ELIS NINA HERLIYANA
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Dengan ini saya menyatakan bahwa disertasi yang berjudul: “Potensi Ligninolitik Jamur Pelapuk Kayu Kelompok Pleurotus” adalah gagasan atau hasil penelitian disertasi karya saya sendiri dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apapun pada perguruan tinggi manapun. Semua data dan informasi yang digunakan telah dinyatakan secara jelas dan dapat diperiksa kebenarannya.
Bogor, Oktober 2007
© Hak cipta milik Institut Pertanian Bogor, tahun 2007
Hak cipta dilindungi
1. Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumber
a. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik atau tujuan suatu masalah
b. Pengutipan tidak merugikan kepentingan yang wajar untuk IPB
ELIS NINA HERLIYANA. Potensi Ligninolitik Jamur Pelapuk Kayu Kelompok Pleurotus. Dibimbing oleh DODI NANDIKA, ACHMAD, LISDAR I. SUDIRMAN dan ARIEF BUDI WITARTO.
Jamur diketahui mampu menguraikan substrat menjadi bahan-bahan organik sederhana melalui proses hidrolisis enzimatik, sehingga dapat dimanfaatkan oleh mikroorganisme lain termasuk tumbuhan. Beberapa jamur pelapuk kayu seperti jamur kelompok Pleurotus berpotensi dalam proses biodelignifikasi, yang dapat dimanfaatkan oleh industri pulp dan kertas. Meskipun demikian kajian lebih mendalam belum banyak dilakukan terhadap jamur kelompok Pleurotus khususnya yang berasal dari Indonesia. Penelitian ini merupakan penelitian dasar yang menyeluruh tentang jamur pelapuk kayu kelompok Pleurotus asal Bogor.
Tujuan utama penelitian ini adalah untuk mengetahui potensi ligninolitik jamur pelapuk putih dari kelompok Pleurotus asal Bogor. Penelitian ini dimulai dengan eksplorasi jamur kelompok Pleurotus di lapangan, isolasi, karakterisasi fisiologi dan ligninolitik pada beberapa chip kayu bahan baku pulp yaitu kayu akasia, pinus dan sengon dan media serbuk gergajian kayu sengon, pengukuran aktivitas enzim ligninase (manganese peroksidase/MnP, lignin peroksidase/LiP dan lakase) dan purifikasi MnP serta terakhir adalah identifikasi secara morfologi yang ditunjang dengan karakter kultur dan fisiologi.
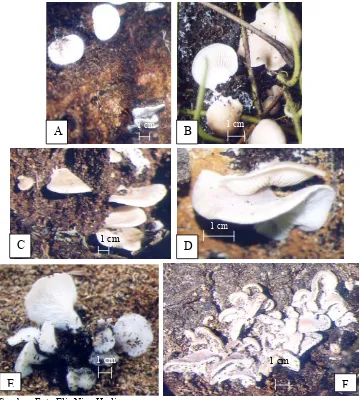
Hasil eksplorasi jamur di berbagai wilayah di Bogor menunjukkan bahwa jenis jamur pelapuk kayu kelompok Pleurotus hanya ditemukan di areal kebun dan tempat penggergajian kayu di kecamatan Ciherang, Bogor (+ 501 m dpl). Sebanyak 24 isolat kelompok Pleurotus yang berhasil diisolasi dari lapangan dan 17 isolat diantaranya kemudian dikultivasi pada media serbuk gergajian kayu sengon. Sebanyak 6 isolat diantaranya dapat membentuk tubuh buah yaitu Pleurotus EB9 (pink), Pleurotus
EB14-2 (cokelat-muda), Pleurotus EB24 (cokelat keabu-abuan), Pleurotus EA4 (cokelat muda), Pleurotus EAB7 (cokelat keabu-abuan), dan Pleurotus EB6 (cokelat keabu-abuan).
P. ostreatus HO merupakan isolat yang paling tinggi laju dekomposisinya kemudian diikuti Pleurotus EB9.
Hasil pengamatan mikroskopik menunjukkan bahwa pada tahap awal kolonisasi oleh jamur pada chip, diketahui miselium mendiami saluran resin dan jari-jari kayu pinus. Hasil pengamatan terhadap kayu akasia terlihat bahwa miselium mendiami pembuluh dan jari-jari. Umumnya miselium melakukan penetrasi melalui noktah-noktah sebagai salah satu cara penyebaran ke sel-sel kayu yang lain. Proses pelapukan selanjutnya terlihat pada kerusakan dan perubahan bentuk pada lamela tengah dan dinding sel sekunder.
Kadar zat-zat ekstraktif total secara umum meningkat setelah diinokulasi oleh masing-masing isolat kelompok Pleurotus baik pada fase vegetatif maupun reproduktif. Setiap isolat kelompok Pleurotus yang diuji menunjukkan variasi rata-rata penurunan kadar lignin (10,7-89,7%) dan selulosa (18,9-87,4%). Pleurotus EB9 mempunyai kemampuan menurunkan kadar lignin substrat terbesar (89,7%) dan kadar selulosa terbesar (87,4%). Antar satu isolat dengan isolat lainnya mempunyai karakter ligninolitis yang berbeda. Diantara isolat yang diuji, Pleurotus EB9 merupakan satu isolat kandidat yang baik untuk agens biopulping dan biobleaching.
Selanjutnya ekspresi MnP setelah 6 hari menunjukkan bahwa Pleurotus EB9 memproduksi MnP paling tinggi pada substrat kayu sengon. Pleurotus EB9 juga memproduksi lakase, sementara produksi LiP tidak terdeteksi. SDS-PAGE hasil kromatografi kolom penukar ion (DEAE-Sepharose) terhadap isolat Pleurotus EA4 dengan pemekatan 40% amonium sulfat, ternyata tidak diperoleh pita-pita yang menunjukkan keberadaan MnP. Hal ini menunjukkan bahwa Pleurotus EA4 tidak memproduksi MnP.
SDS-PAGE hasil kromatografi kolom penukar ion pada contoh Pleurotus EB9 dengan pemekatan 40% amonium sulfat, diperoleh pita dengan bobot molekul 43 kDa. Hasil pemurnian dengan kolom kromatografi gel diperoleh puncak pada fraksi ke 96. SDS PAGE hasil kromatografi kolom gel terhadap sampel pemekatan 40% amonium sulfat dari Pleurotus EB9 dipastikan bahwa Pleurotus EB9 menghasilkan MnP yang telah berhasil dimurnikan. Hasil rangkuman data pemurnian MnP dari
Pleurotus EB9 menunjukkan bahwa tingkat keberhasilan pemurnian masih sangat rendah dengan tingkat kemurnian 3,8. Aktivitas spesifik MnP yang diperoleh adalah sebesar 4,133 U/mg protein.
Hasil identifikasi terhadap enam isolat yang dapat membentuk tubuh buah diketahui lima isolat (Pleurotus EB14-2, EB24, EA4, EAB7 dan EB6) adalah
Hohenbuehelia petaloides dan satu isolat (Pleurotus EB9) adalah Pleurotus djamor
EB9. Analisis kelompok isolat berdasarkan karakterisasi fisiologi dan morfologi menunjukkan bahwa P. djamor EB9 berbeda dengan kelima isolat H. petaloides
tersebut. Produksi enzim ligninase khususnya MnP P. djamor EB9 dan H. petaloides
EA4 juga berbeda.
Kata-kata Kunci: Jamur pelapuk kayu, kelompok Pleurotus, biodegradasi, kayu akasia, pinus dan sengon, P. djamor EB9, H. petaloides,
ELIS NINA HERLIYANA. Ligninolytic Potency of Wood Rot Fungi of
Pleurotus Groups. Under Supervision of DODI NANDIKA, ACHMAD, LISDAR I. SUDIRMAN and ARIEF BUDI WITARTO.
Pleurotus groups of white rot fungi are wood decaying fungi, which able to decompose wood substrate into simple organic materials through enzymatic hydrolytic. This can be used by other microorganisms including plants. They are also potential used in biodelignification process, especially for pulp and paper industry. So far, study on Pleurotus groups especially from Indonesia in detail has not been done. The present study is a comprehensive basic research for white-rot fungi Pleurotus groups especially from Bogor.
The purpose of this research is to examine the ligninolytic potency of
Pleurotus groups from District and City of Bogor. The research activities were started with exploration of Pleurotus in the field, isolation, physiological characterization test, ligninolytic activity, measurement and purification of enzyme produced by Pleurotus groups and finally, identification of isolated
Pleurotus groups morphologically and physiologically. The physiological characterization was done based on the growth rate, oxidation reaction, vegetative and reproductive phases, total biomass and biological efficiency. The ligninolytic character of these six fungi isolates was measured based on biodegradation of sengon sawdust substrate by the fungi and degradation value on acacia, pine and sengon wood. The ligninolytic characterization also was identified through enzyme activity of manganese peroxidase (MnP), laccase and lignin peroxidase (LiP). Furthermore, MnP enzyme from isolate represented Bogor was isolated and purified.
The results of field observation showed that Pleurotus groups were found only in garden and sawmill of sub-district Ciherang (+ 501 m asl). Twenty four isolates of Pleurotus groups were isolated, and 17 of those isolates were cultivated on sengon sawdust media. Among them, six isolates are able to form fruit bodies, namely: Pleurotus EB14-2 (light brown), Pleurotus EB24 (gray brown), Pleurotus
EA4 (light brown), Pleurotus EAB7 (gray brown), Pleurotus EB6 (gray brown) and Pleurotus EB9 (pink).
The results showed that performance of colony of Pleurotus groups on various culture media, temperatures and pH as variables were varies. Positive oxidation reaction on AAG and AAT media showing that all isolates including white-rot fungi. Pleurotus EB9 and Pleurotus EA4 were potentially white-rot fungi with moderately strong reaction on media AAG and AAT. Wild Pleurotus
showed that in the early stage of fungi invasion on pine, mycelium lived in resin tunnel and xyllary rays, while on acacia they lived in vessels and xyllary rays. In general, penetration of mycelium through nocti was as a way to spread further in another of wood cells. On the advanced decay process, there were changes in form and cells damage of middle lamella and secondary cell wall.
