the ability to inhibitS. aureusgrowth, there is a potential action of papaya leaves extract, i.e the ability to inhibit SEA production through the alkaloids activity. This study was conducted to investigate the ability of crude alkaloid extract from papaya leaves to inhibit SEA expression.
Crude alkaloid extract was extracted from grounded dried papaya leaves using ultrasonic-assisted extraction.S. aureuswas isolated from ready to eat food and raw cow milk using modified Bacteriological Analytical Manual (BAM) method. Isolates with typical colony of S. aureus on Baird-Parker Agar (BPA) and Mannitol Salt Agar (MSA) and produced coagulase were identified using API Staph test and PCR of 16S rRNA gene, followed by sequencing. Those 16S rRNA gene sequence were analyzed using BLAST. Minimum Inhibitory Concentration (MIC) of crude alkaloid extract were determined by broth macro-dilution method. Furthermore, S. aureus isolate was exposed to crude alkaloid extract at 0, 1, and 2 folds of MIC, and subsequently the expression of sea gene was analyzed using a quantitative reverse transcription PCR (qRT-PCR).
against S. aureus which was not only inhibited the bacteria but also the toxin formation by suppressing the expression ofseagene.
© Hak Cipta Milik IPB, Tahun 2013
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB
AKTIVITAS EKSTRAK KASAR ALKALOID DAUN PEPAYA
TERHADAP
Staphylococcus aureus
DAN EKSPRESI GEN
STAPHYLOCOCCAL ENTEROTOXIN
A
LITA HANDAYANI
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains
pada
Program Studi Ilmu Pangan
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Judul Tesis : Aktivitas Ekstrak Kasar Alkaloid Daun Pepaya terhadap Staphylococcus aureus dan Ekspresi GenStaphylococcal EnterotoxinA
Nama : Lita Handayani NRP : F251110331
Disetujui
Komisi Pembimbing
Dr Ir Harsi Dewantari Kusumaningrum Dr Didah Nur Faridah, STP MSi Ketua Anggota
Diketahui
Ketua Program Studi Dekan Sekolah Pascasarjana IPB Ilmu Pangan
Prof Dr Ir Ratih Dewanti-Hariyadi, MSc Dr Ir Dahrul Syah, MScAgr
PRAKATA
Puji dan syukur kepada Tuhan Yang Maha Esa atas berkat dan bimbingan-Nya sehingga penulis dapat menyelesaikan penelitian tesis ini. Tesis dengan judul
“Aktivitas Ekstrak Kasar Alkaloid Daun Pepaya terhadap Staphyloccous aureus dan Ekspresi Gen Staphylococcal Enterotoxin A” ini dilaksanakan pada bulan
Januari hingga September 2013.
Dalam kesempatan ini, penulis mengucapkan terima kasih dan penghargaan kepada Dr Ir Harsi Dewantari Kusumaningrum selaku ketua komisi pembimbing dan Dr Didah Nur Faridah, STP MSi selaku anggota komisi pembimbing atas bantuannya sehingga penelitian ini dapat didanai oleh Direktorat Jendral Pendidikan Tinggi, serta atas waktu dan kesempatan yang telah diluangkan dalam memberikan bimbingan, ilmu, arahan, motivasi, dan masukkan selama penulis mengikuti pendidikan, penyusunan proposal, pelaksanaan penelitian, pembuatan artikel jurnal hingga penyusunan tesis. Ucapan terima kasih juga disampaikan kepada Direktorat Jendral Pendidikan Tinggi, Kementerian Pendidikan dan Kebudayaan Republik Indonesia yang telah mendanai penelitian ini melalui program Hibah Desentralisasi dengan Skim Penelitian Unggulan Perguruan Tinggi tahun 2013.
Terima kasih kepada keluarga dan sahabat terkasih atas doa dan dukungannya. Ucapan terima kasih juga penulis sampaikan kepada Bapak Hadi Januar atas kesempatan yang diberikan sehingga penulis dapat menempuh pendidikan di program studi ilmu pangan IPB. Di samping itu, terima kasih kepada Dr Ir Utut Widyastuti, MSi atas arahan dan bimbingan dalam penentuan metode pada tahapan analisis molekuler dan masukkannya selaku penguji luar komisi pembimbing pada ujian tesis, kepada seluruh teman, dosen, teknisi, karyawan di IPN, Departemen ITP dan SEAFAST, terutama Mbak Ari, Pak Taufik, Teh Yayam, Mas Yerris, dan Mbak Nurul, dan terima kasih kepada rekan-rekan penelitian di laboratorium, terutama Mbak Fenny dan Arum atas bantuan, masukkan, dukungan, dan kerja sama, serta kepada Pak Muksin dan Pak Hendar di University Farm atas bantuannya dalam pengambilan sampel daun pepaya dan kepada semua pihak selama penyelesaian tesis ini.
Akhir kata penulis berharap semoga penelitian dan tesis ini dapat bermanfaat bagi pembaca serta mampu memberikan sumbangan yang berarti bagi ilmu pengetahuan.
Bogor, Desember 2013
DAFTAR ISI
DAFTAR TABEL xiv
DAFTAR GAMBAR xiv
DAFTAR LAMPIRAN xv
1 PENDAHULUAN
Latar Belakang 1
Perumusan Masalah 2
Tujuan Penelitian 2
Hipotesis 2
Manfaat Penelitian 3
2 TINJAUAN PUSTAKA
Daun Pepaya 4
Staphylococcus aureusdanStaphylococcal Enterotoxin 6
Staphylococcus aureus 6
Staphylococcal Enterotoxin 6
Staphylococcal EnterotoxinA 7
Isolasi dan IdentifikasiS. aureus 7 Isolasi dan IdentifikasiS. aureussecara Konvensional 7 IdentifikasiS. aureussecara Molekuler 8 Quantitative Reverse Transcription Polymerase Chain Reaction 9
Kuantifikasi Absolut 10
Kuantifikasi Relatif 11
3 METODE PENELITIAN
Tempat dan Waktu 12
Bahan Penelitian 12
Peralatan Penelitian 13
Prosedur Penelitian 13
Persiapan Daun Pepaya 13
Ekstraksi Alkaloid Daun Pepaya 14 IsolasiS. aureusdari Pangan 14
Identifikasi gensea 16
Penentuan Konsentrasi Hambat Minimum 16 PemaparanS. aureusdengan Ekstrak Kasar Alkaloid Daun Pepaya 17 Pengukuran Tingkat Ekspresi Gensea 17
Analisis Data 19
4 HASIL DAN PEMBAHASAN
Ekstrak Kasar Alkaloid Daun Pepaya 20 IsolatS. aureusPenghasil SEA 22 Isolat TerdugaS. aureusdari Pangan 22 Hasil Identifikasi Isolat TerdugaS. aureusdengan metode
Konvensional 23
Hasil Identifikasi Isolat TerdugaS. aureusdengan PCR 25
Amplikon Gensea 27
Aktivitas Ekstrak Kasar Alkaloid Daun Pepaya terhadap Jumlah
BakteriS.aureus dan Sintesis Gensea 31 Aktivitas Ekstrak Kasar Alkaloid terhadap JumlahS. aureus 31
Kemurnian Isolat RNA 32
Aktivitas Ekstrak Kasar Alkaloid terhadap Sintesis Gensea 32
5 SIMPULAN DAN SARAN 36
DAFTAR PUSTAKA 37
LAMPIRAN 43
DAFTAR TABEL
1 Aktivitas antimikroba berbagai ekstrak daun pepaya 4 2 Mekanisme antimikroba berbagai senyawa bioaktif 4 3 Faktor yang mempengaruhi pertumbuhanS. aureus 6 4 Faktor yang mempengaruhi pembentukan SE 6 5 Primer yang digunakan pada PCR dan qRT-PCR .... 16 6 Hasil uji API Staph isolatS. aureusdari pangan 24 7 Konsentrasi dan kemurnian isolat DNA... 26 8 Hasil analisis sekuen penyandi gen 16S rRNA isolatS. aureusdengan
Program BLAST... 27 9 Nilai ekspresi relatif genseadengan metode2 33
DAFTAR GAMBAR
1 Struktur alkaloid karpain 5
2 Kurva amplifikasi pada reaksi qPCR 9
3 Molekul fluoresens pada qPCR 10
4 Diagram alir tahapan penelitian 14
5 Daun pepaya kering 20
6 Proses ekstraksi alkaloid : (a) setelah sonikasi dan penyaringan; (b) setelah penambahan larutan H2SO4, (c) lapisan yang terbentuk saat ekstraksi dengan kloroform, (d) ekstrak kasar alkaloid di dalam
kloroform 21
7 PertumbuhanS. aureuspada media (a) BPA-kuning telur telurit
(b) MSA 23
8 Uji aktivitas koagulase isolatS. aureus 23 9 Uji API Staph (a) awal, setelah inkubasi 18-24 jam (b) isolat 66.8%
positifS. aureus, (c) 85% positifS. aureus, dan (d) 97.8% positif
S. aureus 24
10 Contoh hasil elektroforesis isolat DNA genomS. aureus 25 11 Hasil elektroforesis produk PCR gen penyandi 16S rRNA 26 12 Hasil elektroforesis produk PCR gensea 28 13 Zona penghambatan ekstrak kasar alkaloid daun pepaya terhadap
S. aureusSJ1 29
14 Diameter zona bening ekstrak kasar alkaloid daun pepaya terhadap
15 Kemampuan penghambatan ekstrak kasar alkaloid daun pepaya pada berbagai konsentrasi, dengan uji pengenceran makro setelah inkubasi 24 jam, terhadap pertumbuhanS. aureusSJ1 dengan jumlah awal
inokulum 4.65 log CFU/ml 30
16 Pengaruh ekstrak kasar alkaloid daun pepaya pada 0, 1, dan 2 KHM
terhadap pertumbuhanS. aureusSJ1 31 17 Kurva pelelehan genseadan gen penyandi 16S rRNAS. aureusSJ1 34 18 Hasil elektoforesis produk qPCR gensea S. aureusSJ1 yang dipaparkan
dengan ekstrak kasar alkaloid daun pepaya pada konsentrasi
(1) 0 mg/ml, (2) 0.25 mg/ml, dan (3) 0.5 mg/ml. 34
DAFTAR LAMPIRAN
1 Sekuen gen penyandi 16S rRNA isolatS. aureusdari pangan 43 2 Hasil uji ANOVA dan Duncan diameter zona hambat ekstrak kasar
alkaloid daun pepaya pada berbagai konsentrasi terhadapS. aureusSJ1 45 3 Hasil uji ANOVA dan Duncan nilai CTgenseadan gen penyandi
16S rRNAS. aureusSJ1 yang telah dipaparkan dengan ekstrak kasar
1 PENDAHULUAN
Latar Belakang
Pepaya (Carica papaya) merupakan tanaman yang banyak ditanam di daerah tropis dan subtropis, termasuk di Indonesia. Pepaya telah menjadi tanaman pekarangan dan tersebar di berbagai wilayah di Indonesia. Selain buahnya, daun pepaya juga dimanfaatkan oleh masyarakat. Daun pepaya digunakan sebagai sayur-mayur, bahan pembantu dalam pangan, maupun sebagai bahan obat-obatan. Sebagai sayur-mayur, daun pepaya dimasak menjadi suatu hidangan, sedangkan sebagai bahan pembantu pangan, daun pepaya dimanfaatkan sebagai pengempuk daging dengan cara membungkus daging dengan daun pepaya sebelum dimasak maupun dengan merendam daging bersama dengan sari daun pepaya (Abdallaet al. 2012). Sebagai obat-obatan, daun pepaya digunakan untuk membungkus luka dan uapnya dimanfaatkan untuk mengobati asma. Infusi daun pepaya digunakan untuk mengobati sakit perut, demam, dan beri-beri (Krishnaet al.2008).
