PENDUGAAN PARAMETER GENETIK JAGUNG DAN PEMILIHAN
LINGKUNGAN SELEKSI UNTUK PEMUPUKAN RENDAH
SUTORO
SEKOLAH PASCA SARJANA
INSTITUT PERTANIAN BOGOR
PERNYATAAN
MENGENAI DISERTASI DAN
SUMBER INFORMASI
Dengan ini saya menyatakan bahwa disertasi Pendugaan Parameter Genetik Jagung dan Pemilihan Lingkungan Seleksi untuk Pemupukan Rendah adalah karya saya sendiri dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir disertasi ini.
Bogor, Desember 2005
ABSTRAK
SUTORO. Pendugaan Parameter Genetik Jagung dan Pemilihan Lingkungan Seleksi untuk Pemupukan Rendah. Dibimbing ABDUL BARI, SUBANDI dan SUDIRMAN YAHYA.
Sebagian besar varietas jagung hasil pemulia tanaman yang sudah dilepas adalah varietas yang responsif terhadap pemupukan, sehingga apabila varietas tersebut ditanam pada lahan yang kurang subur maka hasilnya akan rendah. Untuk mendapatkan varietas yang toleran pada kondisi pemupukan rendah dapat dilakukan melalui perbaikan populasi tanaman. Lingkungan seleksi menentukan keberhasilan pemuliaan untuk mendapatkan varietas yang sesuai pada lingkungan yang menjadi target. Tujuan penelitian ini untuk mengetahui perbedaan besaran parameter genetik antar taraf pemupukan dan pada lingkungan pemupukan yang mana seleksi selayaknya dilakukan agar mendapatkan varietas yang toleran pada pemupukan rendah. Penelitian dilakukan dengan menyilangkan antar galur S1 yang berasal dari varietas Bisma dengan menggunakan rancangan persilangan North Carolina II. Hasil persilangan dievaluasi dan diseleksi pada 3 taraf pemupukan. Pendugaan kemajuan seleksi dilakukan untuk seleksi full-sib dan S1 berdasarkan penampilan half-sib.
Hasil penelitian menunjukkan bahwa semakin rendah taraf pemupukan semakin kecil ragam genetiknya yaitu total ragam genetik untuk lingkungan pemupukan rendah dan sedang berturut-turut sebesar 28.8% dan 64.3% dari total ragam genetki pada lingkungan pemupukan normakl. Sebagian besar komponen ragam untuk bobot biji varietas Bisma merupakan ragam aditif, baik pada kondisi pemupukan rendah (78%), sedang (67%) maupun normal (81%). Perbaikan populasi Bisma yang toleran terhadap pemupukan rendah dapat dilakukan melalui seleksi berulang. Seleksi langsung cenderung memberikan kemajuan seleksi yang terbaik, sehingga seleksi untuk lingkungan pemupukan rendah perlu dilakukan pada lingkungan pemupukan rendah. Seleksi pada lingkungan pemupukan sedang untk lingkungan pemupukan rendah memperoleh kemajuan seleksi bobot biji 1.69%, lebih rendah daripada seleksi pada lingkungan pemupukan rendah yang memperoleh kemajuan seleksi 7.57%. Populasi hasil seleksi bobot biji pada kondisi pemupukan normal menghasilkan respon terkorelasi karakter sekunder yang kurang menguntungkan untuk menunjang bobot biji bila ditanam pada kondisi pemupukan rendah.
ABSTRACT
SUTORO. Estimation of Genetic Parameters and Choice of Selection Environment for Improvement of Corn (Zea mays L.) for Low Fertilization Environment. Under direction of ABDUL BARI, SUBANDI, and SUDIRMAN YAHYA.
Most of the improved corn varieties released in Indonesia are responsive to fertilizer application rate, and they will be low yielding when grown in marginal area. To obtain corn variety which is tolerant to low fertilizer input could be done by improving population through selection. Selection environment determines breeding progress to obtain suitable variety for target environment. The objectives of this study were to estimate genetic parameters and to select for environment in which selection should be conducted when the improved population is to be grown under low fertilization. The experiment was carried out by crossing among S1 lines of Bisma variety using North Carolina Design II. Progenies were evaluated and selected in three levels of fertilizer application. Gain of selection was estimated using full-sib and S1 based on half-sib performance methods.
Result of the study showed that as the fertilizer application rate was decreasing genetic parameter values tended to be declining, total genetic variance under low and medium fertilization resulted in 28.8% and 64.3% of total genetic variance under high fertilization. Major component of the total genetic variance for grain yield was contributed by additive variance component which is 78%, 67% and 81% in low, medium and high fertilization level, respectively. Population improvement for low level of fertilization could be done by recurrent selection program. Direct selection resulted in the highest gain of selection. Therefore, selection under low fertilization was most suitable when the target environment was low fertility. Selection under medium level of fertilization resulted in realized gain of 1.69%, lower than 7.57% under low level of fertilization. Improved population selected under high fertilization level resulted in correlated secondary character responses, which were disadvantage to supporting high grain yield, when grown under low fertilization level.
PENDUGAAN PARAMETER GENETIK JAGUNG DAN PEMILIHAN
LINGKUNGAN SELEKSI UNTUK PEMUPUKAN RENDAH
SUTORO
Disertasi
sebagai salah satu syarat untuk memperoleh gelar Doktor pada
Program Studi Agronomi
SEKOLAH PASCA SARJANA
INSTITUT PERTANIAN BOGOR
Judul Disertasi : Pendugaan Parameter Genetik Jagung dan Pemilihan Lingkungan Seleksi untuk Pemupukan Rendah
Nama : Sutoro NRP : P03600004 Program studi : Agronomi
Disetujui
Komisi Pembimbing
Prof. Dr. Ir. Abdul Bari, MSc Ketua
Dr. Subandi, MSc Prof. Dr. Ir. Sudirman Yahya, MSc Anggota Anggota
Diketahui
Ketua Program Studi Agronomi Dekan Sekolah Pasca Sarjana
Dr. Ir. Satriyas Ilyas, MS Prof. Dr. Ir. Syafrida Manuwoto, MSc
RIWAYAT HIDUP
Penulis dilahirkan di Solo pada tanggal 8 Desember 1953 dari pasangan Sutarman dan Sundari. Pendidikan sarjana ditempuh di Fakultas Pertanian IPB, lulus tahun 1977. Pada tahun 1985 penulis lulus dari Fakultas Pasca Sarjana IPB. Pada tahun 2000 melanjutkan ke program doktor pada program studi Agronomi. Beasiswa pendidkan pasca sarjana diperoleh dari Badan Litbang Departemen Pertanian Republik Indonesia.
Penulis bekerja pada Balai Penelitian di lingkungan Badan Litbang Pertanian sejak tahun 1977 sebagai peneliti sesuai dengan perkembangan mandat Balai Penelitian yaitu bidang agronomi (budidaya), ekofisiologi dan pengelolaan plasma nutfah tanaman pangan. Hasil penelitian yang telah dilakukan telah diterbitkan dalam jurnal ilmiah di antaranya dalam Penelitian Pertanian dan Plasma Nutfah. Penulis sekarang bekerja sebagai peneliti Pengelolaan Sumberdaya Genetik Pertanian di Balai Besar Penelitian Bioteknologi dan Sumberdaya Genetik Pertanian.
Pendidikan jangka pendek yang pernah diikuti di antaranya bidang analisis statistika dan simulasi produksi tanaman padi di IRRI, Filipina, bidang agronomi dan evaluasi plasma nutfah di Lembaga Penelitian dan University of Tokyo, Jepang dan bidang pengelolaan plasma nutfah di NBPGR, India.
PRAKATA
Puji syukur kehadirat Allah SWT yang telah melimpahkan rahmat-Nya kepada penulis, sehingga disertasi ini dapat terwujud.
Disertasi ini memberikan informasi mengenai nilai dugaan parameter genetik jagung dan pemilihan lingkungan seleksi untuk pemupukan rendah.
Pada kesempatan ini penulis mengucapkan banyak terima kasih kepada Prof. Dr. Ir. Abdul Bari,MSc sebagai ketua komisi pembimbing, Dr. Subandi, MSc, dan Prof. Dr. Ir. Sudirman Yahya, MSc sebagai anggota komisi pembimbing yang telah banyak memberikan pengarahan dan bimbingan kepada penulis sehingga desertasi ini dapat tersusun. Di samping itu, penghargaan penulis sampaikan kepada Kepala Balai Besar Penelitian Bioteknologi dan Sumberdaya Genetik Pertanian (BB Biogen), Badan Litbang Pertanian yang telah memberikan kesempatan kepada penulis untuk mengikuti pendidikan doktor di Institut Pertanian Bogor serta fasilitas lainnya sehingga penelitian ini dapat terlaksana dengan baik, dan Badan Penelitian dan Pengembangan Pertanian Departemen Pertanian atas fasilitas yang telah diberikan kepada penulis sehingga dapat menyelesaikan pendidikannya. Ungkapan terima kasih juga disampaikan kepada semua teknisi Kebun Percobaan Cikeumeuh dan Laboratorium Bank Gen BB Biogen yang telah membantu penulis dalam pelaksanaan penelitian, serta isteri (Nita Riswari), putra-putri (Dian Ninditasari, Rianto Yuwono, Handaru Riswantoro, Reza Hanung Pradipta) dan seluruh keluarga penulis atas doa dan kasih sayang serta pengertiannya dalam banyak hal.
Akhirnya, penulis berharap semoga hasil peneltian ini bermanfaat bagi semua pihak yang memerlukannya.
Bogor, Desember 2005
DAFTAR ISI
Halaman
DAFTAR TABEL ………...………..……. vi
DAFTAR GAMBAR ……….…. vii
DAFTAR LAMPIRAN ……….….. viii
PENDAHULUAN Latar Belakang ……….……… 1
Tujuan Penelitian ………. 2
Hipotesis Penelitian………..……. 2
TINJAUAN PUSTAKA Populasi Varietas Bisma ……….. 4
Rancangan Persilangan ……….…. 4
Pendugaan Parameter Genetik ……….…… 5
Metode Seleksi ……….……. 6
Lingkungan Seleksi ………..……... 7
Karakter TanamanToleran Cekaman ……….... 9
BAHAN DAN METODE Pembentukan S1, Persilangan antar S1 dan Evaluasi ...………... 10
Seleksi Famili, Persilangan antar Famili dan Evaluasi ...…………... 13
Analisis Data ………... 16
HASIL PERCOBAAN Percobaan di Lingkungan Seleksi ………..….. 19
Percobaan di Lingkungan Evaluasi ……….… 29
PEMBAHASAN Validitas Asumsi dalam Pendugaan Parameter Genetik ... 33
Ragam Genetik ...
35Heritabilitas dan Korelasi Genetik ... 36
Prediksi dan Realisasi Kemajuan Seleksi Bobot Biji di Lingkungan
Evaluasi ...
39Respon Terkorelasi Karakter Sekunder ...
