www.elsevier.comrlocateranireprosci
Levels of progesterone and changes in
prostaglandin F
2arelease during luteolysis and early
pregnancy in llamas and the effect of treatment with
flunixin meglumine
M.A. Aba
a,b,d,), H. Kindahl
c,d, M. Forsberg
a,d, M. Quiroga
b,
N. Auza
b aDepartment of Clinical Chemistry, Faculty of Veterinary Medicine, Swedish UniÕersity of Agricultural
Sciences, P.O. Box 7038, S-750 07Uppsala, Sweden
b
Department of Physiopathology Campus UniÕersitario, Faculty of Veterinary Sciences, UNCPBA, Paraje
Arroyo Seco srn, Tandil, 7000Buenos Aires, Argentina
c
Department of Obstetrics and Gynaecology, Faculty of Veterinary Medicine, Swedish UniÕersity of
Agricultural Sciences, P.O. Box 7039, S-750 07Uppsala, Sweden
d
Centre for ReproductiÕe Biology, SLU, Uppsala, Sweden
Received 19 March 1999; received in revised form 7 December 1999; accepted 7 December 1999
Abstract
The secretory patterns of progesterone in relation to concentrations of 15-ketodihydro-PGF2a
ŽPGFM during the period of luteolysis or of maternal recognition of pregnancy were determined.
in the blood of llamas mated either with an intact or a vasectomized male. The ability of flunixin
Ž .
meglumine FM to postpone luteolysis in non-pregnant llamas was investigated by injecting the drug intravenously every 6 h at a dose of 2.2 mgrkg from days 6 to 12 post-copulation into a group of non-pregnant llamas. A pulsatile pattern of prostaglandin release was recorded during luteolysis in non-pregnant llamas, giving further support to the hypothesis that PGF2a is the luteolytic agent in llamas. The mean number of peaks per animal rose from 0.3 on day 7 to 3.8 on day 10 and then declined to 1.1 on day 12 with corresponding mean peak amplitude changing from 465 to 1234 and 566 pmol ly1, respectively. In pregnant llamas, prostaglandin pulsatile
release also occurred. The mean number of peaks per animal rose from 0.4 on day 7 to 0.8 on day 10 and then declined to 0.2 on day 11 and 0.6 on day 12, with corresponding mean peak
)Corresponding author. UNCPBA Faculty of Veterinary Sci, Dept. of Physiopathology, Campus
Universi-tario, Paraje Arroyo Seco, 7000 Tandil Buenos Aires, Argentina, Tel.:q54-293-26667; fax:q54-293-28485.
Ž .
E-mail address: [email protected] M.A. Aba .
0378-4320r00r$ - see front matterq2000 Elsevier Science B.V. All rights reserved.
Ž .
amplitude changing from 494 to 676, 388 and 547 pmol ly1, respectively. The transient decrease and subsequent recovery in progesterone concentrations was observed to occur in connection with prostaglandin release during early pregnancy. Oestradiol-17bplasma peak concentrations attained
Ž
after luteolysis were significantly higher than those recorded in early pregnant animals around 30
y1 y1.
pmol l and ll pmol l . Concentrations of PGFM decreased rapidly after the first administra-tion of FM and remained low throughout the first 2 days of treatment. Thereafter, pulsatile release of prostaglandins started, and luteolysis proceeded; but a delay of 1–1.5 days in the progesterone decline was observed. Thus, it might be suggested that a higher dose andror a more intensive injection schedule is required in llamas than in other ruminants to prevent luteolysis. q2000 Elsevier Science B.V. All rights reserved.
Keywords: Llama; Progesterone; Prostaglandins; Flunixin meglumine; Corpus luteum; Pregnancy
1. Introduction
Ž .
In non-pregnant llamas Lama glama a dramatic fall in progesterone concentrations is observed between days 9 and 11 post-mating, in connection with repeated surge
Ž .
release of PGF2a from the uterus Sumar et al., 1988; Aba et al., 1995 . In pregnant animals, a transient decrease and subsequent recovery in progesterone concentrations has been reported during the period when maternal recognition of pregnancy is expected to
Ž .
occur Adams et al., 1991; Aba et al., 1995 . In these studies, no pulsatile release of PGF2a has been reported in association with the transitory decline in progesterone
Ž .
concentrations in llamas, but in the closely related alpaca L. pacos , prostaglandin
metabolite peaks have been detected from days 8 to 13 of pregnancy, indicating a temporal relationship between the progesterone decline and PGF2a pulsatile release ŽAba et al., 1997 ..