Ekstrak daun pepaya dilaporkan memiliki aktivitas antimikroba terhadap bakteri Gram positif maupun Gram negatif, seperti Escherichia coli, Staphylococcus aureus, Bacillus cereus, Klebsiella pneumoniae, dan Pseudomonas aeruginosa (Baskaranet al. 2012). Aktivitas antimikroba tersebut berhubungan dengan komponen fitokimia yang dimiliki oleh daun pepaya, seperti flavonoid, tanin, terpenoid, dan alkaloid. Tanin dan terpenoid mampu menghambat pertumbuhan bakteri dengan cara mengganggu membran sel bakteri, sedangkan alkaloid dapat mengambat sintesis DNA dengan cara menyisip pada utas ganda DNA (Cowan 1999; Baskaran et al. 2012). Oleh karena itu, selain sebagai pengempuk daging, daun pepaya dapat menjadi alternatif pengawet alami di dalam pangan.
S. aureus merupakan salah satu bakteri patogen penyebab keracunan pada pangan. Susu dan produk susu, salad, krim pie, krim pengisi pastri, dan produk telur merupakan pangan yang berkaitan dengan kasus keracunan akibat bakteri ini (FDA 2012). Menurut data Center for Disease Control and Prevention (CDC), terdapat sekitar 49 kasus kejadian luar biasa (KLB) yang disebabkan oleh S. aureuspada tahun 2006-2011, dengan jumlah korban sakit mencapai sekitar 1402 orang. Bakteri ini dapat menyebabkan intoksikasi melalui staphylococcal enterotoxin (SE) yang dihasilkannya. SE tahan terhadap panas sehingga tidak akan hilang walaupun pangan mengalami pemanasan.
2
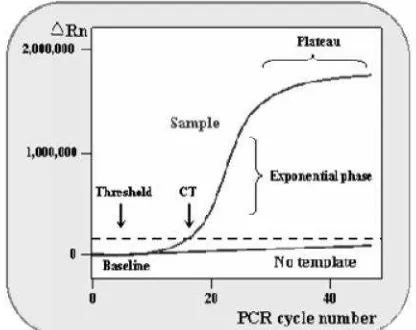
teknik Quantitative Reverse Transcription Polymerase Chain Reaction (qRT-PCR). qRT-PCR meliputi proses transkripsi balik (reverse transcription) yang diikuti dengan real time PCR (qPCR). mRNA ditranskripsi balik membentuk cDNA, yang selanjutnya digunakan sebagai cetakan pada qPCR (Pfaffl et al. 2002). qPCR menggunakan molekul reporter fluoresens untuk memonitor produksi dari produk amplifikasi pada setiap siklus reaksi PCR. Pada qPCR akan diperoleh nilaicycle threshold(CT), yaitu siklus ketika intensitas emisi zat warna fluoresens melewati nilaithreshold. Semakin tinggi jumlah awal kopi target asam nukleat, semakin cepat peningkatan fluoresens sehingga semakin rendah nilai CT.
Perumusan Masalah
Ekstrak daun pepaya diketahui memiliki aktivitas antimikroba terhadap bakteri Gram positif maupun Gram negatif. Aktivitas antimikroba ini berhubungan dengan komponen fitokimia yang terdapat dalam daun pepaya, seperti tanin, terpenoid, dan alkaloid.
S. aureus merupakan patogen pada pangan penghasil enterotoksin yang tahan terhadap pemanasan. SEA adalah enterotoksinS. aureusyang paling sering menyebabkan kasus keracunan pangan. Alkaloid diketahui dapat menghambat sintesis DNA sehingga terdapat kemungkinan ekstrak kasar alkaloid daun pepaya dapat mempengaruhi ekspresi gen penyandi SEA dan menghambat produksi enterotoksin tersebut. Oleh karena itu, kajian mengenai aktvitas ekstrak kasar alkaloid daun pepaya terhadap gen penyandi SEA perlu dilakukan.
Tujuan Penelitian
Tujuan umum dari penelitian ini adalah untuk menguji pengaruh penambahan ekstrak kasar alkaloid daun pepaya dalam berbagai konsentrasi terhadap ekspresi gensea.Tujuan khusus penelitian ini adalah untuk memperoleh ekstrak kasar alkaloid daun pepaya dengan ultrasonic assisted extraction, mendapatkan isolatS. aureuspenghasil SEA dari pangan, mengetahui nilai KHM ekstrak kasar alkaloid daun pepaya terhadap S. aureus, serta pengaruh ekstrak kasar alkaloid terhadap jumlah sel dan ekspresi gensea.
Hipotesis
3
Manfaat Penelitian
Penelitian mengenai pengaruh ekstrak kasar alkaloid daun pepaya terhadap ekspresi gen penyandi SEA pada S. aureus ini diharapkan dapat memiliki manfaat, yaitu:
1. Menambah pengetahuan dan wawasan mengenai kegunaan daun pepaya sebagai senyawa antimikroba alami.
4
2 TINJAUAN PUSTAKA
Daun Pepaya
Daun pepaya mengandung senyawa bioaktif, yaitu alkaloid, saponin, senyawa fenolik, flavonoid, terpenoid, dan tanin (Baskaran et al. 2012). Daun pepaya juga memiliki akitivitas proteolitik karena kandungan enzim papain yang dimilikinya (Nwofia et al. 2012). Ekstrak daun pepaya menunjukkan aktivitas antimikrobial terhadap bakteri Gram positif dan negatif serta kapang (Tabel 1). Aktivitas antimikroba dari ekstrak daun pepaya tersebut berhubungan dengan senyawa bioaktif yang terdapat di dalam daun pepaya. Alkaloid, terpenoid, fenolik, flavonoid, dan tanin memiliki kemampuan menghambat mikroba dengan berbagai mekanisme (Tabel 2).
Tabel 1 Aktivitas antimikroba berbagai ekstrak daun pepaya
Jenis ekstrak Mikroba yang dihambat Pustaka
Ekstrak etanol Escherichia coli, Micrococcus luteus, Pseudomonas aeruginosa, Bacillus cereus, Klebsiella pneumoniae, Staphylococcus aureus, Proteus vulgaris, Enterococcus faecalis, Salmonella typhi, S. paratyphiA, Aspergillus niger, A. flavus, Candida albicans, C. Tropicalis
Anibijuwon dan Udeze 2009; Rahmanet al.2011;
Alabiet al.2012; Baskaranet al.2012
Ekstrak metanol E. coli, M. luteus, P. aeruginosa, B. cereus, S.aureus, A. niger, A. flavus, C. albicans, C. Tropicalis
Baskaranet al.2012
Ekstrak etil asetat E. coli, M. luteus, P. aeruginosa, B. cereus, K. pneumoniae, S. aureus, A. niger, A. flavus, C. albicans, C. Tropicalis
Baskaranet al.2012
Ekstrak kloroform
E. coli, M. luteus, P. aeruginosa, K.
pneumoniae, S. aureus, A. niger, A. flavus, C. albicans, C. Tropicalis
Baskaranet al.2012
Ekstrak aseton E. coli, P. aeruginosa, B. cereus, K.
