KARMANAH
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN SUMBER INFORMASI
Dengan ini saya menyatakan bahwa tesis : ” Peningkatan Ketahanan Batang Bawah Jeruk Pada Lahan Masam Melalui Kultur In Vitro adalah karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apapun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutif dari karya ilmiah yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Bogor, Februari 2009
ABSTRACT
KARMANAH. The Improvement of Citrus Resistance on Acid Soil throughIn Vitro Culture. Under supervisions of SLAMET SUSANTO and MIA KOSMIATIN.
One of the citrus breeding method to obtain rootstock variant tolerance to Al and acid soil stress can be done through in vitro selection technique. This research aim to improve the genetic of Japansche citroen (JC) rootstock through irradiated induce mutation and in vitro selection on selective media Al and low pH to obtain a tolerance varieties to Al and acid soil. This research was conducted in three stages of trial, these were : 1). Irradiated of mutation to increase genetic varieties with gamma rays dose 0, 250, 500, 750 and 1000 rad and embryogenic callus multiplication in MS media with B5 vitamin and 2.4-D 0,5 mg/l + BAP 0,05 – 0,15 mg/l, and NAA 5 mg/l + Kinetin 0,5 mg/l combination ; 2) In vitro selection on Al selective media 0, 250, 500, 750 and 1000 ppm AlCl3.6H2O in pH 4 ; 3). AlCl3.6H2O-selected callus regeneration in media MS + VMW + BAP 3 mg/l + Malt extract 500 mg/l. The result showed that genetic varieties of citrus rootstock increased with 750 rad irradiated induction mutation and in vitro selection on MS media + VB5 + 2.4-D 0,5 mg/l + BAP 0,05 – 0,15 mg/l. 65 the candidate citrus varian tolerance to Al and low pH was produced from the mutant embryo with gamma rays dose 250 rad (19 varian), 500 rad (15 varian), 750 rad (9 varian) and 1000 rad (22 varian). Whereas irradiated of mutation 750 rad and Al selected media in 250 ppm AlCl3.6H2O concentration was produced the best plant growth response.
RINGKASAN
KARMANAH. Peningkatan Ketahanan Batang Bawah Jeruk pada Lahan Masam melalui Kultur In Vitro. Dibimbing oleh SLAMET SUSANTO dan MIA KOSMIATIN.
Penelitian ini bertujuan untuk meningkatkan keragaman genetik batang bawah jeruk melalui induksi mutasi iradiasi dan seleksi secarain vitro dengan menggunakan media yang selektif dan efektif untuk mendapatkan varian yang toleran pada Al dan lahan masam. Kegiatan penelitian meliputi tiga tahap percobaan yaitu : 1). Mutasi iradiasi dan perbanyakan kalus embriogenik ; 2). SeleksiIn-vitro pada media Al dan pH rendah 3). Regenerasi kalus terseleksi AlCl3.6H20.
Hasil percobaan 1, mutasi iradiasi sinar gamma pada embrio muda dapat menurunkan persentase embrio yang hidup. Mutasi iradiasi sinar gamma dengan dosis yang paling tinggi (1000 rad) pada umur 30 hari setelah iradiasi menunjukkan persentase embrio yang hidup paling rendah (56,5%). Embrio yang diiradiasi dengan dosis 250 rad dan dikulturkan di media MS + VB5 + 2,4-D 0,5 mg/l + BAP 0,1 mg/l menghasilkan diameter kalus terbesar (0.95 cm) dan jumlah induksi bakal tunas terbanyak (21 bakal tunas). Sedangkan diameter kalus terkecil 0.35 cm diperoleh dari embrio mutan 1000 rad pada media yang sama. Secara visual media MS yang dikombinasikan dengan 2.4-D 0.05 mg/l dan BAP 0.05-0.15 mg/l dapat menginduksi pembentukan dan pertumbuhan kalus yang lebih baik (100%), dan kalus yang terbentuk umumnya berwarna putih hingga putih kekuningan dan remah.
Hasil percobaan 2, kalus embrio mutan 250, 500 dan 750 rad pada media seleksi AlCl3.6H2O 250 ppm menunjukkan persentase kalus yang hidup paling tinggi (100%), sedangkan embrio mutan 1000 rad menunjukkan persentase kalus yang hidup paling rendah (50%). Perlakuan dosis iradiasi dan media seleksi Al berpengaruh nyata terhadap pertambahan diameter kalus. Diameter kalus terbesar (1.30 cm) diperoleh dari embrio mutan 750 rad pada media seleksi AlCl3.6H2O 750 ppm. Sedangkan diameter kalus terkecil (0.45 cm) diperoleh dari embrio mutan 500 rad pada media tanpa AlCl3.6H2O. Embrio mutan 750 rad yang diseleksi pada media Al juga dapat membentuk tunas dengan jumlah paling banyak (4 tunas). Secara visual kalus yang diseleksi pada media Al mengalami perubahan warna dari putih kekuningan hingga kecoklatan.
Hasil percobaan 3, menunjukkan jumlah kalus beregenerasi dari kalus embrio mutan 250 rad dan kalus embrio yang tidak diiradiasi masing-masing 7 kalus. Dari kalus embrio mutan 750 rad (6 kalus), kalus embrio mutan 500 rad (5 kalus) dan paling sedikit dari kalus embrio mutan 1000 rad (3 kalus).
Hasil analisis kromosom pada beberapa varian terseleksi Al menunjukkan jumlah kromosom yang sama dengan kromosom individu normalnya yaitu 18 artinya tidak terjadi mutasi kromosom akibat iradiasi sinar gamma dan seleksi Al.
Dari hasil penelitian ini dapat disimpulkan bahwa 1) Keragaman genetik batang bawah jeruk meningkat dengan induksi mutasi iradiasi sampai 750 rad dan seleksi in vitro pada media MS + VB5 +2.4-D 0,5 mg/l + BAP 0,05 - 0,15 mg/l. 2) Secara keseluruhan telah dihasilkan 65 varian jeruk kandidat toleran Al dan pH rendah, dimana perlakuan mutasi iradiasi 750 rad dan media seleksi AlCl3.6H2O 250 ppm menghasilkan varian jeruk dengan respon pertumbuhan yang paling baik.
© HAK cipta milik IPB, tahun 2009 Hak cipta dilindungi undang-undang
Dilarang mengutip sebahagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik atau tinjauan suatu masalah ; dan pengutipan tersebut tidak merugikan kepentingan yang wajar IPB
PENINGKATAN KETAHANAN BATANG BAWAH JERUK PADA
LAHAN MASAM MELALUI KULTUR
IN - VITRO
KARMANAH
Tesis
Sebagai salah satu syarat untuk memperoleh gelar Magister Sains pada
Departemen Agronomi dan Hortikultura
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Judul Tesis : Peningkatan Ketahanan Batang Bawah Jeruk Pada Lahan Masam Melalui KulturIn Vitro
Nama : Karmanah
NRP : A151060061
Program Studi : Agronomi
Disetujui Komisi Pembimbing
Prof. Dr. Ir. Slamet Susanto, MSc Mia Kosmiatin, SSi . MSi Ketua Anggota
Diketahui
Ketua Program Studi Agronomi Dekan Sekolah Pascasarjana
Dr. Ir. Munif Ghulamahdi, MS Prof. Dr. Ir. Khairil A. Notodiputro, MS
PRAKATA
Alhamdullilah, segala puji dan syukur penulis panjatkan kepada Allah SWT yang senantiasa melimpahkan Rahmat dan KaruniaNya, sehingga penulis dapat menyelesaikan tesis yang berjudul ”Peningkatan Ketahanan Batang Bawah Jeruk Pada Lahan Masam Melalui KulturIn Vitro”.
Penelitian ini merupakan langkah awal yang dilakukan pada penelitian batang bawah jeruk dalam kondisi cekaman lingkungan (Cekaman Al dan pH rendah) secara in vitro, sehingga diharapkan dapat dikembangkan lagi penelitian-penelitian lanjutan untuk memperoleh batang bawah jeruk yang toleran pada Al dan pH rendah (lahan masam).
Dalam melaksanakan penelitian dan penulisan tesis ini penulis banyak mendapatkan bimbingan dan bantuan dari berbagai pihak. Untuk itu penulis menyampaikan penghargaan dan ucapan terima kasih kepada Prof. Dr. Ir. Slamet Susanto, MSc dan Mia Kosmiatin, SSi. MSi selaku ketua dan anggota komisi pembimbing yang telah banyak meluangkan waktu untuk memberikan arahan serta bimbingan dalam pelaksanaan penelitian maupun dalam penulisan tesis ini. Secara khusus ucapan terima kasih kepada Dr. Ir. Agus Purwito, MSc selaku penguji luar komisi atas saran dan kritiknya.
Ucapan terima kasih disampaikan pula kepada :
1. Rektor Universitas Nusa Bangsa dan Ketua Yayasan PKMK Nusantara yang telah memberikan ijin kepada penulis untuk menempuh pendidikan S2 di Program Studi Agronomi Sekolah Pascasarjana Institut Pertanian Bogor.
2. Direktorat Jenderal Pendidikan Tinggi (DIKTI), Departemen Pendidikan Nasional yang telah memberikan beasiswa BPPS kepada penulis sehingga dapat menempuh dan menyelesaikan pendidikan S2.
3. Prof. Dr. Sudirman Yahya, MSc., Dr. Pasril Wahid, MS. APU dan Dr. Eny Widjayani, MS yang telah memberikan rekomendasi kepada penulis untuk menempuh pendidikan S2 di Sekolah Pascasarjana Institut Pertanian Bogor. 4. Ketua Kelompok Peneliti Biologi Sel dan Jaringan BB-Biogen atas ijinnya juga
KARMANAH
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN SUMBER INFORMASI
Dengan ini saya menyatakan bahwa tesis : ” Peningkatan Ketahanan Batang Bawah Jeruk Pada Lahan Masam Melalui Kultur In Vitro adalah karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apapun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutif dari karya ilmiah yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Bogor, Februari 2009
ABSTRACT
KARMANAH. The Improvement of Citrus Resistance on Acid Soil throughIn Vitro Culture. Under supervisions of SLAMET SUSANTO and MIA KOSMIATIN.
