PERSEBARAN DAN NERACA KEHIDUPAN KUTUDAUN AKAR
PADI Rhopalosiphum rufiabdominalis (Sasaki) DAN Tetraneura
nigriabdominalis (Sasaki) (HEMIPTERA: APHIDIDAE)
DI JAWA BARAT
HARLENI
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA
Dengan ini saya menyatakan bahwa tesis berjudul Persebaran dan Neraca Kehidupan Kutudaun Akar Padi Rhopalosiphum rufiabdominalis (Sasaki) dan
Tetraneura nigriabdominalis (Sasaki) (Hemiptera: Aphididae) di Jawa Barat adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, Juni 2016
Harleni
RINGKASAN
HARLENI. Persebaran dan Neraca Kehidupan Kutudaun Akar Padi
Rhopalosiphum rufiabdominalis (Sasaki) dan Tetraneura nigriabdominalis
(Sasaki) (Hemiptera: Aphididae) di Jawa Barat. Dibimbing oleh PURNAMA HIDAYAT dan HERMANU TRIWIDODO.
Kutudaun merupakan serangga fitofag yang umum ditemukan di bagian tanaman di atas tanah seperti daun, bunga, buah, tunas dan ranting. Namun ada beberapa kutudaun yang hidup pada akar tanaman diantaranya Rhopalosiphum
rufiabdominalis (Sasaki) dan Tetraneura nigriabdominalis (Sasaki). Kutudaun
tersebut mendapatkan nutrisi dari akar tanaman padi. Informasi dasar mengenai sebaran dan sejarah kehidupan kutudaun tersebut di Jawa barat masih sedikit diketahui. Penelitian ini bertujuan untuk mengetahui persebaran dan neraca kehidupan kutudaun pada akar padi di Jawa Barat.
Pengambilan sampel untuk studi persebaran dilakukan pada pertanaman padi di beberapa kabupaten di Jawa Barat. Kutudaun yang digunakan untuk penelitian neraca kehidupan diambil dari sawah di Leuwiliang Bogor. Identifikasi spesies dan penelitian neraca kehidupan dilakukan di Laboratorium Biosistematika Serangga, Departemen Proteksi Tanaman, Fakultas Pertanian, Institut Pertanian Bogor. Penelitian dilakukan sejak Juni 2014 sampai dengan Juli 2015. Pengambilan sampel dilakukan dengan metode purposive sampling pada lahan pertanaman padi kering dengan cara pencabutan rumpun padi dan dilihat bagian akarnya. Jika kutudaun ditemukan maka pada bagian batang padi dekat akar dipotong dan dimasukan ke dalam kotak pemeliharaan, diberi label lokasi dan tanggal pengambilan sampel. Sampel yang diambil sebanyak sepuluh rumpun padi untuk setiap lahan pengamatan.
Tanaman padi yang digunakan untuk pemeliharaan kutudaun di laboratorium adalah ratun padi varietas Ciherang yang berasal dari sawah Situgede, Bogor. Panjang akar ratun padi yang digunakan adalah 4 cm (3 cm di bawah pangkal akar + 1 cm di atas pangkal akar). Masing-masing spesies yaitu 2 imago R. rufiabdominalis dan 2 imago T. nigriabdominalis dimasukkan ke dalam 5 cawan petri yang berisi 3 akar ratun padi untuk memperoleh nimfa instar 1. Sebanyak 60 ekor nimfa instar 1 diepilihara di akar ratun padi. Selanjutnya diamati kematian serta perkembangannya. Data hasil pengamatan individu dicatat setiap hari untuk analisis neraca kehidupan (life table).
Hasil penelitian ini menunjukan bahwa kutudaun R. rufiabdominalis memiliki siklus hidup mulai dari menjadi nimfa instar 1 hingga melahirkan nimfa instar 1 untuk pertama kalinya adalah 4.98 hari. Lama hidupnya sejak menjadi imago hingga imago tersebut mati adalah 15.94 hari. Keperidiannya jumlah nimfa yang dilahirkan oleh setiap imago selama hidupnya adalah 67.44 nimfa. Sedangkan siklus hidup, lama hidup dan keperidian T. nigriabdominalis masing-masing adalah 5.25 hari, 18.11 hari, dan 11.11 nimfa. Kedua spesies kutudaun memiliki 4 instar pada nimfanya. Hasil kajian neraca kehidupan menunjukan bahwa R. rufiabdominalis memiliki tingkat pertambahan intrinsik (r) 0.46 individu/induk/hari dan waktu penggandaan populasi menjadi dua kali lipat (DT) 1.51 hari, sedangkan T. nigriabdominalis memiliki tingkat pertambahan intrinsik (r) 0.16 individu/induk/hari dan waktu penggandaan populasi menjadi dua kali lipat (DT) 4.33 hari. Berdasarkan analisis neraca kehidupan menunjukkan bahwa populasi R. rufiabdominalis lebih cepat tumbuh dibandingkan dengan populasi T.
nigriabdominalis pada akar tanaman padi. Kedua spesies kutudaun ini memiliki
siklus hidup yang pendek dan keperidian yang tinggi, sehingga berpotensi menjadi hama penting pada pertanaman padi di daerah kering
SUMMARY
HARLENI. Distribution and Life Table Rice Root Aphids Rhopalosiphum rufiabdominalis (Sasaki) and Tetraneura nigriabdominalis (Sasaki) (Hemiptera: Aphididae) in West Java. Supervised by PURNAMA HIDAYAT and HERMANU TRIWIDODO.
Aphids are phytopagous insects which usually live on plant parts above the ground, such as leaves, flowers, fruits, shoots, and branches. However, several aphids have been reported live on rice roots, such of Rhopalosiphum rufiabdominalis (Sasaki) and Tetraneura nigriabdominalis (Sasaki). These aphids obtain the nutrition from rice root. The basic information about the rice root aphids are limited. This study was aimed to know the distribution and life table of rice root aphids from West Java.
Distribution study of rice root aphids were conducted on rice fields in saveral districts in West Java. Rice root aphids used for life table study were collected from rice field in Lewiliang, Bogor. Spesies identification and life table study were conducted in the Insect Biosystematic Laboratory, Plant Protection Department, Faculty of Agriculture, IPB. Research was conducted from June 2014 until July 2015. Sampling was done by purposive sampling method on dried rice field by pulling off some clumps of rice hills to observe the presence of aphids on the rice root. The rice roots that contained aphids were cropped near steam base, labeled, and then placed into rearing boxs. Ten rice hills were sampled and collected for each observation field.
Aphids rearing were done on the root of Ciherang variety rice ratoons collected from Situgede Village, West Bogor, and were put into a petri dish. The length of the rice ratoons used for rearing were 4 cm (3 cm below the root base + 1 cm above the root base). This experiment used five petri dishes, that contained two adults of aphids and three rice ratoons root to obatain the same age first instar nymphs. As many as 60 first instar nymphs were reared in the root of rice ratoons.The mortality and fecundity of aphids were observed every day until all aphids dead. This data were used to analysis the life table of aphids.
The results of identification showed that there were two spesies of aphids that infested rice root collected from West Java namely R. rufiabdominalis and T. ngriabdominalis. Both species were collected from the root of Ciherang, Cisadane, IR-64, and Rojelele rice varieties. Aphid R. rufiabdominalis and T. ngriabdominalis had been spread in several areas West Java, included Bandung, Bogor, Cianjur, Cirebon, Garut, Karawang, Sukabumi, and Tasikmalaya Districts. The absence of natural border among provinces and similar rices varieties that planted in West Java and other Java areas, caused possibility of both aphids species can be found in another Java areas.
intrinsic rate of increase (r) and doubling time for it‟s population (DT) was 0.46 individual per parent per day and 1.50 days, respectively while for T.
nigriabdominalisintrinsic rate of increase (r) and doubling time for its population
(DT) was 0.14 individual per parent per day and 4.99 days respectively. Based on the life table analysis, the population of R. rufiabdominalis was growing faster than the population of T. nigriabdominalis on rice root. Both aphids species of the have short life cycle and high fecundity, thereby they are potentially to become important pests on rice plant in dry areas.
© Hak Cipta Milik IPB, Tahun 2016
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB.
13
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains
pada
Program Studi Entomologi
PERSEBARAN DAN NERACA KEHIDUPAN KUTUDAUN
AKAR PADI Rhopalosiphum rufiabdominalis (Sasaki) DAN
Tetraneura nigriabdominalis (Sasaki) (HEMIPTERA:
APHIDIDAE) DI JAWA BARAT
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
BOGOR 2016
PRAKATA
Puji dan syukur penulis panjatkan kepada Allah Subhanahu Wa Ta’ala atas segala karunia-Nya sehingga penelitian ini berhasil diselesaikan. Tesis yang berjudul “Persebaran dan Neraca Kehidupan Kutudaun Akar Padi
Rhopalosiphum rufiabdominalis (Sasaki) dan Tetraneura nigriabdominalis
(Sasaki) (Hemiptera: Aphididae) di Jawa Barat” ini merupakan salah satu syarat untuk memperoleh gelar Magister Sains pada Sekolah Pascasarjana Institut Pertanian Bogor.
