i
ASPEK BIOLOGI DAN GENETIKA PADA SISTEM
REPRODUKSI MINDI (
Melia azedarach
Linn.)
DIDA SYAMSUWIDA
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
iii
PERNYATAAN MENGENAI DISERTASI DAN
SUMBER INFORMASI
Dengan ini saya menyatakan bahwa disertasi Aspek Biologi dan Genetik Pada Sistem Reproduksi Mindi (Melia azedarach L) adalah karya saya sendiri dengan arahan komisi pembimbing dan sebagian sudah diajukan dalam bentuk tulisan di jurnal ilmiah. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir disertasi.
Bogor, Juli 2012
v
ABSTRACT
DIDA SYAMSUWIDA. Biological and Genetical Aspects of
Reproductive System of Mindi (
Melia azedarach
Linn.).
Under academic supervision of ANDRY INDRAWAN, ISKANDAR Z SIREGAR andENDAH R PALUPI.
The study was aimed to determine biological and genetical aspects of reproductive system of mindi in relation to securing high quality of seeds. The study was conducted at four sites in West Java for the periods of 2009-2011. Direct observation over the trees (n=5-20 trees) were conducted on the periods of flowering and fruiting as well as floral morphology. Representative buds, flowers and fruits were sampled for microscopic dissection and reproductive study. The level of genetic diversities of maintained parental and produced offspring were evaluated either phenotipically or genotipically. Molecular characters were determined by using microsatellite markers. The genotipe deviation was evaluated based on Hardy-Weinberg and inbreeding structures. The observation revealed that floral initiation had proceeded for more than three months and the highest percentage of floral initiation occured in September. Reproductive cycle was proceeded for 5-6 months within the year, first observation commenced from generative buds to flower burst in September-October. Fruits reached maturity in January-February that took about 2-3 months after flower burst. The flower was hermaphroditic with position of anther was closed to stigma that selfing might be happened. Stigma receptivity lasted from 08.00 am to 11.00 am, rate of receptif was 66% in average, at 12.00 the receptivity was decreased to 40% and pollen viability was fluctuated from 4,89%-7,03% observed from 07.00 am-11.00 am. Controlled pollination exposed that mindi could be able to do both of cross- and self-pollination with the percentage of fruit set ranged of 70-77% and 42-57%, respectively. The genetic variation of mother trees and progenies for all loci and population were at range of 0,49-0,51 and 0,288-0,426, respectively. Genotipe deviation in progeny was detected at a small part of loci in two population with an average fixation index of -0,0680 and thus inbreeding might have been occured in a mating system of mindi in a low level. Assortative mating was detected in a very low degree. Three characters of seedling growth i.e length of bud, length of petiole and ratio of petiole to leaf length, might have to be used as keys to effective indicator for seedling selection. Key words: Genetic diversity, genotipe deviation, hermaprodititic, inbreeding,
vii
RINGKASAN
DIDA SYAMSUWIDA. Aspek Biologi dan Genetika Pada Sistem
Reproduksi Mindi (
Melia azedarach
Linn.) di bawah bimbingan
ANDRY INDRAWAN, ISKANDAR Z SIREGAR DAN ENDAH R PALUPI
Mindi (Melia azedarach Linn.) adalah salah satu jenis tanaman cepat tumbuh dari keluarga Meliaceae. Jenis ini memiliki kayu yang berkualitas baik dan sudah banyak dikembangkan pada hutan rakyat khususnya di Jawa-Madura. Penyediaan benih bermutu tinggi sangat diperlukan untuk mendukung pengembangan hutan rakyat yang dapat dimulai dengan melakukan pemilihan pohon plus sebagai sumber benih. Namun demikian, perlu pemahaman sistem reproduksi individu pohon sebagai bagian dari populasi sumber benih.
Sistem reproduksi tanaman mindi baik dari aspek biologi maupun genetika sejauh ini belum diketahui dan dipahami dengan jelas dan benar.Oleh karena itu penelitian bertujuan mengkaji aspek-aspek biologi dan genetik pada sistem reproduksi tanaman mindi dalam rangka penyediaan benih berkualitas tinggi untuk peningkatan produktivitas tegakan hutan rakyat. Secara khusus tujuan penelitian adalah 1) menganalisis siklus reproduksi termasuk inisiasi bunga, morfologi bunga-buah dan perkembangannya, masa resptivitas stigma dan viabilitas polen serta tipe penyerbukan 2) Menganalisis tingkat keragaman genetik populasi pohon induk dan anakannya serta menganalisis tingkat inbreeding berdasarkan kesetimbangan Hardy-Weinberg dan struktur in-breeding.
Penelitian dilaksanakan pada tahun 2009 s/d 2011 pada tegakan mindi di Jawa Barat yaitu di Desa Gambung-Bandung, Padasari-Sumedang, Wanayasa-Purwakarta dan Megamendung-Bogor. Siklus reproduksi diamati mulai dari inisiasi bunga, munculnya tunas bunga, bunga mekar hingga buah masak. Pengamatan inisiasi bunga dilakukan terhadap sampel tunas dengan sayatan secara mikroskopis. Pengujian masa reseptivitas stigma menggunakan aktivitas peroksidase dan pengujian viabilitas polen menggunakan media kecambah Brewbaker dalam inkubator suhu 25±1OC, kedua pengujian dilakukan setiap satu
jam mulai dari pk. 07.00-08.00 s/d pk.11.00-12.00.
viii
menggunakan struktur genotipe Hardy-Weinberg dan struktur in-breeding. Parameter genetik: Persentase polimorfik (PLP), jumlah alel obervasi (Na), alel efektif (Ne), heterosigotas observasi (Ho), heterosigotas ekspetasi (He) dan koefisien inbreeding, indeks fiksasi (Fis) dan perbedaan genetik antar populasi (Fst). Analisis data : ANOVA dilanjutkan dengan chi-square, AMOVA (Analysis of Molecular Variance). Pengolahan data menggunakan perangkat lunak PopGen versi 1.31 dan GenAlEx 6.3. Parameter fenotipik: ukuran morfologi benih, kapasitas perkecambahan dan morfologi semai. Analisis data menggunakan metode statistik peubah ganda dengan pendekatan PCA, PGM dan cluster analysis serta penghitungan nilai duga heritabilitas (Hbs) dan nilai duga kemajuan genetik (KG).
Hasil penelitian menunjukkan bahwa inisiasi bunga di Gambung-Bandung Selatan berlangsung lebih dari 3 bulan dimana persentase terbesar terjadi pada bulan September (70%) dan Oktober (60%). Siklus reproduksi berlangsung selama 5 – 6 bulan, mulai dari terjadinya tunas generatif (September), bunga kuncup (September), bunga mekar/anthesis (Oktober), buah muda (Okt-Nop) sampai buah masak panen (Januari-Februari). Mindi memiliki bentuk malai yang disebut panicle, satu malai terdiri atas 30-80 bunga dan mekar secara bergantian. Tipe bunga adalah hermaprodit. Perkembangan bunga terdiri atas lima fase dan fase ke lima ditandai dengan antera yang membuka dan polen sudah berhamburan keluar dan stigma yang berwarna hijau terletak di tengah antera yang memberikan indikasi periode penyerbukan. Stigma dan polen diambil dari bunga yang mekar pada hari pengamatan. Persentase reseptivitas stigma bervariasi mulai dari pengumpulan pukul 08.00 s/d 12.00 yaitu berkisar antara 40-73,33%. Pada pukul 12.00 reseptivitas stigma sudah menurun mencapai 40%. Viabilitas polen mindi berkisar antara 4,89-7,03%. Pengamatan viabilitas polen pada pukul 07.00 mencapai 7% yang tidak meningkat sampai pukul 11.00. Diduga periode penyerbukan terjadi pada pukul 08.00-11.00. Tanaman mindi memiliki kemampuan melakukan penyerbukan silang (outcrossing) atau penyerbukan sendiri (selfing) dengan kisaran nilai fruit set 70-77%.
ix
induk dan anak yang tersimpan di dalam populasi mencapai 67% dan 33% keragaman disumbang dari populasi lain. Terjadi penurunan kualitas genetik yang diturunkan oleh tetua terhadap keturunan. Penyimpangan genotipe dari struktur genetik Hardy-Weinberg dan struktur inbreeding terdeteksi pada sebagian lokus di kedua populasi. Secara keseluruhan nilai indeks fiksasi (Fis) rata-rata -0,0680. Hal ini mengkonfirmasi terjadi inbreeding pada sistem perkawinan tanaman mindi pada tingkat yang masih rendah. Populasi Sumedang memiliki tingkat penyimpangan genotipe yang lebih tinggi daripada populasi Bandung. Perbedaan genetik antar populasi mencapai 17%. Fenomena perkawinan berpilih (assortative mating) terdeteksi pada populasi Bandung pada lokus yang menyimpang dari struktur Hardy-Weinberg.
Karakter daya berkecambah, waktu mulai berkecambah, panjang pucuk semai, panjang petiole daun semai dan ratio panjang petiol terhadap panjang daun, dapat menjadi penciri dalam seleksi pohon induk dan pembibitan pada populasi Wanayasa-Purwakarta. Pohon induk P2, P15 dan P24 pada umumnya memperlihatkan keberagaman karakter fenotipe yang tinggi dibandingkan dengan pohon induk lain.
xi
@ Hak Cipta milik IPB, tahun 2012 Hak Cipta dilindungi Undang-undang
1. Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa
mencantumkan atau menyebutkan sumber
a. Pengutipan hanya untuk kepentingan pendidikan, penelitian,
penulisan karya ilmiah, penyusunan laporan, penulisan kritik atau
tinjauan suatu masalah
b. Pengutipan tidak merugikan kepentingan yang wajar IPB
2. Dilarang mengumumkan dan memperbanyak sebagian atau seluruh
xiii
ASPEK BIOLOGI DAN GENETIKA PADA SISTEM
REPRODUKSI MINDI (
Melia azedarach
Linn.)