One of the citrus breeding method to obtain rootstock variant tolerance to Al and acid soil stress can be done through in vitro selection technique. This research aim to improve the genetic of Japansche citroen (JC) rootstock through irradiated induce mutation and in vitro selection on selective media Al and low pH to obtain a tolerance varieties to Al and acid soil. This research was conducted in three stages of trial, these were : 1). Irradiated of mutation to increase genetic varieties with gamma rays dose 0, 250, 500, 750 and 1000 rad and embryogenic callus multiplication in MS media with B5 vitamin and 2.4-D 0,5 mg/l + BAP 0,05 – 0,15 mg/l, and NAA 5 mg/l + Kinetin 0,5 mg/l combination ; 2) In vitro selection on Al selective media 0, 250, 500, 750 and 1000 ppm AlCl3.6H2O in pH 4 ; 3). AlCl3.6H2O-selected callus regeneration in media MS + VMW + BAP 3 mg/l + Malt extract 500 mg/l. The result showed that genetic varieties of citrus rootstock increased with 750 rad irradiated induction mutation and in vitro selection on MS media + VB5 + 2.4-D 0,5 mg/l + BAP 0,05 – 0,15 mg/l. 65 the candidate citrus varian tolerance to Al and low pH was produced from the mutant embryo with gamma rays dose 250 rad (19 varian), 500 rad (15 varian), 750 rad (9 varian) and 1000 rad (22 varian). Whereas irradiated of mutation 750 rad and Al selected media in 250 ppm AlCl3.6H2O concentration was produced the best plant growth response.
RINGKASAN
KARMANAH. Peningkatan Ketahanan Batang Bawah Jeruk pada Lahan Masam melalui Kultur In Vitro. Dibimbing oleh SLAMET SUSANTO dan MIA KOSMIATIN.
Penelitian ini bertujuan untuk meningkatkan keragaman genetik batang bawah jeruk melalui induksi mutasi iradiasi dan seleksi secarain vitro dengan menggunakan media yang selektif dan efektif untuk mendapatkan varian yang toleran pada Al dan lahan masam. Kegiatan penelitian meliputi tiga tahap percobaan yaitu : 1). Mutasi iradiasi dan perbanyakan kalus embriogenik ; 2). SeleksiIn-vitro pada media Al dan pH rendah 3). Regenerasi kalus terseleksi AlCl3.6H20.
Hasil percobaan 1, mutasi iradiasi sinar gamma pada embrio muda dapat menurunkan persentase embrio yang hidup. Mutasi iradiasi sinar gamma dengan dosis yang paling tinggi (1000 rad) pada umur 30 hari setelah iradiasi menunjukkan persentase embrio yang hidup paling rendah (56,5%). Embrio yang diiradiasi dengan dosis 250 rad dan dikulturkan di media MS + VB5 + 2,4-D 0,5 mg/l + BAP 0,1 mg/l menghasilkan diameter kalus terbesar (0.95 cm) dan jumlah induksi bakal tunas terbanyak (21 bakal tunas). Sedangkan diameter kalus terkecil 0.35 cm diperoleh dari embrio mutan 1000 rad pada media yang sama. Secara visual media MS yang dikombinasikan dengan 2.4-D 0.05 mg/l dan BAP 0.05-0.15 mg/l dapat menginduksi pembentukan dan pertumbuhan kalus yang lebih baik (100%), dan kalus yang terbentuk umumnya berwarna putih hingga putih kekuningan dan remah.
Hasil percobaan 2, kalus embrio mutan 250, 500 dan 750 rad pada media seleksi AlCl3.6H2O 250 ppm menunjukkan persentase kalus yang hidup paling tinggi (100%), sedangkan embrio mutan 1000 rad menunjukkan persentase kalus yang hidup paling rendah (50%). Perlakuan dosis iradiasi dan media seleksi Al berpengaruh nyata terhadap pertambahan diameter kalus. Diameter kalus terbesar (1.30 cm) diperoleh dari embrio mutan 750 rad pada media seleksi AlCl3.6H2O 750 ppm. Sedangkan diameter kalus terkecil (0.45 cm) diperoleh dari embrio mutan 500 rad pada media tanpa AlCl3.6H2O. Embrio mutan 750 rad yang diseleksi pada media Al juga dapat membentuk tunas dengan jumlah paling banyak (4 tunas). Secara visual kalus yang diseleksi pada media Al mengalami perubahan warna dari putih kekuningan hingga kecoklatan.
Hasil percobaan 3, menunjukkan jumlah kalus beregenerasi dari kalus embrio mutan 250 rad dan kalus embrio yang tidak diiradiasi masing-masing 7 kalus. Dari kalus embrio mutan 750 rad (6 kalus), kalus embrio mutan 500 rad (5 kalus) dan paling sedikit dari kalus embrio mutan 1000 rad (3 kalus).
Hasil analisis kromosom pada beberapa varian terseleksi Al menunjukkan jumlah kromosom yang sama dengan kromosom individu normalnya yaitu 18 artinya tidak terjadi mutasi kromosom akibat iradiasi sinar gamma dan seleksi Al.
Dari hasil penelitian ini dapat disimpulkan bahwa 1) Keragaman genetik batang bawah jeruk meningkat dengan induksi mutasi iradiasi sampai 750 rad dan seleksi in vitro pada media MS + VB5 +2.4-D 0,5 mg/l + BAP 0,05 - 0,15 mg/l. 2) Secara keseluruhan telah dihasilkan 65 varian jeruk kandidat toleran Al dan pH rendah, dimana perlakuan mutasi iradiasi 750 rad dan media seleksi AlCl3.6H2O 250 ppm menghasilkan varian jeruk dengan respon pertumbuhan yang paling baik.
© HAK cipta milik IPB, tahun 2009 Hak cipta dilindungi undang-undang
Dilarang mengutip sebahagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik atau tinjauan suatu masalah ; dan pengutipan tersebut tidak merugikan kepentingan yang wajar IPB
PENINGKATAN KETAHANAN BATANG BAWAH JERUK PADA
LAHAN MASAM MELALUI KULTUR
IN - VITRO
KARMANAH
Tesis
Sebagai salah satu syarat untuk memperoleh gelar Magister Sains pada
Departemen Agronomi dan Hortikultura
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Judul Tesis : Peningkatan Ketahanan Batang Bawah Jeruk Pada Lahan Masam Melalui KulturIn Vitro
Nama : Karmanah
NRP : A151060061
Program Studi : Agronomi
Disetujui Komisi Pembimbing
Prof. Dr. Ir. Slamet Susanto, MSc Mia Kosmiatin, SSi . MSi Ketua Anggota
Diketahui
Ketua Program Studi Agronomi Dekan Sekolah Pascasarjana
Dr. Ir. Munif Ghulamahdi, MS Prof. Dr. Ir. Khairil A. Notodiputro, MS
PRAKATA
Alhamdullilah, segala puji dan syukur penulis panjatkan kepada Allah SWT yang senantiasa melimpahkan Rahmat dan KaruniaNya, sehingga penulis dapat menyelesaikan tesis yang berjudul ”Peningkatan Ketahanan Batang Bawah Jeruk Pada Lahan Masam Melalui KulturIn Vitro”.
Penelitian ini merupakan langkah awal yang dilakukan pada penelitian batang bawah jeruk dalam kondisi cekaman lingkungan (Cekaman Al dan pH rendah) secara in vitro, sehingga diharapkan dapat dikembangkan lagi penelitian-penelitian lanjutan untuk memperoleh batang bawah jeruk yang toleran pada Al dan pH rendah (lahan masam).
Dalam melaksanakan penelitian dan penulisan tesis ini penulis banyak mendapatkan bimbingan dan bantuan dari berbagai pihak. Untuk itu penulis menyampaikan penghargaan dan ucapan terima kasih kepada Prof. Dr. Ir. Slamet Susanto, MSc dan Mia Kosmiatin, SSi. MSi selaku ketua dan anggota komisi pembimbing yang telah banyak meluangkan waktu untuk memberikan arahan serta bimbingan dalam pelaksanaan penelitian maupun dalam penulisan tesis ini. Secara khusus ucapan terima kasih kepada Dr. Ir. Agus Purwito, MSc selaku penguji luar komisi atas saran dan kritiknya.
Ucapan terima kasih disampaikan pula kepada :
1. Rektor Universitas Nusa Bangsa dan Ketua Yayasan PKMK Nusantara yang telah memberikan ijin kepada penulis untuk menempuh pendidikan S2 di Program Studi Agronomi Sekolah Pascasarjana Institut Pertanian Bogor.
2. Direktorat Jenderal Pendidikan Tinggi (DIKTI), Departemen Pendidikan Nasional yang telah memberikan beasiswa BPPS kepada penulis sehingga dapat menempuh dan menyelesaikan pendidikan S2.
3. Prof. Dr. Sudirman Yahya, MSc., Dr. Pasril Wahid, MS. APU dan Dr. Eny Widjayani, MS yang telah memberikan rekomendasi kepada penulis untuk menempuh pendidikan S2 di Sekolah Pascasarjana Institut Pertanian Bogor. 4. Ketua Kelompok Peneliti Biologi Sel dan Jaringan BB-Biogen atas ijinnya juga
5. Prof. Dr. Nad Darga T, MS., dan Dr.Ir. H. Arie Lastario K, MSc atas arahannya serta Ir. Hj Tyas Pratiwi, MSi yang banyak membantu penulis selama menempuh pendidikan S2 ini.
6. Ucapan terima kasih juga penulis sampaikan kepada rekan-rekan Program Studi Agronomi Angk 2006 : Sari, Elni, Yosi, Usi Imelda, Fahmi, B’Zul, M’Mei, B’Robi, B’Ami, B’Ifa, P’Ali dan rekan-rekan lainnya atas segala dukungan dan semangatnya.
7. Terakhir penulis sampaikan ucapan terima kasih yang setinggi-tingginya kepada kedua orang tua penulis, bapak dan ibu mertua, suami tercinta dan anakku tersayang Ikbal Ramadhan, kakakku A’Yadi dan T’Elah, adik-adikku Ajat dan Ibeng serta seluruh keluarga atas segala do’a restu dan kasih sayangnya.
Penulis menyadari akan segala keterbatasan dalam penulisan tesis ini. Semoga tesis ini bermanfaat bagi perkembangan ilmu pengetahuan dan pertanian khususnya.
Bogor, Februari 2009
RIWAYAT HIDUP
Penulis dilahirkan di Bogor pada tanggal 7 Juni 1970, sebagai anak kelima dari Bapak H. Sukarna dan Ibu Hj. Ika. Tahun 1989 penulis lulus dari Sekolah Menengah Atas Tajur Bogor. Pada tahun yang sama penulis melanjutkan pendidikan pada Jurusan Budidaya Pertanian Fakultas Pertanian Universitas Nusa Bangsa. Penulis memperoleh gelar sarjana Pertanian pada tahun 1996.
Tahun 1999 penulis diterima sebagai staf pengajar di Fakultas Pertanian Universitas Nusa Bangsa Bogor. Pada tahun 2001 penulis menikah dengan Musa dan dikaruniai seorang putra bernama Ikbal Ramadhan.