43KESIMPULAN DAN SARAN ………..……….... 51
DAFTAR PUSTAKA ……….…… 52
DAFTAR TABEL
Halaman
1. Analisis ragam bagi rancangan persilangan NCD II untuk satu lingkungan... 18 2. Nilai kuadrat tengah bobot biji (kg/plot) pada lingkungan seleksi pemupukan
rendah, sedang dan normal ... 19 3. Nilai penduga ragam bobot biji tetua jantan, betina dan interaksinya serta
kovarian famili halfsib dan fullsib pada lingkungan seleksi pemupukan rendah, sedang dan normal………..…... 20 4. Nilai dugaan ragam genetik bobot biji (kg/plot) populasi varietas Bisma (Co)
pada lingkungan seleksi pemupukan rendah, sedang dan normal... ... 21 5. Nilai dugaan ragam galat percobaan, ragam antar plot, ragam dalam plot
dan ragam lingkungan mikro dalam plot dari bobot biji pada lingkungan seleksi ... 23 6. Nilai dugaan ragam fenotipik dan heritabilitas famili fullsib dan halfsib bobot
biji (kg/plot) populasi varietas Bisma (Co) pada lingkungan seleksi pemupukan rendah, sedang dan normal... 23 7. Rata-rata famili FS dan HS serta diferensial seleksi pada lingkungan seleksi
pemupukan rendah, sedang dan normal ... 24 8. Korelasi genetik bobot biji antar lingkungan pemupukan ... 24 9. Ragam aditif dan dominan karakter sekunder pada lingkungan seleksi
pemupukan rendah, sedang dan normal ... 25 10. Hertabilitas karakter sekunder pada lingkungan seleksi pemupukan rendah,
sedang dan normal ... 26 11. Korelasi genetik bobot biji dengan karakter sekunder pada lingkungan
pemupukan yang sama atau berbeda ... 28 12. Bobot biji hasil persilangan famili terpilih pada pemupukan
rendah, sedang dan normal ... 30 13. Rata-rata ASI, bobot 200 butir, klorofil, jumlah daun dan LAI saat
pembungaan pada lingkungan evaluasi pemupukan rendah, sedang dan normal... 31 14. Rata-rata jumlah daun,LAI , porsi daun menua saat panen dan jumlah biji
tiap tongkol pada lingkungan evaluasi pemupukan rendah, sedang dan
normal... ... 32 15. Nilai prediksi dan realisasi persentase kemajuan seleksi bobot biji ... 40 16. Respon terkorelasi karakter sekunder pada lingkungan pemupukan rendah
akibat seleksi bobot biji pada lingkungan pemupukan rendah, sedang dan normal ... 44 17. Respon terkorelasi jumlah daun dan LAI saat pembungaan pada
lingkungan pemupukan rendah akibat seleksi bobot biji pada lingkungan
pemupukan rendah, sedang dan normal ... 46 18. Respon terkorelasi jumlah daun dan LAI saat panen pada lingkungan
pemupukan rendah akibat seleksi bobot biji pada lingkungan pemupukan rendah, sedang dan normal ... 47 19. Respon terkorelasi porsi daun menua dan jumlah biji pada lingkungan
pemupukan rendah akibat seleksi bobot biji pada lingkungan pemupukan
DAFTAR GAMBAR
Halaman 1. Diagram pelaksanaan kegiatan penelitian ………..………. 15 2. Rata-rata heritabilitas terealisasi pada seleksi langsung dan tak langsung
(R-R, S-S, N-N menunjukkan seleksi langsung pemupukan rendah (R), sedang (S) dan normal (N)) ... 37 3 Prediksi dan realisasi kemajuan seleksi langsung dan tak langsung
famili fullsib (R-R, S-S, N-N menunjukkan seleksi langsung pemupukan rendah (R), sedang (S) dan normal (N)) ... 41 4. Prediksi dan realisasi kemajuan seleksi langsung dan tak langsung famili
DAFTAR LAMPIRAN
Halaman 1. Nilai kuadrat tengah bobot biji/tanaman pada lingkungan seleksi
pemupukan rendah, sedang dan normal ... 57 2. Nilai kuadrat tengah ASI pada lingkungan seleksi pemupukan
rendah, seang dan normal ... 57 3. Nilai kuadrat tengah bobot 200 butir bijipada lingkungan seleksi
seleksi pemupukan rendah, sedang dan normal ... 57 4. Nilai kuadrat tengah klorofil pada lingkungan seleksi pemupukan
rendah, sedang dan normal ... 58 5. Nilai kuadrat tengah jumlah daun saat pembungaan pada
lingkungan seleksi pemupukan rendah, sedang dan normal ... 58 6. Nilai kuadrat tengah LAI saat pembungaan pada lingkungan
seleksi pemupukan rendah, sedang dan normal ... 58 7. Nilai kuadrat tengah jumlah daun saat panen pada lingkungan
seleksi pemupukan rendah, sedang dan normal ... 59 8. Nilai kuadrat tengah LAI saat panen pada lingkungan seleksi
pemupukan rendah, sedang dan normal ... 59 9. Nilai kuadrat tengah porsi daun menua pada lingkungan
seleksi pemupukan rendah, sedang dan normal ... 59 10. Nilai kuadrat tengah jumlah biji tiap tongkol pada lingkungan
seleksi pemupukan rendah, sedang dan normal ... 59 11. Famili fullsib terpilih pada lingkungan seleksi pemupukan rendah,
sedang dan normal ... 60 12. Galur S1 terpilih berdasarkan penamplan famili HS pada
lingkungan seleksi pemupukan rendah, sedang dan normal ... 61 13. Respon terkorelasi ASI akibat seleksi bobot biji pada lingkungan
pemupukan yang sama atau berbeda ... 62 14. Respon terkorelasi bobot 200 butir akibat seleksi bobot biji pada
lingkungan pemupukan yang sama atau berbeda ... 63 15. Respon terkorelasi klorofil akibat seleksi bobot biji pada
lingkungan pemupukan yang sama atau berbeda ... 64 16. Respon terkorelasi jumlah daun saat pembungaan akibat seleksi
bobot biji pada lingkungan pemupukan yang sama atau berbeda ... 65 17. Respon terkorelasi LAI saat pembungaan akibat seleksi bobot
biji pada lingkungan pemupukan yang sama atau berbeda ... .... 66 18. Respon terkorelasi jumlah daun saat panen akibat seleksi bobot
biji pada lingkungan pemupukan yang sama atau berbeda ... 67 19. Respon terkorelasi LAI saat panen akibat seleksi bobot biji pada
lingkungan pemupukan yang sama atau berbeda ... 68 20. Respon terkorelasi porsi daun menua akibat seleksi bobot biji pada
lingkungan pemupukan yang sama atau berbeda ... 69 21. Respon terkorelasi jumlah biji tiap tongkol akibat seleksi bobot biji
pada lingkungan pemupukan yang sama atau berbeda ... 70 22. Iustrasi perhitungan penduga ragam ... 71 23. Ilustrasi perhitungan penduga ragam lingkungan mikro ... 72 24. Jumlah kuadrat tetua betina, janayan, interaksi betina x janan dan galat
PENDAHULUAN
Latar Belakang
Pemuliaan tanaman jagung pada umumnya dilakukan pada lahan yang dipupuk dengan taraf optimal, baik pada fase seleksi, uji daya hasil pendahuluan maupun uji daya hasil lanjutan. Proses pemuliaan pada lingkungan optimal ini berakibat varietas unggul yang dihasilkan menjadi hanya adaptif pada lahan subur. Produktivitas tanaman jagung yang rendah di antaranya disebabkan oleh kemampuan petani dalam memberikan pupuk dalam jumlah yang kurang memadai pada tanaman yang tumbuh di lahan marjinal. Varietas jagung yang dihasilkan oleh pemulia tanaman sebelumnya yaitu varietas yang responsif terhadap pemupukan, sehingga apabila varietas tersebut ditanam pada lahan yang kurang subur maka hasilnya akan rendah dan bahkan lebih rendah daripada varietas lokal. Oleh karena itu, perlu diperoleh varietas jagung yang toleran terhadap pemupukan rendah. Untuk mendapatkan varietas yang toleran pemupukan rendah dapat diperoleh melalui perbaikan populasi tanaman. Perbaikan populasi tanaman dapat diduga berdasarkan informasi parameter genetik dari populasi tanaman.
Komponen ragam genetik suatu populasi terdiri dari komponen ragam genetik aditif dan non-aditif (dominan dan epistasis), dapat digunakan untuk menduga kemajuan genetik dari metode seleksi yang akan digunakan. Suatu populasi yang memiliki ragam genetik aditif tinggi dapat diperbaiki melalui seleksi intra-populasi, sedangkan seleksi inter-populasi digunakan bila ragam genetik dominan tinggi (Malvar et al., 1996). Besaran ragam genetik jagung berbeda antar lingkungan cekaman kekeringan (Hallauer dan Miranda, 1985), hara N (Banziger et al., 1997) dan garam (Rao dan McNeilly, 1999). Di samping itu, besaran parameter genetik juga berbeda antar populasi, sehingga untuk memperbaiki suatu populasi jagung yang sesuai dengan lingkungan target perlu diketahui besaran parameter genetiknya.
biji tinggi pada kondisi hara N cukup, ternyata hibrida-hibrida tersebut memberikan hasil yang bervariasi pada kondisi hara N rendah (Yasin et al., 1998).
Telah banyak varietas jagung bersari bebas yang telah dilepas oleh pemulia tanaman. Varietas Bisma yang dilepas tahun 1995 telah populer karena keunggulannya dalam beberapa pengujian di lahan kering (Subandi et al.,1998). Jagung varietas Bisma memiliki latar belakang genetik yang luas, sehingga masih mungkin mendapatkan varietas yang toleran terhadap pemupukan rendah dengan cara menyeleksi dari populasi varietas Bisma ini. Parameter genetik tanaman jagung varietas Bisma, sebagai populasi jagung yang akan diperbaiki sifat toleransinya terhadap pemupukan rendah, perlu diketahui untuk menduga kemajuan genetik yang diperoleh akibat seleksi.
Lingkungan seleksi menentukan keberhasilan pemuliaan untuk mendapatkan varietas yang cocok dengan lingkungan yang menjadi target. Seleksi sering dilakukan pada kondisi tanpa cekaman (lingkungan optimum) karena banyak pendapat yang menyatakan pada kondisi optimum umumnya memiliki heritabilitas bobot biji yang lebih tinggi daripada heritabilitas di lingkungan tanpa cekaman (Ceccarelli, 1994). Nampaknya seleksi pada lingkungan yang mirip dengan lingkungan target akan menghasilkan kemajuan seleksi yang lebih besar daripada seleksi tak langsung atau seleksi pada lingkungan yang sangat berbeda dengan lingkungan target (Banziger et al., 1997). Lebih lanjut Banziger et al. (1997) menyatakan masih terdapat perbedaan pendapat pada lingkungan mana sebaiknya seleksi itu dilakukan. Oleh karena itu, pada lingkungan cekaman pemupukan yang mana seleksi selayaknya dilakukan agar mendapatkan varietas yang toleran pemupukan rendah perlu diketahui.
Tujuan Penelitian
Penelitian ini bertujuan untuk menyediakan informasi bagi pemulia tanaman dalam memilih lingkungan seleksi, berdasarkan nilai besaran ragam genetik sifat kuantitatif tanaman dalam populasi jagung varietas Bisma untuk mendapatkan varietas yang toleran lingkungan pemupukan rendah.