Disertasi
sebagai salah satu syarat untuk memperoleh gelar Doktor pada
Program Studi Mayor Silvikultur Tropika
DIDA SYAMSUWIDA
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN
xiv
Penguji Luar :
Ujian tertutup : 1. Dr. Ir. Cahyo Wibowo M.Sc 2. Dr. Ir. Yulianti, M.Si.
xv
xvii
PRAKATA
Alhamdulillah puji syukur penulis panjatkan kehadirat Alloh SWT atas rakhmat dan karuniaNya yang diberikan kepada penulis untuk dapat menyelesaikan tugas akhir di Sekolah Pascasarjana, Institut Pertanian Bogor, Program Studi Mayor Silvikultur Tropika. Tugas akhir tersebut adalah tersusunnya disertasi yang berjudul ”Aspek Biologi dan Genetika pada Sistem Reproduksi Mindi (Melia azedarach Linn.)”.
Dengan tersusunnya disertasi ini, penulis menyampaikan terima kasih dan penghargaan kepada Prof.Dr.Ir. Andry Indrawan, MS selaku Ketua Komisi Pembimbing, serta Prof.Dr.Ir. Iskandar Z. Siregar, M.For.Sc. dan Dr. Ir. Endah R. Palupi MSc.selaku Anggota Komisi Pembimbing yang telah membimbing dan mengarahkan penulis mulai dari penyusunan proposal, selama penelitian dan analisis hingga selesainya penulisan disertasi.
Penyelesaian disertasi ini tidak terlepas juga dari bantuan semua pihak. Pada kesempatan ini penulis menyampaikan ucapan terima kasih kepada Kepala Badan Litbang Kehutanan, Kepala Balai Penelitian Teknologi Perbenihan Tanaman Hutan, yang telah memberikan ijin dan kesempatan untuk dapat mengikuti studi program S-3 di Sekolah Pascasarjana, IPB serta Direktorat Jendral Pendidikan Tinggi, Kementerian Pendidikan Nasional, yang telah memberikan dana Hibah Penelitian Tim Pascasarjana, tahun 2009-2010 (No kontrak: 78/13.24.4/SPK/BG/-PD/2009 dan 4/13.24.4/SPK/PD/2010), sehingga penelitian ini dapat berjalan. Terima kasih disampaikan pula kepada pengelola Program Studi Silvikultur Tropika, seluruh staf Fakultas Kehutanan dan adik-adik di Laboratorium Genetika-Silvikultur khususnya Laswi dan Azizah, yang telah membantu kelancaran penyelesaian studi.
Terima kasih yang tak terhingga disampaikan pula untuk suami tercinta (Aam P Permana), anak-anak terkasih (Ghina Amanda Putri dan M Faisal Adha) atas do’a dan keikhlasannya, serta seluruh keluarga dan teman-teman atas dorongan moril maupun materil.
Akhirnya, semoga hasil penelitian ini dapat bermanfaat bagi pengembangan ilmu pengetahuan khususnya bagi pengembangan program hutan tanaman di tanah air.
Bogor, Juli 2012
xix
RIWAYAT HIDUP
Penulis dilahirkan di Bandung 3 Juli 1957 sebagai anak terakhir dari enam bersaudara, dari ayah R.O Kosasih (Alm) dan ibu Sudewi Striretna (Almh). Penulis menikah dengan Aam Padma Permana dan dianugerahi satu orang putri dan satu orang putra : Ghina Amanda Putri dan Moch. Faisal Adha.
xxi
DAFTAR ISI
Halaman
DAFTAR ISI ……….. xxi
DAFTAR TABEL ………..……… xxiii
DAFTAR GAMBAR ……….. xxv
DAFTAR LAMPIRAN ……….……….. xxvii
I. PENDAHULUAN .……….. 1
1.1 Latar Belakang……… 1
1.2
Perumusan Masalah
………
41.3
Tujuan Penelitian
………..
61.4
Ruang Lingkup ...
61.5 Hipotesis……….. 7
1.6
Manfaat Penelitian
………...
71.7
Kerangka Pemikiran
……….
81.8
Kebaruan (Novelty)
……….
101.9
Sistematika Penelitian
………..
101.10 Risalah Lokasi ………. 12
II. TINJAUAN PUSTAKA ………. 15
2.1 Deskripsi Umum Mindi ……… 15
2.1.1 Sebaran dan Tempat Tumbuh Mindi ………. 15
2.1.2 Karakteristik Pohon, Taksonomi dan Pemanfaatan …... 15
2.1.2.1 Karakteristik Pohon ………. 15
2.1.2.2 Taksonomi ……… 17
2.1.2.3 Pemanfaatan tanaman ……….. 18
2.2 Biologi Reproduksi ……… 19
2.3 Keragaman Genetik dan Sistem Perkawinan ……… 23
2.3.1 Keragaman Genetik ……….. 23
2.3.2 Sistem Perkawinan ……….. 23
2.3.3 Penyimpangan Genotipe dalam Sistem Perkawinan (Inbreeding) ……… 26
2.4 Penanda Genetik Mikrosatelit ………. 29
2.5 Perbenihan Tanaman Mindi ………. 30
2.5.1 Sumber benih ………. 30
2.5.2 Produksi benih ………... 31
III. ASPEK BIOLOGI REPRODUKSI ……….. 33
3.1 Pendahuluan ………... 33
3.2 Bahan dan Metode ………. 34
3.3 Hasil dan Pembahasan ………. 39
3.3.1 Hasil ………. 39
3.3.2 Pembahasan ……….. 48
IV. ASPEK GENETIKA REPRODUKSI ……….. 53
4.1 Struktur Genetik Populasi Pohon Induk dan Keturunan ……….. 54
4.1.1 Bahan dan Metode ……… 54
4.1.2 Hasil dan Pembahasan ………... 56
4.2 Keragaman Fenotipik Keturunan ………. 70
xxii
4.2.2 Hasil dan Pembahasan ………... 76
V. PEMBAHASAN UMUM ………... 95
VI. SIMPULAN DAN SARAN ……… 103
6.1 Simpulan ……….. 103
6.2 Saran ……… 104
DAFTAR PUSTAKA ……….. 105
xxiii
DAFTAR TABEL
Halaman
1 Uraian tentang topik penelitian/sub-penelitian, kegiatan dan metode pada penelitian Aspek Biologi dan Genetik pada Sistem
Reproduksi Mindi (Melia azedarach L) ... 9 2 Rekapitulasi risalah lokasi penelitian dan kegiatan penelitian
yang dilakukan pada setiap lokasi ... 12 3 Periode dan waktu perkembangan pembungaan dan pembuahan
mindi... 42 4 Tahap perkembangan bunga mindi ... 44 5 Persentase reseptivitas stigma dan perkecambahan polen mindi
... 47 6 Delapan pasang sekuen primer mikrosatelit beserta urutan
basanya dan rentang ukuran yang digunakan, dikembangkan dari jenis Azadirachta indica (Meliaceae) (Bontoong et al. 2008)
………... 56 7 Rangkuman F-statistik untuk semua lokus ... 58 8 Rangkuman F-statistik untuk semua lokus pada dua populasi ... 59 9 Hasil analisis keragaman molekuler (AMOVA) ... 60 10 Keragaman genetik pada dua populasi pohon induk dan anakan
untuk semua lokus ... 61 11 Keragaman genetik lokus mikrosatelit pada sampel pohon induk
dan anakan mindi dari dua populasi ... 63 12 Hasil pengujian statistik struktur genetik dan dua acuan struktur
(Hardy- Weinberg dan struktur inbreeding) ...………… 64 13 Parameter alel preferensi Uij untuk lokus dengan genotipe yang
menyimpang dan tidak menyimpang dari struktur
Hardy-Weinberg……….. 66
14 Pengamatan karakter fenotipik keturunan mindi (diadopsi dari
Hidayat 2010) ……… 71
15 Varians fenotipe, varians genotipe, heritabilitas (H) dan kemajuan
genetik (KG) progeni mindi berdasarkan pertumbuhan semai. .... 77 16 Varians fenotipe, varians genotipe, heritabilitas (H) dan kemajuan
xxiv
morfologi semai mindi ... 80 18 Nilai eigenvalue dan persentase total variasi dari karakter
morfologi benih, perkecambahan dan morfologi semai mindi ..… 82 19 Matrik hasil klasifikasi dugaan kedekatan antar pohon induk
berdasarkan karakter morfologi benih mindi ... 85 20 Matrik hasil klasifikasi dugaan kedekatan antar pohon induk
berdasarkan kapasitas perkecambahan benih mindi ... 87 21 Matrik hasil klasifikasi dugaan kedekatan antar pohon induk
xxv
DAFTAR GAMBAR
Halaman
1 Alur kerangka pemikiran penelitian Aspek Biologi dan Genetika
pada Sistem Reproduksi Mindi (Melia azedarach ) ... 9 2 Peta lokasi penelitian Aspek Biologi dan Genetik pada Sistem
Reproduksi Mindi (Melia azedarach L) di Jawa Barat ... 13 3 Tegakan pohon dan bentuk daun, rangkaian bunga dan buah mindi
(insert) ... 16 4 Papan mebel kayu mindi bercorak indah ... 18 5 Irisan longitudinal tunas vegetatif memperlihatkan meristem apikal
(ma) mindi: tunas generatif memperlihatkan primordia bunga (pb), primordia daun (pd). A. tunas vegetatif : meristem apikal
membentuk primordial daun dan B.meristem apikal sudah
membentuk primordia bunga ... 39 6 Grafik terjadinya bakal bunga dan bakal daun pada irisan sampel
