Journal of Experimental Marine Biology and Ecology 252 (2000) 181–198
www.elsevier.nl / locate / jembe
Acute and acclimated digestive responses of the cockle
Cerastoderma edule (L.) to changes in food quality and
quantity
I. Feeding and absorption of biochemical components
*
I. Ibarrola , E. Navarro, M.B. Urrutia
´ ´ ´
Departamento de Biologıa Animal y Genetica, Facultad de Ciencias, Universidad del Paıs Vasco /Euskal
Herriko Unibertsitatea, Apartado 644, E-48080 Bilbao, Spain
Received 15 February 2000; received in revised form 14 March 2000; accepted 31 May 2000
Abstract
3 21
Cockles Cerastoderma edule were fed two different concentrations (| 0.8 and 2 mm l ) of
two diets with different qualities (|10 and 60% of organic content) which were achieved by
mixing different proportions of ashed silt particles with cells of the microalgae Tetraselmis
suecica. Clearance, ingestion and absorption rates of organic matter and biochemical components
were measured after 3 days (acute response) and 11 days (acclimated response) of exposure to the diets. With low quality diets cockles were found to reject part of the filtered matter (|25–35%)
through pseudofaeces production both in the acute and acclimated responses. In the acute response, absorption rate of organic matter was positively dependent on food quality and quantity, but the physiological response to increasing food concentrations differed with food quality: with low qualities, increasing absorption rate resulted from the simultaneous increase of clearance (|2
times) and ingestion rate (|4 times) as well as absorption efficiency of organic matter (|22%).
However, those fed high qualities, were found to compensate increasing food concentration by reducing (|50%) clearance rate. The resulting moderate increase of ingestion rate (|1.6 times)
was accompanied with a reduction in absorption efficiency (|20%). Irrespective of food quality
and quantity, protein and lipids were absorbed, respectively, with the highest (from 61.7 to 80.0%) and the lowest (from 42.6 to 66.8%) efficiency. Acclimated response was entirely affected by food quality: with low qualities, cockles greatly improved the energetic intake from available ration (|4 and 2 times, with low and high food concentrations, respectively). Both preingestive and
digestive mechanisms were involved in this response: at the preingestive level, clearance rate and preingestive selection efficiency were significantly increased. At the digestive level, cockles were capable of maintaining absorption efficiency of organic matter with rising ingestion rate. On the contrary, acclimation to high quality diets brought about no significant increase in organic
*Corresponding author. Tel.:134-94-601-2728.
E-mail address: [email protected] (I. Ibarrola).
absorption rate: with low ration, clearance rate was kept constant, whereas with high ration the increase in clearance and ingestion rate (|2 times) promoted a compensatory reduction in
absorption efficiency. However, the biochemical composition of the absorbed matter was found to be absolutely modified, both at low and high food rations, due to an strong reduction of lipid absorption efficiency. The observed modifications of absorption rate and / or the biochemical composition of the absorbed matter suggests the capability of cockles to adjust the digestive performance. 2000 Elsevier Science B.V. All rights reserved.
Keywords: Absorption; Acclimation; Bivalves; Cockles; Feeding
1. Introduction
It has been generally recognized that the ability to regulate rates of food acquisition and digestion constitutes an important factor allowing marine bivalves to inhabit highly variable trophic environments such as intertidal zones. The necessity for a regulatory control of the feeding activity in bivalves stems from the existence of a trade-off between absorption efficiency and ingestion rate which is determined by two fundamen-tal digestive relationship: (i) absorption efficiency is a positive function of food retention time (Bayne et al., 1987; Navarro et al., 1994) which is the time available for the digestive structures to hydrolyse and absorb organic matter, and (ii) food retention time is inversely related with ingestion rate (Bayne et al., 1987, 1989; Navarro and Iglesias, 1993; Navarro et al., 1994). Two conclusions emerge from above assumptions: the first one is that the function of ingestion rate regulation in bivalves is to provide for an optimal gut retention time allowing maximum rates of absorption; and the second one is that optimal retention times, and hence, feeding behaviour would be affected by possible changes in the digestive potential of the digestive structures. Although there is a general agreement regarding the first point, the second one remains poorly investigated despite of great amount of early studies concerning enzymatical (see Vonk and Western, 1984 for review) and histological (see Morton, 1983 for review) aspects of digestion.
Below the threshold for pseudofaeces production, Ibarrola et al. (1996, 1998) described two fundamental relationships between short-term digestive performance and diet characteristics in cockles Cerastoderma edule submitted to different food qualities
3
and quantities: (a) gut content (the amount of food held in the gut: mm ) increases with rising ingestion rate of low quality food, whereas is kept constant with high qualities which can lead to different relationships between absorption efficiency and ingestion rate for foods of different qualities. (b) Carbohydrase activity in the digestive gland positively depends on food organic content (Ibarrola et al., 1996, 1998), which can lead to increased absorption of carbohydrates at the expenses of lipids in cockles fed on high quality diets.
I. Ibarrola et al. / J. Exp. Mar. Biol. Ecol. 252 (2000) 181 –198 183
biochemical composition of the absorptive balance: net absorption efficiencies of lipids is specifically reduced due to the high lipidic fraction of the endogenous faecal losses (Ibarrola et al., 2000).