Ž .
Flunixin meglumine FM , a non-steroidal anti-inflammatory drug, has the ability to reduce prostaglandin biosynthesis by inhibiting the enzyme cyclo-oxygenase. In conse-quence, when used intensively, the drug has shown the ability to inhibit the synthesis of PGF2a to the extent that luteolysis is prevented in cattle, without interfering irreversibly
Ž .
with the pulsatile mechanism of prostaglandin Aiumlamai et al., 1990 .
The objective of the present study was twofold: first, to investigate the plasma concentrations of progesterone in relation to concentrations of 15-ketodihydro-PGF2a ŽPGFM during the period of luteolysis and maternal recognition of pregnancy in llamas. and, second, to evaluate the ability of FM to inhibit luteolysis in non-pregnant llamas.
2. Materials and methods
2.1. Animals
Ž .
Eleven sexually mature female llamas L. glama , 3–5 years old with an average
Ž .
weight of 105 kg range 95–120 , were used. The animals were kept in a natural pasture
Ž X
at the Faculty of Veterinary Sciences, UNCPBA, Tandil, Argentina, 37817 S.L., sea .
to successive trials, a complete rest period of 2 months was provided to the animals between treatments in order to circumvent possible carry-over effects of the previous treatment to the outcome of the following.
2.2. Treatments
Ž . Ž
Intra-vaginal sponges containing medroxyprogesterone acetate MPA 120 mg, .
Syntex, Buenos Aires, Argentina were inserted and left in the vagina for 9 days. Six
Ž .
days after removal of the sponges day 15 , 10 animals were bred with an intact male, while 11 llamas were mated with a vasectomized male. Five of the animals mated with
Ž w
the vasectomized male were treated with FM Finadyne vet., Schering-Plough, Union,
. Ž .
NJ, USA . FM was injected intravenously every 6 h 06:00, 12:00, 18:00 and 24:00 h at a dose of 2.2 mgrkg from days 6 to 12 post-copulation.
2.3. Blood sampling
Blood samples were collected daily from days 3 to 6 post-mating. Thereafter, samples
Ž .
were obtained every second hour during the day 08:00–20:00 h and every fourth hour
Ž .
during the night 20:00–08:00 h until day 16 post-mating. In animals treated with FM, the first sample was collected on day 5 post-mating, and the following samples were collected at 2-h intervals from 06:00 to 24:00 h from days 6 to 12 after mating. All blood samples were collected by venipuncture and immediately drawn into heparinized tubes. Plasma was separated by centrifugation and stored at y208C until analysed. In order to minimise possible damage to the jugular veins from the sampling protocol, puncture was performed according to a schedule at high, medium and low positions, on both the left and right sides of the neck.
2.4. Hormone assays
Progesterone was assayed with an enhanced luminescence immunoassay technique ŽAmerlite; Kodak Clinical Diagnostics, England previously validated for llama plasma. ŽAba et al., 1995 . The intra-assay coefficient of variation calculated from the precision. profiles was below 8% for concentrations between 2 and 160 nmol ly1. The inter-assay
Ž y1.
coefficients of variation, for three quality-control samples were 7% 2 nmol l , 8%
Ž y1. Ž y1. y1
18 nmol l and 6% 54 nmol l . The sensitivity of the assay was 0.2 nmol l . The plasma metabolite of PGF , PGFM, was analysed by RIA according to Kindahl2a
Ž . Ž .
et al. 1976 and Granstrom and Kindahl 1982 . The intra-assay coefficients of variation
¨
were below 8% for samples containing 240 and 485 pmol ly1. The correspondinginter-assay coefficients of variation were 5.5% and 8.4%, respectively. The practical detection limit of the assay was 30 pmol ly1.
Ž
Oestradiol-17b concentration was determined using RIA kit Diagnostic Products,
. Ž
Los Angeles, CA, USA , reported for use with bovine plasma Sirois and Fortune,
. Ž
1990 , and validated for use with llama plasma after minor modifications Aba et al., .
y1 Ž
l . The inter-assay coefficients of variation for three control samples were 22% 13
y1. Ž y1. Ž y1.
pmol l , 6% 39 pmol l and 10% 84 pmol l . The lowest amount of
Ž .
oestradiol-17b detectable defined as the intercept of maximal bindingy2 SD was 4 pmol ly1.