DAFTAR ISI Induksi Mutasi dengan Iradiasi Sinar Gamma ... . 9 SeleksiIn Vitro... 11 Pengaruh Kemasaman Lahan terhadap Tanaman ... 12
DAFTAR TABEL
Halaman 1. Pengaruh dosis iradiasi sinar gamma dan jenis media terhadap
diameter kalus, jumlah bakal tunas dan warna kalus ... 22
2. Pengaruh dosis iradiasi sinar gamma dan media seleksi Al terhadap
diameter kalus, jumlah tunas dan visual biakan kalus ... 27
3. Jumlah kalus beregenerasi pada media regenerasi MS + VMW +
BAP 3 mg/l + Ekstrak malt 500 mg/l ... 30
4. Jumlah tunas dan Rerata tinggi tunas pada media regenerasi MS+VMW + BA 3 mg/l + Ekstrak malt 500 mg/l setelah
4 kali subkultur ... 31
5. Jumlah daun dan jumlah akar pada media regenerasi MS+VMW
DAFTAR GAMBAR
Halaman 1. Buah Jeruk JC yang digunakan ... 17
2. Biji dari buah jeruk JC dan Embrio ... 18
3. Kerangka Alur Penelitian ... 20
4. Persentase embrio yang hidup pada berbagai tingkatan dosis iradiasi
sinar gamma umur 7, 30 dan 60 hari setelah iradiasi ... 21
5. Pembentukan dan Pertumbuhan Kalus Embriogenik pada Beberapa
Jenis Media ... 24
6. Pembentukan kalus embrio mutan dan embrio yang tidak diiradiasi ... 25
7. Persentase kalus yang hidup pada beberapa media seleksi Al, 4 MSP ... 26
8. Respon pertumbuhan kalus pada beberapa media seleksi Al ... 28
9. Pertumbuhan tunas embrio mutan 750 rad pada beberapa media
seleksi Al ... 34
10. Pertumbuhan tunas embrio yang tidak diiradiasi dan embrio mutan
yang terseleksi AlCl3.6H2O 500 ppm ... 35
11. Hasil analisis kromosom varian dari embrio yang tidak diiridiasi dan
DAFTAR LAMPIRAN
Halaman 1. Sasaran produksi buah jeruk untuk memenuhi kebutuhan dalam
negeri, ekspor dan pemenuhan bahan industri pengolahan
tahun 2005 -2025 ... 45
2. Luas Lahan pengembangan jeruk di 10 Proponsi di Indonesia ... 46
3. Kesesuaian Lahan untuk Tanaman Jeruk (Citrus sp) ... 47
4. Kriteria Penilaian Sifat-Sifat Kimia Tanah ... 48
5. Keragaan batang bawah Jeruk JC ... 49
6. Komposisi Media MS ... 50
PENDAHULUAN
Latar belakangJeruk (Citrus sp) merupakan buah subtropik yang telah berkembang luas di Indonesia. Jeruk menjadi salah satu komoditas unggulan dalam program pengembangan usaha agribisnis buah diantara komoditas buah lainnya. Jeruk dapat tumbuh dan berproduksi baik di dataran rendah sampai dataran tinggi dan pada lahan sawah ataupun tegalan. Tercatat areal pertanaman jeruk di Indonesia tahun 2004 mencapai 72.306 ha dengan produktivitas rata-rata 17,829 ton/ha, produksi nasional jeruk pada tahun 2004 sebesar 1.600.000 ton dan masih mengimpor sebanyak 94.696 ton sedangkan ekspornya sebesar 1.261 ton (Departemen Pertanian, 2005). Kondisi tersebut di atas menunjukkan bahwa peluang pasar domestik masih sangat terbuka. Selain itu diperkirakan permintaan buah jeruk akan terus meningkat oleh karenanya pemacuan produksi jeruk nasional menjadi sangat penting (Direktorat Jenderal Bina Produksi Hortikultura, 2005). Sasaran produksi buah jeruk untuk memenuhi kebutuhan dalam negeri dapat dilihat pada Lampiran 1.
Pada tahun 2010, kebutuhan produksi buah jeruk diprediksi sebesar 2.355.550 ton dan jika produktivitasnya 17 - 20 ton per ha, maka pada tahun tersebut diperlukan luas panen kurang lebih 127.327 ha dari sekitar 70.000 ha luas panen yang tersedia pada tahun 2004. Penambahan luas areal untuk mencapai total produksi yang telah ditetapkan hingga tahun 2010 diperkirakan seluas 30.060 ha. Dari luasan ini, maka keperluan bibit jeruk diperkirakan sebanyak 15.030.000 (populasi 500 bibit/ha) (Badan Litbang Departemen Pertanian, 2005).
Berdasarkan kondisi agribisnis jeruk saat ini dan yang ingin diwujudkan masa mendatang terutama pada tahun 2010, Departemen Pertanian melalui kebijakan program revitalisasi agribisnis jeruk melaksanakan beberapa kegiatan antara lain Pengembangan kawasan sentra produksi buah jeruk di 10 Propinsi seluas 5.651.388 ha (Lampiran 2) (Badan Litbang Departemen Pertanian, 2005). Pengembangan areal tersebut meliputi lahan kering masam dan lahan pasang surut di Kalimantan, Sulawesi dan Sumatera.
sehingga untuk mengatasinya diperlukan jenis batang bawah jeruk yang kompatibel dan toleran pada lahan masam.
Menurut Masyarakat Jeruk Indonesia (2004), pemilihan jenis batang bawah jeruk yang kompatibel dan adaptif pada beberapa jenis lahan merupakan faktor penting dalam pengembangan agribisnis jeruk di Indonesia karena akan mempengaruhi bentuk pohon, pola produksi dan produktivitas, kualitas buah serta lama masa produksi. Lebih lanjut dikatakan bahwa kriteria batang bawah yang ideal adalah : perakaran harus cukup (lebat), toleran terhadap kondisi tanah yang tidak menguntungkan, tahan terhadap serangan hama dan penyakit, berkecepatan tumbuh sesuai dengan batang atas yang digunakan dan dapat memberikan produksi hasil yang baik dan berkualitas. Hingga kini belum ada satu varietas batang bawah yang dianggap ideal yang mempunyai seluruh sifat seperti tersebut di atas.
Saat ini lebih dari 95% pertanaman jeruk yang ada di Indonesia menggunakan batang-bawah Rough lemon (Citrus jambhiri Lush) atau sering disingkat RL, dan Japansche citroen (Citrus limonia Osbeck) atau sering disebut JC (Balitjestro, 2006). Kelebihan RL dan JC adalah sesuai dengan hampir semua batang atas jeruk yang ada. Batang bawah berperan terhadap kecepatan pertumbuhan batang atas dan mempengaruhi kualitas buah yang dihasilkan. Hasil penelitian Fallahi et al (1989) dalam Barus (2000), batang bawah mempengaruhi produksi, kandungan air, kandungan juice dan kandungan asam pada buah. Selain itu batang bawah juga mempengaruhi transpirasi, fiksasi CO2 dan kandungan asam amino (Moreno et al, 1994).
Meskipun JC dapat memberikan beberapa sifat baik, namun menurut Triatminingsih dan Karsinah (2004), jeruk JC sebaiknya tidak ditanam atau digunakan pada daerah endemis penyakit busuk akar Phytopthora sp dan lahan masam, karena sifatnya memang kurang toleran. Untuk mengatasi hal tersebut, perlu dilakukan perakitan jeruk dengan menggunakan teknik kultur jaringan guna memperoleh sifat yang toleran pada Al dan pH rendah.
meningkatkan ketahanan sel terhadap Al telah dilakukan pada tanaman tomat dan kentang (Stavarek dan Rains, 1984) ; untuk mendapatkan varian somaklon tenggang Al pada sorghum (Foy et al.,1993). Dall’Agnel et al (1996), melakukan skrining plasma nutfah alfalfa untuk ketenggangan terhadap Al secarain vitro.Seleksiin vitro untuk ketenggangan terhadap aluminium pada empat genotipe jagung juga dilakukan oleh Sutjahjo (2006), dan pada tanaman kedelai (Mariska.,et al, 2004).
Menurut Mariska et al (2004), seleksi in vitro dapat digunakan untuk meningkatkan ketahanan tanaman kedelai terhadap cekaman Al dan pH rendah, dimana sifat-sifat yang dihasilkan dapat diturunkan pada generasi berikutnya. Pengujian di lapang menghasilkan beberapa nomor yang memiliki ketahanan yang lebih baik terhadap Al dan pH rendah dibandingkan varietas yang toleran.
Mariska et al (2004), mengatakan bahwa dalam melakukan seleksi in vitro diperlukan metode regenerasi dengan tingkat keberhasilan yang cukup tinggi. Lebih lanjut dikatakan bahwa keberhasilan pengembangan metode seleksi in vitro memerlukan tersedianya (a) metode kultur jaringan yang efektif untuk regenerasi tanaman dari sel varian dalam jumlah banyak, (b) agens penyeleksi yang dapat menginduksi perkembangan dan poliferasi jaringan varian tetapi mematikan jaringan normal, dan (c) adanya korelasi antara fenotipik hasil seleksi pada tingkat sel dengan fenotipik pada tingkat tanaman.
Tujuan
Penelitian ini bertujuan untuk meningkatkan keragaman genetik batang bawah jeruk melalui induksi mutasi iradiasi dan seleksi secarain vitro dengan menggunakan media yang selektif dan efektif untuk mendapatkan varian yang toleran pada Al dan lahan masam.
Hipotesis
1. Keragaman genetik batang bawah jeruk dapat ditingkatkan dengan perlakuan induksi mutasi iradiasi sinar gamma dan media seleksiin vitro
TINJAUAN PUSTAKA
Ekofisiologi Tanaman JerukJeruk (Citrus sp) banyak dibudidayakan di daerah tropik dan sub tropik. Terdapat banyak seleksi kultivar jeruk yang berasal dari berbagai cara hibridisasi, mutasi dan poliploidi yang terjadi dalam species Citrus. Asal-usul jeruk yang terseleksi tersebut tidak jelas, sehingga sulit untuk melakukan pengelompokkan dan klasifikasi kultivar jeruk yang ada. Menurut Karsinah (1999) klasifikasi jeruk (‘true citrus’) sebagai berikut :
Umumnya jeruk diperbanyak secara komersial dengan biji. Biji tanaman jeruk mempunyai sifat poliembrional, artinya dari satu buah benih jeruk yang ditanam dapat tumbuh menjadi lebih dari satu kecambah jeruk (Rismunandar, 1981). Cara lain yang dilakukan dalam perbanyakan tanaman jeruk adalah dengan melakukan penyambungan antara batang atas dan batang bawah. Batang atas biasanya berasal dari tunas vegetatif pohon induk, sedangkan bibit batang bawah biasanya merupakan hasil perbanyakan melalui biji. Perbanyakan tanaman ini dipengaruhi oleh faktor lingkungan.