Therefore, we have shown that the enzyme activity released by the digestive gland is under physiological control. The aim of present study was (i) to determine the effects elicited by food quantity and quality on food ingestion and digestive investments at different stages of diet acclimation (discussed in this paper), and (ii) to identify the digestive structures involved in the physiological response at the level of the digestive gland (discussed in the companion paper). Physiological and enzymatic measurements were combined with a cytohistological study of the digestive gland of cockles submitted to four different food conditions (two qualities3two rations). To study the time-course of responses, these measurements were performed both after 3 days (short-term response) and 11 days (acclimated response) of exposure to the diets.
2. Material and methods
2.1. Experimental set-up and manipulation of cockles
A total of 140 cockles (ranging in size 24–28 mm) were collected from an intertidal mud-flat in the Mundaka estuary (Biscay, N. Spain) during February 1993. In the laboratory, 120 cockles were divided in four groups, each fed, for approximately 12 days, one of the four diets described in the experimental protocol, and the remaining 20 cockles were submitted to starvation. Individuals were glued to glass slides by the exterior shell surface with the exhalant siphons pointing downwards before being placed in individual chambers which were immersed in sea water containing tanks. Temperature was ambient (t5188C). After 3 and 11 days of exposure to the diets, samples of food suspensions and faeces produced by individuals were collected to compute the rates of food acquisition and absorption of both organic matter and biochemical components. Furthermore, the digestive gland of 15 cockles was dissected out to determine digestive enzyme activities and histological parameters (discussed in the companion paper).
2.2. Elaboration of experimental diets
Diets were prepared by mixing cells of the microalgae Tetraselmis suecica with ashed silt particles (,45mm). Two different qualities (L: low, and H: high) were achieved by mixing algae and silt in different proportions based as particle volume: 10% algae: 90% silt for low quality diets and 90% algae: 10% silt for high quality diets. Concentrated stocks of both food qualities were dosed at one of two particle concentrations (l: low
3
2.3. Characteristics of diets
On two occasions (i.e. during the 3rd and 11th day |60–70 and 240–260 h,
respectively, after the start of the exposure to diets), all faeces produced by each individual during a time period (|12 h: biodeposition time) and four samples of food
suspensions were collected and processed to calculate physiological measurements following the procedure described below (see Section 2.4). Total particulate matter
21
(TPM: mg l ) in food suspensions was determined by filtering known volumes of suspensions onto preweighed and ashed Whatman GF / C filters and reweighing them after rinsing with ammonium formate isotonic with sea water and drying at 1108C.
21
Particulate inorganic matter (PIM: mg l ) was given as the weight remaining after ignition at 4508C during 6 h in a muffle furnace. Particulate organic matter (POM: mg
21
l ) was calculated as the difference TPM2PIM. Food organic content ( f ) was calculated as f5POM / TPM. Food concentration (VOL) varied slightly over the biodeposition time; thus, in order to estimate the average values of TPM, POM and PIM we used the following procedure: (i) departing from the continuous determination of particle concentration with the Coulter Multisizer, an average food concentration (VOL) was estimated for each four conditions. (ii) Departing from suspension samples, regression lines relating TPM, POM and PIM with VOL were separately established for each four condition. (iii) Thus, from the calculated average food concentration (VOL) and regression lines, average values of TPM, POM and PIM were estimated for each four experimental condition. These results are given in Table 1. No variance could thus be assigned to such estimations of diet characteristics.
2.4. Physiological measurements
Total and inorganic weights of the biodeposition were obtained by filtering aliquots of them onto preweighed ashed Whatman GF / C filters and processing as indicated for
21
suspension samples. Following this procedure, egestion rates (mg h ) of total, inorganic and organic matter were measured (Iglesias et al., 1998). Since absorption of inorganic
Table 1
a
Diet characteristics compared between days 3 and 11
Diet Day q VOL f POM PIM TPM
Ll 3 0.248 0.772 9.9 0.192 1.724 1.916
11 0.248 0.773 10.0 0.192 1.727 1.918
Lh 3 0.160 1.999 7.8 0.320 4.043 4.336
11 0.155 2.187 7.2 0.339 4.337 4.706
Hl 3 0.574 0.659 57.9 0.379 0.275 0.654
11 0.573 0.673 58.1 0.385 0.278 0.663
Hh 3 0.510 2.573 66.1 1.313 0.673 1.986
11 0.518 1.896 64.9 0.982 0.532 1.514
a 23 3 21
q, food quality (mg POM mm ); VOL, particle packed volume per volume of suspension (mm l ); f, 21
food organic content (%); POM, particulate organic matter (mg l ); PIM, particulate inorganic matter (mg
21 21
I. Ibarrola et al. / J. Exp. Mar. Biol. Ecol. 252 (2000) 181 –198 185
matter in the digestive system of bivalves is negligible, inorganic egestion rate equals inorganic ingestion rate which, in the absence of pseudofaeces production, represents
21 21
inorganic filtration rate (mg h ). Clearance rate (CR: l h ) was computed dividing inorganic filtration rate by particulate inorganic matter in the food suspension (PIM).
21 21
Ingestion rate of total (IR: mg h ) and organic matter (OIR: mg h ) were then calculated as CR3TPM, and CR3POM, respectively. Finally, absorption rate (AR: mg
21
h ) was obtained as the difference between ingestion and egestion rates of organic matter, and absorption efficiency was calculated as AR / OIR.
With low quality diets cockles produced pseudofaeces. Separate collection and processing of these biodeposits allowed computation of rejection rate of particulate
21
matter (RR: mg h ). In these cases, inorganic filtration rate was calculated as the sum of rejected and egested inorganic particulate matter. Ingestion rate of organic matter (OIR) was obtained as the difference between filtration and rejection rates of organic matter and allowed computation of absorption rate and absorption efficiency as indicated above. Since pseudofaeces production in bivalves comprises preferential rejection of inorganic particles, the organic content of the ingested matter (i ) is altered as compared with food organic content ( f ). Organic content of the ingested matter was estimated as OIR / IR. The efficiency of the preingestive selection process (SE) was estimated as SE512( p /f ), where p represents the organic content of pseudofaeces.
