ADAPTASI TANAMAN LEGUM PAKAN TERHADAP
CEKAMAN KEKERINGAN DAN INOKULASI MIKORIZA
SIMEL SOWMEN
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI DISERTASI DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA*
Dengan ini saya menyatakan bahwa disertasi berjudul “Adaptasi tanaman legum pakan terhadap cekaman kekeringan dan inokulasi mikoriza” adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir disertasi ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
SUMMARY
SIMEL SOWMEN. Forage Legumes Crops Adaptation to Drought Stress and Mycorrhizal Inoculation. Under direction of LUKI ABDULLAH, PANCA DEWI MANU HARA KARTI, and DIDY SOPANDIE.
One of the major problems to ruminant production sustainable is forage availability. The problem is more influenced by the seasons, especially the dry season. Lack of soil water causes plants experiencing drought stress. To cope with drought stress, plants respond with physiological and biochemical changes. It’s causes plants undergo morphological and physiological disorders, and it will
hampered their growth and productivity. Utilization of mycorrhizae may assist
plants to overcome drought stress.
The study was conducted to recognize adaptation mechanisms of legume inoculated with mycorrhiza on drought stress, investigate effect of mycorrhizal inoculation to leguminous adaptability to overcome drought stress, and to obtain forage legume species that adaptive and productive in drought condition. Ten tropical forage legumes species consisting of Leucaena leucocephala, Indigofera zollingeriana, Desmodium sp, Calopogonium mucunoides, Macroptilium bracteatum, Centrocema pascuorum, Pueraria javanica, Clitoria ternatea, Centrocema pubescen, and Stylosanthes seabrana) were tested in different drought condition (optimum watering, drought stress) as factor. The other factor was innoculation of mychorizal fungi (no mycorrhiza, with mycorrhiza), which expected to interact with the plants in reducing drought stress.
Soil water content (swc), leaf water potential (lwp), leaf relative water content (lrwc), leaf proline, leaf water soluble carbohydrate (lwsc), a n d plant dry weight were observed. Drought stress decreased swc, lwp, lrwc and total dry weight. Drought stress also increased leaf proline and lwsc content, but lwsc was declined in Centrocema pascuorum. Mycorrhizal fungi inoculation effect was varied among legume species. Inoculation of mychorrizal fungi was increased leaf proline and lwsc content of all legume species. However, it was tended to decline proline content in tree legumes. Indigofera zollingeriana, Centrocema pascuorum and Clitoria ternatea were productive species that recommended for forage development in dry area.
RINGKASAN
SIMEL SOWMEN. Adaptasi Tanaman Legum Pakan terhadap Cekaman Kekeringan dan Inokulasi Mikoriza. Dibimbing oleh LUKI ABDULLAH, PANCA DEWI MANU HARA KARTI, dan DIDY SOPANDIE.
Salah satu permasalahan utama dalam usaha produksi ternak ruminansia adalah ketersediaan hijauan pakan. Ketersediaan hijauan pakan yang fluktuatif merupakan salah satu permasalahan besar dalam suatu usaha peternakan ruminansia. Permasalahan tersebut lebih dipengaruhi oleh musim, terutama musim kemarau. Produktifitas dari tanaman pakan sangat tergantung pada ketersediaan air tanah. Kondisi air tanah yang defisit dapat mengganggu proses morfologi dan fisiologis tanaman sehingga berakibat pada terhambatnya pertumbuhan dan produktivitas tanaman. Salah satu alternatif yang dapat digunakan untuk mengatasi permasalahan akibat cekaman kekeringan tersebut adalah dengan pemanfaatan mikoriza.
Penelitian bertujuan untuk mengetahui mekanisme adaptasi tanaman legum pakan yang diinokulasi dengan mikoriza saat cekaman kekeringan, mengetahui pengaruh inokulasi mikoriza pada adaptasi tanaman legum pakan terhadap cekaman kekeringan, dan untuk mendapatkan jenis tanaman legum pakan yang adaptif dan produktif saat cekaman kekeringan. Sepuluh jenis tanaman legum pakan terdiri dari: Leucaena leucocephala, Indigofera zollingeriana, Desmodium sp, Calopogonium mucunoides, Macroptilium bracteatum, Pueraria javanica, Clitoria ternatea, Centrocema pascuorum, Centrocema pubescen dan Stylosanthes seabrana diuji ketahanan kekeringannya dengan kondisi penyiraman yang berbeda (penyiraman optimal, cekaman kekeringan) sebagai salah satu faktor. Faktor lainnya adalah inokulasi dengan mikoriza (tanpa mikoriza, dengan mikoriza), dengan harapan dapat membantu tanaman mengurangi pengaruh cekaman kekeringan. Parameter yang diukur dalam penelitian ini adalah: 1) kadar air tanah (kat), 2) potensial air daun (pad), 3) kadar air relatif daun (kar), 4) kadar prolin daun, 5) karbohidrat terlarut air pada daun, 6) bobot kering per bagian tanaman, dan 7) bobot kering tanaman total.
Hasil penelitian menunjukkan bahwa cekaman kekeringan menurunkan kadar air tanah, potensial air daun, kadar air relatif daun dan bobot kering total, meningkatkan prolin dan kandungan karbohidrat terlarut (wsc) daun, tetapi wsc daun pada C. pascuorum mengalami penurunan. Pengaruh fungi mikoriza bervariasi pada masing-masing tanaman legum pakan. Inokulasi mikoriza meningkatkan kandungan prolin dan wsc daun pada semua jenis tanaman yang diuji. Namun terlihat adanya kecenderungan penurunan prolin daun akibat adanya inokulasi mikoriza pada tanaman legum pohon.
Berdasarkan hasil yang didapatkan pada penelitian ini, dapat ditentukan bahwa jenis tanaman legum pakan yang cukup produktif untuk dijadikan sebagai pakan ternak pada daerah dengan musim kemarau panjang adalah: I. zollingeriana, C. ternatea dan C. pascuorum
© Hak Cipta Milik IPB, Tahun 2013
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB
Disertasi
sebagai salah satu syarat untuk memperoleh gelar Doktor
pada
Program Studi Ilmu Nutrisi dan Pakan
ADAPTASI TANAMAN LEGUM PAKAN TERHADAP
CEKAMAN KEKERINGAN DAN INOKULASI MIKORIZA
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
BOGOR 2013
Penguji pada Ujian Tertutup : Dr Ir Suryahadi, DEA Dr Nurhayati D. Purwantari
Judul Disertasi : Adaptasi Tanaman Legum Pakan terhadap Cekaman Kekeringan dan Inokulasi Mikoriza
Nama : Simel Sowmen
NRP : D162080041
Disetujui oleh Komisi Pembimbing
Dr Ir Luki Abdullah, MScAgr Ketua
Dr Ir Panca Dewi MH Karti, MSi Anggota
Prof Dr Ir Didy Sopandie, MAgr Anggota
Diketahui oleh Ketua Program Studi
Ilmu Nutrisi dan Pakan
Dr Ir Dwierra Evvyernie A, MS,MSc
Dekan Sekolah Pascasarjana
Dr Ir Dahrul Syah, MScAgr
PRAKATA
Syukur Alhamdulillah penulis ucapkan kepada Allah SWT atas segala karunia-Nya sehingga Disertasi ini dapat diselesaikan. Tema yang dipilih dalam penelitian yang dilaksanakan sejak bulan Maret 2010 ini ialah kekeringan, dengan judul Adaptasi Tanaman Legum Pakan terhadap Cekaman Kekeringan dan Inokulasi Mikoriza.
Bagian dari disertasi ini telah diterbitkan pada Jurnal Media Peternakan, Institut Pertanian Bogor Volume 35 No. 2, Agustus 2012 dengan judul Physiological adaptation and biomass production of Macroptilium bracteatum inoculated with AMF in drought condition. Bagian lain dengan judul Adaptation of early growing phase leguminous feed to drought and AMF inoculation akan diajukan ke Media Peternakan.
Terima kasih penulis ucapkan kepada Bapak Dr. Ir. Luki Abdullah, MScAgr, Ibu Dr. Ir. Panca Dewi Manu Hara Karti, MSi dan Bapak Prof. Dr. Ir. Didy Sopandie, M.Agr selaku komisi pembimbing yang telah banyak memberikan sumbangan saran, waktu dan fikiran dengan kesabaran dan keikhlasan dalam proses pembimbingan saat penulis mengikuti pendidikan S3, Ucapan terima kasih kepada Dekan Fakultas Peternakan Universitas Andalas, Rektor Universitas Andalas, Rektor Institut Pertanian Bogor dan pengelola Beasiswa Program Pascasarjana Direktorat Jenderal Pendidikan Tinggi, yang telah memberikan kesempatan belajar, bantuan biaya pendidikan dan penelitian.
Penulis juga mengucapkan terima kasih kepada Dekan Sekolah Pascasarjana Institut pertanian Bogor, Dekan Fakultas Peternakan Institut Pertanian Bogor, Ketua Departemen Ilmu Nutrisi dan Teknologi Pakan dan Ketua Mayor Ilmu Nutrisi dan Pakan (INP) Pascasarjana Institut Pertanian Bogor atas bantuannya dalam kelancaran penyelesaian studi. Terima kasih juga penulis sampaikan kepada Ibu Nuril beserta staf di Laboratorium Fisiologi Stres, Puslit Biologi, LIPI, Cibinong atas bantuannya selama analisa di Laboratorium; teknisi Lab. Agrostologi, Fapet IPB dan Green House Cikabayan, University Farm, IPB atas bantuannya selama penelitian di lapangan; Mba’ Dian yang telah membantu penulis analisa di Lab; Kak Ade’, Mas Iwan, Bu Widya, Pak Iwan, Kak Irma, Ni Dwi, Ni Nu, Ni Echi, Kak Yati, Bang Icad dan Mas Supri yang banyak memberikan motivasi, masukan dan bantuan selama penulisan disertasi dan penyelesaian studi, serta Ilham dan Risty yang ikut membantu selama penelitian.
Terima kasih penulis sampaikan kepada kedua orang tua dan adik-adik atas doa, dukungan dan motivasinya. Penghargaan penulis sampaikan kepada suamiku Rusdimansyah serta putri-putriku tersayang Raihannah dan Hanifatul Humaira atas motivasi, pengertian, kesabaran dan kasih sayangnya selama penulis dalam pendidikan dan penelitian.
Semoga Disertasi ini bermanfaat untuk pengembangan ilmu pengetahuan dan dapat dijadikan sebagai bahan acuan dalam bidang peternakan dan pertanian.
