PRODUKSI DAN INAKTIVASI IN
VITRO
TOKSIN ISOLAT
Corynespora cassiicola
(Berk.
&
Curt.) Wei
ASAL DAUN KARET
OLEH:
SUWARTO
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGORSUWARTO. Produksi dan Inaktivasi in Vitro Toksin Isolat Corynespora cassiicola (Berk.&Curt.) Wei Asal Daun Karet. Di bawah bimbingan MEITY SURADJI SMAGA, RUSMILAH SUSENO, SRI HENDRASTUTI HIDAYAT, ASRIL DARUSSAMM dan SISWANTO.
ABSTRAK
Penyakit Gugur Daun Corynespora pada tanaman karet yang disebabkan oleh Corynespora cassiicola [Berk. & Curt.] Wei telah menjadi ancaman bagi produksi karet di Indonesia. Cendawan patogenik tersebut menghasilkan toksin yang dinamakan cassiico!ine yang berperan penting dalam patogenisitasnya. Hingga saat ini masih belum diketahui strategi pengendalian yang efektif dan efisien untuk nlencegah epidemi penyakit tersebut. Penelitian ini bertujuar? (1) menganalisis potensi produksi toksin irt vitro isolat-isolat C . cassiicola dari inang karet dan non karet, (2) menganslisis faktor-faktor yang mempengaruhi aktivitas biologi toksin dalam kaitannya dengan mekanisme pertahanan tanaman terhadap penyakit tersebut, dan (3) mengevaluasi potensi senyawa-senyawa pereduksi untuk menginaktifkan cassiicoline. Fitotoksisitas toksin diuji dengan cara baru yang dinamakan metode bioasai in planta (BiP) sebagai modifikasi dari metode bioasai derigan cara pencelupan pangkal daun. Cara baru ini sangat sederhana dan efisien untuk mengevaluasi toksin dan bahan-bahan kimia lain dengan cara mengaplikasikan bahan langsung in situ pada daun karet tanpa dipetik. Untuk mengetahui pembentukan toksin, enam isolat C. cassiicola yang berasal dari lima klon karet yang berbeda respons resistensinya, tiga isolat C. cassiicola dari pepaya dan satu isolat Cercosporidium henningsii dari ubikayu ditumbuhkan pada medium alternaria yang dimodifikasi (modified alternaria mediumlMAM). Studi menunjukkan bahwa tidak semua isolat C. cassiicola asal karet menghasilkan toksin pada kondisi in vitro. Isolat Cc1~~873~~0400 dari klon moderat rentan IAN 873 me~ghasilkan toksin yang sangat aktif. Walaupun demikian, toksin hampir tidak dihasilkan oleh isolat-isolat C ~ ~ ~ 1 ~ 1 0 3 ~ ~ 0 4 0 0 dan C ~ 1 1 ~ 1 ~ 7 2 5 ~ ~ 0 4 0 0 asal klon rentan RRIC 103 dan RRIM 725, dan isolat C C G T I S U O ~ O ~ dan CCRRIM 600SU0702 asal klon moderat rentan GT 1 dan RRIM 600. Toksin tidak dihasilkan oleh isolat-isolat dari
pepaya dan isolat C. henningsii dari ubikayu. Aktivitas toksin C C ~ ~ . ~ ~ ~ ~ ~ ~ ~ ~ ~ ~ terdeteksi di dalam filtrat kultur pada medium MAM mulai pada empat hari setelah
menginaktifkan toksin. Namun demikian, senyawa antitoksin tersebut belum diidentifikasi lebih spesifik dan disarankan untuk diverifikasi lebih lanjut.
SUWARTO. Production and Inactivation in Vitro of Toxin of Corynespora cassiicola (Berk.&Curt.) Wei from Hevea Leaf. Supervised by MEITY SURADJI SINAGA, RUSMILAH SUSENO, SRI HENDRASTUTI HIDAYAT, ASRIL DARUSSAMIN and SIS WANTO.
ABSTRACT
The Corynespora leaf fall disease on rubber caused by Corynespora cassiicola (Berk. & Curt.) Wei is a serious threat for rubber production in Indonesia. The fungus produces toxin called cassiicoline which play a role in pathogenicity of the disease. Up to now an effective and efficient strategy to control the epidemic of the disease has not been obtained yet. The objectives of this study are (1) to evaluate capability of isolates of C. cassiicola from rubber and non rubber hosts to produce toxin in vitro, and (2) to analyze factors that affecting the biological activity of toxin in relation with the plant defense to the disease, and (3) to evaluate the effectivity of several reducing agents to inactivate cassiicoline. Phytotoxicity of the toxin was determined by using a modified bioassay procedure namely bioassay in planta (BiP) to substitute test by using leaf cuttings. This new method was simple and efficient to evaluate phytotoxicity of toxin or other chemical substances that applied directly to rubber leaf. The C. cassiicola isolated from differential rubber clones and from papaya and Cercosporidium henningsii isolated from cassava were grown in the modified alternaria medium (MAM) to detect the toxin activation. It was found that the isolate of C ~ 1 ~ ~ 8 7 3 ~ ~ 0 4 0 0 isolated from IAN 873, a moderately susceptible clone, produce a very active toxin. However, isolates of C C R R I C ~ O ~ S U O ~ O O and C~RR1~725~~0400, isolated from susceptible clones RRIC 103 and RRIM 725, and isolates of C C G T I S U O ~ O ~ and c ~ R ~ 1 ~ 6 0 0 ~ ~ 0 7 0 2 , isolated from the moderately susceptible clones GT 1 and RRIM 600, produce a low toxin. The activity of the toxin of Cc1~~873~~0400 in MAM was detected four days after incubation and reached the peaks after 14 days with concentration about 1 pg proteinlml. The t ~ x i n is thermo stable. The activity was retained when the toxin solution was heated up to 100 O C for 30 min, however. it will complete!^ lost at 120 O C for 10 min. It was showed that certain reducing agent could inactivate cassiicoline. The phytotoxicity of the toxin was totally blocked by sodium dithionite (Na2S204) and strongly affected by 2-mercaptoethanol, methylene blue or amido black, but not affected by ascorbic or citric acids. The toxin was inactivated by B-serum of IAN 873 latex, however, surprisingly the activity was not affected by B-serum of resistant clones such as AVROS 2037 and BPM 1. The phenomenon was an indication that there was a substance in B-serum among rubber clones that could inactivate cassiicoline, although, it was not specifically identified yet. Therefore, hrther studies to verify the antitoxin substance have to be carried out.
SURATPERNYATAAN
Saya menyatakan dengan sebenar-benamya bahwa segala pernyataan dalam disertasi saya yang berjudul:
"Produksi dan Inaktivasi in Viiro Toksin Isolat Corynespora cassiicola (Berk & Curt.) Wei Asal Daun Karet"
merupakan gagasan atau hasil penelitian disertasi saya sendiri, dengan pembimbingan para Kornisi Pembimbing, kecuali yang dengan jelas ditunjukkan rujukannya. Disertasi ini belum pernah diajukan untuk memperoleh gelar pada progranl sejenis di perguruan tinggi lain.
Semua data dan infornlasi yang digunakan telah dinyatakan secara jelas dan dapat diperiksa kebenarannya.