Terima kasih penulis ucapkan kepada Dr Ir Purnama Hidayat, MSc sebagai Ketua Komisi Pembimbing dan Dr Ir Hermanu Triwidodo, MSc sebagai Anggota Komisi Pembimbing yang telah banyak memberikan pengarahan, bimbingan, saran, motivasi dan masukan selama penelitian, terimakasih juga kepada Dr Ir Ali Nurmansyah, MSi sebagai dosen komisi luar dan Dr Ir Pudjianto, MSi sebagai ketua Program Studi Entomologi. Selain itu, penulis juga menyampaikan ucapan terimakasih kepada Ayahanda A. Dt. Marajo dan Ibunda Khairanis atas doa tulus ikhlas, perjuangan dan pelajaran hidup yang sangat berharga kepada penulis serta uncu Ratna Sari Dewi, Kakanda Insan Kamil Muhammad, Khairusil, Adinda Hardinel. Penulis juga mengucapkan terimakasih kepada Ibu Aisyah, Mba Atiek, Rekan-rekan di laboratorium Biosistematika Serangga Irfan, Ciptadi, Mba Yani, Heri, Herni, Mba Hapsah, Mba Dika, Suryadi, Nia dan teman-teman Entomologi 2013 Mas Ichsan, Laila, Dila, Ihsan. Terimakasih juga kepada Gunawan wibisono, Ilma Satriana Dewi dan teman-teman lain yang tidak dapat penulis tuliskan satu persatu yang telah membantu dan mendukung penulis dalam menyusun karya tulis ini.
Semoga hasil penelitian ini bermanfaat.
Bogor, Juni 2016
17 Taksonomi dan Morfologi Kutudaun di Akar Padi 2 Biologi Kutudaun 4
DAFTAR TABEL
1. Jumlah Kutudaun yang Ditemukan pada Masing-masing Kabupaten 10
2. Biologi R. rufiabdominalis dan T. nigriabdominalis 13
3. Neraca Kehidupan R. rufiabdominalis dan T. nigriabdominalis 16
4. Biologi R. rufiabdominalis di Batang dan Akar Daun Padi 17
5. Neraca Kehidupan R. rufiabdominalis di Akar dan Batang Padi 19
DAFTAR GAMBAR
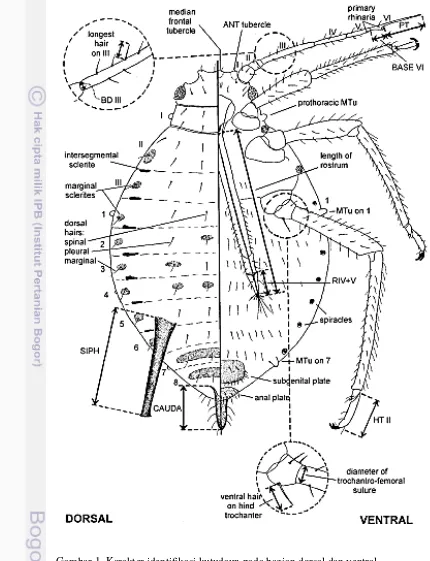
1. Karakter Identifikasi Kutudaun pada Bagian Dorsal dan Ventral 32. Peta Lokasi Pengambilan Sampel 6
3. Kutudaun Pada Akar Padi 8
4. Gejala yang di Timbulkan oleh Kutudaun di Akar 11
5. Morfologi Kutudaun 12
6. Karakter Morfologi R. rufiabdominalis 12
7. Karakter Morfologi T. nigriabdominalis 13
8. Peluang Hidup dan Rataan Keperidian Harian R. rufiabdominalis dan T. nigriabdominalis 15
9. Peluang Hidup dan Keperidian Harian R. rufiabdominalis pada Akar dan Batang 18
DAFTAR LAMPIRAN
1. Neraca kehidupan R. rufiabdominalis pada akar padi 272. Neraca kehidupan T. nigriabdominalis pada akar padi 28
3. Neraca kehidupan R. rufiabdominalis pada akar padi 29
PENDAHULUAN
Latar Belakang
Kutudaun umumnya hidup pada bagian tanaman yang berada di atas permukaan tanah seperti pucuk, daun, ranting, bunga, dan buah. Namun, beberapa spesies kutudaun dapat hidup pada akar tanaman di dalam tanah. Spesies kutudaun yang diketahui hidup di bagian akar tanaman padi adalah
Rhopalosiphum rufiabdominalis (Sasaki) dan Tetraneura nigriabdominalis
(Sasaki) (Pike et al. 1990; Kindler et al. 2004).
Persebaran kutudaun akar padi belum banyak dilaporkan di Indonesia, sehingga perlu dilakukan penelitian untuk melihat persebarannya. Di Indonesia Kalshoven (1950) melaporkan bahwa kutudaun yang menyerang akar padi adalah
Byrsocrypta hirsuta Bak yang ditemukan di Pulau Jawa. Rahmah (2013) melaporkan bahwa T. nigriabdominalis ditemukan pada akar padi di Desa Cijedil, Kecamatan Cugenang, Kabupaten Cianjur. Persebaran T. nigriabdominalis meliputi berbagai negara yaitu Afrika, Amerika, Australia, Bangladesh, Fiji, Indonesia, Itali, Jepang, Korea, Laos, Malaysia, Nepal, New Zealand, Pakistan, Philipina, Sri Lanka, Slovenia, dan Tonga (Pathak dan khan 1994; Galli dan Manicardi 1998; Saito et al. 2006; Modic et al. 2011)
Kutudaun ini memperoleh nutrisi dengan cara menghisap bagian dari akar padi. Kutudaun ini hidup berkoloni pada setiap rumpun padi. Populasi kutudaun yang tinggi pada akar padi dapat menyebabkan kerugian secara ekonomi. Informasi mengenai kutudaun akar padi belum banyak dilaporkan di Indonesia, sehingga diperlukan adanya penelitian mengenai pertumbuhan, perkembangan, keperidian, dan kematian kutudaun tersebut masih sangat terbatas.
Peluang hidup dan keperidian kutudaun di akar padi dapat diketahui dengan metode neraca kehidupan. Neraca kehidupan merupakan suatu metode untuk menggambarkan dan mengukur kematian dalam populasi (Price 1995). Data yang diperoleh dari metode neraca kehidupan dapat digunakan sebagai informasi dasar yang dibutuhkan dalam upaya pengendalian populasi kutudaun di akar padi. Dalam hal ini, neraca kehidupan dapat membantu untuk memutuskan teknik pengendalian yang sesuai dengan mengetahui strategi kehidupan dari hama tersebut.
Tujuan Penelitian
Penelitian ini bertujuan mengetahui persebaran dan neraca kehidupan kutudaun akar padi di Jawa Barat.
Manfaat Penelitian
Taksonomi dan Morfologi Kutudaun Akar Padi
Kutudaun termasuk ke dalam golongan Phytopthires atau kutu tanaman (Saragih 1994). Borror dan Delong (1971) menggolongkan kutudaun tersebut ke dalam Ordo Hemiptera, Subordo Sternorryncha, Superfamili Aphidoidea, Famili Aphididae. Menurut Blackman dan Eastop (2000), Super Famili Aphidoidea terdiri dari tiga famili yaitu, Adelgidae, Phylloxeridae, dan Aphididae. Famili Aphididae terdiri atas delapan subfamili di antaranya Erisomatinae, Hormaphidinae, Anoeciinae, Calaphidinae, Chaitophorinae, Greenideinae, Aphidinae, dan Lachininae. antena, rostrum, kauda, dan tungkai biasanya mempunyai rambu-rambut halus (Gambar 1) Blackman dan Eastop (2006).
Kutudaun T. nigriabdominalis masuk ke dalam famili Aphididae, subfamili Eriosmatinae, tribe Eriosomatini (Blackman dan Eastop 2006). Imago T. nigriabdominalis berwarna jingga dengan ukuran tubuh 1.8 mm dengan bentuk tubuh bulat. Karakter T. nigriabdominalis adalah memiliki antena pendek kurang dari 0.2 kali panjang tubuhnya dengan terminal proses lebih pendek dari segmen dasar antena, terdapat 10 helai rambut pada segmen antena yang ketiga, memiliki sifunkuli yang pendek dan agak gelap dibandingkan tubuhnya. Sifunkuli membentuk seperti pori atau kerucut, tidak memiliki pola polygonal retikulasi, sedikit membengkak, tarsi 1 ruas dan memiliki rostrum yang pendek dan berambut.