2.5. Absorption of biochemical components
Subsamples of faeces collected for determination of individual feeding rates as well as samples of concentrated stocks of diet, were filtered onto ashed Whatman GF / C. A modification of a micro-analytical fractionated extraction scheme developed by Holland and Gabbot (1971) was followed for determination of the contents of biochemical components in the faeces and diets.
Filtered samples were homogenised in a glass tissue grinder with 4 ml of distilled water. Lipid content was established in a 2-ml aliquot following Marsh and Weinstein (1966), after extraction of lipids with mixtures of chloroform–methanol–water accord-ing to Bligh and Dyer (1959). Another aliquot of 2 ml was used to analyze carbohydrate and protein contents. Extraction of carbohydrates was carried out by incubating (30 min) in an ultrasonic bath and boiling the extracts (20 min) with 5% TCA. After centrifuga-tion (4000 rpm, 10 min), the supernatant was separated and the remaining pellet was subjected to further extraction (this was repeated twice). The carbohydrate content in the combined supernatants was determined following Dubois et al. (1956) using glycogen as standard. The pellet was resuspended in 1 ml of 0.5 N NaOH and incubated in an ultrasonic bath for 30 min. Protein content was then determined according to Lowry et al. (1951).
2.5.1. Ingestion rates
Ingestion rates of biochemical components (IR , IRP C and IR for proteins, carbohy-L 21
2.5.2. Absorption rates and efficiencies
21 Absorption rates (AR , AR and AR for proteins, carbohydrates and lipids: mg hP C L ) and efficiencies (AE , AEP C and AE : %) of different biochemical components wereL calculated, after computation of egestion rates of biochemical components (as the product of organic egestion rate and the proportion of the components in the faeces), as the difference between their ingestion and egestion rates. Finally, absorption efficiency of biochemical components was calculated as the ratio of absorption to ingestion.
2.6. Statistical procedures
Significant effects promoted by food quality ( ql ) and quantity ( qn) on physiological measurements and absorption of biochemical components were analyzed by performing two-factor analysis of variance (Zar, 1984). The effect promoted by acclimation to the different diets was analyzed by testing for significant differences (t-test) between the mean values obtained for the acute and acclimated responses.
3. Results
3.1. Characteristics of the diets
Average food characteristics for all experimental conditions were maintained between days 3 and 11 (Table 1), with the exception of Hh diet for which a reduction of 21% in food concentration (TPM, POM and PIM) was obtained in day 11 in comparison with the third day.
3.2. Physiological measurements
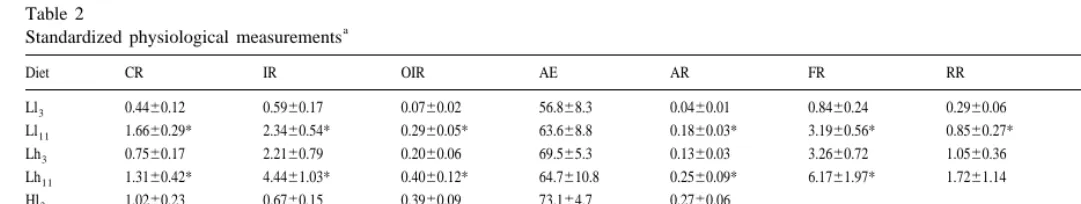
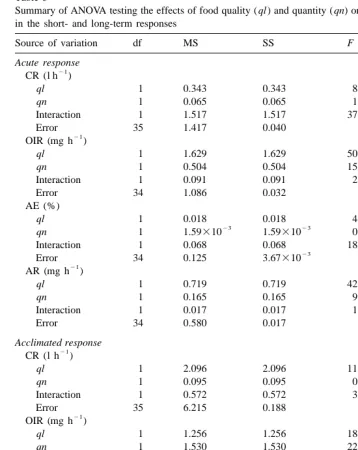
Both in the acute and acclimated responses, cockles fed low quality diets rejected part of the filtered material by means of pseudofaeces production (Table 2). Thus, the table also shows the values of rates of food filtration (FR) and rejection (RR), preingestive selection efficiency (SE) and organic content of ingested matter (i ) obtained for cockles fed low quality diets. Significant differences exerted by food quality ( ql ) and food quantity ( qn) on physiological measurements in the acute and acclimated responses were determined by performing three factor variance analysis (ANOVA) shown in Table 3.
3.2.1. Acute response
I.
Ibarrola
et
al
.
/
J.
Exp
.
Mar
.
Biol
.
Ecol
.