Hipotesis Penelitian
TINJAUAN PUSTAKA
Populasi Varietas Bisma
Varietas jagung unggul dapat diperoleh melalui perbaikan populasi yang memiliki latar belakang genetik yang luas. Varietas Bisma merupakan varietas yang memiliki latar belakang genetik sebagai persilangan antara Gen Pool-4 dengan bahan introduksi yang disertai seleksi massa selama 5 generasi (Anonim, 1999). Gen Pool-4 merupakan populasi dasar yang dibentuk dari sejumlah koleksi plasma nutfah jagung lokal dan introduksi dengan biji yang berwarna kuning dan berumur lebih dari 100 hari (Subandi, 1984).
Varietas Bisma merupakan jagung bersari bebas telah meluas pengembangannya selain berdaya hasil tinggi, juga tahan terhadap penyakit bulai, penyakit karat dan bercak daun serta toleran terhadap kekeringan (Subandi et al., 1998). Perbaikan karakter tertentu dari populasi varietas Bisma yang memiliki latar belakang genetik yang luas ini masih memungkinkan, seperti perbaikan varietas untuk sifat toleran terhadap pemupukan rendah.
Rancangan Persilangan
dengan membuat galur S1 untuk mewakili So. Tongkol yang dihasilkan dari biji-biji galur S1 dapat digunakan sebagai tetua betina dalam persilangan (Hallauer dan Miranda, 1985). Berbeda dengan rancangan persilangan III, keturunan dihasilkan dari silang balik antara turunan F2 dengan kedua tetua galur murni yang menghasilkan F2. Dalam pengujian keturunan hasil persilangan dari NCD I, NCD II dan NCD III di lapangan, maka untuk menghindari ukuran blok yang lebih luas, turunan dari sejumlah tetua (hallf-sib dan full-(hallf-sib) dapat dikelompokkan ke dalam set (Comstock, 1996) untuk mengurangi galat percobaan.
Berdasarkan informasi genetik yang diperoleh dari ketiga rancangan ini, maka informasi dari NCD III lebih banyak dari NCD II, dan NCD II lebih banyak dari NCD I. Pada NCD I , ragam dari penduga ragam dominan (ó2D) biasanya besar, karena merupakan fungsi kuadrat tengah yang kompleks. NCD I dan NCD II tidak dapat digunakan untuk pengujian adanya epistasis (Hill et al., 1998). Pada tanaman jagung, epistasis tidak mengakibatkan penyimpangan pendugaan ragam aditif dan dominan yang serius (Hallauer dan Miranda, 1985). Selanjutnya Hallauer dan Miranda (1985) menyatakan bahwa NCD II efektif untuk menduga komponen ragam genetik populasi jagung. Dalam penelitian ini, tetua-tetua S1 dipilih secara acak yang dibuat dari populasi varietas Bisma sebagai populasi referen. Tanaman-tanaman dari galur S1 ini digunakan sebagai tetua persilangan dalam NCD II untuk menduga parameter genetik populasi jagung Bisma pada 3 taraf pemupukan.
Pendugaan Parameter Genetik
pada 2 populasi jagung Spanyol (Ribadumia yang berbiji warna putih dan Tuy yang berbiji warna kuning) mendapatkan penduga ragam dominan lebih besar (0,90-1,11) daripada ragam aditif (0,10-0,16) untuk kedua populasi tersebut, sehingga ragam dominan lebih penting pada kedua populasi jagung tersebut.
Dalam pendugaan parameter genetik aditif dan dominan sering diasumsikan tidak ada epistasis. Hallauer dan Miranda (1985) menduga ragam epistasis ó2AA dengan menggunakan data hasil biji dari dua percobaan dalam NCD I dan NCD II pada populasi BBBS. Dari dua persamaan Cov HS= ¼ ó2A + 1/16 ó2AA = 34,8 (berasal dari NCD I) dan Cov HS= ½ ó2A + ¼ ó2AA = 60.8 (berasal dari NCD II), maka diperoleh nilai penduga ó2AA sebesar –70,8 dengan simpangan baku 223,5. Berdasarkan penduga ragam bernilai negatif, maka ragam epistasis ó2AA dapat dianggap 0 (nol) atau tidak nyata kontribusinya. Eta-Ndu dan Openshaw (1999) yang menganalisis keberadaan epistasis bagi hasil jagung pada famili F2 dari persilangan A679xW6005 dan A679xFR902 menunjukkan adanya epistasis pada kedua populasi tersebut.
Pada umumnya lingkungan cekaman mengakibatkan perubahan terhadap besaran ragam genetik. Dari hasil evaluasi pada lingkungan cekaman air, menunjukkan bahwa cekaman menyebabkan penurunan keragaman genetik (Hallauer dan Miranda, 1985). Rao dan McNeilly (1999) yang menduga parameter genetik panjang akar jagung pada 3 lingkungan cekaman garam menunjukkan bahwa ragam aditif konsisten lebih besar daripada ragam non aditif pada ketiga cekaman garam. Nisbah ragam dominan-aditif pada kondisi cekaman garam lebih besar daripada kondisi tanpa cekaman garam. Nilai penduga parameter genetik berbeda dari populasi satu dengan populasi lainnya. Oleh karena itu, untuk memperbaiki suatu populasi tertentu diperlukan informasi besaran parameter genetik bagi populasi yang akan diperbaiki karakternya.
Metode Seleksi
sebagai populasi dasar dari siklus pertama dapat meningkatkan hasil 23,6% (Subandi, 1982), seleksi half-sib dengan varietas Arjuna sebagai pejantan dan 3 varietas Penjalinan Y(C1), ICS2 Genjah Kretek, dan BC10 MS4 sebagai tetua betina dari siklus satu ke silkus kedua meningkatkan hasil masing-masing berturut-turut 7,9%, 7,5% dan 0,5% (Subandi, 1987). Stojsin dan Kannenberg (1994) yang mengevaluasi 5 populasi jagung menyatakan bahwa umumnya seleksi famili selfing merupakan prosedur seleksi yang paling efektif lalu diikuti oleh seleksi famili half-sib. Respon seleksi famili selfing terutama diakibatkan oleh perubahan frekuensi allel yang memiliki pengaruh aditif, sedangkan famili half-sib dari pengaruh aditif dan dominan. Populasi jagung toleran kekeringan La Posta Sequia dan Pool 26 Sequia yang diseleksi berdasarkan S1 memperoleh kemajuan genetik 12,4-12,7 % tiap siklus, sedangkan Tuxpeno Sequia yang juga toleran kekeringan diseleksi dengan famili full-sib memperoleh kemajuan genetik 3,8% tiap siklus pada kondisi kekeringan (Edmeades et al., 1999).
Seleksi massa nampaknya efektif pada dua silkus pertama pada populasi jagung Perta dan Metro, karena siklus selanjutnya respon tidak menentu dan di antaranya disebabkan ragam aditif yang relatif kecil (Subandi et al., 1980). Seleksi massa pada jagung prolifik BC10 dapat meningkatkan bobot biji 4,4% tiap siklus, dan setelah 10 siklus bobot biji meningkat 32% dari populasi dasar (Subandi, 1990).
Lingkungan Seleksi
Varietas unggul yang telah dilepas, diperoleh melalui seleksi pada kondisi pemupukan optimum dan dievaluasi dengan pemupukan yang optimum pula. Varietas yang beradaptasi luas sebenarnya hanya secara geografis, karena dievaluasi dan diseleksi pada lingkungan optimum. Oleh karena itu, keunggulan varietas pada kondisi optimum tidak diekspresikan pada lingkungan sub-optimal (Ceccareli, 1994).
(2001) menyatakan strategi pemuliaan untuk memperoleh peningkatan produksi pada kondisi masukan rendah dengan PPB lebih efektif dari pada pemuliaan formal, karena langsung pada lingkungan target.
Pada populasi jagung toleran kekeringan, kemajuan seleksi dapat diperoleh pada kondisi kekeringan maupun tanpa kekeringan. Kemajuan seleksi pada kekeringan lebih tinggi dibandingkan dengan tanpa kekeringan. Populasi jagung DTP2-C2 yang toleran kekeringan ternyata sangat baik pada kondisi tanpa cekaman air. Hal ini memberi petunjuk bahwa populasi jagung tersebut yang toleran kekeringan tidak berkurang potensi hasilnya (Edmeades et al., 1999). Seleksi hasil tinggi pada lingkungan subur memacu hilangnya adaptasi spesifik untuk kondisi cekaman, sedangkan seleksi hasil tinggi pada lingkungan cekaman tidak menghilangkan potensi hasil (Ceccareli et al., 1998 dan Edmeades et al., 1999). Seleksi toleran kekeringan pada jagung dapat pula mengarahkan suatu perubahan yang adaptif menguntungkan pada kondisi cekaman N (Laffite dan Edmeades, 1995 dan Banziger et al., 1999).
Efisensi seleksi pada kondisi N tinggi lebih rendah dibandingkan dengan seleksi pada kondisi N rendah, bila lingkungan target pada kondisi yang mengakibatkan hasil jagung berkurang lebih dari 43% (Banziger et al., 1997). Hal ini memberi petunjuk bahwa seleksi jagung pada N rendah dapat memperbaiki populasi jagung yang cocok pada kondisi N rendah hingga sedang. Korelasi genetik antara hasil jagung pada kondisi N rendah dan N tinggi menurun dengan meningkatnya pengurangan hasil relatif pada N rendah, sehingga seleksi tak langsung pada N tinggi menjadi kurang efisien (Banziger et al. 1997).
Efisiensi seleksi langsung atau tak langsung pada lingkungan target tergantung dari karakteristik populasi dan intensitas cekaman pada lingkungan target tersebut. Pada penelitian ini akan diuji pengaruh lingkungan seleksi langsung dan tak langsung pada lingkungan pemupukan rendah, sedang dan tinggi atau normal yang biasa dilakukan oleh pemulia tanaman.
Karakter Tanaman Toleran Cekaman
Seleksi dengan menggunakan karakter sekunder dan hasil biji dibandingkan
mempertimbangkan kriteria yang diperkirakan berhubungan dengan perbaikan respon tanaman terhadap cekaman. Berdasarkan korelasi genetik hasil biji dengan ASI (anthesis silking interval), jumlah tongkol tiap tanaman, konsentrasi klorofil, jumlah daun hijau dan daun menua (senescence), maka karakter sekunder tersebut umumnya berkaitan dengan toleran cekaman N dan kekeringan (Banziger et al., 1997; Chapman dan Edmeades , 1999). Jumlah daun hijau saat panen berbeda antar genotip jagung pada kondisi kekeringan dan dapat diturunkan secara moderat (Chapman and Edmeades , 1999).
Akumulasi N sebelum fase pembungaan merupakan karakter penting pada lingkungan N rendah, karena merupakan refleksi dari potensi varietas untuk mengambil dan mengirimkan N untuk pengisian biji pada kondisi cekaman hara (Cox et al., 1991). Tanaman jagung kekurangan P mengurangi laju penuaan daun yang berada di bawah tongkol (Colomb et al., 2000).
BAHAN DAN METODE
Pembentukan S
1, Persilangan antar S
1dan Evaluasi
Untuk menduga parameter genetik varietas Bisma sebagai populasi dasar dilakukan 3 kegiatan yaitu pembentukan galur S1, persilangan antar galur S1 dan evaluasi hasil persilangan.
