Fine structure, carbohydrates and photosynthetic pigments
of sugar maize leaves under UV-B radiation
Michael Barsig *, Ralf Malz
Institut fu¨r O8kologie u.Biologie,TU Berlin,D-10587Berlin,Germany
Received 27 July 1999; received in revised form 7 October 1999; accepted 7 October 1999
Abstract
Leaves ofZea mayswere subjected to different scenarios of ultraviolet-B radiation in a sun simulator to determine the cellular vitality at the microscopic level and the contents of carbohydrates and photosynthetic pigments. The results show that the leaf morphology and fine structure of sugar maize leaves are only slightly affected by UV cut-off wavelengths down to 288 nm. At a microscopic level, a number (about one third) of epidermal cells, predominantly below leaf tips, are affected by supplemental UV-B. They showed partially collapsed or deformed cell walls and membranes. Underneath damaged epidermal cells, a slight enlargement of intercellular space could be observed. However, other cell components including thylakoid membranes and the epicuticular wax layer on the adaxial leaf surface remained unaffected. The leaf dry weight was not influenced by enhanced UV-B. The amount and structure of starch grains in leaf chloroplasts did not differ between UV treatments. There was no clear impact of enhanced UV-B on sucrose content. Carbohydrate partitioning was more significantly influenced by leaf exposure than by UV treatment. Only glucose was decreased under high UV-B. Changes in photosynthetic pigments were limited to a slight destructive effect of UV-B on chlorophyllb. The results presented in this study may be interpreted that sugar maize leaves have adapted efficiently to cope with supplemental UV-B radiation. Changes in epidermal cell layer due to their shield function may indicate that this remarkable resistance against enhanced UV-B radiation is not unlimited. © 2000 Elsevier Science B.V. All rights reserved.
Keywords:Carbohydrates; Epidermis; Maize leaf; Photosynthetic pigments; Ultrastructure; UV-B; Wax layer;Zea mays
www.elsevier.com/locate/envexpbot
1. Introduction
Global changes in the chemical composition of the atmosphere with a resulting thinning of the protective ozone layer can have considerable
im-pacts on plant life (Caldwell and Flint, 1994). Considerable higher intensity of ultraviolet-B (UV-B) radiation (280 – 320 nm) at the Earth’s surface, resulting from anthropogenic depletion of the stratospheric ozone, can have potentially dam-aging effects to a wide range of plant species (Teramura, 1990). UV-B effects on plants can comprise alterations of the genome, the chemical quality and the photosynthetic capacity of the * Corresponding author. Tel.:+49-30-31473197; fax:+
49-30-31473177.
E-mail address:[email protected] (M. Barsig)
plant tissue (Tevini, 1993). In some studies, it was shown that the ultrastructure of photosynthetic plant organs may be changed by UV-B radiation (Santos et al., 1993; Lu¨tz et al., 1997; Barsig et al., 1998). However, the information on ultrastruc-tural changes by increasing UV-B is limited. Even though Zea mays is one of the most important cereal crops, knowledge of its resistance to en-hanced UV radiation is limited. Previous investi-gations have shown growth reductions or leaf damage in different maize cultivars (Santos et al., 1993; Mark and Tevini, 1996).
The sun simulator at the GSF Research Center near Munich, Germany, provides realistic white light conditions which is a prerequisite for ecolog-ically relevant UV experiments (Meindl and Lu¨tz, 1996). For this study a realistic scenario of UV-B enhancement under intensive light conditions was simulated. Unrealistic high UV-B doses as used in previous investigations were avoided. The aim of this study is to describe UV-B effects on cell fine structure and on carbohydrate and pigment pools simultaneously. We investigated the impact of different UV-B irradiances on epicuticular wax structure, on the epidermis and the underlying cell layers. The structural stability of leaf components under enhanced UV-B is a prerequisite for their functional vitality. Therefore, carbohydrate and pigment content may be a possible marker. The initial hypothesis of this study was that the sugar maize cultivar (Z. mays cv. saccharata Koern ‘Tasty Sweet’) is a UV-sensitive plant species at the level of leaf vitality.