pneumoniae, S. aureus, E. faecalis, A. niger, A. flavus, C. albicans, C. Tropicalis
Alabiet al.2012; Baskaran et al.2012
Ekstrak air panas S. aureus, P. aeruginosa, E. coli, K. Pneumoniae, Proteus mirabilis
Anibijuwon dan Udeze 2009; Baskaranet al.2012
Tabel 2 Mekanisme antimikroba berbagai senyawa bioaktif
Senyawa bioaktif Subkelas Mekanisme antimikroba Pustaka
Alkaloid - Menyisip pada utas ganda DNA Cowan 1999;
Caoet al2007 Mempengaruhi enzim
topoisomerase dan enzim dalam proses perbaikan DNA yang salah
Caoet al2007
Terpenoid - Merusak membran sel Cowan 1999;
Senyawa bioaktif Subkelas Mekanisme antimikroba Pustaka
Fenolik Flavonoid Berikatan dengan adhesin,
membentuk kompleks dengan dinding sel
Cowan 1999
Tanin Berikatan dengan adhesin,
menghambat enzim, membentuk kompleks dengan dinding sel, merusak membran sel
Cowan 1999
Fenolik sederhana (tymol, eugenol, carvacrol)
Mengganggu membran sel, menghambat ATPase, menghambat pelepasan ATP intraseluler
Cetin-Karaca 2011
Quinones Berikatan dengan adhesin,
membentuk kompleks dengan dinding sel
6
Staphylococcus aureusdanStaphylococcal Enterotoxin
Staphylococcus aureus
S. aureusmerupakan bakteri Gram positif berbentuk kokus, non-motil, tidak membentuk spora, dan bersifat anaerob fakultatif. S. aureus termasuk sebagai bakteri mesofilik dan mampu bertahan pada kondisi kering dalam waktu yang cukup lama. Bakteri ini memiliki ketahanan yang tinggi terhadap garam dan gula ([FDA] 2012). S. aureus mampu memproduksi pigmen karotenoid sehingga menghasilkan koloni berwarna keemasan (Schelin et al. 2011). Berbagai faktor lingkungan mempengaruhi pertumbuhan bakteri ini (Tabel 3).
Tabel 3 Faktor yang mempengaruhi pertumbuhanS. aureus
Faktor Batas pertumbuhan Pertumbuhan optimum
Suhu 6 - 48°C 35 - 41°C
pH 4–10 6–7
aw 0.83–0.99 0.99
NaCl 0 - 20% 0%
Oksigen anaerobik-aerobik aerobik
Sumber: Schelinet al.2011
S. aureusdapat ditemukan pada tanah, air, dan udara. Bakteri ini juga dapat ditemukan pada hidung dan kulit hewan berdarah panas. S. aureus merupakan patogen pada manusia yang dapat menyebabkan staphylococcal food poisoning (SFP), toxic shock syndrome, pneumonia, dan infeksi pada luka. Pada kasus keracunan pangan, S. aureus yang mengontaminasi pangan mampu tumbuh dan menghasilkan toksin (Schelinet al.2011).
Staphylococcal Enterotoxin
Terdapat 21 enterotoksin S. aureus yang telah diidentifikasi dan dikelompokkan menjadi enterotoksin klasik (SEA, SEB, SEC, SED, SEE) dan enterotoksin non klasik (SE1G, SElH, SEl, SElJ, SElK, SElL, SElM, SElN, SElO, SElP, SElQ, SER, SES, SET, SElU, SElV) (Schelinet al.2011). SE bersifat tahan panas dan resisten terhadap protease gastrointestinal. Faktor lingkungan yang mempengaruhi pembentukan SE memiliki batas yang berbeda dengan batas faktor yang mempengaruhi pertumbuhanS. aureus(Tabel 4).
Tabel 4 Faktor yang mempengaruhi pembentukan SE
Faktor Batas pembentukan SE Pembentukan SE optimum
Suhu 10 - 46°C 34 - 40°C
pH 5–9.6 7–8
aw 0.86–0.99 0.99
NaCl < 12% 0%
Oksigen anaerobik-aerobik Aerobik
Sumber: Schelinet al.2011
7
paling banyak ditemukan sebagai penyebab SFP adalah SEA (Balaban dan Rasooly 2000).
Staphylococcal EnterotoxinA (SEA)
SEA merupakan polipeptida yang terdiri atas 233 asam amino dan disintesis dari gen sea, yang tersusun atas 774 pasang basa. Gen sea dibawa oleh bakteriofag yang disisipkan pada kromosom bakteri sebagai profag dan berperilaku seperti bagian dari genom bakteri. Transkripsi sea berkaitan dengan siklus hidup dari profag penyandi SEA (Schelinet al. 2011). Polimorfisme alami pada profag ditemukan mempengaruhi jumlah SEA yang diproduksi oleh bakteri pembawa profag. Hasil analisis sekuen daerah promotor menunjukkan bahwa strain yang memproduksi sea dapat dikelompokkan menjadi 2 kelompok, yaitu strain yang memproduksi SEA dalam jumlah tinggi (kelompok SEA1) dan strain yang memproduksi SEA dalam jumlah rendah (kelompok SEA2) (Borst dan Betley 1994). Derzelle et al. (2009) menunjukkan bahwa ekspresi sea tidak dipengaruhi oleh pertumbuhan bakteri. Akan tetapi, produksi SEA pada susu terdeteksi ketika selS. aureustelah mencapai konsentrasi 106.5CFU/ml (Fujikawa dan Morozumi 2006).
SEA mulai diproduksi pada pertengahan fase eksponensial pertumbuhan (Balaban dan Rasooly 2000). Hanya sedikit jumlah SEA yang dibutuhkan untuk menimbulkan penyakit, yaitu kurang dari 1 µg. SEA bersifat seperti superantigen, yang dapat berinteraksi dengan banyak sel T secara non spesifik. Interaksi ini menyebabkan terjadinya pelepasan yang tidak terkontrol dari berbagai sitokin dan menyebabkan terjadinya inflamasi akut dan shock sehingga menimbulkan gejala keracunan, seperti mual dan muntah-muntah (Proft dan Fraser 2003).
Isolasi dan IdentifikasiS. aureus
Isolasi dan IdentifikasiS. aureussecara Konvensional
Isolasi S. aureus dimulai dengan mendeteksi keberadaan S. aureus pada sampel pangan menggunakan media BPA. Media BPA yang disuplementasi dengan kuning telur dan kalium telurit merupakan media selektif diferensial yang direkomendasikan oleh International Organization for Standardization (ISO), Official Analytical Chemists International (AOAC), dan Bacteriological Analytical Manual(BAM), untuk enumerasi dan isolasi S. aureusdari pangan. S. aureus akan membentuk koloni berwarna abu-abu hingga hitam dengan zona bening atau opaque disekeliling koloni. Akan tetapi, BPA tidak cukup selektif untuk menganalisis keberadaan S. aureus pada pangan, terutama pangan dengan flora kontaminan yang tinggi. Beberapa mikroorganisme, seperti staphylococci selain S. aureus, Bacillus spp., dan micrococci membentuk koloni dengan morfologi yang mirip denganS. aureuspada BPA (Capitaet al.2001).
8
menggunakankit, sepertiStaphylase test kit(Oxoid) untuk uji koagulase dan API Staph (bioMérieux) untuk uji biokimia (Di Giannataleet al.2011).
IdentifikasiS. aureussecara Molekuler
Identifikasi isolat terduga S. aureus dapat dilakukan secara molekuler dengan menggunakan teknik PCR dan sekuensing. DNA dari isolat terduga S. aureusdiisolasi dan isolat DNA yang diperoleh digunakan sebagai cetakan untuk mengamplifikasi gen penyandi 16S rRNA dengan PCR. Produk PCR gen penyandi 16S rRNA selanjutnya disekuensing dan sekuen yang diperoleh dibandingkan dengan basis data genom.
Prinsip Isolasi DNA Bakteri Secara umum, terdapat 3 prinsip dalam isolasi DNA bakteri, yaitu perusakan sel bakteri, ekstraksi DNA dengan pelarut organik, serta presipitasi DNA dengan menggunakan alkohol (Mooreet al.2004). Perusakan sel bakteri dapat dilakukan melalui digesti menggunakan enzim dan lisis dengan detergen. Lisozim merupakan enzim yang mengatalisis hidrolisis ikatan β-1,4-glikosidik di antara N-asetilmuramic acid-N-acetylglucosamine pada lapisan peptidoglikan bakteri. Selain enzim lisozim, juga dapat digunakan enzim proteinase-K, yaitu protease serin yang dihasilkan oleh fungiTritirachium album. Enzim ini memotong di dekat gugus karboksil dari asam amino alifatik dan aromatik, yang terlibat dalam ikatan peptida, termasuk pada ikatan sebrang silang peptida yang terdapat pada peptidoglikan dinding sel bakteri.
Detergen juga digunakan untuk mengganggu sel bakteri. Detergen dapat membentuk ikatan yang kuat dengan protein dan menyebabkan denaturasi yang tidak dapat dibalik (irriversible). Detergen terutama efektif untuk mengganggu bakteri yang telah mengalami kerusakan pada dinding selnya. Oleh karena itu, pada proses isolasi DNA, perlakuan dengan detergen diberikan setelah dinding sel diberi perlakuan dengan enzim. Detergen yang dapat digunakan, yaitu sodium dodecylsulfate (SDS) dan cetyl trimethylammonium bromide (CTAB). SDS merupakan detergen anionik, yang pada konsentrasi rendah dapat berikatan pada protein yang terikat pada sel dan lipoprotein. SDS efektif dalam mendenaturasi protein tersebut. CTAB merupakan detergen kationik yang dapat digunakan dalam mengekstraksi DNA bakteri. CTAB dapat mendenaturasi dan mempresipitasi lipopolisakarida dan protein dinding sel. Penggunaan NaCl bersamaan dengan CTAB berfungsi untuk menyediakan kation monovalen (Na+) sehingga mencegah DNA terpresipitasi. Keberadaan kation monovalen dengan konsentrasi di atas 0.5 M akan membuat DNA tetap terlarut.