Pembentukan galur S1
Pembentukan galur S1 dilakukan dengan menanam jagung varietas Bisma pada bulan April 2001. Populasi varietas Bisma diperoleh dari Laboratorium Bank Gen, Balai Penelitian Bioteknologi dan Sumber Daya Genetika Pertanian ditanam di Kebun Percobaan Cikeumeuh, Bogor. Benih yang ditanam sebagai contoh acak dari populasi Bisma, ditanam dengan jarak 75 cm x 20 cm dengan satu tanaman tiap lubang dan dipupuk 300 Kg Urea, 100 kg SP36 dan 50 kg KCl per hektar. Selanjutnya setiap tanaman dilakukan penyerbukan diri-sendiri (selfing) yaitu menyerbuki bunga betina dengan pollen pada tanaman yang sama untuk memperoleh galur S1 untuk mewakili So dari populasi varietas Bisma. Galur S1 yang dibentuk sebanyak 250 galur, namun yang memiliki biji dalam jumlah cukup sebanyak 216 galur S1. Biji hasil panen tiap tongkol (galur S1) dipipil, dikumpulkan dalam kantong dan dikeringkan lalu disimpan dalam ruang dingin. Benih galur S1 selanjutnya akan digunakan untuk persilangan.
Persilangan antar galur S
1Persilangan antara galur S1 dilaksanakan pada bulan Mei 2002. Pertanaman dipupuk dengan 300 kg Urea, 100 kg SP36 dan 50 kg KCl tiap hektar. Persilangan antar galur S1 dilakukan menurut rancangan persilangan NCD II. Galur S1 yang telah dibentuk dibagi menjadi 36 set, masing-masing set terdiri dari 6 galur S1 (3 galur sebagai tetua jantan dan 3 galur sebagai tetua betina). Semua kemungkinan pasangan tetua jantan dan betina dari setiap set disilangkan sehingga diperoleh 9 persilangan. Tetua jantan atau betina digunakan sebagai salah satu tetua, jantan atau betina saja, sehingga kombinasi persilangan (Xij) dapat disusun sebagai berikut:
Tetua betina 4 5 6 1 X14 X15 X16 2 X24 X25 X26 3 X34 X35 X36
Setiap galur S1 ditanam sebanyak 20 tanaman dan persilangan tetua jantan dan betina di antara galur S1 dilakukan untuk mendapatkan 5-10 persilangan (Hallauer dan Miranda, 1985). Dari 36 set persilangan akan dipilih 30 set yang memiliki kombinasi persilangan yang lengkap dan akan dievaluasi pada 3 taraf pemupukan.
Evaluasi hasil persilangan
Biji hasil persilangan antar galur S1 yang telah dilakukan sebelumnya yang terdiri dari 30 set persilangan ditanam dengan rancangan acak kelompok tak lengkap (Incomplete randomized block design) dengan 3 ulangan yang dilakukan pada 3 taraf pemupukan yaitu pemupukan rendah, sedang dan normal. Pemupukan normal diberikan dengan taraf yang biasa dilakukan oleh pemulia tanaman jagung. Ketiga taraf pemupukan tersebut adalah:
a. Pupuk dosis normal : 300 kg Urea- 100 kg SP36- 50 kg KCl tiap hektar. b. Pupuk dosis sedang: 150 kg Urea – 50 kg SP36 tiap hektar.
c. Pupuk rendah: 75 kg Urea dan 25 kg SP36 tiap hektar.
Untuk mendapatkan data yang lebih baik, maka penempatan perlakuan (set) menurut strip plot (set yang sama berdampingan pada kondisi pupuk normal, sedang dan rendah). Setiap set persilangan ditambah varietas Bisma sebagai pembanding, yang akan digunakan sebagai kriteria pemilihan full-sib dan half-sib terbaik pada kegiatan penelitian berikutnya.
pertanaman jagung sebelumnya adalah 0,127 % N, 0,408 mg P/100 g dan 0,432 me K/100 g dan bekas pertanaman kedelai sebelumnya adalah 0,178 % N, 0,621 P mg/100 g dan 0,379 me K/100 g.
Jagung ditanam dengan jarak 80 cm x 20 cm dengan 1 tanaman tiap lubang. Masing-masing progeni ditanam sebanyak 20 tanaman dalam satu baris/plot. Biji masing-masing progeni diambil sejumlah biji yang sama dari tiap tongkol (balance composite) progeni yang diperoleh dari persilangan. Sepertiga dosis pupuk Urea, seluruh dosis pupuk SP36 dan KCl diberikan pada saat tanam dan sisanya dua-pertiga pupuk Urea diberikan pada saat tanaman berumur 1 bulan. Pupuk diberikan sekitar 7 cm di samping tanaman dengan cara ditugal. Perlindungan tanaman terhadap serangan hama-penyakit dan kompetisi dengan gulma dilakukan secara optimum. Karakter sekunder tanaman jagung yang diamati adalah karakter yang diduga dapat mempengaruhi peningkatan bobot biji jagung. Karakter tersebut adalah ASI (anthesis silking interval), jumlah daun dan LAI (leaf area index) pada saat pembungaan dan panen, porsi daun menua, kadar klorofil, jumlah biji tiap tongkol dan bobot 200 butir. ASI yang cepat dapat menghasilkan jumlah biji yang relatif lebih banyak, karena peluang bunga betina diserbuki oleh polen lebih besar. Tanaman jagung yang memiliki jumlah daun dan LAI serta klorofil yang tinggi diharapkan dapat memberikan hasil fotosintat yang tinggi, yang selanjutnya ditranslokasikan ke dalam biji.
Karakter tanaman jagung diamati dengan cara sebagai berikut:
1. Umur berbunga jantan dan betina yang ditetapkan bila 50% dari pertanaman telah muncul polen bunga jantan (anthesis) dan rambut bunga betina (silking). Selanjutnya ASI = selisih umur munculnya polen dengan rambut bunga betina. 2. Jumlah dan luas daun hijau tiap tanaman pada fase pembungaan dan saat
panen. Luas daun fase pembungaan dan saat panen diduga menurut Sutoro (1992) dan Sutoro (2003). LAI merupakan nisbah luas daun dengan luas lahan tempat tanaman itu berada.
3. Luas daun menua dihitung dengan mengurangi luas daun pada fase pembungaan dengan luas daun saat panen. Porsi daun menua merupakan nisbah luas daun menua dengan luas daun pada fase pembungaan.
4. Kadar klorofil daun pada tongkol yang diukur dengan Chlorophylmeter Minolta SPAD-502 pada saat 2 minggu setelah pembungaan (silking).
5. Jumlah butir biji tiap tongkol dan bobot 200 butir biji.
Pengamatan karakter jumlah daun hijau dan luasnya tiap tanaman, kadar klorofil daun, jumlah bji tiap tongkol dilakukan pada 5 contoh acak tanaman yang kompetitif.
Seleksi Famili, Persilangan antar Famili dan Evaluasi
Seleksi full-sib dan galur S
1Metode seleksi yang diplih berdasarkan efisiensi waktu pelaksanaan penelitian dan ketersediaan benih untuk keperluan seleksi. Benih yang telah dimiliki yaitu benih full-sib dan S1 yang telah dihasilkan dari kegiatan pembentukan galur S1 dan persilangan antar galur S1.
Dari percobaan evaluasi hasil persilangan pada 3 taraf pemupukan yang dilaksanakan sebelumnya, famili full-sib dan half-sib terbaik dipilih berdasarkan selisih hasil biji yang terbesar dengan varietas Bisma pada setiap taraf pemupukan. Dari setiap taraf pemupukan dipilih 10% full-sib (27 FS) dan galur S1 (18 S1) berdasarkan 10% half-sib terbaik menurut bobot bijinya. Full-half-sib dan S1 terpilih ini digunakan sebagai bahan persilangan selanjutnya untuk membentuk populasi hasil seleksi siklus pertama (C1).
Persilangan antar FS dan antar S1 terpilih pada 3 taraf pemupukan
Evaluasi hasil persilangan antar FS dan antar S
1terpilih
Hasil persilangan antar FS dan antar S1 pada 3 taraf pemupukan yang telah dibentuk sebelumnya dan varietas Bisma sebagai pembanding, selanjutnya dievaluasi pada taraf pemupukan rendah, sedang dan normal. Evaluasi dilakukan dalam rancangan acak kelompok dengan 5 ulangan. Kelima ulangan ditanam di kebun percobaan Cikeumeuh, Bogor pada waktu yang berbeda antara bulan Februari hingga Maret 2004 pada lahan bekas pertanaman jagung. Hasil analisis tanah dari 2 contoh tanah menunjukkan kandungan hara tanah tergolong rendah dengan 0,14 % N, 0,35 mg /100 g, 0,35 me K/100 g dan 0,19 % N, 0,34 mg P/100 g, 0,16 me K/100 g. Luas plot tiap perlakuan 3.75 m x 5 m. Jarak tanam 75 x 20 cm dengan 1 tanaman tiap lubang. Karakter tanaman yang diamati dan pemberian pupuk sesuai dengan perlakuan dan dengan cara pemupukan yang sama dengan percobaan evaluasi hasil persilangan antar S1 di atas.
Kombinasi perlakuan dari dua faktor yang diuji yaitu: Faktor I :Varietas/populasi terdiri dari
1. Bisma
2. Hasil persilangan FS pada pupuk rendah (FS-r) 3. Hasil persilangan FS pada pupuk sedang (FS-s) 4. Hasil persilangan FS pada pupuk normal (FS-n)
5. Hasil persilangan S1 berdasarkan HS pada pupuk rendah (S1(HS)-r) 6. Hasil persilangan S1 berdasarkan HS pada pupuk sedang (S1(HS)-s) 7. Hasil persilangan S1 berdasarkan HS pada pupuk normal (S1(HS)-n)
Faktor II: Pemupukan terdiri dari 3 taraf pemupukan 1. Pemupukan rendah
2. Pemupukan sedang 3. Pemupukan normal
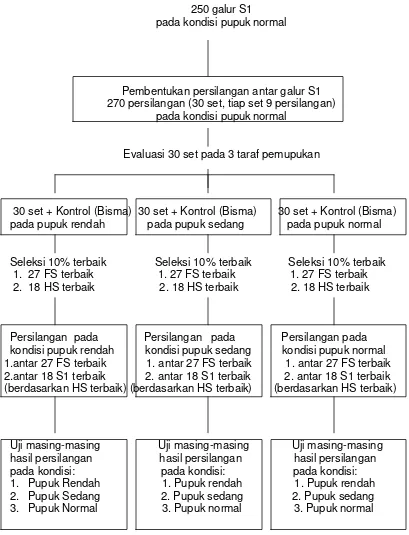
Diagram pelaksanaan kegiatan penelitian disajikan pada Gambar 1.