Faktor lingkungan yang sangat mempengaruhi adalah jenis tanah, suhu, curah hujan serta ketinggian dari permukaan laut. Jeruk dapat tumbuh dengan baik di wilayah dataran sedang – tinggi ( 500 m dpl), pada tanah yang berdrainase baik, solum tanah minimum 50 cm, banyak mengadung bahan organik dan tekstur tidak terlalu liat (Djaenudinet al, 2000). Tingkat kemasaman tanah 4.0 – 7.8 (optimum 6), temperatur 13 – 35oC (optimum 22-23oC) dengan curah hujan 1000 – 3000 mm/thn (Masyarakat Jeruk Indonesia, 2004).
sangat penting. Kesesuaian lahan adalah tingkat kecocokan suatu bidang lahan untuk penggunaan tertentu. Kesesuaian tersebut dapat dinilai untuk kondisi saat ini atau setelah diadakan perbaikan. Lebih spesifik lagi kesesuaian lahan tersebut ditinjau dari sifat-sifat fisik lingkungannya yang terdiri dari tanah, iklim, topografi, hidrologi dan atau drainase sesuai untuk suatu usahatani atau komoditas tertentu yang produktif Dalam menilai kesesuaian lahan ada beberapa cara antara lain dengan perkalian parameter, penjumlahan atau meggunakan hukum minimum yaitu mencocokan (matching) antara kualitas dan karakteristik lahan sebagai parameter dengan kriteria kelas kesesuaian lahan yang telah disusun berdasarkan persyaratan penggunaan atau persyaratan tumbuh tanaman atau komoditas lainnya yang dievaluasi.
Struktur klasifikasi kesesuaian lahan menurut FAO (1976) adalah :
1. Ordo : Kesesuaian lahan secara global. Pada tingkat ini kesesuaian lahan dibedakan atas sesuai (S) dan tidak sesuai (N).
2. Kelas : Lahan yang tergolong sesuai dibedakan antara lahan yang sangat sesuai (S1), cukup sesuai (S2) dan sesuai margina (S3).
Kelas S1 sangat sesuai artinya lahan tidak mempunyai faktor pembatas yang berarti atau nyata terhadap penggunaan secara berkelanjutan.
Kelas S2 cukup sesuai artinya lahan mempunyai faktor pembatas dan faktor pembatas ini akan berpengaruh terhadap produktivitas sehingga memerlukan tambahan masukan (input).
Kelas S3 sesuai marginal artinya lahan mempunyai faktor pembatas yang berat sehingga memerlukan tambahan masukan yang lebih banyak daripada lahan yang tergolong S2.
Kelas N tidak sesuai artinya lahan yang tidak sesuai karena mempunyai faktor pembatas yang sangat berat dan sulit diatasi.
3. Subkelas : keadaan tingkatan dalam kelas kesesuaian lahan. Kelas kesesuaian lahan dibedakan menjadi subkelas berdasarkan kualitas dan karakteristik lahan yang menjadi faktor pembatas. Tergantung peranan faktor pembatas pada masing-masing subkelas, kemungkinan kelas kesesuaian lahan yang dihasilkan bisa diperbaiki dan ditingkatkan kelasnya sesuai masukan yang diberikan.
diperlukan dan sering merupakan perbedaan detil dari faktor pembatasnya. Dalam praktek evaluasi kesesuaian lahan pada kategori unit ini jarang dilakukan.
Persyaratan kesesuaian lahan untuk jeruk dan kriteria sifat-sifat kimia tanah dapat dilihat pada Lampiran 3 dan 4.
Jeruk Japansche Citroen (Citrus limonia Osbeck)
Jeruk Japansche Citroen (Citrus limonia Osbeck) merupakan varietas hybrida yang dihasilkan dari persilangan antara Citroes nobilis (keprok) X Citroes medica (lemon). Japansche Citroen (JC) bersifat mirip dengan Rough Lemon, tahan terhadap kekeringan, dapat merangsang pembentukan buah lebih awal dari biasanya dan menghasilkan produksi tinggi dengan kualitas yang baik. Jenis ini peka terhadap serangan Exocortis (Sugiyarto, 1994). Exocortis adalah sejenis virus yang dapat menyebabkan terganggunya kulit tanaman dan menyebabkan tanaman menjadi kerdil (Niyomdhan, 1997).
Menurut Masyarakat Jeruk Indonesia (2004), jeruk Japansche Citroen mempunyai ciri-ciri sebagai berikut :
1. Pohon tegar dan produktif, ukuran sedang, cabang menyebar dan merunduk, duri kecil dan sedikit
2. Daun berwarna hijau gelap, aroma daun menyengat, pupus warna ungu
3. Bunga berukuran kecil hingga sedang, putik dan kelopak bunga berwarna ungu tua.
4. Buah berukuran kecil hingga sedang, warna kulit buah bila masak kekuningan sampai jingga kemerahan.
5. Biji jumlahnya banyak, berukuran kecil dan warna keping biji hijau muda, setiap buah berisi 8-10 biji, 100 kg buah diperoleh 8.000-10.00 biji yang baik, derajat poliembrioninya rendah yaitu 18.3%-50% semaian generatif 35%.
6. Tahan kekeringan
7. Daya dukung terhadap batang atas baik dan cepat menghasilkan buah yang berkualitas sedang hingga baik.
Menurut Purbiati et al (2002) Japansche Citroen memiliki kevigoran yang tinggi, ukuran biji sedang (diameter 0,5 cm), mudah beradaptasi tetapi buahnya sangat masam dan kurang layak untuk dikonsumsi, oleh karenanya direkomendasikan sebagai batang bawah. Keragaan batang bawah jeruk Japansche Citroen dapat dilihat pada Lampiran 5.
Batang bawah Japansche Citroen memiliki tingkat kompatibelitas yang baik. Hasil penelitian Susanto (2003) menunjukkan penggunaan batang bawah Japansche Citroen bersifat lebih mendorong pertumbuhan vegetatif batang atas dibandingkan Rangpur Lime dan Rough Lemon. Sedangkan hasil penelitian Rahayuni dan Hadijah (1996) menunjukkan tanaman yang berbatang bawah Japansche Citroen tumbuh lebih vigor sehingga berukuran lebih besar dibandingkan dengan batang bawah Rough Lemon,Citrus aurantifolia,Citrus amblycarpadanCitroen nobilis.
Kultur Jaringan Tanaman Jeruk
Program pengembangan tanaman jeruk menuntut adanya penyediaan bibit tanaman jeruk yang bebas penyakit, baik batang atas maupun batang bawah (Triatminingsih dan Karsinah, 2004). Pemilihan jenis batang bawah jeruk yang kompatibel dan adaptif pada beberapa jenis lahan juga merupakan faktor penting dalam pengembangan agribisnis jeruk di Indonesia karena akan mempengaruhi bentuk pohon, pola produksi dan produktivitas, kualitas buah serta lama masa produksi (Masyarakat Jeruk Indonesia, 2004).
Kulturin vitro dapat dimanfaatkan tidak hanya untuk perbanyakan tetapi juga untuk perbaikan tanaman termasuk perakitan varian yang dapat adaptif pada lahan marjinal dan penyimpanan plasma nutfah. Selain itu untuk dapat merakit varietas baru, sebaiknya penelitian tidak hanya dilakuan di laboratorium tetapi harus dilanjutkan dengan pengujian lapang, oleh karenanya sangat diperlukan kerja sama dengan para pemulia konvensional. George dan Sherington (1984) menyatakan bahwa keunggulan teknik kultur jaringan yaitu (a) dapat memperbanyak varietas hibrida baru untuk tujuan komersial, (b) dapat memperoleh tanaman baru yang bebas virus, (c) dapat memperbanyak tanaman yang sukar diperbanyak dengan biji, (d) dapat memperoleh tanaman induk yang sama sifat genetiknya dalam jumlah banyak dan (e) dapat menghasilkan tanaman baru sepanjang tahun tanpa dipengaruhi oleh musim.
Pada prinsipnya metode kultur jaringan merupakan cara untuk memperbanyak protoplas atau sel atau organ dalam media tumbuh aseptik (yang mengandung formulasi hara buatan) dengan lingkungan yang terkendali. Lestari (2008) menyatakan bahwa prinsip yang mendasari penggunaan metode kultur jaringan adalah pembuktian teori totipotensi sel.
Totipotensi sel adalah kemampuan sel untuk tumbuh dan berkembang menjadi individu yang sempurna jika ditempatkan dalam lingkungan yang sesuai dan terkendali (Lestari, 2008). Arah pertumbuhan dan perkembangan suatu sel sangat dipengaruhi oleh lingkungan tumbuhnya, zat pengatur tumbuh yang ditambahkan, serta dipengaruhi oleh media tumbuhnya. Ketepatan pemilihan dan penggunaan media kultur sangat menentukan keberhasilan penggunaan teknik kultur jaringan.
Media kultur jaringan yang paling banyak digunakan untuk berbagai tujuan kultur jaringan adalah media dasar Murashige dan Skoog (1962). Walaupun pada awal mulanya unsur-unsur makro dalam media MS dibuat untuk kultur kalus tembakau, tetapi komposisi MS ini juga mendukung dan sesuai untuk hampir semua kultur jaringan tanaman. Media MS mengandung unsur makro dengan konsentrasi relatif tinggi terutama unsur N yang diberikan dalam bentuk NO3- dan NH4+ (nitrat dan amonium).
media, sedangkan hara mikro berbeda-beda komposisinya sesuai jenis media. Lebih lanjut dijelaskan bahwa unsur hara yang disediakan dalam media sebenarnya merupakan kebutuhan pokok bagi tanaman yang tumbuh di lapangan yang tersedia di dalam tanah. Unsur-unsur hara ini diberikan ke dalam media dalam bentuk garam-garam anorganik. Selanjutnya sejalan dengan perkembangan teknik kutur jaringan tanaman, komposisi media bervariasi sesuai dengan tujuan kultur.
Dalam menggunakan metode kultur jaringan, media sangat berkaitan erat dengan penggunaan zat pengatur tumbuh (ZPT) yang ditambahkan ke dalam media tersebut. Zat pengatur tumbuh yang banyak digunakan dalam kultur jaringan adalah golongan auksin, giberelin dan sitokinin. Dalam memilih dan menggunakan ZPT, Watimmena (1987), menjelaskan beberapa hal penting yaitu ZPT harus sampai dan berada cukup lama di jaringan target. Selain itu ZPT eksogen yang diberikan harus berinteraksi dengan fitohormon endogen, yang menimbulkan reaksi sinergis atau antagonis sehingga interaksinya akan tampil dalam pertumbuhan. Pemilihan jenis dan konsentrasi ZPT yang diberikan harus diperhatikan, karena pada prinsipnya pemberian ZPT eksogen akan merubah nisbah atau keseimbangan ZPT endogen (Winata, 1988).
Zat pengatur tumbuh yang ditambahkan dapat mempengaruhi arah pertumbuhan dan perkembangan suatu eksplan, serta dapat memacu terjadinya perubahan-perubahan metabolisme sel eksplan. Menurut Wattimena (1987), prinsip kerja ZPT adalah memacu suatu perubahan-perubahan metabolisme sel yang seterusnya menjurus pada respon fisiologi tertentu. Lebih lanjut dijelaskan bahwa penting untuk mengetahui respon fisiologis suatu tanaman, sebab respon fisiologi tersebut tidak hanya dipengaruhi oleh satu ZPT, tetapi juga merupakan kerjasama beberapa ZPT. Masing-masing ZPT akan berada dalam keadaan seimbang agar pertumbuhan berjalan normal.