Proses perusakan sel bakteri tersebut dilakukan dalam larutan bufer (pH 8-9) yang mengandung agen pengelat logam, seperti ethylenediamine-tetraacetic acid (EDTA). pH basa akan mengurangi interaksi elektrostatik antara DNA dan protein yang bersifat basa sehingga memfasilitasi denaturasi protein seluler lain dan menghambat aktivitas nuklease. EDTA dapat mengikat kation divalen (Mg2+ and Mn2+) sehingga mengurangi stabilitas dari dinding dan membran serta menghambat nuklease yang membutuhkan kation logam.
dan lipoprotein serta me sel yang terdenaturasi ak sentrifugasi. Secara umu kloroform lebih efektif digunakan untuk mengur target. Semakin tinggi peningkatan fluoresens s produk PCR dan intens jumlah DNA cetakan pad
menyebabkan denaturasi dan presipitasi. Presipita akan berada pada fase organik dan dapat dipisahka umum, fenol efektif dalam mendenaturasi protein, ktif untuk polisakarida (Moore et al. 2004). Isoam ngurangi pembentukan buih selama proses ekstraksi
h terekstrak kemudian dipresipitasi menggunaka nol) pada suhu rendah. Suhu rendah dapat me ga DNA lebih mudah mengendap. Presipitat DN an etanol 70% untuk menghilangkan sisa-sisa reag
ative Reverse TranscriptionPCR (qRT-PCR)
k (reverse transcription) yang diikuti denganreal metode yang dapat digunakan untuk mende NA. mRNA ditranskripsi balik membentuk cDNA, n sebagai cetakan pada qPCR. Metode ini merupak
mendeteksi dan menguantifikasi tingkat ekspresi g
unakan molekul reporter fluoresens untuk m amplifikasi pada setiap siklus reaksi PCR. Pada q uoresens yang dihasilkan secara proporsional selam DNA. Pengujian real time ini menentukan titik wak
asi produk PCR pertama kali dideteksi. Hal ini di (cycle number) saat intensitas emisi zat warn ound noise(Gambar 2).
urva amplifikasi pada reaksi qPCR (NCBI 2012).
ni dikenal sebagai threshold cycle (CT). CT ditent ksi PCR dan berbanding terbalik dengan jumlah gi jumlah awal kopi target asam nukleat, sema ns sehingga semakin rendah nilai CT. Korelasi line
11
Kuantifikasi Relatif
Pada kuantifikasi relatif, perubahan ekspresi gen sampel diukur relatif terhadap gen referensi (kontrol internal) dan sampel kalibrator. Nilai CT dari sampel target secara langsung dibandingkan dengan nilai CT referensi dan hasilnya dinyatakan sebagai rasio antara target dengan referensi (Wong dan Medrano 2005). Berbagai metode dapat digunakan untuk menganalisis data ekspresi gen relatif. Salah satunya adalah dengan menggunakan metode perbandingan CTatau metode2 .
12
3 METODE PENELITIAN
Tempat dan Waktu
Penelitian dilakukan di Laboratorium Mikrobiologi dan Laboratorium Kimia Pangan PAU IPB, Laboratorium Mikrobiologi Pangan Departemen Ilmu dan Tekonologi Pangan IPB, serta Laboratorium Mikrobiologi dan Laboratorium Bioteknologi Pangan SEAFAST Center IPB, pada bulan Januari hingga September 2013.
Bahan Penelitian
Bahan-bahan yang digunakan untuk mengekstraksi alkaloid dari daun pepaya, antara lain daun pepaya calina (IPB 9), sodium dodecylsulfate (SDS) (Merck & Co., New Jersey, USA), kertas saring Whatman no.1, akuades, H2SO4 (Merck & Co., New Jersey, USA) 2% (v/v), reagen Mayer (HgCl2, KI), Na2CO3 (Merck & Co., New Jersey, USA) 5% (b/v), kloroform (J.T. Baker, Pennsylvania, USA), fenolftalein, Na2SO4(Merck & Co., New Jersey, USA), dan gas N2.
Isolasi S. aureus dilakukan dari beberapa pangan, yaitu ayam suwir, sate jeroan, bakso, telur balado, tumis usus ayam, dan susu sapi mentah. Bahan dan media yang digunakan pada proses isolasi ini, yaitu NaCl 0.85%, baird-parker agar(BPA) (63 g/liter, Oxoid Ltd., Hampshire, UK), kuning telur, kalium telurit 1%, mannitol salt agar (MSA) (111 g/liter, Oxoid Ltd., Hampshire, UK), Staphylase test kit (Oxoid Ltd., Hampshire, UK), dan APIStaph (bioMérieux Inc., North Carolina, USA). Bahan yang digunakan pada penentuan nilai konsentrasi hambat minimum (KHM) ialahtriptone soya agar (TSA) (40 g/liter; Oxoid Ltd., Hampshire, UK), triptone soya broth (TSB) (30 g/liter; Oxoid Ltd., Hampshire, UK), mueller hinton agar (MHA) (38 g/liter; Oxoid Ltd., Hampshire, UK), dimethyl sulfoxide(DMSO) (Merck & Co., New Jersey, USA). Selain itu bakteri S. aureusATCC 25923 juga digunakan sebagai pembanding.
13
Bahan-bahan yang digunakan dalam menganalisis ekspresi gen sea, antara lain bahan untuk ekstraksi RNA, yaitu peqGOLD Bacterial RNA Kit (PEQLAB Biotechnologie GmbH, Erlangen, Jerman) dan DNAse I, RNase free (Thermo Fisher Scientific, Massachusetts, USA); bahan untuk sintesis cDNA, yaitu RevertAid First Strand cDNA Synthesis Kit (Thermo Fisher Scientific, Massachusetts, USA), primer SEA (SEA1 dan SEA2), primer 16S rRNA (16sF dan 16sR3); serta bahan untuk real-time PCR, yaitu KAPA SYBR® FAST qPCR Kit Master Mix (Kappa Biosystems, Massachusetts, USA).
Peralatan Penelitian
Alat-alat yang digunakan dalam penelitian ini adalah alat-alat gelas, pipet mikro 10 ml, pipet mikro 1 ml, pipet mikro 100 µl, pipet mikro 10 µl, blender, oven vakum (VWR A143 A-143, Sheldon Manufacturing, Inc., Oregon, USA), ultrasonic bath (Bransonic Ultrasonic Cleaner model 8510E MTH, Branson Ultrasonic Corporation, Connecticut, USA), rotari evaporator (Butchi Rotavapor R-210, BÜCHI Labortechnik, Flawil, Switzerland),stomacher (BagMixer®400P, Interscience, Perancis), sentrifuge (Hermle Z383K; Hermle Labortechnik GmbH, Wehingen, Saint Nom, Jerman), spektrofotometer UV-1800 (Shimadzu, Jepang), perangkat elektroforesis DNA (Bio-Rad, Bio-Rad Laboratories Pte. Ltd, Singapore), Thermal Cycler 2720 (Applied Biosystems, California, USA),gel doc (Bio-Rad, Bio-Rad Laboratories Pte. Ltd, Singapore), Swift Spectrum Themal Cycler 48 (Esco Healthcare Pte. Ltd., Singapore).
Prosedur Penelitian
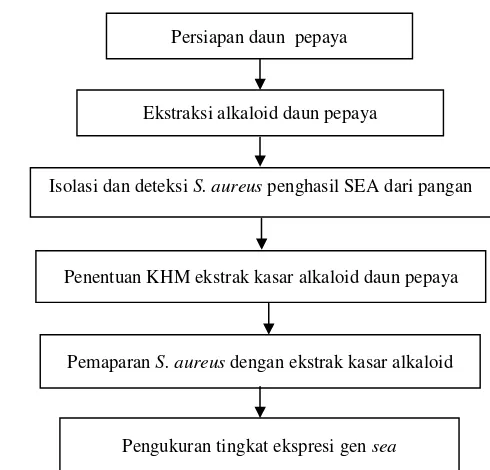
Tahapan di dalam prosedur penelitian ini dapat dilihat pada Gambar 4. Di dalam tahap isolasi dan deteksi S. aureus penghasil SEA dari pangan, deteksi keberadaan genseahanya dilakukan pada 10 isolatS. aureusyang diperoleh dari sampel pangan. Dari isolat-isolatS. aureuspenghasil SEA yang diperoleh, hanya 1 isolat yang digunakan pada tahapan penentuan nilai Konsentrasi Hambat Minimum (KHM), pemaparanS. aureusdengan ekstrak kasar alkaloid, serta pada pengukuran tingkat ekspresi gensea.
Persiapan Daun Pepaya
14
Ekstraksi alkaloid daun pepaya Persiapan daun pepaya
Isolasi dan deteksiS. aureuspenghasil SEA dari pangan
Gambar 4 Diagram alir tahapan penelitian.