Kutudaun R. rufiabdominalis masuk ke dalam famili Aphididae, subfamili Aphidinae, tribe Aphidini (Blackman dan Eastop 2006). Imago R. rufiabdominalis
berwarna hijau kecoklatan dengan ukuran tubuh 1.7 mm. Karakter R. rufiabdominalis adalah memiliki kepala hitam dengan mata merah gelap hampir hitam. Rostrum bagian pangkal ke tengah hijau pucat, bagian tengah ke ujung kehitaman, antena hitam, toraks hitam, vena sayap depan coklat kehitaman dengan tepi coklat, kauda hijau gelap, pangkal femur dan tibia coklat terang, tarsus hitam dan bagian tengah tubuh ke ujung abdomen kehitaman sampai hitam (Blackman dan Eastop 2000).
3
Biologi Kutudaun
Di negara empat musim kutudaun berkembangbiak dengan cara seksual (kawin) dan aseksual (tidak kawin). Pada musim gugur populasi terdiri dari individu berkelamin jantan dan betina. Kutudaun betina meletakkan telur yang akan menetas di musim semi dan menjadi serangga betina. Pada musim semi dan panas semua serangga adalah betina dan berkembangbiak secara partenogenetis. Pada musim gugur berikutnya akan timbul lagi individu yang berkelamin jantan dan perkembangbiakan secara kawin. Kutudaun ini bereproduksi secara vivipar partenogenesis. Perkembangan kutudaun terdiri atas empat instar yaitu masing-masing instar 1-4 adalah 2.2, 2.7, 3.8, dan 5.7 hari. Kutudaun dewasa hidup 15 sampai 20 hari dan menghasilan 35 sampai 45 nimfa. Nimfa biasanya dilahirkan tidak lama setelah kutudaun mencapai tahap dewasa (Pathack dan khan 1994).
Reproduksi kutudaun di Indonesia (daerah tropis) dengan aseksual yaitu partenogenesis dan vivipar. Nimfa yang baru lahir dapat berkembang cepat menjadi imago dan siap melahirkan nimfa baru. Bentuk dewasa dari kutudaun di akar ada yang bersayap dan tidak bersayap. Kutudaun akar padi yang dewasa bersayap dengan panjang 1.5 sampai 2.5 mm dan bentuk yang tidak bersayap dengan panjang 1.5 sampai 2.3 mm. Serangga bersayap dihasilkan apabila terdapat jumlah populasi yang berdesakan pada saat populasi memuncak (Kalshoven 1981).
Nilai Ekonomi
Kerusakan tanaman akibat serangan kutudaun dapat terjadi secara langsung maupun tidak langsung. Kerusakan secara langsung berkisar antara 6-25%. Kerusakan secara tidak langsung dapat sebagai vektor yang menyebabkan kerugian mencapai lebih dari 80% (Miles 1987).
Imago dan nimfa T. nigriabdominalis & R. rufiabdominalis mengambil cairan tanaman yang mengakibatkan warna daun menjadi kuning dan tanaman menjadi kerdil (Pathack dan Khan 1994; Shepard 1995). Kehilangan hasil produksi padi yang disebabkan oleh kutudaun tersebut di Jepang mencapai 50% (Heinrichs dan Barrion 2004). Di India T. nigriabdominalis merupakan spesies umum yang menyerang akar padi pada persemaian sampai musim tanam. Populasi kutudaun ini sangat tinggi pada padi varietas IR8 (Heinrichs dan Barrion 2004). Meskipun beberapa spesies kutudaun akar padi dilaporkan sebagai vektor virus
Tobacco Vein-banding Virus (TBMV), tetapi T. nigriabdominalis tidak menularkan virus tersebut berdasarkan hasil percobaan di laboratorium. Jedlinsky (1981) dan Chapin (2001) melaporkan bahwa R. rufiabdominalis merupakan vektor Barley Yellow Dwarf Virus pada tanaman gandum yang mengakibatkan kehilangan hasil sebesar 20.1% di Carolina Selatan.
Neraca Kehidupan
5
mortalitas dan kelangsungan hidup suatu populasi. Pertumbuhan populasi suatu spesies dapat diketahui dengan melihat jumlah keturunan yang dihasilkan seekor betina pada interval umur selama hidupnya, dan jumlah individu pada setiap interval umur tersebut. Untuk mengetahui hal tersebut, pendekatannya dapat disederhanakan dengan hanya mengetahui betina saja dalam satu populasi (Price 1995). Informasi tersebut merupakan informasi dasar yang diperlukan dalam menelaah perubahan kepadatan dan laju pertumbuhan atau penurunan suatu populasi. Tabel kehidupan berisi nilai-nilai berikut (Begon et al. 2008):
1. x adalah titik tengah umur kohort (hari, minggu, bulan dll); 2. lx adalah peluang hidup setiap individu pada umur x;
3. mx adalah rata-rata jumlah keturunan yang dihasilkan oleh serangga betina
pada umur x;
4. lxmxadalah banyaknya anak yang dilahirkan pada kelas umur x.
Parameter neraca kehidupan yang digunakan untuk melihat hubungan preferensi kutudaun terhadap tanaman yang diujikan adalah lama stadia pradewasa, lama stadia imago dan keperidian imago betina. Parameter demografi yang dihitung Birch (1948) meliputi :
1. Laju reproduksi bersih (R0) dihitung dengan rumus:
R0 = ∑lxmx
2. Laju pertambahan intrinsik (r) dihitung dengan rumus: r = ln R0/ T
3. Rataan masa generasi (T) dihitung dengan rumus: T = ∑xlxmx/∑lxmx
4. Populasi berlipat ganda (DT) dihitung dengan rumus: DT = ln (2/r)
Laju reproduksi bersih (R0) merupakan rata-rata jumlah keturunan betina
Tempat dan Waktu Penelitian
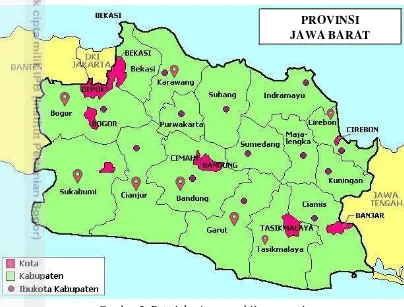
Pengambilan sampel dilakukan pada pertanaman padi di beberapa kabupaten di Jawa Barat yaitu Bogor, Garut, Sukabumi, Tasikmalaya, Bandung, Cirebon, Karawang dan Cianjur (Gambar 2). Kutudaun untuk pengamatan neraca kehidupan diambil dari sawah di Leuwiliang Bogor. Identifikasi morfologi serta pengamatan neraca kehidupan dilakukan di Laboratorium Biosistematika Serangga, Departemen Proteksi Tanaman, Fakultas Pertanian, Institut Pertanian Bogor. Penelitian dilakukan sejak bulan Juni 2014 sampai dengan Juli 2015.
Gambar 2 Peta lokasi pengambilan sampel
Alat dan Bahan
Alat-alat yang digunakan antara lain mikroskop nikon SMZ-U ZOOM 1:10, mikroskop stereo, mikroskop cahaya, kamera Dino-eye, GPS, objek glass, cover glass, hotplate fisher scientific slide warmer, wadah plastik, tisu, plastik bening, kuas, jarum mikro, cawan petri, alat tulis, kamera digital, hand-counter, alkohol (50, 70, 80, 95 dan 100 %), larutan KOH 10%, balsam kanada dan minyak cengkeh.
7
Pengambilan Sampel Kutudaun
Pengambilan sampel dilakukan dengan metode purposive sampling pada suatu lahan pertanaman padi kering. Pada lahan tersebut dilakukan pencabutan rumpun padi, kemudian diamati bagian akarnya. Jika terdapat kutudaun maka pada bagian batang padi dekat akar dipotong dan dimasukan ke dalam kotak pemeliharaan, diberi label lokasi dan tanggal pengambilan sampel. Sampel yang diambil sebanyak sepuluh rumpun padi.
Setiap rumpun padi yang dicabut diperhatikan keberadaan kutudaun dengan mengukur jarak antara kutudaun dari pangkal akar keberadaan kutudaun tersebut. Kelimpahan populasi kutudaun dilakukan dengan cara menghitung jumlah kutudaun pada semua tanaman contoh. Selain itu, diamati dan dicatat fase tanam dan varietas yang ada pada masing-masing plot pengambilan sampel. Posisi geografis dan ketinggian lokasi survei diukur dengan menggunakan bantuan (Global Positioning System) GPS.
Pembuatan Preparat Slide Kutudaun
Pembuatan preparat slide kutudaun dilakukan dengan metode preparat permanen yang bertujuan mempermudah proses identifikasi dan penyimpanan dalam waktu yang lama. Spesimen yang digunakan dalam pembuatan preparat slide kutudaun adalah fase imago (dewasa).
Imago kutudaun dipilih berdasarkan kelengkapan bagian tubuh (antena, kepala, tungkai, sifunculi, dan kauda). Kutudaun direbus dalam tabung reaksi yang sudah diisi alkohol 95% selama tiga menit. Spesimen yang telah direbus diletakkan di cawan petri, kemudian bagian abdomennya ditusuk dengan jarum mikro untuk mengeluarkan cairan tubuhnya. Kemudian spesimen direbus dalam tabung reaksi berisi KOH 10% hingga berwarna bening (transparan). Setelah itu spesimen diletakkan di dalam cawan petri dan ditekan bagian abdomennya untuk mengeluarkan sisa isi bagian tubuhnya.