Induksi Mutasi dengan Iradiasi Sinar Gamma
bagian tanaman namun lebih banyak terjadi pada jaringan yang masih aktif mengadakan pembelahan sel seperti tunas, biji, dan sebagainya.
Mutasi sebenarnya dapat terjadi secara alamiah di alam, tetapi peluang kejadiannya sangat kecil yaitu 10-7 – 10-6 (Syarifah, 2006). Untuk meningkatkan terjadinya mutasi alami, dilakukan mutasi buatan dengan menggunakan mutagen.
Menurut Poespodarsono (1988), mutagen dapat dikelompokkan kedalam 3 golongan yaitu : (1) mutagen kimia, seperti EMS (Ethyl Methane Sulfonate), DES (Diethyl Sulfate) dan NMU (Nitrosomethyl Urea), (2) mutagen fisik iradiasi, seperti sinar x, sinar , dan sinar , (3) mutagen fisik non-radiasi, seperti sinar UV. Akhir-akhir ini mulai dikembangkan mutagen biologi (transposom).
Salah satu cara induksi yang saat ini banyak dilakukan yaitu induksi mutasi fisik iradiasi. Induksi iradiasi dapat menyebabkan mutasi karena sel yang diiradiasi dibebani oleh tenaga kinetik yang tinggi sehingga dapat mempengaruhi atau mengubah reaksi kimia yang ada dalam jaringan tanaman, yang pada akhirnya menyebabkan perubahan susunan kromosom (Poespodarsono, 1988). Sumber iradiasi yang sering digunakan yaitu sinar gamma dari Cobalt 60. Kelebihan sinar gamma antara lain karena energi dan daya tembusnya yang relatif tinggi, serta secara global telah terbukti paling efektif dan efisien dalam menghasilkan varietas unggul bermacam jenis tanaman (Maluszynski,et al, 2000).
Faktor yang mempengaruhi terbentuknya mutan antara lain adalah besarnya dosis iradiasi. Dosis iradiasi diukur dalam satuan Gray (Gy), 1 Gy sama dengan 0,10 krad yaitu 1 J energi per kilogram iradiasi yang dihasilkan (Soedjono, 2003). Dosis iradiasi dibagi tiga yaitu : Tinggi (>10 k Gy), sedang (1-10 k Gy) dan rendah (<1 k Gy).
Dosis optimum untuk mendapatkan mutan yang diinginkan selain dipengaruhi oleh teknik iradiasi dan jenis tanaman juga dipengaruhi oleh sinar iradiasi serta bentuk bahan tanaman yang diiradiasi. Bahan tanaman yang bisa digunakan antara lain ; biji, stek, umbi, planlet (tanaman kultur jaringan), polen, kalus, dan sebaginya. Iradiasi sinar gamma sebaiknya dilakukan pada tingkat kalus karena sel-selnya masih bersifat meristematik sehingga lebih responsif terhadap radioaktif dibandingkan dengan sel-sel dewasa (Soedjono, 2003).
hidup atau tunas, dapat memperpanjang waktu kemasakan pada buah-buahan dan sayuran, serta meningkatkan kadar pati, protein dan kadar minyak pada biji jagung, kacang dan bunga matahari. Tanaman mutan juga memiliki daya tahan yang lebih baik terhadap serangan patogen dan kekeringan (Soedjono, 2003).
Iradiasi sinar gamma dapat memunculkan berbagai mutan baru sebagai sumber keragaman, diantaranya untuk mengembangkan tanaman yang toleran terhadap Al dan pH rendah. Hasil penelitian iradiasi sinar gamma pada kalus yang mengalami subkultur lama dapat meningkatkan keragaman genetik dan mendapatkan somaklonal nilam yang kadar minyaknya lebih tinggi dari tanaman induknya (Mariska et al, 1997).
Menurut Delta et al (1995), iradiasi sinar gamma dapat menyebabkan terjadinya perubahan kandungan klorofil sebagai akibat perubahan jalur biosintesis pigmen klorofil dan waktu yang dibutuhkan untuk sintesis klorofil dalam jaringan palisade dan spongi sel-sel mesofil. Beberapa hasil penelitian dan publikasi mengungkapkan bahwa iradiasi sinar gamma dapat menyebabkan ketidakseimbangan fungsi fisiologis tanaman, hal ini sebagai kontribusi iradiasi sinar gamma yang merusak DNA sehingga menyebabkan terjadinya perubahan ekspresi gen (Duncan, 1997). Menurut Harten (1998), semakin tinggi dosis iradiasi sinar gamma, semakin meningkatkan efek kerusakan pada DNA, bahkan dosis tertentu dapat mengakibatkan perubahan struktur kromosom atau aberasi kromosom.
Menurut Kadir (2007), berdasarkan hasil penelitiannya iradiasi sinar gamma yang dilanjutkan dengan seleksi in vitro dapat meningkatkan sifat toleransi tanaman nilam dari yang peka menjadi toleran terhadap kekeringan. Hasil penelitian iradiasi sinar gamma 400 rad dan seleksi in vitro pada taraf Al 0 –500 ppm pada pH 4 diperoleh beberapa tanaman kedelai yang diduga tahan terhadap cekaman Al dan kemasaman tanah (Mariskaet al, 2001b).
SeleksiIn Vitro
yang diinginkan. Seleksiin vitro diketahui sangat berguna untuk menghasilkan varian somaklonal yang mempunyai populasi dengan karakteristik unggul tertentu, diantaranya mengembangkan klon yang toleran terhadap faktor biotik (hama dan penyakit) dan abiotik (cekaman kekeringan, pH rendah dan Al tinggi). Munculnya variasi somaklonal dapat disebabkan oleh berbagai faktor, yaitu (1) jenis eksplan yang digunakan, (2) tipe kultur antara lain kultur kalus, kultur suspensi dan kultur protoplasma, (3) zat pengatur tumbuh, (4) lamanya fase pertumbuhan kalus, (5) komposisi bahan kimia dalam media (6) genotipe yang digunakan (Jacobsen, 1987).
Namun masalah yang sering dihadapi dalam seleksiIn Vitro adalah sulitnya meregenerasikan massa sel yang tahan Al dan pH rendah, padahal regenerasi tanaman sangat penting untuk pembuatan klon yang mampu mempertahankan susunan genetik sehingga diperoleh tanaman yang tahan pada cekaman Al dan pH rendah (Purnamaningsih dan Mariska, 2003). Dengan demikian sistem regenerasi dari masa sel perlu dikuasai terlebih dahulu.
Pengaruh Kemasaman Lahan terhadap Tanaman
Kemasaman lahan adalah faktor stres terbesar yang mempengaruhi pertumbuhan tanaman dimana keberadaan aluminium merupakan faktor pembatas pertumbuhan pada tanah masam. Kelarutan ion Al yang tinggi pada tanah masam, sering berada pada pH dibawah 5.5 yang telah lama diketahui memberi efek negatif terhadap pertumbuhan tanaman. Al telah dapat bersifat racun bagi tanaman meskipun konsentrasinya masih sangat rendah (mikromolar). Disamping ada sebagian Al membentuk ikatan dengan ligand atau dalam bentuk tidak beracun seperti aluminium silikat. Bentuk Al yang bersifat toksik bagi tanaman adalah ion trivalent Al 3+ yang dominan pada kondisi masam (Delhaizeet al., 1995).
Ryanet al., (1993) menyatakan bahwa hanya ujung 2-3 mm dari akar jagung meliputi tudung akar dan meristem yang perlu diperlakukan dengan Al untuk menghambat pertumbuhan, sedangkan jika Al secara selektif digunakan pada zona pemanjangan atau pada semua bagian akar kecuali ujungnya, pertumbuhan tidak terganggu.
Menurut Jones dan Kochian (1995) akar yang diperlakukan secara cepat dengan Al akan terhambat pemanjangan sel-sel akarnya dan mengakibatkan ujung-ujung akar membengkak, namun ketika tanaman diperlakukan dengan Al lebih lama (lebih dari 24 jam) terjadi penghambatan pemanjangan dan pembelahan sel-sel akar.
Pengaruh kerusakan Al pada tanaman diawali dengan adanya gangguan terhadap tudung akar yang mempunyai sinyal dan merupakan detektor gaya gravitasi dan hambatan mekanis sehingga pada gilirannya akan mengurangi sekresi mucilage sel tudung akar dimana sel tersebut merupakan sumber pengatur endogen pertumbuhan.
Pada tingkat molekular, Al berhubungan dengan DNA sehingga interaksi Al dengan DNA akan menghentikan sifat-sifat fitokomia dan fungsi biologis seperti menghentikan pembelahan sel pada meristem akar, perpanjangan sel, sintesis DNA dan RNA (Matsumoto, 1991). Lebih lanjut dikatakan bahwa pada dinding sel, penghambatan terjadi karena Al menggantikan kedudukan Ca 2+ pada lamela tengah. Ca2+ merupakan second messenger dalam aktivitas H+-ATPase dengan bantuan protein regulator calmodulin. Dalam hal ini dengan digantikannya Ca2+ yang melekat pada calmodulin akan terjadi perubahan aktivitas enzim. Ikatan Al dengan karboksil (RCOO-) membentuk ikatan kuat sehingga sel tidak mampu membesar. Selain itu Ryanet al., (1997) melaporkan hasil penelitiannya bahwa Al juga berhubungan dengn membran lipida bilayer pada sel dimana Al dapat memblok Ca2+ dan saluan K+ sehingga mengganggu proses penyerapan hara tanaman. Selanjutnya pada tingkat selular ion Al mempengaruhi permeabilitas dan aktivitas transpor membran plasma. Gejala yang umum diketahui dimana Al menghambat pertumbuhan akar dapat digunakan sebagai alat pengukur untuk mengidentifikasi pengaruh keracunan Al. Pada larutan hara dengan konsentrasi Al beberapa mikromolar dalam waktu 60 menit telah menghambat pertumbuhan akar.
kondisi pH <5, sedangkan Al(OH)2+ dan Al(OH)2+merupakan bentuk yang dominan dengan makin tingginya pH. Pada keadaan tanah yang bereaksi netral, Al berbentuk Al(OH)3– atau gibsit, sedangkan pada tanah alkalin dijumpai bentuk Al(OH)4-. Kation Al monomer membentuk ikatan dengan berbagai ligand asam organik dan anorganik seperti PO43-, SO42-, asam organik, protein dan lemak.
Pada tanah dengan pH rendah maka kapasitas ion H+ meningkat. Hal ini menyebabkan penyerapan unsur-unsur lainnya menjadi berkurang dan unsur Al berkelebihan. Meningkatnya konsentrasi Al terlarut akibat kemasaman tanah mengakibatkan terjadinya defisiensi P, K dan hara mikro seperti seng, tembaga dan molybdenum. Hasil penelitian Yamamoto et al., (1992) mendapatkan bahwa toksisitas Al selain mengakibatkan tanaman kekurangan nutrien juga mengubah struktur dan fungsi dari membran plasma dan menghalangi pembelahan sel pada ujung-ujung akar. Pada akhirnya tanaman akan mengurangi sistem perakarannya dan menunjukkan berbagai gejala kekurangan nutrien akibat keracunan Al (MacDiarmid dan Gardner, 1996).