Having applied of each wild Pleurotus group isolates resulted in increasing of extractives total compounds both in vegetative and reproductive phases. Each wild Pleurotus group isolates shows variation in decreasing average of lignin content (10,7-89,7%) and cellulose content (18,9-87,4%). Pleurotus EB9 are able to decrease the highest lignin (89,7%) and cellulose (87,4%) content of substrate. Classification based on ligninolytic character is different with classification based on morphological and physiological characters. Pleurotus
EB9 seems to be separated from other isolates. This shows that there was different ligninolytic character among the isolates. The best isolate for biopulping and biobleaching agent is Pleurotus EB9 on vegetative phase.
The expression of MnP (U/ml) after six days showed that, extracellularly,
Pleurotus EB9 was the highest in producing MnP on sengon-wood substrate in swing medium condition. Pleurotus EB9 was also produced laccase, while LiP was not detected. The SDS-PAGE chromatography result of ionic exchange column (DEAE-Sepharose) of Pleurotus EA4 sample with 40% solid concentration of ammonium sulphate did not produce ribbons that indicate the presence of MnP. Its activity did not produce good result too. This fact proves that
Pleurotus EA4 did not produce MnP.
The SDS-PAGE chromatography result of ionic exchange column of
Pleurotus EB9 sample with 40% solid concentration of ammonium sulphate produced a ribbon with 43 kDa molecule weight. Furthermore, the purification result using high resolution HiPrep 16/60 Sephacryl s-200 gel chromatography column (GE Biosciences) with flow rate of 0.3 ml/min and fractionation rate of 1.5, and using buffer solution of 10mM potassium phosphate pH 7.0 and HPLC AKTA Purifier (GE Biosciences), reached a peak at 96th fraction. The SDS PAGE gel column chromatography result of Pleurotus EB9 sample with 40% solid concentration of ammonium sulphate proved that Pleurotus EB9 produced MnP which has been purified with a molecule weight of 43 kDa. The summary result of MnP purification data of Pleurotus EB9 showed that the success level of purification is still low with purification rate of 3.8. The specific activity of MnP obtained is 4.133 U/mg of protein.
The identification result of six isolates that were able to form fruit body are five isolates of Hohenbuehelia petaloides, which are EB14-2, EB24, EA4, EAB7 and EB6, and also Pleurotus djamor EB9. The analysis result of isolate group based on physiological and morphological characters showed that P. djamor EB9 is different with H. petaloides. The production of ligninase enzyme, especially MnP, of P. djamor EB9 and H. petaloides EA4 are also different. Keywords: Wood rot fungi, Pleurotus groups, biodegradation, acacia, pine and sengon
ELIS NINA HERLIYANA
Disertasi
sebagai salah satu syarat untuk memperoleh gelar
Doktor pada
Program Studi Ilmu Pengetahuan Kehutanan
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Penguji pada Ujian Tertutup : Prof. Dr. Ir. Wasrin Syafii
Penguji pada Ujian Terbuka : 1. Prof. Dr. Ir. Yusuf Sudohadi
PRAKATA
Segala puji bagi Allah SWT yang telah memberikan rahmat, karunia, berkah dan hidayah-Nya kepada penulis, sehingga dapat menyelesaikan studi hingga tahap penyusunan disertasi ini.
Penulis menyampaikan rasa terima kasih yang tidak terhingga kepada Prof. Dr. Ir. Dodi Nandika, M.S., Dr. Ir. Achmad, M.S., Dr. Ir. Lisdar Manaf I. Sudirman dan Dr. Ir. Arief Budi Witarto, M.Eng. atas bimbingan, kesabaran, pengkayaan wawasan, kritik, saran dan dukungan moril yang sangat besar peranannya dalam penyelesaian penulisan disertasi ini.
Rasa hormat yang setinggi-tingginya penulis sampaikan kepada kedua orang tua tercinta, ayahanda Atjeng Kamaludin dan ibunda Imas Maryani, juga kepada ayahanda mertua H. Kusno dan ibunda H. Hindun dan keluarga ayahanda Mu’zi (Alm.), yang telah memberi dukungan do’a dan kasih sayangnya. Terima kasih yang mendalam penulis haturkan kepada suami tercinta Dr. Ir. Abdul Munif, M.Sc.Agr. serta ananda tersayang Muhammad Hidayatul Mustofa, Abdullah Fathan Taufik dan Ziyad Muhammad Imani, atas do’a, kasih sayang, pengertian, kesabaran dan pengorbanan serta semua dukungan yang sangat berarti hingga terselesaikannya studi doktor ini. Ucapan terimakasih juga disampaikan untuk adik-adik, Kompol Asep Hendradiana, dr, Sp.An., M.Kes., Dewi Pudjiati, S.I.P., Ir. Deden Hidayatullah, Rita Yustika, Amd, Nizar Budiaji, Amd dan Nando Sugawa, S.T., Ir. Rohmad Mu’zi, Masruroh Mu’zi, Umdzatull Millah Mu’zi, Imron Mudzakir Mu’zi, SE, Sri Rahayu dan Khoiron, SE atas dorongan semangat, pengertian dan do’anya.
Penghargaan dan terima kasih juga disampaikan kepada Prof. Dr. Ir. Wasrin Syafii, yang bersedia menjadi Penguji Luar Komisi pada Ujian Tertutup, Prof. Dr. Ir. Yusuf Sudohadi dan Dr. Siswanto, D.E.A., yang bersedia menjadi Penguji Luar Komisi pada Ujian Terbuka dan Pimpinan Sekolah Pasca Sarjana, Program Studi Ilmu Pengetahuan Kehutanan, Fakultas Kehutanan IPB, Departemen Manajemen Hutan, Departemen Silvikultur, yang telah memberi kesempatan studi. Disampaikan juga ucapan terimakasih kepada tim manajemen BPPS Dikti yang telah memberikan dukungan dana selama perkuliahan dan penelitian.
Finkeldey, Ir. Deded Nawawi, M.Si., dan Prof. Dr. Sipon Mulyadi yang telah bersedia memberikan saran. Ucapan terimakasih juga kepada Dr. Lee Su She dan Dr. Sri Rahayu di UPM Malaysia, serta Dr. Dewi Siti Latifah dan Sarah Reinecke selama di Jerman atas saran dan pustaka-pustaka yang sangat berharga.
Penghargaan dan terimakasih penulis sampaikan kepada Bapak Iwa dari Laboratorium Mikrobiologi dan Biokimia, Pusat Studi Ilmu Hayat, Bapak Engkus dari Rumah Jamur Laboratorium Mikologi di Tajur, Departemen Biologi, Bapak Atien dari Laboratorium Kimia Kayu dan Esti, Amd., S.Si. dari Laboratorium Kayu Solid, Fakultas Kehutanan, IPB; Atie Sunarti, S.T.P., M.S. dan Ir. Desriyani, M.Si. dari Laboratorium Kelompok Penelitian Rekayasa Protein, Pusat Penelitian Bioteknologi dan Dra. Endang dari Laboratorium Anatomi dan SEM, Pusat Penelitian Zoologi, serta Dra. Atik Retnowati, M.Sc. dari Herbarium Bogoriense, LIPI; Ir. Alina, M.Si. dari Laboratorium Protein dan Enzim, Balitbiogen, Bogor; Taufan, Amd. dan Riana dari Laboratorium Bioteknologi dan Biologimolekuler, Lembaga Riset Perkebunan Indonesia, Bogor; Ir. Farid, M.Si. dari Dept. Statistik, FMIPA; Bapak Dr. Widodo beserta staf di Laboratorium Mikologi dan Damayanti, S.P. di Laboratorium Nematologi, Departemen Proteksi Tanaman, Fakultas Pertanian; Ibu Pepi, Ibu Emi dan Ibu Ika di PAU Bioteknologi; Tutin Suryatin, B.Sc.F., M. Alam F., S.Hut., M.Si dan alumni serta anggota Mushroom Studies, Luci Andini, S.Hut., Anang P.H., S.Hut., Osica Ayu, S.Hut., Jenal Mutaqin, S.Hut., Wartaka, S.Hut., Lendi F., S.Hut., Fiki R, S.Hut., Deka F., S.Hut., Alwiah, Ope P., S.Hut., Reni M., S.Hut., Resa, Indah D.P. dan Desti di Laboratorium Penyakit Hutan; Ir. Dorli, M.Si. di Laboratorium Anatomi dan Morfologi Tumbuhan, Departemen Biologi, FMIPA, dan Ir. Noverita, M.Si., Ir. Trisna Priyadi, M.Sc., serta Oemi Fadhillah, S.P., selama pengumpulan data, serta kepada Bapak Sapri, Sukaesih dan keluarga Yuli atas bantuan dan kerjasamanya kepada penulis dan keluarga. Ucapan terima kasih juga disampaikan kepada semua pihak yang tidak dapat disebutkan satu persatu yang telah memberikan bantuan sehingga penelitian dan penyusunan disertasi ini terwujud. Semoga Allah SWT selalu memberikan ampunan dan maghfirah-Nya kepada kita semua, amien.
Semoga Allah SWT membalas seluruh kebaikan yang telah diberikan dan semoga disertasi ini bermanfaat bagi perkembangan ilmu dan pembaca yang memerlukannya.
Bogor, Oktober 2007
Penulis dilahirkan di Cianjur pada tanggal 21 April 1967, sebagai anak pertama dari tujuh bersaudara dari pasangan Atjeng Kamaludin dan Imas Maryani. Penulis menikah dengan Dr. Ir. Abdul Munif, MSc.Agr. pada tahun 1990, dan telah dikaruniai tiga orang putra bernama Muhammad Hidayatul Mustofa (tahun 1991), Abdullah Fathan Taufik (tahun 1996) dan Ziyad Muhammad Imani (tahun 2002).
Pada tahun 1986, penulis menempuh pendidikan sarjana di Institut Pertanian Bogor pada Jurusan Hama dan Penyakit Tumbuhan, Fakultas Pertanian, lulus pada tahun 1990. Pada tahun 1993, penulis melanjutkan pendidikan Magister di Institut Pertanian Bogor pada Program Studi Entomologi/Fitopatologi Program Pascasarjana dan menamatkannya pada tahun 1997. Pada tahun 1998, penulis menjadi peneliti tamu di Institut Planzenkrankheiten, Faculty of Agriculture, University of Bonn, Germany. Pada tahun 2000-2001, penulis mengambil program khusus pada the Faculty of Agriculture, University of Bonn, Germany.