Bogor, Desember 2003
PRODUKSI DAN INAKTIVASI IN
U T R O
TOKSIN ISOLAT
Corynespora cassiicola
(Berk&Curt.) Wei
ASAL DAUN KARET
Oleh:
SUWARTO
Disertasi
Sebagai salah satu syarat untuk memperoleh gelar Doktor pada
Program Studi Entomologi-Fitopatologi
SEKOLAH PASCASARJANA
PRODUKSI DAN INAKTIVASI
INVITRO
TOKSIN ISOLAT
Corynespora cassiicola
(BerkdkCurt.) Wei
ASAL DAUN KARET
Oleh:
SUWARTO
Disertasi
Sebagai salah satu syarat untuk memperoleh gelar Doktor pada
Program Studi Entomologi-Fitopatologi
SEKOLAH PASCASARJANA
Judul Disertasi : Produksi dan Inaktivasi in Vitro Toksin Isolat Corynespora cassiicola (Berk. & Curt.) Wei Asal Daun Karet
Nama Mahasiswa : Suwarto
Nomor Pokok : 975037lFIT
Program Studi : Entomologi-Fitopatologi
1. K o ~ i s i Pembimbing
Dr. Ir. ~ e i t 4 s u r a u Sinaea. MSc.
n I Ketua
Prof. Dr. Ir. Rusmilah Suseno, MSc. Dr. Ir. Sri Hendrastuti Hidayat, MSc.
Anggota Anggota
*
.
Dr. Sis anto DEA Anggota
Dr. Asril arussamin MS
I!!!-
Mengetahui:
2. Ketua Program Studi
&u&&
Dr. Ir. Sri Hendrastuti Hidayat, MSc. Manuwoto, MSc.
RIWAYAT HIDUP
Penulis dilahirkan di Desa Bendungan, Kabupaten Kulon Progo pada tanggal 15 November 1957 sebagai anak kedua diantara enam bersaudara dari pasangan
Hadisumarto dan Ratjik. Penulis menikah dengan Ir. Ermina Muhayati tahun 1985 dan dikaruniai lima orang anak, yaitu Radityo Andi Dharma, Putri Widha Sari &
Putri Wika Sari (kembar), Rofif Tyo Zaidan Fajar dan Farhan Tyo Zahid Akbar.
Pendidikan sarjana ditempuh di Jurusan Ilmu Hama dan Penyakit Tumbuhan,
Fakultas Pertanian IPB, lulus pada tahun 1981. Pada tahun 1984, penulis diterima
di Program Studi Fitopatologi pada Fakultas Pascasarjana IPB untuk mengikuti
pendidikan S2 dan menamatkannya pada tahun 1988. Kesempatan untuk melanjutkan ke program doktor pada program studi yang sama aiperoleh pada tahun 1997. Biaya pendidikan pascasarjana diperoleh dari Balai Penelitian Sungei
Putih, Pusat Penelitian Karet. Lembaga Riset Perkebunan Indonesia.
Penulis bekerja di Balai Penelitian Sungei Putih, Pusat Penelitian Karet dan
ditempatkan di Desa Sungei Putih, Kecamatan Galang, Kabupaten Deli Serdang,
Provinsi Sumatera Utara sejak tahun 1982 sampai sekarang. Bidang penelitian yang menjadi tanggung jawab peneliti ialah perlindungan tanaman khususnya
tanaman karet. Penulis aktif mengikuti seminar, lokakarya, konferensi, simposium
dan konggres ilmiah baik sebagai pemrasaran maupun sebagai peserta. Di bidang
administrasi, penulis pernah berperlgalaman sebagai anggota Dewan Redaksi jurnal ilmiah dan sebagai Pemimpin UnidBagian Proyek Penelitian. Untuk meningkatkan
kemampuan di bidangnya dalam tahun 1990 penulis pernah menjadi peserta Short
viii
Malaysia dan mengikuti LatihadMagang Penelitian Pertanian dan Bioteknologi
Pertanian selama 6 bulan yang diselenggarakan oleh Badan Penelitian dan
Pengembangan Pertanian di Sukamandi, Jawa Barat.
Selama mengikuti program S3, penulis menjadi anggota Perhimpunan
Fitopatologi Indonesia. Karya ilmiah berjudul Assays isolates of Corynespora cassiicola originated from papaw and differential rubber clones telah disampaikan
pada simposium International Rubber Research and Development Board dan
diterbitkan di dalam prosiding pada tahun 2000. Karya ilmiah tersebut merupakan
PRAKATA
Puji dan syukur penulis panjatkan ke hadirat Allah SWT atas ridha yang
diberikan sehingga penyusunan disertasi ini dapat diselesaikan. Tema yang dipilih
dalam penelitian ini ialah "Produksi dan Inaktivasi in Vitro Toksin Isolat
Corynespora cassiicola Asal Daun Karet". Cendawan patogenik C. cassiicola
yang mampu menghasilkan toksin ini adalah penyebab penyakit penting pada daun karet yang mengakibatkan klon-klon karet rentan tidak dapat disadap karena
pertumbuhan batang tanaman sangat terhambat setelah terjadi gugur daun secara
terus menerus. Disertasi ini disusun sebagai salah satu syarat untuk menempuh
pendidikan doktor pada Program Pascasarjana di Institut Pertanian Bogor. Harapan
peaillis, tulisan dalam disertasi ini dapat memberikan konstribusi dalam ilmu pengetahuan hubungan interaksi antara tanaman inang dan patogen dan membuka
jalan dalam upaya melakukan penelitian-penelitian yang mendasari teknik penanggulangan penyakit khususnya penyakit daun tanaman karet.
Pada kesempatan ini penulis ingin mengucapkan terima kasih kepada berbagai
fihak yang telah memberikan bantuan dan dukungan dalam pelaksanaan dan penyelesaian tugas belajar dan penyusunan disertasi yang ditempuh di Institut
Pertanian Bogor. Ucapan terima kasih ini berturut-turut disampaikan kepada:
1. Dr. Ir. Meity Suradji Sinaga, MSc. sebagai Ketua Komisi Pembimbing yang
X
2. Prof. Dr. Ir. Rusmilah Suseno, MSc. sebagai Anggota Komisi Pembimbing yang dengan penuh kesabaran telah memberikan petunjuk-petunjuk ilmiah dan
bimbingan yang sangat baik dalam pelaksanaan penelitian dan penyelesaian
disertasi.
3. Dr. Ir. Sri Hendrastuti Hidayat, MSc. sebagai Anggota Komisi Pembimbing yang telah memberikan saran-saran dengan kritis dan konsultatif baik dalam
penyusunan proposal, pelaksanaan penelitian dan pen-yusunan disertasi.
4. Dr. Asril Darussamin, MS sebagai Anggota Komisi Pembimbing dan manian Direktur Pusat Penelitian Karet yang telah banyak meluangkan waktu, tenaga
dan pemikiran, memberikan pengarahan-pengarallan yang sangat baik dan bimbingan penuh selama dalam pelaksanaan penelitian, telah mengupayakan penulis untuk mengikuti studi S3 di IPB serta selalu mendorong penyelesaian
studi.
5. Dr. Siswanto, D.E.A. sebagai Anggota Komisi Pembimbing yang telah memberikan kesempatan dan fasilitas penuh untuk bekerja di Laboratorium
Biokiinia dan Molekuler dan memberikan arahan-arahan yang sangat baik dalam pelahanaan penelitian dan penyusunan disertasi.