DAFTAR ISI
DAFTAR TABEL xi
DAFTAR GAMBAR xii
DAFTAR LAMPIRAN xiii
1 PENDAHULUAN 1
Latar Belakang 1
Perumusan Masalah 2
Tujuan Penelitian 3
Manfaat Penelitian 3
2 TINJAUAN PUSTAKA 4
Tanaman Legum Pakan Tropis 4
Leucaena leucocephala 4
Indigofera zollingeriana 5
Desmodium sp 5
Calopogonium mucunoides 6
Macroptilium bracteatum 6
Clitoria ternatea 6
Centrocema pascuorum 7
Pueraria javanica 7
Centrocema pubescens 7
Stylosanthes seabrana 8
Peranan Air bagi Tanaman 8
Respon Tanaman terhadap Cekaman Kekeringan 9
Prolin dan Cekaman Kekeringan 12
Mikoriza 13
3 METODE 15
Bahan Penelitian 15
Peralatan Penelitian 15
Rancangan Percobaan 16
Pelaksanaan Penelitian 16
Peubah yang Diukur 17
4 HASIL DAN PEMBAHASAN 20
Adaptasi Tanaman Legum Pohon Fase Pertumbuhan Awal terhadap Cekaman Kekeringan dan Inokulasi Mikoriza 20
Kadar air tanah (KAT) 20
DAFTAR ISI (lanjutan)
Kadar air relatif (KAR) daun 23
Kadar prolin daun 25
Karbohidrat terlarut (WSC) daun 28
Bobot kering tanaman (BK) 29
Adaptasi Tanaman Legum Herba terhadap Cekaman Kekeringan dan Inokulasi Mikoriza 31
Kadar air tanah (KAT) 31
Potensial air daun (PAD) 32
Kadar air relatif (KAR) daun 36
Prolin daun 36
Karbohidrat terlarut (WSC) daun 41
Bobot kering tanaman (BK) 44
5 PEMBAHASAN UMUM 48
6 SIMPULAN DAN SARAN 55
7 DAFTAR PUSTAKA 56
DAFTAR TABEL
3.1 Tempat pengambilan bibit tanaman legum pakan yang digunakan
pada penelitian 15 4.1 Rataan kadar air tanah (%) pada tanaman legum pohon fase
pertumbuhan awal yang diinokulasi dan tanpa inokulasi mikoriza pada
dua perlakuan penyiraman 20
4.2 Rataan potensial air daun (MPa) pada tanaman legum pohon fase pertumbuhan awal yang diinokulasi dan tanpa inokulasi mikoriza pada
dua perlakuan penyiraman 21
4.3 Rataan kadar air relatif (%) daun pada tanaman legum pohon fase pertumbuhan awal yang diinokulasi dan tanpa inokulasi mikoriza pada
dua perlakuan penyiraman 24
4.4 Rataan prolin daun (µ mol/g bb) pada tanaman legum pohon fase pertumbuhan awal yang diinokulasi dan tanpa inokulasi mikoriza pada
dua perlakuan penyiraman 25
4.5 Rataan karbohidrat terlarut (WSC) daun pada tanaman legum pohon fase pertumbuhan awal yang diinokulasi dan tanpa inokulasi mikoriza
pada dua perlakuan penyiraman 29
4.6 Rataan bobot kering total (g/pot) pada tanaman legum pohon fase pertumbuhan awal yang diinokulasi dan tanpa inokulasi mikoriza pada
dua perlakuan penyiraman 30
4.7 Rataan bobot kering (akar, batang, daun) pada tanaman legum pohon fase pertumbuhan awal yang diinokulasi dan tanpa inokulasi mikoriza
pada dua perlakuan penyiraman 30
4.8 Rataan kadar air tanah (%) pada tanaman legum herba yang diinokulasi dan tanpa inokulasi mikoriza pada dua perlakuan penyiraman 34 4.9 Rataan potensial air daun (MPa) pada tanaman legum herba yang
diinokulasi dan tanpa inokulasi mikoriza pada dua perlakuan
penyiraman 35
4.10 Rataan kadar air relatif (%) daun pada tanaman legum herba yang diinokulasi dan tanpa inokulasi mikoriza pada dua perlakuan
penyiraman 39
4.11 Rataan prolin daun (µ mol/g bb) pada tanaman legum herba yang diinokulasi dan tanpa inokulasi mikoriza pada dua perlakuan
penyiraman 42
4.12 Rataan karbohidrat terlarut (WSC) daun pada tanaman legum herba yang diinokulasi dan tanpa inokulasi mikoriza pada dua perlakuan
penyiraman 43
4.13 Rataan bobot kering total (g/pot) pada tanaman legum herba yang diinokulasi dan tanpa inokulasi mikoriza pada dua perlakuan
penyiraman 44
4.14 Rataan bobot kering (akar, batang, daun) pada tanaman legum herba yang diinokulasi dan tanpa inokulasi mikoriza pada dua perlakuan
penyiraman 47
5.2 Penurunan bobot kering total pada tanaman legum pakan yang
mengalami cekaman kekeringan 50
5.3 Respon relatif mikoriza terhadap bobot kering tanaman legum pakan
akibat cekaman kekeringan (%) 51
5.4 Pengaruh inokulasi mikoriza terhadap tanaman legum pakan 53
DAFTAR GAMBAR
2.1 Tanaman Leucaena leucocephala, Indigofera zollingeriana, Desmodium sp yang termasuk tanaman legum pohon yang digunakan
dalam penelitian 5
2.2 Tanaman Calopogonium mucunoides, Macroptilium bracteatum,Pueraria javanica, Clitoria ternatea, Stylosanthes eabrana, Centrocema pascuorum, Centrocema pubescens yang termasuk
tanaman legum herba yang digunakan pada penelitian 8 2.3 Jalur biosintesis prolin pada tanaman ditandai dengan anak panah garis
tegas ( ▬► ), dan pada organisme rendah ditandai dengan garis putus -putus (---►)(Kavi Kishor et al., 2005) 12 3.1 WP4, Dewpoint PotentioMeter, alat pengukur potensial air daun 17 4.1 Interaksi faktor penyiraman dan mikoriza terhadap potensial air daun
(MPa) pada hari akhir pengamatan tanaman legum pohon fase pertumbuhan awal. –M= tanpa mikoriza, +M= mikoriza, □ = disiram tiap hari (watering), ■ = cekaman kekeringan (drought) 23 4.2 Interaksi faktor penyiraman dan mikoriza terhadap kadar air relatif
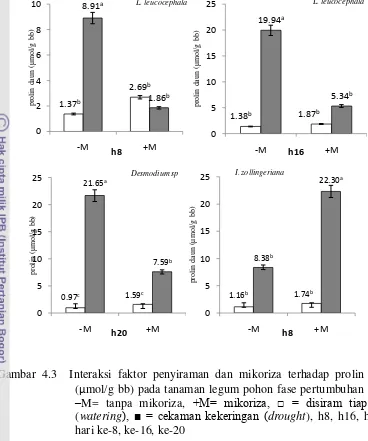
(kar) daun (%) pada hari akhir pengamatan tanaman legum pohon fase pertumbuhan awal. –M= tanpa mikoriza, +M= mikoriza, □ = disiram tiap hari (watering), ■ = cekaman kekeringan (drought) 25 4.3 Interaksi faktor penyiraman dan mikoriza terhadap prolin daun
(µmol/g bb) pada hari akhir pengamatan tanaman legum pohon fase pertumbuhan awal. –M= tanpa mikoriza, +M= mikoriza, □ = disiram
tiap hari (watering), ■ = cekaman kekeringan (drought) 27 4.4 Interaksi faktor penyiraman dan mikoriza terhadap karbohidrat terlarut
(wsc) daun (mg/g bk) pada hari akhir pengamatan tanaman legum pohon fase pertumbuhan awal. –M= tanpa mikoriza, +M= mikoriza, □ =
disiram tiap hari (watering), ■ = cekaman kekeringan (drought) 28 4.5 Interaksi faktor penyiraman dan mikoriza terhadap kadar air tanah (%)
Clitoria ternatea hari pengamatan ke-16 –M= tanpa mikoriza, +M= mikoriza, □ = disiram tiap hari (watering), ■ = cekaman kekeringan
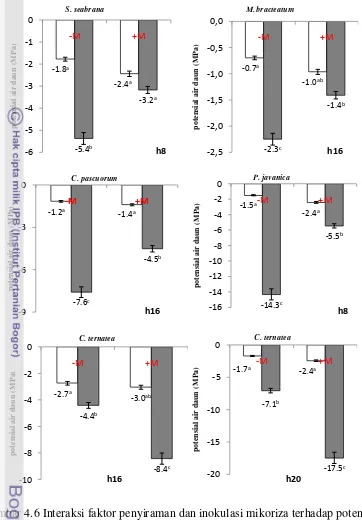
(drought) 31 4.6 Interaksi faktor penyiraman dan mikoriza terhadap potensial air daun
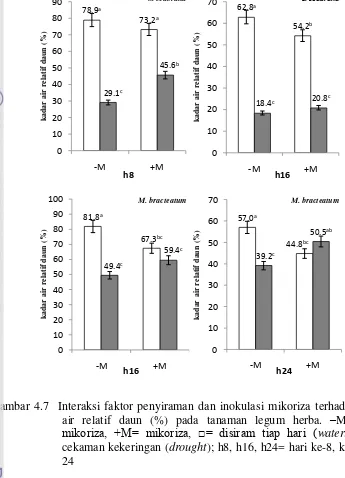
(MPa) pada tanaman legum herba –M= tanpa mikoriza, +M= mikoriza, □ = disiram tiap hari (watering), ■ = cekaman kekeringan (drought) 36 4.7 Interaksi faktor penyiraman dan mikoriza terhadap kadar air relatif
daun (%) pada tanaman legum herba –M= tanpa mikoriza, +M= mikoriza,□ = disiram tiap hari (watering), ■ = cekaman kekeringan
4.8 Interaksi faktor penyiraman dan mikoriza terhadap prolin daun (µmol/g bb) pada tanaman legum herba –M= tanpa mikoriza, +M= mikoriza, □ = disiram tiap hari (watering), ■ = cekaman
kekeringan (drought) 40
4.9 Interaksi faktor penyiraman dan mikoriza terhadap karbohidrat terlarut (wsc) daun (mg/g bk) pada tanaman legum herba –M= tanpa mikoriza, +M= mikoriza, □ = disiram tiap hari (watering), ■ =
cekaman kekeringan (drought) 43 4.10 Interaksi faktor penyiraman dan mikoriza terhadap bobot kering
(g/pot) pada tanaman legum herba –M= tanpa mikoriza, +M= mikoriza, □ = disiram tiap hari (watering), ■ = cekaman
kekeringan (drought) 45
DAFTAR LAMPIRAN
1 Analisis ragam kadar air tanah pada tanaman legum pohon 65 2 Analisis ragam potensial air daun pada tanaman legum pohon 65 3 Analisis ragam kadar air relatif daun pada tanaman legum pohon 65 4 Analisis ragam prolin daun pada tanaman legum pohon 65 5 Analisis ragam karbohidrat terlarut (WSC) daun pada tanaman legum
pohon 66 6. Analisis ragam bobot kering total pada tanaman legum pohon 66 7 Analisis ragam bobot kering (akar, batang, dan daun) pada tanaman
legum pohon 66 8 Analisis ragam kadar air tanah daun pada tanaman legum herba 67 9 Analisis ragam potensial air daun pada tanaman legum pohon 67 10 Analisis ragam kadar air relatif daun pada tanaman legum pohon 68 11 Analisis ragam prolin daun pada tanaman legum pohon 68 12 Analisis ragam karbohidrat terlarut (WSC) daun pada tanaman legum
pohon 69
13 Analisis ragam bobot kering total pada tanaman legum pohon 69 14 Analisis ragam bobot kering (akar, batang, dan daun) pada tanaman
legum pohon 70
1
PENDAHULUAN
Latar Belakang
Produktivitas usaha peternakan ruminansia sangat tergantung pada keberadaan hijauan pakan yang merupakan makanan utama bagi ternak tersebut. Sumber hijauan yang umum dimanfaatkan sebagai tanaman pada umumnya adalah berasal dari rumput-rumputan (graminae), legum dan sisa pertanian. Legum umumnya kaya akan protein bila dibandingkan dengan rumput. Penggunaan legum dalam pakan menjadi sangat penting karena merupakan sumber protein murah. Bahan pakan sumber protein pada umumnya sulit diperoleh dan sangat mahal. Oleh karena itu peternak banyak beralih mengandalkan tanaman legum untuk memperbaiki kualitas pakan.
