PRODUKSI PROTEIN REKOMBINAN HBsAg100-GST
SEBAGAI MODEL IMUNOGEN UNTUK
MENGHASILKAN ANTIBODI
PADA MENCIT
SLAMET RIYADI
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI DISERTASI DAN
SUMBER INFORMASI
Dengan ini saya menyatakan bahwa disertasi yang berjudul “Produksi
Protein HBsAg Rekombinan sebagai Model Imunogen untuk Menghasilkan
Antibodi pada Mencit” adalah karya saya dengan arahan dari komisi pembimbing
dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir disertasi ini.
Bogor, Pebruari 2012
SLAMET RIYADI. Production of HBsAg100-GST Recombinant Protein as an Immunogen Model for Generating Antibody in Mice. Under direction of RARAH R.A. MAHESWARI, MIRNAWATI SUDARWANTO, FRANSISKA R. ZAKARIA, and MUHAMAD ALI.
Since years ago, a new paradigm of vaccine design is emerging. Instead of attenuated virulent microorganisms or killed virulent microorganisms, effective subunit vaccines were developed using recombinant DNA technology. Biosynthesis of recombinant protein in Escherichia coli may offer an alternative procedure to generate therapeutic protein free from human protein. In this research, hydrophilic domain of S protein (aa 100-164)-encoding gene of hepatitis B surface antigen was cloned for vaccine candidate production. The gene was ligated with pGEX-4T-2 vector and sequenced. Sequences alignment of the amplified fragment with genome of hepatitis B virus indicated that the sequences were identical. In this research, cloned DNA fragment of Hepatitis B surface antigen was placed downstream from the gluthatione S-transferase (GST) protein-encoding gene in expression plasmid pGEX-4T-2 and expressed in Escherichia coli cells. A polypeptide of 34.8 kDa molecular weight was synthesized and identified as HBsAg100-GST fusion proteins. The recombinant proteins were then purified using GSTrap and HiTrap column and could be used for vaccine candidate or for antibody generation. The purified protein was tried to trigger cell immune to produce antibody in mice. Results indicated that the immunogenicity of HBsAg100-GST was higher than GST protein in elicit the levels of HBsAg100-specific IgG antibody in mice. These results suggest that the HBsAg100 produced in E. coli has immunogenicity. A major result achieved from this research was clones carrying S antigens-encoding gene that could be used further for production of recombinant hepatitis B vaccine candidates.
RINGKASAN
SLAMET RIYADI. Produksi Protein Rekombinan HBsAg100-GST sebagai Model Imunogen untuk Menghasilkan Antibodi pada Mencit. Dibimbing oleh RARAH R.A. MAHESWARI, MIRNAWATI SUDARWANTO, FRANSISKA R. ZAKARIA, dan MUHAMAD ALI.
Kemajuan teknologi molekuler dalam beberapa dekade terakhir, terutama sejak ditemukannya sekuen genom lengkap dari mikroba-mikroba patogen, telah menemukan jalan baru bagi dihasilkannya berbagai jenis protein rekombinan, baik vaksin, antibodi, maupun peptide sintetik yang memiliki manfaat tertentu. Pada saat ini, vaksin telah dihasilkan dengan teknologi DNA rekombinan, yaitu melalui kloning gen penyandi protein tertentu pada mikroorganisme patogen yang dilanjutkan dengan ekspresi gen tersebut pada sel hewan, sel tanaman, ataupun pada bakteri.
Penggunaan mikroorganisme virulen yang dilemahkan ataupun yang
dimatikan telah diganti dengan penggunaan vaksin sub unit yang lebih efektif
dengan teknologi DNA rekombinan. Melalui penggunaan teknologi tersebut, gen
tertentu dari mikroorganisme virulen dapat dikloning, diekspresi dan dievaluasi
penggunaannya sebagai vaksin. Tersedianya bioteknologi rekayasa genetika yang dilahirkan pada tahun 1973, telah memungkinkan manusia untuk mengisolasi gen (serangkaian molekul DNA) serta memanipulasinya, kemudian memindahkan gen tersebut dari satu organisme ke organisme lain. Peranan bioteknologi dirasakan semakin bertambah besar dalam menunjang kegiatan pembangunan industri di berbagai sektor, terutama sektor kesehatan dan pertanian termasuk sub sektor peternakan.