tunas. ... 40 7 Siklus reproduksi tanaman mindi di Bandung Selatan-Jawa Barat
... 42 8 Deskripsi organ bunga A] petal (p), B] pistil (ps) dan sepal (sp), C]
anther (a), stigma (s)………. 43 9 Deskripsi polen (P), lubang pori (lp), dinding luar (ex), dinding dalam
(in) dan polen yang berkecambah (KP) serta tabung polen (tp)…… 43 10 Perkembangan ovarium menjadi buah muda ... 45
11 Pembesaran ovarium [ovr], stilus [st] A) irisan melintang ovarium memperlihatkan 5 ovul [ovl] B) dan penampang melintang buah (bh)
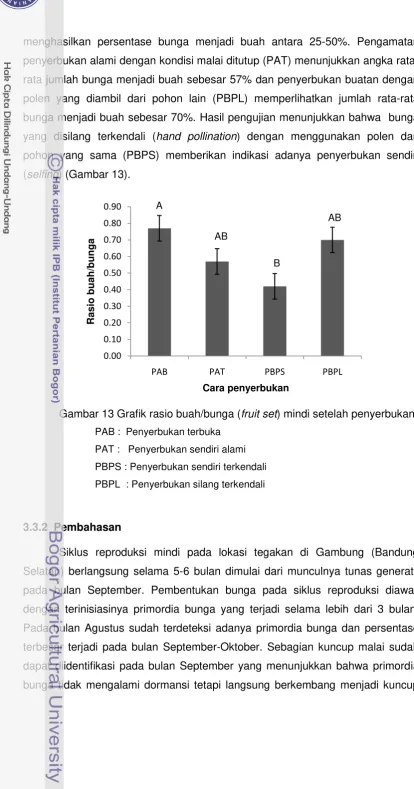
memperlihatkan lokus [lk], endokarp [end] dan biji [s] C) ... 46 12 Buah masak berwarna kuning (A) dan benih kering (B) ... 46 13 Grafik rasio buah/bunga (fruit set) mindi setelah penyerbukan ... 48 14 Pola polimorfik DNA mikrosatelit pada mindi dengan rentang
ukuran 100-220 bp ………. 57
15 Dimensi karakter semai mindi yang diukur ……… 73 16 Analisis komponen utama/PCA pohon induk mindi berdasarkan
xxvi
karakter perkecambahan ……… 83
18 Analisis komponen utama/PCA pohon induk mindi berdasarkan
karakter morfologi semai ……… 84
19 Dendrogram hasil clustering pohon induk mindi menggunakan
karakter morfologi benih... 90 20 Dendrogram hasil clustering pohon induk mindi menggunakan
karakter perkecambahan benih... 91 21 Dendrogram hasil clustering populasi pohon induk mindi
xxvii
DAFTAR LAMPIRAN
1 Tabel data skoring inisiasi pembungaan mindi di lokasi
Gambung-Bandung ... 115 2 Reseptivitas stigma dan Viabilitas Polen... 116 3 Penyerbukan mindi di Megamendung-Bogor... 119 4 Produksi buah mindi di lima lokasi di Jawa Barat ... 120 5 Data Pengukuran Karakter Fenotipik Pohon Induk Mindi... 121 6
7
Prosedur Laboratorium Analisis DNA dengan Penanda Mikrosatelit.. Analisis Data Keragaman Genetik...
1
I. PENDAHULUAN
1.1 Latar Belakang
Mindi (Melia azedarach Linn.) adalah salah satu jenis dari keluarga Meliaceae yang termasuk jenis tanaman cepat tumbuh. Jenis ini memiliki potensi besar untuk dikembangkan dalam pembangunan hutan rakyat karena bersifat serbaguna (multipurposes tree species). Bagian tanaman lain seperti kulit batang, daun, buah dan biji dapat dimanfaatkan sebagai obat sakit kepala, demam, antiseptik, pestisida atau obat HIV dan kanker karena kandungan antibiotiknya (Khan et al. 2008). Tanaman ini memiliki tiga komponen kimia yang bersifat komersil yaitu azadirachtin, selanin dan meliantriol (Hanum & van der Maesen 1997). Mindi juga merupakan salah satu kayu alternatif pengganti kayu berkualitas yang saat ini sudah mulai sulit ditemukan dan berharga mahal karena permintaan pasar yang semakin meningkat. Kayu dapat digunakan dalam bentuk kayu utuh untuk bahan bangunan, bahan furniture dan barang kerajinan, sebagai kayu lapis dan vinir laminar karena kayu bercorak indah dan mudah dikerjakan (Basri & Yuniarti 2006). Jenis ini juga menghasikan kualitas kayu yang dapat diandalkan untuk bahan pulp dan kayu bakar dengan nilai kalor 5100 kcal/kg (ICRAF database).
Tanaman mindi memiliki sifat mudah menyesuaikan diri dengan lingkungan tempat tumbuh karena dapat tumbuh pada zona ekologi yang cukup luas. Mindi diduga merupakan jenis eksotik yang berasal dari India (Asia Utara) dengan keragaman tinggi sehingga mudah beradaptasi dengan baik di wilayah Indonesia dan ditemukan tersebar di Pulau Jawa, Bali, Nusa Tenggara Barat dan Nusa Tenggara Timur. Jenis ini tumbuh pada ketinggian 200 – 1400 m dpl dengan curah hujan > 900 mm/thn, termasuk tipe iklim A-C (Martawijaya, et al. 1989, Soerianegara & Lemmens 1995).
Di Jawa Barat mindi tumbuh pada daerah dengan curah hujan rata-rata cukup tinggi yaitu > 2000 mm/th (Pramono et al. 2008). Memerlukan tanah dengan drainase yang baik, subur dan sedikit asam (pH 5,5 -6,5). Musim berbuah terjadi pada bulan Desember – Januari setiap tahunnya, walaupun kadang-kadang ada yang berbuah pada bulan Juni (Danu 2000).
(Pramono et al. 2008). Melihat kenyataan ini maka ada kecenderungan masyarakat menggunakan jenis mindi yang dapat beradaptasi luas dengan lingkungan untuk memenuhi kebutuhan papannya.
Di Kabupaten Bogor dilaporkan bahwa dari luasan hutan yang ada, 4% (13.320 ha) diantaranya adalah hutan rakyat dan jenis mindi mempunyai peluang yang besar untuk dikembangkan sebagai bahan pasokan kayu yang dipasarkan secara internasiona. Satu perusahaan eksportir furnitur memerlukan bahan baku kayu mindi sekitar 250 m³ sawn timber setiap bulan (Karyono & Hariyatno 2001). Dengan demikian, tantangan yang perlu diperhatikan adalah upaya untuk meningkatkan produktivitas tegakan mindi pada lahan masyarakat agar menghasilkan nilai ekonomi yang lebih tinggi.
Penyediaan benih bermutu tinggi sangat diperlukan untuk mendukung pengembangan hutan rakyat yang dapat dimulai dengan melakukan pemilihan pohon plus sebagai sumber benih. Namun demikian, perlu pemahaman sistem reproduksi individu pohon sebagai bagian dari populasi sumber benih.
Keragaman genetik tanaman mindi pada beberapa lokasi hutan rakyat di Jawa Barat tergolong rendah yaitu antara 0,16-0,19 dan 70% keragaman terdapat dalam populasi (Yulianti 2011). Produksi benih mindi pada lokasi-lokasi tersebut pada tahun 2009-2010 dilaporkan berkisar antara 5,1 – 9,9 kg/pohon (Atmandhini 2011). Produksi ini dinilai cukup rendah dibandingkan dengan produksi benih yang dilaporkan Nurhasybi & Danu (1997) yakni 15-20 kg/pohon dari beberapa lokasi di Jawa Barat. Keragaman genetik yang cukup rendah sudah diteliti pada beberapa tanaman. Pada pinus keragaman genetik yang rendah dikaitkan dengan sistem perkawinan (Siregar 2000), pada Veratum nigrum (Liliaceae) disebabkan oleh terbatasnya kelimpahan polen dan tidak efektifnya penyerbukan (Liao et al. 2006), dan pada tanaman Trillium camschatcense (Trilliaceae) kurangnya alokasi sumber pembungaan (Tomimatsu & Ohara 2006). Pola sebaran tanaman juga menjadi salah satu penyebab tinggi rendahnya keragaman genetik dalam populasi terutama pada hutan rakyat yang umumnya bercampur dengan tanaman lain dengan pola penanaman yang tersebar serta jumlah dan luasan yang terbatas (Hamid et al. 2008).
3
reproduksi tanaman untuk mendukung peningkatan kualitas benih tanaman mindi yang ditanam pada areal milik masyarakat.
Tanaman dengan variasi genetik yang tinggi dapat menghasilkan benih (keturunan) dengan berbagai sifat yang menguntungkan. Irwanto (2006) menyatakan bahwa metode pemuliaan (breeding system) yang tepat untuk suatu jenis tanaman tergantung kepada sistem penyerbukan, tingginya variabilitas, tujuan pemuliaan serta produksi benih.
Faegri and van de Pijl (1979), Sedgley (1986) dan Willian et al. (2001) juga mengemukakan bahwa untuk memperoleh strategi pemuliaan yang berhasil secara efisien, reproduksi suatu jenis perlu dipelajari melalui pengamatan yang spesifik terhadap fenologi pembungaan, karakteristik perkembangan bunga, arsitektur pembungaan, penyebaran polen dan vektornya, struktur organ seksual, sistem perkawinan, sistem inkompatibilitas dan implikasi genetik dengan adanya in-breeding serta hubungan paternalistik.