Spesimen yang telah direbus kemudian dicuci dengan akuades sebanyak dua kali. Selanjutnya direndam dengan alkohol secara bertahap dimulai dari alkohol 50%, 80%, 95% dan terakhir dengan alkohol 100% masing-masing selama 10 menit. Perendaman ini bertujuan agar spesimen tidak mengkerut. Kemudian spesimen direndam dengan minyak cengkeh selama 10 menit untuk menghilangkan kadar air yang masih tersisa di tubuh spesimen selanjutnya spesimen diletakkan di atas kaca objek untuk ditata atau dilakukan perentangan, sehingga bisa terlihat jelas bagian-bagian tubuhnya. Kemudian kanada balsam diteteskan secara perlahan dari bagian atas spesimen agar tidak merusak posisinya, lau dibiarkan beberapa hingga kemudian spesimen ditutup dengan
cover glass. Preparat yang telah selesai dikeringkan di dalam oven pengering serangga atau pada Hotplate fischer scientific slider warmer selama dua minggu.
Identifikasi Kutudaun
dihubungkan dengan sebuah komputer PC. Identifikasi dilakukan hingga tingkat spesies.
Karakteristik Spesies Kutudaun
Karakter setiap spesies dari kutudaun digambar dengan menggunakan mikroskop Nikon SMZ-U ZOOM 1:10 sehingga terbentuk gambar yang mirip dengan karakter yang diinginkan. Karakter yang digambar adalah karakter spesifik yang dimiliki oleh setiap spesies. Karakter tersebut dapat digunakan sebagai pembeda antar spesies.
Pemeliharaan Kutudaun untuk Pengamatan Neraca Kehidupan
Kutudaun di akar padi yang diperoleh dari sawah di Leuwiliang Bogor dimasukkan ke dalam kotak pemeliharaan yang berisi tanaman padi untuk perbanyakan. Kotak pemeliharaan yang digunakan berupa wadah plastik berdiameter 10 cm dengan tinggi 8 cm.
Tanaman padi yang digunakan untuk pemeliharaan adalah ratun padi Varietas Ciherang yang berasal dari sawah Situgede, Bogor. Panjang akar ratun padi yang digunakan adalah 4 cm (3 cm di bawah pangkal akar + 1 cm di atas pangkal akar) (Gambar 3). Sebanyak dua imago dari masing-masing spesies R. rufiabdominalis dan T. nigriabdominalis dimasukkan ke dalam cawan petri yang berisi akar ratun padi. Perlakuan ini diulang sebanyak lima kali.
Populasi kohort merupakan sejumlah individu yang memiliki umur seragam. Nimfa instar 1 R. rufiabdominalis dan T. nigriabdominalis dalam populasi kohort diinfestasi pada akar padi ratun. Nimfa instar 1 kutudaun tersebut diinfestasikan masing-masing ke dalam 60 wadah plastik berdiameter 5 cm dengan tinggi 4.8 cm. Setiap wadah berisi satu nimfa kutudaun.
Gambar 3 Kutudaun pada akar padi: (a) koloni yang diperoleh dari lapang (b) Individu pada akar ratun padi di wadah pemeliharaan
Pengamatan Pemeliharaan Kutudaun di Akar, Batang dan Daun Padi
Pengamatan ini bertujuan untuk melihat kesesuaian habitat kutudaun pada akar, batang, dan daun padi. Tanaman padi yang digunakan untuk pemeliharaan adalah ratun padi Varietas Ciherang yang berasal dari sawah di Situgede Bogor.
Akar, batang, dan daun padi yang digunakan berukuran 3 cm. Nimfa instar 1
9
kutudaun diinfestasikan ke dalam 15 wadah plastik untuk setiap pakan. Wadah yang digunakan berdiameter 5 cm dengan tinggi 4.8 cm.
Pengamatan Neraca Kehidupan
Peubah yang diamati yaitu lamanya waktu tiap instar, siklus hidup, praoviposisi, lama hidup dan keperidian kutudaun di akar padi. Siklus hidup kutudaun tersebut diamati mulai dari nimfa instar 1 yang diinfestasi hingga melahirkan nimfa instar satu untuk pertama kalinya. Praoviposisi kutudaun diamati dari waktu yang dibutuhkan sejak menjadi imago pertama hingga melahirkan nimfa instar satu untuk pertama kalinya. Lama hidup kutudaun dimulai dari hari pertama menjadi imago hingga imago tersebut mati. Pengamatan keperidian kutudaun dihitung dari banyaknya nimfa yang dilahirkan oleh setiap imago selama hidupnya. Data hasil pengamatan disusun dalam tabel biologi kutudaun di akar padi.
Pengamatan peluang hidup (lx) dilakukan dengan cara menghitung jumlah
individu kutudaun di akar padi yang hidup tiap harinya. Pengamatan keperidian harian (mx) yaitu rata-rata jumlah nimfa kutudaun yang dilahirkan oleh imago
setiap harinya pada umur (x). Data peluang hidup dan keperidian harian dapat digambarkan dalam bentuk kurva dan diperoleh neraca kehidupan.
Neraca kehidupan kohort merupakan neraca kehidupan yang mengikuti perkembangan kohort dimulai dari nimfa instar 1 sampai imago terakhir yang mampu bertahan hidup. Data mengenai pengamatan kohort kutudaun di akar padi disusun dalam tabel neraca kehidupan. Penentuan parameter demografi lainnya dapat ditentukan dengan menggunakan data neraca kehidupan kutudaun tersebut.
Birch (1948) menyatakan bahwa parameter demografi yang dihitung meliputi: 1. Laju reproduksi bersih (R0)
(mx) = keperidian spesifik individu-individu pada kelas umur ke-x
Laju reproduksi bersih (R0) merupakan jumlah individu betina yang akan
Persebaran Kutudaun Akar Padi di Daerah Jawa Barat
Pengambilan sampel kutudaun akar padi dilakukan di beberapa kabupaten Jawa Barat. Terdapat dua jenis kutudaun yang ditemukan pada penelitian ini yaitu
R. rufiabdominalis danT. nigriabdominalis.
Kutudaun R. rufiabdominalis ditemukan di Kabupaten Bandung (Kec Pangalengan), Bogor (Kec. Tenjolaya, Ciampea, Kemang, Cibungbulang, Dramaga, Leuwiliang), Cianjur (Kec. Cipanas, Bocong Picung), Cirebon (Kec. Palimanan), Garut (Kec. Limbangan, Malangbong, Samarang), Kerawang (Kec. Purwasari), Sukabumi (Kec. Sukamulya) dan Tasikmalaya (Kec. Gunung Cupu, Cikangere, Samarang). Kutudaun tersebut ditemukan pada ketinggian tempat antara 12 sampai 1438 mdpl (Tabel 1). R. rufiabdominalis merupakan serangga kosmopolitan pada daerah tropis khususnya di Indonesia (Hill 1971), selain itu daya tahan R. rufiabdominalis terhadap musim dan cekaman lingkungan juga merupakan salah satu penyebab ditemukannya spesies ini pada semua lokasi pengamatan (Shepard et al.1995).
Tabel 1 Jumlah total kutudaun yang ditemukan pada masing-masing kabupaten
Lokasi
Kutudaun R. rufiabdominalis ditemukan pada padi Varietas Ciherang, IR-64, Cisadane dan Rojolele. Jumlah koloni terbanyak ditemukan pada bagian pangkal akar sekitar 1 sampai 5 cm dari permukaan tanah. Koloni yang terbentuk terdiri dari imago dan nimfa. Pada akar yang kedalamannya 6 sampai 10 cm jumlah koloni yang ditemukan lebih sedikit dan di dalam koloni ini ditemukan musuh alami yakni Staphylinidae. Hal ini sama dengan yang dilaporkan Dixon (1985) yang menyatakan bahwa musuh alami menyebabkan kutudaun dimangsa sebelum sempat melahirkan anak atau anak yang dilahirkan dimangsa sebelum dewasa.
11
Limabangan) dan Kerawang (Kec. Purwasari). Kutudaun tersebut ditemukan pada ketinggian tempat antara 12 sampai 600 mdpl (Tabel 1). Kutudaun T. nigriabdominalis ditemukan pada padi varietas Ciherang, IR-64, Cisadane dan Rojolele.
Ketinggian di atas 1300 mdpl yaitu di Pangalengan, T. nigriabdominalis
tidak ditemukan sedangkan R. rufiabdominalis ditemukan, pada daerah dataran rendah, yaitu di Cirebon (12 mdpl) kedua kutudaun ini ditemukan. Hal ini dapat membuktikan bahwa pada tempat dengan ketinggian rendah kutudaun di akar padi dapat berkembangbiak dan melakukan penyebaran. Hill (1971) menyatakan bahwa suhu merupakan pembatas bagi persebaran kutudaun. Semakin tinggi suatu tempat maka suhu pada tempat tersebut akan semakin rendah, maka dapat diasumsikan bahwa T. nigriabdominalis tidak dapat berkembangbiak pada daerah yang memiliki suhu rendah, sedang R. rufiabdominalis masih dapat berkembang biak pada suhu rendah.