Pada tahun 2002, penulis diterima sebagai mahasiswa Program Studi Ilmu Pengetahuan Kehutanan Sekolah Pascasarjana program Doktor di Sekolah Pascasarjana IPB. Penulis memperoleh beasiswa BPPS dari Ditjen Pendidikan Tinggi, Departemen Pendidikan Nasional Republik Indonesia.
DAFTAR ISI
Hipotesis Penelitian ………..….…. Kebaruan Penelitian ……… TINJAUAN PUSTAKABahan Lignoselulolitik dan Gambaran Umum Tiga Jenis Kayu … Penguraian Komponen Kayu Secara Enzimatik oleh Jamur
Pelapuk Putih ……….…. Jamur Pelapuk Putih Kelompok Pleurotus ……… 1. EKSPLORASI JAMUR PELAPUK PUTIH KELOMPOK
Pleurotus DARI DAERAH BOGOR
Hasil Penelitian ………..… Pembahasan ………... Simpulan ………...
2. KARAKTER FISIOLOGIS JAMUR PELAPUK PUTIH KELOMPOK Pleurotus ASAL BOGOR
3. KARAKTER LIGNINOLITIK KELOMPOK Pleurotus BERDASARKAN BIODEGRADASI PADA TIGA JENIS KAYU BAHAN PULP
Hasil Penelitian ………... Pembahasan ………..….… Simpulan ………...…..
4. KARAKTER LIGNINOLITIK KELOMPOK Pleurotus
BERDASARKAN BIODEGRADASI SUBSTRAT GERGAJIAN KAYU SENGON
Hasil Penelitian ……….. Pembahasan ………... Simpulan ………...………..
5. EKSPRESI ENZIM LIGNINOLITIK KELOMPOK Pleurotus PADA SUBSTRAT CAIR KAYU SENGON SERTA
PEMURNIAN PARSIAL MANGANESE PEROKSIDASE
Abstrak ………....
Abstract ……….….. Pendahuluan ……… Bahan dan Metode ……….. Hasil dan Pembahasan
Hasil Penelitian ………..…………... Pembahasan ………..….… Simpulan ………..……..
6. IDENTIFIKASI BERDASARKAN KARAKTER FISIOLOGIS, LIGNINOLITIK DAN MORFOLOGIS ENAM ISOLAT KELOMPOK Pleurotus ASAL BOGOR
PEMBAHASAN UMUM ………... SIMPULAN DAN SARAN ……….. DAFTAR PUSTAKA ………..….. LAMPIRAN ……….…..
Halaman
Persentase kadar selulosa, hemiselulosa, lignin dan pektin serta pati kayu berdasarkan bobot kering ...
Penelitian 1
Beberapa genus jamur pleurotoid pada beberapa lokasi di Kabupaten dan Kota Bogor pada bulan Mei 2004 ... Isolat kelompok Pleurotus yang ditemukan di daerah Ciherang Bogor ...
Penelitian 2
Media, suhu dan pH optimum, serta reaksi pada AAG dan AAT ketujuh isolat kelompok Pleurotus ... Bobot basah tubuh buah hasil panen jamur ketujuh isolat kelompok Pleurotus ... Nilai rata-rata total panen tubuh buah pada ketujuh isolat jamur isolat jamur Pleurotus ... Perbandingan nilai EB jamur pada ketujuh isolat kelompok
Pleurotus ... Fase vegetatif, fase reproduktif dan jumlah panen jamur isolat kelompok Pleurotus ...
Penelitian 3
Tingkat degradasi pada kayu bahan pulp pinus, akasia dan sengon setelah diinokulasi oleh isolat kelompok Pleurotus ... Rata-rata tingkat degradasi berdasarkan lama inkubasi ... Rata-rata tingkat degradasi berdasarkan isolat ... Rata-rata tingkat degradasi berdasarkan jenis kayu dan lama inkubasi ... Laju dekomposisi pada kayu bahan pulp pinus, akasia dan sengon setelah diinokulasi oleh isolat kelompok Pleurotus ... Tanda-tanda kerusakan kayu secara mikroskopis pada kayu pinus, akasia dan sengon setelah diinokulasi dengan isolat kelompok
Pleurotus ... Penelitian 4
Kadar air dan kadar zat-zat ekstraktif substrat (%) setelah diinokulasi dengan masing-masing isolat kelompok Pleurotus ...
4.2
Kadar lignin, holoselulosa, selulosa dan hemiselulosa substrat (%) setelah diinokulasi dengan masing-masing isolat kelompok
Pleurotus ... Peningkatan zat ekstraktif, penurunan bobot kering, kadar lignin, selulosa dan hemiselulosa pada fase vegetatif dan reproduktif setelah diinokulasi dengan masing-masing isolat kelompok
Pleurotus ... Penelitian 5
Ekspresi enzim MnP (U/ml) isolat-isolat jamur kelompok
Pleurotus secara ekstraseluler ……… Ekspresi MnP (U/ml) setelah 6 hari inkubasi secara ekstraseluler Ekspresi MnP, LiP dan lakase (U/ml) oleh isolat Pleurotus EB9 secara ekstraseluler pada substrat kayu sengon dengan kondisi media digoyang dalam bioreaktor ……….. Ekspresi enzim MnP, LiP dan Lakase (U/ml) secara ekstraseluler oleh isolat Pleurotus EB9 dan Pleurotus EA4 yang sudah dipekatkan dengan amonium sulfat (As) ……… Lama penyimpanan enzim dari mulai panen sampai analisis ekspresi enzim Pleurotus EB9, Pleurotus EA4, Pleurotus EB6, P. chrysosporium PC dan P. ostreatus HO ……….. Lama penyimpanan enzim kasar dari mulai panen sampai analisis ekspresi enzim Pleurotus EB9 dan Pleurotus EA4 ……… Ekspresi MnP, LiP dan lakase (U/ml) Pleurotus EB9 dengan substrat kayu sengon dengan kondisi media digoyang dalam bioreaktor dengan Pleurotus EB9-AS, Pleurotus EB9-F dan isolat
Pleurotus EA4-AS ……… Aktivitas enzim MnP dari Pleurotus EB9 pada tahapan pemurnian parsial ……….
Penelitian 6
Komposisi masing-masing larutan Johansen dan lama perendaman pada metode parafin tahap dehidrasi dan penjernihan Isolat-isolat jamur kelompok Pleurotus yang ditemukan di daerah Ciherang, Bogor ... Tipe koloni kultur ketujuh isolat kelompok Pleurotus pada beberapa jenis media ... Pola kunci H. petaloides (H. petaloides EB14-2, H. petaloides
EB24, H. petaloides EA4, H. petaloides EAB7, H. petaloides
EB6), P. djamor EB9 dan P. ostreatus HO pada media MEA ... Ekspresi enzim MnP, LiP dan Lakase (U/ml) secara ekstraseluler oleh isolat Pleurotus EB9 dan Pleurotus EA4 yang sudah dipekatkan dengan amonium sulfat (As) ……….. Rata-rata jumlah tangkai jamur kelompok Pleurotus pada panen pertama, kedua, ketiga dan keempat ……….
Halaman
A. Pelapukan kayu spruce (Picea abies) oleh jamur pelapuk putih.. B. Bagan kemungkinan jalur pertumbuhan hifa... Mekanisme degradasi selulosa ...
Penelitian 1
Denah lokasi eksplorasi jamur pleurotoid (huruf hitam) dan jenis jamur yang ditemukan (huruf merah)... Kondisi tempat ditemukannya jamur kelompok Pleurotus...
Penampakan tubuh buah Schizophillum spp. di lapangan ... Penampakan tubuh buah Lentinus spp. di lapangan ... Penampakan tubuh buah Panellus spp. di lapangan ... Penampakan tubuh buah Crepidotus spp.di lapangan asal hutan Gunung Luhur Cisarua, Bogor ... Penampakan tubuh buah isolat Pleurotus EA1 sampai EA6 di lapangan ... Penampakan tubuh buah isolat Pleurotus EB6 sampai EA10 di lapangan ... Penampakan tubuh buah isolat Pleurotus EA11 sampai EB16 di lapangan ... Penampakan tubuh buah isolat Pleurotus EB17 sampai EBB28 di lapangan ... Kultur murnikelompok Pleurotus setelah inkubasi 10 hari pada media MEA ... Tubuh buahkelompok Pleurotus di lapangan ... Isolat kelompok Pleurotus yang berhasil membentuk tubuh buah seperti Pleurotus ...
Penelitian 2
Penampakan kultur keenam isolat kelompok Pleurotus dan P. ostreatus HO ... Laju pertumbuhan koloni rata-rata isolat kelompok Pleurotus
pada media, suhu dan pH optimum ... Penampakan visual uji reaksi oksidasi isolat kelompok
Pleurotus setelah tujuh hari inkubasi pada media AAG (atas) dan AAT (bawah) ...
Pleurotus EB24, Pleurotus EB9 dan P. ostreatus HO pada media serbuk gergajian kayu sengon ...
2.5
Tahap-tahap perkembangan Pleurotus EB6 ... Tahap-tahap perkembangan Pleurotus EB9 ... Dendogram berdasarkan karakter fisiologis dari tujuh isolat kelompok Pleurotus asal Bogor...
Penelitian 3
A.Kayu yang digunakan. a. Pinus. b. Akasia. c. Sengon. B. Pengujian biodegradasi kayu pinus ... Pengaruh jenis isolat yang berbeda terhadap rata-rata tingkat degradasi kayu pinus dan akasia ... Bobot kering sisa kayu pinus setelah diinokulasi dengan isolat kelompok Pleurotus ... Bobot kering sisa kayu akasia setelah inokulasi oleh tujuh isolat kelompok Pleurotus ... Bobot kering sisa kayu sengon setelah inokulasi oleh Pleurotus
EB9 dan P. ostreatus HO ... A. Bidang transversal kayu pinus kontrol ... B. Bidang radial kayu pinus setelah diinokulasi dengan P. ostreatus HO. Mikrograf cahaya dan SEM ... Bidang transversal dan tangensial kayu pinus setelah diinokulasi dengan P. ostreatus HO. Mikrograf SEM ... Bidang transversal kayu akasia.... Mikrograf cahaya ... Bidang tangensial kayu akasia.. Mikrograf cahaya dan SEM .... Bidang radial kayu akasia setelah diinokulasi dengan P. ostreatus HO .... Mikrograf SEM ... Bidang radial kayu akasia setelah diinokulasi dengan P. ostreatus HO. Mikrograf SEM ... Bidang transversal dan tangensial kayu sengon ... Mikrograf cahaya ...