Sistem perkawinan acak biasa terjadi pada jenis tanaman hutan dan penyimpangan terhadap genotipe perkawinan acak banyak terbukti pada populasi alam. Fenomena ini dapat dijelaskan dengan membandingkan struktur genetik populasi pohon induk dan keturunannya (progeny) (Muller-Starck & Liu 1989). Penyimpangan genotipe yang terjadi umumnya disebabkan oleh adanya inbreeding yang merupakan akibat dari perkawinan kerabat (sendiri).
Sistem reproduksi tanaman mindi baik dari aspek biologi maupun genetika sejauh ini belum diketahui dan dipahami dengan jelas dan benar.Oleh karena itu dalam penelitian ini telah dilakukan pengamatan terhadap beberapa aspek reproduksi biologi meliputi pengamatan siklus reproduksi, morfologi bunga dan perkembangannya, reseptivitas stigma dan viabilitas polen dan aspek genetika pada sistem reproduksi yang meliputi keragaman genetik pohon induk dan turunannya serta penyimpangan struktur genetik pada sistem perkawinan mindi.
1.2 Perumusan Masalah
Studi tentang reproduksi tanaman mindi sejauh ini masih sangat terbatas. Hal ini menyulitkan pengembangan sumber benih mindi karena dalam pembangunan dan pengelolaannya diperlukan informasi dasar yang berkaitan dengan biologi reproduksi termasuk organ reproduksi, siklus reproduksi dan periode perkembangan pembungaan-pembuahan. Dari satu siklus reproduksi tanaman dalam satu periode pembungaan dan pembuahan dapat diprediksi waktu yang efektif melakukan penyerbukan dan kapan buah masak fisiologis dapat dipanen. Siklus reproduksi tanaman mindi belum diketahui secara detail mulai dari inisiasi bunga, tunas generatif, bunga mekar (anthesis) hingga terjadinya pembentukan buah dan buah masak. Kajian terhadap inisiasi bunga sangat diperlukan karena informasi yang diperoleh dapat digunakan untuk menduga waktu pembungaan. Waktu inisiasi bunga juga merupakan informasi yang diperlukan ketika akan mempercepat atau menstimulasi pembungaan.
5
dan viabilitas polen akan menentukan keberhasilan sistem penyerbukan. Informasi reseptivitas stigma dan viabilitas polen pada tanaman mindi belum diketahui. Oleh karena itu perlu dikaji lebih jauh tentang masa reseptif dan viabilitas polen. Waktu terjadinya stigma yang reseptif dan polen yang viabel juga merupakan informasi yang sangat bermanfaat ketika akan melakukan penyerbukan buatan yang efektif. Struktur organ reproduksi tanaman mindi adalah hermaprodit, dimana organ jantan dan betina berada dalam satu bunga yang sama, sehingga berpeluang terjadinya penyerbukan sendiri (selfing). Dengan demikian perlu dipelajari tipe penyerbukannya dalam rangka pembangunan sumber benih dan meningkatkan produksi benih.
Beberapa jenis tanaman hutan yang satu famili dengan mindi (Meliaceae) seperti mahoni (Swietenia macrophylla) dan mimba (Azadirachta indica) memiliki derajat penyerbukan silang yang tinggi yaitu masing-masing 96,6% (Lemes et al. 2007) dan 90,4% (Kundu 1999). Tanaman dengan pola penyerbukan silang akan memiliki keragaman yang luas sebagai akibat dari persilangan acak antar individu didalam populasi. Dengan demikian maka diduga keragaman pada keturunannya (progeni) akan mengikuti pola keragaman pohon induk. Akan tetapi pada jenis mindi informasi keragaman genetik pohon induk dan keturunan masih sangat terbatas, dengan demikian perlu mengevaluasi keragaman populasi pohon induk dan keturunannya melalui analisis terhadap variasi fenotipik dan variasi genetik.
Sistem reproduksi tanaman mindi dapat dianalisis secara genetik..
Penyimpangan genotipe pada sistem perkawinan sering terjadi pada populasi alam dan umumnya terjadi karena adanya inbreeding yang merupakan akibat dari perkawinan kerabat (sendiri). Oleh karena itu perlu analisis terhadap struktur inbreeding pada sistem reproduksi mindi untuk mengetahui kemungkinan adanya penyimpangan pada sistem perkawinan mindi.
Dari perumusan masalah di atas maka beberapa pertanyaan muncul yaitu sebagai berikut:
1. Kapan dan berapa lama terjadinya siklus reproduksi dan kapan inisiasi bunga mindi terjadi dalam satu periode waktu pada suatu populasi tegakan?
2. Kapan terjadinya stigma yang reseptif dan polen yang viabel? 3. Bagaimana tipe penyerbukan tanaman mindi?
dan keturunan pada jenis mindi dan bagaimana tingkat inbreeding yang terjadi berdasarkan kesetimbangan Hardy-Weinberg dan struktur inbreeding?
ontinuing interpretations is the lacot begenetic variatio
1.3 Tujuan Penelitian
Tujuan utama penelitian ini adalah mengkaji aspek-aspek biologi reproduksi dan genetik pada sistem reproduksi tanaman mindi dalam rangka penyediaan benih berkualitas tinggi.
Tujuan spesifiknya adalah:
1. Mengetahui aspek biologi reproduksi tanaman mindi yang meliputi siklus reproduksi, morfologi bunga dan tahap perkembangannya, masa reseptivitas stigma dan viabilitas polen serta tipe penyerbukan
2. Menganalisis tingkat keragaman genetik populasi pohon induk dan keturunan serta menganalisis tingkat inbreeding berdasarkan kesetimbangan Hardy-Weinberg dan struktur inbreeding
1.4 Ruang lingkup
Kegiatan penelitian meliputi dua aspek besar yang terkait biologi reproduksi dan genetika reproduksi. Aspek lingkungan bukan merupakan bagian dari penelitian studi reproduksi karena aspek ini sudah diteliti oleh Atmandhini (2011) terkait faktor tempat tumbuh dan produksi benih.
Penelitian aspek biologi dan genetika pada sistem reproduksi mindi meliputi:
1. Biologi reproduksi tanaman mindi Kegiatan penelitian termasuk: a. pengamatan siklus reproduksi
b. penentuan waktu inisiasi pembungaan c. pengamatan morfologi bunga
d. pengamatan reseptivitas stigma dan viabilitas polen e. penentuan tipe penyerbukan
7
a. Menganalisis keragaman genetik pohon induk dan keturunan (single tree progeny)
b. Menganalisis penyimpangan genetik pada perkawinan mindi menurut struktur Hardy-Weinberg dan struktur inbreeding
c. Menganalisis keragaman fenotipik keturunan
1.5 Hipotesis
Hipotesis yang akan diuji dalam penelitian ini adalah:
1. Reseptivitas stigma dan viabilitas polen terjadi pada rentang waktu yang bersamaan, sehingga penyerbukan dapat terjadi secara efektif.
2. Keragaman genetik populasi tanaman mindi yang rendah dengan tipe penyerbukan sendiri cenderung terindikasi adanya penyimpangan genetik pada sistem perkawinan.
1.6 Manfaat Penelitian
Hasil penelitian diharapkan memberikan manfaat dalam peningkatan produktivitas tegakan mindi melalui penyediaan benih bermutu dalam suatu manajemen sumber benih. Informasi yang diperoleh dari penelitian ini diantaranya yaitu:
1. Pemahaman tentang biologi reproduksi tanaman yang sangat bermanfaat dalam pengelolaan suatu tegakan atau sumber benih berkaitan dengan manajemen penyerbukan, waktu inisiasi bunga dan pembuahan sehingga masa panen buah mindi dapat di prediksi.
2. Mengetahui tingkat keragaman genetik populasi pohon induk dan anakan mindi pada lokasi yang berbeda sehingga benih atau bibit yang dikumpulkan dari lokasi tersebut dapat diterima tingkat kualitasnya secara genetik sebagai bahan pertimbangan dalam perencanaan pengembangan kebun benih rakyat. 3. Mengetahui tingkat struktur inbreeding tanaman mindi dalam suatu populasi
1.7 Kerangka Pemikiran
Hutan berbasis masyarakat terutama di pulau Jawa saat ini berkembang cukup pesat (Departemen Kehutanan 2003) dan salah satu jenis potensial yang dapat dikembangkan pada hutan yang dikelola masyarakat atau hutan rakyat adalah mindi. Dalam pengembangannya perlu dukungan dari berbagai sektor yang terkait termasuk diantaranya adalah ilmu pengetahuan dan teknologi (IPTEK).
Produktivitas tegakan hutan sangat kuat dipengaruhi oleh faktor lingkungan dan faktor genetik serta kombinasi keduanya (Zobel & Talbert 1984). Salah satu upaya untuk menghasilkan produktivitas tegakan hutan yang tinggi terutama pada luasan yang terbatas adalah melalui penggunaan dan penyediaan benih berkualitas tinggi secara berkelanjutan. Benih berkualitas tinggi secara genetis dapat diperoleh dari sumber benih terkatagori (kelas sumber benih) apabila sudah tersedia, selain itu benih berkualitas secara fisik dan fisiologik diperoleh melalui penanganan benih yang optimal.
IPTEK perbenihan sangat diperlukan untuk mendukung peningkatan kualitas benih dan produksi yang memadai dalam rangka penyediaan bahan pertanaman. Dengan demikian, sangat penting untuk memahami dan menguasi sistem reproduksi pohon induk sebagai bagian dari populasi sumber benih.