250 galur S1
pada kondisi pupuk normal
Pembentukan persilangan antar galur S1 270 persilangan (30 set, tiap set 9 persilangan)
pada kondisi pupuk normal
Evaluasi 30 set pada 3 taraf pemupukan
30 set + Kontrol (Bisma) 30 set + Kontrol (Bisma) 30 set + Kontrol (Bisma) pada pupuk rendah pada pupuk sedang pada pupuk normal
Seleksi 10% terbaik Seleksi 10% terbaik Seleksi 10% terbaik 1. 27 FS terbaik 1. 27 FS terbaik 1. 27 FS terbaik 2. 18 HS terbaik 2. 18 HS terbaik 2. 18 HS terbaik
Persilangan pada Persilangan pada Persilangan pada kondisi pupuk rendah kondisi pupuk sedang kondisi pupuk normal 1.antar 27 FS terbaik 1. antar 27 FS terbaik 1. antar 27 FS terbaik 2.antar 18 S1 terbaik 2. antar 18 S1 terbaik 2. antar 18 S1 terbaik (berdasarkan HS terbaik) (berdasarkan HS terbaik) (berdasarkan HS terbaik)
Uji masing-masing Uji masing-masing Uji masing-masing hasil persilangan hasil persilangan hasil persilangan pada kondisi: pada kondisi: pada kondisi: 1. Pupuk Rendah 1. Pupuk rendah 1. Pupuk rendah 2. Pupuk Sedang 2. Pupuk sedang 2. Pupuk sedang 3. Pupuk Normal 3. Pupuk normal 3. Pupuk normal
Analisis D ata
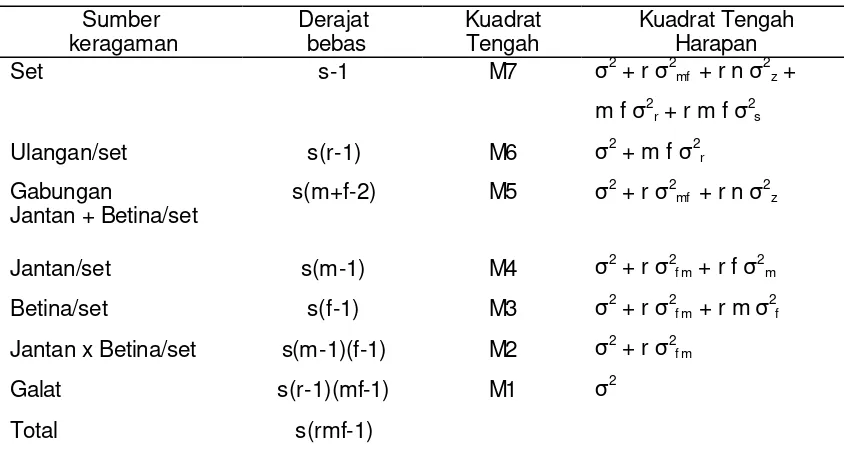
Untuk menduga ragam genetik dapat diperoleh melalui nilai kuadrat tengah dengan menggabungkan derajat bebas dan jumlah kuadrat yang berasal dari jantan dan betina (Comstock dan Robinson, 1952). Tabel analisis ragam yang akan digunakan untuk menduga besaran parameter genetik pada setiap taraf pemupukan disajikan pada Tabel 1 (Subandi, 1972).
Penduga ragam σ2z sebagai penduga peragam di antara half-sib dan σ2 mf sebagai selisih peragam full-sib dengan peragam half-sib (Hallauer dan Miranda, 1985). Dalam rancangan NCD II, tetua jantan dan betina yang digunakan merupakan individu dari F2 yang berasal dari persilangan 2 galur murni. Dalam penelitian ini tetua jantan dan betina yang digunakan adalah individu dari galur S1 yang mewakili So dari varietas Bisma. Oleh karena itu, untuk menduga ragam genetik diperlukan faktor koreksi. Penduga ragam aditif dan dominan dihitung mengikuti prosedur yang dilakukan Subandi dan Compton (1974) sebagai berikut:
σ2z = Cov. HS =¼ [ (1+Fi) + (Fj-Fi)/m1 ] σ2A dan σ2mf = Cov. FS – 2 Cov. HS
( Fj-Fi) 1 1 1 1
= [ --- ( --- - --- ) ] σ2A + --- (1+ Fi)2 σ2D + (----) (Fj-Fi) (2 + Fi + Fj)σ2D , 2 m2 m1 4 4m2
sedangkan Fi= koefisien inbreeding S0 = 0, Fj=koefisien inbreeding S1=0,5; m1= rata-rata harmonik dari banyaknya tanaman S1 untuk mewakili setiap S0 pada famili saudara tiri (half-sib), dan m2=rata-rata harmonik dari banyaknya tanaman S1 untuk mewakili So pada famili saudara kandung (full-sib). Dalam penelitian ini m1=18,6 dan m2=5,7. Dari persamaan di atas maka penduga ragam aditif dapat dihitung dari σ2z =[ ( 1 + 1/2m1)/4] σ2A. Dengan demikian penduga ragam aditif σ2A = 4 σ2z /(1 + 1/2m1). Ragam dominan diduga menurut hubungan
σ2mf = ¼ [ (1/ m2 - 1/ m1) σ2A + ( 1 + 5/4m2) σ2D ], maka penduga ragam dominan
1 1 4
σ2D = [ ( 4σ2mf - ( --- - ---- ) ( --- σ2z )] / (1 + 5/4m2) m2 m1 1+(1/2m1)
V(σi2) = -- --- , f2 df+2
Mi = kuadrat tengah yang digunakan untuk menduga komponen ragam, df = derajat bebas dari kuadrat tengah Mi,
f = koefisien komponen ragam.
Penduga ragam bagi σ2A dapat dihitung berdasarkan persamaan diatas, 4
sehingga penduga ragam σ2A = V(σ2A )= [ --- ] 2 V(σz2) dan 1+ (1/2m1)
2 Mi2 V(σz2) = --- [ Ó --- ] (k1)2 vi+2
Penduga ragam bagi σ2D = V(σ2D ) diduga mengikuti Subandi (1972) yaitu: 4 2 Mj2 (1/m2 - 1/m1)2 2 Mi2 V(σ2D ) = [--- ]2 [ ---- Ó --- + --- --- Ó --- + (1 + 5/4m2) k22 j vj + 2 ( 1 + 1/2m1)2 k12 i vi + 2 ( 1/m2 – 1/m1) 4 Mp
--- --- Ó --- ] , sedangkan ( 1+ 2m1) k1 k2 p vp + 2
Mi = kuadrat tengah ke-i yang berhubungan dengan σz2 vi = derajat bebas dari Mi
Mj = kuadrat tengah ke-j yang berhubungan dengan σ2mf vj = derajat bebas dari Mj
Mp = kuadrat tengah ke-p yang berhubungan dengan σz2 dan σ2mf vp = derajat bebas dari Mp
k1 = koefisien dari σz2 pada nilai harapan kuadrat tengah k2= koefisien dari σ2mf pada nilai harapan kuadrat tengah.
Pengujian kesamaan dua nilai ragam aditif dan dominan antar taraf pemupukan berdasarkan selang kepercayaan (1-á)100% yaitu (σ2A ± z á/2 SE(σ2A )) bagi ragam aditif dan (σ2D ± z á/2 SE(σ2D )) bagi ragam dominan, SE (σ2A ) dan SE (σ2D ) berturut-turut adalah akar kuadrat dari V(σ2A ) dan V(σ2D ).
Heritabilitas (h2) dalam arti sempit (
narrow sense) berdasarkan plot diduga dengan persamaan h2 = ó2A/ ( ó2A + ó2D+ ó2 /r) dan simpangan bakunya SEh2=SE(ó2A )/(ó2 + ó2A + ó2D) (Hallauer dan Miranda, 1985)
karakter i, dan pada seleksi langsung i=j, k= koefisien diferensial seleksi,rA(ij) =korelasi genetik karakter i dan j, óA(i) = akar ragam aditif karakter i, óA(j) = akar ragam aditif karakter j, óP(i) =akar ragam fenotip karakter i.
Korelasi genetik antar dua karakter i dan j yaitu Covz (ij)
rA (ij) = --- , dimana Covz (ij) = peragam antara karakter i dan j, (σ2z(i) σ2z(j))1/2
σ2z(i) = ragam karakter i dan σ2z(j) = ragam karakter j. Simpangan baku korelasi genetik dihitung berdasarkan Mode dan Robinson (1959).
[image:30.612.85.507.301.528.2]Tabel 1 Analisis ragam bagi rancangan persilangan NCDII untuk satu lingkungan
Sumber
keragaman Derajat bebas Kuadrat Tengah Kuadrat Tengah Harapan
Set s-1 M7 σ2 + r σ2mf + r n σ2z +
m f σ2r + r m f σ2s
Ulangan/set s(r-1) M6 σ2 + m f σ2r
Gabungan
Jantan + Betina/set s(m+f-2) M5 σ
2 + r σ2mf + r n σ2z
Jantan/set s(m-1) M4 σ2 + r σ2f m + r f σ2m Betina/set s(f-1) M3 σ2 + r σ2f m + r m σ2f Jantan x Betina/set s(m-1)(f-1) M2 σ2 + r σ2f m
Galat s(r-1)(mf-1) M1 σ2
Total s(rmf-1)
s = jumlah set persilangan; r= jumlah ulangan; m = jumlah tetua jantan; f= jumlah tetua betina; wi = jumlah tanaman contoh pada setiap plot ; n=m=f.
Percobaan di Lingkungan Seleksi
Pendugaan ragam genetik bobot biji varietas Bisma
Percobaan yang telah dilaksanakan dengan menggunakan 30 set persilangan ternyata ada 3 set persilangan yang tidak lengkap datanya. Ketidaklengkapan data ini karena jumlah tanaman yang ada dalam plot relatif sedikit (tanaman ada yang tidak tumbuh dan tongkol jagung yang hilang). Oleh karena itu, data yang dianalisis dalam rancangan persilangan NCDII dalam penelitian ini mencakup 27 set persilangan.
Untuk menentukan strategi pemuliaan yang paling sesuai diperlukan informasi besaran parameter genetik yaitu ragam genetik (aditif dan dominan) dan heritabilitas karakter kuantitatif. Besaran parameter genetik dapat digunakan untuk mendapatkan petunjuk besarnya kemajuan seleksi.