Penelitian 4
Bobot kering substrat setelah diinokulasi dengan masing-masing isolat kelompok Pleurotus ... Kadar zat-zat ekstraktif total substrat setelah diinokulasi dengan masing-masing isolat kelompok Pleurotus ... Penurunan kadar lignin substrat setelah diinokulasi dengan masing-masing isolat kelompok Pleurotus ……… Dendogram karakter ligninolitik dari tujuh isolat kelompok
Pleurotus asal Bogor yaitu Pleurotus EB6, P. ostreatus HO,
Pleurotus EB24, Pleurotus EA4, Pleurotus EB14-2, Pleurotus
EAB7 dan Pleurotus EB9 ……….. Dendogram karakter ligninolitik dari tujuh isolat kelompok
Pleurotus asal Bogor yaitu Pleurotus EB14-2, P. ostreatus HO,
Pleurotus EB6, Pleurotus EAB7, Pleurotus EA4, Pleurotus
5.1
Ekspresi MnP, LiP dan lakase (U/ml) oleh isolat Pleurotus EB9 secara ekstraseluler dengan substrat kayu sengon dengan kondisi media digoyang dalam bioreaktor………... Reaksi secara visual pada uji aktivitas MnP (A), LiP (B) dan lakase (C) pada Pleurotus EB9 setelah inkubasi satu sampai 6 hari (H0 sampai H6) dibandingkan dengan kontrol (K)……….. Ekspresi MnP, LiP dan lakase isolat Pleurotus EB9 (A) dan
Pleurotus EA4 (B)……….. Tingkat ekspresi MnP (A), LiP (B) dan lakase (C) Pleurotus
EB6, P. chrysosporium PC dan P. ostreatus HO ……… Tingkat ekspresi MnP (A), LiP (B) dan lakase (C), serta bobot kering miselium (D) Pleurotus EB9 dan Pleurotus EA4 ……… SDS-PAGE hasil kromatografi kolom gel hidrofobik (Phenyl
-Sepharose) terhadap sampel pemekatan 40% amonium sulfat isolat Pleurotus EA4 ……….. SDS-PAGE hasil kromatografi kolom penukar anion (DEAE-Sepharose) terhadap sampel pemekatan 40% amonium sulfat isolat Pleurotus EB9 ……….. Kromatogram pemurnian protein sampel Pleurotus EB9-amonium sulfat 40% dengan gel kromatografi……….. SDS-PAGE hasil kromatografi kolom gel terhadap sampel pemekatan 40% amonium sulfat dari Pleurotus EB9 ………….
Penelitian 6
Penampakan dan tipe koloni kultur. A. H. petaloides EB14-2, B. H. petaloides EB24, C. H. petaloides EA4, D. H. petaloides
EAB7, E. H. petaloides EB6, F. P.djamor EB9, G. P. ostreatus
HO... Ekspresi MnP, LiP dan lakase (U/ml) secara ekstraseluler oleh isolat P. djamor EB9 dengan substrat kayu sengon dengan kondisi media digoyang dalam bioreaktor...
6.14
Penampakan mikroskopik. A. H. petaloides EB14-2. B. H. petaloides EB24. C. H. petaloides EA4. D. H. petaloides EAB7. E. H. petaloides EB6. F. P. djamor EB9. G. P. ostreatus HO .... Sistidia berdinding tipis pada P. djamor EB9 (A) dan Sistidia berdinding tebal (metuloid) pada H. petaloides (B) ... Karakter morfologi pada irisan melintang tudung ... Sistidia pada lapisan gelatin (A dan B) dan Sistidia pada
pileipelisH. petaloides EB6 (C dan D) ... Plot antara panjang dengan lebar basidiospora ketujuh isolat .... Panjang dan lebar basidiospora rata-rata dari tujuh isolat jamur kelompok Pleurotus ... Plot antara panjang dengan lebar basidia ketujuh isolat ... Lebar dan panjang basidia rata-rata dari tujuh isolat jamur kelompok Pleurotus ... Panjang sistidia rata-rata dari tujuh isolat jamur kelompok
Latar Belakang
Industri pulp dan kertas Indonesia memiliki prospek yang cerah dan Indonesia dinilai potensial menjadi salah satu negara produsen terbesar pulp dan kertas dunia. Permintaan pulp dan kertas di tingkat Nasional maupun dunia terus meningkat baik dari segi kualitas produk maupun kuantitasnya. Data Asosiasi Pulp dan Kertas Indonesia (APKI 2006 dalam Tambunan 2006) menunjukkan kapasitas produksi pulp Indonesia tercatat 6,41 juta ton per tahun, 45% di antaranya diekspor, sedangkan kapasitas produksi kertas Indonesia tercatat 10,3 juta ton, 30% di antaranya diekspor ke luar negeri.
Produksi pulp di sebagian besar negara di dunia saat ini masih merupakan pulp kimia yang dihasilkan melalui proses sulfatisasi. Cara kimia tersebut dianggap memberikan banyak keuntungan, diantaranya waktu pemasakan yang relatif singkat dan kualitas pulp yang dihasilkan relatif tinggi. Namun pada saat ini pulp dengan proses sulfat kurang diminati oleh masyarakat, selain karena besar investasi awal dan warna gelap pulp yang dihasilkan, juga karena bahaya pencemaran lingkungan yang dapat ditimbulkan terutama polusi udara. Gas berbau belerang berupa hidrogen sulfida (H2S), metil merkaptan (CH3SH), dimetil
sulfida (CH3SCH3) dan dimetil disulfida (CH3SSCH3) mendominasi bagian dari
limbah yang dihasilkan dan dapat menyebabkan terjadinya hujan asam (Casey 1980). Proses pulping konvensional baik dengan cara mekanik maupun cara kimia juga membutuhkan energi yang sangat tinggi (Higley dan Dashek 1998).
Industri kertas menghasilkan limbah cair utama berupa black liquor dan senyawa klorin sisa pemutihan yang mengandung senyawa yang bersifat toksik, mutagenik, persisten dan bioakumulasi, yang dapat mengganggu lingkungan khususnya lingkungan perairan, dan resiko terpaparnya masyarakat oleh buangan zat kimia berbahaya. Diantara zat kimia berbahaya tersebut adalah hasil reaksi klorin dengan senyawa organik dalam kayu yang disebut dioksin, furan,
1
polychlorinated biphenyls (PCBs) dan yang paling toksik
2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD). Organoklorin dapat menimbulkan berbagai gangguan kesehatan seperti kanker, cacat lahir, endometriosis, penurunan jumlah spermatozoa dan gangguan perkembangan janin, juga dapat menyebabkan kerusakan genetis dan penurunan daya tahan ikan (Muladi 2000; Rini 2002).
Kini sejumlah besar perusahaan di dunia sedang mengupayakan keuntungan melalui suatu pendekatan pencegahan lingkungan yang dikenal sebagai produksi bersih. Sebagai salah satu pendekatan dalam pengelolaan lingkungan, penerapan produksi bersih masih baru di Indonesia. Dari sisi proses produksi, ternyata pelaksanaan produksi bersih pada industri pulp dan kertas banyak terkait dengan upaya recovery bahan kimia pemasak serta penggunaan bahan pemutih yang aman bagi lingkungan, disamping upaya konservasi bahan baku, air, dan energi (Hasibuan 2007). Oleh karena itu berbagai upaya untuk mengurangi dampak negatif proses-proses tersebut perlu terus diusahakan. Penerapan bioteknologi yaitu dengan memanfaatkan proses biologi menggunakan jamur pendegradasi lignin dalam proses teknologi biobleaching dan biopulping, merupakan salah satu alternatif dan terobosan besar yang perlu dikaji (Oriaran et al. 1990; Higley dan Dashek 1998).
endapan meningkat, kekuatan regang, pecah dan lipat lembaran meningkat dan nilai sobek menurun dengan peningkatan lama inkubasi. Keadaan di atas, menurut Oriaran et al. (1990) disebabkan proses biodegradasi oleh jamur yang menghasilkan penurunan biomassa kayu dan struktur pori tambahan pada dinding sel yang mempertinggi penetrasi cairan pemasak dan meningkatkan nisbah holoselulosa terhadap lignin (H/L). Oriaran (1989 dalam Oriaran et al. 1990) mengemukakan bahwa produksi ligninase jamur dapat meningkatkan nisbah H/L pada kayu. Hal ini disebabkan penurunan kandungan lignin serpih kayu yang didegradasi jamur. Namun Oriaran et al. (1990) mengemukakan adanya penurunan derajat putih pulp dengan meningkatnya lama inkubasi jamur. Yuliansyah et al. (2007) juga mengemukakan hasil pemutihan dengan jamur yaitu bilangan kappa menurun namun rendemen dan kekuatan kertas juga menurun drastis. Ho et al. (1990) mengemukakan bahwa penambahan 0,25% pulp pada media mendorong kecepatan pertumbuhan Coriolus versicolor dengan memperpendek periode lag bleaching dari 2 hari menjadi 1 hari dan mempertinggi proses rangkaian pemutihan pulp.
Pleurotus ostreatus diduga berpotensi untuk industri pulp karena diketahui mendegradasi lignin lebih efisien dibanding P. chrysosporium (Kerem et al. 1992; Hadar et al. 1993). Beberapa aktivitas enzim ekstraseluler Pleurotus spp. telah dipelajari, diantaranya MnP (Kerem et al. 1992). Mekanisme degradasi lignin oleh
Pleurotus belum banyak dipelajari seperti pada P. chrysosporium. Namun dari beberapa studi yang dilakukan, terlihat enzim yang bertanggungjawab untuk degradasi lignin dalam Pleurotus cukup bervariasi.