252
(2000
)
181
–
198
187
Table 2
a
Standardized physiological measurements
Diet CR IR OIR AE AR FR RR S.E. i
Ll3 0.4460.12 0.5960.17 0.0760.02 56.868.3 0.0460.01 0.8460.24 0.2960.06 0.4060.09 11.960.9 Ll11 1.6660.29* 2.3460.54* 0.2960.05* 63.668.8 0.1860.03* 3.1960.56* 0.8560.27* 0.5960.09* 12.361.2 Lh3 0.7560.17 2.2160.79 0.2060.06 69.565.3 0.1360.03 3.2660.72 1.0560.36 0.4860.10 10.461.8 Lh11 1.3160.42* 4.4461.03* 0.4060.12* 64.7610.8 0.2560.09* 6.1761.97* 1.7261.14 0.6260.09* 9.862.1 Hl3 1.0260.23 0.6760.15 0.3960.09 73.164.7 0.2760.06
Hl11 0.9360.53 0.6260.35 0.3660.21 64.669.9 0.2360.18 Hh3 0.5460.25 1.0860.50 0.7160.33 58.869.9 0.4560.24 Hh11 0.9760.28* 1.6460.68 0.9560.27 51.165.6* 0.5260.22
a 21 21 21
CR, clearance rate (l h ); IR, ingestion rate of TPM (mg h ); OIR, ingestion rate of POM (mg h ); AE, absorption efficiency (%); AR, absorption rate (mg
21 21 21
Table 3
Summary of ANOVA testing the effects of food quality ( ql ) and quantity ( qn) on physiological measurements in the short- and long-term responses
Source of variation df MS SS F P
Acute response 21
CR (l h )
ql 1 0.343 0.343 8.459 0.0063**
qn 1 0.065 0.065 1.597 0.2147
Interaction 1 1.517 1.517 37.410 0.0001***
Error 35 1.417 0.040
21
OIR (mg h )
ql 1 1.629 1.629 50.381 0.0001***
qn 1 0.504 0.504 15.772 0.0004***
Interaction 1 0.091 0.091 2.859 0.1000
Error 34 1.086 0.032
AE (%)
ql 1 0.018 0.018 4.834 0.0348*
23 23
qn 1 1.59310 1.59310 0.433 0.5148
Interaction 1 0.068 0.068 18.449 0.0001***
23
Error 34 0.125 3.67310
21
AR (mg h )
ql 1 0.719 0.719 42.142 0.0001***
qn 1 0.165 0.165 9.679 0.0038**
Interaction 1 0.017 0.017 1.021 0.3194
Error 34 0.580 0.017
Acclimated response 21
CR (l h )
ql 1 2.096 2.096 11.129 0.0021**
qn 1 0.095 0.095 0.503 0.4830
Interaction 1 0.572 0.572 3.038 0.0903
Error 35 6.215 0.188
21
OIR (mg h )
ql 1 1.256 1.256 18.452 0.0001***
qn 1 1.530 1.530 22.468 0.0001***
Interaction 1 0.810 0.810 11.894 0.0016**
Error 33 2.247 0.068
AE (%)
ql 1 0.038 0.038 4.495 0.0416*
qn 1 0.072 0.072 8.500 0.0063**
Interaction 1 0.028 0.028 3.302 0.0783
23
Error 33 0.278 8.42310
21
AR (mg h )
ql 1 0.248 0.248 10.679 0.0025**
qn 1 0.287 0.287 12.389 0.0013**
Interaction 1 0.118 0.118 5.072 0.0311*
Error 33 0.765 0.023
I. Ibarrola et al. / J. Exp. Mar. Biol. Ecol. 252 (2000) 181 –198 189
content of ingested matter was enhanced 20 and 23% (with Ll and Lh , respectively)3 3 with respect to food organic content.
In contrast with cockles fed low quality diets, those fed high qualities reduced (t-test,
P50.0002) clearance rate (|47%) with increasing particle concentration (Hh vs. Hl ).3 3 This reduction of CR was not as high as to preclude the increment of ingestion rate which was found to be approximately two times higher for cockles fed Hh than those3 fed Hl diet. Therefore, in spite of existence of different trends in CR, increasing food3 ration brought about enhanced ingestion rates irrespective of food quality, thus, ingestion rate of organics was positively dependent on both food quantity and food quality (Table 3).
3.2.1.2. Absorption rate and efficiency of organic matter. The increment of ingestion rate brought about by increasing food concentrations exerted differential effects on absorption efficiency of cockles fed different food qualities: whereas with low quality diets AE was significantly increased (from 56.868.3 to 69.565.3%, t-test, P50.0131). With high quality diets, AE became significantly reduced (from 73.164.7 to 58.869.9%, t-test, P50.0008). This reduction was not intense enough as to preclude significantly higher absorption rates in Hh as compared to Hl diet. Thus, in spite of the3 3 differential relationship between AE and OIR, absorption rate was increased with rising food rations irrespective of food quality.
3.2.2. Acclimated response
3.2.2.1. Food acquisition. Acclimation to low food quality brought about a signifi-cant increment of clearance, filtration and rejection rates (Table 2). The increment on clearance rate with low quality diets was strong enough (33.8 and 1.7 for Ll and Lh, respectively) to yield significantly higher values than those of cockles fed on high quality diets (ANOVA on Table 3). Since cockles acclimated to the low ration of low food quality (Ll ) increased clearance rate more than those acclimated to the high ration11 (Lh ), the difference in ingestion rate between these two groups were partially reduced11 after acclimation (Lh / Ll3 350.203 / 0.071 vs. Lh11/ Ll1150.399 / 0.288). In addition, and irrespective of food quantity, the proportion of filtered matter rejected as pseudofaeces was reduced (from 34 to 26%) and the selection efficiency increased (from 0.40 and 0.48 for Ll and Lh to 0.59 and 0.61 for Ll3 3 11 and Lh ) significantly after acclimation.11 Conversely, with high food qualities only cockles fed on the concentrated ration (Hh) enhanced clearance rate (31.8), whereas those acclimated to Hl did not modify clearance rate. In consequence, the difference in OIR between cockles fed high and low
21
quantities increased after acclimation (Hh / Hl3 350.715 / 0.387 mg h vs. Hh11/ Hl115
0.950 / 0.361).
enhanced clearance rate during acclimation (Hl), and was significantly reduced in those having increased rates of clearance (Hh). This reduction of AE compensated for the
21
slight (from 0.71560.331 to 0.95060.272 mg h ), though not significant (t-test,
P50.0617), enhancement of OIR promoted by the increase of CR during acclimation to Hh diet. Thus, absorption rates for cockles fed high quality diets were not significantly affected by acclimation to either low or high food concentrations.