Sistem reproduksi tanaman mindi sejauh ini belum diketahui dan dipahami dengan jelas dan benar. Sistem reproduksi meliputi studi tentang aspek biologi reproduksi dan aspek genetika pada sistem reproduksi. Informasi ini sangat bermanfaat dalam pengelolaan suatu tegakan benih atau pembangunan sumber benih berkaitan dengan perolehan benih berkualitas.
Pengamatan biologi reproduksi yang meliputi siklus reproduksi, morfologi bunga dan tahap perkembangannya, masa reseptifitas stigma dan viabilitas polen serta tipe penyerbukan menghasilkan informasi yang sangat bermanfaat terkait prediksi masa panen buah dan efektivitas penyerbukan. Hasil kajia terhadap karakteristik reproduksi secara biologis akan saling mendukung dengan hasil analisis reproduksi secara genetis, sehingga dapat dievaluasi untuk dijadikan bahan pertimbangan perencanaan pembangunan atau pengelolaan sumber benih.
9
diduga melalui pendekatan marka genetik dan marka morfologi. Penyimpangan genotipe yang mungkin terjadi akibat perkawinan kerabat (selfing) pada tanaman mindi yang mempunyai tipe bunga hermaprodit, dapat diketahui dengan menganalisis tingkat inbreeding berdasarkan kesetimbangan struktur Hardy-Weinberg dan struktur inbreeding.
Seluruh informasi yang didapatkan dari hasil analisis genetika dan biologi terkait sistem reproduksi tersebut tentu akan sangat bermanfaat dalam mengevaluasi pengelolaan sumber benih secara efisien dengan memperhatikan aspek reproduksi yang efektif untuk menghasilkan benih yang berkualitas tinggi secara berkelanjutan. Selain itu informasi yang diperoleh dapat menjawab permasalahan yang berkaitan dengan rendahnya keragaman genetik populasi pohon mindi dan produksi benih pada hutan rakyat di Jawa Barat.
[image:37.595.106.512.260.741.2]Kerangka pemikiran untuk penelitian ini disajikan alurnya pada Gambar 1 .
1.8 Kebaruan (Novelty)
Kebaruan informasi yang dapat disampaikan dari hasil penelitian ini adalah:
1. Diperolehnya informasi mengenai sistem biologi reproduksi tanaman mindi berdasarkan pengamatan siklus reproduksi yang intensif mulai dari inisiasi bunga hingga ke pembentukan bunga dan buah serta diketahuinya tipe penyerbukan mindi.
2. Diketahuinya tingkat keragaman genetik tanaman mindi berdasarkan marka molekuler mikrosatelit.
3. Diketahuinya tingkat inbreeding populasi tegakan mindi berdasarkan evaluasi terhadap penyimpangan struktur genetik yang terjadi dan selanjutnya dapat dijadikan dasar pertimbangan dalam penggunaan tegakan sebagai sumber benih.
[image:38.595.59.489.60.777.2]1.9 Sistematika Penelitian
Tabel 1 Uraian tentang topik penelitian/sub-penelitian, kegiatan dan metode pada penelitian Aspek Biologi dan Genetika pada Sistem Reproduksi Mindi (Melia azedarach L)
Topik penelitian/
Sub penelitian Kegiatan Penelitian Metode Penelitian
1. Aspek Biologi Reproduksi:
Siklus reproduksi dan
inisiasi bunga Mengamati siklus reproduksi dan periode
perkembangan pembungaan-pembuahan
Pengamatan langsung di plot penelitian di
Gambung-Bandung Selatan
Mengambil tunas, menyayat jaringan dan mengamati primordia bunga di bawah mikroskop.
Pengamatan mikroskopis jaringan tunas
Morfologi bunga dan tahap
perkembangannya
Mengamati, mengukur dan membuat foto/ sketsa struktur organ reproduksi. Mengamati dan membuat foto tahap perkembangan bunga
Pengamatan langsung di plot penelitian di
11
Lanjutan Tabel 1
Topik penelitian/
Sub penelitian Kegiatan Penelitian Metode Reseptivitas stigma
dan viabilitas polen Mengamati stigma reseptif berdasarkan kriteria tahap
pemekaran bunga dengan interval waktu pengambilan sampel bunga setiap 1 jam selama 5 jam
Pengamatan reaksi permukaan stigma terhadap H2O2
(Kearns & Inouye 1993) Mengamati polen viabel berdasarkan kriteria tahap pemekaran bunga dengan interval waktu pengambilan sampel bunga setiap 1 jam selama 5 jam
Pengamatan perkecambahan polen pada kultur media Brewbaker (Brewbaker&Kwack 1963)
Tipe penyerbukan Melakukan
penyerbukan buatan dan alami
Penyerbukan sendiri dan silang terkendali,
penyerbukan sendiri secara alami dan penyerbukan terbuka (Palupi 2006)
Penelitian
2. Aspek Genetika: Struktur genetik pohon induk dan keturunan
Menganalisis keragaman genetik pohon induk dan keturunannya (single tree progeny) dengan pendekatan marka DNA dan menentukan tingkat struktur inbreeding berdasarkan kesetimbangan Hardy-Weinberg Dugaan tingkat keragaman genetik menggunakan penanda mikrosatelit dan menghitung frekuensi inbreeding observasi dan harapan (Muller-Starck&Liu 1989) Keragaman fenotipik
1.10 Risalah Lokasi Penelitian
[image:40.595.69.494.268.817.2]Penelitian merupakan suatu rangkaian kegiatan penelitian yang dimulai pada tahun 2009 dan diakhiri tahun 2011. Kegiatan penelitian dilaksanakan di Jawa Barat yaitu pada tegakan mindi yang ada di kebun campuran baik milik masyarakat maupun areal perkebunan BUMS. Pemilihan lokasi berdasarkan hasil survey penyebaran tanaman mindi di Jawa Barat yang dilakukan oleh Pramono et al. (2008) yakni daerah penyebaran mindi yang mewakili tipe tapak dengan ketinggian yang berbeda.
Tabel 2 Rekapitulasi risalah lokasi penelitian dan kegiatan penelitian yang dilakukan pada setiap lokasi
No Lokasi Kondisi umum Kegiatan
penelitian 1 Ds. Tegalmindi, Kec.
Megamendung, Kab. Bogor
Letak pada 06O40’ 477” S-106O53’635’’E. Jenis tanah podsolik coklat, topografi bergelombang. Curah hujan rata-rata 2200-4500 mm/th, ketinggian 710 m dpl. Suhu rata-rata berkisar antara 25 0C - 29 0C dengan kelembaban 70 % - 75 %. Merupakan kebun campuran dengan beberapa jenis tanaman pangan, sayuran dan buah-buahan. Tanaman mindi ditanam pada bagian pematang sebagai tanaman peneduh.
Percobaan tipe
penyerbukan
Ds.Nagrak, Kec.
Sukaraja, Kab. Bogor Letak pada 06
O40’ 472” S-106O53’615’’E. Podsolik merah kunig, topografi sedikit bergelombang. Curah hujan rata-rata 2200-4500 mm/th, ketinggian 250-350 m dpl. Suhu rata-rata berkisar antara 26 0C - 27 0C dengan kelembaban 70 %.
Percobaan reseptivitas stigma dan viabilitas polen
2 Kab. Purwakarta, Kec.Wanayasa, Ds. Legokhuni
Letak pada 06O39’ 378” S-106O32’479’’E. Jenis tanah podsolik merah kuning, topografi sedikit bergelombang dengan kelerengan 5 O– 10 O danketinggian 620 meter dpl. Suhu rata-rata 26 – 28 0C dengan kelembaban 70-75 %, pencahayaan 15.000 – 62.500 flux.Lahan merupakan kebun campuran yang didominasi oleh tanaman teh serta beberapa tanaman keras seperti kopi, pisang, sengon, aren, manggis, cengkeh, pala, suren, kayu afrika, duren, rambutan, kelapa, nangka dan limus. Tanaman mindi merupakan tanaman peneduh yang cukup dominan, ditanam pada tahun 2001.
13
Lanjutan Tabel 2
No Lokasi Kondisi umum Kegiatan
penelitian Pemilik memanfaatkan kayu mindi
sebagai bahan pembuatan mebel terutama lemari dan kursi. Kayu mindi dinilai lebih tinggi harganya dibandingkan dengan kayu sengon/jeungjing.
3 Ds. Padasari, Kec.Cimalaka, Kab.Sumedang
Letak pada 06 O 47” S-107O56’E. Jenis tanah podsolik merah kuning, topografi bergelombang dengan ketinggian 600-700 meter dpl. Suhu berkisar antara 26-30 OC dan kelembaban 80-85%. Plot penelitian berada di areal kebun milik masyarakat, dimana mindi tumbuh bercampur dengan jenis tanaman lain seperti buah-buahan, tanaman pangan dan jenis hutan lainnya seperti suren, sengon, kayu afrika dlsb.
Struktur genetik pohon induk dan keturunan
4 Ds. Gambung, Kec.Ciwidey, Kab. Bandung
Letak pada 07 O14’ S-107O56’E . Jenis andosol, topografi bergelombang dengan ketinggian 1340 m dpl, curah hujan rata-rata 1200-1600 mm/th 700, suhu berkisar 15 O C – 28 O C dengan kelembaban 40 % - 50 %.Tegakan mindi berada pada areal perkebunan teh sebagai pohon pelindung.