Perumusan Masalah
Sampai saat ini penelitian mengenai jamur pelapuk kayu kelompok
Sebagai langkah awal diperlukan suatu penelitian untuk mencari isolat jamur dari kelompok Pleurotus lokal yang unggul sebagai pendegradasi lignin. Isolat lokal merupakan plasma nutfah kekayaan alam Indonesia yang harus digali potensi ligninolitiknya. Preservasi plasma nutfah dan pendekatan genetik serta pemanfaatan isolat-isolat lokal ini adalah dasar untuk pengembangan industri jamur di sejumlah negara dan juga di Indonesia sehingga layak menjadi prioritas dalam menghadapi persaingan bebas dalam era globalisasi saat ini
Dalam penelitian ini dilakukan eksplorasi dan isolasi jamur kelompok
Pleurotus di lapangan di sekitar Bogor. Isolasi dilakukan dengan teknik kultur jaringan dari daging tubuh buah jamur dengan media MEA. Kultivasi dengan media serbuk gergajian kayu sengon dalam kantong dilakukan untuk mengetahui pembentukkan tubuh buah. Isolat-isolat kelompok Pleurotus yang dapat membentuk tubuh buah pada media serbuk gergajian kayu sengon, kemudian dipelajari karakter fisiologisnya, karakter ligninolitiknya pada jenis kayu yang berbeda, aktivitas ligninolitiknya pada media serbuk gergajian kayu sengon, aktivitas enzim ligninasenya yaitu manganese peroksidase (MnP), lignin peroksidase (LiP) dan lakase. Selanjutnya enzim MnP dari dua isolat yang mewakili asal Bogor dipurifikasi. Terakhir, jamur pelapuk putih kelompok
Pleurotus tersebut diidentifikasi. Sebagai pembanding standar digunakan
Tujuan Penelitian
Tujuan dari penelitian ini adalah untuk mengetahui keberadaan jamur kelompok Pleurotus di lapangan di sekitar Bogor dan mempelajari potensi ligninolitiknya dengan melakukan eksplorasi dan isolasi, karakterisasi fisiologis, ligninolitik dan morfologis serta identifikasi. Aktivitas ligninolitik jamur diukur berdasarkan pada daya degradasinya pada beberapa jenis kayu bahan baku pulp yaitu kayu akasia, pinus dan sengon dan biodegradasi media serbuk gergajian kayu sengon. Potensi ligninolitik juga diamati dengan melihat aktivitas MnP, LiP dan lakase kemudian memurnikan enzim MnP dari isolat yang mewakili asal Bogor.
Manfaat Penelitian
Manfaat penelitian ini adalah untuk memperoleh informasi ilmiah yang menyeluruh mengenai karakter fisiologi dan morfologi jamur kelompok Pleurotus
asal Bogor dan potensinya dalam mendegradasi lignin, yang diharapkan dapat menjadi landasan pengetahuan untuk memanfaatkan potensi jamur tersebut sebagai agen biobleaching dan biopulping, khususnya potensinya sebagai penghasil enzim ligninase seperti MnP, LiP dan lakase.
Hipotesis Penelitian
1. Isolat spesies jamur yang berbeda memiliki karakter fisiologi yang berbeda 2. Isolat spesies jamur yang berbeda memiliki kemampuan yang berbeda dalam
mendegradasi satu jenis media
3. Isolat spesies jamur yang sama memiliki kemampuan yang berbeda dalam mendegradasi jenis kayu yang berbeda
4. Isolat spesies jamur yang berbeda memiliki aktivitas enzim ligninase yang berbeda
Kebaruan Penelitian
Penelitian yang dilakukan merupakan penelitian dasar yang menyeluruh tentang jamur pelapuk putih kelompok Pleurotus asal Bogor dari mulai eksplorasi, isolasi, karakterisasi fisiologi, karakterisasi potensi ligninolitiknya pada chip kayu bahan baku pulp dan media serbuk gergajian kayu sengon, sampai pengukuran aktivitas enzim ligninase (MnP, LiP dan lakase) serta purifikasi MnP dari isolat yang mewakili asal Bogor. Terakhir adalah identifikasi secara morfologi baik makroskopik maupun mikroskopik yang ditunjang dengan karakter kultur dan fisiologi. Kajian mendalam ini belum pernah dilakukan terhadap jamur kelompok
Bahan Lignoselulolitik dan Gambaran Umum Tiga Jenis Kayu
Bahan lignoselulolitik merupakan selulosa yang berasosiasi secara erat
dengan lignin dan hemiselulosa (Enari 1983). Perbandingan komponen selulosa,
hemiselulosa dan lignin pada kebanyakan padatan selulosa secara kasar adalah
4:3:3 (Kollmann dan Cote 1968 dalam Haygreen dan Bowyer 1993) (Tabel 1).
Tabel 1 Persentase kadar selulosa, hemiselulosa, lignin dan pektin serta pati kayu berdasarkan bobot kering
Tipe kayu Selulosa Hemiselulosa Lignin Pektin dan pati Kayu daun jarum
Kayu daun lebar
40-44 40-44
20-32 15-35
25-35 18-25
6 6 Sumber: Kollman dan Cote (1968) dalam Haygreen dan Bowyer (1993)
Wenzl (1970) menyatakan kandungan zat ekstraktif kayu daun lebar
(hardwood) lebih tinggi dibanding kayu daun jarum (softwood). Komponen zat ekstraktif terdiri atas resin, terpen, asam lemak, fenol dan tannin, yang tersusun
dari senyawa asam-asam resin, asam-asam lemak, ester-ester lemak dan bahan
yang tidak tersabunkan. Scheffer (1973 dalam Tambunan dan Nandika 1989)
menyatakan zat ekstraktif berfungsi sebagai bahan pengawet alami kayu dan
keberadaannya mempengaruhi ketahanan kayu terhadap deteriorasi oleh jamur.
Kayu Pinus. Pinus merkusii Jungh. et de Vriese (pinus/damar/tusam) termasuk jenis kayu daun jarum. Kayu pinus ini mempunyai kayu teras berwarna
coklat kuning muda dengan pita dan gambar yang berwarna
putih/kekuning-kuningan dan tekstur kayu yang halus, permukaan kayunya licin serta
mengandung damar dan berbau terpentin. Rata-rata bobot jenisnya adalah 0,55
(0,40-0,75) dan termasuk kelas awet IV dan kelas kuat III (Pandit dan Ramdan
2002), bobot jenisnya adalah 0,44-0,59 dan termasuk kelas awet IV dan kelas kuat
mengandung selulosa 54,9%, lignin 24,3%, pentosa 14,0%, abu 1,1% dan silika
0,2% (Abdurahim et al. 1989).
Kayu Akasia. Acacia mangium Willd (mangium/akasia) termasuk jenis kayu daun lebar yang cepat tumbuh (fast growing species). Kayu dengan pori tata lingkar ini mempunyai warna kayu teras dan gubal yang dapat dilihat jelas; bagian
teras berwarna lebih gelap, sedangkan gubalnya berwarna putih dan lebih tipis.
Kayu teras berwarna agak kecoklatan, hampir mendekati kayu jati, kadang-kadang
mendekati warna jati gembol. Arah seratnya terlihat lurus sampai berpadu
(Ginoga et al. 1999 dan Ginoga 1997 dalam Malik et al. 2007).
Kayu ini mempunyai bobot jenis antara 0,46 sampai 0,54 (Ginoga 1997
dalam Malik et al. 2007). Kayu akasia dari hutan tanaman (asal Jawa Barat) relatif memiliki sifat keawetan lebih buruk (kelas awet II – III) dibanding kayu akasia
dari hutan alam (asal Maluku). Berdasarkan bobot jenis, keteguhan lentur statis
dan tekan sejajar arah serat, maka kayu akasia ini termasuk kelas kuat II – III.
Menurut klasifikasi komponen kimia kayu Indonesia (Deptan 1976 dalam
Malik et al. 2007), kayu akasia ini termasuk kelompok sedang (40–44 %) dalam hal kandungan selolusa, kadar lignin sedang (18–32 %), kadar pentosan, silika dan
abu termasuk rendah dan zat ekstraktif termasuk tinggi. Menurut Siagian et al.
(1999 dalam Malik et al. 2007) menyatakan bahwa kayu akasia ini baik dijadikan sebagai bahan baku pulp karena memiliki kadar selulosa tinggi, lignin sedang,
pentosan rendah, ekstraktif tinggi dan abu sedang. Akan tetapi pembuatan pulp
dari kayu akasia ini perlu diperhatikan karena kadar ekstraktifnya tinggi.
Perbedaan umur pohon memberikan pengaruh yang berbeda terhadap
komposisi kimia kayu. Kadar selulosa, lignin, kelarutan dalam alkohol-benzena
dan air dingin, secara umum menunjukkan kecenderungan menurun dengan
bertambahnya umur pohon sedangkan kadar pentosan cenderung meningkat.
Untuk kadar abu, silika, kelarutan dalam NaOH 1% dan air panas, memberikan
respon yang berfluktuatif dengan bertambahnya umur tanaman (Malik et al. 2007).
Kayu Sengon. Kayu sengon (Paraserianthes falcataria L. Nielsen) merupakan kayu daun lebar tropis yang mempunyai warna kayu teras hampir
kayu terasnya, mempunyai tekstur kayu yang agak kasar dan merata, arah serat
yang lurus, bergelombang lebar atau terpadu. Kayu sengon memiliki bobot jenis
yang rendah yaitu berkisar antara 0,24-0,49 dengan rata-rata 0,33. Kekuatannya
digolongkan sebagai kayu kelas kuat IV-V dan keawetannya digolongkan sebagai
kayu kelas IV-V (Widarmana 1984). Kayu sengon mempunyai kadar selulosa
48,33%, lignin 27,28% dan pentosan 16,34%, dengan nisbah C/N 53,17/0,25
(Nurhayati 1988).
Penguraian Komponen Kayu Secara Enzimatik oleh Jamur Pelapuk Putih
Jamur pelapuk putih (white-rot fungi) merombak lignin dan polisakarida. Kayu yang terdegradasi menjadi putih, kuning atau coklat terang dan lunak.
Umum menyerang kayu daun lebar. Kerugian lain adalah keuletan kayu dan
derajat polimerasi menurun, namun pengerutan, pulp yang dihasilkan, kualitas
serat serta kelarutan dalam alkali hampir sama dengan kayu yang normal (Fengel
dan Wegener 1989; Tambunan dan Nandika 1989). Contoh penampakan sel kayu
yang lapuk oleh jamur pelapuk putih ditunjukkan pada Gambar 1A. Miselium
tumbuh dan menyebar di dalam kayu dengan bantuan enzim-enzimnya
kemungkinan melalui beberapa jalur seperti lumina, noktah atau langsung
menembus dinding sel (Gambar 1B) (Fengel dan Wegener 1989).