Ekstraksi Alkaloid Daun Pepaya
Daun pepaya kering sebanyak 10 g disuspensikan dalam 400 ml SDS 0.2 % dan disonikasi selama 2.5 jam di dalam ultrasonic bath pada suhu 25 - 35°C. Ekstrak dipisahkan dengan kain saring dan residu pada kain saring dicuci dengan 20 ml akuades. Selanjutnya, ekstrak disaring dengan kertas Whatman no. 1. Ke dalam filtrat ditambahkan larutan H2SO42% hingga diperoleh pH 3-4 dan alkaloid dipresipitasi dengan 15 ml reagen Mayer. Presipitat dipisahkan dengan sentrifugasi pada 2 400 × g selama 10 menit dan dilarutkan dengan Na2CO35% lalu diekstraksi dengan CHCl3. Lapisan organik yang terbentuk dicuci dengan akuades hingga pH menjadi netral dan dilewatkan pada Na2SO4. Larutan dievaporasi dengan rotari evaporator dan dikeringkan dengan gas N2 untuk memperoleh alkaloid (Djilani et al. 2006). Ekstrak alkaloid dilarutkan di dalam DMSO sebelum digunakan dalam analisis.
IsolasiS. aureusdari Pangan
S. aureusdiisolasi dari beberapa jenis pangan, yaitu ayam suwir, sate jeroan, telor balado, tumis usus, susu sapi mentah, dan bakso. Isolat S. aureus diperoleh melalui beberapa tahapan, yaitu isolasi bakteri terduga dari sampel pangan, identifikasi isolat terduga S. aureus secara konvensional, dan identifikasi isolat terdugaS. aureussecara molekuler dengan menggunakan PCR. Identifikasi isolat secara molekuler terdiri atas isolasi DNA bakteri, amplifikasi gen penyandi 16S rRNA, dan sekuensing.
Isolasi Bakteri Isolasi bakteriS. aureusdilakukan dengan menggunakan metode BAM (Bennet dan Lancette 2001) dengan modifikasi. Sebanyak 25 g atau 25 ml sampel pangan dimasukkan ke dalam plastik steril dan ditambahkan 225 ml larutan garam fisiologis 0.85% sebagai pengencer. Sampel dihancurkan dengan
Penentuan KHM ekstrak kasar alkaloid daun pepaya
PemaparanS. aureusdengan ekstrak kasar alkaloid
15
menggunakan stomacher selama 2 menit. Selanjutnya, dibuat seri pengenceran sampel, yaitu 10-1, 10-2, dan 10-3. Dua puluh lima gram sampel yang telah dihancurkan di dalam 225 ml larutan pengencer merupakan pengenceran 10-1.
Sebanyak 1 ml sampel pengenceran 10-1 dipipet dan dibagi ke dalam 3 cawan media BPA, yang disuplementasi dengan 50 ml/L larutan kuning telur–1% kalium telurit (kuning telur: NaCl 0.85%: kalium telurit 1% = 2:2:1). Selanjutnya, sebanyak 0.1 ml sampel pengenceran 10-1, 10-2, dan 10-3 masing-masing disebar ke dalam 1 cawan media BPA. Sampel diinkubasi pada 37°C selama 18 - 24 jam. Koloni dengan ciri-ciri tipikal S. aureus (bulat, cembung, berdiameter 2-3 mm pada pertumbuhan yang tidak padat, berwarna hitam, dan dikelilingi dengan zona buram (opaque) atau bening diambil dengan ose tusuk dan digoreskan pada media MSA dan diinkubasi pada 37°C selama 18 -24 jam. Koloni yang dapat mengubah warna media MSA menjadi kuning selanjutnya digunakan pada uji koagulase. Uji koagulase dilakukan menggunakan Staphylase Test Kit. Isolat yang mampu menghasilkan koagulase akan membentuk aglutinasi saat dicampurkan dengan reagen.
Identifikasi Isolat Terduga S. aureus dengan Metode Konvensional Isolat dengan hasil koagulase positif selanjutnya diidentifikasi dengan uji biokimia menggunakan analytical profile index (API) Staph. Hasil API kemudian dibaca dengan menggunakanapiwebTMidentification sofware.
Isolasi DNA Isolasi DNA S. aureus menggunakan metode Masonet al. (2001), dengan modifikasi, yaitu penggunaan lysostaphin digantikan dengan lisozim 10 mg/ml serta tanpa pemakaian RNase. Isolat yang menunjukkan positif S. aureus pada API ditumbuhkan pada media TSB dan diinkubasi pada 37°C selama 18–24 jam. Sebanyak 2 ml kulturS. aureuspada media TSB disentrifugasi pada 21 000 × g selama 1 menit. Supernatan dibuang dan pelet diresuspensi menggunakan 560 μ l bufer TE 1x. Selanjutnya, ditambahkan 100 μ l lisozim (10 mg/ml) kemudian dicampurkan hingga homogen dan diinkubasi pada 37°C selama 1 jam (tabung dibolak-balik setiap 15 menit). Sebanyak 30 μ l SDS 10% dan 10 μ l proteinase K (10 mg/ml) ditambahkan dan diinkubasi kembali pada 37°C selama 1 jam (tabung dibolak-balik setiap 15 menit).
Setelah itu, 100 μ l NaCl 5M dan 80μ l CTAB-NaCl (10% CTAB di dalam 0.7 M NaCl) yang telah dipanaskan pada 65°C ditambahkan kemudian diinkubasi selama 10 menit pada 65°C. Kloroform dengan volume yang sama dengan suspensi ditambahkan lalu divorteks dan disentrifugasi pada 21 000 × g selama 5 menit. Supernatan dipindahkan ke dalam tabung baru dan diekstraksi 2 kali dengan PCI (25:24:1) dan 1 kali dengan CI, dengan volume yang sama dengan suspensi.
16
Kuantifikasi DNA Konsentrasi dan kemurnian isolat DNA ditentukan dengan mengukur absorbansi pada panjang gelombang 260 nm dan 280 nm. Isolat DNA diencerkan 1:1000 menggunakan bufer TE 1x dan diukur absorbansinya (A260dan A280). Konsentrasi DNA (ng/µl) dapat dihitung dengan rumus A260x 50 x faktor pengenceran (1 OD A260= 50 ng/µl dsDNA). Kemurnian DNA dapat ditentukan dengan mengukur rasio A260/A280. Kemurnian DNA yang baik memiliki nilai rasio antara 1.8 sampai 2.0 (Johnson & Tyler 1993).
Amplifikasi Gen Penyandi 16S rRNA Amplifikasi gen penyandi 16S rRNA dilakukan dengan menggunakan Thermal Cycler 2720. Isolat DNA diamplifikasi dengan primer 16sF dan 16sR3 (Tabel 5). Campuran reaksi PCR sebanyak 25 μ l terdiri atas 12.5 µl DreamTaq Green master mix, 1 µl setiap primer (10 µM), 2 µl DNA cetakan (1000 ng/µl), dan 8.5 µl air bebas nuklease. Siklus PCR yang digunakan, yaitu 1 siklus denaturasi selama 5 menit pada 95°C, 30 siklus amplifikasi (denaturasi 1 menit pada 95°C, annealing 1 menit pada 55°C, dan extension1 menit pada 72°C), dan terminasi selama 5 menit pada 72°C (Leeet al. 2007).
Produk hasil amplifikasi divisualisasikan pada gel agarosa 1.5% dengan elektroforesis pada tegangan 120 V selama 35 menit. Sebagai marker digunakan 100-bp plus DNAladder. Gen penyandi 16S rRNA menghasilkan pita berukuran 240 bp.
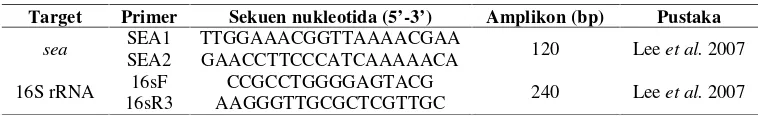
Tabel 5 Primer yang digunakan pada PCR dan qRT-PCR
Target Primer Sekuen nukleotida (5’-3’) Amplikon (bp) Pustaka
sea SEA1SEA2 TTGGAAACGGTTAAAACGAAGAACCTTCCCATCAAAAACA 120 Leeet al.2007
16S rRNA 16sR316sF AAGGGTTGCGCTCGTTGCCCGCCTGGGGAGTACG 240 Leeet al.2007
Sekuensing Sekuensing dilakukan untuk mengetahui urutan basa DNA dari gen penyandi 16S rRNA isolat S. aureus dari pangan. Sekuensing dilakukan dengan mengirimkan hasil PCR gen penyandi 16S rRNA ke 1st Base Pte Ltd, Singapura, melalui PT. Genetika Science Indonesia. Sekuen yang diperoleh dianalisis dengan program Basic Local Alignment Search Tool (BLAST) (http://blast. ncbi.nlm.nih.gov/).
Identifikasi gensea
Identifikasi isolat S. aureus penghasil SEA dilakukan secara molekuler dengan menggunakan teknik PCR. Amplifikasi genseadilakukan dengan metode yang sama dengan amplifikasi gen penyandi 16S rRNA. Primer yang digunakan adalah primer SEA1 dan SEA2 (Tabel 5). Gen sea ditunjukkan dengan pita berukuran 120 bp. Isolat S. aureus penghasil SEA diinokulasikan pada TSA miring sebagai stok.
Penentuan Konsentrasi Hambat Minimum
17
0.85% hingga mencapai kekeruhan setara dengan kekeruhan standar 0.5 McFarland (1.5 × 108CFU/ml). Kekeruhan diukur dengan spektrofotometer pada panjang gelombang 625 nm dengan nilai absorbansi 0.08. Nilai absorbansi tersebut setara dengan 0.5 standar McFarland (Andrews 2005; Oliveira dan de Lencastre 2011).