6. Dr. Ir. Soekirman Pawirosoemardjo, MS sebagai Direktur Pusat Penelitian
Karet yang telah memberikan kesempatan dan pembiayaan untuk
menyelesaikan tugas belajar dan mengupayakan pendanaan semaksimal
xi
7. Dr. Ir. Karyudi sebagai Kepala Balai Penelitian Sungei Putih
-
Pusat Penelitian Karet, yang telah mengupayakan dana untuk pelaksanaan penelitian danmemberikan dorongan untuk penyelesaian studi.
8. Ir. Sujatno dan Saudara Soleh Suryaman di Kelompok Proteksi Tanaman Balai Penelitian Sungei Putih, Saudara Ahmad Topany dan Mamak Suryani masing- masing di Laboratorium Biokimia & Molekuler dan di Laboratorium Mikroba
Balai Penelitian Bioteknologi Perkebunan Bogor serta Saudara Edi Supardi di
Laboratorium Virologi Jurusan Ilmu Hama & Penyakit Tumbuhan, Fakultas
Pertanian IPB, yang telah membantu pelaksanaan penelitian.
9. Dr. Ir. Darmono Taniwiryono, MSc. sebagai Kepala Balai Penelitian Bioteknologi Perkebunan Bogor, dan ir. Basuki, MS sebagai mantan Kepala Unit Penelitian Bioteknologi Perkebunan Bogor yang telah memberi izin dan
fasilitas dalam pelaksanaan penelitian di lingkup instansi tersebut.
10. Dr. Djoko Santosa peneliti di Balai Penelitian Bioteknologi Perkebunan Bogor
yang telah membantu membran dialisis berukuran 3,5 kDa untuk dapat
melanjutkan analisis di laboratorium.
11. Rektor dan Direktur Pasca sarjana IPB serta jajarannya, para staf pengajar dail karyawan yang telah memberikan andil dalam proses belajar dan pelaksanaan
penelitian.
12. Ir. Heri Dwi Basuki dan keluarga yang telah banyak memberikan dukungan
moral dan finansial.
13. Isteri saya Ir. Ermina Muhayati yang selalu penuh pengertian dan kesabaran
xi i
yang selalu memberi arti dan penghiburan tersendiri sehingga penulis selalu
berupaya dapat menyelesaikan tugas belajar ini.
Penelitian dan penyusunan disertasi ini tidak mungkin dapat terlaksana tanpa
dukungan dari berbagai fihak. Oleh karena itu, kepada semua fihak yang telah memungkinkan pelaksanaan penelitian dan penyelesaian disertasi ini dapat
berlangsung dengan baik, penulis menyatakan berhutang budi. Selain itu, penulis menyampaikan permohonan ma'af kepada fihak-fihak yang tidak mungkin
disebutkan namanya satu persatu di sini. Akhirnya, semoga karya ilmiah ini
bermanfaat.
Bogor, Desember 2003
DAFTAR IS1
AB STRAK
...
ABSTRACT...
SURAT PERNYATAAN...
RI WAY AT HIDUP
...
PRAKAT A...
DAFTAR IS1...
DAFTAR TABEL...
DAFTAR GAMBAR...
PENDAHULUAN
...
Latar Belakang...
Perumusan Masalah
...
Tujuan Penelitian...
TINJAUPLW PUSTAKA...
Etiologi dan Karakterisasi C . cassiicola
...
...
Kisaran Inang
Sebaran Geografi
...
Spesifisitas lnang...
Toksin
(:
cassiicola...
Inaktivasi Toksin
...
STUD1 1 PRODUKSI IN VITRO TOKSIN ISOLAT Corynespora cassiicola
DAN TOKSISITASNYA PADA DAUN KARET
Halaman
. .
11
...
111
iv v vii
...
X l l l
xv xvii 1 1 5 7 8 8 10 11 13 14 16
1.1 Pendahuluan
...
1.2 Metode Penelitian
...
1.2.1 Monitoring Penyakit Gugur Daun Corynespora di Lapangan
...
1.2.2 Pengukuran konidia C . cassiicola
1.2.3 Preparasi Isolat
...
1.2.4 Produksi Toksin
...
...
1.2.5 Bioasai in Planta untuk Uji Fitotoksisitas Toksin
...
1.2.6 Pengaruh Tingkat Keasaman
...
1.2.7 Pemisahan Toksin dan Penentuan Berat Molekul
...
1.3 Hasil dan Pembahasan
...
1.3.1 Kisaran Inang dan Kejadian Penyakit
1
.
3.2 Keragaman Konidia...
1.3.3 Uji Fitotoksisitas Toksin dengan Metode Bioasai in Planta..
.
...
1.3.4 Produksi Toksin Isolat C cassiicola
...
1.3.5 Pengaruh Tingkat Kemasaman
STUD1 2 INAKTIVASI IN VITRO TOKSIN ISOLAT Corynespora cassiicola
ASAL DAUN KARET
2.1 Pendahuluan
...
2.2 Metode Percobaan...
...
2.2.1 Preparasi Serum B dan C dari Lateks Karet
2.2.2 Penetapan Kadar Fen01
...
2.2.3 Inaktivasi dengan Pemanasan...
2.2.4 Inaktivasi dengan Senyawa Pereduksi
...
2.2.5 Inaktivasi dengan Serum B dan C dari Lateks
...
2.3 Hasil dan Pembahasan
...
2.3.1 Pengaruh Pernanasan...
2.3.2 Pengaruh Senyawa Pereduksi
...
2.3.3 Pengaruh Serum B d m C dari Lateks...
2.4 Kesimpulan...
PEMBAHASAN IJMUM
Inang Non Karet
...
.
Toksin dan Patogenisitas C cassiicola
...
Toksin C
.
cassiicola dan Inaktivasinya...
KESIMPULAN DAN SARAN
Kesimpulan
...
Saran
...
DAFTAR PUSTAKA
...
DAFTAR TABEL
Deskripsi kategori kerusakan daun akibat aktivitas toksin Corynepora cassiicola
...
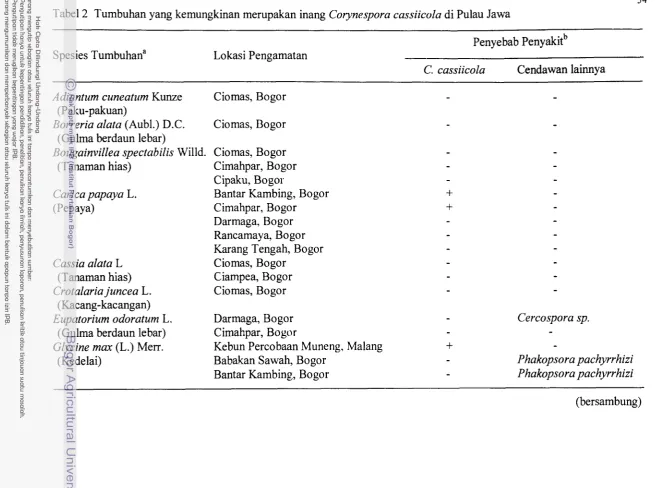
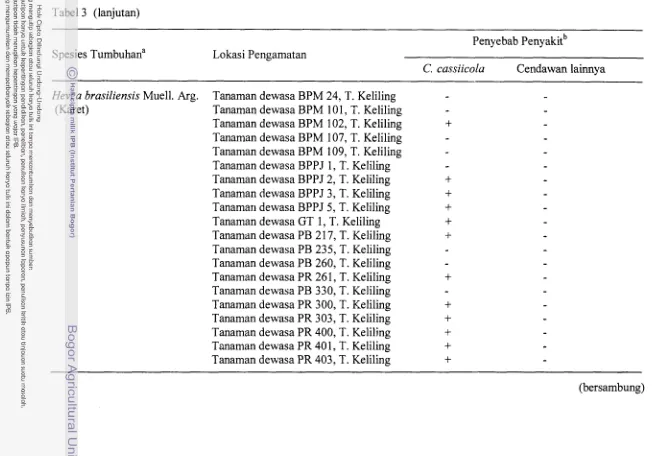
Tumbuhan yang kemungkinan merupakan hang Corynespora
...