2. Material and methods
2.1. Plant material and growth conditions
Seeds of sugar maize (Z.maysL. cv. saccharata Koern ‘Tasty Sweet’ supplied by Agrisaaten) were planted in 20-cm pots containing a loam/quartz sand mixture and grown under Central European mid summer conditions measured at Neuherberg (48.2° N, 11.5° E, altitude 500 m) in July. After emergence of the sixth leaf, plants were exposed in the GSF sun simulator. The GSF sun simulator provides light conditions of ambient Central
Eu-ropean mid summer conditions. A combination of different lamps simulates the visible light, com-pared to spectroradiometric field data obtained on a cloudless summer day at 50° N with approx. 60° maximum solar elevation (Do¨hring et al., 1996; Thiel et al., 1996). By the lighting technology in the GSF Phytotron the self-shading of the leaves was minimized, thus possible interactions with the response of maize plants to UV-B can be neglected.
The climate used in the simulators was a weekly repeated radiation, temperature and humidity regime with temperature variation of 16 – 28°C (with a range of daily maxima between 22 and 28°C). The relative humidity was kept at 90% (at night) and continuously reduced to a minimum of 60% (in the afternoon). As a UV-B free control (UV-BB0.1 W m−2), a WG 360 filter (Schott) was used. All irradiation data were obtained from spectroradiometry using a double monochroma-tor system (Bentham DM 300) with a cosine corrected input optics (Thiel et al., 1996).
2.2. UV-B simulation
has been conducted in two sun simulator cham-bers for 16 days.
2.3. Macroscopic obser6ations
Immediately after the end of their exposure in the sun simulator, leaf samples from the treat-ments were observed for growth characteristics and vitality under a binocular microscope (25×).
2.4. Light and electron microscopy
For transmission and scanning electron mi-croscopy, segments (5 mm below the tip and of the middle part) of the fourth leaf of each treated plant were fixed in 2.5% glutaraldehyde and postfixed in 2% osmium tetroxide in 0.1 M phos-phate buffer, pH 7.1. Dehydration was accom-plished in a graded acetone series. The samples were infiltrated and embedded in Epon (Spurr). Semi-thin sections (1-mm cross sections) were cut with a Reichert OM U3 ultratome and stained with azur-II-methylene-blue for preliminary screening with a light microscope. For total leaf and epidermal cell thickness they were measured with a light microscope connected to the imaging analysis system SYS®. Thin sections were cut with a diamond knife, stained with Reynold’s lead citrate and examined with a Siemens Elmi 1 A TEM at 80 kV. For scanning electron microscopy, leaf segments were fixed in glutaraldehyde, dehy-drated in acetone, critical-point dried, sputtered
with gold and examined in a Hitachi-S-4000 REM at 20 kV.
2.5. Determination of dry weight
Dry weight of all leaves was determined by freeze-drying of the material.
2.6. Determination of carbohydrate metabolites
Leaf samples were separated according to their phyllotaxis into three groups (a: leaf 2 – 3; b: leaf 4; c: leaf 5 – 6), were immediately quick-frozen in liquid nitrogen, freeze-dried for 24 h (Christ Al-pha I-5), homogenized by grinding in a ball mill (Retsch) and stored at −25°C.
2.6.1. Starch
Five hundred milligrams lyophilized powder was extracted at room temperature with 80% (v/v) ethanol. Following centrifugation (10 000×g, 10 min), the supernatant (containing the low-molecu-lar, soluble carbohydrates) was discarded. Follow-ing two further treatments with 80% ethanol, the pellet was hydrolyzed by heating in 25 – 30 ml bidistilled water (95°C, 1 h) and cooled. The pH was adjusted to 4.68 – 4.74 by addition of sodium acetate. Amyloglucosidase (Aspergillus niger L., EC 3.2.1.3) was added (incubation for 1 h, 58°C, water-bath), followed by boiling (95°C), addition of charcoal (75 mg) and centrifugation (10 000×
g, 10 min, twice). After filtration, an aliquot of the supernatant was used for enzymatic determination with equipment by Boehringer according to Beut-ler (1985a).
Table 1
Integrated data of the irradation regimes used in the UV-B simulation
Spectrum no.