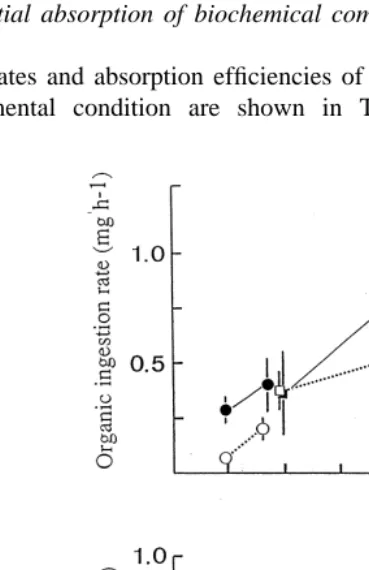
In Fig. 1, rates of ingestion of organic matter and rates of absorption have been plotted against POM concentration. Due to the intense increase of AR in cockles acclimated to low quality diets and lack of it in those acclimated to high quality diets, there seems to be a linear relationship between AR and POM irrespective of food quality.
3.3. Differential absorption of biochemical components
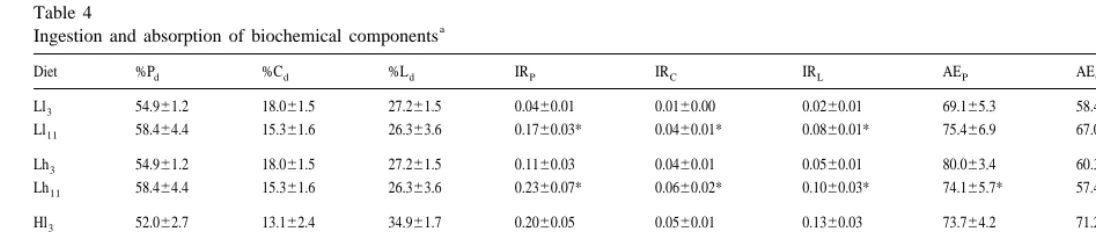
Ingestion rates and absorption efficiencies of biochemical components recorded with each experimental condition are shown in Table 4. The table also contains the
21 21
I.
Ibarrola
et
al
.
/
J.
Exp
.
Mar
.
Biol
.
Ecol
.
252
(2000
)
181
–
198
191
Table 4
a
Ingestion and absorption of biochemical components
Diet %Pd %Cd %Ld IRP IRC IRL AEP AEC AEL
Ll3 54.961.2 18.061.5 27.261.5 0.0460.01 0.0160.00 0.0260.01 69.165.3 58.469.8 45.169.3 Ll11 58.464.4 15.361.6 26.363.6 0.1760.03* 0.0460.01* 0.0860.01* 75.466.9 67.065.7* 42.4617.0
Lh3 54.961.2 18.061.5 27.261.5 0.1160.03 0.0460.01 0.0560.01 80.063.4 60.365.6 42.669.8 Lh11 58.464.4 15.361.6 26.363.6 0.2360.07* 0.0660.02* 0.1060.03* 74.165.7* 57.466.6 19.6616.8*
Hl3 52.062.7 13.162.4 34.961.7 0.2060.05 0.0560.01 0.1360.03 73.764.2 71.265.7 66.865.2 Hl11 69.761.6 8.860.7 21.561.3 0.2560.14 0.0360.02* 0.0860.04* 79.768.7 50.0611.3* 11.2626.6*
Hh3 52.062.7 13.162.4 34.961.7 0.3760.17 0.0960.04 0.2560.12 61.766.9 59.068.9 61.269.0 Hh11 69.761.6 8.860.7 21.561.3 0.6660.19* 0.0860.02 0.2060.06 62.863.8 48.464.2* 6.8613.6*
a 21
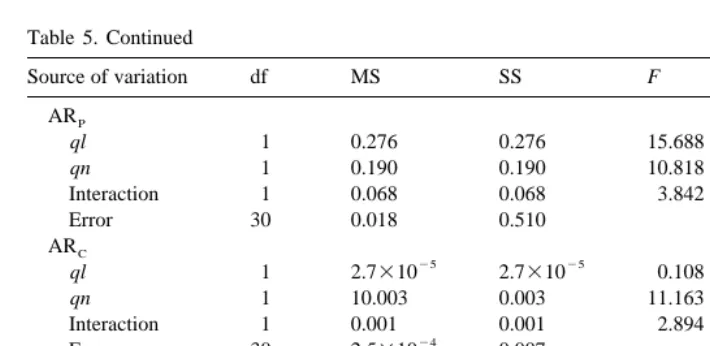
biochemical composition of the diet suspensions. In all feeding conditions protein and lipids were absorbed, respectively, with the highest and the lowest efficiency (Hh being3 the exception, Table 4), with greatest significant differences following acclimation. Resulting absorption rates of biochemical components are plotted in Fig. 2. Table 5 shows the results of the two factor variance analyses, food quality ( ql ) and quantity ( qn), on absorption efficiencies and rates of biochemical components.