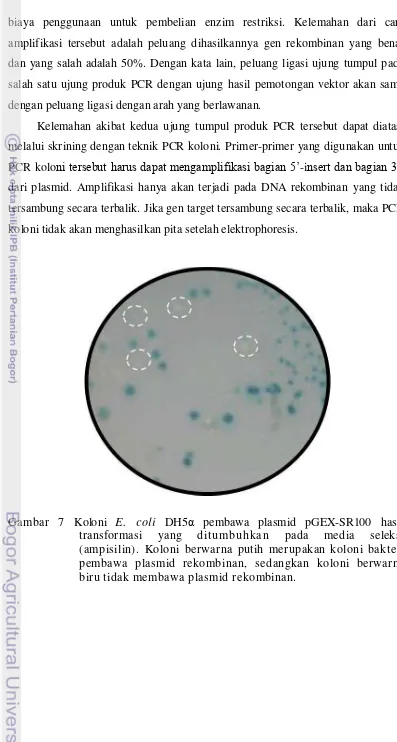
plasmid, sehingga dipastikan tidak terjadi kesalahan arah insert. Amplifikasi hanya akan terjadi pada DNA rekombinan yang tidak tersambung secara terbalik. Adanya pita tunggal DNA dari gambar hasil elektroforesis merupakan indikasi bahwa klon yang diamplifikasi mengandung plasmid rekombinan.
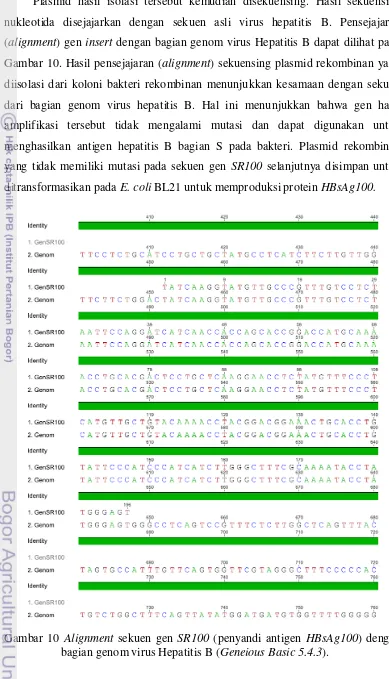
Koloni yang mengandung plasmid rekombinan dengan hasil PCR koloni pita tunggal kemudian dikultur dari replika pada media LB pada suhu 37oC selama 12 jam dengan shaker untuk isolasi plasmid rekombinan. Plasmid hasil isolasi tersebut kemudian disekuensing. Hasil pensejajaran (alignment) sekuensing plasmid rekombinan yang diisolasi dari koloni bakteri rekombinan menunjukkan kesamaan dengan sekuen dari bagian genom virus hepatitis B. Hal ini menunjukkan bahwa gen hasil amplifikasi tersebut tidak mengalami mutasi dan dapat digunakan untuk produksi antigen hepatitis B bagian S pada bakteri. Plasmid rekombinan yang tidak memiliki mutasi pada sekuen insert selanjutnya disimpan untuk ditransformasikan pada E. coli BL21 dan produksi protein HBsAg100-GST.
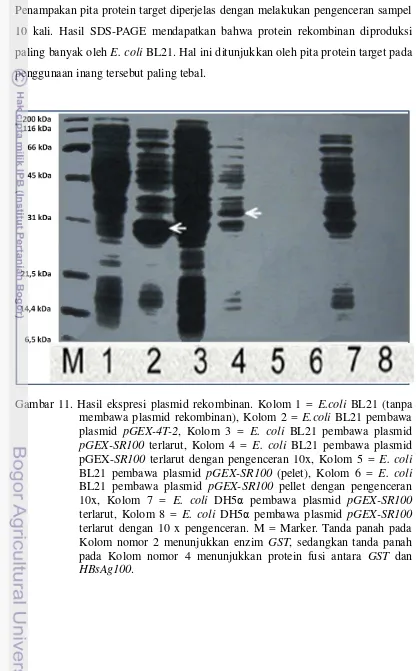
Pemisahan terhadap hasil sonikasi untuk mengetahui bahwa protein rekombinan dalam bentuk terlarut (soluble) menggunakan sentrifugasi dan filterisasi (filter ukuran 0.22 µm). Kelarutan protein rekombinan sangat penting untuk mempermudah proses pemurnian. Hasil yang diperoleh baik larutan maupun pelet dimasukkan ke dalam gel akrilamid. Kelarutan dari protein rekombinan diperlihatkan oleh adanya pita-pita protein target pada bagian supernatan. Sebaliknya hasil SDS-PAGE dari pelet bakteri yang tidak memperlihatkan adanya pita-pita dari protein target menjadi indikator bahwa protein rekombinan tersebut berada dalam bentuk tak larut (insoluble). E. coli BL21 yang membawa plasmid rekombinan pGEX-SR100 memiliki protein dengan ukuran sekitar 34.8 kDa karena merupakan gabungan antara GST yang memiliki berat 28 kDa dengan antigen S dengan ukuran 6.8 kDa.