Hormone concentrations are expressed in SI units. To convert from pmol ly1 to pg
mly1 and from nmol ly1 to ng mly1 the following factors should be used: PGF 2a
metabolite: 2.8; oestradiol-17b: 3.7 and progesterone: 3.2.
2.5. Analysis of data
Basal concentrations of PGF2a metabolite were calculated by averaging the values obtained on each particular day and removing values higher than two standard devia-tions from the mean value, until the baseline remained unchanged. Concentradevia-tions that exceeded the baseline by more than two standard deviations between days 7 and 12 were defined as peaks. The amount of PGF2a released during each peak was estimated by calculating the areas under the release curve according to the formula:
PGF2a releasesÝ
Ž
Ž
PGFMiqPGFMiq120.
r2.
=120 min,where is0, 120, 240, . . . , min and where PGFMisbasal concentration was consid-ered equal to 0. Daily progesterone concentrations were estimated by averaging the concentrations measured in the sample collected at 08:00, 16:00 and 24:00 h for each
Ž .
individual animal. Analysis of variance, using a repeated measures within-SS design, was applied to detect differences in hormone concentrations. In all cases, a
least-signifi-Ž .
cant difference test LSD was used to determine differences between means. Because of variations in the time required to attain peak progesterone concentrations in pregnant animals, values were normalised against the highest concentration during days 7, 8 and 9. The mean peak value was further compared with mean concentrations attained during the following days. All statistical analyses were carried out using the StatisticarW,
Ž .
release 4.0, software package Statsoft, USA Statistica for Windows, 1993 . Results are expressed as mean"SEM.
3. Results
The progesterone profiles showed that all animals ovulated in response to copulation. Increasing concentrations of progesterone were recorded by day 4 post-mating. Five out
Ž .
of 10 llamas mated with an intact male became pregnant 50% . By day 10–11 after mating, all non-pregnant llamas showed progesterone concentrations close to the detec-tion limit of the assay.
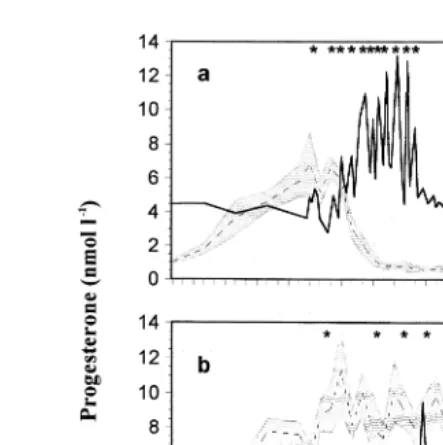
Visual examination of the results for each individual confirmed that a pulsatile pattern of prostaglandin release occurred during luteolysis in non-pregnant llamas. Fig.
Ž . Ž .
Ž . Ž .
Fig. 1. Plasma concentrations of progesterone mean, dotted line,"SEM, shaded area in non-pregnant a
Ž . Ž .
and pregnant b llamas and a prostaglandin metabolite secretory profile solid line in a representative animal from days 3 to 16 post-mating. Values identified as significant pulses of PGF2a metabolite are indicated by asterisks. Note the logarithmic scale for the prostaglandin metabolite values.
in most of the animals. PGFM pulses were registered between days 8 and 9 and day 13 after mating in non-pregnant animals. Since no significant differences in the hormonal secretory patterns were registered between non-pregnant llamas, whether mated with an intact or a vasectomized male, all animals were considered as one group for further analysis of PGFM profiles. Table 1 shows the characteristics of luteolytic pulses of
Table 1
Ž .
Characterisation of normal luteolytic release of PGF2a as determined by PGFM analysis in llamas ns11 . Numbers within parentheses in the second column represent the mean number of peaks detected on each particular day. Ms08:00 h, As16:00 h, Ns24:00 h
Days Cumulative Area under Peak-to-peak Mean peak Progesterone y1
Ž . Ž . Ž .
after number curve peaks interval h amplitude pmol l
y1
y1 Ž .
mating of peaksr Žpmol l . Ž .% pmol l M A N
animal
7 0.3 2.6 0.1 – 465 7.9 8.3 9.3
Ž .
8 1.4 1.1 49.7 3.6 19.6 748 8.9 7.6 6.5
Ž .