(1) (2) (3) (4)
Sanalux
Filter 1 Sanalux Sanalux Sanalux
Filter 2 Plexiglas Sanalux Pyran Float glass
4.73 2.15
UV-B (280–320 nm) (W m−2) 1.27 0.03
35.4 33.2
UV-A (320–400 nm) (W m−2) 30.0 28.2
Max. 1800 PAR (400–700 nm) (mol m−2s−1)
103 105 102 100
Max. illuminance (IL) (kLux)
281 288 293 306
2.6.2. Sucrose,glucose and fructose
One hundred milligrams of maize-powder was extracted in 65% aqueous ethanol (v/v) at 68°C for 1 h after pH-adjustment to ]6 to avoid hydrolysis of sucrose. After centrifugation as above the starch-containing sediment was re-moved and charcoal was added to reduce the blank reading. Following centrifugation, aliquots were assayed according to Beutler (1985b) and Boehringer (1989). All extraction procedures de-scribed here were optimized in order to obtain a recovery of external standards of more than 90%.
2.7. Pigment analysis
Pigments were analysed in the fourth leaves. They were lyophilized as described above. Chloro-phylls and carotenoids were extracted in 80% acetone. The extract was centrifuged twice at 5300×g for 10 min and analysed spectrophoto-metrically (Shimadzu UV-160A) at 646 and 663 nm for chlorophylls and at 470 nm for carotenoids. Chlorophylls and carotenoids were calculated per unit dry weight according to Licht-enthaler and Wellburn (1983).
2.8. Statistical analyses
Significance of differences between treatments was determined by either Mann – Whitney U-test for nonparametric data or by one-way and multi-ple ANOVA followed by LSD or Scheffe´ means tests for parametric data.
3. Results
3.1. Leaf morphology and leaf dry weight
No morphological changes or discolouration of leaves independent of UV irradiation were ob-served. Leaf length was not affected by enhanced UV-B (not shown in detail). The leaf dry weight varied between 12.4 and 14.9% of fresh weight and was not influenced by different UV-B irradiances.
3.2. Light and electron microscopic obser6ations
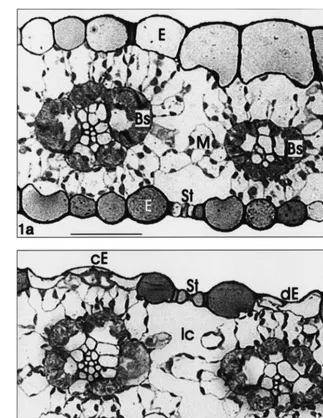
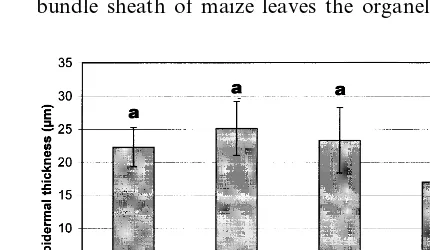
In comparison to controls (Fig. 1a) most of the leaf tissues were intact even under high UV-B. The only remarkable deviation consisted of de-formed or collapsed cells predominantly in the adaxial epidermal layer under enhanced UV-B. At the ultrastructural level all organelles and mem-branes in such cells were affected. This pattern was accompanied by a slight enlargement of the neighbouring intercellular space (Fig. 1b). This was confirmed by measurements revealing a sig-nificantly (P50.001) reduced thickness of epider-mal cells under enhanced UV-B radiation (Fig. 3). Because only the thin epidermal layer was af-fected, the total leaf thickness was not signifi-cantly changed under high UV-B. By light microscopic measurements, 33.9912.5% (com-pared to 11.7917.2% under ambient UV-B) of epidermal cells were damaged (significant differ-ence between variables with P50.01). On this occasion mostly adaxial cells were affected and showed deformed cell walls and reduced cell lu-mina. The proportion of affected epidermal cells was especially increased in leaf samples below the tip (with an increase of 130% compared to the middle parts of leaves under high UV-B).
At the ultrastructural level (Fig. 2a), no devia-tions have been observed in shape, size or struc-ture of organelles in assimilation and bundle sheath cells. Chloroplasts and nuclei remained intact even after enhanced UV-B radiation. The conducting tissues showed no alterations and structure or amount of vacuolar contents ap-peared independent of UV-B exposure.
SEM observations revealed that the epicuticular wax layer was intact. Small regions with slight modifications showing erosion in tubular waxes were found in all UV-B treatments and controls. Wax structures in and around stomata were unaf-fected (Fig. 2b).