Gambar 4 Gejala yang ditimbulkan oleh kutudaun di akar, a) batang kerdil dan daun kekuningan; b) koloni kutudaun T. nigriabdominalis
Kutudaun di akar padi menyerang perakaran yang fungsinya untuk menyerap unsur hara yang akan diolah dan dijadikan sumber makanan bagi tanaman. Serangan yang dilakukan oleh kutudaun akan menimbulkan dampak bagi tanaman padi. Menurut Shepard et al. (1995), kutudaun yang menyerang akar padi menimbulkan gejala kekuningan pada daun padi. Gejala ini timbul apabila populasi kutudaun mencapai puncaknya, sedangkan populasi kutudaun yang ditemukan pada masing-masing tempat pengamatan tidak terlalu banyak, sehingga gejala yang terlihat tidak terlalu signifikan hanya sedikit kekuningan pada batang dan terdapat beberapa tanaman yang terlihat seperti kerdil. Menurut Patack dan Khan (1994), kutudaun di akar umumnya menyebabkan kerusakan berupa perubahan warna dan pertumbuhan tanaman padi yang terganggu (Gambar 4).
Identifikasi Kutudaun
Identifikasi spesies dilakukan untuk memastikan jenis kutudaun yang digunakan dalam penelitian. Berdasarkan hasil identifikasi kutudaun yang ditemukan pada akar padi merupakan spesies R. rufiabdominalis dan T. nigriabdominalis (Gambar 5).
Kalshoven (1950) melaporkan bahwa kutudaun yang menyerang akar padi adalah Byrsocrypta hirsuta Bak yang ditemukan di pulau Jawa. Rahmah (2013)
a
menemukan kutudaun akar padi di Kabupaten Cianjur, Jawa Barat dan setelah di identifikasi ternyata spesies tersebut adalah spesies T. nigriabdominalis. Secara morfologi kutudaun ini menunjukan kesamaan dengan yang dilaporkan oleh Kalshoven.
Gambar 5 Morfologi kutudaun akar padi: (a) R. rufiabdominalis, (b) T. nigriabdominalis
Blackman dan Eastop (2000) juga melaporkan bahwa terdapat beberapa kutudaun akar padi, dua diantaranya adalah R. rufiabdominalis dan T. nigriabdominalis. Spesies R. rufiabdominalis sampai saat ini belum pernah dilaporkan ditemukan pada akar padi di Indonesia.
Imago R. rufiabdominalis dicirikan dengan imago pada bagian toraks berwarna hijau gelap. Abdomen berwarna hijau kecoklatan dengan panjang tubuh 1.7 mm. Bentuk tubuh bulat sedikit memanjang. Antena panjang dengan lima ruas (Gambar 6c), antena dari ruas 3 sampai 5 berambut, terdapat rambut halus pada tubuhnya, sifunkuli berbentuk seperti tabung (Gambar 6b), kauda berbentuk kerucut (Gambar 6a), rostrum panjang (Gambar 6d), tarsus dengan 4 ruas.
Gambar 6 Karakter morfologi dari R. rufiabominalis: (a) bentuk kauda; (b) sifunkuli; (c) ruas antena; (d) rostrum
0.5 mm 0.5 mm
0.2 mm
13
Imago T. nigriabdominalis dicirikan dengan imago berwarna jingga (orange) sampai coklat kekuningan dengan ukuran tubuh 1.8 mm dengan bentuk tubuh bulat. Antena pendek (Gambar 7c), antena ruas ketiga memiliki rambut sebanyak 10 helai. Bagian abdomen terdapat rambut-rambut halus. Sifunkuli pendek berbentuk seperti pori atau kerucut (Gambar 7f). Kauda berbentuk bulat dan berambut 2-5 helai (Gambar 7b). Tarsi satu ruas (Gambar 7d) dan memiliki rostrum yang pendek dan berambut (Gambar 7e).
Gambar 7 Karakter morfologi dari T. nigriabdominalis, (a) bentuk kepala; (b) kauda; (c) ruas antena; (d) tarsus; (e) rostrum; (f) sifunkuli
Biologi R. rufiabdominalis dan T. nigriabdominalis
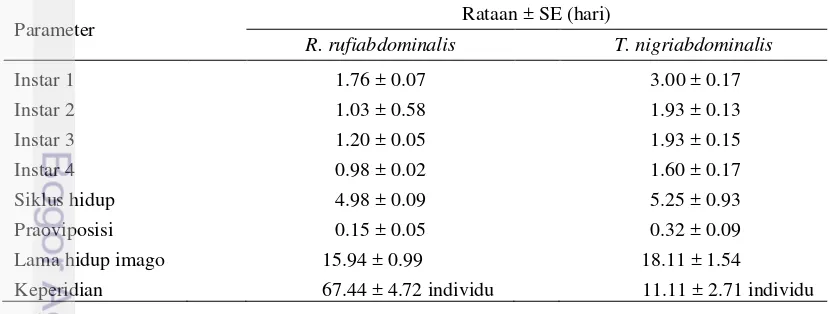
Hasil analisis nilai rataan biologi R. rufiabdominalis berbeda dengan T. nigriabdominalis. Spesies T. nigriabdominalis mengalami perkembangan lebih lama dari pada spesies R. rufiabdominalis (Tabel 2). Siklus hidup R. rufiabdominalis dan T. nigriabdominalis berturut-turut adalah 4.98 hari dan 5.25 hari. Perbedaan siklus hidup serangga menurut Morgan et al. (2001), dipengaruhi oleh spesies serangga, suhu, tanaman inang, serta metode perbanyakan serangga yang digunakan.
Tabel 2 Biologi R. rufiabdominalis dan T. nigriabdominalis di akar padi
Parameter Rataan ± SE (hari)
R. rufiabdominalis T. nigriabdominalis
Instar 1 1.76 ± 0.07 3.00 ± 0.17
Instar 2 1.03 ± 0.58 1.93 ± 0.13
Instar 3 1.20 ± 0.05 1.93 ± 0.15
Instar 4 0.98 ± 0.02 1.60 ± 0.17
Siklus hidup 4.98 ± 0.09 5.25 ± 0.93 Praoviposisi 0.15 ± 0.05 0.32 ± 0.09 Lama hidup imago 15.94 ± 0.99 18.11 ± 1.54
Keperidian 67.44 ± 4.72 individu 11.11 ± 2.71 individu
Lama hidup merupakan selang waktu sejak imago pertama kali muncul hingga imago tersebut mati. Setiap organisme mempunyai variasi jangka hidup yang terbatas. Rata-rata lama hidup imago R. rufiabdominalis dan T. nigriabdominalis berturut-turut adalah 15.94 hari dan 18.11 hari. Lama hidup T. nigriabdominalis lebih panjang dari R. rufiabdominalis tetapi R. rufiabdominalis
memiliki keperidian yang lebih tinggidari T. nigriabdominalis.
Lamanya perkembangan siklus hidup T. nigriabdominalis secara tidak langsung akan berpengaruh terhadap waktu reproduksinya. Semakin lama waktu yang diperlukan untuk mencapai fase imago, maka semakin lama pula waktu untuk bereproduksi. Kozlowski (1992) menyebutkan bahwa tertundanya waktu reproduksi akan berdampak terhadap peningkatan mortalitas sebelum bereproduksi, penurunan masa reproduksi, penurunan output reproduksi dan waktu generasi menjadi lebih lama.
Waktu praoviposisi kedua kutudaun ini berbeda, R. rufiabdominalis lebih cepat dibandingkan T. nigriabdominalis. Cepatnya waktu praoviposisi akan berpengaruh pada banyaknya jumlah keperidian karena penundaan masa praoviposisi akan mengakibatkan proses tertundanya imago untuk melahirkan nimfa. Salah satu faktor eksternal yang mepengaruhi adalah terpenuhinya nutrisi kutudaun. Kuo et al. (2006) melaporkan bahwa nutrisi yang diserap oleh T. nigriabdominalis yang rendah, termasuk nitrogen dapat menyebabkan keperidiannya menurun.
Keperidian kutudaun dihitung dari banyaknya nimfa yang dilahirkan oleh setiap imago selama hidupnya. Semakin banyak nimfa yang dilahirkan imago maka semakin tinggi nilai keperidian. Banyaknya nimfa yang dilahirkan imago memperlihatkan tingkat kesesuaian individu tersebut pada tanaman inangnya. Rataan keperidian imago R. rufiabdominalis dan T. nigriabdominalis berturut-turut adalah 67.44 nimfa dan 11.11 nimfa (Tabel 2). Keperidian imago R. rufiabdominalis lebih tinggi dari keperidian imago T. nigriabdominalis. Menurut Kuswanto dan Budi (2007), menyatakan bahwa meningkatnya jumlah nimfa yang dilahirkan oleh imago kutudaun dapat meningkatkan populasinya secara cepat, terutama dipengaruhi oleh faktor makanan yang tidak terbatas. Selain faktor makanan, menurut Subagyo dan Hidayat (2014), meningkatnya keperidian juga dipengaruhi oleh suhu.