Penelitian ini menggabungkan fragmen DNA dari antigen permukaan virus
Hepatitis B dengan gen penyandi enzim gluthation-S-transferase (GST) di dalam
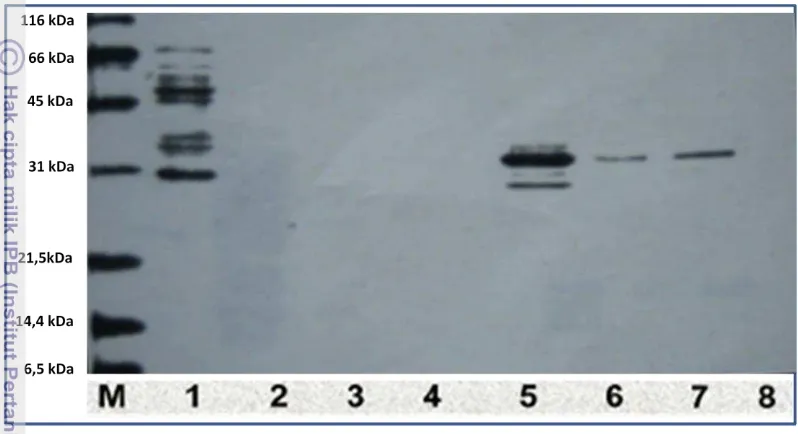
protein gabungan HBsAg100-GST. Protein gabungan tersebut kemudian dimurnikan menggunakan kolum GSTrap yang disambung dengan kolum HiTrap.
Hasil pemurnian fusi HBsAg100 dan GST dalam penelitian ini menunjukkan bahwa antigen rekombinan yang diperoleh setelah pemurnian relatif murni dan dalam jumlah yang cukup untuk digunakan dalam aplikasi (assay) selanjutnya. Keberhasilan isolasi ini tidak terlepas dari sifat meningkatnya kelarutan protein rekombinan karena fusi dengan GST. Berdasarkan uji antigenisitas pada mencit BALB/c, protein HBsAg100-GST hasil purifikasi dapat menghasilkan antibodi
anti HBsAg100-GST yang berpotensi sebagai vaksin.
Nilai optikal densiti (OD) dari serum mencit yang diperoleh dari darah mencit sebelum dan setelah dilakukan vaksinasi dengan HBsAg100-GST pada kelompok A dan dengan GST pada kelompok B menunjukkan, bahwa rerata respon humoral mencit yang diimunisasi dengan fusi protein meningkat setelah dilakukan imunisasi maupun setelah dilakukan booster. Namun, seiring dengan penambahan waktu pemeliharaan, respon humoral mencit tersebut menurun sedikit demi sedikit sampai akhir minggu ke 12.
Berdasarkan hasil penelitian ini dapat disimpulkan bahwa gen SR100
berhasil diamplifikasi, kemudian diligasi dengan vektor pGEX-4T-2, dan
ditransformasikan ke dalam bakteri E. coli DH5α. Hasil sekuensing menunjukkan
tidak terdapat mutasi pada gen hasil kloning. Uji lanjut untuk konfirmasi
imunogenisitas protein antigen HBsAg100-GST masih perlu dilakukan pada
hewan lain seperti kelinci, kambing dan kuda.
© Hak Cipta milik IPB, tahun 2012
Hak Cipta dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan yang wajar IPB.