Ada berbagai kriteria yang ditetapkan untuk menentukan apakah suatu tanaman toleran atau tidak terhadap cekaman Al. Samuel et al., (1997) menetapkan suatu kriteria bagi tanaman yang toleran terhadap cekaman Al yaitu : (1). Akar mampu untuk tumbuh terus dan ujung akarnya tidak mengalami kerusakan dan (2). Ion Al sedikit yang ditranslokasikan ke bagian atas dan sebagian besar ditahan di akar.
Menurut Marschner (1995) ada 2 kelompok mekanisme toleransi tanaman terhadap stres Al yaitu :
1. Mekanisme eksternal (exclution tolerance mechanism) adalah sistem toleransi yang dibangun oleh tanaman dengan cara mencegah Al untuk tidak masuk ke dalam sistem simplas. Bentuknya dapat berupa : immobilisasi Al di dinding sel, permeabilitas selektif dari membran plasma, barier pH di rhizosfer, eksudasi ligand pengkelat Al, efluks Al-fosfat.
BAHAN DAN METODE
Waktu dan Tempat
Penelitian dilaksanakan di Laboratorium Kultur In vitro Biologi Sel dan Jaringan, Balai Besar Penelitian dan Pengembangan Bioteknologi dan Sumberdaya Genetik Pertanian (BB-BIOGEN) Bogor. Iradiasi sinar gamma dilakukan di Pusat Aplikasi Teknologi Isotop dan Radiasi (PATIR), Badan Tenaga Atom Nasional (BATAN) Jakarta. Analisis jumlah kromosom dilakukan di Laboratorium Biologi Lembaga Ilmu dan Penelitian Indonesia (LIPI) Cibinong. Penelitian dilaksanakan dari bulan Agustus 2007 sampai dengan Desember 2008.
Bahan dan Alat
Bahan tanaman yang digunakan dalam penelitian ini adalah biji dari buah jeruk Japansche Citroen (JC) yang masih muda, diperoleh dari Balai Penelitian Jeruk dan Buah Subtropis Tlekung – Batu, Jawa Timur. Media dasar yang digunakan adalah media Murashige dan Skoog (1962) yang disingkat dengan MS. Gula 30 -50 g/l dan bahan pemadat phytagel sebanyak 2,5 g/l ditambahkan ke dalam media MS. Zat pengatur tumbuh yang digunakan adalah Napthalene Acetic Acid (NAA), Benzil Adenin Purin (BAP), 2,4 Dichlorophenoxyacetic Acid (2,4-D) dan Kinetin. Vitamin yang digunakan adalah Vitamin Gamborg - B5, Vitamin MS, serta Vitamin Morel dan Weitmore (VMW). Komponen penyeleksi yang digunakan adalah AlCl3.6H2O. Bahan-bahan lain yang digunakan alkohol 70% dan 96 %, NaOH, HCl, dan Spirtus.
Metodologi Penelitian
Penelitian dibagi dalam tiga tahap percobaan yaitu 1). Mutasi Iradiasi dan perbanyakan kalus embriogenik ; 2). Seleksi In vitro pada media Al dan pH rendah 3). Regenerasi kalus terseleksi AlCl3.6H2O.
1. Mutasi iradiasi dan perbanyakan kalus embriogenik
Eksplan yang digunakan untuk mutasi iradiasi dan perbanyakan kalus embriogenik ini berasal dari embrio biji jeruk yang masih muda, berukuran± 1 mm . Biji jeruk yang digunakan berasal dari buah muda yang berdiameter antara 1- 2 cm (Gambar 1).
Gambar 1 Buah jeruk JC yang digunakan
Gambar 2 (A) Biji dari buah jeruk JC dan (B) Embrio yang akan diiradiasi
Embrio dalam media ½ MS tadi diiradiasi dengan menggunakan sinar gamma Cobalt 60 di Pusat Aplikasi Teknologi Isotop dan Radiasi (PATIR), Badan Tenaga Atom Nasional (BATAN) Jakarta. Setelah diiradiasi embrio diinduksi pembentukkan kalusnya dengan cara disubkulturkan pada media MS ditambah dengan Vitamin Gambrog-B5 (VB5) dan diperkaya dengan ZPT 2,4-D dan BAP dengan konsentrasi berdasarkan perlakuan. Setiap botol kultur diisi dengan 4 eksplan. Botol kultur yang sudah ditanami disimpan dalam ruang kultur dengan temperatur ruangan 19oC - 21oC. Penyinaran menggunakan lampu TL 100 watt dengan intensitas penyinaran 16 jam per hari.
Percobaan menggunakan Rancangan Acak Lengkap (RAL) dengan 2 faktor. Faktor pertama yaitu dosis iradiasi dengan 5 taraf : 0, 250, 500, 750 dan 1000 rad. Faktor kedua yaitu jenis media dengan 4 kombinasi perlakuan :
A. MS + VB5 + 2,4-D 0,5 mg/l + BAP 0,15 mg/l B. MS + VB5 +2,4-D 0,5 mg/l+ BAP 0,1 mg/l C. MS + VB5 +2,4-D 0,5 mg/l+ BAP 0,05 mg/l D. MS + VB5 + NAA 5 mg/l + K 0,5 mg/l.
Komposisi media MS dapat dilihat pada Lampiran 6.
Setiap perlakuan diulang 4 kali. Uji lanjut pengaruh iradiasi dan media induksi kalus dilakukan dengan menggunakan Uji berganda Duncan taraf 5 %. Peubah yang diamati meliputi : Persentase embrio yang hidup, diameter kalus yang terbentuk, jumlah induksi bakal tunas dan warna kalus.
2. SeleksiIn vitro pada media Al dan pH rendah
Pada percobaan ini media seleksi Al yang akan digunakan terlebih dahulu disiapkan. Media yang digunakan adalah media MS yang dimodifikasi ditambah dengan Al dalam bentuk AlCl3.6H2O dengan konsentrasi sesuai perlakuan, selanjutnya media ditera pada pH 4 (Mariska et al, 2004). MS yang dimodifikasi dimaksudkan untuk memunculkan sifat toksisitas dari Al pada media seleksi, caranya garam-garam makro dari media MS dimodifikasi (MS md), yaitu kandungan NH4NO3 ditingkatkan dari 1.650 mg/l menjadi 2.400 mg/l, CaCl2.2H2O diturunkan dari 440 mg/l menjadi 15 mg/l, dan KH2PO4 diturunkan dari 170 mg/l menjadi 13 mg/l. Sebagai sumber Fe digunakan FeSO4 28 mg/l.
Kalus embriogenik yang dihasilkan dari percobaan sebelumnya, kemudian diseleksi secaraIn vitro pada media seleksi Al dan pH 4. Caranya kalus dipindahkan ke dalam botol yang mengandung media seleksi Al tersebut, selanjutnya botol disimpan dalam ruang kultur dengan temperatur ruangan 19oC - 21oC dengan intensitas penyinaran 16 jam per hari.
Percobaan menggunakan Rancangan Acak Lengkap (RAL) dengan 2 faktor. Faktor pertama yaitu dosis iradiasi dengan 5 taraf : 0, 250, 500, 750 dan 1000 rad. Faktor kedua adalah media MS modifikasi (Ms md) dengan konsentrasi perlakuan AlCl3.6H2O (ppm) yaitu :
Setiap perlakuan diulang berdasarkan jumlah kalus yang dihasilkan dari percobaan sebelumnya. Uji lanjut pengaruh iradiasi dan media seleksi Al dilakukan dengan menggunakan Uji berganda Duncan taraf 5 %.
Percobaan 3. Regenerasi kalus terseleksi AlCl3.6H2O
Kalus-kalus dari percobaan kedua yang dapat membentuk bakal tunas (organogenesis) dan kalus yang masih hidup meskipun tidak membentuk bakal tunas kemudian diregenerasikan pada media regenerasi MS yang ditambah dengan Vitamin Morel dan Weitmore (VMW), diperkaya dengan BAP 3 mg/l dan Ekstrak malt 500 mg/l (Husni, 2007). Untuk mendapatkan tunas yang banyak dan vigor maka tunas yang terbentuk dapat disubkulturkan pada media yang sama setiap 4 minggu.
Percobaan ini menggunakan Rancangan Acak Lengkap (RAL) dengan 2 faktor yaitu berdasarkan taraf dosis iradiasi dan seleksi in vitro pada media Al dan pH rendah (hasil dari percobaan 2). Peubah yang diamati yaitu : Jumlah kalus beregenerasi, jumlah tunas, tinggi tunas, jumlah daun, jumlah akar dan visual pertumbuhan kalus. Pada percobaan ini dilakukan pula analisis kromosom pada varian jeruk yang tumbuh sempurna untuk melihat jumlah kromosom jeruk JC setelah diiradiasi dan diseleksi pada media seleksi Al. Analisis dilakukan pada bagian meristem akar yang baru tumbuh karena bagian ini masih aktif membelah. Prosedur analisis kromosom menggunakan Metode Pra-Perlakuan Lengkap (Sastrosumarjo, 2006) dapat dilihat pada Lampiran 7.
Gambar 3 Kerangka Alur penelitian 1. Mutasi iradiasi dan
perbanyakan kalus embriogenik
Varian toleran Al dan pH rendah 3. Regenerasi kalus terseleksi AlCl3.6H2O
2. SeleksiIn vitro pada media Al dan pH
rendah Eksplan tanaman (embrio) dari biji buah
HASIL DAN PEMBAHASAN
1. Mutasi Iradiasi dan induksi kalus embriogenik
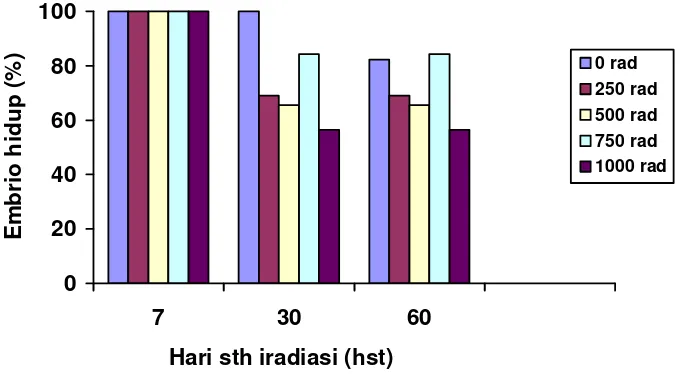
Mutasi iradiasi sinar gamma yang dilakukan pada embrio muda dengan taraf dosis 0, 250, 500, 750 dan 1000 rad memperlihatkan pengaruh yang berbeda terhadap persentase embrio yang hidup sejak umur 7 hari, 30 hari dan 60 hari setelah iradiasi (Gambar 4). Hasil pengamatan menunjukkan bahwa mutasi iradiasi sinar gamma yang dilakukan pada embrio muda dengan taraf dosis berbeda ini dapat menurunkan persentase embrio yang hidup. Dosis iradiasi yang paling tinggi yaitu 1000 rad pada umur 30 hari setelah iradiasi menyebabkan kematian sebagian besar eksplannya, sehingga persentase embrio yang hidup paling rendah dibandingkan dengan dosis iradiasi yang lebih rendah. Namun penurunan persentase embrio yang hidup sejak 30 hari setelah iradiasi tidak menunjukkan penurunan yang signifikan. Hal ini mengindikasikan bahwa pengaruh iradiasi sinar gamma terhadap persentase eksplan yang hidup tidak menunjukkan perbedaan yang berarti pada umur eksplan sejak 30 hari setelah iradiasi.