3.3. Carbohydrates
Fig. 1. Light microscopy of semi-thin transverse sections ofZ.maysleaf blades. (a) Intact tissues grown under low UV-B; Bs, bundle sheath; E, epidermal cells; Ic, intercellular space; M, mesophyll cells; St, stomata. (b) UV-B treated leaf (70% enhanced in relation to ambient radiation). Some epidermal cells are deformed or collapsed and below the intercellular space is slightly enlarged; mesophyll cells, leaf bundles and stomata are undisturbed; cE, collapsed epidermis; dE, deformed epidermis. Scale bars=50mm.
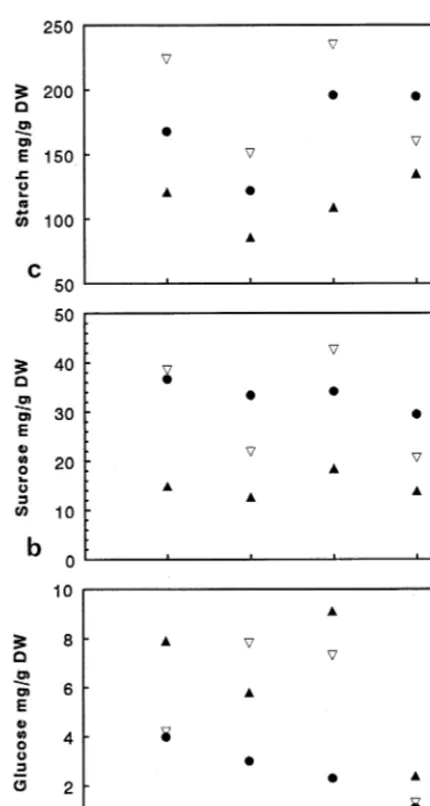
glucose are compared in Fig. 4a – c for leaves 2/3 and 4, 5/6, respectively. The content of fructose was very low (B1 mg g−1 dry mass) in all treatments. The sucrose and starch partitioning varied more in dependence of the leaf exposure without showing a significant impact of UV-B treatment (Fig. 4b,c). Otherwise, the glucose
Fig. 2. (a) Transmission electron micrograph of leaf under supplemental UV-B. Normal structure of mesophyll (asterisks) and of bundle sheath chloroplasts (with numerous starch grains). Other organelles are also intact. Scale bar=1mm. (b) Scanning electron micrograph of adaxial leaf surface under enhanced UV-B. The epicuticular wax fine structure is predominantly unaffected, also around the stomatal opening. Scale bar=20mm.
3.4. Pigment changes
There was no significant impact of variable UV levels simulated in this study on chlorophylla or carotenoid contents. Under enhanced UV-B, a slight decrease of chlorophyll b occurred. The ratio between chlorophyll a and b was increased under enhanced UV-B but was decreased under ambient UV-B in comparison to low UV-B radia-tion (Table 2).
4. Discussion
Leaves ofZ.mays cv. saccharata Koern ‘Tasty Sweet’ were not heavily affected by enhanced UV-B radiation in a phytotron experiment with careful shaping of the spectral irradiance. This result is in accordance with a report of Meindl and Lu¨tz (1996), where green algae were only UV-sensitive when lowering the cut-off wave-length to 280 or 275 nm (already UV-C). Cross-sections revealed that maize leaf tissues, except for the epidermis, were formed normally (similar to descriptions for other maize cultivars by Russell and Evert, 1985; Bosabalidis et al., 1994; Bon-gard-Pierce et al., 1996; Evert et al., 1996; Russin et al., 1996) independent of UV treatment. More drastic changes in structure and function of
mem-brane systems were observed inZ.mayscv. LG12 by Santos et al. (1993) or in the mossPolytrichum commune by Barsig et al. (1998). Santos et al. (1993) described UV-B effects on maize leaves by vesiculation of dictyosomes, damage in mitochon-dria and an increase in rough ER (already at minor UV-B enhancement compared to our ex-perimental design). Contradictory to this study our findings have shown that in mesophyll and bundle sheath of maize leaves the organelles and
Fig. 3. Epidermal leaf thickness (means9S.D.) of corn grown under different UV irradiation (0.03 – 4.73 W m−2 UV-B).