Sumber: Foto 1A (von Aufsess dalam Fengel dan Wegener 1989) dan Foto 1B (Fengel dan Wegener 1989)
Gambar 1 A. Pelapukan kayu spruce (Picea abies) oleh jamur pelapuk putih. a,b. Degradasi oleh Heterobasidium annosum, mikrograf cahaya. c. Degradasi oleh Peniophora gigantea, mikrograf SEM. B. Bagan kemungkinan jalur pertumbuhan hifa. 1. lumina sel parenkim. 2. lumina sel pembuluh. 3. Jalur sederhana, noktah setengah berbatas dan noktah berbatas. 4. Melalui dinding-dinding sel. 5. lamela tengah majemuk dan dinding-dinding sel.
Enzim-enzim perombak selulosa dan lignin pada umumnya merupakan
enzim adaptif yaitu enzim-enzim yang dihasilkan hanya pada suatu substrat
tempat dia aktif.
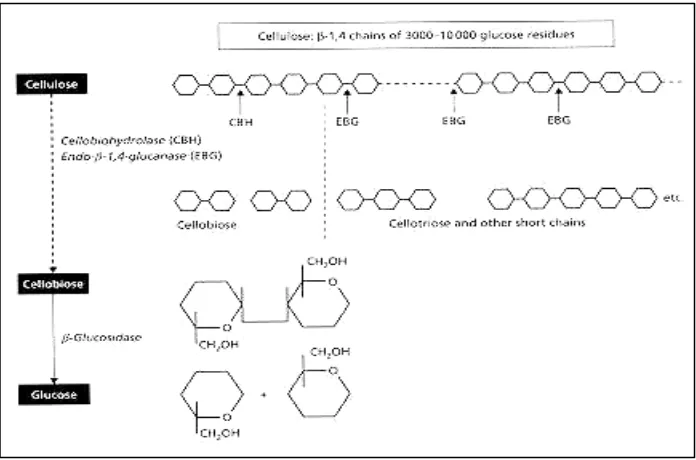
Mekanisme Degradasi Selulosa. Degradasi pada selulosa kristal oleh jamur pelapuk putih P. chrysosporium, mirip pada selulase-selulase jamur lainnya, yaitu dilakukan oleh sebuah kompleks enzim multikomponen dimana tiap-tiap
komponen berinteraksi secara sinergi untuk mendegradasi selulosa menjadi
glukosa. Endoglukanase (EGs) bekerja secara acak pada permukaan luar
mikrofibril selulosa, yaitu dengan membuka ujung non-reduksi yang kemudian
oleh cellobiohidrolase (CBHs) dihidrolisis dan menghasilkan selobiose. Selobiose dipotong oleh β-glukosidase, menghasilkan glukosa. Selobiose, suatu produk dari
kerja selulase, dapat menginduksi dan juga menginhibisi selulase pada P. chrysosporium (Eriksson dan Hamp 1978 dalam Highley dan Dashek 1998) (Gambar 2).
Sumber: Eriksson dan Hamp 1978 dalam Highley dan Dashek 1998
Gambar 2 Mekanisme degradasi selulosa.
Hemiselulosa adalah sebuah grup homopolimer dan heteropolimer yang
mengandung sebagian besar ikatan-ikatan utama anhidro–β-(1→ 4)D-xylopyranosa, mamnopiranosa, glukopiranosa dan galaktopiranosa. Enzim-enzim yang mendegradasi hemiselulosa juga kompleks yang umum disebut hemiselulase.
Hemiselulase menunjukkan sebagai agen bleaching yang menjanjikan dalam produksi pulp dan kertas. Degradasi hemiselulosa oleh jamur pelapuk putih
kemudian dianalogikan secara kasar dengan selulosa, tetapi mekanisme
serangannya telah dipelajari secara lebih detail oleh Kirk dan Cowling (1984).
Ikatan hemiselulosa diserang pertamakali oleh endoenzim-endoenzim (mannanase
dan xilanase) yang menghasilkan secara intensif ikatan-ikatan pendek yang
dihidrolisis menjadi gula sederhana oleh glukosidase (mannosidase, xilosidase dan
glukosidase). Sejauh ini belum diketahui jenis eksoenzim yaitu enzim-enzim yang
dapat mengendalikan sisi-ikatan substitusi (arabinosa, asam uronik dan asetil)
yang terlibat (Kirk dan Cowling 1984). Seperti dengan selulase, gula-gula
sederhana membatasi produksi sebagian besar enzim-enzim pendegradasi
hemiselulosa oleh jamur pelapuk putih. Selulosa diduga menjadi sumber karbon
penting untuk mendorong terbentuknya enzim-enzim pendegradasi hemiselulosa
oleh jamur.
Degradasi Lignin. Tidak seperti selulosa dan hemiselulosa, lignin prinsipnya tidak berikatan linear tetapi merupakan senyawa kompleks. Polimer
heterogen, dengan senyawa aromatik non-stereoregular yang disusun oleh unit
fenilpropanoid. Jamur pelapuk putih adalah satu-satunya organisme yang dikenal
mampu mendegradasi lignin secara sempurna menjadi karbondioksida dan air.
Perkembangan terbaru saat ini telah menunjukkan kaitannya dengan biokimia dan
genetik molekuler biodegradasi lignin yang umumnya menggunakan penelitian
jamur pelapuk putih P. chrysosporium (Buswell dan Odier 1987, Alic dan Gold 1991, Kuan et al. 1991 dalam Highley dan Dashek 1998; Cullen dan Kersten 1992 dan 1996).
Ligninolitik berhubungan dengan produksi enzim ekstraseluler
pendegradasi lignin yang dihasilkan oleh jamur pelapuk putih. Dua enzim yang
(lignin peroksidase (LiP) dan manganese peroksidase (MnP)) (Howard et al. 2003; Kirk et al. 1980).
Lakase merupakan enzim multi-copper yang dapat mengkatalis reaksi oksidasi beberapa substrat seperti polifenol, substituen penol, diamin dan
beberapa senyawa anorganik (Thurston 1994). Enzim ini pertama kali ditemukan
dalam getah pohon pernis Jepang Rhus vernicifera pada tahun 1988 dan pada umumnya banyak ditemukan pada jamur dan tanaman tingkat tinggi (Anonim
2006; Thurston 1994; Palonen 2004).
Lakase (E.C.1.10.3.2; benzendiol: oksigen oksidoreduktase) sebagian
besar merupakan glikoprotein ekstraseluler yang mengandung atom tembaga
dengan bobot molekul antara 60-80 kDa dan juga merupakan salah satu grup
terkecil enzim yang dinamakan oksidase tembaga biru (Thurston 1994). Lakase
mengandung empat atom protein pada tiap molekulnya (Palonen 2004).
Mekanisme reaksi enzimatik yang terjadi oleh lakase adalah reaksi
oksidasi satu elektron. Dibutuhkan peranan molekul oksigen sebagai penerima
elektron dan kemudian membentuk molekul air. Ketika reaksi oksidasi
berlangsung, substrat kehilangan satu elektronnya dan biasanya terbentuk radikal
fenoksi bebas (Thurston 1994) yang berperan sebagai intermediet. Radikal bebas
yang tidak stabil tersebut dapat melangsungkan reaksi oksidatif enzimatik
selanjutnya atau reaksi non-enzimatik seperti hidrasi, disproporsionasi dan polimerisasi (Thurston 1994).
Lakase telah banyak menjadi subyek penelitian untuk dimanfaatkan secara
luas oleh karena lakase sifat spesifiknya yang rendah terhadap
substrat-substratnya (Cavallazzi et al. 2004; Thurston 1994). Hidrokuinin, katekol, guaiakol, 2,6-dimetoksifenol, p-fenildiamin dan siringaldazin merupakan
substrat-substrat yang cukup bagus bagi lakase. Substrat tiruan lakase seperti ABTS
(2,2-azinobis-3-etilbenzthiazolin-6-sulfonat) dapat berperan sebagai mediator yang
memungkinkan oksidasi komponen non fenolik pada lignin yang tidak dapat
dioksidasi oleh lakase sendiri (Bourbonnais dan Paice 1990).
Pada jamur, lakase berperan dalam proses degradasi lignin (Hatakka 1994)
dan beberapa fungsi lain diantaranya pigmentasi, pembentukan badan buah,
berbeda memiliki perbedaan yang sangat nyata pada substrat spesifik, bobot
molekul, pH optimumnya seperti yang dihasilkan oleh beberapa jamur yakni
Panaeolus sphinctrinus, Panaeolus papilionaceus, Coprinus friesii (Heinzkill et al. 1998), Lepista sordida (Cavallazzi et al. 2004), Trametes versicolor (Han et al.
2004), Pycroporus cinnabarinus (Eggert et al. 1996), Pleurotus eringii (Muñoz et al. 1997) dan Coriolopsis gallica (Pickard et al. 1999).
Pemanfaatan lakase sangat luas diterapkan dalam berbagai bidang antara
lain dalam proses bioremediasi dan biodegradasi polutan organik pada tanah
seperti klorofenol (Ahn et al. 2002), dan polisiklik aromatik hidrokarbon (Han et al. 2004), pada proses dekolorisasi dan detoksifikasi pada pewarna tekstil (Abadulla et al. 2000) serta digunakan sebagai bleaching pada proses biodelignifikasi pada pulp industri kertas (Bourbonnais dan Paice 1990).
Lignin peroksidase (EC.1.11.1.14; diarilpropan: oksigen, hidrogen
peroksida oksidoreduktase; bobot molekul antara 38 dan 43 kDa) dan MnP
(EC.1.11.1.13; Mn(II): H2O2 oksidoreduktase; bobot molekul antara 43 dan 49
kDa) merupakan glikoprotein yang memiliki sebuah protoporfirin IX sebagai
gugus prostetik dan membutuhkan hidrogen peroksida sebagai oksidan (Hatakka
1994; Tien dan Kirk 1984; Gold dan Alic 1993).
Enzim ekstraseluler LiP dan MnP memiliki peranan yang sangat penting
dalam proses biodelignifikasi. LiP memiliki kemampuan mengkatalis beberapa
reaksi oksidasi antara lain pemecahan ikatan Cα-Cβ rantai samping propil non
fenolik komponen aromatik lignin, oksidasi benzil alkohol, oksidasi fenol,
hidroksilasi benzylic methylenegroups dan pemecahan cincin aromatik komponen non phenolik senyawa lignin (Tien dan Kirk 1984). Sedangkan MnP diketahui
memiliki kemampuan mengoksidasi baik komponen fenolik maupun non fenolik
senyawa lignin.