3.3.1. Acute response
Both food quality and the interaction term ( ql3qn) exerted significant effects on
absorption efficiencies of protein and carbohydrates (Table 5). In other words, absorption efficiency of these two components presented a similar tendency of absorption efficiency for total organics: they are increased with rising rations of low quality food (from 69.165.3 to 80.063.4%, t-test P50.0025; and from 58.469.8 to 60.365.6%, t-test P50.3225 (not significant) for P and C, respectively) and sig-nificantly reduced with increasing concentrations of high quality (from 73.764.2 to 61.766.9%, t-test P50.0001; and from 71.265.7 to 59.068.9% t-test P50.0009, for P and C, respectively). These reductions were not intense enough as to preclude significantly higher absorption rates of proteins and carbohydrates in Hh as compared3 to Hl . Conversely, absorption of lipids followed a different trend: AE was increased3 L with increasing quality and decreased with increasing food quantities (Table 4).
3.3.2. Acclimated response
Acclimation brought about different effects on the absorption efficiencies of the different biochemical components. With regard to proteins, the high values of absorption efficiency recorded in the acute response were maintained (with the exception of cockles acclimated to Lh for which an slight but significant reduction, from 80.063.4 to 74.165.7%, Table 4, was recorded). This is especially remarkable for cockles accli-mated to low qualities which significantly increased ingestion rate, as well as for those acclimated to Hh which suffered a significant reduction of absorption efficiency of organic matter. Thus, absorption rate of proteins followed a similar pattern to that described for ingestion, and absorption rates of organic matter were positively dependent on food quality ( ql ) and quantity ( qn) (Table 5).
Conversely, carbohydrate and lipid absorption efficiencies were significantly modified during acclimation. An overall decrease in AEL was recorded. With low quality diets, this reduction tends to compensate, in terms of absorption rate, the increased ingestion rate of lipid achieved after acclimation; however, with high qualities, AEL reduction promoted significant reductions in ARL with even negative absorption rates being recorded, a result that indicates that some individuals were loosing endogenous lipids through the faeces.
I. Ibarrola et al. / J. Exp. Mar. Biol. Ecol. 252 (2000) 181 –198 193
21
Table 5
Summary of ANOVA testing the effects of food quality ( ql ) and quantity ( qn) on the absorption efficiency of the different biochemical components
Source of variation df MS SS F P
Short-term
AEP
ql 1 0.038 0.038 14.128 0.0007***
24 24
qn 1 2.61310 2.61310 0.097 0.7578
Interaction 1 0.105 0.105 38.985 0.0001***
23
Error 30 0.081 2.70310
AEC
ql 1 0.027 0.027 4.778 0.0368*
qn 1 0.022 0.022 3.769 0.0617
Interaction 1 0.041 0.041 7.165 0.0119*
23
Error 30 0.171 5.71310
AEL
ql 1 0.331 0.331 47.669 0.0001***
qn 1 0.013 0.013 1.882 0.1802
23 23
Interaction 1 1.95310 1.95310 0.283 0.5990
23
Error 30 0.208 6.93310
ARP
ql 1 0.150 0.150 26.715 0.0001***
qn 1 0.044 0.044 7.762 0.0092**
Interaction 1 0.002 0.002 0.372 0.5457
Error 30 0.008 0.168
ARC
ql 1 0.008 0.008 25.559 0.0001***
qn 1 0.002 0.002 6.922 0.0133*
24 24
Interaction 1 1.0310 1.0310 0.333 0.5682
24
Error 30 3.7310 0.010
ARL
ql 1 0.093 0.093 42.096 0.0001***
qn 1 0.013 0.013 5.698 0.0235*
Interaction 1 0.006 0.006 2.554 0.1205
Error 30 0.002 0.066
Long-term
AEP
ql 1 0.010 0.010 2.407 0.1312
qn 1 0.068 0.068 16.126 0.0004***
Interaction 1 0.050 0.050 11.785 0.0018**
23
Error 30 0.126 4.20310
AEC
ql 1 0.138 0.138 26.596 0.0001***
qn 1 0.026 0.026 4.961 0.0336*
Interaction 1 0.013 0.013 2.492 0.1249
23
Error 30 0.156 5.18310
AEL
ql 1 0.392 0.392 10.826 0.0026**
qn 1 0.151 0.151 4.175 0.0499*
Interaction 1 0.069 0.069 1.911 0.1771
I. Ibarrola et al. / J. Exp. Mar. Biol. Ecol. 252 (2000) 181 –198 195 Table 5. Continued
Source of variation df MS SS F P
ARP
ql 1 0.276 0.276 15.688 0.0004***
qn 1 0.190 0.190 10.818 0.0026**
Interaction 1 0.068 0.068 3.842 0.0596
Error 30 0.018 0.510
ARC
25 25
ql 1 2.7310 2.7310 0.108 0.7450
qn 1 10.003 0.003 11.163 0.0023**
Interaction 1 0.001 0.001 2.894 0.0996
24
Error 30 2.5310 0.007
ARL
ql 1 0.002 0.002 2.210 0.1479
24 24
qn 1 1.2310 1.2310 0.172 0.6816
27 27
Interaction 1 8.0310 8.0310 0.001 0.9734
Error 30 0.001 0.020
* P,0.05. ** P,0.01. *** P,0.001.
be pointed out, however, that this effect could have resulted from the existence of lower carbohydrate contents in the microalgae used to prepare the diets on day 11 as compared to that used on day 3 (see Table 4).