Fig. 4. Intermediates of carbohydrate metabolism (means) in relation to UV-B irradiation in sugar maize at different leaf stage (n=16). (a) Glucose (S.D. 90.77), (b) sucrose (S.D.
95.2), (c) starch (S.D. 917.4).
of UV-B radiation and ageing or damage of leaves (Steinmu¨ller and Tevini, 1985; Barnes et al., 1996). UV-B induced changes may alter wax com-position and structure in a number of crop plants (Steinmu¨ller and Tevini, 1985; Barnes et al., 1994). But the results of this study have shown that the leaf epicuticular wax layer was struc-turally unchanged under enhanced UV-B as in experimentally UV-B radiated barley (Tevini et al., 1986).
The majority of UV radiation is absorbed by the epidermis. Under UV-B irradiation in a sun simulator the epidermal cells contained about 95% of leaf DNA damage (Stapleton et al., 1997). Supplemental UV-B may induce a collapse of adaxial epidermis cells (Bornman et al., 1983; Tevini et al., 1986; Santos et al., 1993). This effect was accompanied by reduced intercellular spaces in cucumber leaves (Tevini et al., 1986). All these studies reported a more pronounced epidermal collapse. In our investigation, under increased UV-B, we also observed collapse as only variably deformed cells without a distinct change of inter-cellular space area. We suggest that even ambient UV-B combined with high PAR also affects some adaxial epidermal cells. Under such conditions the partial damage of the epidermis was revealed by deformed cells with reduced lumina whereas col-lapsed cells were exclusively observable under supplemental UV-B. However, this pattern did not significantly lead to a reduction in total leaf thickness (not shown in detail) or dry weight, contrary to the findings of Santos et al. (1993), or effects on general plant leaf growth. In agreement with Tevini et al. (1991a) and Stapleton et al. (1997), our study indicates that epidermal cells appear to shield effectively mesophyll and con-ducting cells from most UV in the solar spectrum at the cost of alterations in epidermal structure. About one third of epidermal cells were partially damaged under enhanced UV-B especially below the tip. It would be interesting to investigate if the proportion of damaged epidermal cells would in-crease in a long-term experiment or if repair sys-tems may be efficient.
An UV-induced alteration of starch and sucrose contents could not be observed. At an ultrastruc-tural level we observed a higher starch content in endomembrane system remained undisturbed
af-ter UV-B exposure.
bundle sheath chloroplasts compared to other ul-trastructural reports (Santos et al., 1993; Evert et al., 1996). UV-B induced an accumulation of starch in cucumber leaves (Britz and Adamse, 1994). Santos et al. (1993) reported that bundle sheath chloroplasts contained more starch after enhanced UV-B exposure in relation to controls. Our results revealed no corresponding differences between leaves under low or high UV-B irradia-tion. We suggest that sugar maize generally con-tains more starch than other cultivars. A high starch content could be related to its function as a carbohydrate storage pool for the period of sugar accumulation in the kernels. The only significant effect in UV-B exposed maize leaves was a de-crease in glucose under supplemental UV-B whereas the sucrose content was not reduced. Supplemental levels of UV-B induced a decrease of soluble carbohydrates in leaves of Triticum aesti6um(Yue et al., 1998) and decreased sucrose
and starch contents in leaves of the moss Poly
-trichum commune(Barsig et al., 1998). According to Garrard et al. (1977), UV-B sensitive phanerogams are characterised by reduced amounts of sucrose. These authors concluded that maize leaves were tolerant to enhanced UV-B if carbohydrates were used as a marker. Apparently the influence of leaf exposure on carbohydrate content of leaves was not sufficiently considered by other investigators. In this study drastic changes in starch and sucrose partitioning appear to be due to leaf exposure. By corresponding ultrastructural observations of intact thylakoids in
mesophyll and bundle sheath chloroplasts, a nega-tive impact of enhanced UV-B on photosynthetic capacity of maize leaves cannot be deduced.