Seperti enzim peroksidase lainnya, LiP memiliki siklus katalitik yang
dinamakan mekanisme ping-pong. Reaksi yang terjadi yakni H2O2 mengoksidasi enzim pada keadaan awal (resting enzyme) dengan dua elektron membentuk senyawa intermediet I, senyawa tersebut kemudian mengoksidasi substrat
aromatik dengan menggunakan satu elektron membentuk senyawa intermediet II
mengoksidasi substrat lainnya sehingga terbentuk enzim awal dan produk radikal
bebas (Cullen dan Kersten 1992). Terbentuknya radikal bebas secara spontan atau
bertahap inilah yang mengakibatkan lepasnya ikatan antar molekul dan beberapa
inti pada cincin aromatik.
Prinsip fungsi MnP adalah bahwa enzim tersebut mengoksidasi Mn2+
membentuk Mn3+ dengan adanya H2O2 sebagai oksidan. Aktivitasnya dirangsang
oleh adanya asam organik yang berfungsi sebagai pengkelat atau penstabilkan
Mn3+. Mekanisme reaksi yakni MnP pada keadaan awal dioksidasi oleh H2O2
membentuk MnP-senyawa I yang dapat direduksi oleh Mn2+ dan senyawa fenol
membentuk MnP-senyawa II. Senyawa tersebut kemudian direduksi kembali oleh
Mn2+ tetapi tidak oleh fenol membentuk enzim keadaan awal dan produk
(Wariishi et al. 1989). Adanya Mn2+ bebas sangat penting untuk menghasilkan siklus katalitik yang sempurna.
MnP + H2O2 MnP-senyawa I + H2O
MnP-senyawa I + Mn2+ MnP-senyawa II + Mn3+
MnP-senyawa I + AH MnP-senyawa II + A- + H+
MnP-senyawa II + Mn2+ MnP + Mn3+
Manganese peroksidase dihasilkan oleh P. ostreatus (Sarkar et al. 1997) dan juga oleh P. crysosporium (Brown et al. 1990) dan oleh Phlebia radiata
(Vares et al. 1995). Masing-masing jamur menghasilkan kombinasi enzim yang berbeda-beda, misalnya ada yang hanya menghasilkan LiP dan MnP, MnP dan
lakase, atau jamur yang menghasilkan LiP dan lakase (Kerem dan Hadar 1998).
Aktivitas ligninolitik jamur pelapuk putih seperti pada T. versicolor dan P. chrysosporium meningkat pada media tumbuh yang kandungan karbon sederhana dan nitrogennya rendah (Eaton dan Hale 1993; Katagiri et al. 1995). Menurut Leatham et al. (1983) dan Griffin (1994), penambahan asam glutamat, glutamin, histidin dan sikloheksimida akan menekan aktivitas ligninolitiknya. Menurut
Heinzkill dan Messner (1997), jamur pelapuk putih yang mendegradasi lignin
tidak dapat menggunakan lignin sebagai satu-satunya sumber karbon dan energi.
Namun berdasarkan hasil penelitian Sugipriatini (1998) diketahui bahwa
penambahan glukosa dapat menekan pertumbuhan maupun aktivitas ligninolitik
memiliki kemampuan mendegradasi lignin yang tinggi dan dalam
pertumbuhannya menggunakan lignin sebagai sumber karbon (Artiningsih et al.
2000).
Sampai saat ini isolat jamur pelapuk putih P. chrysosporium banyak menjadi obyek para peneliti dikarenakan menghasilkan aktivitas LiP dan MnP
yang tinggi. Seperti halnya pada lakase selain potensi dalam proses
biodelignifikasi, lignin peroksidase dan mangan peroksidase berpotensi dalam
proses biobleaching dan biopulping pulp serta proses degradasi senyawa-senyawa berbahaya.
Aktivitas suatu enzim dipengaruhi oleh beberapa faktor diantaranya
konsentrasi substrat, konsentrasi enzim, pH, suhu dan beberapa faktor lain.
Karakteristik dari reaksi yang dikatalisis enzim yakni pada konsentrasi enzim
yang digunakan, laju awal reaksi akan meningkat sebanding dengan peningkatan
konsentrasi substrat sampai mencapai harga tertentu. Mula-mula laju naik dengan
pesat, namun pada konsentrasi substrat yang tinggi laju reaksi menjadi tidak
berpengaruh dan relatif konstan. Hal ini disebabkan adanya interaksi sistem yang
dideteksi oleh substrat yakni pada konsentrasi substrat yang besar, substrat dapat
mengihibisi reaksinya sendiri.
Enzim-enzim dengan derajat kemurnian yang tinggi dalam batas-batas
tertentu, memiliki suatu hubungan linier antara konsentrasi enzim dan taraf
aktivitasnya. Hubungan tersebut menjelaskan bahwa aktivitas enzim merupakan
ukuran hilangnya reaktan atau terbentuknya produk dari reaksi yang dikatalisis.
Aktivitas enzim bertambah dengan naiknya suhu sampai aktivitas optimumnya
tercapai. Kenaikan suhu lebih lanjut berakibat dengan berkurangnya aktivitas dan
pada akhirnya terjadi kerusakan enzim (terdenaturasi). Demikian juga halnya
untuk pH, ada pH tertentu yang memberikan aktivitas optimum, di atas atau di
bawah pH tersebut aktivitas enzim akan menjadi berkurang.
Aktivitas enzim yang dihasilkan P. chrysosporium pada kultur awal lambat (Kirk 1988 dalam Trotter 1990), tetapi setelah diperoleh strain yang cukup
baik dan optimalisasi parameter kultur, aktivitas enzim telah diperbaiki sampai
Jamur Pelapuk Putih Kelompok Pleurotus
Beberapa jamur pleurotoid adalah jamur pelapuk kayu yang diketahui
mempunyai kemampuan sebagai pendegradasi lignin. Jamur pleurotoid
merupakan salah satu dari spesies yang dibedakan berdasarkan stature types yang memudahkan dalam identifikasi jamur di lapangan, yang ciri-ciri pentingnya
adalah: a) keberadaan cincin, b) keberadaan volva, c) konsistensi tangkai,
d) attachment tangkai, e) attachment lamela, f) bentuk (shape) dari pileus
(tudung) dan g) tipe pinggiran pileus. Spesies berdasarkan tipe stature lainnya diantaranya adalah Amanitoid, Vaginatoid, Lepiotoid, Pluteotoid, Tricholomatoid,
Naucoroid, Clitocyboid, Almillarioid, Mycenoid dan Collibioid (Largent 1973).
Jamur pleurotoid umumnya mempunyai ciri-ciri di lapangan: bentuk
tudung agak membulat, lonjong dan melengkung seperti cangkang tiram, jamur
berlamella melanjut (deccurent), berukuran kecil sampai besar, berwarna bervariasi (putih, krem, abu-abu, violet, sampai hitam), lunak, licin, daging
basidiokarp tebal, berbau sedap; tangkai pendek atau absen (pinggir, kurang dari
2 cm), eksentrik atau lateral; spora bulat-elips, mempunyai dinding tipis dan halus, spora non-amiloid, jejak spora umumnya putih; kadang-kadang jamur ini dapat tumbuh tunggal, biasanya ditemukan banyak tubuh buah pada satu kali
pengamatan, berkelompok, berkerumun, bersusun seperti rak; habitatnya
umumnya pada kayu konifer dan kayu daun lebar; dan diketahui beberapa
spesiesnya bersifat edible, diantaranya yang terkenal adalah kelompok Pleurotus
(Brown 1981; Largent 1973). Hohenbuehelia termasuk kelompok Pleurotus, karena ada yang mempunyai nama sinonim Pleurotus, juga belum banyak dilaporkan dan diduga mempunyai potensi mendegradasi lignin karena di alam
banyak tumbuh pada substrat kayu. Terdapat lebih kurang 67 spesies kelompok
Pleurotus yang berhasil dikumpulkan dan dilaporkan oleh para peneliti seluruh dunia, sejak Roussel (1805) pertama kali memberi nama Pleurotus (LR 2004).
Jamur Pleurotus diklasifikasikan oleh beberapa peneliti dalam Alexopoulos et al. (1996) dan Chang dan Miles (1989) adalah sebagai berikut: Super Kingdom: Eukariota; Kingdom: Mycetes (Fungi); Divisi: Mycota;
(Basidiomycetes); Sub Klas: Holobasidiomycetidae; Ordo: Agaricales; Famili:
Tricholomataceae; Genus : Pleurotus; Species: Pleurotus spp..
Menurut Segedin et al. (1995), Pleurotus diklasifikasikan ke dalam famili tersendiri yaitu Pleurotaceae. Menurut Moncalvo et al. (2002), Hohenbuehelia
juga mempunyai famili yang sama dengan Pleurotus yaitu Pleurotaceae. Dalam famili ini hanya ada dua genus yaitu Pleurotus dan Hohenbuehelia, hal ini berdasarkan penelitian terbaru dengan menggunakan uji genetik molekuler RNA.
Di lapangan, keduanya sulit dibedakan dan dapat dikelompokkan menjadi
(Exploration of White-rot Fungi Pleurotus Group from Bogor)
Abstrak
Eksplorasi jamur pelapuk putih lokal kelompok Pleurotus telah dilakukan pada bulan Mei 2004 dari berbagai wilayah di Kabupaten dan Kota Bogor. Kondisi iklim saat ekplorasi adalah akhir musim hujan dengan suhu rata-rata minimum dan maksimum berturut-turut 18,3 dan 26,3oC, kelembaban nisbi rata-rata 85%, hari hujan 17 hari dan curah hujan 245 mm/bulan. Isolasi jamur dilakukan dengan teknik kultur jaringan dari daging tubuh buah jamur dengan media MEA. Kultivasi dilakukan pada media serbuk gergajian kayu sengon (Paraserianthes falcataria) dalam kantong untuk mengetahui pembentukkan tubuh buah.