Keberhasilan budidaya tanaman legum pakan sepanjang tahun merupakan faktor kunci ketersediaan hijauan pakan, yang berdampak langsung pada produksi ternak. Ketersediaan hijauan sangat dipengaruhi oleh musim terutama musim kemarau karena pada saat itu pertumbuhan dan produktivitas tanaman pakan berkurang dan menyebabkan kelangkaan hijauan pakan. Hal ini dapat terjadi karena pada saat kemarau kandungan air tanah tidak mencukupi untuk hidup pokok (maintenance) dan pertumbuhan tanaman. Air tanah merupakan faktor penting dalam pertumbuhan dan perkembangan tanaman. Kebutuhan air tanaman berbeda-beda tergantung pada jenis tanamannya. Defisit air merupakan salah satu faktor utama yang dapat menghambat produktifitas tanaman. Apabila jumlah air yang tersedia di tanah tidak mencukupi kebutuhan tanaman maka tanaman akan mengalami cekaman kekeringan yang ditunjukkan oleh gejala gangguan morfologi dan fisiologis, sehingga pertumbuhan dan produktivitasnya akan terhambat.
Salah satu respon tanaman terhadap defisit air tanah adalah pengurangan pertumbuhan aerial (pengurangan produksi daun baru, mengurangi area kanopi daun) dan peningkatan pertumbuhan akar. Kondisi ini menyebabkan akar secara morfologi akan memperluas jangkauannya dalam tanah, namun daun akan kehilangan banyak air akibat transpirasi yang sangat diperlukan untuk pertukaran gas dalam fotosintesis (Scott, 2008). Tanaman melakukan pengaturan osmotik dan ketika potensial osmotik sel turun, maka terjadi akumulasi solute kompatibel seperti sukrosa, sorbitol, prolin atau glycin betain untuk membantu mempertahankan turgor (Taiz dan Zeiger, 2002). Pengaturan osmotik merupakan mekanisme fisiologis utama yang berhubungan dengan pemeliharaan turgor sel dalam merespon cekaman kekeringan, dan pengaturan osmotik berhubungan juga dengan akumulasi compatible solute seperti karbohidrat terlarut dan prolin (Da Costa dan Huang, 2006).
mikoriza arbuskula (CMA) merupakan kelompok endomikoriza yaitu suatu cendawan tanah yang bersifat simbiotik obligat dengan akar tanaman, mempunyai pengaruh yang menguntungkan bagi pertumbuhan tanaman, karena dapat meningkatkan serapan hara dan air. Struktur yang terbentuk akibat kerjasama yang saling menguntungkan antara hyfa dengan akar tanaman, mempunyai kemampuan untuk meningkatkan masukan air dan hara dari tanah ke dalam jaringan tanaman.
Penelitian tentang pemanfaatan mikoriza pada tanaman pakan yang mengalami kekeringan telah dilakukan di antaranya pada rumput Setaria splendida Stapf (Karti, 2004), Gliricidia sepium dan Leucaena leucocephala (Fagbola et al., 2001), alfalfa (Goicoechea et al., 1996; Djebali et al., 2010), serta Centrosema pubescen, Calopogonium mucunoides dan Pueraria phaseoloides oleh Rahman (2006). Namun sampai sekarang masih sedikit informasi yang terkait dengan efektifitas mikoriza dan mekanisme tanaman legum pakan tropis dalam menghadapi cekaman kekeringan. Berdasarkan permasalahan tersebut dilakukan penelitian untuk melihat efektifitas mikoriza terkait dengan respon tanaman legum pakan terhadap kekeringan serta mendapatkan jenis tanaman legum pakan yang adaptif dan cukup poduktif pada kondisi cekaman kekeringan.
Perumusan Masalah
Ketersediaan hijauan pakan sepanjang tahun merupakan jaminan penting dalam meningkatkan produktivitas ternak. Sampai saat ini ketersediaan hijauan pakan sangat dipengaruhi oleh musim terutama musim kemarau di mana pada musim tersebut ketersediaan air tanah menjadi berkurang dan mengakibatkan tanaman tidak mendapatkan asupan air yang mencukupi sehingga mengalami cekaman kekeringan.
Respon awal tanaman yang mengalami cekaman kekeringan adalah melakukan penutupan stomata sehingga pemasukan C02 terhambat dan aktifitas
fotosintesis terganggu, mengakibatkan terjadinya penurunan produksi fotosintesis yang merupakan sumber energi untuk pertumbuhan dan perkembangan tanaman sehingga hal ini berdampak langsung terhadap penurunan produksi (bobot kering tanaman). Jaleel et al., (2008) menyatakan bahwa cekaman kekeringan dikarakterisasi dengan penurunan kandungan air, turgor, potensial air total, pelayuan, penutupan stomata dan pengurangan perluasan dan pertumbuhan sel. Respon tanaman dalam menghadapi cekaman kekeringan dengan mekanisme penghindaran (avoidance mechanisms) seperti; mengurangi perkembangan daun, penurunan kadar air relatif dan jumlah cabang dan juga bisa melalui mekanisme toleran (tolerance mechanisms); peningkatan rasio akar:tajuk dan penurunan ukuran sel mesofil (Li et al., 2010; Calvet et al., 2004; Meyer dan Boyer, 1981). Banyak penelitian menghasilkan bahwa mikoriza dianggap dapat membantu tanaman dalam mengatasi kekeringan melalui hypanya yang dapat membantu efisiensi penyerapan air pada akar tanaman inangnya.
Meteorologi dan Geofisika (BMG) membagi curah hujan di Indonesia menjadi 3 pola yaitu: 1) Pola Monsoon, dengan distribusi curah hujan 1 puncak musim hujan dan 6 bulan masa peralihan (curah hujan sedang); 2) Pola Ekuator, dengan dua puncak musim hujan (curah hujan tinggi); dan 3) Pola Lokal, dengan 1 puncak musim hujan (curah hujan rendah) dan memiliki pola yang berlawanan dengan pola monsoon.
Tanaman legum pakan ini belum diketahui bagaimana mekanisme adaptasinya dalam menghadapi cekaman kekeringan dan berapa lama ketahanannya terhadap cekaman kekeringan. Mikoriza dianggap dapat membantu ketahanan tanaman terhadap stres tetapi belum diketahui apakah mikoriza selalu efektif dalam membantu menghadapi cekaman kekeringan pada semua jenis tanaman legum pakan.
Respon fisiologis dan bobot kering dari tanaman legum pakan serta pengaruh pemberian mikoriza terhadap cekaman kekeringan, diharapkan dapat dijadikan sebagai dasar pengambilan jenis tanaman legum pakan yang dapat dikembangkan pada daerah dengan musim kemarau panjang.
Tujuan Penelitian
Tujuan dari penelitian ini adalah untuk: (1) menjelaskan mekanisme adaptasi tanaman legum pakan terhadap cekaman kekeringan, (2) menjelaskan pengaruh pemberian mikoriza terhadap daya adaptasi tanaman legum pakan pada kondisi cekaman kekeringan, dan (3) memperoleh tanaman legum pohon dan herba yang adaptif dan berproduksi tinggi pada kondisi cekaman kekeringan.
Manfaat Penelitian
2
TINJAUAN PUSTAKA
Tanaman Legum Pakan Tropis
Kebutuhan pakan ternak ruminansia terbesar adalah berasal dari hijauan. Hijauan pakan dimanfaatkan untuk hidup pokok dan produksi oleh ternak ruminansia. Sumber hijauan yang umum dimanfaatkan sebagai tanaman pada umumnya adalah berasal dari rumput-rumputan (graminae), legum dan sisa pertanian. Untuk memenuhi kebutuhan hijauan pakan yang besar maka perlu diperhatikan ketersediaannya agar tidak terjadi kekosongan sumber hijauan pakan. Ketersediaan hijauan pakan di daerah tropis lebih dipengaruhi oleh musim. Pada musim kemarau, kondisi air tanah menjadi defisit sehingga berpengaruh langsung terhadap pertumbuhan dan perkembangan tanaman.