In this study no clear changes of chlorophylls and carotenoids due to UV-B radiation could be revealed. These results are consistent with a previ-ous UV-B radiation study (Barsig et al., 1998). The slight reduction in chlorophyllb correspond-ing with changes in the chlorophyll a/b ratio in sugar maize leaves under supplemental UV-B may be explained by a slight destructive irradiation effect in the light harvesting domains while chlorophyll a and b-carotene seem to be more stable against UV-B stress (Lu¨tz et al., 1997). The effects of UV-B radiation on chlorophyll a/b ra-tios were quite distinct in different UV studies and this heterogeneity probably reflects species and cultivar differences (Teramura, 1983). Our study suggests that these compounds do not appear to correspond to morphological patterns. These re-sults are in contrast to another investigation showing a significant decrease of total chlorophyll content combined with ultrastructural changes in leaves of the maize cultivar LG12 (Santos et al., 1993). Tevini et al. (1991b) noted that maize seedlings contained more total chlorophyll based on leaf area under enhanced UV-B. This effect was explained by a decrease in leaf area. Other-wise, these changes did not influence the growth of these maize seedlings and no sign of leaf senes-cence was observed (Tevini et al., 1990). Only further long-term investigations under field condi-tions can show if alteracondi-tions in leaf pigments are
Table 2
Concentrations of photosynthetic pigments in leaf 4 ofZ.maysafter treatment with four UV-B levels in two sun simulatorsa
Spectrum no.
Cut-off wavelength (nm) 288 293 306
5.0390.18 4.7690.30
important markers for growth and yield reduc-tions caused by a delay of plant development and flowering under higher solar UV-B radiation (Tevini, 1996).
Experimental UV conditions may influence the possible degree of leaf damage in such studies. It is interesting that changes in chloroplast thy-lakoids or other membrane systems can be more remarkable in UV experiments under greenhouse conditions (Santos et al., 1993) than under field conditions (Gehrke, personal communication). Such contradictory results may often be explained by unbalanced UV-A/UV-B/PAR ratios, which are important for the repair processes of cells (Holm-Hansen et al., 1993; Meindl and Lu¨tz, 1996). Most UV-B research experiments in the past two decades have been conducted in growth chambers and greenhouses where the unnatural spectral balance of radiation may have changed plant sensitivity to UV-B (Yue et al., 1998).
According to Stapleton et al. (1997), green maize tissues had detectable UV-induced damage but also efficient mechanisms for photorepair. Maize plants have adapted to the natural UV radiation. This is supported by our findings that leaves under ambient UV-B did not show differ-ent structural or biochemical patterns compared to low or without UV-B radiation. Protection against photooxidants and photorepair of DNA damage probably reduces deleterious effects of enhanced UV-B. In contrast to our initial hypoth-esis we conclude that even supplemental UV-B, simulating 170% of the ambient UV-B radiation in a period of 16 cloudless summer days, cannot effect a marked structural and functional distur-bance on sugar maize leaves. However, we could not measure effects on growth and yield of our plant material as in other studies. In different UV-B treatments other maize cultivars developed growth and yield reductions under enhanced solar UV-B radiation (Mark and Tevini, 1996; Tevini, 1996). Other investigations have shown a distinct intraspecific variation in sensitivity to UV-B radi-ation. Differences in response to supplemental UV-B have been observed both within and be-tween many food plant species (Teramura and Murali, 1986; Barnes et al., 1990, 1993; Ziska et al., 1992). There is a high genotypic variability in
maize cultivars for other stress factors (drought and salinity) and this range, perhaps, has strong implications for maize plantation under changed UV-B solar radiation.
Acknowledgements
We wish to thank Cornelius Lu¨tz and Britta Firsching for their support at the GSF research center in Oberschleißheim, Germany. We are also grateful to Iris Pieper for expert technical assistance.
References
Barnes, J., Paul, N., Percy, K., Broadbent, P., McLaughlin, C., Mullineaux, P., Creissen, G., Wellburn, A., 1994. Ef-fects of UV-B radiation on wax biosynthesis. In: Percy, K.E., Cape, J.N., Jagels, R., Simpson, C.J. (Eds.), Air Pollutants and the Leaf Cuticle. Springer, Berlin, pp. 195 – 204.
Barnes, J.D., Percy, K.E., Paul, N.D., Jones, P., McLaughlin, C.K., Mullineaux, P.M., Creissen, G., Wellburn, R., 1996. The influence of UV-B radiation on the physicochemical nature of tobacco (Nicotiana tabacumL.) leaf surfaces. J. Exp. Bot. 47, 99 – 109.