cassiicola di Pulau JawaTumbuhan yang kemungkinan merupakan inang Corynespora
...
cassiicola di Provinsi Sumatera Utara
Kejadian Penyakit Gugur Daun Corynespora di pembibitan batang
...
bawah (rootstocks) karet
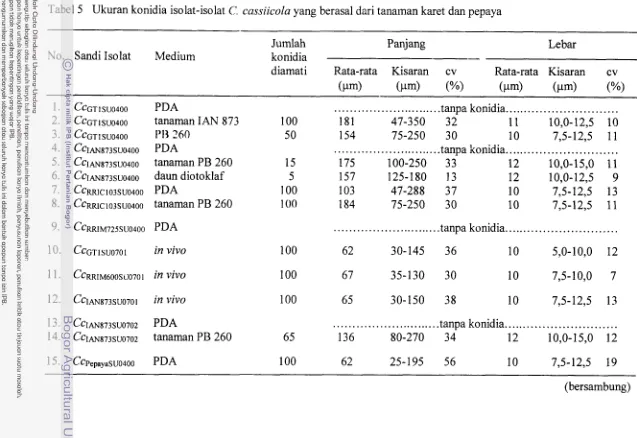
U h a n konidia isolat-isolat Corynesporn cassiicola yang berasal
...
dari tanaman karet dan pepaya
Perbandingan uji fitotoksisitas toksin antara metode bioasai in planta
...
(BiP) dan metode pencelupan pangkal daun
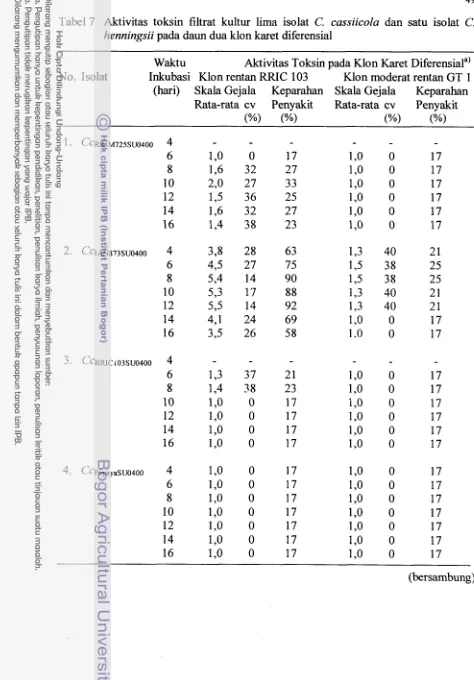
Aktivitas toksin filtrat kultur lima isolat Corynespora cassiicola dan satu isolat Cercosporidium henningsii pada daun dua klon karet
...
diferensial RRIC 103 dan GT 1
Pertumbuhan lima isolat Corynespora cassiicola pada 14 hari
setelah inkubasi dan aktivitas toksin filtrat yang dihasilkan pada klon
...
rentan RRIC 103
Berat kering miselium dan pH filtrat kultur dua isolat Corynespora cassiicola asal klon moderat rentan IAN 873 padal4 hari setelah inkubasi
...
Pengaruh pH bufer fosfat 100 rnM pada filtrat kultur isolat
CclAN873~~0400 dan Cc1~~873~~0702 terhadap aktivitas toksin pada daun klon rentan RRIC 103
...
Pengaruh perlakuan separasi dan dialisis filtrat kultur isolat
C ~ ~terhadap aktivitas toksin pada daun klon rentan ~ ~ 7 ~ ~ ~ ~ 4 ~ ~ PPN 2447
...
Halaman 29 3 4 36 3 9 42 46
C Cterhadap aktivitas toksin pada daun klon rentan ~ ~ ~ ~ ~ ~ ~ ~ ~ ~ ~
PPN 2447
...
7 513 Pengaruh pemanasan toksin yang dimurnikan parsial dari filtrat
kultur isolat C ~ 1terhadap aktivitas toksin pada daun klon ~ ~ ~ ~ ~ ~ ~ 4 ~ ~
...
rentan PPN 244714 Pengaruh serum B dan C lateks AVROS 2037 yang telah diencerkan dengan natrium ditionit 0,08% terhadap aktivitas toksin pada daun
...
klon rentan RRIC 103 80
15 Pengaruh serl-lm B dan C lateks karet klon moderat rentan IlZN 873
terhadap aktivitas toksin pada daun klon rentan PPN 2447
...
8316 Pengaruh serum B lateks lima klon karet terhadap aktivitas toksin filtrat kultur isolat dan C ~ 1 ~ ~ 8 7 ~ ~ ~ 0 7 0 2 pada daun klon
rentan RRIC 103
...
8317 Kadar fen01 serum B lateks lima klon karet diferensial AVROS 2037,
...
DAFTAR GAMBAR
Halaman
Gejala yang diakibatkan oleh toksin isolat C ~ 1 ~ ~ 8 7 3 ~ ~ 0 4 0 0 pada daun
karet
...
45Gejala kerusakan yang ditimbulkan oleh filtrat kultur urnur 4, 6, 8, 10, 12, 14 dan 16 hari setelah inkubasi dari lima isolat Corynespora cassiicola dan satu isolat Cercosporidium henningsii pada daun karet
klon rentan RRIC 103 dan klon moderat GT 1
...
5 1Warna filtrat yang dihasilkan oleh isolat-isolat C. cassiicola dan
gejala infeksi yang ditimbulkan oleh isolat C ~ ~pada daun ~ ~ ~ ~ ~ ~ 0 4 ~ ~ karet kion moderat GT 1 dan k b n rentan RRIC 103
...
5 1Pertunlbuhan kultur isolat C C I A N ~ ~ ~ ~ U O ~ O O yang lebih lambat
dibandingkan dengan isolat-isolat lainnya
...
5 6Pemurnian toksin cassiicoline isolat C~1~~873~~0400 pada kolom
...
sephacn.1 S-200-HR presipitat kultur 58
Elektroforegram SDS-PAGE (1 5% poliakrilamid) protein dari filtrat kultur isolat C ~ ~ ~ 8 7 ~ ~ ~ 0 4 0 0 setelah pemekatan dengan rotary evaporator
...
pada 50 OC dan dialisis dH20 melalui membran berporositas 3,5 kDa 5 9
Aktivitas toksin filtrat kultur isolat C C I A N ~ ~ ~ S ~ ~ ~ ~ ~ dan fiaksi yang diperoleh dari pemisahan dengan separator Vivaspin 500
...
bermembran ! 0 kDa pada dam klon rentan RRlC 103 60
Gejala toksin yang dimurnikan secara parsial berkadar protein 5 pglml sebelum dan setelah pemanasan 120 OC 10 menit pada tiga taraf pengenceran dengan dHzO 1 : 1, 1 :3 dan 1 :7 pada daun
klon rentan RRIC 103
...