Sekitar 75% konsumsi hijauan pakan di daerah tropis berasal dari rumput sedangkan pemanfaatan legum tidak begitu menonjol karena pemeliharaan yang cukup sulit pada pasture campuran (Martin, 1993). Legum merupakan jenis tanaman yang mempunyai kandungan protein kasar yang tinggi sehingga dapat digunakan sebagai sumber pakan hijauan ternak herbivora (Purbajanti, 2013). Legum mempunyai kualitas yang lebih tinggi dibanding rumput, karena memiliki serat kasar rendah, protein tinggi serta lebih disukai oleh ternak. Legum pada umumnya memiliki daun yang lebih lebar dibandingkan dengan rumput dan mempunyai kemampuan mengikat nitrogen dari udara karena bersimbiosis dengan Rhizobium yang terdapat pada bintil akarnya sehingga legum memiliki kualitas yang lebih tinggi dibanding rumput.
Martin (1993) menyatakan beberapa manfaat tanaman legum pada pastura yaitu: legum mengandung protein tinggi dan dapat memperkaya pakan bila dikombinasikan dengan rumput serta memiliki kemampuan untuk meningkatkan nitrogen pada tanah. Beberapa jenis tanaman legum yang umum dimanfaatkan sebagai sumber hijauan pakan ternak di Indonesia di antaranya adalah Leucaena leucocephala, Centrocema pubescens, Stylosanthes, Clitoria ternatea, Calopogonium, dan lain-lain. Legum tersebut merupakan sumber hijauan pakan berprotein tinggi dan murah.
Leucaena leucocephala
Leucaena berasal dari Amerika, memiliki palatabilitas tinggi, produksi tinggi, kaya protein dan toleran terhadap kekeringan (Jones, 1979, diacu dalam, Masafu, 2006; Gutteridge dan Shelton, 1998). Leucaena leucocephala termasuk dalam family Mimosacea, merupakan jenis tanaman pohon yang tumbuh bisa mencapai tinggi 7 – 18 m. Bagian daun dari tanaman ini mempunyai nilai nutrisi yang cukup tinggi bagi ternak ruminansia. Tanaman ini mengandung asam amino mimosine yang bisa menyebabkan rontok bulu pada ternak (ECHO, 2006).
2-6% tannin, merupakan senyawa fenolik phenolic yang mengikat dan melindungi protein pakan dari degradasi di dalam rumen. Produksi hijauan bervariasi tergantung kesuburan tanah, curah hujan, ketinggian, densitas dan frekuensi pemotongan 1-15 t/ha/tahun. Produksi daun maksimal didapatkan pada saat pemotongan dengan interval 6-12 minggu selama periode pertumbuhan. Sebagai pagar tanaman ekstensif pada daerah tropis dan subtropis, produksinya berkisar 2-6 t/ha/tahun (Cook et al., 2005).
Indigofera zollingeriana
Indigofera sp merupakan salah satu legum pohon yang dapat menghasilkan hijauan sepanjang tahun (Suharlina dan Abdullah, 2012). Indigofera sp dapat beradaptasi pada kondisi tanah kering dan salin, dan merupakan sumber pakan alternatif bagi ternak ruminansia (Ginting et al., 2010). Indigofera sp. memiliki produktivitas yang tinggi dan kandungan nutrien yang cukup baik, terutama kandungan proteinnya yang tinggi yakni 24,17% (Sirait et al., 2012).
Desmodium sp
Tanaman legum ini berasal dari Meksiko Barat dan Amerika, disebarluaskan ke Asia Selatan. Desmodium merupakan jenis legum semak pohon dengan tinggi 1-3 m dan bisa hidup selama 2-3 tahun, memiliki daun tipis dengan panjang sekitar 5-7 cm, bunga ungu, biji kecil dan keras. Desmodium biasa dimanfaatkan sebagai kontur pagar dan alley cropping. Kelebihan dari Desmodium ini adalah pertumbuhannya yang cepat, bisa sebagai pagar tanaman dan sumber pakan yang berkualitas bagus. Sedangkan keterbatasan Desmodium adalah hidup sebentar (2-3 tahun), tidak toleran terhadap cekaman kekeringan dan cekaman dingin serta penanaman harus dari biji (Cook et al., 2005).
Gambar 2.1 Tanaman Leucaena leucocephala,Indigofera zollingeriana, Desmo-dium sp yang termasuk tanaman legum pohon yang digunakan dalam penelitian
Calopogonium mucunoides
tahunan dengan masa hidup yang pendek, memiliki daun trifoliolate, berbunga biru atau ungu, menghasilkan 65.000-70.000 biji/kg. Merupakan sumber pupuk hijau, penutup tanah, mengurangi erosi dan meningkatkan kesuburan tanah. Meskipun secara umum palatabilitasnya rendah, ternak memakan Calopo saat musim kering di Asia tropis dan Afrika. Dapat beradaptasi pada daerah tropis basah dengan curah hujan musiman 1.500 mm. Tidak terlalu tahan terhadap kekeringan, sangat tahan terhadap genangan. Kelebihan tanaman ini adalah perkecambahannya cepat, sebagai cover crop (Cook et al., 2005).
Macroptilium bracteatum
Tanaman ini biasa disebut Burgundy bean berasal dari Amerika Selatan, terdapat di Argentina, Bolivia, Brazil, Paraguay, Peru dan Venezuela dengan curah hujan bervariasi antara 400 sampai 1600 mm. Tanaman ini tumbuh pada musim panas, tahan terhadap kekeringan, merupakan legum yang tidak menyebabkan kembung dan dapat memproduksi sampai 10 ton bahan kering per hektar. Dapat tumbuh sampai tinggi 80 cm, daunnya yang berbulu membuat tanaman ini tahan terhadap kekeringan tetapi tidak mempengaruhi palatabilitas, dapat tumbuh pada temperatur rendah dibandingkan dengan legum tropis dan subtropis lainnya, sehingga mempunyai musim tumbuh yang lama (Di P, 2010).
Kelebihan Burgundy bean di antaranya adalah: dapat mengatasi kesuburan tanah yang menurun, dapat berkecambah dan tumbuh pada kondisi dingin, tidak menyebabkan kembung pada ternak. Merupakan tanaman pakan yang berkualitas tinggi, daunnya mengandung 3.2% N, 0.23% P dan ADF 29.3%, (Whitbread dan Lawrence, 2006). M. bracteatum digunakan sebagai tanaman pastura dengan masa pendek pada tanah alkaline di daerah subtropis, tahan terhadap suhu dingin. M. bracteatum dapat berproduksi sampai 5–8 t/ha/tahun bahan kering pada daerah subtropis. Kelebihan dari tanaman ini adalah dapat berkecambah dan tumbuh pada kondisi dingin, sangat disukai, beregenerasi dengan baik dari biji setiap tahun (Cook et al., 2005).
Clitoria ternatea
Legum ini biasa disebut Butterfly pea, merupakan jenis legum yang mempunyai banyak manfaat. Clitoria ternatea merupakan tanaman pakan yang memiliki palatabilitas tinggi, mempunyai batang yang sangat kecil dengan daun yang lebar, tidak menyebabkan kembung dan tidak mengandung toksik membuat tanaman ini ideal sebagai hijauan. Tanaman ini tahan terhadap dingin dan kekeringan. Kandungan protein kasar mencapai 10.5% - 25.5% (Gomez dan Kalamani, 2003). Manfaat lain C. ternatea disamping sebagai tanaman pakan dan penutup tanah, ternyata C. ternatea juga sangat baik sebagai tanaman obat. Potensi tanaman C. ternatea sebagai tanaman obat didukung oleh kandungan senyawa kimia seperti saponin, flavonoid, alkaloid, Ca-oksalat, dan sulfur (Suarna, 2012).
pertumbuhan, sedangkan produksi saat musim kering adalah 2-6 t/ha/tahun bahan kering (Cook et al., 2005).
Centrosema pascuorum
Tanaman pakan ini berasal dari Meksiko dan didistribusikan ke Brazil, Ekuador, Venezuela. Centrosema pascuorum merupakan tanaman semusim, dan dapat beradaptasi pada daerah tropis dengan musim kering sampai 8 bulan, cocok pada daerah dengan curah hujan 700-1.500 mm per tahun. Tanaman individu mati pada kondisi kekeringan, tetapi populasinya bertahan melalui mekanisme drought escape dengan percepatan tumbuh dan pembungaan serta produksi biji yang tinggi. Palatabilitas cukup bagus, kandungan protein kasar dan kecernaan bervariasi tergantung umur tanaman dan musim, berturut-turut berkisar antara 6-27% dan 42-79%. Produksi bahan kering 4-9 t/ha/tahun (Cook et al., 2005). Pueraria javanica
Legum ini biasa disebut Kudzu tropis dengan nama latin Pueraria javanica (Roxb.) Benth, merupakan tanaman pakan yang secara tradisional sebagai pakan sapi dan penutup tanah (Cordial et al., 2006). Pueraria javanica merupakan legum tahunan yang memanjat, sedikit berkayu, berbulu, diameter batang utama 6 mm dan panjangnya bisa mencapai 10 m. Pucuk muda biasanya dilapisi oleh bulu coklat. Cukup adaptif terhadap berbagai tipe tanah tetapi tidak bagus pada tanah liat. Adaptasi bagus pada tanah masam (pH 3.5-5.5), tetapi membutuhkan media tanah dengan kesuburan tinggi. Tidak tahan terhadap salinitas, cukup tahan terhadap genangan air tapi dalam waktu pendek. Legum ini kurang tahan terhadap kekeringan, dapat bertahan selama 4-5 bulan, tetapi akan mengalami kehilangan daun cukup banyak. Tanaman ini memiliki kandungan protein kasar 12-24%, kecernaan 60-70%. Kualitas nutrisi menurun saat musim kering. Di Villavicencio, Kolombia, karakteristik kimia tanaman umur 6 bulan 50% KCBK, 22.6% PK, 0.30% P dan 0.65% Ca pada musim hujan; dibandingkan dengan 55.5% KCBK, 19.8% PK , 0.23% P dan 0.52% Ca pada musim kemarau (Cook et al, 2005). Centrocema pubescens
Gambar 2.2 Tanaman Calopogonium mucunoides, Macroptilium bracteatum, Pueraria javanica, Clitoria ternatea. Stylosanthes seabrana, Centrocema pascuorum, Centrocema pubescens yang termasuk tanaman legum herba yang digunakan pada penelitian
Stylosanthes seabrana
Legum ini dapat dimanfaatkan sebagai sumber hijauan pakan untuk sapi, kerbau, domba, kambing dan babi; untuk reklamasi lahan dan sebagai tanaman campuran pada lahan kering (Chandra et al., 2006). Stylosanthes seabrana pertama dikenalkan pada tahun 1980 oleh CSIRO’s Plant Introduction group, Australia dan CIAT’s Genetic Resources group, Colombia (Vanni dan Fernandez, 2011). Stylosanthes berasal dari Brazil, biasa digunakan sebagai legum pastura dan merupakan legum musim panas. Tanaman ini dapat hidup pada daerah dengan curah hujan 400-1.190 mm per tahun dan 2-7 bulan musim kering, cukup adaptif pada lingkungan tropis dan subtropis dengan curah hujan 500-1.000 mm per tahun, sangat tahan terhadap kekeringan, dan tidak dapat tumbuh pada tanah yang tergenang (Cook et al., 2005).