9 4.3 2.9 358.9 28.6 7.5 1112 4.9 3.3 1.9
Ž .
10 8.2 3.8 520.8 45.7 7.1 1234 1.0 0.7 0.6
Ž .
11 10.5 2.3 227.2 19.9 9.1 779 0.5 0.7 0.5
Ž .
Table 2
Ž .
Mean "SEM plasma progesterone concentrations from days 10 to 16 after mating in pregnant llamas
compared with the mean maximum concentration between days 7 and 9 y1
Ž .
Mean days 7–9 Plasma progesterone concentrations nmol l
Day 10 Day 11 Day 12 Day 13 Day 14 Day 15 Day 16
UU UU UU UU UU U
13.00 9.00 8.93 9.26 8.83 8.38 9.66 9.15
"1.90 "0.72 "0.61 "0.70 "0.63 "0.64 "0.94 "0.83
PGFM during luteolysis days 7–12 after mating in non-pregnant animals ns11 . In
Ž y1.
all cases, the initial pulses were lower in amplitude 525.9"36.7 pmol l than those
Ž .
recorded later P-0.01 . The mean peak concentration of the luteolytic pulses was
y1 Ž .
1019"48.5 pmol l . On average, 11.6 range 10–14 peaks per animal were recorded between days 7 and 12 post-mating. Decreasing concentrations of progesterone were already registered by the afternoon of day 8, when only one or two prostaglandin metabolite peaks were recorded in each individual animal. Although all animals had reached basal plasma concentrations of progesterone on day 10 after mating, PGFM peaks were recorded during the following 2 days.
In pregnant animals, plasma concentrations of progesterone remained high until the
Ž Ž ..
end of the experiment Fig. 1 b . Peak plasma progesterone concentrations were attained between days 7 and 9. When the mean progesterone concentrations registered on those days were compared with the mean concentrations registered from days 10 to
Ž .
16, a significant drop in progesterone was seen after day 9 Table 2 . In connection with the progesterone decline, PGF2a pulsatile release was observed in all pregnant animals
Ž .
from days 7 to 15 after mating. Fig. 1 b shows the PGFM concentrations in a representative llama. The characteristics of prostaglandin release during early pregnancy Ždays 7–12 after mating are shown in Table 3. The overall mean frequency of PGF. 2a
Table 3
Ž .
Characterisation of PGF2arelease during early pregnancy in llamas as determined by PGFM analysis ns5 . Numbers between parentheses in the second column represent the mean number of peaks detected on each particular day. Ms08:00 h, As16:00 h, Ns24:00 h
Days Cumulative Area under Peak-to-peak Mean peak Progesterone y1
Ž . Ž . Ž .
after number curve peaks interval h amplitude nmol l
Ž . Ž .
Fig. 2. Plasma concentrations of oestradiol-17b mean"SEM, shaded area in non-pregnant a and pregnant
Ž .b llamas from days 3 to 16 post-mating.
metabolite peaks detected in pregnant animals was 0.64 peaksranimalrday, while the mean peak amplitude was 503.8"49.1 pmol ly1.
Oestradiol-17b plasma concentrations increased slowly from days 3 to 8 after mating in all lamas. Thereafter, concentrations sharply increased in non-pregnant llamas, until
Ž y1.
peak concentrations 30.9"3.8 pmol l were attained on day 15. Conversely, a slow oestradiol-17b rise was recorded in pregnant animals in which the highest
concentra-Ž y1. Ž Ž . Ž .
tions 11.2"4.9 pmol l were observed 12 days after mating Fig. 2 a and b .
respectively .
Ž .
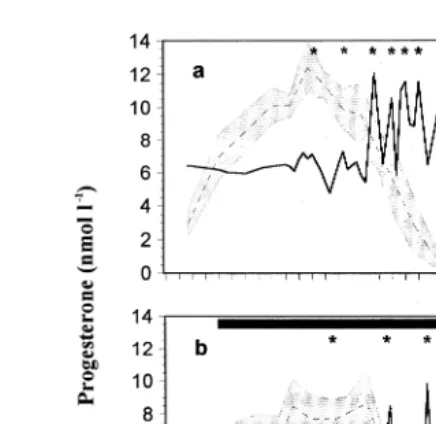
Fig. 3. Plasma concentrations of progesterone mean, dotted line,"SEM, shaded area in non-pregnant llamas
Ž . Ž .
mated with a vasectomized male a and in non-pregnant llamas treated with FM black bar four times a day
Ž .b , and a prostaglandin metabolite secretory profile solid line in a representative animal from days 5 to 12Ž .