Pengaruh asam askorbat, asam sitrat dan natrium ditionit rnasing- masing pada tingkat konsentrasi 0,0%, 0,2%, 0,4%, 0,6%, 0,8% dan
1,0% terhadap aktivitas toksin filtrat kultur isolat C C I A N ~ ~ ~ S U O ~ ~ ~ pada
...
daun klon rentan RRIC 1 03
Pengaruh empat senyawa pereduksi terhadap aktivitas toksin filtrat kultur (pH 5,3) isolat Cc1AN873~u0400 pada daun klon rentan
...
1 1 Pengaruh empat senyawa pereduksi pada pH 7,O terhadap aktivitas
toksin filtrat kultur isolat
C
C
I
pada daun klon rentan~
N
~
~
~
~
~
~
~
~
~
PPN 2444
...
8 112 Pengaruh serum C lateks enam klon karet diferensial terhadap aktivitas toksin filtrat kultur isolat pada daun klon
rentari PPN 2444
...
8213 Elektroforegram protein serum B dan C dari lateks dengan penvarnaan
Coomassie blue
...
8 714 Warna serum B lateks karet klon resisten (AVROS 2037, BPM I), Moderat rentan (RRIM 600, IAN 873) dan klon sangat rentan
xix
DAFTAR LAMPIRAN
Halaman
1. Dafiar turnbuhan inang Corynespora cassiicola
. . .
.
.
. .
. . .
. . .
. . . ...
1202. Sebaran geografi Corynespora cassiicola pada tumbuhan inangnya
. .
.
1253. Isolat-isolat Corynespora cassiicola dan isolat Cercosporidium
PENDAHULUAN
Latar Belakang
Penyakit Gugur Daun Corynespora (PGDC), Corynespora Leaf Fall Disease atau
Corynespora Leaf Spot, yang disebabkan oleh Corynespora cassiicola (Berk. & Curt.)
Wei merupakan penyakit penting pada tanaman karet (Hevea brasiliensis Muell. Arg.).
Pada tanaman karet PGDC berkernbang pada dam-dam muda dan daun-dam tua baik
pada tanaman di pembibitan maupun pada tanaman dewasa. Keberhasilan infeksi
patogen tersebut ditunjukkan oleh gejala nekrosis yang spesifik pada tulang-tulang
daun yang mengakibatkan pzngguguran daun sehingga tanaman dapat menjadi
meranggas. Semenjak epidemik pertama kali diketahui pada dekade 1980an di negara-
negara utama penghasil karet alam termasuk Indonesia, kini PGDC menjadi ancaman
utarna bagi kelangsungan budidaya karet. Setelah berlangsung epidemi C. cassiicola
tarnpaknya mengalami perubahan menjadi ras-ras yang lebih virulen setelah
beradaptasi dengan beberapa klon yang ada. Hal ini terbukti bahwa beberapa klon
karet yang pada muianya masih bebas ternyata kemudian terkena oleh penyakit
tersebut.
Beberapa klon karet yang berasal dari negara penghasil karet alam, antara lain
RRIC 103 (Sri Lanka), KRS 21 (Thailand), IAN 873 (Brasil) dan RRIM 725
(Malaysia), ternyata sangat rentan terhadap PGDC (Soepena 1 983). Klon-k!on hasil
pemuliaan dalam negeri di Indonesia, yaitu PPN 2058, PPN 2444 dan PPN 2447, yang
pernah menjadi unggulan karena produktivitasnya yang tinggi ternyata juga sangat
2
Klon RRIC 103 yang telah ditanam secara komersial di Sri Lanka, dilaporkan
mengalami kerusakan sangat berat pada Desember 1985 (Chee 1988). Sebelum terjadi
epidemi produktivitas klon tersebut sangat tinggi dengan pencapaian hasil karet kering
pada tahun sadap ke delapan sebesar 2.298 k g h .
Di Sri Lanka kerusakan perkebunan karet akibat PGDC mencapai 4.600 ha dalam
tahun 1987 sehingga tanaman yang rusak berat terpaksa dibongkar dan diganti dengan
klon yang lebih resisten (Jayasinghe & Silva 1996).
Di Indonesia epidemi PGDC pertama kali diketahui pada tahun 1980 di Sumatera
Utara dan Surnatera Selatan (Soepena 1983). Klon RRIC 103 dan KRS 21 yang
ditanam di Kebun Percobaan Sungei Putih, Sumatera Utara, tidak pernah dapat disadap
karena lingkar batangnya tidak mencapai kriteria minimal yang ditetapkan sebesar 45
cm di atas pertautan okulasi. Hal ini terjadi karena klon-klon tersebut mengalami
pengguguran daun secara terus-menerus selama pertumbuhannya.
Kunci keberhasilan budidaya karet diawali oleh ketepatan pemilihan klon tanaman
yang unggul produksinya dan merniliki sifat-sifat sekunder lain yailg baik. Sifat-sifat
sekunder itu antara lain tahan terhadap gangguan angin dan resisten terhadap penyakit
penting karet. Penggunaan bahan tanam karet yang resisten terhadap penyakit penting
pada daun merupakan langkah jangka panjang dalam mengatasi eksplosi penyakit. Hal
ini sejalan dengan upaya penerapan konsep pengendalian hama terpadu (PHT) seperti
yang tertuang dalam Undang-undang Nomor 12 tabun 1992 tentang Sistem Budidaya
Tanaman. Penggunaan tanaman resisten sebagai komponen utama PHT dalam
3
dikemukakan oleh Fry (1982), yaitu dapat bersifat (i) kompatibel bila dipadukan
dengan cara pengendalian penyakit lain, (ii) praktis dan tidak memerlukan penanganan
secara khusus selama satu siklus pertumbuhan tanaman, (iii) tidak menimbulkan
masalah pencemaran lingkungan, dan (iv) dengan hanya menggunakan tanaman
resisten secara tepat memungkinkan masalah suatu penyakit dapat diatasi sampai di
bawah ambang kerusakan ekonomi. Dalam budidaya karet keragaman klon resisten
dalam areal pertanaman yang luas perlu diperthnbangkan karena pertanaman
monokonal dapat mengakibatkan tekanan seleksi pada patogen sehingga resistensi
dapa dipatahkan.
Upaya penelitian untuk mengantisipzti perkembangan PGDC saat ini sangat
diperlukan. Penggunaan fingisida untuk pengendalian PGDC selama ini dirasakan
tidak e fektif karena secara teknis aplikasi h n g isida pada tanaman dewasa mengalami
kendala yaitu ketinggian pohon yang dapat mencapai lebih dari 10 m tidak mudah
dijangkau oleh hngisida yang diaplikasikan dari bawah. Alat-alat aplikasi pestisida
yang tersedia tidak dapat menjangkau ketinggian pohon dan kondisi lahan perkebunan
yang sering terjal menghambat pelaksanaan aplikasi. Selain itu, penggunaan hngisida
tidak memecahkan masalah bahkan akan menstirnulasi pembentukan ras baru yang
lebih virulen. Oleh karena itu, perlu dicari substitusi sebagai upaya solusi dalam
mengatasi masalah PGDC, antara lain dengan menggunakan klon-klon karet yang
resisten.