Peranan Air bagi Tanaman
(mis. sukrosa pada floem), ion inorganik (nutrient dari akar ke daun pada xylem); dan atmosfer gas (difusi oksigen untuk respirasi) (Fitter dan Hay, 2002).
Noggle dan Frizt (1983) menjelaskan fungsi air bagi tanaman yaitu sebagai: (1) senyawa utama pembentuk protoplasma, (2) senyawa pelarut bagi masuknya mineral-mineral dari larutan tanah ke tanaman dan sebagai pelarut mineral nutrisi yang akan diangkut dari satu bagian sel ke bagian sel lain, (3) media terjadinya reaksi-reaksi metabolik, (4) reaktan pada sejumlah reaksi metabolisme seperti siklus asam trikarboksilat, (5) penghasil hidrogen pada proses fotosintesis, (6) menjaga turgiditas sel dan berperan sebagai tenaga mekanik dalam pembesaran sel, (7) mengatur mekanisme gerakan tanaman seperti membuka dan menutupnya stomata, membuka dan menutupnya bunga serta melipatnya daun-daun tanaman tertentu, (8) berperan dalam perpanjangan sel, (9) bahan metabolisme dan produk akhir respirasi, serta (10) digunakan dalam proses respirasi.
Kebutuhan air pada tanaman dipengaruhi oleh beberapa faktor antara lain jenis dan umur tanaman, kadar air tanah dan kondisi cuaca. Fungsi air adalah sebagai: (a) komponen essensial tumbuhan, (b) pelarut, di dalamnya terdapat gas. garam dan zat terlarut lainnya yang bergerak keluar masuk sel, (c) pereaksi dalam fotosintesis dan pada berbagai proses hidrolisis, dan (d) air essensial untuk menjaga turgiditas di antaranya dalam pembesaran sel dan pembukaan stomata (Griffin et al., 2004; Kramer dan Boyer, 1995). Ketersediaan air dalam tanaman diperoleh melalui proses fisiologis dan hilangnya air dari permukaan bagian tanaman melalui proses evaporasi dan transpirasi.
Pergerakan air pada tanaman diatur oleh perbedaan potential air, yaitu: air mengalir dari potensial tinggi menuju potensial rendah. Tujuan pergerakan air pada tanaman adalah: (1) sebagai alat transport nutrisi dari tanah menuju organ tanaman yang memerlukan, (2) agar sel tanaman tetap tegak (disebut turgor), (3) untuk transpirasi tanaman, dan (4) tanaman dapat secara aktif mengatur laju aliran air (transpirasi) melalui pengaturan ukuran dari pembukaan stomata (Bouman et al., 2007).
Respon Tanaman terhadap Cekaman Kekeringan
Cekaman kekeringan merupakan salah satu faktor lingkungan yang berdampak sangat buruk terhadap pertumbuhan tanaman sehingga dapat menyebabkan penurunan produksi tanaman (Jun-Feng et al., 2010). Cekaman kekeringan terjadi ketika ketersediaan air tanah menurun dan kondisi atmosfir menyebabkan kehilangan air terus menerus melalui transpirasi atau evaporasi (Jaleel et al., 2009; Taiz dan Zeiger, 2002). Salah satu penyebab terjadinya cekaman kekeringan adalah tingginya kecepatan evaporasi yang melebihi laju absorbsi air oleh akar. Lebih lanjut dijelaskan bahwa cekaman kekeringan ditandai dengan rendahnya kadar air, penyusutan potensial air daun dan tekanan turgor, penutupan stomata serta berkurangnya pembesaran dan pertumbuhan sel (Borges, 2003).
dikelompokkan pada 2 mekanisme utama yaitu drought tolerance (mentolerir kekeringan) dan drought avoidance (menghindari kekeringan) (Salisbury dan Ross, 1992). Respon tanaman dalam menghadapi cekaman kekeringan dengan mekanisme penghindaran (avoidance mechanisms) seperti; mengurangi perkembangan daun, penurunan kadar air relatif dan jumlah cabang dan juga bisa melalui mekanisme toleran (tolerance mechanisms); peningkatan rasio akar:tajuk (Li et al., 2010; Calvet et al., 2004; Meyer dan Boyer, 1981). Mekanisme drought avoidance dikarakterisasi dengan nilai potensial air daun lethal dan kadar air relatif (RWC) yang relatif masih tinggi dan osmotik adjustment yang relatif kecil, tanaman yang mengembangkan mekanisme drought tolerance dikarakterisasi dengan status air daun (potensial air daun dan RWC) lethal yang rendah dan meningkatkan akumulasi solute aktif saat mengalami cekaman kekeringan (Auge et al., 1998). Jenk dan Hasegawa (2005) menyatakan bahwa tanaman yang menghindari kekeringan berusaha untuk mempertahankan potensial air tetap tinggi dan tanaman yang toleran terhadap kekeringan adalah dapat mentolerir saat terjadinya defisit air. Mekanisme toleran terhadap kekurangan air bisa dihubungkan dengan akumulasi dari osmoprotektan seperti gula terlarut (Salwa dan Heba, 2011; Tatar dan Gevrek, 2008; Parida et al., 2007).
Menurut Hamim (2004), pengaruh cekaman kekeringan bergantung pada genetik tanaman, di mana perbedaan morfologi, anatomi dan metabolisme akan menghasilkan respon yang berbeda terhadap cekaman kekeringan. Ketika jumlah absorbsi air mulai terbatas, maka tanaman memiliki mekanisme untuk mencegah kehilangan air dengan melakukan penutupan stomata. Perubahan pada ketahanan mekanisme stomata sangat diperlukan untuk mengatur kehilangan air oleh tanaman dan untuk mengatur pengambilan karbondioksida (CO2) yang penting
untuk ketersediaan fiksasi CO2 selama proses fotosintesis (Taiz dan Zeiger, 2002).
Jaleel et al., (2008) menyatakan bahwa cekaman kekeringan dikarakterisasi dengan penurunan kandungan air, turgor, potensial air total, pelayuan, penutupan stomata dan pengurangan perluasan dan pertumbuhan sel. Cekaman kekeringan yang parah dapat menyebabkan fotosintesis terhenti, menghambat metabolisme dan akhirnya mati. Kekeringan selain menurunkan laju fotosintesis, juga menyebabkan penurunan laju pertumbuhan akibat rendahnya potensial air dan turgor tumbuhan (Tezara et al., 2002).
Tanaman yang mengalami cekaman kekeringan merespon dengan perubahan ditingkat seluler dan molekuler seperti perubahan pada pertumbuhan tanaman. volume sel menjadi lebih kecil, penurunan luas daun, daun menjadi tebal, adanya rambut pada daun, peningkatan ratio akar-tajuk, sensitivitas stomata, penurunan laju fotosintesis, perubahan metabolisme karbon dan nitrogen, perubahan produksi aktivitas enzim dan hormon (Pugnaire et al., 1999). Cekaman kekeringan dapat menghambat pertumbuhan tanaman, salah satunya dapat dilihat pada perluasan daun. Penurunan luas daun merupakan respon pertama tanaman terhadap kekeringan. Keterbatasan air akan menghambat pemanjangan sel yang secara perlahan akan menghambat pertumbuhan luas daun. Kecilnya luas daun akan menyebabkan rendahnya transpirasi, sehingga menurunkan suplai air dari akar ke daun. Jika kondisi ini dibiarkan terus menerus lama kelamaan akan terjadi absisi daun (Taiz dan Zeiger, 2002).
respirasi; (d) berkurangnya perubahan asimilasi terus menerus pada organ pertumbuhan; (e) mempercepat penuaan daun; (f) meningkatkan rasio akar tajuk (Banziger et al., 2000). Vurayai et al. (2011) menyatakan bahwa cekaman kekeringan menurunkan laju ekpansi relatif daun, jumlah daun, tinggi tanaman, dan rasio tajuk:akar tergantung pada tahap perkembangan dari tanaman itu sendiri pada saat terjadi cekaman kekeringan.
Kadar air relatif / Relatif Water Content (RWC) yang menggambarkan kadar relatif air daun merupakan salah satu parameter ketahanan tanaman menghadapi cekaman kekeringan. Proses fotosintesis pada sebagian besar tanaman akan mulai tertekan bila nilai RWC kurang dari 70%, sehingga tanaman memerlukan pengaturan dalam tubuhnya diantaranya dengan melakukan penutupan stomata (Quilamboo, 2004). Menurut Ashri (2006) cekaman kekeringan selama 14 hari pengamatan akan menurunkan nilai RWC menjadi 43-33% pada varietas kedelai budidaya dan 24 hari sebesar 30% pada varietas kedelai liar. RWC dapat dijadikan ukuran ketahanan tanaman terhadap cekaman kekeringan.
Shao et al. (2008) menyatakan bahwa potensial air daun dan RWC menurun pada setiap tahap pertumbuhan shorgum yang mengalami cekaman kekeringan. Kadar air relatif berhubungan dengan jumlah air yang tersisa setelah dehidrasi yang didapatkan saat turgid jaringan penuh dan memberi informasi tentang sebagian kecil kandungan air jaringan. Menurut Stoyanov (2005) cekaman kekeringan secara signifikan mempengaruhi perubahan pada RWC dan potensial air daun pada daun tanaman young bean. Setelah kekeringan hari ke-21, stem water potensial, RWC, rasio berat turgid dan berat kering, dan konsentrasi pati menurun secara signifikan dibandingkan dengan kontrol, sedangkan jumlah prolin dan potassium meningkat secara signifikan yang mengindikasikan bahwa kemampuan dalam pengaturan osmotik pada Miniature rose (Meshkinjan et al., 2010).
Akumulasi dari beberapa compatible solute seperti gula, betain dan proline yang mengatur potensial osmotik sel merupakan reaksi awal tanaman pada saat mengalami cekaman kekeringan (Tatar dan Gevrek, 2008). Compatible solute berperan penting dalam toleran kekeringan, karena dapat melindungi tanaman dari stres melalui mekanisme berbeda termasuk pengaturan osmotik selular, detoksifikasi ROS, melindungi integritas membran dan penstabil protein enzim (Ashraf dan Foolad, 2007).