PRODUKSI PROTEIN REKOMBINAN HBsAg100-GST
SEBAGAI MODEL IMUNOGEN UNTUK
MENGHASILKAN ANTIBODI
PADA MENCIT
SLAMET RIYADI
Disertasi
sebagai salah satu syarat untuk memperoleh gelar Doktor pada
Program Studi Ilmu Ternak
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Penguji Luar Komisi Pembimbing
Penguji Luar Komisi Pembimbing Ujian Tertutup (25 Januari 2012) 1. Prof. Dr. drh. Retno Damayanti Soeyoedono, MS
2. Dr. Jakaria, S.Pt., M.Si
Penguji Luar Komisi Pembimbing Ujian Terbuka (30 Januari 2012) 1. Prof. Dr. Ir. Cece Sumantri, M.Agr.Sc
Judul Disertasi : Produksi Protein Rekombinan HBsAg100-GST sebagai Model Imunogen untuk Menghasilkan Antibodi pada Mencit
Nama : Slamet Riyadi NRP : D061030061 Program Studi : Ilmu Ternak
Disetujui Komisi Pembimbing
Dr. Ir. Rarah R.A. Maheswari, DEA Prof. Dr. drh. Mirnawati Sudarwanto
Ketua Anggota
Prof. Dr. Ir. Fransiska R. Zakaria, M.Sc Muhamad Ali, S.Pt., M.Si., Ph.D
Anggota Anggota
Diketahui
Ketua Program Studi Dekan Sekolah Pascasarjana IPB Ilmu Ternak
Dr. Ir. Rarah R.A. Maheswari, DEA Dr. Ir. Dahrul Syah, M.Sc.Agr
Atas tersusunnya disertasi ini dengan judul “Produksi Protein Rekombinan HBsAg100-GST sebagai Model Imunogen untuk Menghasilkan Antibodi pada
Mencit”, Penulis panjatkan puji syukur kepada Allah SWT., Tuhan yang maha mengetahui dan maha menolong sehingga karya ilmiah ini berhasil diselesaikan.
Disertasi ini memuat tiga bab yang merupakan pengembangan dari naskah artikel yang diajukan ke jurnal ilmiah. Bab 1 berjudul Kloning Gen SR100 dalam rangka Produksi Protein Rekombinan sebagai Model Imunogen untuk Menghasilkan Antibodi sedang menunggu penerbitan di Jurnal Peternakan Indonesia Fakultas Peternakan Universitas Andalas, Padang. Bab 2 berjudul Biosintesis Antigen Permukaan HBsAg100 pada E. coli dalam rangka Produksi Protein Rekombinan sebagai Model Imunogen untuk Menghasilkan Antibodi juga sedang menunggu penerbitan di Jurnal Kedokteran YARSI Fakultas Kedokteran Universitas YARSI, Jakarta. Bab 3 berjudul Imunogenisitas Protein Rekombinan HBsAg100-GST dalam Memicu Sel Imun untuk Menghasilkan Antibodi pada Mencit sedang disiapkan untuk dikirim ke penerbit Jurnal Ilmiah.
Penulis menyampaikan ucapan terima kasih yang setinggi-tingginya kepada Komisi Pembimbing, Dr. Ir. Rarah Ratih Adji Maheswari, DEA (Ketua); Prof. Dr. drh. Mirnawati Sudarwanto (Anggota); Prof. Dr. Ir. Fransiska R. Zakaria, M.Sc. (Anggota), dan Dr. Muhamad Ali, M.Si. (Anggota). Ungkapan terima kasih juga disampaikan kepada istri dan anak-anak tercinta atas segala do’a dan kasih sayangnya. Penulis juga menyampaikan ucapan terima kasih yang sebesar-besarnya kepada semua pihak yang telah membantu dalam proses penulisan disertasi ini.
Kritik dan saran yang sifatnya membangun demi penyempurnaan disertasi ini sangat diharapkan. Semoga karya ilmiah ini dapat mendukung pengembangan bioteknologi di Indonesia pada khususnya dan bermanfaat bagi para pembaca.