Budidaya yang tepat klon karet yang resisten dapat dipertirnbangkan sebagai
4
karet di Indonesia cukup tersedia, yakni terdiri atas 20 klon anjuran dan 18 klon
unggul harapan (Azwar et al. 1995), 8.000 jenis koleksi plasma nutfah dari Amerika
Selatan hasil ekspedisi International Rtrbber Research and Development Board
(IRRDB) tahun 198 1, ortet-ortet baru hasil persilangan, dan beberapa spesies selain H.
brasiliensis. Oleh karena its, perlu dianalisis faktor yang mendasari resistensi bahan
tanam karet terhadap penyakit-penyakit penting khususnya PGDC, sehingga pemilihan
jenis bahan tanarrl untuk keperluan budidaya secara komersial dapat dilakukan dengan
tepat.
Hingga kini faktor yang mendasari ketahanan klon-klon karet terhadap C.
cassiicola belum banyak diketahui. Beberapa klon karet tertentu tampaknya
memperlihatkan sifat resistensi secara umum terhadap beberapa pepyakit daun dan
beberapa klon lainnya memperlihatkan si fat resistensi secara khusus terhadap penyakit
daun tertentu. Tingkat patogenisitas C. cassiicola sangat ditentukan oleh produksi atau
aktivitas toksin yang dapat menimbulkan kerusakan dengan cepat pada jaringan daun
sehingga daun-daun yang terinfeksi menjadi gugur. Oleh sebab itu, fenomena
inaktivasi roksin oleh senyawa antitoksin dalarn tanaman karet yang kemungkinan
terjadi di dalam lateks danlatau d a m karet klon-klon tertentu menjadi penting.
Mekanisme ini akan mempunyai prospek yang baik untuk digunakan sebagai dasar
dalam pengembangan budidaya karet dengan klon-klon tertentu yang resisten terhadap
PGDC di suatu wilayah endemi sehingga menekan perkembangan C. cassiicola.
Secara umum pertahanan metabolik tanaman berupa protein-protein yang
5
proteins) telah banyak diteliti dalam upaya mencari solusi masalah penyakit. Contoh
PR protein misalnya kitinase,
P-
1,3-glukanase, dan protein penghambatpoligalakturonase (polygalacturonase-inhibitingproteidPGIP). Aktivitas kitinase dan
P-
1,3-glukanase telah banyak dipelajari pada berbagai tanaman, antara lain pada karet(Breton et al. 1997), tomat (Wubben et al. 1996), jagung (Cordero et al. 1994), kedelai
(Yi & Hwang 1996) dan cabai (Kim & Hwang 1996), sedangkan PGIP telah diketahui
terdapat pada kacang buncis (Nuss et al. 1996). Namun demikian, senyawa-senyawa
di dalam tanaman karet yang kemungkinan dapat mengakibatkan inaktivasi toksin C.
cassiicola belum pernah Ciketahui.
Perurnusan Masalah
Cendawan patogenik C. cassiicola tergolong ke dalam famili Dematiaceae yang
beberapa spesiesnya melakukan patogenesis pada inangnya dengan toksin yang
diproduksinya. Diketahui bahwa toksin yang diproduksi C. cassiicola dapat
menirnbulkan kerusakan parah pada daun-daun karet yang rentan. Hal yang rnenjadi
masalah adalah bahwa C. cassiicola memiliki kisaran tumbuhan inang yang sangat
luas, namun di Indonesia khususnya di lingkungan pertanaman karet, kisaran
tumbuhan hang tersebut belun~ banyak diketahui. Oleh karena itu, jenis tumbuhan
inang C. cassiicola yang ada masih perlu dipahami dengan baik, sedangkan ras
fisiologi C. cassiicola yang telah ada di lapangan diduga berbeda-beda kemampuannya
6
Toksin yang dihasilkan C. cassiicola yang dinamakan cassiicoline tergolong
sebagai glikoprotein dengan dua jembatan disulfida pada rantai protein (Breton 1997).
Toksin iiii belum pernah dilaporkan dapat diinaktifkan oleh senyawa-senyawa kirnia,
baik oleh senyawa-senyawa kimia sintetik maupun oleh senyawa-senyawa biokimia
yang terdapat di dalam turnbuhan hang. Sebagai glikoprotein yang bersifat fitotoksik,
cassiicoline diduga dapat diinaktihn melalui berbagai kemungkinan mekanisme,
antara lain (i) reaksi enzimatik, (ii) pengikatan protein (protein-binding), (iii)
pengubahan molekul pada gugus fbngsiorial tertentu pada toksin, dan (iv) pemblokiran.
Dalam penelitian ini akan dianalisis beberapa faktor yang mungkin berpengaruh
terhadap aktivitas biologi toksin, seperti suhu, tingkat kernasaman, senyawa-senyawa
pereduksi, dan fiaksi-fraksi lateks tmaman karet. Bagian gugils tertentu pada
cassiicoline diduga dapat direduksi oleh senyawa-senyawa kirnia pereduksi tertentu
sehingga aktivitas biologinya dapat dihambat. Beberapa jenis senyawa pereduksi yang
akan dipelajari pengaruhnya terhadap aktivitas biologi toksin, yaitu natrium ditionit
(sodium dithionite; sodium hydrosulfite; sodium hyposulfite; Na2S204), asam askorbat
(L-ascorbic acid; vitamin C; C6HsO6), asam sitrat (citric acid; C6H8O7), 2-
merkaptoetanol (~mercaptoethanol; hydroxymethylmercaptan; C2H60S), metilen
biru (methylene blue; tetramethylthionine; C 1 6H S), dan arnido hitam (amido
7
Tujuan Penelitian
Dalam penelitian ini telah dilakukan suatu seri percobaan yang bertujuan (1)
Menganalisis tingkat aktivitas toksin yang dihasilkan in vitro oleh isolat-isolat C.
cassiicola yang berasal dari hang karet
dan
non-karet, (2) Menganalisis faktor-faktoryang berpengaruh terhadap aktivitas biologi toksin yang dihasilkan oleh C. cassiicola,
dan (3) Mengevaluasi potensi senyawa-senyawa pereduksi, serum B dan C dari lateks
TINJAUAN PUSTAKA
Etiologi dan Karakterisasi C. cassiicola
Cendawan patogenik Corynespora cassiicola (Berk. & Curt.) Wei termasuk ke
dalam famili Dematiaceae, kelas Deuteromycetes. Nama genus Corynespora pertama
kali digunakan oleh Giissow untuk menamakan cendawan yang diperoleh dari daun
dan buah mentimun di Inggris tahun 1906 (Wei 1950). Nama C. cassiicola juga
sinonim dengan Hclminthosporium cassiicola Berk. & Curt. yang pertama kali
digunakan dalam tahun 1869, Cercospora melonis Cooke (1 896), Cor)wespora mazei
Giissow (1 906), Helminthosporium papuyae H. Sydow (1 923), Cercospora vignicola
Kawamura (193 1) d m Helminthosporium vignae Olive apud, Bain & Lefebvre (1945).