[image:31.612.89.487.331.506.2]
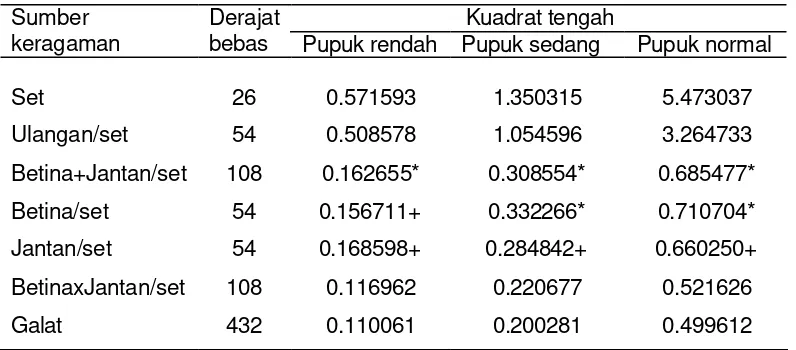
Tabel 2 Nilai kuadrat tengah bobot biji (kg/plot) pada lingkungan seleksi pemupukan rendah, sedang dan normal
Kuadrat tengah Sumber
keragaman Derajat bebas Pupuk rendah Pupuk sedang Pupuk normal
Set 26 0.571593 1.350315 5.473037
Ulangan/set 54 0.508578 1.054596 3.264733
Betina+Jantan/set 108 0.162655* 0.308554* 0.685477*
Betina/set 54 0.156711+ 0.332266* 0.710704*
Jantan/set 54 0.168598+ 0.284842+ 0.660250+
BetinaxJantan/set 108 0.116962 0.220677 0.521626
Galat 432 0.110061 0.200281 0.499612
*nyata taraf 5%, +nyata taraf 10%
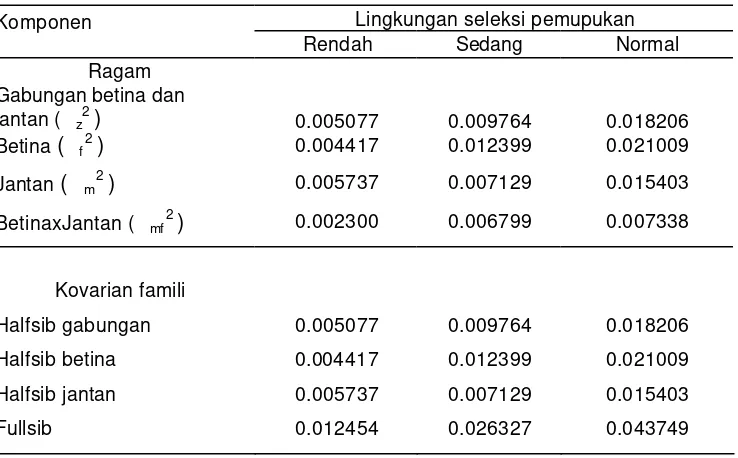
halfsib berbanding langsung dengan ragam aditif, sehingga dapat diperkirakan ragam aditif nyata. Penduga ragam interaksi betinaxjantan dan penduga ragam tetua betina dan jantan merupakan penduga kovarian fullsib menurut hubungan Cov FS= f2 + m2 + mf2 (Matter dan Jinks, 1982). Kovarian fullsib (Cov FS) terdiri dari komponen ragam aditif dan dominan, sehingga besaran penduga ragam dominan tergantung besaran ragam interaksi tetua betinaxjantan. Besaran dugaan nilai ragam tetua dan kovarian disajikan pada Tabel 3 dan dapat digunakan untuk menduga besaran ragam aditif dan dominan.
Tabel 3 Nilai penduga ragam bobot biji (kg/plot) tetua jantan, betina dan interaksinya serta kovarian famili halfsib dan fullsib pada lingkungan seleksi pemupukan rendah, sedang dan normal
Lingkungan seleksi pemupukan Komponen
Rendah Sedang Normal Ragam
Gabungan betina dan
jantan ( z2 ) 0.005077 0.009764 0.018206
Betina ( f2 ) 0.004417 0.012399 0.021009
Jantan ( m2 ) 0.005737 0.007129 0.015403
BetinaxJantan ( mf2 ) 0.002300 0.006799 0.007338
Kovarian famili
Halfsib gabungan 0.005077 0.009764 0.018206 Halfsib betina 0.004417 0.012399 0.021009 Halfsib jantan 0.005737 0.007129 0.015403 Fullsib 0.012454 0.026327 0.043749
dan normal berturut-turut 78%, 67% dan 81% dari total ragam genetik. Hasil pengujian ragam menunjukkan bahwa ragam aditif bobot biji nyata, sedangkan ragam dominan tidak nyata pada semua taraf pemupukan. Ragam aditif pada pemupukan rendah cenderung lebih kecil daripada pemupukan yang lebih tinggi. Penduga ragam dominan yang diperoleh melalui rancangan persilangan NCDII menghasilkan simpangan baku ragam dominan yang lebih besar daripada simpangan baku ragam aditif. Simpangan baku ragam dominan besar kerena penduga ragam dominan merupakan fungsi dari sejumlah nilai kuadrat tengah (Hallauer dan Miranda, 1985).
Tabel 4 Nilai dugaan ragam genetik bobot biji (kg/plot) populasi varietas Bisma (Co) pada lingkungan seleksi pemupukan rendah, sedang dan normal
Pupuk rendah Pupuk sedang Pupuk normal Parameter
genetik
Ragam Simpangan
baku Ragam Simpangan baku Ragam Simpangan baku
Ragam
aditif ( 2A) 0.019775+
(0.78) 0.011692 0.038032+ (0.67) 0.022138 0.070913* (0.81) 0.030269
Ragam dominan ( 2D)
0.005572
(0.22) 0.020969 0.018512 (0.33) 0.039359 0.017000 (0.19) 0.093265
Ragam genetik ( 2G)
0.025349
(1.00) 0.023758 0.056544 (1.00) 0.045158 0.087913 (1.00) 0.098054
*nyata taraf 5%, +nyata taraf 10%. Angka dalam tanda kurung merupakan porsi.
Ragam fenotipik bobot biji dan heritabilitas famili FS dan S1(HS)
ragam genetik dalam plot ó2wg= ó2G - Cov FS (Hallauer dan Miranda, 1985). Berdasarkan hubungan ó2w = ó2wg+ ó2We, maka ragam lingkungan dalam plot ó2We = ó2w - (ó2G – Cov FS). Nilai ó2, ó2G dan Cov FS tiap tanaman dapat diperoleh dari hasil analisis data plot yang dibagi dengan jumlah tanaman tiap plot. Besaran nilai penduga komponen ragam dalam dan antar plot dari bobot biji disajikan pada Tabel 5. Dari data yang diperoleh menunjukkan ragam antar tanaman dalam plot sebagian besar disebabkan oleh ragam lingkungan mikro pada ketiga taraf pemupukan, sedangkan ragam genetik antar tanaman dalam plot kecil.
Ragam fenotipik bobot biji untuk FS diduga dengan
ó2P = ½ ó2A + ¼ ó2D + [ó2pl + (½ ó2A+ ¾ ó2D + ó2We)/ w] /r, sedangkan ragam fenotipik famili HS dari S1 yaitu
ó2P= ¼ ó2A + ¼f (ó2A + ó2D) + [ó2pl +{ (¾ - ¼f) ó2A+ (1- ¼f) ó2D + ó2We}/w]/r (Subandi dan Compton, 1974). Ilustrasi perhitungan ragam ó2We dan ó2P disajikan pada Lampiran 23. Nilai dugaan ragam fenotipik famili FS dan S1(HS) menunjukkan semakin banyak pupuk yang diberikan cenderung semakin besar ragam fenotipiknya (Tabel 6). Ragam fenotipik merupakan komponen dalam perhitungan pendugaan kemajuan seleksi yang berbanding terbalik dengan kemajuan seleksi, sehingga makin besar ragam fenotipik semakin kecil kemajuan seleksi yang akan diperoleh.
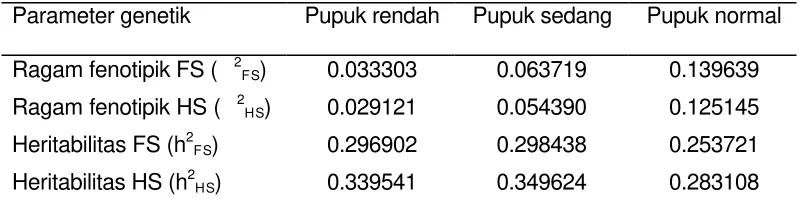
Heritabilitas (h2) suatu karakter merupakan besaran yang menunjukkan karakter tersebut dapat diwariskan ke keturunannya, yang merupakan porsi dari total keragaman fenotipe yang disebabkan oleh faktor genetik. Oleh karena itu, keberhasilan seleksi dapat dicerminkan oleh besaran heritabilitas. Heritabilitas famili FS dan S1(HS) merupakan rasio ragam aditif dengan ragam fenotipiknya (½ó2A/ó2P) dan hasil nilai dugaan dari penelitian ini disajikan pada Tabel 6. Heritabilitas pada pemupukan rendah dan sedang cenderung lebih tinggi dari pada heritabilitas pada pemupukan normal. Di samping itu, heritabilitas HS lebih tinggi daripada heritabilitas FS.
Pemupukan
Galat percobaan
ó2 Antar plot ó2pl
Dalam plot ó2W/w
Lingkungan mikro dalam plot
ó2We/w
g/tanaman
Rendah 275.15 174.00 101.15 99.51
Sedang 500.70 377.09 123.61 119.74
[image:35.612.88.483.73.193.2]Normal 1249.09 1146.76 102.33 96.72
Tabel 6 Nilai dugaan ragam fenotipik dan heritabilitas famili fullsib dan halfsib bobot biji (kg/plot) populasi varietas Bisma (Co) pada lingkungan seleksi pemupukan rendah, sedang dan normal
Parameter genetik Pupuk rendah Pupuk sedang Pupuk normal Ragam fenotipik FS ( 2FS) 0.033303 0.063719 0.139639 Ragam fenotipik HS ( 2HS) 0.029121 0.054390 0.125145 Heritabilitas FS (h2FS) 0.296902 0.298438 0.253721 Heritabilitas HS (h2HS) 0.339541 0.349624 0.283108
FS=fullsib, HS=halfsib
Seleksi famili terbaik
Untuk mengetahui kemajuan seleksi maka dipilih bobot biji famili FS dan S1 berdasarkan penampilan HS 10% terbaik. Seperti telah dinyatakan sebelumnya, dasar pemilihan famili FS dan HS dengan pertimbangan ketersediaan benih yang telah dimiliki. Famili-famili yang terpilih disajikan pada Lampiran 11 dan Lampiran 12, sedangkan besaran diferensial seleksi pada setiap famili dan lingkungan seleksi disajikan pada Tabel 7. Berdasarkan famili-famili yang terpilih pada pemupukan rendah dan sedang terdapat 4 famili FS dan 1 galur S1 yang sama. Pada pemupukan sedang dan normal terdapat 3 famili FS dan 3 galur S1 yang sama, sedangkan pada pemupukan rendah dan normal diperoleh 1 famili FS dan 1 galur S1 yang sama. Di samping itu, terdapat 1 famili FS dan 1 galur S1 yang sama terpilih pada ketiga taraf pemupukan.
Tabel 7 Rata-rata famili FS dan HS serta diferensial seleksi pada lingkungan seleksi pemupukan rendah, sedang dan normal
[image:35.612.84.484.281.384.2]famili (kg/plot) terpilih (kg/plot) (kg/plot)
Pupuk rendah
FS 1.0342 1.4609 0.4267
HS 1.0342 1.2941 0.2599
Pupuk sedang
FS 1.4615 2.0064 0.5449
HS 1.4615 1.7444 0.2829
Pupuk normal
FS 2.2346 2.9578 0.7232
HS 2.2346 2.6740 0.4394
FS=fullsib, HS=halfsib
Korelasi genetik bobot biji antar lingkungan pemupukan
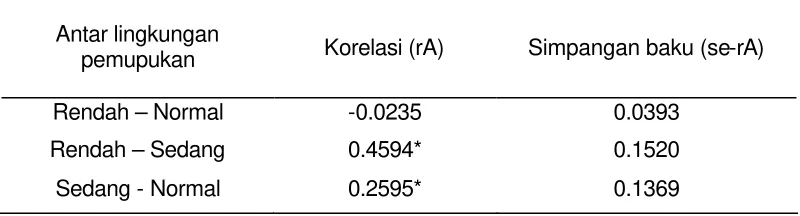
[image:36.612.84.484.472.580.2]Dalam pendugaan kemajuan seleksi tak langsung memerlukan informasi besaran korelasi genetik. Besaran korelasi genetik bobot biji antar taraf pemupukan sebagai komponen pendugaan kemajuan seleksi tak langsung. Korelasi bobot biji antar lingkungan pemupukan disajikan pada Tabel 8. Korelasi bobot biji yang nyata diperoleh antar lingkungan pemupukan rendah-sedang dan sedang-normal, sedangkan antara lingkungan pemupukan rendah dan normal tidak nyata. Korelasi genetik bobot biji antar dua kondisi pemupukan cenderung semakin rendah dengan semakin besarnya perbedaan taraf pemupukan.