Gambar 4 Persentase embrio yang hidup pada berbagai tingkatan dosis iradiasi sinar gamma umur 7, 30 dan 60 hari setelah iradiasi.
mg/l + BAP 0,1 mg/l menunjukkan nilai diameter kalus yang paling besar (0.95 cm) walaupun hasilnya tidak berbeda nyata dengan media MS + VB5 + 2,4-D 0,5 mg/l + BAP 0,05 mg/l (0.94 cm). Diameter kalus terkecil 0.35 cm diperoleh dari dosis iradiasi 1000 rad meskipun dikulturkan pada media MS + VB5 + 2,4-D 0,5mg/l + BAP 0,1 mg/l.
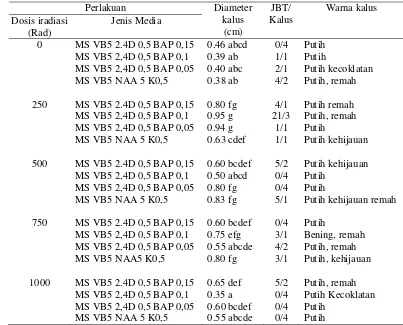
Tabel 1 Pengaruh dosis iradiasi sinar gamma dan jenis media terhadap diameter kalus, jumlah bakal tunas dan warna kalus
Perlakuan
Keterangan : Angka yang diikuti oleh huruf yang sama pada kolom yang sama tidak berbeda nyata pada uji Duncan taraf 5%.
2,4 Dichlorophenoxyacetic Acid (2,4-D) ; Benzil Adenin Purin (BAP) ; Napthalene Acetic Acid (NAA) danKinetin(K) dalam satuan mg/l. JBT = jumlah bakal tunas.
sinar gamma ditingkatkan walaupun dikulturkan pada media yang mengandung rasio auksin dan sitokinin yang cukup seimbang, formulasi ini tidak dapat menginduksi dan meningkatkan pertumbuhan sel-sel pada kalus. Peningkatan dosis iradiasi pada embrio muda ternyata dapat menurunkan kemampuan pertumbuhan sel-selnya dalam membentuk kalus. Charbaji dan Nabulsi (1999), mengungkapkan bahwa iradiasi sinar gamma dapat menghambat pembelahan dan pertambahan jumlah sel. Kalus yang mengalami kerusakan sel dapat mematikan selnya serta menurunkan kemampuan regenerasinya (Biswas,et al. 2002).
Pada Tabel 1 juga tampak jumlah induksi bakal tunas pada kalus embrio mutan memberikan hasil yang lebih banyak dibandingkan dengan kalus embrio yang tidak diiradiasi. Jumlah bakal tunas terbanyak adalah 21 bakal tunas dari kalus embrio mutan 250 rad pada media MS + VB5 +2,4-D 0,5 mg/l + BAP 0,1 mg/l. Hasil ini mendukung hasil penelitian Triatminingsih dan Karsinah (2004) pada jeruk JC, dimana induksi tunas terbanyak juga terbentuk pada media MS yang dikombinasikan dengan 2,4-D dan BAP 0,1 mg/l. Menurut Ling and Iwamasa (1997), bahwa peningkatan BAP pada batas tertentu dapat memacu kecepatan proliferasi kalus.
Secara keseluruhan jumlah induksi bakal tunas adalah dari kalus embrio yang tidak diiradiasi (7 bakal tunas), embrio mutan 250 rad (27 bakal tunas), embrio mutan 500 rad dan 750 rad (masing-masing10 bakal tunas) dan 1000 rad (5 bakal tunas). Hasil ini mengindikasikan bahwa sampai dosis iradiasi sinar gamma 750 rad kemampuan regenerasi eksplan masih baik, tetapi pada dosis 1000 rad terjadi kerusakan dan penurunan kemampuan regenerasinya. Hasil ini sejalan dengan penelitian Kosmiatin (2007), dimana dengan perlakuan dosis iradiasi yang sama pada kalus jeruk, menunjukkan peningkatan kemampuan regenerasi sampai dosis 750 rad dan terjadi penurunan kemampuan regenerasi pada dosis yang lebih tinggi yaitu 1000 rad.
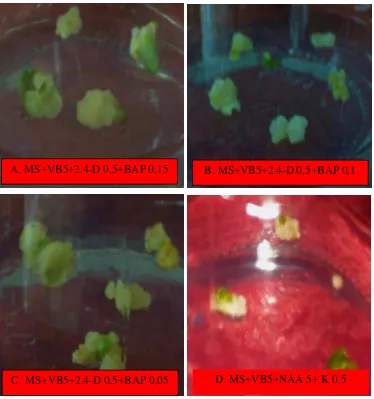
Gambar 5 Pembentukan dan pertumbuhan kalus embriogenik pada beberapa jenis media
Secara umum, induksi kalus embriogenik dari embrio yang diiradiasi (embrio mutan) dan yang tidak diiradiasi setelah dikulturkan pada berbagai formulasi media perlakuan menunjukkan pembentukan kalus yang hampir sama (Gambar 6). Hasil pengamatan secara visual terlihat bahwa kalus yang terbentuk dari embrio mutan maupun embrio yang tidak diiradiasi umumnya berwarna putih hingga putih kekuningan, remah, jika terinduksi bakal tunasnya nampak kalus berwarna putih kehijauan (membentuk spot-spot warna hijau).
A. MS+VB5+2.4-D 0,5+BAP 0,15
C. MS+VB5+2.4-D 0,5+BAP 0,05
B. MS+VB5+2.4-D 0,5+BAP 0,1
Gambar 6 Pembentukan kalus : (A) embrio mutan ; (B) embrio yang tidak diiradiasi (C) Embrio yang terinduksi bakal tunas dan (D) Embrio yang tidak membentuk kalus
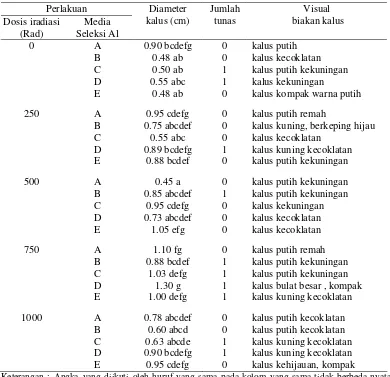
2. Seleksi In vitro pada media Al dan pH rendah
Hasil seleksiIn vitro pada media yang mengandung AlCl3.6H20 dengan pH 4 (asam) terlihat bahwa kalus yang berasal dari embrio mutan 250, 500 dan 750 rad yang diseleksi pada media seleksi AlCl3.6H20 250 ppm menunjukkan persentase kalus yang hidup paling tinggi (100%) dibanding dengan kalus yang berasal dari embrio yang tidak diiradiasi. Sedangkan untuk embrio mutan 1000 rad menunjukkan tingkat kehidupan kalus yang paling rendah walaupun dikulturkan pada media perlakuan dengan dosis AlCl3.6H20 yang paling rendah (Gambar 7). Hasil ini diduga
A
C
B
bahwa dosis iradiasi yang tinggi dapat menyebabkan kerusakan sel-sel dan mengakibatkan kematian sel-sel jaringan sehingga sel-sel tidak dapat bertahan pada perlakuan Al.
Media seleksi Al juga memberikan pengaruh yang berbeda terhadap persentase kalus yang hidup baik pada embrio mutan maupun embrio yang tidak diiradiasi. Makin tinggi konsentrasi Al, cenderung menurunkan persentase kalus yang hidup. Menurut Sutjahjo (2006), bahwa kalus-kalus yang tidak tenggang pada konsentrasi Al yang tinggi akan mengalami kematian sejak dini.
0
Konsentrasi AlCl3.6H2O (ppm)
Kalu
Gambar 7 Persentase kalus yang hidup pada beberapa media seleksi Al, 4 MSP
Tabel 2 Pengaruh dosis iradiasi sinar gamma dan media seleksi Al terhadap diameter kalus, jumlah tunas dan visual biakan kalus
Perlakuan
Keterangan : Angka yang diikuti oleh huruf yang sama pada kolom yang sama tidak berbeda nyata pada uji Duncan taraf 5%.
Media A = MS md tanpa AlCl3.6H20 ; B = MS md + AlCl3.6H20 250 ppm ; C = MS md + AlCl3.6H20 500 ppm ; D = MS md + AlCl3.6H20 750 ppm ; E = MS md + AlCl3.6H20 1000 ppm . MS md = MS yang dimodifikasi
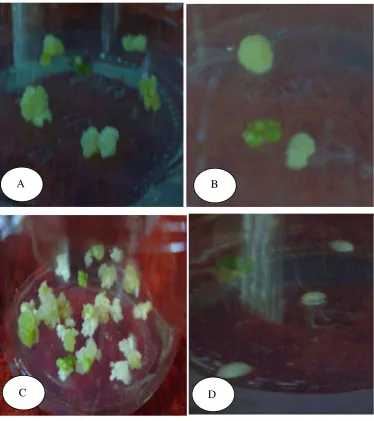
Secara visual perubahan warna kalus pada saat diseleksi dengan Al dan pH rendah juga berbeda-beda, umumnya berwarna putih kekuningan hingga kecoklatan (Gambar 8).
Gambar 8 Respon pertumbuhan kalus pada beberapa media seleksi Al
C. AlCl3.6H2O 500 ppm
D. AlCl3.6H2O 750 ppm E. AlCl3.6H2O 1000 ppm
Secara umum kalus melakukan mekanisme pertahanan terhadap cekaman, salah satunya dengan pembentukan senyawa fenol yang biasanya diikuti dengan perubahan warna kalus menjadi coklat (George dan Sherrington, 1984). Menurut Mariskaet al, (2004), Respon yang berbeda pada kalus yang diseleksi Al bergantung pada ketahanan masing-masing sel terhadap komponen seleksi serta taraf konsentrasi Al. Sel-sel yang tidak mempunyai sifat ketahanan berwarna hitam serta tidak mampu tumbuh, sedangkan sel-sel yang tahan tetap tumbuh dan berwarna hijau.