Suspensi bakteri diinokulasikan menggunakan cotton swab steril pada seluruh permukaan MHA. Setelah itu, sumur dibuat menggunakan pembolong agar steril berdiameter 4 mm. Sebanyak 20 µl ekstrak kasar alkaloid daun pepaya (10 mg/ml, 8 mg/ml, 6 mg/ml, 4 mg/ml, 2 mg/ml, 1 mg/ml) dipipet ke dalam lubang pada media. Media didiamkan selama 1 jam pada suhu ruang dan diinkubasi tanpa dibalik pada 37 ºC selama 18-24 jam (Ehsan et al. 2009; Subramanian dan Saratha 2010). Sebagai kontrol negatif digunakan DMSO.
Penentuan KHM dilakukan dengan metode pengenceran makro. Sebanyak 100 μ l suspensi S. aureus berumur 18-24 jam dengan konsentrasi 106 CFU/ml diinokulasikan ke dalam 1 ml media TSB yang mengandung ekstrak kasar alkaloid daun pepaya pada berbagai konsentrasi (kisaran konsentrasi berdasarkan hasil penapisan). Kultur bakteri kemudian diinkubasi pada 37 °C selama 24 jam dan digoyang dengan kecepatan 150 rpm (Mazzolaet al.2009; Subramanian dan Saratha 2010). Selanjutnya, dibuat seri pengenceran dari kultur bakteri pada setiap konsentrasi ekstrak alkaloid dan disebar pada media TSA. Media tersebut diinkubasi pada 37°C selama 48 jam dan dilakukan penghitungan jumlah bakteri. KHM90 merupakan konsentrasi ekstrak kasar alkaloid daun pepaya terendah yang dapat menghambat 90% pertumbuhan bakteri dibandingkan dengan jumlah awal inokulum (Fazeli dan Salehi 2007).
PemaparanS. aureusdengan Ekstrak Kasar Alkaloid Daun Pepaya
Sebanyak 100 μ l suspensiS. aureuspenghasil SEA, dengan konsentrasi 108 CFU/ml, dipipet ke dalam 5 ml media TSB yang telah ditambahkan dengan ekstrak kasar alkaloid daun pepaya dengan konsentrasi 0, 1, dan 2 KHM. Campuran tersebut diinkubasi pada suhu ruang selama 2 jam dan digoyang dengan kecepatan 150 rpm. Jumlah sel bakteri, sebelum dan setelah diinkubasi selama 2 jam, ditentukan dengan menggunakan metode agar sebar pada TSA.
Pengukuran Tingkat Ekspresi Gensea
Tingkat ekspresi gen sea dari bakteri S. aureus yang telah dipaparkan dengan ekstrak kasar alkaloid daun pepaya dapat diketahui melalui beberapa tahapan, yaitu, isolasi mRNA bakteri, sintesis cDNA, serta pengukuran ekspresi genseamenggunakanreal-timePCR.
18
telah dipasangkan dengancollection tube(CT) dan disentrifugasi pada 10 000 × g selama 2 menit.
Etanol 70% 1x vol (sekitar 550 µl) ditambahkan ke dalam sampel pada CT dan dicampurkan dengan pipet. Sebanyak 650 µl sampel dipindahkan ke dalam PerfectBind RNA column yang telah dipasangkan dengan CT dan disentrifugasi pada 10 000 × g selama 1 menit. Residu sampel yang terdapat pada CT kembali dimasukkan ke dalam PerfectBind RNA column yang sama dan disentrifugasi 10 000 × g selama 1 menit.
PerfectBind RNA column tersebut kemudian dipasangkan dengan CT baru dan ditambahkan 500 µl RNA wash buffer I lalu disentrifugasi selama 1 menit
PerfectBind RNA columndipasangkan dengan CT dan disentrifugasi selama 2 menit pada 10 000 × g untuk mengeringkan matriks kolom. Selanjutnya, PerfectBind RNA column tersebut dipasangkan dengan tabung 1.5 ml dan sebanyak 30 – 50 µl air bebas nuklease ditambahkan langsung pada matriks, diinkubasi 1 menit, dan disentrifugasi selama 1 menit pada 6 000 × g untuk mengelusi RNA. Kemurnian RNA ditentukan dengan mengukur rasio A260/A280 dengan spektofotometer UV.RNA yang diperoleh diberi perlakuan denganDNAse I, RNase freeuntuk mendegradasi DNA yang terdapat ekstrak RNA.
Sintesis cDNA Sintesis cDNA menggunakan RevertAid First Strand cDNA Synthesis Kit dengan primer spesifik untuk gensea dan gen penyandi 16S rRNA sebagai gen referensi (Tabel 5). Prosedur sintesis cDNA dilakukan sesuai dengan petunjuk yang terdapat padakit. Campuran reaksi terdiri atas 1 µg RNA total, 20 pmol setiap primer, dan air bebas nuklease hingga volume 12 µl. Campuran diinkubasi pada 65ºC selama 10 menit. Selanjutnya, ditambahkan 4 µl bufer reaksi 5x, 1 µl RiboLock RNase Inhibitor (20 u/µl), 2 µl dNTP 10 mM, dan RevertAid M-MuLV Reverse Transcriptase (200 u/µl). Campuran reaksi diinkubasi pada suhu 42ºC selama 60 menit. Reaksi dihentikan melalui pemanasan pada 70ºC selama 5 menit.
19
Analisis Data
20 yang baik, yaitu berkisa Ekstrak kasar alkaloid d merupakan surfaktan solubilisasi, dispersi, dan SDS dapat meningkatka udara pada permukaan antara larutan dengan da ekstrak berwarna hijau
4 HASIL DAN PEMBAHASAN
kstrak Kasar Alkaloid Daun Pepaya
lkaloid diperoleh dari daun pepaya yang telah dik dengan kadar air 78.17 ± 0.10% dikeringkan meng 22 jam hingga diperoleh kadar air daun kering, yai
kering diblender hingga membentuk bubuk (Ga ya kering adalah 20.62 ± 1.77%. Pada Nwofiaet al
emiliki kadar air sebesar 81 – 85%. Perbedaan rjadi karena adanya perbedaan jenis pepaya, mus
pat penanaman pohon.
Gambar 5 Daun pepaya kering.
s ekstraksi alkaloid, daun pepaya kering ditamba 0.2% dan disonikasi sehingga diperoleh rendeme kisar antara 0.48% – 1.82% per berat kering daun
d diperoleh dari 7 kali proses ekstraksi yang berbe n anionik yang memiliki sifat membasahi
dan emulsifikasi (Djilani et al. 2006). Sifat memb tkan kemampuan dari air atau larutan untuk meng
n zat cair atau padat. Hal ini akan meningkatk n daun pepaya kering. Solubilisasi merupakan keada
cair) larut secara spontan melalui interaksi dengan larut. Interaksi tersebut dapat menurunkan
at terlarut sehingga dapat meningkatkan kelarutan memungkinkan suatu zat dapat larut pada pelarut, tidak dapat larut (Rosen 2004).
berperan di dalam proses dispersi padatan di dal kan membasahi daun pepaya kering dan memindahka
rmukaan daun pepaya kering, memecah kelompok atuan kembali partikel yang telah terdispersi un pepaya kering dapat terdispersi di dalam larutan
n. Selain itu, kemampuan SDS dalam emulsifi ntara dua fase cair yang tidak dapat dicampur. Ke ahi, mendispersi, menyolubilisasi, dan mengem kelarutan senyawa di dalam daun pepaya kering pada
pepaya kering dengan pe alkaloid dari garam me diekstraksi dengan kl ekstrak kasar alkaloid konvensional membutuh
n penyaringan menggunakan kain saring dan ker elanjutnya, pH dari ekstrak diturunkan me n H2SO4. Perlakuan dengan larutan asam dapat me
DS. Alkaloid akan bersatu dengan asam membent larutan asam, warna larutan berubah dari hijau tu bar 6b). Setelah itu, reagen Mayer ditambahkan ndapkan alkaloid. Reagen Mayer terdiri atas merkur Atom merkuri di dalam reagen ini akan membentuk
yang terdapat pada alkaloid. Pasangan ion ters yang bersifat tidak larut.
an disentrifugasi untuk memisahkan endapan alk loid dilarutkan dengan larutan Na2CO3 untuk me
apan yang bersifat asam. Kondisi basa ini akan menjadi bentuk bebas. Larutan basa tersebut s kloroform. Proses ekstraksi dengan klorofor an, yaitu lapisan air di atas dan lapisan organik
(b) (c) (d)
kstraksi alkaloid: (a) setelah sonikasi dan penyar penambahan larutan H2SO4, (c) lapisan yang terbe
dengan kloroform, (d) ekstrak kasar alkaloid .
n memisahkan alkaloid dari pengotor dan garam apisan air. Ekstrak kasar alkaloid di dalam larutan ngan Na2SO4 untuk mengikat sisa air yang ter vaporasi serta gas N2untuk menguapkan kloroform
ak kasar alkaloid yang diperoleh pada penelitian i bandingkan ekstrak kasar alkaloid yang diekstraks konvensional. Sudsai (2006) menggunakan metode
mengekstraksi alkaloid dari daun pepaya dan oid daun pepaya sebesar 0.284%. Metode a butuhkan pelarut bersifat asam dan basa yang lebi
yang lebih lama. Selain karena perbedaan jenis pe enggunaan sonikasi dapat meningkatkan rendemen s ekstraksi dari material tanaman kering, materia
pelarut yang akan memfasilitasi terjadiny hidrasi. Selanjutnya, terjadi transfer massa seny
22
dapat larut, dari material tanaman ke dalam pelarut melalui proses difusi dan osmosis. Pada Ultrasonic Assisted Extraction (UAE), ultrasonik dapat memfasilitasi proses hidrasi dan pengembangan sehingga menyebabkan pembesaran pori pada dinding sel. Hal ini akan meningkatkan proses difusi dan transfer massa (Vinatoru 2001).