Bogor, Pebruari 2012
RIWAYAT HIDUP
Dilahirkan di Pemalang pada tanggal 29 Maret 1960 sebagai anak kedua dari enam bersaudara dari pasangan Bapak Kampyun (alm) dan Ibu Maryati (alm). Menikah dengan Rahma Jan dikaruniai seorang putri, Lisantiyas Nurani mahasiswi semester lima Fakultas Kedokteran UNRAM, dan dua orang putra, Abdillah Rahmadiputra mahasiswa semester tiga jurusan Teknik Mesin Fakultas Teknik UNRAM dan Abdul Ghoffar Triatmojo mahasiswa semester tiga Fakultas Kedokteran UNRAM. Pada saat ini, bertugas sebagai Staf Pengajar di Fakultas Peternakan Universitas Mataram di Mataram.
Halaman
Aplikasi Rekayasa Genetik di Bidang Peternakan... 23
IMUNOGENISITAS PROTEIN REKOMBINAN HBsAg100-GST DALAM MEMICU SEL IMUN UNTUK MENGHASILKAN ANTIBODI PADA MENCIT
Abstrak ... 49
Abstract ... 49
Pendahuluan ... 49
Bahan dan Metode ... 51
Hasil dan Pembahasan ... 54
Simpulan ... 57
Daftar Pustaka ... 57
PEMBAHASAN UMUM ... 59
SIMPULAN DAN SARAN ... 65
DAFTAR PUSTAKA ... 67
Halaman 1 Ukuran genome dari beberapa makhluk hidup dalam bentuk haploid ... 16 2 Daftar primer yang digunakan dalam penelitian ... 30 3 Nilai optikal densiti (OD) serum mencit yang diperoleh dari darah mencit
DAFTAR GAMBAR
Halaman 1 Sekelompok virus Hepatitis B (Sumber: Stannard 1995) ... 10 2 Pembesaran dari dua buah core yang ditunjuk dengan tanda panah
(Sumber: Stannard 1995) ... 10 3 Representasi diagram dari virus Hepatitis B dan komponen antigen
permukaan (Sumber: Stannard 1995) ... 10 4 Ilustrasi virus Hepatitis B dengan capsid dan internal density yang
tampak pada irisan melintang (Sumber: Dryden et al. 2006) ... 11 5 Diagram struktur dari bagian DNA heliks ganda (Sumber: Andre
2006) ... 14 6 Diagram organisasi genome virus hepatitis B (Sumber: Wagner
2004) ... 18 7 Koloni E. coli DH5α pembawa plasmid pGEX-SR100 hasil
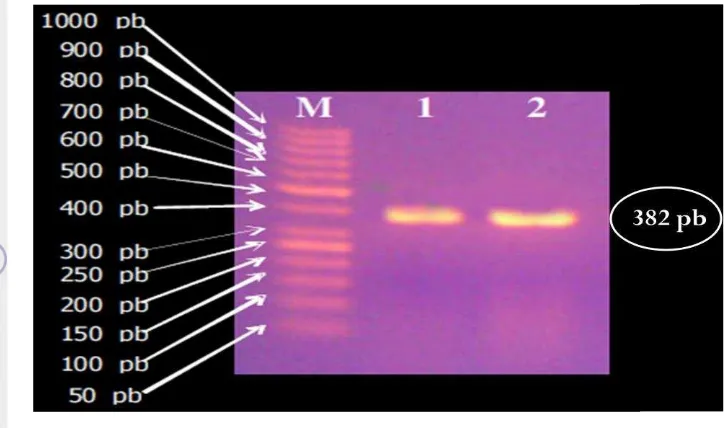
transformasi yang d itumbuhkan pada media seleksi (ampisilin). Koloni berwarna putih merupakan koloni bakteri pembawa plasmid rekombinan, sedangkan koloni berwarna biru tidak membawa plasmid rekombinan ... 33 8 Hasil elektrophoresis dari PCR koloni. M = Marker (1000 pb), 1
dan 2 = E. coli DH5α pembawa plasmid pGEX-SR100 sebagai cetakan ... 34 9 Pita DNA plasmid pGEX-4T-2 rekombinan hasil elektroforesis
dalam 1% agrosa M : marker DNA λ. Lajur 1: Pita DNA plasmid utuh pGEX-4T-2 rekombinan. Lajur 2, 3, 4, 5, 6, 7 : pita DNA plasmid pGEX-4T-2 rekombinan yang dipotong dengan enzim HindIII ... 34 10 Alignment sekuen gen insert (penyandi antigen HBsAg100) dengan
bagian genom virus Hepatitis B (Geneious Basics 5.4.3) ... 35 11 Hasil ekspresi plasmid rekombinan. Kolom 1 = E.coli BL21 (tanpa