[image:41.595.100.518.102.728.2]1.Pengamatan siklus reproduksi, inisiasi bunga dan morfologi bunga 2.Struktur genetik pohon induk dan keturunan
Gambar 2 Peta lokasi penelitian Aspek Biologi dan Genetika pada Sistem Reproduksi Mindi (Melia azedarach L) di Jawa Barat KAB. BOGOR
KAB. BANDUNG
1
4 3
KAB. SUMEDANG
II. TINJAUAN PUSTAKA
2.1 Deskripsi Umum Mindi
2.1.1 Sebaran dan Tempat Tumbuh Mindi
Mindi merupakan jenis eksotik yang tumbuh baik dan tersebar luas di Indonesia. Di tempat asalnya yaitu Asia Utara (India dan Pakistan) dapat tumbuh hingga ketinggian 2.200 m dpl di kaki pegunungan Himalaya, menyebar hingga ke Solomon, Malaysia dan Indonesia. Kemudian menyebar antar benua yaitu Amerika (Amerika Serikat bagian selatan dan Hawai), Amerika Latin ( Argentina), Eropa (Croatia dan Perancis), China dan Australia (bagian timur dan selatan) (Hanum & van der Maesen 1997).
Jenis ini dapat beradaptasi pada perbedaan kondisi yang luas mulai dari dataran rendah (1 – 1.200 m dpl) di Papua Nugini hingga ke dataran tinggi Himalaya (1.200-2.200 m dpl) dengan kisaran suhu dari – 5 0C hingga 39 0C dan
curah hujan 600 – 2.000 mm/th (Hanum & van der Maesen 1997). Di Afrika ditanam sebagai tanaman peneduh yang toleran kekeringan dan di Amerika bagian selatan dan barat daya tumbuh liar pada daerah kering dengan curah hujan dibawah 600 mm/tahun.
Menurut Heyne (1987) di Jawa mindi tidak tumbuh liar, tapi awalnya ditanam untuk pohon peneduh pada perkebunan kopi pada zaman pendudukan Belanda. Namun di perkebunan teh dan kina Gambung (Bandung Selatan) juga ditemukan mindi sebagai pohon peneduh tanaman teh yang berasal dari India yang ditanam dengan jarak 2,5 m x 2,5 m hingga umur > 10 tahun, kemudian dijarangi dan kayu hasil penjarangan digunakan untuk bahan bakar pada proses pengeringan di pabrik pengolahan teh. Selain itu penanaman mindi juga ditujukan untuk mengurangi serangan hama pada tanaman teh, karena daun dan biji mindi dapat digunakan sebagai biopestisida (Erwan Johan1) 2009 ,
komunikasi pribadi).
2.1.2 Karakteristik Pohon, Taksonomi dan Pemanfaatan
2.1.2.1 Karakteristik Pohon
beralur, halus atau berbintil. Bentuk tajuk relatif simetris dengan percabangan melebar, berdaun ringan, tipe daun majemuk, bentuk anak daun bulat lonjong dengan bagian tepi bergerigi atau kadang halus (Gambar 3 insert). Umumnya menggugurkan daun pada musim kering (decidoeus), bertunas setelah masa rontok daun yang diikuti dengan pembungaan. Berakar tunggang yang dalam dengan akar cabang yang banyak (Heyne 1987).
Dokumentasi: Dida Syamsuwida
Gambar 3 Tegakan pohon dan bentuk daun, rangkaian bunga dan buah mindi (insert)
[image:44.595.45.470.52.643.2]17
Sifat kayu:
Secara fisik kayu mindi tergolong kelas kuat III dengan berat jenis 0,53 (0,42-0,65) dan keawetan terhadap jamur pelapuk termasuk kelas II-III (Basri & Yuniarti 2006), kekuatan kayu mindi setara dengan mahoni, sungkai, meranti merah. Kayu teras pada pohon mindi berwarna merah coklat muda keunguan dengan kayu gubal berwarna putih kemerahan dan mempunyai batas yang jelas dengan kayu teras. Serat lurus atau agak berpadu, berat jenis rata-rata 0,53 g/cm3, kerapatan 510-660 kg/m3. Besarnya penyusutan dari keadaan basah
sampai kering tanur adalah 3,3% (radial) dan 4,1% (tangensial). Pengeringan alami, pada papan tebal 2,5 cm dari kadar air 37% sampai 15% memerlukan waktu 47 hari, dengan kecenderungan pecah ujung dan melengkung. Pengeringan dalam dapur pengering dengan bagan pengeringan yang dianjurkan adalah suhu 60-80% dengan kelembaban nisbi 80-40% (Martawidjaja et al. 1989).
2.1.2.2 Taksonomi
Klasifikasi tanaman mindi menurut sistematika (taksonomi) tumbuhan adalah sebagai berikut (Stanley & Ross 1983):
famili : Meliaceae genus : Melia
species : azedarach Linn. Sinonim: Melia australasica A. Juss.
Melia composita Willd. Melia dubia Cavanilles Melia japonica Don. Melia bukayan Royle. Melia sempervirens (L.) Sw.
2.1.2.3 Pemanfaatan Tanaman
Tanaman mindi dalam bentuk tegakan dapat dimanfaatkan sebagai tanaman ornamental, peneduh, penahan angin dan rehabilitasi hutan dan lahan. Sifatnya yang cepat tumbuh dan mampu tumbuh pada kondisi yang tidak optimal sangat cocok untuk penghijauan dan reklamasi lahan marjinal di area semi arid pada daerah tropis dan subtropis.
Kayunya dapat diolah menjadi kayu papan, kayu bakar, pembuatan mebel, pertukangan, veneer, kayu lapis dan bubur kertas (pulp) (Florido et al. 2002). Pemanfaatan mindi untuk kayu bakar sangat baik karena dapat menghasilkan nilai kalor sebesar 5100 kcl/kg. Kayu mindi terbukti baik sebagai bahan baku mebel untuk ekspor dan dikembangkan secara domestik. Sifat kayu mindi yang mudah dikerjakan, dapat mengering tanpa cacat, sangat sesuai untuk pembuatan mebel (Gambar 4). Selain itu kayunya mengkilap dan menghasilkan venir bercorak indah seperti kayu mewah yang terdapat pada kayu sonokeling atau kayu kuku (Martawidjaia et al. 1989).
Dokumentasi: Dida Syamsuwida
Gambar 4 Papan mebel kayu mindi bercorak indah
Ekstrak daun mindi mengandung insektisida (azadirachtin) yang dapat digunakan sebagai bahan untuk mengendalikan hama pada pakaian dan belalang. Kulit mindi dipakai sebagai penghasil obat untuk mengeluarkan cacing usus. Kulit daun dan akar mindi telah digunakan sebagai obat reumatik, demam, bengkak dan radang. Selain itu, suatu glycopeptide yang disebut meliacin diisolasi dari daun dan akar mindi yang berperan dalam menghambat perkembangan beberapa DNA dan RNA dari beberapa virus misalnya virus polio (Khan et al. 2008).
19
Minyak biji adalah merupakan produk penghasil bahan obat paling aktif pada tanaman mindi yang dapat digunakan sebagai antiseptik untuk radang. Juga digunakan untuk rematik dan penyakit kulit serta pengobatan bagian dalam seperti demam malaria dan leprosi (Florido et al. 2002, Khan et al. 2008).
2.2 Biologi Reproduksi
Seperti halnya terjadi pada semua makhluk hidup tingkat tinggi, umumnya tanaman secara alami memperbanyak diri melalui alat reproduksi yang terdiri dari organ jantan dan betina. Perkawinan antara organ jantan dan betina menghasilkan individu baru yang memiliki sifat gabungan antara kedua organ tersebut.
Tanaman memproduksi biji karena secara alami tanaman memiliki kemampuan untuk mempertahankan jenis agar keberadaannya tetap lestari. Selain itu juga untuk menghasilkan individu baru yang beragam agar memiliki berbagai sifat yang menguntungkan. Biji sendiri adalah merupakan ovul yang sudah masak setelah dibuahi yang mengandung embrio, nutrisi tersimpan, integumen dan testa (Esau 1976).
Proses reproduksi dimulai dari inisiasi atau induksi pembungaan. Inisiasi biasanya dibedakan dengan induksi. Inisiasi adalah suatu transisi dari meristem vegetatif (memproduksi primordia daun) menjadi apikal reproduktif (primordia bunga) yang akan berkembang menjadi bunga (Owens & Blake 1985). Perubahan ini dilihat secara anatomi dengan pemeriksaan mikroskopis, sedangkan induksi, perubahannya dilihat dari kandungan biokimia seperti asam nukleus, total protein, sintesis RNA, formasi ribosom dan indeks mitosis (Esau 1976, Sedgley & Griffin 1989). Perubahan terjadi beberapa hari, minggu atau bulan sebelum munculnya primordia (bakal) bunga. Inisiasi pembungaan terjadi setelah tanaman melewati fase juvenilitas. Faktor-faktor lingkungan yang mempengaruhi terjadinya proses inisiasi pembungaan diantaranya : suhu, intensitas cahaya, panjang hari, kelembaban, mineral dan hara serta faktor cekaman (stress air, pelukaan, pencekikan dll) (Sedgley & Griffin 1989).
Siklus reproduksi pada tanaman sayuran misalnya berbeda dengan tanaman keras, secara genetis tanaman sayuran berumur lebih pendek daripada tanaman keras, sehingga siklusnya berlangsung lebih cepat.
Tipe organ reproduksi pada individu tanaman bervariasi seperti hermaprodit (♂ + ♀ dalam satu bunga), monoecious (♂ + ♀ dalam satu tanaman), androecious (hanya menghasilkan bunga ♂), gynoecious (hanya menghasilkan bunga ♀) serta kombinasi antara karakteristik diatas (Schmidt 2000). Tanaman mindi termasuk tipe hermaprodit dan monoecious, sementara jenis Meliaceae lainnya seperti suren (Toona chinensis) memiliki tipe hermaprodit dan androecious (Hidayat 2008).