UAE merupakan teknik ekstraksi yang mudah, efisien, dan tidak mahal. Gelombang ultrasonik (20 kHz – 100 MHz) melewati medium dengan membentuk tekanan dan ekspansi. Proses ini menghasilkan fenomena yang disebut peronggaan (cavitation), yaitu terjadinya produksi, pengembangan, dan pemecahan gelembung. Hanya material cair dan material cair dengan padatan yang memiliki efek peronggaan. Energi ultrasonik dapat memfasilitasi pelepasan komponen organik dan inorganik dari matriks tanaman. Gelombang ultrasonik dapat meningkatkan transfer massa dan mempercepat akses pelarut menuju material sel pada bagian tanaman. Mekanisme ekstraksi oleh gelombang ultrasonik melibatkan 2 tipe fenomena fisik, yaitu proses difusi melewati dinding sel dan pelepasan isi sel setelah dinding mengalami kerusakan (Azmir et al. 2013).
UAE dapat memberikan beberapa keuntungan, seperti mempersingkat waktu ekstraksi serta mengurangi penggunaan energi dan pelarut. Selain itu, penggunaan UAE dapat meningkatkan jumlah ekstrak yang dihasilkan. Djilaniet al. (2006) menunjukkan bahwa penggunaan UAE dan larutan SDS dapat peningkatkan jumlah total alkaloid yang diekstraksi dari daun Ruta graveolens and Hyoscyamus muticus, dibandingkan dengan metode konvensional (dengan pelarut) atau dengan menggunakan soxhlet.
IsolatS. aureusPenghasil SEA
Isolat TerdugaS. aureusdari Pangan
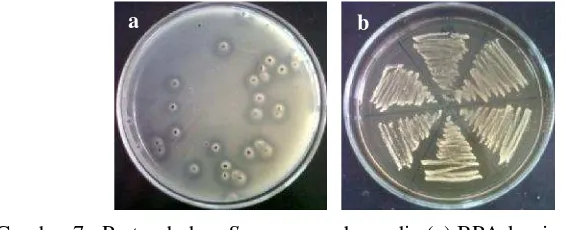
Isolat dengan bentuk koloni tipikal S. aureus pada media BPA hanya ditemukan pada sampel sate jeroan, telur balado, tumis usus, dan susu sapi mentah, sedangkan pada sampel ayam suwir dan bakso, ciri-ciri koloni sulit untuk diamati. Terdapat 66 isolat terdugaS. aureuspada media BPA. Akan tetapi, hanya 62 isolat yang dapat tumbuh dan menunjukkan ciri koloni S. aureuspada media MSA.
BPA dengan kuning telur dan telurit merupakan media yang mengandung karbon dan nitrogen sebagai sumber nutrisi untuk pertumbuhan. Glisin, litium klorida, dan kalium telurit merupakan agen selektif yang akan menghambat pertumbuhan berbagai bakteri pada pangan, tanpa menghambat pertumbuhan S. aureus.S. aureusmemiliki kemampuan dalam mereduksi telurit menjadi telurium. Hal ini menyebabkan koloniS. aureusmemiliki warna abu-abu hingga hitam pada media ini. Kemampuan S. aureusdalam menghasilkan lesitinase dapat memecah lesitin yang terdapat dalam kuning telur. Pemecahan lesitin ditunjukkan dengan terbentuknya zona bening disekeliling koloni (Gambar 7a) (Capitaet al. 2001).
23
termasuk S. aureus dapat tumbuh pada kadar NaCl 7.5%.S. aureus juga mampu memfermentasikan manitol dan menghasilkan asam. Kondisi asam akan mengubah merah fenol pada media dari merah menjadi kuning (Gambar 7b) (Shields dan Tsang 2013).
Gambar 7 PertumbuhanS. aureuspada media (a) BPA-kuning telur telurit (b) MSA.
Seluruh isolat dengan ciri koloni tipikal S. aureus pada media MSA kemudian diuji kemampuannya dalam memproduksi koagulase. Produksi koagulase merupakan salah satu uji yang digunakan untuk membedakan isolat S. aureusdari bakteri lain, termasuk dari staphylococci lain, seperti Staphylococcus epidermidis. S. aureus dapat memproduksi koagulase bebas maupun terikat. Kemampuan S. aureusdalam menghasilkan koagulase dapat diuji menggunakan Staphylase Test Kit. Reagen yang digunakan pada uji ini mengandung sel darah merah domba yang disensitisasi dengan fibrinogen kelinci. Semua isolat yanng diuji memberikan hasil positif koagulase. Bakteri yang mampu menghasilkan koagulase, seperti S. aureus dan isolat S1, akan membentuk aglutinasi saat direaksikan dengan reagen ini. Aglutinasi ditunjukkan dengan terbentuknya titik-titik berwarna coklat (Gambar 8). Berbeda dengan S. aureus, Escherichia coli adalah bakteri yang tidak mampu memproduksi koagulase sehingga tidak membentuk aglutinasi.
Gambar 8 Uji aktivitas koagulase isolatS. aureus. 1=S. aureusATCC 25923, 2 = isolat S1, 3.Escherichia coli.
Isolat klinis S. aureus dapat mensekresikan dua faktor penggumpalan (clotting factor), yaitucoagulase (Coa) danvon-Willebrand factor binding protein (vWbp). Coa dan vWbp dapat mengikat dan mengaktifkan protrombin, yang akan mengubah fibrinogen menjadi fibrin. Hal ini akan menyebabkan terjadinya penggumpalan pada plasma atau darah (Chenget al.2010).
Hasil Identifikasi Isolat TerdugaS. aureusdengan Metode Konvensional Isolat dari pangan yang memiliki ciri positif S. aureus pada media BPA, MSA, dan memproduksi koagulase selanjutnya digunakan pada uji biokimia
No. Isolat Sumber pangan API Staph
1 S1 susu sapi mentah 85% positifS. aureus
2 S4 susu sapi mentah 97.8 % positifS. aureus
3 S10 susu sapi mentah 97.8 % positifS. aureus
4 TB1 telur balado 85% positifS. aureus
5 TB10 telur balado 97.8 % positifS. aureus
6 UA1 tumis usus 85% positifS. aureus
7 UA2 tumis usus 97.8 % positifS. aureus
8 UA13 tumis usus 97.8 % positifS. aureus
9 SJ1 sate jeroan 66.8 % positifS. aureus
Hasil Identifikasi Isolat analisis profil plasmid p aureus tersebut memili
olat TerdugaS. aureusdengan PCR
DNA yang diperoleh divisualisasikan dengan elek , dengan voltase 120 V, selama 45 menit. Pada ha us, diperoleh 3 pita DNA, yaitu pita yang terdapa 1500 bp, di antara marker 2000 bp dan 3000 bp, se
bar 10).
Contoh hasil elektroforesis isolat DNA genom S. aureus. M= marker 100 bp plus DNA ladder, 1=S. aureusATCC 25293, 2= Isolat UA1.
us memiliki ukuran yang besar, yaitu berkisar a bp (Suzukiet al. 2012). Pita yang berada di atas pi
enunjukkan DNA kromosom, sedangkan kedua pi daan DNA plasmid. Pyzik dan Marek (2013) m
d pada isolat S. aureus yang diisolasi dari telur iliki plasmid dengan ukuran kecil, yaitu antara
n kemurnian DNA ditentukan melalui nilai absorb ng gelombang 260 dan 280 nm. Kemurnian is
3.7, sedangkan konsentrasi DNA berkisar antara Isolat UA1 dan UA13 memiliki kemurnian DNA
sio A260/A280di antara 1.8–2.0.
ngan nilai rasio A260/A280 di bawah 1.8 mengindi sedangkan nilai rasio lebih besar dari 2.0 menu ng ikut terekstrak (Moore et al. 2004). Sebagian b h memiliki nilai rasio lebih besar dari 2.0. Hal
s isolasi DNA tidak digunakan RNase ya Walaupun memiliki kemurnian yang kurang ba h dapat digunakan sebagai cetakan dalam PCR de
ampu mendeteksi keberadaan gen penyandi 16S r antara isolat yang memiliki atau tidak memiliki g
26
Tabel 7 Konsentrasi dan kemurnian isolat DNA
No Isolat A260/280 Konsentrasi (ng/µl)
1 S1 3.7 1387.5
2 S4 2.9 1662.5
3 S10 3.7 1375
4 TB1 1.7 4600
5 TB10 3.2 1575
6 UA1 1.9 1487.5
7 UA2 3.5 1500
8 UA13 1.8 1450
9 SJ1 2.2 1237.5
10 SJ4 3.4 1525
Sekuen Gen Penyandi 16S rRNA IsolatS. aureus Hasil elektroforesis produk
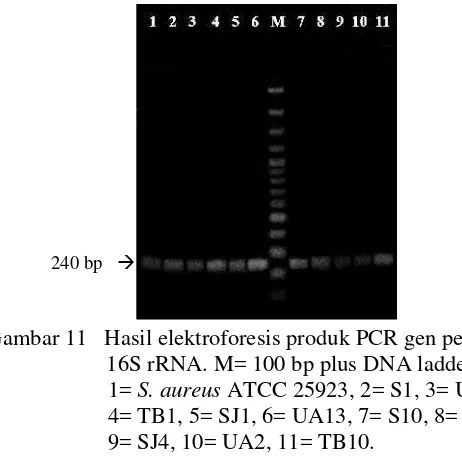
PCR gen penyandi 16S rRNA dengan primer 16sF dan 16sR3 menunjukkan bahwa sama seperti S. aureus ATCC 25923, kesepuluh isolat S. aureus dari pangan menghasilkan amplikon dengan ukuran 240 bp (Gambar 11).