Spesies C. cassiicola (Berk. & Cu1-t.) Wei comb. nov. dipakai oleh Wei ( 1950)
untuk merevisi nama C'ercospora melonis, C. vignicola atau Helminthosporium vignae,
koleksi cendawan penyebab bercak daun karet (Hevea leaf spot) yang diberi nama
Helminthosporium heveae, dan 16 koleksi dalam berkas map Helminthosporium
cassiicola di Herbarium I.M.I. (!nternational Mycological Irzstitute) Inggris yang
diperoleh dari tumbuhan inang yang berbeda di negara-negara tropis.
Helminthosporium papuyae dari pepaya termasuk di antara 16 koleksi yang direvisi.
Nama Corynespora digunakan untuk mengatasi kerancuan yang ditimbulkan oleh
perbedaan antara nama tipikal He1min;hosp~rium (H. velutinum) dan Cercospora (C.
apii), sedangkan nama cassiicola digunakan pertama kali pada Helminthosporium
9
Menurut Wei (1950) karakterisasi spesies C. cassiicola ialah sebagai berikut:
Cendawan ini bersifat parasitik pada batang, daun dan buah (tetapi kebanyakan pada
daun), menyebabkan gejala bercak dengan ukuran diameter yang beragam pada kisaran
mulai kurang dari 1 mm sampai 1,5 cm dan bahkan mencapai 2 cm, berwarna pucat
sampai coklat kekuning-kuningan dengan zonasi atau bagian tepi benvarna coklat
keungu-unguan, dan sering bersifat sekunder.
Konidiofora kebanyakan bersifat hipofilus, tegak lurus ke permukaan substratum
dari miselium yang muncul melalui epidermis atau biasanya dari hifa udara,
kebanyakan tunggal atau kadang-kadang membentuk klaster, sederhana, lurus, agak
kaku, bersekat, benvarna coklat gelap, dengan atau tanpa bagian dasar yang
membentuk pentolan (bulbous base). sedikit atau sama sekali tidak membengkak pada
ujung yang berdinding tebal, proliferasi terjadi secara minimal melalui bekas
terlepasnya konidia atau kadang-kadang melalui konidiofora atau konidia yang rusak,
mencapai panjang hingga 600 pm atau lebih dan berdiameter 3,8-11,3 pm; proliferasi
bergabllng hingga panjangnya beragam, yakni 10- 100 x 5,2- 1 1,4 pm, warna bertambah
terang pada bagian ujung .
Konidia terbentuk secara tunggal pada bagian ujung, atau membentuk rantai dua
sampai enam spora pada kondisi tertentu yang kelembabannya tinggi, dan kadang-
kadang dihubungkan oleh bagian genting yang hialin ke konidiofora atau ke konidia di
bawahnya, berbentuk seperti gada (obclavate), kadang-kadang silindris, lurus atau
sering melengkung sedikit, meruncing ke arah ujung yang biasanya berdinding tipis,
10
umur, bersekat 6-14, tetapi kadang-kadang 1-2 atau hingga 20 atau lebih, dengan satu
pellicle yang berwarna dan eksospora hialin dan tebal, dan suatu hilum yang mencolok
terletak melintang 3,6-7,9 pm, berukuran 32-320 x 8,4-22,4 pm, umurnnya 65,s-181 x
13,6-20 pm, berkecambah dengan tabung kecambah polar. Konidia yang dibentuk
pada jaringan turnbuhan hang yang sukulen (polong atau buah muda) berukuran lebih
panjang daripada yang berasal dari buah, berwarna terang atau bahkan hialin,
ukurslnnya sangat beragam, mencapai panjang 520 pm, tetapi diametemya tetap 9- 1 3
pm, memiliki pseudosepta 4-20, dan sering membentuk rantai dua sampai tiga konidia.
Kisaran Inang
Kisaran tumbuhan inang C. cassiicola sangat luas. Kurang lebih 122 spesies
tumbuhan yang tergolong ke dalam 39 famili telah dilaporkan dapat menjadi inangnya
(Tabel Lampiran 1). Sebanyak 16 koleksi isolat C. cassiicola yang dipelajari oleh Wei
(1 950) berasal dari berbagai spesies tumbuhan inang, sebagai berikut: Carica papaya
L., Cassia alata L., Corchorus capsularis L., Crotalaria juncea L., Cucumis sativus
L., Glycine max (L.) Merr., Hevea brasiliensis Muell. Arg., Hibiscus esculentus L., H
sabdariffa L., Impatiens balsamina L., Lycopersicon esculentum Mill., Norantea
guianensis, Phaseolus vulgaris L., Sesamum indicum L., Tithonia speciosa dan V i p a
sinensis L.
Di Indonesia jenis turnbuhan hang C. cassiicola yang telah dilaporkan masih
terbatas. C. cassiicola dilaporkan pertama kali ditemukan pada tanaman karet dalam
1 1
(Hardaningsih et al. 1987). Cendawan patogen ini juga pernah dilaporkan terdapat
pada tanaman pepaya, ubi jalar dan tomat di Irian Jaya (Johnston 1961). Dua spesies
gulrna, Borreria alata dan Melastoma af$ne, yang diinokulasi dengan isolat C.
cussiicola dapat terinfeksi dengan intensitas kerusakan cukup tinggi, berturut-tumt 60
dan 36%. Sedangkan lima spesies lainnya, Axonopus compresus, Euputorium
odoratum, Paspalum conjugatum, Mikania cordata dan Lantana camara, terinfeksi
dengan intensitas kerusakan ringan, yakni kurang dari 20% (Budiman & Suryaningtyas
1992).
Sebaran Geografi
Sebaran geografi C. cassiicola sangat luas, meliputi negara-negara di berbagai
belahan dunia, yakni di wilayah Asia, Afiika, Amerika Utara, Arnerika Tengah,
Karibia, Pasifk, Arnerika Selatan, Eropa dan Australia. Dafiar sebaran geografi C.
cassiicola pada tumbuhan inangnya ditunjukkan pada Tabel Lampiran 2. Sebaran
geografi C. cassiicola pada tanaman pepaya mencakup areal paling luas dibandinghan
dengan tumbuhan inang lainnya dan telah dilaporkan dapat ditemukan di berbagai
negara, seperti Australia (Silva et ul. 19959, Fiji (Graham 1971), Filipina (Wei 1950),
Gold Coast (Ellis 1957), Indonesia (Johnston 1961), Kongo Belgia (Ellis 1957),
Malaysia (Thompson & Johnston 1953), Nigeria (Onesirosan et al. 1974), Sierra
Leone (Wei 1950), Singapura (Singh 1980), Sri Lanka (Silva et al. 1997), Sudan (Ellis
1957), Thailand (Giatgong 1980) dan Venezuela (Albornett & Sanabria de Albarracin
13
Spesifisitas Inang
Inang spesifik dari C. cassiicola yang menyerang tanaman karet dan tumbuhan
lain belum banyak diketahui. Pengujian patogenisitas yang pernah dilakukan dengan
menggunakan beberapa isolat dari karet dan isolat dari tanaman lainnya menunjukkan
data yang berbeda-beda. Perbedaan tersebut diperkirakan karena cendawan patogenik
h i memiliki keragaman dalam patogenisitas. Di Malaysia isolat C. cassiicola dari
karet tidak menirnbulkan geja!a pada pepaya, tomat, slada, kedelai, kakao dan kelapa
sawit, sedangkan isolat dari pepaya hanya menginfeksi pepaya saja (Chee 1988).