Secara umum faktor-faktor yang memengaruhi biologi kutudaun ini adalah suhu, nutrisi tanaman, umur inang, struktur permukaan inang dan komposisi kimia yang ada pada tanaman (Weathersbee dan Hardee 1994). Selain faktor eksternal, faktor internal juga memberikan pengaruh terhadap biologi kutudaun. Cara adaptasi serangga tersebut terhadap lingkungan sekitar menjadi faktor penting dalam laju siklus hidupnya (Razmjou et al. 2006).
Neraca Kehidupan R. rufiabdominalis dan T. nigriabdominalis
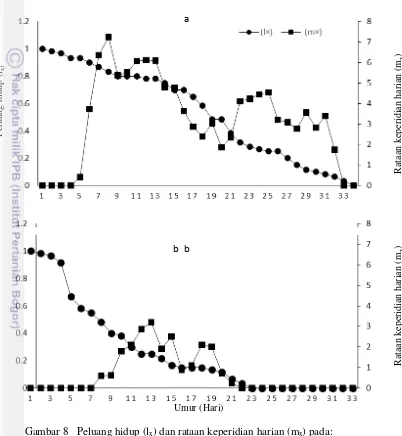
Nilai rataan jumlah nimfa yang dilahirkan oleh imago R. rufiabdominalis
berbeda dengan imago T. nigriabdominalis (Gambar 8). Nilai rataan keperidian harian tertinggi pada R. rufiabdominalis adalah 7 nimfa, tetapi untuk T. nigriabdominalis adalah 3.2 nimfa. Puncak keperidian R. rufiabdominalis
sebanyak 5 kali, sedangkan pada T. nigriabdominalis 3 kali. R. rufiabdominalis
15
Tipe bertahan hidup R. rufiabdominalis menunjukan kurva tipe II dan T. nigriabdominalis menunjukkan kurva tipe III. Menurut Price (1997), kurva tipe I adalah kematian populasi organisme yang rendah pada umur muda dan dalam jumlah besar pada umur tua, tipe II adalah kematian populasi suatu individu yang konstan dan tipe III adalah tingginya kematian populasi suatu individu yang
(a) R. rufiabdominalis, (b) T. nigriabdominalis
berlipat ganda yang tinggi pada suatu individu dapat menyebabkan penurunan sumber daya lingkungan dan mempengaruhi nilai laju pertambahan intrinsik (r).
Tabel 3 Neraca Kehidupan R. rufiabdominalis dan T. nigriabdominalis
Parameter R. rufiabdominalis T. nigriabdominalis Satuan instrinsik, (T) rataan lama generasi, (DT) waktu populasi berlipat ganda
Nilai rataan lama generasi (T) diartikan sebagai rataan waktu yang dibutuhkan sejak nimfa diletakkan hingga imago betina menghasilkan separuh keturunannya. Nilai rataan lama generasi pada R. rufiabdominalis dan T. nigriabdominalis berturut-turut adalah 8.93 hari dan 11.85 hari.
Nilai GRR (laju reproduksi kotor) R. rufiabdominalis lebih besar dari T. nigriabdominalis. Nilai GRR dan R0 yang tinggi pada R. rufiabdominalis
memperlihatkan tingkat kesesuaian hidup terhadap tanaman inang. Nilai R0 (laju
reproduksi bersih) pada R. rufiabdominalis menunjukkan bahwa generasi berikutnya akan meningkat sebanyak 62.18 kali dari generasi sebelumnya, sedangkan nilai R0 pada T. nigriabdominalis hanya meningkat sebanyak 5.18 kali.
Laju pertambahan intrinsik merupakan kapasitas suatu populasi untuk peningkatan. Nilai yang diperoleh ditentukan oleh berbagai aspek yang berhubungan dengan sejarah kehidupan organisme, yaitu kematian, kelahiran dan waktu perkembangan (Kurniawan 2007). Nilai r T. nigriabdominalis lebih rendah daripada R. rufiabdominalis. Nilai r pada T. nigriabdominalis berkisar antara 0.14 nimfa per hari, sedangkan pada R. rufiabdominalis 0.46 nimfa per hari. Siklus hidup yang panjang pada T. nigriabdominalis menyebabkan laju pertambahan intrinsiknya menjadi rendah (Tabel 3).
Tingginya nilai r ini disebabkan oleh tingginya keperidian dan rendahnya mortalitas pada pradewasa dan dewasa. Akan tetapi menurut Birch (1948), nilai laju pertambahan intrinsik (r) yang tinggi pada suatu spesies tidak selalu diartikan sebagai tingkat keberhasilan dalam suatu habitat. Hal tersebut berdasarkan adanya proses seleksi dari spesies tersebut agar nilai r-nya menjadi relatif tinggi sehingga mampu berkompetisi dengan spesies lain. Laju pertambahan intrinsik dapat digunakan untuk memprediksi pertumbuhan populasi serangga dalam jangka waktu yang panjang. Laju pertambahan intrinsik yang rendah dapat diartikan bahwa populasi suatu organisme memiliki sedikit kemungkinan untuk terus tumbuh.
Biologi R. rufiabdominalis pada Daun, Batang dan Akar Padi
Kutudaun R. rufiabdominalis merupakan kutudaun di akar padi, tetapi pada penelitian ini dicoba dibiakkan pada daun, batang dan akar padi untuk melihat kecocokan dan ketahanan hidup. Pada daun padi R. rufiabdominalis tidak bisa diamati biologinya secara keseluruhan karena pada daun padi R. rufiabdominalis
17
bertahan hidup di daun padi dikarenakan daun padi sedikit lebih keras sehingga instar satu tidak bisa menusukan stilet pada daun padi.
Menurut Hsieh (1970); Kindler et al. (2004); Zilahi et al. (2005), kutudaun di akar ini dapat bertahan hidup di wadah pemeliharaan dengan pakan akar dan batang tanaman. Ada beberapa kutudaun yang memang hidup pada daun, seperti
Rhopalosiphum maidis pada jagung (Carena dan Glogoza 2004); Macrosiphum rosae pada daun mawar (Kmiek 2007), Macrospihum euphorbiae pada daun tomat (Hummel et al. 2004), Myzus persicae pada daun tanaman kentang atau dari famili Solanaceae (Musa et al. 2004; Troncoso et al. 2005). Penelitian kutudaun di akar yang dibiakkan pada daun sampai saat ini belum dilakukan, sehingga pada penelitian ini dilakukan pemeliharaan dengan menggunakan daun padi sebagai pakan, namun tidak ada kutudaun yang berhasil menyelesaikan siklus hidupnya.
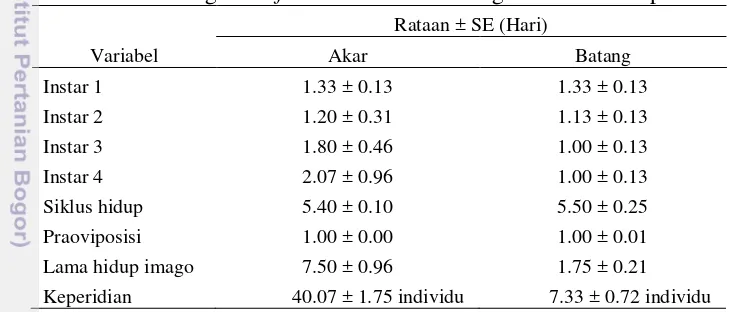
Hasil rataan biologi R. rufiabdominalis di akar dan batang tidak jauh berbeda (Tabel 4). Siklus hidup R. rufiabdominalis di akar dan batang berturut-turut adalah 5.40 hari dan 5.50 hari. Pada batang padi serangga ini akan masuk ke dalam lipatan batang, karena dalam lipatan batang kondisi pakannya lebih lunak. Salah satu kunci perkembangan hidup yang cepat bagi serangga herbivora adalah kecocokan inangnya (Awmack dan Leather 2002). Semakin tidak cocok inang yang diberikan kepada serangga herbivora, akan berdampak pada penurunan populasi dan penurunan kualitas hidupnya (Legrand & Barbosa 2000).
Tabel 4 Biologi R. rufiabdominalis di batang dan akar daun padi Variabel
Kutudaun lebih suka dengan permukaan tanaman yang tidak terlalu keras karena serangga ini memakan bagian floem tanaman dengan menggunakan stiletnya (Kindler et al. 2004). Akar tanaman memiliki bagian yang lebih lunak dibandingan dengan bagian lainnya, sehingga banyak kutudaun jenis ini yang menyerang pada bagian tersebut (Emden dan Bashford 1969; Weathersbee dan Hardee 1994). Menurut Hsieh (1970), ketika kondisi lahan dalam keadaan kering kutudaun ini dapat terlihat bergerombol berada pada akar tanaman padi, sedangkan ketika kondisi lahan dalam keadaan basah atau tergenang air dan populasi kutudaun tinggi, maka kutudaun ini dapat berpindah untuk menghisap atau makan pada bagian batang.
pendek dari pada di akar karena kurangnya nutrisi makanan untuk kelangsungan hidup sehingga mortalitas tinggi dan sedangkan di akar lebih lama karena nutrisi makanan sehingga mortalitas nya tidak terlalu tinggi.