Prostaglandin metabolite concentrations decreased rapidly in all animals after the first administration of FM from about 450 pmol ly1 to concentrations of around 180 pmol
y1 Ž .
l P-0.01 . Concentrations of the metabolite remained low throughout the first 2
Ž .
days of FM treatment days 6 and 7 post-mating . Thereafter, pulsatile release of PGF2a
started, and concentrations similar to those recorded during the first 2 days of treatment were recorded between peaks.
Fig. 3 shows the changes in hormone concentrations in non-pregnant llamas mated
Ž . Ž .
with a vasectomized male during normal luteolysis a and during FM treatment b . In
Ž .
llamas treated with FM, the cumulative number of peaksranimal were 1 day 8 , 2.2 Žday 9 , 4.4 day 10 , 5.8 day 11 and 6 day 12 . The mean peak prostaglandin. Ž . Ž . Ž .
y1 Ž . y1 Ž . y1
metabolite amplitudes were 261 pmol l day 8 , 547 pmol l day 9 , 561 pmol l Žday 10 , 993 pmol l. y1 Žday 11 and 864 pmol l. y1 Žday 12 . Increasing concentrations.
of progesterone were recorded from days 5 to 7 post-mating in both groups. Thereafter, progesterone concentrations declined and were close to the detection limit of the assay by day 9 post-copulation in untreated animals. Progesterone concentrations remained high until day 8 post-mating, after which, they started to decrease until basal concentra-tions were attained by day 10 or 11 post-mating.
4. Discussion
The PGFM, pattern registered during luteolysis, correlates well with previous reports showing the close temporal association between pulsatile release of PGF2a and
termina-Ž .
tion of the corpus luteum function in llamas Sumar et al., 1988; Aba et al., 1995 . The sampling schedule used in this study, allowed a more detailed study of the luteolytic pulses of PGF2a in llamas, although some peaks could have been missed. As has been
Ž
shown in cattle, sheep and goat Fredriksson et al., 1984; Basu and Kindahl, 1987; Zarco
. Ž .
et al., 1988b , the first peaks registered on days 7 and 8 had consistently lower amplitudes than those recorded during the following days. Calculating the area under the peaks revealed that about 35% of the total release of PGF2a occurred during days 8 and 9, when luteolysis was at its highest, while about 65% of the total release was seen after progesterone levels had decreased below 1 nmol ly1. Sampling was not frequent enough to permit an accurate estimation of the duration of pulses. However, based on the observation that increased PGFM concentrations were registered in two successive samples in most cases during the period of maximum secretion, an estimate of 4–5 h for peak duration may be suggested. Moreover, it was observed during this period that in several cases, concentrations did not reach basal values between two consecutive peaks, indicating that the interval between the end of one peak and the beginning of the next could be shorter than 2 hours. This suggestion is supported by the observation that most
Ž .
of the animals had at least four peaks during 1 day day 10 . The overall secretory pattern of PGF2a appears to be similar to that observed in other ruminants. However, PGF2a pulses during the period of luteolysis seemed to be somewhat shorter in duration
Ž and to be produced at a higher frequency in llamas than those reported in cattle Basu
Ž
The luteolytic release of PGF2a starts on day 7 or 8 post-mating approximately day .
5–6 after ovulation and is completed by day 9 or 10 after mating in non-pregnant llamas. It has also been shown that in South American camelids, the first significant
Ž
increase in progesterone concentrations is observed 3–4 days after mating Sumar et al., .
1988; Bravo et al., 1996 . Exposure to an environment characterised by high proges-terone concentrations is claimed to play a key role in the regulation of uterine oxytocin
Ž
receptors and, consequently, of prostaglandin pulsatile release in ruminants Silvia et al., .
1991; Lamming and Mann, 1995 . Such a period lasts only 3–4 days in llamas, while it lasts around 3 and 5 times longer in the sheep and in the cow, respectively. In addition, it is apparent that luteal activity after a sterile mating in llamas, is considerably shorter than that observed in other induced ovulators. Thus, using the length of the normal pregnancy for each specie as 100%, the lifespan of the CL in non-pregnant animals
Ž .
varies from 100% in mink and ferret Møller, 1973; Heap and Hammond, 1974 , to
Ž .
approximately 55–60% in rabbit and cat Paape et al., 1975, Rowlands and Weir, 1984 and to about 3% in llamas. The mechanism behind this rapid increase in sensitivity and the responsiveness of the developing CL to prostaglandins remains to be determined in llamas.