Barnes, P.W., Flint, S.D., Caldwell, M.M., 1990. Morphologi-cal responses of crop and weed species of different growth forms to Ultraviolet-B radiation. Am. J. Bot. 77, 1354 – 1360.
Barnes, P.W., Maggard, S., Holman, S.R., Vergara, B., 1993. Intraspecific variation in sensitivity to UV-B radiation in rice. Crop Sci. 33, 1041 – 1046.
Barsig, M., Gehrke, C., Schneider, K., 1998. Effects of UV-B radiation on fine structure, carbohydrates and pigments in
Polytrichum commune. Bryologist 101, 357 – 365.
Beutler, H.O., 1985a. Starch. In: Bergmayer, H.U. (Ed.), Methods of Enzymatic Analysis, vol. VI, Metabolites 1: Carbohydrates. Verlag Chemie, Weinheim/Bergstr, pp. 1 – 10.
Beutler, H.O., 1985b.D-Fructose. In: Bergmeyer, H.U. (Ed.), Methods of Enzymatic Analysis VI, Metabolites 1: Carbo-hydrates. Verlag Chemie, Weinheim/Bergstr., pp. 321 – 327. Boehringer, 1989. Methoden der enzymatischen
Lebensmittel-analytik mit Einzelreagentien. Boehringer, Mannheim. Bongard-Pierce, D.K., Evans, M.M., Poethig, R.S., 1996.
Het-eroblastic features of leaf anatomy in maize and their genetic regulation. Int. J. Plant Sci. 157, 331 – 340. Bornman, J.F., Evert, R.F., Mierzwa, R.J., 1983. The effect of
Bosabalidis, A.M., Evert, R.F., Russin, W.A., 1994. Ontogeny of the vascular bundles and contiguous tissues in the maize leaf blade. Am. J. Bot. 81, 745 – 752.
Britz, S.J., Adamse, P., 1994. UV-B-induced increase in spe-cific leaf weight of cucumber as a consequence of increased starch content. Photochem. Photobiol. 60, 116 – 119. Caldwell, M.M., Flint, S.D., 1994. Stratospheric ozone
reduc-tion, solar UV-B radiation and terrestrial ecosystems. Clim. Change 28, 375 – 394.
Do¨hring, T., Ko¨fferlein, M., Thiel, S., Seidlitz, H.K., 1996. Spectral shaping of artificial UV-B irradiation for vegeta-tion stress research. J. Plant Physiol. 148, 115 – 119. Evert, R.F., Russin, W.A., Bosabalidis, A.M., 1996.
Anatomi-cal and ultrastructural changes associated with sink-to-source transition in developing maize leaves. Int. J. Plant Sci. 157, 247 – 261.
Garrard, L.A., Van, T.K., West, S.H., 1977. Plant response to middle ultraviolet (UV-B) radiation: carbohydrate levels and chloroplast reactions. Crop Sci. 36, 184 – 188. Gehrke, C. 1999. Personal Communication.
Holm-Hansen, O., Lubin, D., Helbling, E.W., 1993. Ultravio-let radiation and its effects on organisms in aquatic envi-ronments. In: Young, A.R., Bjo¨rn, L.O., Moan, J., Nultsch, W. (Eds.), Environmental UV Photobiology. Plenum, New York, pp. 379 – 425.
Huttunen, S., 1994. Effects of air pollutants on epicuticular wax structure. In: Percy, K.E., Cape, J.N., Jagels, R., Simpson, C.J. (Eds.), Air Pollutants and the Leaf Cuticle. Springer, Berlin, pp. 81 – 96.
Lichtenthaler, H.K., Wellburn, A.R., 1983. Determinations of total carotenoids and chlorophyllsaandbof leaf extracts in different solvents. Biochem. Soc. Trans. 11, 591 – 592. Lu¨tz, C., Seidlitz, H.K., Meindl, U., 1997. Physiological and
structural changes in the chloroplast of the green alga
Micrasterias denticulata induced by UV-B simulation. Plant Ecol. 128, 54 – 64.