Lama hidup R. rufiabdominalis berpengaruh terhadap keperidian, karena kekurangan nutrisi akan menggangu reproduksi. R. rufiabdominalis yang kekurangan nutrisi meyebabkan keperidian yang rendah sehingga mengurangi jumlah populasi. Keperidian di batang lebih rendah dari pada di akar. Keperidian di akar 40.07 nimfa dan keperidian di batang 7.33 nimfa. Menurut Zilahi-Balogh
et al. (2005), ketika dalam populasi kutudaun sudah terdapat generasi yang memiliki sayap, maka kutudaun bersayap tersebut akan berpindah ke batang. Akan tetapi, hanya berpindah sampai dengan pangkal batang yang memiliki permukaan yang lebih lunak dibandingkan dengan permukaan batang lainnya.
Kecilnya parameter siklus biologi yang didapat kutudaun yang dipelihara pada batang mungkin disebabkan karena pemilihan batang yang kurang lunak sehingga kutudaun tersebut kesulitan untuk mendapatkan makanan dan mengalami gagal tumbuh. Selain itu, siklus hara dari akar lebih banyak ditransport ke daun, sehingga floem pada akar lebih banyak megandung nutrisi atau unsur hara dan hal ini dapat memberikan nutrisi yang cukup bagi pertumbuhan kutudaun yang dipelihara pada bagian akar. Walaupun memiliki sayap, akan tetapi menurut Chapin et al. (2001), pergerakannya tidak terlalu aktif dibandingkan dengan
Rhopalosiphum padi dan biasanya hanya terbawa oleh angin atau agen pembawa lainnya (hewan & manusia). Menurut Rusli (1999), Aphis glycine pada kedelai lebih banyak ditemukan pada daun dibandingkan pada batang.
Neraca Kehidupan R. rufiabdominalis pada Batang dan Akar Padi
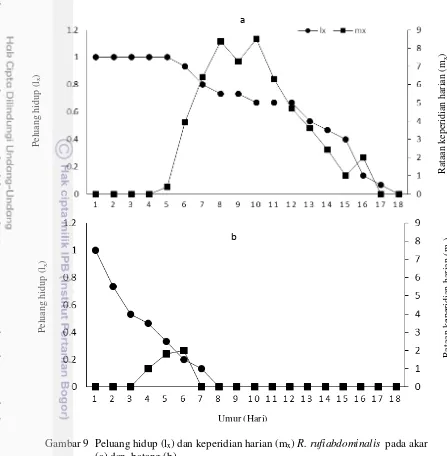
Rataan jumlah nimfa yang dilahirkan oleh setiap imago R. rufiabdominalis
di akar setiap harinya berbeda dengan imago R. rufiabdominalis di batang (Gambar 9). Keperidian harian tertinggi pada R. rufiabdominalis di akar mencapai 8.5 nimfa, sedangkan pada batang hanya 2 nimfa. Puncak keperidian R. rufiabdominalis di akar sebanyak 3 kali, sedangkan pada batang hanya 1 kali. R. rufiabdominalis di akar tidak menghasilkan nimfa dari umur 17 hari, sedangkan pada batangtidak menghasilkan nimfa dari umur 7 hari.
19
Gambar 9 Peluang hidup (lx) dan keperidian harian (mx) R. rufiabdominalis pada akar (a) dan batang (b)
Nilai GRR R. rufiabdominalis di akar lebih besar dari pada di batang. Nilai GRR dan R0 yang tinggi pada akar padi memperlihatkan tingkat kesesuaian hidup
terhadap tanaman inang. Nilai R0 pada R. rufiabdominalis di akar menunjukkan
bahwa generasi berikutnya akan meningkat sebanyak 37.40 kali dari generasi sebelumnya, sedangkan nilai R0 pada batang meningkat sebanyak 1.47 kali. Nilai r
R. rufiabdominalis di batang lebih rendah daripada di akar. Nilai r pada batang berkisar antara 0.08 nimfa per hari, sedangkan pada R. rufiabdominalis 0.46 nimfa per hari. Laju pertambahan intrinsik merupakan kapasitas suatu populasi untuk peningkatan, nilai yang diperoleh ditentukan oleh berbagai aspek yang berhubungan dengan sejarah kehidupan organisme, yaitu kematian, kelahiran dan waktu perkembangan (Kurniawan 2007).
Parameter neraca kehidupan yang diukur dari kutudaun R. rufiabdominalis
pada akar dan batang menunjukan bahwa kelima parameter menunjukan hasil
yang berbeda (Tabel 5). Hal ini dikarenakan kesesuaian pakan atau inang yang diberikan. Kutudaun ini lebih memilih untuk menghisap cairan floem dari akar (Hsieh 1970; Blackman & Eastop 2000; Kindler et al. 2004). Kutudaun yang dipelihara pada akar memiliki nilai parameter statistika demografi yang lebih baik dibandingkan kutudaun yang dipelihara pada batang. Nilai T pada batang lebih rendah daripada pada akar, nilai T pada batang 4.45 hari sedangkan nilai T pada akar 8.82 hari. Nilai DT pada batang 8.01 hari dan nilai DT pada akar 1.49 hari (Tabel 5).
Tabel 5 Neraca Kehidupan R. rufiabdominalis di akar dan batang No Variabel Akar Batang Satuan 1. GRR 54.93 4.80 Individu/generasi 2. Ro 37.40 1.47 Individu/induk/generasi 3. r 0.46 0.08 IiIndividu/induk/hari
4. T 8.82 4.45 Hari
5. DT 1.49 8.01 Hari
Keterangan: (GRR) Laju reproduksi kotor, (R0) laju reproduksi bersih, (r) laju pertambahan instrinsik, (T) rataan lama generasi, (DT) waktu populasi berlipat ganda
Hal ini menunjukan bahwa kutudaun tersebut juga dapat hidup pada batang padi dan menunjukan nilai yang lebih bagus daripada di akar. Seperti disebutkan di atas bahwa ketika populasi kutudaun meningkat atau ketika kondisi lahan dalam keadaan tergenang air, maka kutudaun ini akan naik ke batang dan hidup pada bagian tersebut (Hsieh 1970; Zilahi-Balogh et al. 2005). Akan tetapi apabila dilihat dari preferensi makan, maka kutudaun yang dipelihara pada akar lebih cepat menghabiskan makanannya dibandingkan yang dipelihara pada batang. Hal ini dapat dilihat pada nilai DT yang kecil. Menurut Birch (1948), serangga yang memiliki waktu berlipat ganda yang cepat maka akan mempercepat penurunan sumberdaya makanan. Perkembangan dan pertumbuhan kutudaun dipengaruhi oleh inang yang diberikan selama masa pemeliharaan (Kersting et al. 1998; Satar
SIMPULAN DAN SARAN
Simpulan
Kutudaun R. rufiabdominalis dan T. nigriabdominalis telah ditemukan di beberapa daerah di Jawa Barat. Tidak adanya pembatas alami antar provinsi dan kesamaan penggunaan varietas padi Ciherang dan IR-64 di berbagai sentra pertanaman padi di pulau Jawa, kemungkinan besar kutudaun ini juga telah terdapat pada pertanaman padi di pulau Jawa.
Hasil kajian neraca kehidupan R. rufiabdominalis memiliki tingkat pertambahan intrinsik (r) 0.46 hari dan waktu penggandaan populasi menjadi dua kali lipat (DT) 1.51 hari, sedangkan T. nigriabdominalis memiliki tingkat pertambahan intrinsik (r) 0.16 hari dan waktu penggandaan populasi menjadi dua kali lipat (DT) 4.33 hari. Kedua spesies kutudaun ini memiliki siklus hidup yang pendek dan keperidian yang tinggi, sehingga berpotensi menjadi hama penting pada pertanaman padi di daerah kering.
Saran
Awmack CS, Leather SR. 2002. Host plant quality and fecundity in herbivorous insects. Annu Rev Entomol. 47: 817-844.
Begon M, Townsend CR dan Herper JL. 2008. Ecology: From Individuals to Ecosystems. 4th edition. Oxford. (UK): Blackwell puslishing.
Birch LC. 1948. The intrinsic rate of natural increase of an insect population. J Anim. Ecol. 17(1):15-28
Borror DJ, Delong DM. 1971. An Introduction to the Study of Insects. Third Ed. Holt, New York (US) Rinehart and Winston Inc.
Blackman RL, Eastop VF. 2000. Aphids on the World’s Crops. Chicester (GB): John Wiley & Sons.