12 Protein rekombinan hasil pemurnian. M = marker (ukuran berat molekul pada masing-masing pita dari atas ke bawah: 116 kDa, 66 kDa, 45 kDa, 31 kDa, 21,5 kDa, 14,4 kDa, 6,5 kDa), Kolom 1 = protein bakteri (unbound protein), Kolom 2 = protein bakteri (unbound protein) diencerkan 10x, Kolom 3 = protein bakteri (unbound protein) 2, Kolom 4 = protein bakteri (unbound protein) 2 yang diencerkan 10x, Kolom 5 = protein rekombinan (bound protein), Kolom 6 = protein rekombinan (bound protein) dengan pengenceran 10x, Kolom 7 = protein rekombinan (bound protein) 2, Kolom 8 = protein rekombinan (bound protein) 2 dengan pengenceran 10x ... 46 13 Imunisasi terhadap mencit dilakukan dengan penyuntikan secara
subcutaneus ... 52 14 Respon humoral mencit terhadap vaksinasi HBsAg100-GST
DAFTAR LAMPIRAN
Halaman
1 Topologi dan peta fisik plasmid pGEM-T Easy ... 73
2 Genom lengkap dari isolat virus hepatitis B “2059 Java” ... 74
3 Program PCR yang berhasil digunakan untuk amplifikasi gen SR100.. 77
4 Topologi dan peta fisik plasmid pGEX-4T-2 ... 78
5 Situs-situs pemotongan dan sekuen lengkap pGEX-4T-2 ... 79
6 Hasil sekuensing gen SR100 dengan menggunakan primer pGEX-5’... 83
7 Hasil sekuensing gen SR100 dengan menggunakan primer pGEX-3’... 85
8 Mesin Thermal Cycler untuk mengamplifikasi segmen DNA 87
9 Alat elektroforesis untuk memisahkan segmen DNA ... 87
10 Alat elektroforesis (BIO-RAD) untuk memisahkan molekul protein berdasarkan berat molekulnya (tampak depan) ... 88
11 Alat elektroforesis (BIO-RAD) untuk memisahkan molekul protein berdasarkan berat molekulnya (tampak atas) ... 88
12 Kelompok kandang mencit dalam penelitian ... 89
13 Keadaan mencit di dalam kandang percobaan ... 89
14 Proses mencampur HBsAg100-GST dengan Freund’s Adjuvant sebagai bahan vaksin ... 90
15 Menyiapkan mencit untuk vaksinasi ... 90
16 Proses vaksinasi terhadap mencit sedang berlangsung ... 91
17 Pengambilan darah mencit melalui ujung ekor ... 91
18 Hasil elisa dalam penentuan konsentrasi serum mencit untuk menguji 92 19 Mesin Elisa Photoreader yang digunakan untuk membaca hasil elisa.. 92
20 Printer yang terhubung dengan Mesin Elisa ... 93
Latar Belakang
Sejak tahun 1972 telah berkembang usaha rekayasa genetika yang
memberikan harapan bagi industri peternakan, baik yang berkaitan dengan
masalah reproduksi, pakan maupun kesehatan hewan. Old dan Primrose (1989)
menjelaskan, bahwa teknik rekayasa genetika telah ditemukan pada waktu yang
hampir bersamaan, yaitu pertama kali dilaporkan pada tahun 1972 oleh Jackson et
al. Selanjutnya dilaporkan pula oleh Lobban dan Kaiser pada tahun 1973 dengan
melakukan pengklonan suatu fragmen DNA asing, atau DNA penumpang, atau
DNA sasaran dalam suatu vektor. Winarno dan Agustinah (2007), menegaskan
bahwa dengan adanya penemuan tersebut menunjukkan awal dimulainya revolusi
bioteknologi modern.