4. Discussion
In the short-term, cockles were found to increase ingestion rate with rising rations of both low and high food qualities. However, ingestion rate increased by different means in cockles fed on different food qualities: with low qualities, clearance rate was increased, whereas with high qualities a partial regulation of ingestion rate was displayed by reducing clearance rate. Contrasting effects of increasing particle concentration on the clearance rate of cockles fed on different diets may be explained by different effects of ingestion rate (IR) on absorption efficiency (AE): Whereas a simultaneous enhancement of IR and AE occurs with increasing TPM of low qualities, with high qualities both factors were inversely related. In a previous experiment (Ibarrola et al., 1998), similar results were recorded for cockles fed on a similar combination of diets. In that occasion we found that the trend shown by the gut capacity (GC: the amount of food held in the
3
explained behaviour of gut content (i.e. increment from Ll to Lh and maintenance in high quality diets) as reported separately (see Part II: Enzymatic digestion, in this issue). After 11 days of acclimation, cockles significantly changed absorption rate of total organics and / or the biochemical composition of the absorbed material, suggesting that further modifications of the functional gut capacity took place. With low qualities acclimation brought about an outstanding increase of net absorption rates by factors of
|4 and 2 with Ll and Lh diets, respectively. Both preingestive and digestive mechanisms
were involved in this response: at the preingestive level, clearance rate (CR) and selection efficiency (SE) were significantly increased. One possibility is that SE enhancement could have resulted as a mere consequence of increasing filtration rate, which would allow a broader scope for labial palps to select among different food items. Alternatively it could be the result of a time-dependent optimization of the selective mechanisms. The possibility of this second hypothesis would need further research.
At the digestive level, maintenance of absorption efficiency in spite of the increase in ingestion rate suggests that cockles developed a higher digestive capacity during acclimation to low quality diets. As a result of this strategy, net absorption rates, per unit of particulate organic matter in the diet, of cockles acclimated to low qualities achieved similar values to that obtained in the short-term by cockles fed high qualities. Thus, cockles tends to compensated for the negative effects of reduced food quality on absorptive balance. This response brought about no clear changes in the biochemical composition of the absorbed material except with diet Lh, for which an intense (significant) drop AE of lipids was found.
Cockles acclimated to high quality diets (Hl and Hh) showed very different behaviour: no significant increments of absorption rates of organic matter were measured, but the biochemical composition of the absorbed material was entirely affected, protein absorption being largely increased at the expenses of lipid. In our previous experiments (Ibarrola et al., 1996, 1998) the reduction in the absorption efficiency of lipids was interpreted as a consequence of the intensification of digestive action and the high lipid proportion of metabolic faecal losses (Ibarrola et al., 2000). In these previous occasions, an increased energy income in the form of proteins and carbohydrates was achieved as a counterpart of the reduction in the lipid absorption. However, in the present experiments, a similar modification of the biochemical balance was uncoupled to enhancement of net absorption rate. This suggests that the control of digestive performance in Cerastoderma
edule might be specifically sensitive to the absorptive balances of proteins and / or
carbohydrates as much as to the energy balance itself.
I. Ibarrola et al. / J. Exp. Mar. Biol. Ecol. 252 (2000) 181 –198 197
1996; protein and carbohydrates: Kreeger et al., 1995, 1996); (ii) in most of the studies, individuals were submitted to a single feeding condition, so that a range of variation of absorption efficiencies associated to the changes in food characteristics was not provided and, (iii) in several cases, bivalves were fed artificial diets (microcapsules) that are absorbed with low efficiencies (Kreeger, 1993, Kreeger et al., 1995, 1996). However, from the studies of Kreeger a preference of protein over carbohydrate absorption seems to occur in winter in mussels. This has been suggested to be linked to the high protein demands of gametogenesis, a process that normally takes place in that season in the mussel (reviewed by Newell and Bayne, 1980). Since the present experiment was performed in February, the results might reflect a similar behaviour in cockles to that found in mussels.
Alternatively, the status of metabolic reduction of cockles in the present experiment should be considered. In previous experiments using similar diets (both in composition
21 and concentration) recorded net absorption rates ranged from 0.44 to 0.9 mg h in June
21
(our unpublished results), and from 0.2 to 0.7 mg h in November (Ibarrola et al., 21
1998) that when compared with 0.05–0.45 mg h obtained in present study reveals a reduced capability of the digestive system of cockles for food digestion and absorption. A depleted digestive capability could induce modifications also in the biochemical balance because the net absorption efficiency of biochemical components is a function of (i) the type of enzymes participating in the digestive response (Ibarrola et al., 1996, 1998) and (ii) the rate of loss of endogenous biochemical components through the metabolic faecal losses (Ibarrola et al., 2000); both factors being entirely dependent of the digestive status of the animals. Further evidences of that come from the comparison of present results with those obtained by Navarro et al. (1994) in a similar experiment performed in spring. They found no variations in clearance and absorption from the 3rd to the 10th day of cockles exposed to various concentrations of natural sediment. Since the short-term response was reported to involve adjustments of gut content and throughput time, the absence of an acclimation-effect was interpreted as a fast completion (less than 3 days) of digestive adaptation. Evidence for long-term digestive acclimation recorded in the present experiment suggests the existence of a seasonally dependent, slower time-course of the digestive response.
The analysis of the histological and enzymatic parameters of the digestive gland will show the digestive mechanisms available for the acute and acclimated responses of cockles to the changes in the food quantity and quality. Whatever the mechanisms are, their characteristics should be consistent with (i) a preferential absorption of protein among the different biochemical components and (ii) the evidences for a time-dependent intensification of intracellular digestion, irrespective of food quality.