Tabel 8 Korelasi genetik bobot biji antar lingkungan seleksi pemupukan
Antar lingkungan
pemupukan Korelasi (rA) Simpangan baku (se-rA)
Rendah – Normal -0.0235 0.0393
Rendah – Sedang 0.4594* 0.1520
Sedang - Normal 0.2595* 0.1369
* nyata taraf uji 5%
Pendugaan ragam genetik dan heritabilitas karakter sekunder
Hasil analisis menunjukkan bahwa ragam aditif semua karakter sekunder nyata, kecuali untuk karakter jumlah biji tiap tongkol.
Tabel 9 Ragam aditif dan dominan karakter sekunder pada lingkungan seleksi pemupukan rendah, sedang dan normal
Lingkungan seleksi pemupukan
Rendah Sedang Normal Karakter Ragam
Ragam SE Ragam SE Ragam SE aditif 1.3411* 0.2618 0.6714* 0.1498 0.6091* 0.1346 ASI (hari)
dominan 0.1649 0.2699 0.1755 0.1739 0.1119 0.1561
aditif 5.8221* 2.6856 10.1475* 3.4402 9.1041* 3.0075 Bobot
200 butir
(g) dominan 2.9111 4.5025 3.9833 5.1777 1.0242 4.5388
aditif 4.4416* 1.8288 3.2179* 1.5940 3.9128* 1.6346 Klorofil
dominan 0 3.0244 0.3475 2.7611 1.5644 2.6669
aditif 0.1493* 0.0417 0.0478* 0.0284 0.0795* 0.0310 Jumlah
daun tiap tanaman saat berbunga
dominan 0.0418 0.0571 0.0079 0.0511 0.0473 0.0491
aditif 0.0258* 0.0124 0.0335* 0.0114 0.0343* 0.0112 LAI saat
berbunga dominan 0.0276 0.0206 0.0021 0.0175 0 0.0169
aditif 0.1116 0.1132 0.1516+ 0.0840 0.1149 0.1137 Jumlah
daun tiap tanaman saat panen
dominan 0.8851* 0.2022 0.5568* 0.1382 0.9266* 0.2022
aditif 0.0254* 0.0107 0.0364* 0.0139 0.0413* 0.0255 LAI saat
panen dominan 0.0510* 0.0167 0.0688* 0.0209 0.1454* 0.0431
aditif 0.0021 0.0012 0.0018+ 0.0009 0.0041* 0.0019 Porsi
daun
menua dominan 0.0079* 0.0020 0.0051* 0.0016 0.0084* 0.0032 aditif 380.85 290.22 324.69 232.33 397.74 268.29 dominan 0 547.91 0 435.70 0 497.33 Jumlah
biji tiap tongkol
*nyata taraf 5%, +nyata taraf 10%. ASI=anthesis silking inteval. LAI=leafarea index
lebih besar daripada ragam aditif. Hasil pengujian selanjutnya menunjukkan bahwa ragam aditif semua karakter tidak berbeda nyata antar taraf pemupukan.
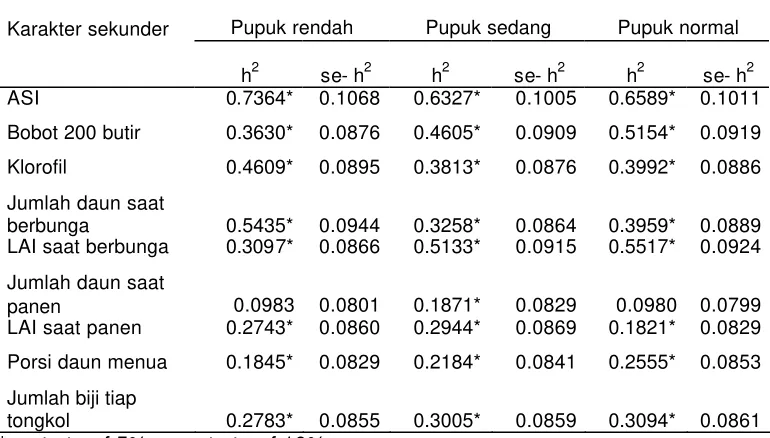
Karakter sekunder yang relatif besar heritabilitasnya dibandingkan dengan karakter sekunder lainnya adalah ASI. Heritabilitas ASI pada pemupukan rendah, sedang dan normal berturut-turut 0,7364, 0.6323 dan 0,6589 (Tabel 10).
Heritabilitas ASI yang diperoleh lebih tinggi daripada heritabilitas bobot biji yang memiliki heritabilitas h2 < 0,35 baik pada lingkungan pemupukan rendah, sedang maupun normal. Heritabilitas karakter sekunder semua nyata, kecuali pada jumlah daun saat panen pada pemupukan rendah dan normal.
[image:38.612.91.476.430.649.2]Hasil pengujian heritabiltas antar taraf pemupukan tidak menunjukkan perbedaan yang nyata. Apabila karakter sekunder memiliki korelasi genetik tinggi dengan karakter primer dan memiliki heritabilitas lebih tinggi daripada karakter primer maka seleksi berdasarkan karakter sekunder tersebut akan lebih efisien. Dengan demikian karakter ASI dapat dipertimbangkan sebagai karakter sekunder dalam proses seleksi baik pada kondisi pemupukan rendah, sedang maupun normal. Namun perlu dipertimbangkan besaran korelasi genetik karakter sekunder tersebut dengan bobot biji.
Tabel 10 Hertabilitas karakter sekunder pada lingkungan seleksi pemupukan rendah, sedang dan normal
Karakter sekunder Pupuk rendah Pupuk sedang Pupuk normal
h2 se- h2 h2 se- h2 h2 se- h2 ASI 0.7364* 0.1068 0.6327* 0.1005 0.6589* 0.1011 Bobot 200 butir 0.3630* 0.0876 0.4605* 0.0909 0.5154* 0.0919 Klorofil 0.4609* 0.0895 0.3813* 0.0876 0.3992* 0.0886 Jumlah daun saat
berbunga 0.5435* 0.0944 0.3258* 0.0864 0.3959* 0.0889 LAI saat berbunga 0.3097* 0.0866 0.5133* 0.0915 0.5517* 0.0924 Jumlah daun saat
panen 0.0983 0.0801 0.1871* 0.0829 0.0980 0.0799 LAI saat panen 0.2743* 0.0860 0.2944* 0.0869 0.1821* 0.0829 Porsi daun menua 0.1845* 0.0829 0.2184* 0.0841 0.2555* 0.0853 Jumlah biji tiap
tongkol 0.2783* 0.0855 0.3005* 0.0859 0.3094* 0.0861
Korelasi genetik bobot biji dengan karakter sekunder
Karakter sekunder dapat digunakan sebagai kriteria seleksi untuk meningkatkan kemajuan seleksi bobot biji, jika memiliki korelasi genetik dengan bobot biji tinggi dan memiliki heritabilitas yang lebih tinggi dibandingkan dengan bobot biji. Besaran korelasi genetik antara karakter sekunder dengan bobot biji pada lingkungan pemupukan yang sama dapat digunakan untuk menduga respon terkorelasi karakter sekunder pada lingkungan pemupukan yang sama. Korelasi genetik bobot biji pada lingkungan pemupukan tertentu dengan karakter sekunder pada lingkungan pemupukan yang lain dapat digunakan untuk menduga respon terkorelasi karakter sekunder akibat seleksi bobot biji pada lingkungan pemupukan tertentu. Besaran korelasi genetik bobot biji dengan karakter sekunder pada lingkungan pemupukan yang sama atau berbeda disajikan pada Tabel 11.
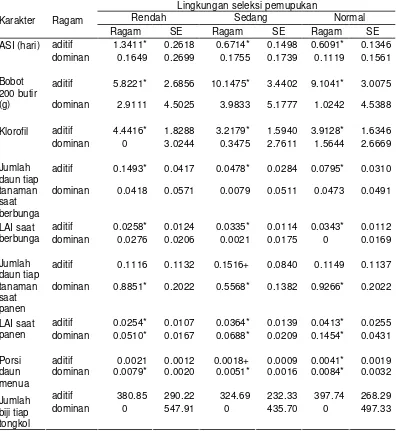
Korelasi genetik bobot biji pada lingkungan pemupukan rendah dengan semua karakter sekunder yang diamati pada pemupukan rendah (rR-R) nyata, kecuali jumlah daun saat panen. Korelasi genetik bobot biji pada lingkungan pemupukan rendah dengan semua karakter sekunder pada lingkungan pemupukan sedang (rR-S) nyata, kecuali jumlah daun saat pembungaan.
Selanjutnya, korelasi genetik bobot biji pada lingkungan pemupukan rendah dengan semua karakter sekunder yang diamati pada lingkungan pemupukan normal (rR-N) nyata, kecuali jumlah daun dan LAI saat panen serta porsi daun menua. Berdasarkan koefisien korelasi yang diperoleh, terdapat perbedaan arah korelasi dari karakter ASI dan klorofil. Korelasi genetik bobot biji dengan ASI diperoleh rR-R dan rR-S negatif, tetapi korelasi rR-N diperoleh positif. Korelasi genetik bobot biji dengan klorofil diperoleh rR-R dan rR-S posittif, tetapi korelasi rR-N diperoleh negatif.
Korelasi genetik bobot biji pada lingkungan pemupukan sedang dengan karakter sekunder pada lingkungan pemupukan rendah, sedang dan normal umumnya memberikan arah yang sama. Korelasi genetik jumlah daun saat pembungaan pada pemupukan rendah dan normal dengan bobot biji pada pemupukan sedang (rS-R dan rS-N) tidak nyata. Korelasi genetik LAI saat pembungaan dan jumlah daun saat panen pada pemupukan rendah dengan bobot biji pada pemupukan sedang tidak nyata.