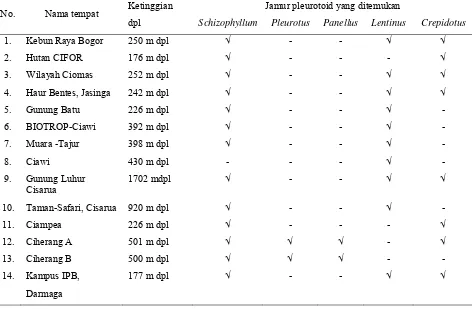
Jenis jamur pleurotoid yang umum ditemukan di lapangan yaitu
Schizophillum spp., Lentinus spp. dan Crepidotus spp., sedangkan kelompok
Pleurotus dan Panellus spp. hanya ditemukan dari satu daerah yaitu di areal kebun dan tempat penggergajian kayu di kecamatan Ciherang (+ 501 m dpl). Sebanyak 24 isolat jamur kelompok Pleurotus telah diisolasi dan 17 isolat diantaranya dikultivasi pada media serbuk gergajian kayu sengon. Enam isolat diantaranya dapat membentuk tubuh buah.
Kata-kata Kunci : Jamur kelompok Pleurotus, Bogor
Abstract
Exploration of white-rot fungi of wild Pleurotus group was conducted in May 2004 from 13 places in District and City of Bogor. The climatic condition at that time of exploration was the end of rainy season with minimum and maximum temperatures of 18.3 and of 26.3oC respectively, relative humidity of 85%, 17 rainy days and rainfall of 245 mm/month. Isolation was carried out using tissue culture method of fruiting bodies of the fungi on MEA medium. Cultivation treatments using sengon (P. falcataria) sawdust media was conducted to know formation of fruiting bodies .
Results showed pleurotoid fungi that commonly found were Schizophillum
spp., Lentinus spp. and Crepidotus spp., whereas Pleurotus group and Panellus
spp. were found only in garden and sawmill of sub-district Ciherang (+ 501 m asl). Twenty four isolates of Pleurotus group were isolated, and 17 of those isolates were cultivated on sengon wood sawdust media. Among them, six isolates are able to form fruit bodies.
Pendahuluan
Penerapan bioteknologi melalui pemanfaatan jamur pendegradasi lignin
dalam proses biobleaching dan biopulping, merupakan salah satu alternatif dan terobosan besar yang perlu dikaji (Higley dan Dashek 1998). Penelitian dimulai
dengan proses eksplorasi pada organisme yang potensial sehingga diperoleh agen
yang maksimal. Beberapa jamur pleurotoid adalah jamur pelapuk kayu yang
diketahui mempunyai kemampuan sebagai pendegradasi lignin. Jamur yang
termasuk pleurotoid diantaranya adalah Schizophyllum, Pleurotus, Panus, Lentinus, Plicatura, Hohenbuehelia, Claudotus, Paxillus dan Crepidotus (Largent 1973). Lebih kurang 67 spesies Pleurotus yang berhasil dikumpulkan dan dilaporkan oleh para peneliti seluruh dunia, sejak Roussel (1805) pertama kali
memberi nama Pleurotus (LR 2004). Penelitian ini lebih difokuskan pada kelompok Pleurotus dan Hohenbuehelia yang sebelumnya mempunyai nama sinonim Pleurotus yang diduga mempunyai potensi mendegradasi lignin.
Sampai saat ini penelitian mengenai jamur pleurotoid kelompok Pleurotus
asal Indonesia masih sangat terbatas meskipun jamur ini sudah dikenal oleh
masyarakat untuk berbagai keperluan. Oleh karena itu penelitian mengenai
keberadaan jamur pleurotoid kelompok Pleurotus merupakan data dasar yang sangat berharga untuk potensi dan pemamfaatannya. Tujuan dari penelitian ini
adalah untuk mengetahui keberadaan jamur pleurotoid kelompok Pleurotus dari lapangan di sekitar Bogor.
Bahan dan Metode
Eksplorasi jamur pleurotoid kelompok Pleurotus dimulai dengan melakukan survei di beberapa wilayah di Kabupaten dan Kota Bogor yang diduga
terdapat populasi jamur. Wilayah yang dikunjungi meliputi daerah pegunungan,
hutan, perkebunan dan bekas tegakan kayu dan sampai ke daerah sentra
penggergajian kayu dan perkampungan. Kegiatan survei dan eksplorasi jamur
dilaksanakan sejak bulan Maret hingga Juni 2004. Jamur kelompok Pleurotus
gambarnya. Deskripsi dan kondisi iklim pada saat jamur ditemukan juga dicatat.
Selanjutnya jamur dikoleksi dan diambil dengan menggunakan plastik untuk
dibawa ke Laboratorium untuk keperluan isolasi. Selanjutnya jamur diidentifikasi
mengacu pada Brown (1981).
Isolasi Basidiokarp Jamur Dari Lapang
Basidiokarp atau tubuh buah jamur yang diperoleh diambil gambarnya dan
dibersihkan dari kotoran. Jamur selanjutnya ditelungkupkan di atas kertas
berwarna gelap dan kertas berwarna putih dan dimasukkan ke dalam wadah
plastik berbentuk nampan tertutup untuk mendapatkan jejak sporanya. Tubuh
buah dalam wadah tersebut kemudian dibawa ke Laboratorium dengan hati-hati
untuk keperluan isolasi.
Tubuh buah jamur diisolasi dengan metode kultur jaringan dengan
menanam jaringan daging (contex) basidiokarp pada medium agar. Permukaan basidiokarp yang baru diambil dari lapangan disemprot dengan alkohol 70%
kemudian dipotong melintang menjadi dua bagian. Selanjutnya bagian daging
basidiokarp tanpa lamela dan jaringan epidermis luar tudung tersebut diambil
secara aseptik dengan bantuan pinset. Potongan tersebut kemudian diinokulasikan
pada medium MEA (Malt Extract Agar) yang telah diberi antibiotik kloramfenicol/ kemisitin 250 mg/l dalam cawan Petri berdiameter 9 cm dan agar
miring pada botol vilial 20 ml dan diinkubasikan pada 29(±1)oC selama 10 hari.
Miselium yang telah tumbuh dimurnikan dan diberi kode isolatnya sesuai dengan
kelompok basidiokarp yang diisolasi. Untuk melihat apakah isolat benar-benar
dari kelompok Basidiomiset maka diamati adanya sambungan apit pada miselium.
Kultivasi Pada Media Serbuk Gergajian Kayu Sengon
Untuk memastikan bahwa yang diisolasi tersebut adalah pleurotoid
kelompok Pleurotus, selanjutnya biakan jamur tersebut dibuat bibit dengan cara menumbuhkan miselium ukuran 100 mm2 pada media bibit dalam botol yang
berisi jewawut 23%, dedak 3%, kapur 0,4% dan serbuk gergajian 73,6% serta air.
Selanjutnya biakan tersebut diinkubasi di tempat gelap pada suhu kamar sampai
sebanyak satu sendok teh dari bibit (+ 10 gram) tersebut untuk diinokulasikan
pada media serbuk gergajian kayu sengon dalam kantong. Serbuk gergajian kayu
sengon diperoleh dari tempat penggergajian kayu di daerah Ciherang Bogor, tanpa
diketahui asal usul kayu, dan disimpan lebih kurang satu minggu sebelum dipakai
dalam penelitian.
Media serbuk gergajian kayu sengon dalam kantong terdiri atas 82,5%
serbuk gergajian yang ditambah 15% dedak, 1,5% gips, dan 1,0% kapur serta air
secukupnya. Tiap kantong plastik diisi sekitar 400 gram media dan disterilisasi
dalam drum kukus selama 7 jam. Media serbuk gergajian kayu sengon yang sudah
diinokulasi dengan bibit disimpan di ruang inkubasi. Kantong yang sudah
dipenuhi dengan miselium selanjutnya disimpan di ruang pemeliharaan/ruang
produksi sampai keluar tubuh buah. Percobaan ini dilakukan dengan 4-10
ulangan.
Hasil dan Pembahasan
Hasil Penelitian
Eksplorasi Jamur Pleurotoid Kelompok Pleurotus
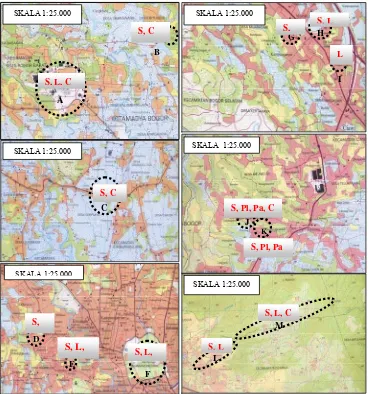
Hasil eksplorasi dan survei jamur selama bulan Mei 2004 di 14
wilayah/lokasi di Bogor disajikan dalam Tabel 1.1. Lokasi eksplorasi jamur dan
denahnya disajikan di Gambar 1.1. Hasil pengamatan di lapangan menunjukkan
jenis jamur pleurotoid yang umum ditemukan adalah genus Schizophillum,
Lentinus dan Crepidotus, kelompok Pleurotus dan genus Panellus hanya ditemukan di kecamatan Ciherang (+ 501 m dpl). Sebanyak 24 isolat jamur
kelompok Pleurotus telah diisolasi (Tabel 1.2).
Kondisi umum areal di kecamatan Ciherang, dimana jamur kelompok
Pleurotus ditemukan merupakan areal kebun bekas areal penggergajian kayu yang sudah ditinggalkan sekitar dua tahun sebelumnya. Tumpukan serbuk gergajian
sudah melapuk tersebut banyak ditanami dengan vegetasi pertanian, seperti pisang
dan ketela pohon (Ciherang A) dan areal penggergajian kayu campuran yang
Sumber:Bakosurtanal 2007
Keterangan: huruf merah: S=Schizophyllum; Pl=Pleurotus; Pa=Panellus; L=Lentinus; dan C=Crepidotus; huruf hitam: A. Kampus Darmaga IPB. B. Hutan CIFOR. C. lokasi penggergajian kayu di Ciampea. D. Areal perkampungan di Gunung Batu. E. Areal sekitar Pusat Penelitian dan Pengembangan Kehutanan. F. Kebun Raya Bogor. G. Areal sekitar Biotrop. H. Kampung Muara. I. Gang Gudang Garam, Ciawi. J. Areal pedesaan dan tempat penggergajian kayu di Ciherang A. K. Ciherang B. L. Batu Kasur Taman Safari. M. Areal hutan di Gunung Luhur Cisarua.
Gambar 1.1 Denah lokasi eksplorasi jamur pleurotoid (huruf hitam) dan jenis jamur yang ditemukan (huruf merah).