Acknowledgements
´
I. Ibarrola was supported by a grant from the Plan de Formacion del Personal ´
Investigador (Ministerio de Educacion y Ciencia, Spain). Part of this research was
´ ´
References
Bayne, B.L., Hawkins, A.J.S., Navarro, E., 1987. Feeding and digestion by the mussel Mytilus edulis L. (Bivalvia: Mollusca) in mixtures of silt and algal cells at low concentrations. J. Exp. Mar. Biol. Ecol. 111, 1–22.
Bayne, B.L., Hawkins, A.J.S., Navarro, E., Iglesias, J.I.P., 1989. Effects of seston concentrations on feeding, digestion and growth in the mussel Mytilus edulis. Mar. Ecol. Prog. 55, 47–54.
Bayne, B.L., Iglesias, J.I.P., Navarro, E., Heral, M., Deslous-Pauli, J.M., 1993. Feeding behaviour of the mussel Mytilus edulis L: responses in variations in both quantity and organic content of seston. J. Mar. Biol. Assoc. UK 73, 813–829.
Bligh, E.G., Dyer, W.J., 1959. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 37, 911–917.
Dubois, M., Gilles, K.A., Hamilton, J.K., Rebers, P.A., Smith, F., 1956. Colorimetric method for determination of sugars and related substances. Anal. Chem. 28, 350–356.
Holland, D.L., Gabbot, P.A., 1971. A micro-analytical scheme for the determination of protein, carbohydrate, lipid and RNA levels in marine invertebrate larvae. J. Mar. Biol. Assoc. UK 51, 659–668.
Ibarrola, I., Iglesias, J.I.P., Navarro, E., 1996. Differential absorption of biochemical components in the diet of the cockle Cerastoderma edule: enzymatic responses to variations in seston composition. Can. J. Zool. 74, 1887–1897.
Ibarrola, I., Navarro, E., Iglesias, J.I.P., 1998. Short-term adaptation of digestive processes in the cockle
Cerastoderma edule exposed to different food quantity and quality. J. Comp. Biochem. Physiol. B. 168,
32–40.
Ibarrola, I., Navarro, E., Iglesias, J.I.P., Urrutia, M.B., 1999. Time-course of digestive enzyme acclimation in the cockle Cerastoderma edule. Mar. Biol. 135, 47–56.
Ibarrola, I., Navarro, E., Iglesias, J.I.P., Urrutia, M.B., 2000. Temporal changes of feeding and absorption of biochemical components in Cerastoderma edule fed algal diet. J. Mar. Biol. Assoc. UK 80, 119–125. Iglesias, J.I.P., Urrutia, M.B., Navarro, E., Ibarrola, I., 1998. Measuring feeding and absorption in
suspension-feeding bivalves: an appraisal of the biodeposition method. J. Exp. Mar. Biol. Ecol. 219, 71–86. Kreeger, D.A., 1993. Seasonal patterns in the utilization of dietary protein by the mussel Mytilus trossulus.
Mar. Ecol. Prog. Ser. 95, 215–232.
Kreeger, D.A., Hawkins, A.J.S., Bayne, B.L., Lowe, D.M., 1995. Seasonal variation in the relative utilization of dietary protein for energy and biosynthesis by the mussel Mytilus edulis. Mar. Ecol. Prog. Ser. 126, 177–184.
Kreeger, D.A., Hawkins, A.J.S., Bayne, B.L., 1996. Use of dual-labeled microcapsules to discern the physiological fates of assimilated carbohydrate, protein carbon, and protein nitrogen in suspension-feeding organisms. Limnol. Oceanogr. 41 (2), 208–215.
Lowry, O.H., Rosebrough, N.J., Farr, A.L., Randall, R.J., 1951. Protein measurements with the Folin phenol reagent. J. Biol. Chem. 193, 265–275.
Marsh, J.B., Weinstein, D.B., 1966. Simple charring method for determination of lipids. J. Lipid Res. 7, 574–576.
Morton, B.S., 1983. Feeding and digestion in Bivalvia. In: Saleuddin, A.S.M., Wilbur, K.M. (Eds.), The Mollusca. Physiology, Part 2, Vol. V. Academic Press, New York, pp. 65–147.
Navarro, E., Iglesias, J.I.P., 1993. Infaunal filter-feeding bivalves and the physiological response to short-term fluctuations in food availability and composition. In: Dame, R.F. (Ed.), Bivalve Filter Feeders in Estuarine and Coastal Ecosystem Processes. NATO ASI Series, Vol. G 33. Springer, New York, pp. 25–56. Navarro, E., Iglesias, J.I.P., Ortega, M.M., Larretxea, X., 1994. The basis for a functional responses to variable
food quantity and quality in cockles Cerastoderma edule (Bivalvia, Cardiidae). Physiol. Zool. 67, 468–498. Newell, R.I.E., Bayne, B.L., 1980. Seasonal changes in the physiology, reproductive condition and carbohydrate content of the cockle Cardium (5Cerastoderma) edule (Bivalvia: Cardiidae). Mar. Biol. 56,
11–19.
Vonk, H.J., Western, J.R.H., 1984. Comparative Biochemistry and Physiology of Enzymatic Digestion. Academic Press, London.
Wang, W.-X., Fisher, N.S., 1996. Assimilation of trace elements by the mussel Mytilus edulis: effects of diatom chemical composition. Mar. Biol. 125, 715–724.