Bioteknologi baru atau bioteknologi modern juga disebut sebagai rekayasa
genetika atau modifikasi genetika. Pada umumnya bioteknologi diasosiasikan
sebagai rekayasa genetik dan biologi molekuler, namun sebenarnya lebih luas dari
itu, yaitu meliputi mikrobiologi, biokimia dan pengetahuan reproduksi
(Wiryosuhanto dan Sudradjat 1992). Menurut Winarno (2004), The European
Federation of Biotechnology pada tahun 1982 telah memberikan definisi bahwa
bioteknologi adalah aplikasi terpadu dari biokimia, mikrobiologi, ilmu teknik atau
rekayasa (engineering) bagi pemanfaatan mikroba, kultur jaringan serta
komponen-komponennya dalam skala industri. Wiryosuhanto dan Sudradjat
(1992) mendefinisikan bioteknologi sebagai serangkaian teknik yang
berhubungan dengan biokimia dan kemampuan genetik dari mahluk hidup untuk
tujuan praktis. Muladno (2002) menyatakan, bahwa semua teknologi yang
memanfaatkan mahluk hidup sebagai salah satu komponen utamanya sering
disebut sebagai bioteknologi, namun dalam arti sempit, bioteknologi diartikan
sebagai teknologi rekayasa genetika yang bekerja pada level molekuler khususnya
DNA.
Wiryosuhanto dan Sudradjat (1992) menjelaskan, beberapa hasil penelitian
bioteknologi peternakan saat ini sudah dapat diaplikasikan dan dimanfaatkan
2
status kesehatan hewan. Selanjutnya dijelaskan bahwa bioteknologi reproduksi
meliputi inseminasi buatan, embryo transfer dan pemuliabiakan ternak dan dalam
upaya peningkatan reproduksi ternak telah dikembangkan penelitian dan aplikasi
bioteknologi sampai dengan generasi keempat, yaitu hewan transgenik. Sebagai
generasi pertama adalah inseminasi buatan dan embryo transfer merupakan
generasi kedua, sedangkan generasi adalah kloning. Bioteknologi di bidang pakan
merupakan teknologi biokimia dan mikrobiologi yang telah diaplikasikan untuk
perbaikan mutu pakan, seperti manipulasi mikroba rumen maupun dengan
perlakuan kimiawi dan mikrobiologi.untuk meningkatkan daya cerna dari hijauan
makanan ternak, jerami dan limbah pertanian yang tinggi kadar selulosanya.
Bioteknologi kesehatan hewan meliputi: (1) produksi komersial berbagai macam
zat penggertak pertumbuhan (growth promotors), seperti produksi hormone
dengan DNA rekombinan memanfaatkan bakteri tertentu. (2) produksi komersial
substansi imunogenik untuk memproduksi vaksin dengan DNA rekombinan yang
lebih baik dan lebih aman dibandingkan dengan antigen konvensional yang
berasal dari bakteri atau mikroorganisme lain yang patogen. Selanjutnya Muladno
(2002) menjelaskan, bahwa dengan tersedianya bioteknologi rekayasa genetika
yang dilahirkan pada tahun 1973, telah memungkinkan manusia untuk
mengisolasi gen (serangkaian molekul DNA) serta memanipulasinya dan
kemudian memindahkan gen tersebut dari satu organisme ke organisme lain.
Perbedaan teknologi ini dibanding dengan teknologi lainnya adalah bahwa
teknologi ini memanfaatkan mahluk hidup sebagai komponen utamanya. Mahluk
hidup yang digunakan bisa berasal dari mikroorganisme, tanaman atau hewan.
Peranan bioteknologi dirasakan semakin bertambah besar dalam menunjang
kegiatan pembangunan industri di berbagai sektor, terutama sektor kesehatan dan
pertanian termasuk sub sektor peternakan. Cakupan bioteknologi ini sangat luas
baik yang baru dalam tahap penelitian maupun yang sudah dapat diaplikasikan. Di
bidang kesehatan dan kedokteran, telah ditemukan berbagai jenis obat-obatan
baru hasil pengembangan bioteknologi modern, antara lain insulin bagi pasien
diabetes yang kini dapat diperoleh lebih mudah dan lebih murah harganya,
hormon pertumbuhan manusia dan vaksin Hepatitis B (Winarno dan Agustinah
berkembang. Virus Hepatitis B merupakan penyebab utama hepatitis akut yang
dapat berlanjut menjadi kronis, sirosis dan kanker hati. Komplikasi akibat virus
ini telah mengakibatkan sekitar 1 juta orang meninggal setiap tahun (Kimura et al.
2005).