In cows, no prostaglandin peaks are seen during maternal recognition of pregnancy ŽBasu and Kindahl, 1987 . Similarly, there is no pulsatile pattern in pregnant goats. during this period, but PGFM concentrations in the peripheral circulation start to
Ž .
increase on day 14 or 15 Fredriksson et al., 1984 . In pregnant sheep, although no pulsatile pattern has been observed, two or three PGF2a pulses of low amplitude, separated by long intervals, can be distinguished during maternal recognition of preg-nancy. However, no changes in plasma progesterone concentrations have been reported
Ž .
in connection with these pulses Zarco et al., 1988a . In the present study, prostaglandin peaks were recorded in all pregnant llamas during the period when maternal recognition of pregnancy was expected to occur. The magnitude and the frequency of the pulses registered during days 7–12 of pregnancy were not comparable to those observed in non-pregnant animals. The amount of PGFM recorded as peaks during early pregnancy averaged about 3% of the total amount of prostaglandin released during luteolysis; nevertheless, this prostaglandin production proved to be effective in inducing a decline in progesterone concentrations. A similar transient decrease in progesterone concentra-tions during maternal recognition of pregnancy has been reported previously in llamas
Ž .
and alpacas Adams et al., 1990; Aba et al., 1995,1997 . Prostaglandin metabolite peaks with relatively low amplitude have been observed in connection with the decline in
Ž .
progesterone concentrations in pregnant alpacas Aba et al., 1997 .
The mechanism by which the pulsatile release of PGF2a from the uterus is depressed Žbut not suppressed in the pregnant llama is not known. Attempts to identify any.
Ž
substance similar to ovine or bovine interferons in camel the related Old World .
camelids conceptus incubates from days 10 to 33 after ovulation have been
unsuccess-Ž .
ful Skidmore et al., 1994 . Moreover, no increase in oestrone sulphate concentrations in urine or oestradiol-17b in plasma has been recorded during early pregnancy in South
Ž .
American camelids Bravo et al., 1991; Aba et al., 1995 .
progesterone from the corpus luteum exerts a negative influence on follicular activity
Ž .
during early pregnancy in llamas Adams et al., 1990; Aba et al., 1995 .
The dose of FM used here, proved to be effective in depressing prostaglandin synthesis. This effect was clearly seen during the first 2 days of the treatment and during the periods between peaks on the successive days. A negative influence on the pulsatile pattern of PGF2awas also observed. Thus, both the amplitude of the PGFM peaks and the cumulative number of peaks per animal were significantly lower in llamas treated with FM than in untreated animals; but this negative effect was not strong enough to completely inhibit the occurrence of luteolytic pulses of PGF , and luteolysis pro-2a
ceeded in these animals. However, the progesterone patterns indicated that luteolysis was delayed for 1–1.5 days in treated llamas, as compared to untreated animals. A similar dose of FM has been previously proven to be effective in other ruminants to prevent luteolysis; however, earlier reports indicate that the FM half-life is shorter in the related camel than it is in the cow. The authors concluded that, using the same dose regimen in camels as has been used in cattle, would result in lower pharmacological
Ž .
effectiveness Oukessou, 1994; Ali et al., 1996 .
Acknowledgements
Ž .
The International Foundation for Science IFS , the Swedish Council for Forestry and
Ž .
Agricultural Research SJFR and the Swedish Agency for Research Co-operation with
Ž .
Developing Countries SAREC , are acknowledged for financial support.
References
Aba, M.A., Bravo, P.W., Forsberg, M., Kindahl, H, 1997. Endocrine changes during early pregnancy in the alpaca. Anim. Reprod. Sci. 47, 273–279.
Aba, M.A., Forsberg, M., Kindahl, H., Sumar, J., Edqvist, L.-E., 1995. Endocrine changes after mating in pregnant and non-pregnant llamas and alpacas. Acta Vet. Scand. 36, 489–498.
Adams, G.P., Sumar, J., Ginther, O.J., 1990. Effect of lactational and reproductive status on ovarian follicular
Ž .
waves in llamas Lama glama . J. Reprod. Fertil. 90, 535–545.