Bobot biji pada lingkungan seleksi
pemupukan
Karakter sekunder pada lingkungan seleksi pemupukan
Rendah Sedang Normal r se-r r se-r r se-r
Rendah
ASI -0.2186* 0.0743 -0.0640* 0.0257 0.4970* 0.1608 Bobot 200 butir 0.4233* 0.1690 0.3068* 0.1084 0.3600* 0.1301 Klorofil 0.1470* 0.0421 0.4137* 0.1518 -0.2365* 0.1019 Jumlah daun
saat berbunga 0.1485* 0.0567 0.0697 0.0555 0.4000* 0.1685 LAI saat
berbunga 0.4307* 0.1662 0.3635* 0.1395 0.2385* 0.0891 Jumlah daun
saat panen 0.2329 0.1663 0.1627* 0.0698 0.1148 0.0710 LAI saat panen 0.3879* 0.1456 0.2573* 0.1001 0.0515 0.0470 Porsi daun
menua -0.3113* 0.1457 -0.1581* 0.0687 0.0300 0.0271 Jumlah biji tiap
tongkol 0.5114* 0.2287 0.1741+ 0.0959 0.3257+ 0.1891
Sedang
ASI -0.2073* 0.0710 -0.2491* 0.0888 -0.1401* 0.0413 Bobot 200 butir 0.3727* 0.1281 0.3618* 0.1135 0.4166* 0.1366 Klorofil -0.2660* 0.1318 -0.0683 0.0947 -0.1073* 0.0490 Jumlah daun
saat berbunga -0.0540 0.0352 0.1929* 0.0876 0.0559 0.0396 LAI saat
berbunga 0.0206 0.0191 0.1878* 0.0606 0.2082* 0.0731 Jumlah daun
saat panen 0.0290 0.0378 0.1422* 0.0518 -0.0205 0.0433 LAI saat panen 0.2516* 0.0798 0.2200* 0.0742 0.3480* 0.1629 Porsi daun
menua -0.5954* 0.2813 -0.1765* 0.0669 -0.2603* 0.1120 Jumlah biji tiap
tongkol 0.1285* 0.0331 0.6433* 0.3312 0.0837 0.0571
Normal
ASI 0.1610* 0.0561 -0.4865* 0.1915 -0.1312* 0.0487 Bobot 200 butir 0.2506* 0.1086 0.6329* 0.2517 0.2994+ 0.1794 Klorofil 0.0222 0.0731 0.3244* 0.1362 0.2528* 0.1013 Jumlah daun
saat berbunga -0.2481* 0.1000 -0.4156* 0.1728 0.0055 0.0293 LAI saat
berbunga -0.2068 0.0961 0.9657* 0.4013 0.1844* 0.0756 Jumlah daun
saat panen 0.1700 0.1324 0.4521+ 0.2572 0.1611 0.1241 LAI saat panen 0.1046 0.0757 0.8740* 0.3966 0.1636* 0.0678 Porsi daun
menua -0.3884 0.2418 -0.7257+ 0.3735 -0.0463 0.0305 Jumlah biji tiap
Korelasi bobot biji pada lingkungan pemupukan normal dengan semua karakter sekunder yang diamati pada lingkungan pemupukan yang sama (normal) nyata, kecuali jumlah daun saat pembungaan dan saat panen serta jumlah biji tiap tongkol. Seperti pada korelasi bobot biji pada lingkungan pemupukan rendah dengan karakter ASI pada pemupukan normal, diperoleh perbedaan arah korelasi. Korelasi genetik bobot biji dengan ASI diperoleh rN-R positif dan rN-S dan rN-N negatif.
Korelasi bobot biji pada pemupukan normal nyata dengan klorofil pada lingkungan pemupukan sedang dan normal, tetapi tidak nyata pada pemupukan rendah. Di samping itu, korelasi bobot biji pada lingkungan pemupukan normal berkorelasi negatif dengan jumlah biji tiap tongkol pada lingkungan pemupukan rendah.
Percobaan di Lingkungan Evaluasi
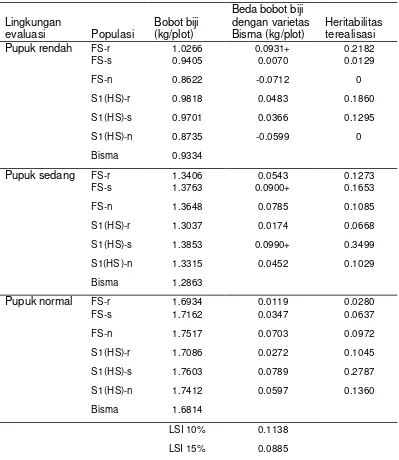
Hasil analisis ragam bobot biji menunjukkan adanya perbedaan yang nyata di antara populasi yang dievaluasi. Rata-rata hasil pengamatan karakter tanaman jagung pada lingkungan evaluasi disajikan pada Tabel 12-14. Pada lingkungan evaluasi pupuk rendah, bobot biji populasi FS-r yang diseleksi pada lingkungan pemupukan rendah cenderung lebih tinggi daripada bobot biji populasi varietas Bisma sebagai populasi dasar. Bobot biji populasi FS-s dan S1(HS)-s hasil seleksi pada lingkungan pemupukan sedang juga cenderung lebih tinggi dari pada varietas Bisma yang dievaluasi pada pemupukan yang sama. Rata-rata bobot biji pada populasi FS dan S1(HS) tidak menunjukkan perbedaan yang nyata
Heritablitas terealisasi bobot biji (realized heritability) diduga dengan h2 = R/S, dimana R=respon nyata akibat seleksi dan S=diferensial seleksi= selisih antara nilai rata-rata famili terseleksi dengan rata-rata populasi yang diseleksi (Falconer dan Mackay, 1996). Nilai heritabilitas terealisasi bobot biji yang diperoleh disajikan pada Tabel 12. Pada lingkungan evaluasi pemupukan rendah, populasi FS-r dan S1(HS)-r lebih tinggi daripada populasi lainnya. Demikian juga, populasi FS-s dan S1(HS)-s pada lingkungan evaluasi pemupukan sedang.
saat panen populasi FS-s nyata lebih banyak daripada populasi Bisma. Pada kondisi pemupukan normal, semua karakter sekunder menunjukkan tidak ada perbedaan yang nyata bila dibandingkan dengan populasi Bisma.
Tabel 12 Bobot biji hasil persilangan famili terpilih pada pemupukan rendah, sedang dan normal
Lingkungan
evaluasi Populasi Bobot biji (kg/plot)
Beda bobot biji dengan varietas
Bisma (kg/plot) Heritabilitas terealisasi
Pupuk rendah FS-r 1.0266 0.0931+ 0.2182
FS-s 0.9405 0.0070 0.0129
FS-n 0.8622 -0.0712 0
S1(HS)-r 0.9818 0.0483 0.1860 S1(HS)-s 0.9701 0.0366 0.1295 S1(HS)-n 0.8735 -0.0599 0
Bisma 0.9334
Pupuk sedang FS-r 1.3406 0.0543 0.1273
FS-s 1.3763 0.0900+ 0.1653
FS-n 1.3648 0.0785 0.1085 S1(HS)-r 1.3037 0.0174 0.0668 S1(HS)-s 1.3853 0.0990+ 0.3499
S1(HS )-n 1.3315 0.0452 0.1029 Bisma 1.2863
Pupuk normal FS-r 1.6934 0.0119 0.0280
FS-s 1.7162 0.0347 0.0637 FS-n 1.7517 0.0703 0.0972 S1(HS)-r 1.7086 0.0272 0.1045
S1(HS)-s 1.7603 0.0789 0.2787 S1(HS)-n 1.7412 0.0597 0.1360
Bisma 1.6814
LSI 10% 0.1138 LSI 15% 0.0885
+ nyata pada taraf 15%
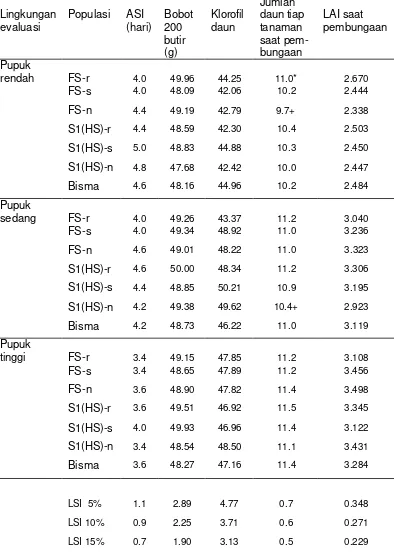
Lingkungan
evaluasi Populasi ASI (hari) Bobot 200 butir (g)
Klorofil daun
Jumlah daun tiap tanaman saat pem-bungaan
LAI saat pembungaan
Pupuk
rendah FS-r 4.0 49.96 44.25 11.0* 2.670
FS-s 4.0 48.09 42.06 10.2 2.444
FS-n 4.4 49.19 42.79 9.7+ 2.338
S1(HS)-r 4.4 48.59 42.30 10.4 2.503
S1(HS)-s 5.0 48.83 44.88 10.3 2.450
S1(HS)-n 4.8 47.68 42.42 10.0 2.447
Bisma 4.6 48.16 44.96 10.2 2.484
Pupuk
sedang FS-r 4.0 49.26 43.37 11.2 3.040
FS-s 4.0 49.34 48.92 11.0 3.236
FS-n 4.6 49.01 48.22 11.0 3.323
S1(HS)-r 4.6 50.00 48.34 11.2 3.306
S1(HS)-s 4.4 48.85 50.21 10.9 3.195
S1(HS)-n 4.2 49.38 49.62 10.4+ 2.923
Bisma 4.2 48.73 46.22 11.0 3.119
Pupuk
tinggi FS-r 3.4 49.15 47.85 11.2 3.108
FS-s 3.4 48.65 47.89 11.2 3.456
FS-n 3.6 48.90 47.82 11.4 3.498
S1(HS)-r 3.6 49.51 46.92 11.5 3.345
S1(HS)-s 4.0 49.93 46.96 11.4 3.122
S1(HS)-n 3.4 48.54 48.50 11.1 3.431
Bisma 3.6 48.27 47.16 11.4 3.284
LSI 5% 1.1 2.89 4.77 0.7 0.348 LSI 10% 0.9 2.25 3.71 0.6 0.271 LSI 15% 0.7 1.90 3.13 0.5 0.229
[image:43.612.90.484.77.630.2]*nyata pada taraf 5%, + nyata pada taraf 10%.ASI=anthesis silking inteval. LAI=leafarea index
biji tiap tongkol pada pemupukan rendah, sedang dan tinggi di lingkungan evaluasi
Lingkungan
evaluasi Populasi
Jumlah daun tiap tanaman saat panen
LAI saat
panen Porsi daun menua
Jumlah biji tiap tongkol Pupuk
rendah FS-r 4.48 1.094 0.588 331.1
FS-s 4.36 0.909 0.622 327.6
FS-n 4.32 0.896 0.621 270.5+
S1(HS)-r 4.32 0.906 0.630 317.9
S1(HS)-s 4.76 1.031 0.577 329.8
S1(HS)-n 3.64 0.772 0.688 314.0
Bisma 4.20 0.919 0.638 318.4
Pupuk
sedang FS-r 4.84 1.268 0.587 343.0
FS-s 5.00+ 1.400 0.576 338.8
FS-n 4.88 1.370 0.589 352.0
S1(HS)-r 3.96 1.166 0.648 344.3
S1(HS)-s 4.56 1.258 0.615 346.9
S1(HS)-n 3.76 1.001 0.657 349.4
Bisma 4.32 1.204 0.621 337.8
Pupuk
tinggi FS-r 5.00 1.324 0.576 345.8
FS-s 5.00 1.449 0.577 345.3
FS-n 5.56 1.639 0.536 352.8
S1(HS)-r 5.08 1.497 0.557 365.7
S1(HS)-s 4.84 1.315 0.575 361.5
S1(HS)-n 4.76 1.394 0.591 345.2
Bisma 5.16 1.489 0.547 346.9
LSI 5% 0.91 0.358 0.095 48.8 LSI 10% 0.70 0.279 0.074 37.9 LSI 15% 0.59 0.235 0.062 32.0
PEMBAHASAN
Hasil pendugaan ragam genetik yang telah diperoleh tentu tidak terlepas dari bias, karena penelitian ini dilakukan hanya pada satu lingkungan (satu lokasi dan satu musim). Di samping itu, ulangan dalam percobaan tidak dapat dilaksanakan pada waktu yang bersamaan, sehingga hasil percobaan mungkin masih dipengaruhi oleh waktu tanam