Hasil tersebut berbeda dengan isolat dari karet asal .Taws Barat, CcCio, yang ternyata
dapat menginfeksi kedelai hingga mencapai tingkat kerusakan daun sebesar 66%
(Suwarto 1989). Isolat yang digunakan oleh Chee (1988) tersebut tampaknya mirip
dengan isolat dari karet asal Sumatera Utara, CcSP, yang hanya spesifik menginfeksi
tanaman karet saja.
Pengujian spesifisitas inang dari 28 isolat C. cassiicola asal 20 tumbuhan inang
dari Amerika Serikat, Meksiko dan Nigeria telah dipelajari oleh Onesirosan et al.
(1974). Beliau menggunakan tanaman uji yang terdiri atas delapan spesies tanaman,
yaitu cowpea (Vigna unguiculata [L.] Walp.), kapas (Gossypium hirsutum L.), kedelai
(Glycine max [L.] Merr.), mentimun (Cucumis sativus L.), pepaya (Carica papaya L.),
terung (Solanum melongena L.), tomat (Lycopersicon esculentum Mill.) dan wijen
(Sesamum indicum L.). Hasil percobaan meilunjukkan bahwa isolat-isolat C.
cassiicola memiliki spesifisitas hang yang berbeda-beda yang dapat digolongkan ke
14
dan satu isolat dari Amerika Serikat, memiliii virulensi yang tinggi pada kedelai,
wijen, terung dan kapas, sedangkan satu isolat lainnya asal kedelai di Amerika Serikat
memiliki virulensi yang tinggi pada tomat dan terung namun bervirulensi rendah pada
kedelai. Beberapa isolat yang berasal dari tumbuhan hang non ekonorni tampaknya
berpotensi sebagai sumber inokulum primer bagi tanaman budidaya yang diuji. Isolat
C. cassiicola dari Calopogonium mucunoides Desv. dan nodeweed (Synedrella
nodlj7ora [L.] Gaertn.) dapat menj adi sumber inohlum yang potensial bagi tanaman
budidaya tornat dan terung, sedangkan isolat dari Aspilia africana (Pers.) C.D. Adarns
dan isolat dari Lepistemon sp. merr~punyai virulensi yang tinggi pada kapas.
Toksin C. cassiicola
Dalam mekanisme patogenisitasnya C. cassiicola diketahui menghasilkan toksin
yang dapat terbentuk secara in vitro setelah isolat cendawan tersebut dikulturkan pada
medium cair sintetik Modified Alterncria Medium (MAM) (Onesirosan et al. 1975,
Suwarto 1989) atau dikulturkan pada medium Czapeck yang dimodifikasi (Breton et
al. 1996). Medium cair sintetik MAM adalah medium cair dengan komposisi senyawa-
senyawa kimia sintetik defmitif untuk pertumbuhan kultur Alternaria raphani, A.
brassicae dan A. brassicicola yang diformulasi oleh Taber, Vanterpool dan Taber
(1968). Untuk menumbuhkan isolat C. cassiicola yang berasal dari tanaman tornat
dalam rangka memproduksi ioksin, MATd dimodifikasi dengan cara menghilangkan
sumber vitamin-vitamin (Onesirosan et al. 1975).
Berdasarkan percobaan Onesirosan et al. (1975), kondisi optimum untuk
15
medium 3-15 % (blv) dan periode inkubasi 12 hari. Percobaan dilakukan tanpa
penggoyangan dengan menggunakan 25 ml MAM dalam tabung Erlenmeyer
bervolume 125 ml atau 50 ml MAM dalam tabung Erlenmeyer bervolume 250 ml.
Produksi toksin isolat C. cassiicola dilaporkan oleh Onesirosan et al. (1975)
berkorelasi positif dengan pertambahan berat kering miselium selama periode inkubasi
antara 4-16 hari. Dari 28 isolat C. cassiicola dengan virulensi yang berbeda-beda
berdasarkan uji patogenisitasnya (enam isolat sangat virulen, delapa~ isolat kurang
virulen dan 14 isolat non-patogenik), isolat C. cassiicola yang sangat virulen
dilaporkan menghasilkan toksin yang sangat &if, sedangkan isolat non-patogenik
tidak menghasilkan toksin yang potensial untuk menimbulkan kerusakan pada jaringan
tmaman.
Dalam percobaan produksi toksin yang dilakukan Suwarto (1989) dengan
menggunakan 50 ml MAM dalam tabung erlenmeyer bervolume 250 ml pada kondisi
optimum yang dilaporkan Onesirosan et al. (1975), toksin yang sangat aktif dapat
terdeteksi dalam filtrat kultur isolat C. cassiicola yang berasal dari tanaman karet dan
kedelai. Fitotoksisitas toksin diuji berdasarkan bioasai secara silang pada daun kedila
tanaman tersebut dan ternyata toksin bersifat spesifik inang. Toksin yang dihasilkan
isolat dari tanaman karet dapat menimbulkan kerusakan berat pada daun karet rentan
tetapi hanya menimbulkan sedikit kerusakan pada daun kedelai. Sebaliknya, toksin
yang berasal dari tanaman kedelai hanya menimbulkan kerusakan pada daun kedelai
dan sama sekali tidak menimbulkan kerusakan pada daun karet. Aktivitas toksin
16
C. cassiicola yang diuji, yakni isolat dari karet dan kedelai, mencapai perturnbuhan
optimum berat kering rniselium pada 12-14 hari setelah inkubasi pada 26 OC.
Dari hasil percobaan Breton et al. (1996), toksin C. cassiicola yang dimurnikan
dan dinamakan cassiicoline, diketahui tergolong sebagai glikoprotein yang larut dalam
air. Penentuan berat molekul toksin tersebut tampaknya mengalami beberapa kendala.
Berdasarkan pemurnian yang dilakukan mereka dengan tahapan filtrasi 0,22 pm,
kromatografi DEAE, filtrasi gel (ACA 202 dan TSK G 3000 SW) dan SDS-PAGE,
toksin dilaporkan mempunyai berat molekul
+
6,5 kDa (Breton, Garcia, Sanier,d'Auzac & Esbach 1996), tetapi pada laporan berikutnya toksin tersebut dinyatakan
memiliki berat molekul21 kDa (Breton & d' Auzac 1999), dan terdiri atas tiga peptida
yang masing-masing non-toksik, yakni 8,8, 7,l dan 5,9 kDa. Kelebihan perkiraan berat
molekul toksin tersebut dinyatakan telah terjadi. Pada penetapan dengan spektrometri
masa, toksin dilaporkan mempunyai berat molekul riil2,886 kDa (Breton 1997).
Inaktivasi Toksin
Berdasarkan penelitian Breton (1997) toksin yang dihasilkan oleh isolat C.
cassiicola diketahui bersifat termo-stabil. Sebanyak 1 pglml casiicoline yang
dipanaskan dalam otoklaf pada 120 OC selama 15 dan 30 menit masih mengakibatkan
fitotoksisitas 60%. Pada percobaan yang pernah dilakukan oleh Suwarto (1989) filtrat
kultur C. cassiicola isolat Cc-SP yang berasal dari tanaman karet mengalami
kehilangan aktivitas toksin setelah diberi perlakuan pemanasan pada 121 OC selama 30