Adams, G.P., Sumar, J., Ginther, O.J., 1991. Form and function of the corpus luteum in llamas. Anim. Reprod. Sci. 24, 127–138.
Aiumlamai, S., Odensvik, K., Stabenfeldt, G., Kindahl, H., 1990. Regulation of prostaglandin biosynthesis with flunixin meglumine in the bovine species. J. Vet. Med. A 37, 16–22.
( Ali, B.H., Oukessou, M., Bashir, A.K., 1996. Pharmacokinetics considerations in the camel Camelus
)
dromedarius : a review. Comp. Biochem. Physiol. 115, 1–9.
Basu, S., Kindahl, H., 1987. Development of a continuous blood collection technique and a detailed study of prostaglandin F2a release during luteolysis and early pregnancy in heifers. J. Vet. Med. A 34, 487–500. Bravo, P.W., Stabenfeldt, G.H., Fowler, M.E., Lasley, B.L., 1991. Urinary steroids in the periparturient and
postpartum periods through early pregnancy in llamas. Theriogenology 36, 267–278.
Bravo, P.W., Stewart, D.R., Lasley, B.L., Fowler, M.E., 1996. Hormonal indicators of pregnancy in llamas and alpacas. J. Am. Vet. Med. Assoc. 208, 2027–2030.
Granstrom, E., Kindahl, H., 1982. Radioimmunoassay of the major plasma metabolite of PGF¨ 2a, 15-keto-13,14-dihydro-PGF2a. Methods Enzymol. 86, 320–339.
Heap, R.B., Hammond, J. Jr., 1974. Plasma progesterone levels in pregnant and pseudopregnant ferrets. J. Reprod. Fertil. 39, 149–152.
Kindahl, H., Edqvist, L.-E., Granstrom, E., Bane, A., 1976. The release of prostaglandin F¨ 2aas reflected by 15-keto-13,14-dihydroprostaglandin F2a in the peripheral circulation during normal luteolysis in heifers. Prostaglandins 11, 871–878.
Lamming, G.E., Mann, G.E., 1995. A dual role for progesterone in the control of cyclicity in ruminants. J. Reprod. Fertil. 49, 561–566.
Ž .
Møller, O.M., 1973. The progesterone concentrations in the peripheral plasma of the mink MustelaÕison
during pregnancy. J. Endocrinol. 56, 121–132.
( )
Oukessou, M., 1994. Kinetic disposition of flunixin meglumine in the camel Camelus dromedarius . Vet. Res. 25, 71–75.
Paape, S.R., Shille, V.M., Seto, H., Stabenfeldt, G.H., 1975. Luteal activity in the pseudopregnant cat. Biol. Reprod. 13, 470–474.
Rowlands, I.W., Weir, B.J., 1984. Reproductive cycles in vertebrates; Mammals: non-primate eutherians. In:
Ž .
Lamming, G.E. Ed. , Marshall’s Physiology of Reproduction. Churchil Livingstone, pp. 458–658. Silvia, W.J., Lewis, G.S., McCracken, J.A., Thatcher, W.W., Wilson, L., 1991. Hormonal regulation of uterine
secretion of prostaglandin F2aduring luteolysis in ruminants. Biol. Reprod. 45, 655–663.
Sirois, J., Fortune, J.E., 1990. Lengthening the bovine estrous cycle with low levels of exogenous proges-terone: a model for studying ovarian follicular dominance. Endocrinology 127, 916–924.
Skidmore, J.A., Allen, W.R., Heap, R.B., 1994. Maternal recognition of pregnancy in the dromedary camel. Proceedings of the 4th International Symposium on Reproduction in Domestic Animals, 46, Townsville, Australia.
Ž .
Statistica for Windows 1993 . Release 4.0, E software package, Statsoft, Tulsa, OK USA.
Sumar, J., Fredriksson, G., Alarcon, V., Kindahl, H., Edqvist, L.-E., 1988. Levels of 15-keto-dihydroPGF2a, progesterone and oestradiol-17b after induced ovulations in llamas and alpacas. Acta Vet. Scand. 29, 339–346.
Zarco, L., Stabenfeldt, G.H., Basu, S., Bradford, G.E., Kindahl, H., 1988a. Modification of prostaglandin F2a synthesis and release in the ewe during the initial establishment of pregnancy. J. Reprod. Fertil. 83, 527–536.