Struktur organ reproduksi suatu jenis sangat kuat dipengaruhi oleh faktor genetika, namun demikian faktor lingkungan dapat merubah struktur reproduksi terutama dalam ukuran dimensinya melalui proses evolusi. Keragaman yang luas dalam hal ukuran dan morfologi bunga serta pola pemanjangan stamen dan stylus selama bunga hidup (anthesis) ditemukan pada jenis Collinsia dan Tonella yang merupakan jenis tanaman tahunan yang self-compatible (Armbruster et al. 2007).
Memahami struktur dan sistem organ reproduksi tanaman hutan sangat penting dalam kaitannya dengan sistem perkawinan yang terjadi, sehingga manajemen penyerbukan dapat dilakukan secara tepat dengan memperhatikan bentuk, ukuran, warna dan aroma bunga yang dapat menarik vektor penyerbuk. Selain memahami struktur bunga dan sebagai bagian dari sistem organ reproduksi, informasi tentang kematangan polen dan reseptivitas stigma sangat penting dalam rangka mempelajari biologi reproduksi. Penyerbukan akan berhasil apabila terjadi sinkronisasi antara polen viabel dengan stigma yang reseptif.
21
Butiran polen tersusun atas empat komponen mendasar: exine atau lapisan dinding terluar, intine atau lapisan dinding dalam (keduanya mengandung protein), pollenkit atau mantel (memberi warna pollen) dan colpi atau lubang germinasi (mengandung lemak) (Esau 1976).
Secara visual, polen yang matang dapat dideteksi dari perubahan warna dan kelekatan (stickiness) butiran-butirannya (Sedgley & Griffin 1989, Ghazoul 1997). Perubahan warna permukaan butiran polen dari kuning pucat menjadi kuning terang mengindikasikan adanya peningkatan sporopollenin – bagian dari exine yang merupakan ciri spesifik dari suatu spesies yang mempengaruhi kenampakan luarnya dan pollenkit yang basah, lengket dan berwarna, mengandung lemak, protein, karbohidrat, pigmen, senyawa fenolik dan enzim.
Peningkatan kelekatan butiran polen mengindikasikan bahwa polen tersebut telah siap untuk berkecambah dengan melakukan proses hidrasi dan melepaskan protein. Mekanisme hidrasi inilah yang dianggap paling menentukan dalam mengawali terjadinya proses penyerbukan, yang merupakan rangkaian dari proses interaksi jantan-betina, perkecambahan polen dan pembentukan tabung polen (Griffin & Sedgley 1989).
Struktur polen beradaptasi dengan cara penyerbukan, apabila bentuk polen lonjong dengan ukuran < 300 µm atau agregat, dinding polen tebal, permukaan berminyak atau lengket maka penyerbukan biasanya dibantu oleh hewan. Sedangkan bentuk polen yang lonjong dengan ukuran> 50 µm, dinding polen tipis dan permukaan tipis tapi mudah menempel, penyerbukan dibantu oleh angin (Faegri and van der Pijl 1979).
Penyebaran polen dan biji tanaman sangat berpengaruh terhadap pergerakan gen yang dibawanya. Polen dan biji tanaman semak sering berpencar dekat dengan pohon induk (Levin 1989). Implikasi dari penyebaran polen yang terbatas menyebabkan hubungan antara individu tanaman menurun, akibatnya penurunan fertilitas sering terjadi pada tanaman yang melakukan penyerbukan dengan pohon tetangga terdekat dibandingkan dengan pohon yang melakukan penyerbukan dengan pohon dengan jarak yang lebih jauh (Soute et al. 2002).
individu, biologi penyerbukan suatu populasi dan sistem persilangan jenis (Wyatt 1983, Kalisz et al. 1999, Heslop-Harrison 2000, Sanzol 2003). Reseptivitas stigma didefinisikan sebagai kemampuan stigma untuk mendukung perkecambahan polen yang mana merupakan tahap yang sangat penting dalam keberhasilan penyerbukan dan sangat bervariasi antar spesies (Heslop-Harrison 2000).
Secara umum masa reseptif stigma biasanya ditandai dengan : perubahan warna putik menjadi lebih terang, pembesaran pori-pori pada kepala putik, tangkai putik berangsur menjadi lurus , permukaan putik memproduksi sekresi. Secara visual, reseptivitas stigma dapat dideteksi dari perubahan kelekatan (stickiness), warna dan bentuk, baik pada kepala maupun tangkai putik (Sedgley &Griffin 1989, Owens et al. 1991).
Pada saat reseptif stigma mengeluarkan sekresi yang berperan sebagai medium yang berfungsi untuk menangkap butiran polen, serta merupakan penentu keberhasilan pembentukan tabung polen yang akan membawa sel gamet jantan menuju ke ovarium (Sedgley & Griffin, 1989). Menurut Owens et al. (1991), sekresi ekstraseluler tersebut mengandung lemak dan protein yang meningkat pada saat reseptif (Ghazoul 1997).
Perubahan warna permukaan putik dari hijau menjadi kuning terang, yang dimulai dari pangkal tangkai putik (stylus) juga menandai reseptifnya stigma. Makin terangnya warna stigma menunjukkan bahwa sel-sel epidermis terluar sedang berkembang untuk meningkatkan produksi sekresi dan pori-pori membesar untuk meningkatkan kemampuan sekresi (Jamsari 2006).
Lamanya reseptivitas stigma berlainan untuk setiap jenis, biasanya penyerbukan yang dilakukan oleh bantuan angin lebih lama daripada jenis yang dibantu oleh serangga (Khadari et al. 1995). Reseptivitas dapat berlangsung selama kurang dari satu jam (jenis Avena) atau lebih dari satu minggu seperti pada Eucalyptus (Heslop-Harrison 2000).
Fase reproduksi khususnya dari penyerbukan ke pembuahan sangat rentan terhadap kondisi lingkungan saat itu termasuk suhu (Stephenson et al. 1992). Suhu lingkungan dapat mempengaruhi reseptivitas stigma dimana kapasitas stigma dalam mendukung perkecambahan polen akan berkurang dengan semakin tingginya suhu (30 OC). Suhu optimal untuk reseptivitas stigma
23
ini merupakan jenis temperate dimana lingkungan habitatnya bersuhu rendah (Hedhly et al. 2003).
2.3 Keragaman Genetik dan Sistem Perkawinan
2.3.1 Keragaman Genetik
Penilaian keragaman genetik di dalam suatu populasi tegakan merupakan langkah pertama untuk mengevaluasi keragaman genetik saat ini dan yang akan datang (Moran et al 1980). Penilaian untuk menduga (nilai duga) keragaman dari suatu populasi dapat berdasarkan variasi fenotipik dan variasi genetik. Penilaian ini dapat menduga tingkat keragaman genetik pada populasi tegakan alam ataupun populasi hasil penanaman. Keragaman genetik pada populasi hasil pertanaman tidak menunjukkan penurunan keragaman genetik yang signifikan dibandingkan dengan keragaman pada populasi tegakan alam (El-Kassaby & Ritland 1996). Hal ini mengindikasikan bahwa seleksi awal dan perlakuan silang (breeding) tidak menurunkan variasi genetik. Walaupun demikian, struktur genetik pohon induk sangat kuat pengaruhnya terhadap keragaman benih (keturunannya) (Nurtjahyaningsih 2008). Karena itu untuk mempertahankan tingkat keragaman genetik yang sama antara populasi pohon induk dengan anakannya, maka derajat perkawinan silang (outcrossing rate) (Moran et al. 1980) dan perkawinan acak yang diimbangi dengan kontribusi semua pohon induk dalam mewariskan gennya (Panmictic Equilibrium) harus tinggi (Chaix et al. 2003) serta harus sinkron dengan pembungaan pohon tetangga disekitar tegakan pohon induk (Gomory et al. 2003).
Tingkat dan pola keragaman genetik pohon hutan sangat kuat dipengaruhi oleh sistem perkawinannya serta pergerakan gen diantara populasi dari jenis yang sama (penyebaran gen/gene flow). Sistem perkawinan pada pohon hutan bervariasi dan ada rentang dari mekanisme untuk melakukan penyerbukan silang (outcrossing) ke mekanisme penyerbukan sendiri (self pollination) (Finkeldey 2005).
2.3.2 Sistem Perkawinan
Oleh karena itu, sistem perkawinan sangat penting dalam membentuk struktur genotipik dari generasi selanjutnya.
Menurut Finkeldey (2005) sistem perkawinan populasi tanaman melibatkan beberapa aspek penting yaitu diantaranya penyerbukan sendiri (selfing), penyerbukan silang (outcrossing), kawin acak (random mating), dan kawin berpilih (assortative mating). Tipe sistem perkawinan pada suatu jenis tanaman memiliki pengaruh yang kuat terhadap pola keragaman dan sistem perkawinannya sangat tergantung pada karakter yang dipilih (mating reference) untuk mengelompokan tipe-tipe perkawinan kedalam kawin acak ataupun berpilih. Pemilihan mating reference tergantung pada keberadaan informasi tentang frekuensi tipe (genotypes) di dalam populasi dan/atau pada aspek khusus dari sistem perkawinan (Starck, Gregorius 1988). Apabila frekuensi dan pemilihan tipe perkawinan yang tepat (mating reference) telah ditentukan maka karakter dasar dari suatu sistem perkawinan disebut sebagai mating preferences (perkawinan yang dikehendaki).