Blackman RL, Eastop VF. 2006. Aphids on the World’s Herbaceous Plants and
Shrubs. Chicester (GB): John Wiley & Sons.
Carena MJ, Glogoza P. 2004. Resistance of maize to the corn leaf aphid: A review. Maydica. 49: 241-254.
Carver M, Gross GF, Woodmard TE. 2000. Hemiptera. Di dalam: Nauman ID, carne PB, Lawrence JF, Nielsen ES, Spradbery JP, Taylor RW, Whitten MJ, Littlejohn MJ, editor. The Insect of Australia. Volume 1. Victoria (AU):
Brevicoryne brassicae and Myzus persicae in relation to plant age and leaf amino acids. Entomolo Experiment et Appl. 14: 349-360.
Galli E, Manicardi GC. 1998. Cytogenetic analysis on the gall generation of two Aphid spesies: Tetraneura nigriabdominalis and T. ulmi. Dipartimeno di Biologia Animale. University di Modeno. Italy. Caryologia. 51:3-4: 235-243.
Heinrichs EA, Barrion AT. 2004. Rice-Feeding Insects and Selected Natural Enemies in West Africa. Manila (PH). International Rice Recearch Institute. Hill A.R. 1971. The reprodictive behavior of metapolophium testucea (Theobald)
at different temperatures and different host Plant. Ann Appl Biol. 67(3). Hsieh C. 1970. The aphids attacking rice plants in Taiwan. Plant Protect Bull. 12:
68-77.
Hummel NA, Zalom FG, Miyao GM, Underwood NC, Villalobos A. 2004. Potato aphid, Moacrosiphum euphorbiae (Thomas), in tomatoes: Plant canopy distribution and binomial sampling on processing tomatoes in California. J Econ Entomol. 97(2): 490-495.
23
Kalshoven LGE. 1950. De Plagen van de Cultuurgewassen in Indonesia.
Bandoeng (ID): Uitgeverij W. Van hoeve-„s-Gravenhage.
Kalshoven LGE. 1981. The Pests of Crops in Indonesia. Laan PA van der, penerjemah. Jakarta (ID): Ichtiar Baru-van Hoeve. Terjemahan dari: De Plagen van de Cultuurgewassen in Indonesia.
Kersting U, Satar S, Uygun N. 1998. Effect of temperature on development rate and fecundity of apterous Aphis gossypii Glover reared on Gossypium hirsutum L. J of Appl Entomol. 123: 23-27.
Kindler D, Hesler L, Elliortt N, Shufran K, Springer T. 2004. Cereal and grass hosts of the rice root Aphid, Rhapolosiphum rufiabdominalis (Sasaki), and a description of an efficient green house rearing techniques. J Agric Urban Entomol. 21 (1):51-59.
Kmiek. 2007. The occurence of Macrosiphum rosae (L.) (Hemiptera: Aphididae) on rose shrubs in Lublin. J of Poland Agricultural. Available at development and poopulation growth of Tetranura nigriabdominalis
(Homoptera: Pemphigidae) on three host plant. J of Eco Entomol. 99(4): 1209-2113.
Kuswanto, Budi W. 2007. Model pendugaan jumlah aphid (A. craccivora Koch) secara in situ pada tanaman kacang panjang (Vigna sesquipedalis). J Ilmu Pertanian Indones. 14(1): 69-77.
Kurniawan HA. 2007. Neraca kehidupan kutukebul, Bemisia tabaci Gennadius (Hemiptera: Aleyrodidae) Biotipe-B dan Non-B pada tanaman mentimun (Curcumis sativus L) dan cabai (Capsicum annuum L) [tesis]. Bogor (ID): Institut Pertanian Bogor.
Legrand A, Barbosa P. 2000. Pea aphids (Hom: Aphididae) fecundity, rate of increase, and within plant distribution unaffected by plant morphology.
Environt Entomol. 29: 987-993.
Miles PW. 1987. Feeding process of Aphidoidea in relation of effects on their food plants. Di dalam: Minks AK, Harrewijin P, editor. Aphids: Their Biology Natural Enemis and Control. Vol 2A. Amsterdam (NL): Elsevies. 321-340.
Modic S, Razinger J, Urek G. 2011. Tetraneura (Tetraneurelia) nigriabdominalis
(sasaki) gall forming aphid found maize roots in slovenia. J Acta Entomol. Slovenica. 20(2):147-152.
Morgan D, Walters KFA, Aegerter JN. 2001. Effect of temperature and cultivar on pea aphid, Acyrthosiphon pisum (Hemiptera: Aphididae) life history. Bull of Entomol Research. 91: 47-52.
Musa FM, Carli C, Susuri LR, Pireva IM. 2004. Monitoring of Myzus persicae
(Sulzer) in potato fields in Kosovo. Acta Agri Slovenica. 83(2): 379-385. Painter RH. 1951. Insect Resistence in Crop Plant. Kansas (USA): University of
Pathak MD, Khan ZR. 1994. Insect Pests of Rice. Manila (PH). International Rice Recearch Institute.
Pike KS, Boydston L, Allison D. 1990. Alate aphid viviparae associated with small grains in North America: a key and morphometric characterization. J Kansas Entomol Soc. 63: 559–602.
Price PW. 1997. Insect Ecology. 3rd ed. New York (US): John Wiley & Sons. Rahmah ST. 2013. Jenis dan karakteristik koloni kutudaun (Hemiptera:
Aphididae) di Bogor dan Cianjur [skripsi]. Bogor (ID) : Institut Pertanian Bogor.
Razmjou J, Moharramipour S, Fathipour Y, Mirhoseini SZ. 2006. Effect of cotton cultivar on performance of Aphis gossypii (Hemiptera: Aphididae) in Iran. J of Econom Entomol 99: 1820-1825.
Rusli R. 1999. Biologi Aphis glycine Matsumura (Homoptera: Aphididae) pada beberapa tingkat umur tanaman kedelai (Glycine max (L) Merrill)
Saito K, Linquist B, Keobualapha K, Phanthabon T, Shiraiwa T, Hone. 2006. Cropping intensity and rainfall effects on upland rice yields in northern laos.
J Plant Soil. 284:175-185. (10 1007/s 11104-006-0049-5).
Saragih, Evarista BR. 1994. Identifikasi dan Biologi Kutudaun pada Brassicaceae Liar. Institut Pertanian Bogor (ID). 20Hal
Satar S, Kersting U, Ulusoy MR. 2005. Temperature dependent life history traits of Brevicoryne brassicae (L.) (Hom., Aphididae) on white cabbage. Turkish J of Agri and Forest. 29(5): 341-346.
Shepard BM, Barrion AT, Litsinger JA. 1995. Rice Feeding Insects of Tropical Asia. Manila (PH). International Rice Research institute.
Subagyo VNO, Hidayat P. 2014. Neraca kehidupan kutu kebul Bemisia tabaci
(Gennadius) (Hemiptera: Aleyrodidae) pada tanaman cabai dan gulma babadotan pada suhu 25°C dan 29°C. J Entomol Indones. 11(1): 11-18. Troncoso AJ, Vargas RR, Tapia DH, Olivares-Donoso R, Niemeyer HM. 2005.
Host selection by the generalist aphid Myzus persicae (Hemiptera: Aphididae) and its subspecies specialized on tobacco, after being reared on the same host. Bull of Entomol Research. 95: 23-28.
Weathersbee III AA, Hardee DD. 1994. Abundance of cotton aphids and associated biological control agents on six cotton cultivars. J of Economic Entomol. 87: 258-265.
Zilahi-Balogh GMG, Foottit RG, Ferguson G, Shipp JL. 2005. New Records for
27
Lampiran 2 Neraca kehidupan T. nigriabdominalis pada akar padi Hari
pengamatan ke kelas umur (x)
individu yang hidup (lx) jumlah keturunan betina (mx) lxmx
00.5 1.00 0.00 0.00
01.5 0.98 0.00 0.00
02.5 0.97 0.00 0.00
03.5 0.92 0.00 0.00
04.5 0.67 0.00 0.00
05.5 0.58 0.00 0.00
06.5 0.55 0.00 0.00
07.5 0.48 0.59 0.28
08.5 0.40 0.63 0.25
09.5 0.38 1.78 0.68
10.5 0.30 2.11 0.63
11.5 0.25 2.87 0.72
12.5 0.25 3.20 0.80
13.5 0.22 1.92 0.42
14.5 0.17 2.50 0.42
15.5 0.15 0.89 0.13
16.5 0.15 1.11 0.17
17.5 0.15 2.11 0.32
18.5 0.13 2.00 0.27
19.5 0.12 0.71 0.08
20.5 0.07 0.25 0.02
21.5 0.03 0.00 0.00
29
Lampiran 3 Neraca kehidupan R. rufiabdominalis pada akar padi Hari pengamtan




![BIMBINGAN DAN KONSELING LINTAS BUDAYA [Compatibility Mode]](data:image/gif;base64,R0lGODlhAQABAIAAAP///wAAACH5BAEAAAAALAAAAAABAAEAAAICRAEAOw==)