Mark, U., Tevini, M., 1996. Combination effects of UV-B radiation and temperature on sunflower (Helianthus annuus
L., cv. Polstar) and maize (Zea mays L., cv. Zenit 2000) seedlings. J. Plant Physiol. 148, 49 – 56.
Meindl, U., Lu¨tz, C., 1996. Effects of UV irradiation on cell development and ultrastructure of the green algaMicraste
-rias. J. Photochem. Photobiol. B Biol. 36, 285 – 292. Russell, S.H., Evert, R.F., 1985. Leaf vasculature inZea mays
L. Planta 164, 448 – 458.
Russin, W.A., Evert, R.F., Vanderveer, P.J., Sharkey, T.D., Briggs, S.P., 1996. Modification of a specific class of plasmodesmata and loss of sucrose export ability in the sucrose export defective maize mutant. Plant Cell 8, 645 – 658.
Santos, I., Almeida, J.M., Salema, R., 1993. Plants of Zea mays L. developed under enhanced UV-B radiation. J. Plant Physiol. 141, 450 – 456.
Stapleton, A.E., Thornber, C.S., Walbot, V., 1997. UV-B component of sunlight causes measurable damage in field-grown maize (Zea mays L.): developmental and cellular heterogeneity of damage and repair. Plant Cell Environ. 20, 279 – 290.
Steinmu¨ller, D., Tevini, M., 1985. UV-B-induced effects upon the cuticular waxes of cucumber, bean, and barley leaves. In: Worrest, R.C., Caldwell, M.M. (Eds.), Stratospheric Ozone Reduction. Solar Ultraviolet Radiation and Plant Life. Springer, Berlin, pp. 271 – 285.
Teramura, A.H., 1983. Effects of ultraviolet-B radiation on the growth and yield of crop plants. Physiol. Plant. 58, 415 – 427.
Teramura, A.H., 1990. Implications of stratospheric ozone depletion upon plant production. Hortic. Sci. 25, 1557 – 1560.
Teramura, A.H., Murali, N.S., 1986. Intraspecific differences in growth and yield of soybean exposed to ultraviolet-B radiation under greenhouse and field conditions. Environ. Exp. Bot. 26, 89 – 95.
Tevini, M., 1993. Effects of UV-B radiation on terrestrial plants. In: Tevini, M. (Ed.), UV-B Radiation and Ozone Depletion: Effects on Humans, Animals, Plants, Microor-ganisms and Materials. Lewis, Boca Raton, FL, pp. 125 – 153.
Tevini, M., 1996. Erho¨hte UV-B-Strahlung: Ein Risiko fu¨r Nutzpflanzen? Biol. unserer Zeit 26, 245 – 254.
Tevini, M., Steinmu¨ller, D., Iwanzik, W., 1986. U8ber die Wirkung erho¨hter UV-B-Strahlung in Kombination mit anderen Stressfaktoren auf das Wachstum und Funktion von Nutzpflanzen. BPT-Bericht, Gesellschaft fu¨r Strahlen-und Umweltforschung, Munich.
Tevini, M., Mark, U., Fieser, G., Saile, M., 1990. Effects of solar and artificial UV-B radiation on growth, function and composition of crop plant seedlings. BPT-Final report. GSF, Munich.
Tevini, M., Braun, J., Fieser, G., 1991a. The protective func-tion of the epidermal layer of rye seedlings against ultravi-olet-B radiation. Photochem. Photobiol. 53, 329 – 333. Tevini, M., Mark, U., Saile-Mark, M., 1991b. Effects of
enhanced solar UV-B radiation on growth and function of crop plant seedlings. Plant Biochem. Physiol. 10, 13 – 31. Thiel, S., Do¨hring, T., Ko¨fferlein, M., Kosak, A., Martin, P.,
Seidlitz, H.K., 1996. A phytotron for plant stress research: How far can artificial lighting compare to natural sunlight? J. Plant Physiol. 148, 456 – 463.
Yue, M., Li, Y., Wang, X., 1998. Effects of enhanced ultravi-olet-B radiation on plant nutrients and decomposition of spring wheat under field conditions. Environ. Exp. Bot. 40, 187 – 196.
Ziska, L.H., Teramura, A.H., Sullivan, J.H., 1992. Physiologi-cal sensitivity of plants along an elevational gradient to UV-B radiation. Am. J. Bot. 79, 863 – 871.