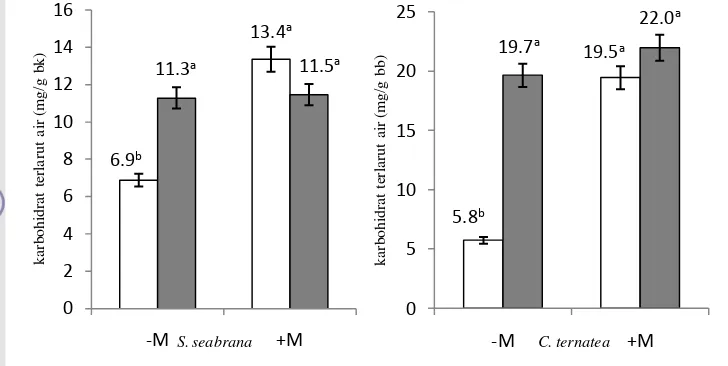
Menurut Volaire dan Thomas (1995) pada perlakuan cekaman kekeringan, total gula terlarut (water soluble carbohydrate) meningkat dari 10%-30% pada hari ke 43. Beberapa tanaman sub-tropis, termasuk Wheat (Triticum sp) menyimpan kelebihan carbon dalam bentuk water soluble carbohydrate (WSC), terutama mengandung fructo-oligosakarida (fruktan) seperti sukrosa dan heksosa (Ruuska et al., 2006). Karena kandungannya dalam jumlah banyak dan secara cepat dapat dimobilisasi, WSC diimplikasikan sebagai sumber karbon dominan untuk mobilisasi pada grains, terutama ketika fotosintesis aktif yang terhambat akibat cekaman kekeringan pada sereal (Foulkes et al., 2007).
Bentgrass (Da Costa dan Huang, 2006), tembakau (Yue et al., 2011) dan kedelai (Hapsoh et al., 2006). Peningkatan akumulasi prolin pada tanaman yang terkena cekaman merupakan parameter yang bagus untuk mengetahui ketahanan tanaman terhadap cekaman kekeringan. Pada tanaman Oryza sativa yang mengalami cekaman kekeringan, terjadi degradasi protein dan konsekuensinya adalah peningkatan kandungan prolin dan penurunan kandungan protein pada daun (Roy et al., 2009). Kondisi cekaman kekeringan meningkatkan konsentrasi beberapa jenis asam amino, jumlah prolin bebas adalah sekitar 60% dari seluruh asam amino yang terakumulasi (Yamada et al., 2005).
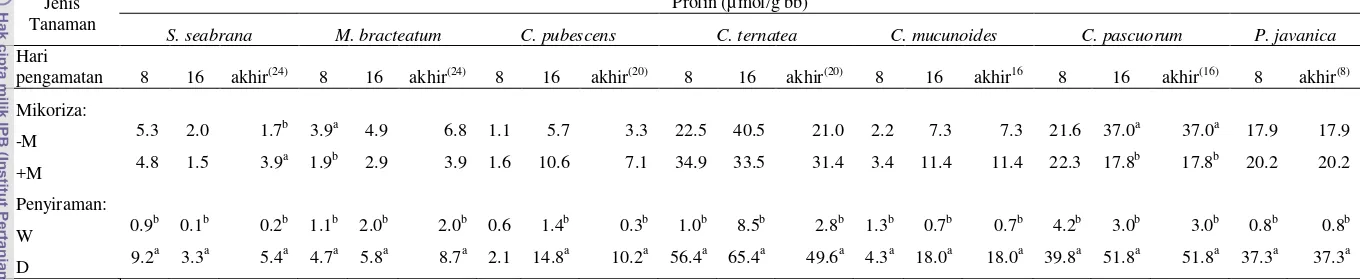
Prolin dan Cekaman Kekeringan
Tanaman pada kondisi cekaman kekeringan mengakumulasi prolin, beberapa peneliti menyatakan tentang peningkatan akumulasi prolin pada tanaman gandum (Johari-Pireivatlou et al., 2010), Petunias (Yamada et al., 2005), Aeluropus (Vaziri et al., 2011), dan tomat (Jurekova et al., 2011) yang mengalami cekaman kekeringan. Pada kondisi cekaman kekeringan moderate (-1 MPa) dan tinggi (-1.5 MPa), tanaman canola meproduksi prolin dalam jumlah tinggi untuk melindungi sel dari cekaman, kandungan prolin pada akar relatif lebih tinggi dibanding tajuk (Omidi, 2010). Peningkatan prolin bertujuan untuk melindungi sel tanaman yang mengalami cekaman kekeringan (Omidi, 2010); membantu melindungi dan perkembangan sel dari kerusakan osmotik akibat cekaman (Khalil dan El-Noameni, 2012).
Peningkatan nilai prolin seiring dengan peningkatan cekaman merupakan salah satu mekanisme pertahanan pada tanaman yang terkena cekaman yang digunakan untuk menurunkan potensial osmotik sel, sehingga meningkatkan asupan air pada sel (Khalil dan El-Noameni, 2012). Akumulasi prolin yang dramatis disebabkan peningkatan biosintesis prolin dan berkurangnya degradasi prolin, sebaliknya penurunan tingkat akumulasi prolin pada tanaman yang dehidrasi disebabkan rendahnya biosintesis prolin dan meningkatnya degradasi prolin (Yoshiba et al., 1997; Kavi Kishor et al., 2005).
Gambar 2.3 menjelaskan bahwa pada tanaman yang mengalami cekaman, prolin tidak hanya disintesis dari glutamate, tetapi juga dari arginin atau ornithin. Arginin dikonversi menjadi ornithin oleh enzim arginase. Pada tanaman, GSA secara langsung berasal dari ornithine melalui enzim ornithine d-aminotransferase (d-OAT) (Kavi-kishor et al., 2005). Sintesis prolin utamanya melalui glutamate, dimana direduksi menjadi glutamate-semialdehyde (GSA) oleh enzim carboxylate synthetase (P5CS) dan secara spontan berubah menjadi pyrroline-5-carboxylate (P5C). Kemudian P5C reductase (P5CR) mereduksi P5C intermediate menjadi proline. Jalur alternatif, prolin dapat juga disintesis dari ornithin, dimana awalnya ditransaminasi oleh ornithine-delta-aminotransferase (OAT) memproduksi GSA dan P5C, kemudian dirubah menjadi prolin (Szabados dan Savoure, 2009).
Mikoriza
Definisi mikoriza pertama kali dikemukakan oleh Frank pada tahun 1885. Mikoriza dapat didefinisikan sebagai hubungan mutualisme antara cendawan dengan tanaman induk, mikoriza mendapatkan karbohidrat dari tanaman dan sebagai gantinya tanaman mendapatkan nutrisi dari mikoriza (Albrecht et al., 1999; Scott, 2008; Pearson et al., 2006). Berdasarkan morfologinya mikoriza dapat digolongkan menjadi: ektomikoriza, endomikoriza (mikoriza arbuskular), arbutoid, monotropoid, ectendo, ericoid dan orchid (Smith dan Read, 2008; Scott, 2008).
Pertumbuhan fungi mikoriza dimulai 2-10 hari setelah diinokulasi pada akar tanaman. Fungi mikoriza mengeluarkan hyfa dan memproduksi jaringan radikal (radical shape network) dengan diameter 2.5 mm. Kontak pertama antara mycelium dan akar terjadi pada 1-3 hari setelah perkecambahan. Tujuh hari setelah kontak antara fungi-induk, beberapa spora sekunder atau strukturnya seperti vesikel terlihat sama dengan spora sebenarnya kecuali pada ukuran (diameter 20-30 µm). Spora asli yang pertama terbentuk adalah 25 hari setelah terjadi kontak dan jumlah spora akan meningkat secara eksponensial. Spora ini hyaline dan keputih-putihan pada awalnya, tetapi berubah menjadi kuning kecoklatan. Setelah 12 minggu, 1000-2500 spora terlihat pada setiap plat (Eskandari dan Danesh, 2010).
Secara hakiki sistem pertahanan tanaman terhadap cekaman, beberapa mikroorganisme tanah terbukti dapat mengurangi symptom cekaman. Mikoriza bersimbiosis dengan akar tanaman dengan cara mengkolonisasi jaringan kortikal akar selama masa pertumbuhan aktif tanaman (Song, 2005). Inokulasi dengan arbuscular mikoriza (AM) tidak mempengaruhi pH tanah tetapi meningkatkan keseimbangan agregat tanah dan menurunkan konsentrasi ion garam pada tanah, meningkatkan pertumbuhan tanaman, kandungan gula terlarut, kandungan klorofil, dan aktifitas akar dibandingkan dengan tanaman tanpa mikoriza (Li et al., 2012).
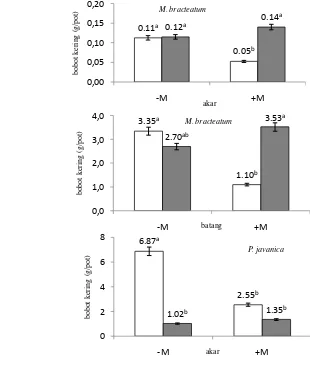
meningkatkan ketahanan tanaman terhadap cekaman kekeringan. Simbiosis dengan AM dapat memperbaiki kapasitas serapan dan meningkatkan pertumbuhan dari tanaman induk, terbukti pada sugarcane, mung bean, apel, jeruk, gandum, tomat dan wild jujube (Wu dan Xia, 2004). Wu et al. (2008) menyatakan bahwa kolonisasi dengan fungi arbuskular mikoriza dapat meningkatkan pertumbuhan pada tanaman Citrus dan dapat mempengaruhi penyimpanan kandungan air tanah melalui glomalin’s pada stabilitas agregat air tanah.
Mekanisme mikoriza dalam meningkatkan ketahanan tanaman terhadap cekaman kekeringan melalui beberapa faktor sebagai berikut: (1) meningkatkan hara tanah di rizosper; (2) memperluas area akar tanaman sehingga meningkatkan efisiensi penyerapan air; (3) mengaktifkan sistem pertahanan tanaman secara cepat; (4) memprotek tanaman dari kerusakan oksidatif karena cekaman kekeringan (Song, 2005). Wu dan Xia (2006) menjelaskan bahwa kolonisasi dengan AM menstimulasi pertumbuhan dan biomassa pada tanaman Poncirus trifoliatartifoliate tanpa memperhatikan status air. Keuntungan dari kolonisasi dengan AM pada kondisi cekaman kekeringan adalah pada peningkatan pengaturan osmotik tanaman. Peningkatan ketahanan kekeringan pada tanaman bermikoriza bisa dihubungkan dengan kemampuan dari hypa untuk mengakses ruang pori yang kecil yang tidak bisa dilakukan oleh akar dan rambut akar dari tanaman induk dan penyerapan air oleh mycelia dari AM untuk mempertahankan aktivitas fisiologis selama cekaman kekeringan (Christopher et al., 2008).