Sistem perkawinan dalam suatu tegakan sumber benih perlu dipahami dengan benar karena akan mempengaruhi kualitas maupun kuantitas generasi keturunannya. Dikatakan bahwa potensi yang besar dari suatu sumber benih tidak akan terjadi secara maksimal pada generasi keturunannya apabila beberapa faktor tidak diperhatikan, seperti tidak seimbangnya produksi bunga betina dan bunga jantan, kurangnya sinkronisasi perkembangan diantara organ reproduksi, perbedaan kompatibilitas antar klon, frekuensi yang signifikan dari penyerbukan sendiri secara alami dan pengaruh yang tidak dikehendaki dari sumber polen ekternal (Burczyk & Chalupka 1997). Menurut El-Kassaby et al. (1984) proses pembungaan akan mempengaruhi keberhasilan perkawinan antar klon di dalam suatu populasi sumber benih, dengan demikian akan berdampak terhadap kualitas maupun kuantitas produksi benih yang dihasilkan. Kang (2000) juga mengindikasikan bahwa kelimpahan bunga merupakan kunci keberhasilan perkawinan, karena komposisi genetik pada suatu tegakan sumber benih sangat tergantung pada jumlah bunga betina dan jantan yang dihasilkan oleh populasi pohon induk. Demikian juga, proporsi relatif bunga betina dan jantan akan mempengaruhi jumlah benih bernas melalui tingkat ketersediaan polen ketika organ betina reseptif (Choi et al. 2004).
25
(Nurtjahjaningsih 2008). Pola transmisi adalah merupakan sistem perkawinan suatu jenis (Hartl & Clark 1997), maka bersama-sama dengan aliran gen yang dimediasi oleh polen dan biji mempunyai pengaruh yang kuat terhadap struktur genetik populasi tanaman (Burczyk et al 2004, Dow & Ashley 1998).
Berbagai tipe sistem perkawinan dijelaskan sebagai berikut (Finkledey 2005):
Perkawinan Acak (random mating):
Dalam populasi perkawinan acak, semua mating preferences adalah sama dengan satu. Sebuah tipe Ti dikatakan berkawin acak jika tidak menunjukkan adanya preferensi terhadap tipe tertentu lainnya. Preferensi kawin dari tipe Ti terhadap tipe Tj berarti tipe Tj lebih sering menjadi pasangan dari tipe Ti dibandingkan dengan ‘yang dikehendaki’ berdasarkan frekuensinya di populasi. Dalam hal ini tipe Ti lebih menyukai tipe Tj sebagai pasangan perkawinan (Finkeldey 2005).
Perkawinan Berpilih (Assortative Mating):
Kawin acak sulit terjadi pada jenis pohon hutan tropis yang mempunyai kerapatan rendah. Struktur spasial dan karakter pembungaan (early atau late flowering) akan menyebabkan adanya preferensi diantara tipe-tipe tertentu atau disebut perkawinan berpilih (assortative mating). Dengan demikian, dapat dikatakan bahwa kawin berpilih adalah merupakan sebuah penyimpangan dari kawin acak.
Perkawinan Tidak Acak (Non-Random Mating):
Dalam populasi pohon hutan perkawinan tidak acak diharapkan akan sering terjadi. Hal ini disebabkan periode pembungaan yang jelas bervariasi pada individu pohon, dan kemungkinan inkompatibilitas prezygotic pada kebanyakan jenis pohon angiospermae, memunculkan berbagai bentuk positif atau negatif dari assortative mating.
Self-incompatibility secara genetis sering terjadi pada banyak pohon hutan tropis, yang ditandai dengan sedikit atau tidak ada pembentukan biji setelah melakukan penyerbukan sendiri (Bawa et al. 1985). Pada jenis konifer, umumnya yang terjadi adalah perbedaan waktu pematangan antara bunga jantan dan bunga betina, namun dapat juga karena pemandulan sendiri (self-sterility) dimana biji gugur sebelum matang dan kematian ini diduga karena matinya gen resesif, yaitu terjadi setelah individu yang berkerabat dekat melakukan penyerbukan (inbreeding) (NAS 1991).
Pada tanaman mindi belum diketahui secara jelas model perkawinan yang terjadi. Walaupun struktur bunga memiliki tipe hermaprodit, namun belum diketahui apakah melakukan perkawinan sendiri atau silang atau campuran keduanya. Memperhatikan potensi reproduksi yang cukup tinggi pada tanaman mindi dengan keberhasilan reproduksi sebelum perkecambahan (KRSP) 34% (Syamsuwida 2009, tidak dipublikasi), maka diduga tanaman mindi banyak melakukan perkawinan silang (outcrossing), sehingga menghasilkan ratio ovul terhadap biji yang relatif tinggi. Walaupun hal ini masih perlu dikaji dengan menganalisis secara genetik pohon induk dan keturunannya untuk mengetahui derajat outcrossing tanaman mindi.
2.3.3 Penyimpangan Genotipe dalam Sistem Perkawinan (Inbreeding)
Ketika polen dari suatu pohon menyerbuki bunga yang terdapat pada pohon itu sendiri, maka disebut penyerbukan sendiri (selfing). Hal ini juga dapat diterapkan pada penyerbukan yang terjadi antara ramet (bahan tanaman berasal dari organ vegetatif) dari famili yang sama. Meskipun ramet-ramet tersebut berbeda tanaman, mereka tetap identik secara genetik.
Selfing sering terjadi pada geitonogami (penyerbukan dari bunga yang berbeda pada satu tanaman) dan autonogami (penyerbukan dari bunga yang sama). Kedua tipe sexual ini termasuk pada tumbuhan hermaprodit yaitu tanaman yang berbunga sempurna dimana gamet jantan maupun betina dihasilkan pada satu bunga yang sama. Penyerbukan sendiri ini tidak mungkin terjadi pada jenis tanaman yang dioecious (berumah dua dimana bunga jantan dan bunga betina dihasilkan pada dua individu yang berbeda).
27
lingkungan. Faktor penentu genetik yang benar-benar menolak selfing adalah system seksual dioecious dan system inkompatibilitas. Faktor genetik lain adalah postzygotic self-sterility (sterilitas otomatis setelah zigot) yaitu suatu mekanisme genetis yang menghasilkan keturunan hasil selfing dalam frekuensi rendah.
Tanaman hutan yang biseksual, banyak yang mampu melakukan penyerbukan selfing dan outcrossing. Derajat selfing dari tanaman yang melakukan system perkawinan campuran tersebut menunjukkan variasi temporal dan spasial. Hal ini disebabkan kuatnya pengaruh kondisi lingkungan pada sistem perkawinan campuran.
Derajat selfing biasanya diduga berdasarkan pola variasi genetik pada lokus-lokus gen penanda yang terdapat pada keturunan dari satu pohon induk dan atau suatu populasi.
Metode yang digunakan adalah pendugaan derajat selfing berdasarkan alelik yang jarang/unik pada lokus gen tertentu. Yang dimaksud dengan alelik yang jarang adalah alelik yang tidak dimiliki oleh pohon lain yang bereproduksi pada populasi yang sama. Alelik yang unik pada lokus gen yang ko-dominan akan mudah dikenali apabila pohon tersebut homozygot. Tetapi kasus ini jarang terjadi pada populasi alam. Pada populasi alam, alelik yang unik biasanya ditemukan pada organisme heterozigot.
Metode lain yang digunakan untuk menduga derajat selfing adalah berdasarkan model perkawinan campuran. Struktur genotipik keturunan yang berasal dari selfing berbeda dalam banyak hal dari struktur genotipik keturunan hasil outcrossing. Keturunan hasil selfing hanya mempunyai alelik-alelik dari pohon induknya, pada seluruh lokus gen, meskipun genotipe sebuah keturunan dapat berbeda dari pohon induknya, apabila pohon induknya heterozigot. Keturunan hasil outsrossing membawa alelik-alelik yang ada di populasi. Penurunan proporsi heterozigot yang khas dapat diduga pada keturunan hasil selfing, bila dibandingkan dengan dengan keturunan dari outcrossing secara penuh.
asumsi lainnya dari model perkawinan campuran kemungkinan adalah jenis tanaman dengan kerapatan rendah. Penyimpangan dari asumsi model tersebut dapat menyebabkan lahirnya pendugaan yang tidak ada artinya secara biologis, seperti derajat outcrossing yang secara nyata lebih tinggi dari 1, sehingga konsekuensinya derajat selfing menjadi negative (suatu hal yang tidak mungkin terjadi) (Finkeldey 2005).
Bentuk penyimpangan genotipe yang sering terjadi dalam sistem perkawinan adalah adanya inbreeding (penyerbukan dalam).
Selfing bukanlah satu-satunya penyebab terjadinya inbreeding. Beberapa mekanisme dapat mengurangi kemungkinan selfing, akan tetapi tidak dapat dilakukan pada inbreeding. Tingkat inbreeding tidak hanya ditentukan oleh sistem reproduksi alami, tetapi juga oleh struktur famili yang mana dipengaruhi oleh karakteristik polen dan penyebaran benih (Finkeldey 2005).
Penyebaran polen dan benih yang melampaui batas area dapat meningkatkan kekerabatan genetik dari individu yang berdekatan didalam sebuah populasi perkawinan jadi berpotensi untuk inbreeding. Ketahanan hidup beberapa pohon dibandingkan dengan yang lainnya dalam sebuah populasi juga dapat mengakibatkan relatif sedikitnya individu yang secara genetik mendominasi gene pool . Hal ini, apabila dikombinasikan dengan faktor-faktor seperti ketidaksesuaian antara bunga jantan dan bunga betina, generasi yang overlapping didalam populasi, dan rasio seksual yang tidak sama, akan lebih berpotensi meningkatkan derajat inbreeding.
Apomiksis, atau reproduksi uniparental yang







![Gambar 11 Pembesaran ovarium [ovr], stilus [st] A) irisan melintang ovarium](https://thumb-ap.123doks.com/thumbv2/123dok/956552.394410/74.595.46.480.44.820/gambar-pembesaran-ovarium-ovr-stilus-irisan-melintang-ovarium.webp)