Hasil penelitian juga memperlihatkan pertumbuhan kalus yang diseleksi pada media seleksi Al yang lebih tinggi cenderung menghasilkan kalus non embriogenik, akibatnya hanya diperoleh sedikit kalus yang embriogenik. Menurut Paterson dan Smith (1991), karakteristik kalus yang embriogenik adalah kalus yang berwarna putih hingga kekuningan, mengkilat dan remah sehingga mudah dipisahkan membentuk fragmen. Kalus yang masih bersifat embriogenik dalam media seleksi Al memungkinkan dapat diregenerasikan menjadi tanaman lengkap, sehingga regeneran yang dihasilkan dari kalus tersebut diharapkan dapat membawa sifat yang toleran terhadap cekaman Al dan pH masam.
Percobaan 3. Regenerasi kalus terseleksi AlCl3.6H2O
Percobaan regenerasi kalus terseleksi Al (hasil percobaan 2), pada media regenerasi MS + VMW + BAP 3 mg/l + Ekstrak malt 500 mg/l, menunjukkan respon jumlah kalus beregenerasi yang berbeda. Pada Tabel 3 terlihat bahwa kalus yang diregenerasikan tidak semuanya dapat beregenerasi. Hal ini kemungkinan disebabkan karena terjadinya dediferensiasi dimana masa sel terus membelah dan tidak terorganisasi. Kemungkinan lain kalus telah mengalami kerusakan atau penurunan kemampuan regenerasi karena pengaruh perlakuan iradiasi dan seleksi Al (Biswaset al, 2002). Menurut Purnamaningsih dan Mariska (2003) kesulitan yang sering dihadapi dalam seleksiin vitro adalah sulitnya meregenerasikan massa sel yang tahan Al dan pH rendah, padahal regenerasi tanaman sangat penting untuk pembuatan klon yang mampu mempertahankan susunan genetik sehingga diperoleh tanaman yang tahan pada cekaman Al dan pH rendah.
diiradiasi masing-masing 7 kalus. Dari kalus embrio mutan 750 rad sebanyak 6 kalus, kalus embrio mutan 500 rad sebanyak 5 kalus dan jumlah kalus beregenerasi paling sedikit diperoleh dari kalus embrio mutan 1000 rad sebanyak 3 kalus. Hasil ini mengindikasikan bahwa kalus dari embrio yang diiradiasi dengan dosis yang lebih rendah (sampai 750 rad) masih memiliki kemampuan beregenerasi yang lebih baik sedangkan dosis iradiasi yang tinggi akan mematikan bahan yang dimutasi atau mengakibatkan sterilitas (Soedjono, 2003).
Tabel 3 Jumlah kalus beregenerasi pada media regenerasi MS + VMW + BAP 3 mg/l + Ekstrak malt 500 mg/l
Perlakuan Awal Jumlah Jumlah Kalus
Kalus-kalus yang dapat beregenerasi disubkultur kembali pada media yang sama untuk menghasilkan tunas. Hasilnya menunjukkan bahwa kalus-kalus yang beregenerasi semuanya dapat menghasilkan tunas. Menurut Husni (2007), formulasi media yang digunakan pada percobaan ini merupakan media yang terbaik untuk regenerasi jeruk siam.
Tabel 4 Jumlah tunas dan Rerata tinggi tunas pada media regenerasi MS + VMW + BAP 3 mg/l + Ekstrak malt 500 mg/l setelah 4 kali subkultur
Perlakuan Awal JT/ JKB Rerata Visual pertumbuhan
Keterangan : MS md = MS modifikasi ; JT = jumlah tunas ; JKB = jumlah kalus beregenerasi ; TT = tinggi tunas
memacu pertumbuhan kalus-kalus mutan dan kalus yang tidak diiradiasi untuk membentuk tunas. Pemberian sitokinin dengan konsentrasi yang cukup tinggi akan menyebabkan terjadinya pemanjangan dan pembesaran sel yang lebih mengarah pada terbentuknya tunas (Gaba, 2002). Hasil penelitian Triatminingsih dan Karsinah (2004), ekstrak malt yang ditambahkan pada media regenerasi dapat meningkatkan jumlah tunas jeruk JC yang terbentuk.
Sementara rerata tinggi tunas dari kalus yang diregenerasikan pada media MS + VMW + BAP 3 mg/l + Ekstrak malt 500 mg/l paling tinggi 1.00 cm masing-masing dihasilkan dari embrio mutan 500 rad pada perlakuan AlCl3.6H2O500 ppm, embrio mutan 750 rad pada perlakuan AlCl3.6H2O0 ppm dan 250 ppm serta embrio mutan 1000 rad pada perlakuanAlCl3.6H2O750 ppm.
Tabel 5 Jumlah daun dan jumlah akar yang terbentuk pada media regenerasi MS + VMW + BAP 3 mg/l + Ekstrak malt 500 mg/l setelah 4 kali subkultur
Pada Tabel 5, jumlah daun yang terbentuk dari masing-masing tunas menunjukkan respon yang berbeda, rata-rata 1-2 daun yang terbentuk per tunas. Selain membentuk daun dan tunas, beberapa varian jeruk sudah terinduksi akarnya. Jumlah akar yang terbentuk umumnya dihasilkan dari varian terseleksi Al dengan konsentrasi AlCl3.6H2O yang lebih rendah ( 500 ppm) dengan rata-rata panjang akar yang terbentuk berkisar 0,5 – 1,0 cm. Hal ini mengindikasikan bahwa varian jeruk tersebut toleran terhadap cekaman Al yang diberikan. Menurut Samuelet al., (1997) salah satu kriteria tanaman yang toleran terhadap cekaman Al yaitu akar mampu untuk tumbuh terus dan ujung akarnya tidak mengalami kerusakan.
Pada Tabel 5 juga menunjukkan beberapa varian jeruk yang lain tidak terinduksi akarnya dan cenderung menunjukkan pertumbuhan yang abnormal. Hal ini mengindikasikan terjadinya penurunan kemampuan regenerasi dan menunjukkan respon morfologi akibat cekaman Al. Menurut Sutjahjo (2006), pada tanaman yang diberi cekaman Al, gejala yang umum dapat dilihat yaitu terjadinya penghambatan pertumbuhan akar akibat pengaruh keracunan Al. Hasil penelitian Sivaguru et al (2003), menunjukkan bahwa perlakuan Al pada tanaman arabidopsi (ecotype Landsberg-O) dapat menghambat 50% pertumbuhan akarnya, ujung akar dan akar cabang menebal sehingga serapan dan translokasi unsur-unsur hara terganggu. Hasil penelitian Yamamoto et al., (1992) mendapatkan bahwa toksisitas Al selain mengakibatkan tanaman kekurangan nutrien juga mengubah struktur dan fungsi dari membran plasma dan menghalangi pembelahan sel pada ujung-ujung akar. Pada akhirnya tanaman akan mengurangi sistem perakarannya dan menunjukkan berbagai gejala kekurangan nutrien akibat keracunan Al (MacDiarmid dan Gardner, 1996).
Gambar 9 Pertumbuhan tunas embrio mutan 750 rad yang terseleksi Al pada konsentrasi (A) AlCl3.6H2O 0 ppm ; (B) AlCl3.6H2O 250 ppm ; dan (C) AlCl3.6H2O 1000 ppm.
Embrio mutan 750 rad yang terseleksi pada media seleksi Al yang lebih rendah (AlCl3.6H2O 250 ppm) juga memperlihatkan pertumbuhan yang lebih baik dibanding dengan embrio yang tidak diiradiasi dan embrio mutan lainnya (Gambar 10). Hasil penelitian Yuliasti (2007), pada tanaman kedelai menunjukkan bahwa penambahan konsentrasi Al dapat menghambat pertumbuhan tanamannya.
A B
Gambar 10 Pertumbuhan tunas embrio yang tidak diiradiasi dan embrio mutan yang terseleksiAlCl3.6H2O500 ppm
Analisis jumlah kromosom pada beberapa varian yang terseleksi Al menunjukkan jumlah kromosom yang sama dengan kromosom individu normalnya yaitu 18 (Gambar 11). Hal ini menunjukkan bahwa tidak terjadi mutasi jumlah kromosom akibat mutasi iradiasi sinar gamma dan seleksiIn vitro pada media seleksi Al, tetapi kemungkinan terjadi mutasi titik sehingga perubahan yang terjadi pada varian jeruk JC dapat meningkatkan ketahanan terhadap Al dan pH rendah. Menurut Taiz dan Zeiger (2002), Peningkatan ini kemungkinan disebabkan perubahan ekspresi gen pada kultur. Perubahan susunan ekspresi gen akan mengakibatkan perubahan fenotip tanaman (Kadir, 2007).
A. Tunas embrio yang tidak diiradiasi B. Tunas embrio mutan 500 rad
Gambar 11 Hasil analisis kromosom varian dari embrio yang tidak diiradiasi dan embrio mutan yang terseleksi Al
Menurut Duncan (1997), iradiasi sinar gamma dapat menyebabkan ketidakseimbangan fungsi fisiologis tanaman. Iradiasi sinar gamma dapat merusak DNA sehingga menyebabkan terjadinya perubahan ekspresi gen. Semakin tinggi dosis iradiasi sinar gamma, semakin meningkatkan efek kerusakan pada DNA, bahkan
A. 0 rad AlCl3.6H2O 500 ppm
C. 750 rad AlCl3.6H2O 0 ppm
B. 500 rad AlCl3.6H2O 500 ppm
D. 750 rad AlCl3.6H2O 250 ppm
dosis tertentu dapat mengakibatkan perubahan struktur kromosom atau aberasi kromosom (Harten, 1998). Sedangkan Hao and Deng (2002), melakukan analisis sitologi dan molekuler menemukan penyimpangan genetik (mitotic irregulated) pada tanaman jeruk, seperti pemanjangan dan pemendekan kromosom serta ketidakseimbangan kromosom sehingga tanaman yang diregenerasikan dari kalus yang dikultur dalam waktu lama (“long term culture”) akan mengalami abnormalitas.
SIMPULAN
1. Keragaman genetik batang bawah jeruk meningkat dengan induksi mutasi iradiasi sampai 750 rad dan seleksiIn vitro pada media MS + VB5 +2.4D 0,5 mg/l + BAP 0,05 - 0,15 mg/l.
2. Dihasilkan sebanyak 65 varian jeruk kandidat toleran Al dan pH rendah, masing-masing dari embrio mutan 250 rad (19 varian), 500 rad (15 varian), 750 rad (9 varian) dan 1000 rad (22 varian).
3. Perlakuan mutasi iradiasi 750 rad dan media seleksi Al (AlCl3.6H2O 250 ppm), menghasilkan varian jeruk kandidat toleran Al dan pH rendah dengan respon pertumbuhan yang paling baik.
SARAN
1. Perlu dilakukan analisis Al dan asam organik untuk mengetahui mekanisme ketahanan terhadap cekaman Al.