3
METODE
Bahan Penelitian
Tanaman pakan yang digunakan dalam penelitian adalah 10 jenis tanaman legum yang umum dipergunakan sebagai pakan ternak di Indonesia dan dipilih dari beberapa daerah di Indonesia (Tabel 3.1). Jenis dari tanaman legum pakan yang diteliti terdiri dari dua kelompok, yaitu kelompok pohon (Leucaena leucocephala, Indigofera zollingeriana dan Desmodium sp) dan kelompok herba (Calopogonium mucunoides, Macroptilium bracteatum, Centrocema pascuorum, Pueraria javanica, Clitoria ternatea, Centrocema pubescens, dan Stylosanthes seabrana).
Bahan-bahan yang digunakan pada penelitian ini adalah: tanah, pupuk kandang, mikoriza dengan merk dagang Mycofer (Gigaspora margarita dan Glomus manihotis) dari Laboratorium bioteknologi kehutanan, PAU, IPB; aquades; es batu, serta bahan-bahan kimia untuk analisa prolin dan karbohidrat terlarut (WSC) daun.
Tabel 3.1 Tempat pengambilan bibit tanaman legum pakan yang digunakan pada penelitian
No. Legum Asal
1. Leucaena leucocephala Jawa Barat
2. Indigofera zollingeriana Jawa Barat
3. Desmodium sp Jawa Barat
4. Calopogonium mucunoides Jawa Timur
5. Centrocema pubescens Jawa Timur
6. Pueraria javanica Jawa Timur
7. Macroptilium bracteatum NTT
8. Clitoria ternatea NTT
9. Centrocema pascuorum NTT
10. Stylosanthes seabrana NTT
Peralatan Penelitian
Rancangan Percobaan
Penelitian ini dirancang menggunakan Rancangan Acak Lengkap (RAL) pola faktorial 2x2 dengan 3 ulangan, yang terdiri dari faktor pemberian mikoriza, yaitu: tanpa mikoriza (-M) dan inokulasi dengan mikoriza (+M); dan faktor penyiraman, yaitu: disiram (W) dan tidak disiram (D). Model matematis mengacu pada Steel dan Torrie (1995).
Model linier analisis keragaman pada penelitian ini adalah: Yijk = µ + αi + βj+ (αβ)ij+ εijk
Keterangan:
Yijk = Nilai pengamatan pada perlakuan mikoriza ke-i, penyiraman ke-j dan ulangan ke-k
µ = rataan umum
αi = pengaruh mikoriza ke-i
βj = pengaruh penyiraman ke-j
(αβ)ij = pengaruh interaksi mikoriza ke-i dan penyiraman ke j
εijk = pengaruh galat
Data yang diperoleh dianalisis dengan ANOVA, jika terdapat pengaruh terhadap peubah yang diukur maka dilanjutkan dengan uji LSMEAN.
Pelaksanaan Penelitian
Penelitian Pelaksanaan penelitian ini dimulai dengan mempersiapkan media tanam menggunakan tanah latosol dan pupuk kandang dengan perbandingan = 9: 1, tanah sebanyak 4.5 kg dan pupuk kandang 0.5 kg. Bibit tanaman legum pakan yang telah disiapkan disemai terlebih dahulu, ditumbuhkan sampai berumur 1 bulan. Selanjutnya tanaman legum pakan dipindahkan ke dalam pot kapasitas 5 kg media tanam. Khusus untuk perlakuan dengan mikoriza, pada media tanam ditambahkan Mycofer (inokulasi mikoriza komersial yang mengandung G. margarita dan G. manihotis) sebanyak 20 gram (±1 sendok makan) pada lubang tanam kemudian baru tanaman dipindahkan. Setiap lubang tanam ditanami dengan 2 individu tanaman legum pakan.
Tanaman dipelihara selama ± 1 bulan. Pemeliharaan dilakukan dengan menyiram tanaman dua hari sekali, membersihkan gulma yang tumbuh secara manual, dan pemupukan dengan NPK dilakukan pada 14 hari setelah tanaman dipindahkan ke dalam pot. Setelah tanaman tumbuh dengan baik selanjutnya dilakukan pemangkasan awal (trimming) untuk menyeragamkan tinggi tanaman. Satu minggu kemudian perlakuan cekaman kekeringan dimulai yaitu dengan tidak melakukan penyiraman sampai tanaman mati (untuk perlakuan cekaman kekeringan = D), sedangkan untuk perlakuan disiram (W) dilakukan penyiraman1 kali sehari yaitu pada pagi hari.
yang tidak diselotip untuk memudahkan penyiraman. Perlakuan dimulai pada keesokan harinya dan dihitung sebagai hari ke-0. Pengambilan sampel kadar air tanah, kadar air relatif daun, dan potensial air daun dilakukan per 4 hari, serta sampel prolin dilakukan per 8 hari dimulai sejak hari ke-0. Penyiraman pot untuk perlakuan W dilakukan setiap pagi hari, sedangkan untuk perlakuan D tidak dilakukan penyiraman sampai tanaman mencapai lethal point dan ini berarti perlakuan dihentikan. Selanjutnya tanaman dipotong (dipanen), dan dapat dilakukan pengambilan sampel tanaman untuk peubah karbohidrat terlarut (WSC) daun dan bobot kering tanaman.
Peubah yang diukur Kadar air tanah (%)
Kadar air tanah diuji secara manual, yaitu dengan cara mengambil 5 gram sampel tanah segar (BS) yang diambil dari tanah pada kedalaman 20 cm, kemudian dioven 1050 C selama 24 jam. Kemudian timbang berat setelah dioven (BO). Kadar air tanah dapat dihitung menggunakan rumus berikut:
KAT = � −�
� � 100%
Kadar air tanah diukur setiap 4 hari sekali, mulai dari hari ke-0 sampai perlakuan cekaman kekeringan selesai yang ditandai dengan tanaman mulai menunjukkan tanda-tanda kematian (lethal point).
Bobot kering akar, batang dan daun (g/pot)
Setelah pemanenan, bagian tanaman seperti akar, batang dan daun segar dikeringkan pada suhu 70º C selama 48 jam, kemudian ditimbang sebagai bobot kering akar, batang dan daun.
Potensial air daun (MPa)
Sampel yang digunakan untuk pengukuran potensial air daun adalah daun segar tanaman legum pakan. Sampel diambil pada pukul 4 WIB dini hari dan langsung dimasukkan ke dalam cup WP4, ditutup dan dimasukkan ke dalam coolbox. Pengukuran potensial air daun legum dilakukan setiap 4 hari sekali.
Potensial air daun pada kondisi tanpa cekaman dan cekaman kekeringan diuji menggunakan WP4 Dewpoint PotentioMeter (Gambar 3.1), dengan cara: sampel daun segar dipotong menjadi beberapa bagian kemudian dimasukkan ke dalam cup WP4. Cup+sampel tanpa tutup dimasukkan ke dalam alat potensiometer dan tunggu sampai di layar menunjukkan nilai seperti ini: “ Ts-Tb=-0,58”, kemudian tombol diputar ke posisi “read”, tunggu hingga terdengar bunyi bip beberapa kali dan lampu keseimbangan menyala. Setelah angka menunjukkan posisi konstan, hasil yang tertera di layar WP4 dicatat sebagai nilai potensial air daun (ICT International, 2010). Contoh hasil pengukuran potensial air daun yang terbaca ditunjukkan pada Gambar 3.1.
Kadar air relatif daun (%)
Pengukuran kadar air relatif daun dilakukan setiap 4 hari sekali sampai tanaman mati. Nilai kadar air relatif daun diukur dengan cara: sampel daun segar ditimbang sehingga didapatkan berat segar (BS), kemudian dimasukkan ke dalam cup. Cup berisi sampel daun segar diberi aquadest sehingga seluruh permukaan daun terendam, ditutup dengan kertas saring, dan disimpan dalam suhu ruang selama 18-24 jam. Air yang masih tersisa dibuang dan sampel ditiriskan dengan tisu, ditimbang sehingga didapatkan berat turgid (BT). Kemudian sampel di oven pada suhu 600 C selama 2x24 jam, ditimbang dan didapatkan berat kering (BK). Nilai kadar air relatif daun didapatkan dengan menggunakan perhitungan:
�� ������ �� � � �= � − ��
� − �� �100%
Kadar prolin daun (µ mol/g bb)
Pengambilan sampel untuk analisa prolin dilakukan setiap 8 hari sekali sampai tanaman mati. Kadar prolin daun diuji mengacu pada metode Bates et al., (1973), dengan prosedur sebagai berikut: sampel daun segar digerus menggunakan nitrogen cair sampai sampel menjadi halus (<1 mm), dan sekitar 100 mg hasil gerusan ditempatkan dalam mikro tube 1.5 ml. Kemudian ditambahkan 1.3 ml asam sulfosalisilat 3% dan diaduk menggunakan vortex. Untuk memisahkan supernatan dengan padatan daun, tabung disentrifuse pada kecepatan 12.000 rpm selama 10 menit pada suhu 40 C. Selanjutnya 200 l supernatan diambil dan dimasukkan ke dalam tabung baru, kemudian direaksikan dengan 200 l larutan asam ninhidril (0,125 gr ninhidril, 3 ml asam asetat glasial, 2 ml 6M fosforic acid dengan agitasi dan pemanasan) dan diaduk menggunakan vortex. Tube diinkubasi dalam waterbath pada suhu 100C, selama 1 jam, kemudian disimpan dalam wadah berisi es batu untuk menghentikan reaksi. Ke dalam setiap tube dimasukkan 400 l toluen kemudian diaduk dengan vortex hingga mengeluarkan cairan merah. Ambil 100 l cairan merah dari hasil reaksi dan dipindahkan ke dalam tube baru, dicampur dengan 900 l toluen. Kemudian dibaca dengan spektrofotometer pada panjang gelombang 520 nm. Konsentrasi prolin ditentukan terhadap kurva standar proline yang dibuat menggunakan bahan prolin murni.