Gambar 11 Hasil elektroforesis produk PCR gen penyandi 16S rRNA. M= 100 bp plus DNA ladder, 1=S. aureusATCC 25923, 2= S1, 3= UA1, 4= TB1, 5= SJ1, 6= UA13, 7= S10, 8= S4, 9= SJ4, 10= UA2, 11= TB10.
Dari hasil sekuensing dapat diketahui sekuen dari gen penyandi 16S rRNA yang dimiliki isolat S. aureus (Lampiran 1). Dengan mengetahui sekuen isolat-isolat tersebut, dapat dilakukan analisis bioinformatika dengan BLAST. Dengan demikian, dapat diketahui sekuen bakteri yang mirip dengan sekuen isolat-isolat tersebut. Semua isolat yang disekuensing memiliki sekuen gen penyandi 16S rRNA yang mirip dengan sekuen genom S. aureus subsp. aureus ST288 yang berukuran 2759510 bp (Tabel 8).
27
Tabel 8 Hasil analisis sekuen penyandi gen 16S rRNA isolat S. aureus dengan program BLAST
Isolat Ukuran sekuen(bp) Sekuen bakteri yang mirip E value Identities(%)
S1 215 genomStaphylococcus aureussubsp.
aureusST288, isolat 10338 2 x 10
-101 99
S4 217 genomStaphylococcus aureussubsp.
aureusST288, isolat 10338 2 x 10
-101 99
S10 215 genomStaphylococcus aureussubsp.
aureusST288, isolat 10338 6 x 10
-102 99
TB1 212 genomaureusStaphylococcus aureusST288, isolat 10338subsp. 1 x 10-98 99
UA1 213 genomStaphylococcus aureussubsp.
aureusST288, isolat 10338 4 x 10-99 99
UA2 217 genomStaphylococcus aureussubsp.
aureusST288, isolat 10338 2 x 10-101 99
UA13 212 genomaureus ST288, isolat 10338Staphylococcus aureussubsp. 1 x 10-98 99
SJ1 210 genomStaphylococcus aureussubsp.
aureusST288, isolat 10338 2 x 10-97 99
SJ4 216 genomStaphylococcus aureussubsp.
aureusST288, isolat 10338 9 x 10-100 99
Berdasarkan basis data genomS. aureussubsp.aureusST288, isolat 10338 (GenBank: HE579059.1), bakteri ini memiliki gen penyandi SEA berukuran 774 bp, yaitu pada basa ke-1988966 hingga basa ke-1989739 (NCBI 2013). Hal ini mengonfirmasikan hasil identifikasi isolat terdugaS. aureusdengan menggunakan API Staph. Identifikasi isolat terduga dengan metode konvensional dan PCR menunjukkan bahwa semua isolat yang diperoleh ialahS. aureus.
Amplikon Gensea
Penggunaan primer SEA1 dan SEA2 akan mengamplifikasi gen sea sehingga menghasilkan amplikon berukuran 120 bp. Amplikon berukuran 120 bp tersebut akan berada di antara marker berukuran 100 bp dan 200 bp. Bakteri yang tidak memiliki gen sea, tidak akan menghasilkan amplikon tersebut. Hasil elektroforesis produk PCR menunjukkan bahwa isolat S1, S4, S10, TB1, UA1, UA2, UA13, SJ1, dan SJ4 memiliki gen sea (Gambar 12). Keberadaan gen ini menunjukkan bahwa isolat-isolat tersebut dapat memproduksi SEA. Di sisi lain, isolat TB10 tidak memiliki gen sea seperti S. aureus ATCC 25923 yang tidak memiliki gen sea sehingga tidak memproduksi SEA. Hal ini mengonfirmasikan bahwaS. aureusATCC 25923 memang tidak menghasilkan SEA.
28 ekstrak, semakin besar diameter zona pengham
sil elektroforesis produk PCR gensea. M= 100 bp us DNA ladder, 1=S. aureusATCC 25923, 2= S1, UA1,4= TB1, 5= SJ1, 6= UA13, 7= S10, 8= S4, SJ4, 10= UA2, 11= TB10.
2008) mendeteksi gen toksin pada isolatS. aureusya n susu pasteurisasi menggunakan teknik PCR. Di a nterotoksin klasik (SEA-SEE), genseamerupakan
temukan pada isolat dari susu tersebut. Sebanyak 4 kuti oleh gen sec (20.5%),sed (3.7%), seb(7.7% pada bahan pangan, S. aureus pembawa gen
n dan rongga hidung dari para pengolah makanan. eksi gen enterotoksin pada isolatS. aureusyang be ngan pengolah makanan yang bekerja di dapur indus
ksi dilakukan dengan teknik PCR dan menggunaka banyak 17 isolat S. aureus diperoleh dan 9 isola emiliki gen sea, diikuti oleh gen see (40.8%) 7%).
bat Minimum Ekstrak Kasar Alkaloid Daun Pe terhadapS. aureus
alkaloid pada setiap konsentrasi menunjukkan apS. aureus SJ1, sedangkan DMSO sebagai pelar
as penghambatan (Gambar 13). Semakin tinggi kon ar diameter zona hambat yang dihasilkan. Secara
hambatan yang dihasilkan oleh ekstrak kasar tidak berbeda nyata dengan konsentrasi 8 mg/ml ( konsentrasi 1 mg/ml masih memberikan zona peng 34 (Gambar 14). Dengan demikian, alkaloid deng
29
Gambar 13 Zona penghambatan ekstrak kasar alkaloid daun pepaya terhadap S. aureusSJ1.
Gambar 14 Diameter zona hambat ekstrak kasar alkaloid daun pepaya terhadapS. aureusSJ1.Nilai dengan huruf yang berbeda adalah berbeda nyata (P < 0.05).
Ekstrak kasar alkaloid daun pepaya pada konsentrasi 10 mg/ml memberikan zona penghambatan yang lebih kecil dibandingkan dengan zona hambat ekstrak etanol daun pepaya. Pada konsentrasi 10 mg/ml, ekstrak etanol tersebut membentuk 15 mm zona hambat terhadapS. aureus(Rahmanet al.2011). Hal ini dapat terjadi karena ekstrak etanol daun pepaya mengandung berbagai komponen fitokimia yang dapat bekerja sama dan memberikan aktivitas antibakterial yang lebih besar terhadap S. aureus. Cowan (1999) melaporkan bahwa etanol dapat digunakan untuk mengekstrak tanin, polifenol, poliasetilena, flavonol, terpenoid, sterol, alkaloid, and propolis. Di sisi lain, ekstrak kasar alkaloid daun pepaya pada konsentrasi 10 mg/ml memiliki aktivitas antibakterial yang lebih kuat dibandingkan ekstrak air dan metanol dari daun Chelidonium majus terhadap S. aureus. Ekstrak air dan metanol pada konsentrasi 100 mg/ml, secara berurutan, membentuk 12 ± 0.81 dan 10 ± 0.81 mm zona hambat (Cruz-Galvezet al.2013).
Nilai KHM ditentukan menggunakan metode pengenceran makro. Pada metode ini, ekstrak kasar alkaloid daun pepaya digunakan dalam berbagai konsentrasi, yaitu konsentrasi dengan seri pengenceran 2 kali. Konsentrasi yang diujikan adalah 1 mg/ml, 0.5 mg/ml, 0.25 mg/ml, dan 0.125 mg/ml. Penambahan ekstrak kasar alkaloid, pada konsentrasi 0.25 mg/ml, 0.5 mg/ml, dan 1 mg/ml
9.21
8.40
3.67
1.88
1.50
0 1 2 3 4 5 6 7 8 9 10
0 0.125 0.25 0.5 1
J
u
m
la
h
b
a
k
te
r
i
(lo
g
CF
U/m
l)
31
Aktivitas Ekstrak Kasar Alkaloid Daun Pepaya terhadap Jumlah Bakteri
S. aureusdan Ekspresi Gensea
Aktivitas Ekstrak Kasar Alkaloid terhadap Jumlah BakteriS. aureus
PemaparanS. aureusSJ1 dengan konsentrasi 0, 1, dan 2 KHM selama 2 jam menghasilkan jumlah sel bakteri yang lebih rendah dibandingkan dengan kontrol (Gambar 16). Pada pemaparan dengan 0 mg/ml ekstrak kasar alkaloid, jumlah bakteri mengalami kenaikan 1.09 log CFU/ml, sedangkan pada konsentrasi 0.25 mg/ml dan 0.5 mg/ml, jumlah bakteri hanya mengalami sedikit kenaikan, yaitu 0.42 log CFU/ml dan 0.21 log CFU/ml. Semakin tinggi konsentrasi ekstrak yang diberikan, semakin kecil kenaikan jumlah bakteri setelah 2 jam waktu inkubasi. Berdasarkan perbandingan dengan pertumbuhan kontrol (0 KHM), ekstrak kasar alkaloid konsentrasi 0.25 mg/ml dapat menghambat 61% pertumbuhan dan pada konsentrasi 0.5 mg/ml mampu menghambat 81% pertumbuhan. Konate et al. (2012) dan Luoet al.(2013) juga menemukan terjadinya penurunan jumlah selS. aureusyang diberi perlakuan dengan alkaloid.
Gambar 16 Pengaruh ekstrak kasar alkaloid daun pepaya pada 0, 1, dan 2 KHM terhadap pertumbuhan S. aureusSJ1. 0 jam, 2 jam.