Kadar karbohidrat terlarut (WSC) daun (mg/g bk)
20 – 30 mg sampel daun kering atau akar kering diekstrak sebanyak 4 kali masing-masing selama 15 menit dalam 10 ml air mendidih. Setelah itu disentrifus pada 3500 rpm selama 10 menit. Supernatant dikoleksi dan dikumpulkan hingga volume akhir mencapai 50 ml. Satu ml supernatant diambil dan dimasukkan ke dalam tabung baru dan ditambahkan satu ml larutan phenol 18 % dan 5 ml konsentrat asam sulfur (H2SO4 concentrate). Campuran tersebut diaduk
menggunakan vortex, kemudian dibaca dengan spektrofotometer pada panjang gelombang 490 nm.
4
HASIL DAN PEMBAHASAN
Adaptasi Tanaman Legum Pohon Fase Pertumbuhan Awal terhadap Cekaman Kekeringan dan Inokulasi Mikoriza
Kadar air tanah (KAT)
Kadar air tanah (KAT) yang tersedia mempengaruhi pertumbuhan dan perkembangan tanaman. Apabila jumlah air yang tersedia di tanah tidak mencukupi bagi kebutuhan tanaman maka tanaman akan mengalami gangguan morfologi dan fisiologis sehingga pertumbuhan dan produktifitasnya akan terhambat, hal ini menyebabkan tanaman mengalami cekaman kekeringan.
Analisis ragam terhadap nilai kadar air tanah menunjukkan terdapat pengaruh nyata (P<0.05) hanya dari faktor tunggal penyiraman, sedangkan faktor tunggal mikoriza serta interaksi penyiraman dan mikoriza tidak berpengaruh nyata (Lampiran 1). Tabel 4.1 menunjukkan pengaruh faktor penyiraman dan inokulasi mikoriza terhadap nilai kadar air tanah yang diuji.
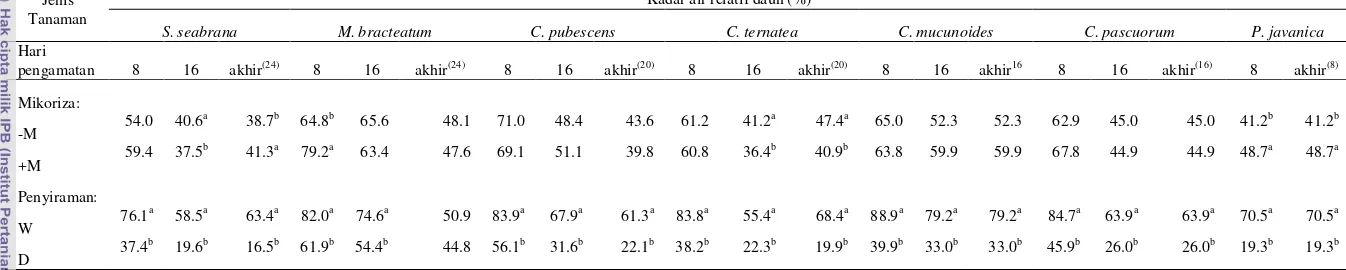
Tabel 4.1 Rataan kadar air tanah (%) pada tanaman legum pohon fase pertumbuhan awal yang diinokulasi dan tanpa inokulasi mikoriza pada dua perlakuan penyiraman
Jenis Tanaman Kadar air tanah (%)
L. leucocephala I. zollingeriana Desmodium sp Hari
pengamatan 8 16 Akhir(28) 8 16 Akhir(20) 8 16 Akhir(16)
Mikoriza:
-M 33.6 31.3 31.7 35.8 31.3 30.5 33.1 30.0 30.0
+M 34.4 30.7 33.5 34.3 30.5 29.0 30.3 27.4 27.4
Penyiraman:
W 36.1 37.7a 40.6a 42.9a 36.7a 36.3a 37.8a 34.8a 34.8a
D 31.9 24.3b 24.6b 27.2b 25.1b 23.3b 25.6b 22.6b 22.6b Keterangan: a, b pada kolom yang sama menunjukkan hasil yang berbeda nyata (P<0.05)
-M = tanpa mikoriza, +M = diinokulasi FMA, W = disiram, D = tidak disiram.
Hasil penelitian menunjukkan bahwa perlakuan cekaman kekeringan menurunkan kadar air tanah berkisar antara 11 - 39% dibandingkan dengan kondisi tanpa cekaman. Hasil serupa juga diperoleh pada 4 jenis Turfgrasses (Midlawn bermudagrass, Prairie buffalograss, Meyer zoysiagrass dan Mustang tallfescue) yang mendapatkan bahwa kandungan air tanah pada hari ke-25 cekaman kekeringan mengalami penurunan sebanyak 78% (kandungan air tanah menjadi 8%) pada Mustang tallfescue (Qian dan Fry, 1997).
memungkinkan air tanah dapat masuk ke dalam sel tanaman (Yoshiba et al., 1997; Kramer dan Boyer, 1995; Taiz dan Zeiger, 2002).
Potensial air daun (PAD)
Aliran air terjadi dari potensial air yang tinggi ke potensial air yang rendah, saat potensial air tanah rendah akibat cekaman kekeringan maka tanaman akan merespon dengan lebih menurunkan potensial airnya sehingga air dari tanah dapat diserap oleh tanaman (Kramer dan Boyer, 1995). Analisis ragam terhadap potensial air daun menunjukkan bahwa terdapat pengaruh nyata (P<0.05) dari faktor penyiraman, faktor mikoriza (L.leucocephala dan I. zollingeriana), serta interaksi faktor mikoriza dan penyiraman (L.leucocephala dan I. zollingeriana) (Lampiran 2). Rataan potensial air daun pada tanaman legum pohon fase pertumbuhan awal ditunjukkan pada Tabel 4.2.
Tabel 4.2 Rataan potensial air daun (MPa) pada tanaman legum pohon fase pertumbuhan awal yang diinokulasi dan tanpa inokulasi mikoriza pada dua perlakuan penyiraman
Jenis Tanaman Potensial air daun (Mpa)
L. leucocephala I. zollingeriana Desmodium sp Hari
pengamatan 8 16 Akhir(28) 8 16 Akhir(20) 8 16 Akhir(20)
Mikoriza:
-M -1.3 -2.8 -4.8b -2.3b -2.4 -3.8a -2.1 -5.3 -4.7
+M -1.4 -2.9 -3.4a -1.8a -3.8 -6.0b -0.8 -5.9 -4.8
Penyiraman:
W -1.1a -1.7a -1.9a -1.5a -1.3a -1.8a -1.3 -1.5a -1.3a
D -1.6
b
-4.0b -6.3b -2.6b -4.9b -7.9b -1.6 -9.7b -8.2b
Keterangan: a, b pada kolom yang sama menunjukkan hasil yang berbeda nyata (P<0.05) -M = tanpa mikoriza, +M = diinokulasi FMA, W = disiram, D = tidak disiram.
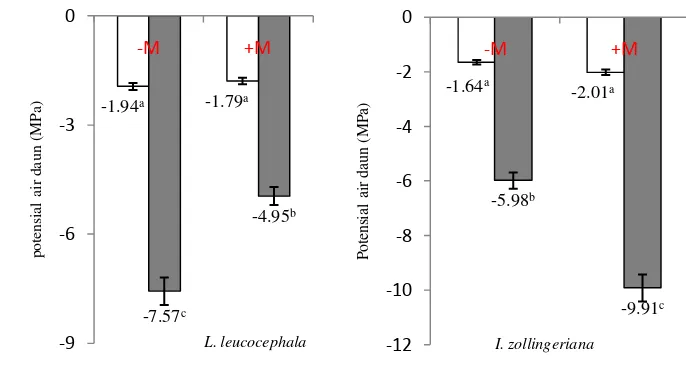
Tabel 4.2 menunjukkan bahwa secara umum perlakuan cekaman kekeringan menurunkan potensial air daun pada ke-3 jenis tanaman legum pohon fase pertumbuhan awal. Berbeda dengan pengaruh cekaman kekeringan, pemberian mikoriza dapat meningkatkan atau bahkan menurunkan potensial air daun yang diuji. Faktor mikoriza berpengaruh nyata pada L. leucocephala (pengamatan hari terakhir) dan I. zollingeriana (pengamatan hari ke-8 dan hari terakhir). Pada L. leucocephala, nilai potensial air daun tanaman yang diinokulasi dengan mikoriza lebih tinggi 29% dibandingkan dengan tanaman tanpa mikoriza. Hal ini diduga karena mikoriza efektif dalam membantu penyerapan air tanah oleh akar sehingga tanaman tidak perlu menurunkan potensial air daun menjadi semakin negatif. Pada tanaman Hasil serupa juga didapatkan oleh Sanchez Blanco et al. (2004) yaitu cekaman kekeringan pada tanaman Rosmarinus officinalis menurunkan potensial air daun pada tanaman yang diinokulasi dengan mikoriza dan juga tanaman tanpa mikoriza, tetapi penurunan ini lebih rendah pada tanaman tanpa mikoriza.
cekaman kekeringan didapatkan nilai potensial air daun tanaman yang diinokulasi dengan mikoriza lebih rendah (-6 MPa) dibandingkan dengan tanaman tanpa mikoriza (-3.8 MPa). Hasil ini diduga terkait dengan adanya kecenderungan peningkatan prolin daun pada tanaman yang diinokulasi dengan mikoriza pada hari terakhir pengamatan cekaman kekeringan (Tabel 4.4). Penurunan potensial air yang semakin negatif tersebut diasumsikan karena adanya mikoriza memperkuat mekanisme penyesuaian osmotik pada tanaman, ditunjukkan dengan adanya peningkatan prolin daun.
Faktor penyiraman berpengaruh nyata terhadap potensial air daun pada ke-3 jenis tanaman legum pohon. Nilai potensial air daun pada ke-3 jenis tanaman legum pohon yang diteliti mengalami penurunan seiring dengan semakin lamanya cekaman kekeringan. Hal ini ditunjukkan dengan nilai potensial air daun pada hari ke-8, dan ke-16 pengamatan berturut-turut dengan kisaran -1.6 MPa sampai -2.6 MPa dan -4.0 MPa sampai -9.7 MPa (Tabel 4.2). Pada hari terakhir pengamatan cekaman kekeringan terjadi penurunan potensial air daun sebesar 3-4x lipat dibandingkan dengan kondisi normal tanpa cekaman.
Beberapa penelitian juga mendapatkan nilai potensial air semakin negatif seiring dengan semakin lamanya durasi cekaman kekeringan seperti Ludlow (1989) yang memperoleh nilai potensial air daun lethal -12 MPa pada beberapa rumput C4. Fageria et al. (2010) menyata