Selubung virus hepatitis B (hepatitis B virus envelope) terdiri dari membran
glikoprotein dimana terdapat 3 bagian protein permukaan yaitu antigen pre-S1
(119 asam amino), pre-S2 (55 asam aminio) dan S (226 asam amino) (Yamada et
al. 2001; Jaoude dan Sureau 2005, Barrera et al. 2005). Beberapa ahli
menggolongkan ketiga protein tersebut sebagai protein kecil (small), sedang
(middle) dan besar (large). Antigen S telah digunakan secara luas sampai saat ini
sebagai vaksin konvensional. Menurut Hu et al. (2004a), asam amino ke 139-147
pada bagian S merupakan epitop utama pada protein S tersebut dan asam amino
Pre-S1 dan Pre-S2 masih dikaji tingkat immunogenisitasnya melalui serangkaian
diagnosa (Maruyama et al. 2000).
Proyek immunisasi massal di Lombok menunjukkan penggunaan vaksin
konvensional mampu menurunkan prevalensi Hepatitis B hanya sampai 70%
(Mulyanto et al. 2002). Hasil immunisasi Hepatitis B tersebut belum optimal,
kemungkinan hal ini disebabkan oleh vaksin konvensional tersebut (Korean
Green Cross) berasal dari plasma darah orang asing sehingga tidak mampu
menstimulasi munculnya antibodi spesifik yang mampu melawan virus Hepatitis
B yang terdapat di Indonesia.
Berdasarkan permasalahan tersebut, penelitian ini dilaksanakan untuk
melakukan rekayasa terhadap gen penyandi antigen permukaan Hepatitis B untuk
menghasilkan antigen rekombinan Hepatitis B bagian S (HBsAg100) pada E. coli.
Bagian gen penyandi epitop yang bersifat hidrophilik (dari asam amino nomor
100-164) dilaporkan dapat meningkatkan ekspresi pada E. coli. Selain itu, gen
penyandi HBsAg100 digabung dengan gen penyandi enzim
gluthation-S-transferase (GST) dapat meningkatkan ekspresi maupun solubilitas antigen yang
sangat penting untuk aktivitas maupun proses purifikasi. Gen penyandi HBsAg100
adalah gen yang diisolasi dari virus Hepatitis B sub tipe adw sebagai sub tipe
utama di Indonesia yang digunakan untuk membuat model dalam memproduksi
4
protein antibodiyang sekaligus dapat dimanfaatkan sebagai kandidat vaksin galur
lokal yang mampu memberikan respon antibodi yang spesifik sesuai dengan
genetik virus Hepatitis B yang terdapat di Indonesia.
Protein HBsAg100 rekombinan yang dihasilkan dengan teknologi rekayasa
DNA menggunakan bakteri ini diharapkan dapat menggantikan metode produksi
vaksin konvensional dari plasma yang banyak memiliki kelemahan, diantaranya,
rendahnya imunogenisitas, sumber plasma yang terus berkurang (karena jumlah
penderita penyakit Hepatitis B menurun sejalan dengan keberhasilan program
vaksinasi), serta kekhawatiran adanya kontaminasi penyakit lain (terutama HIV)
pada serum donor. Antigen ini diharapkan dapat menghasilkan kandidat vaksin
rekombinan Hepatitis B yang sesuai dengan genetik virus tersebut di Indonesia,
karena gen penyandi antigen tersebut diisolasi dari virus HB yang terdapat di
Indonesia.
Perumusan Masalah
Perlunya dikembangkan bioteknologi dalam memenuhi kebutuhan manusia
dalam perkembangan dunia global dapat dipertimbangkan berdasarkan
permasalahan sebagai berikut:
1 Bioteknologi merupakan cabang ilmu pengetahuan yang dibutuhkan dalam
berbagai aspek kehidupan manusia.
2 Perkembangan bioteknologi yang sangat pesat perlu dimanfaatkan secara
maksimal.
3 Aplikasi bioteknologi di Indonesia masih terbatas, sehingga perlu untuk digali
dan dikembangkan, khususnya di bidang peternakan yang meliputi
bioteknologi reproduksi, pakan ternak dan bioteknologi molekuler di bidang
produksi ternak dan kesehatan hewan seperti pembuatan bahan obat dan bahan
vaksin.
4 Di bidang kesehatan hewan, penggunaan vaksin konvensional yang
mempunyai banyak kelemahan bisa diatasi dengan pembuatan bahan vaksin
Tujuan Penelitian
1 Membuat model plasmid rekombinan untuk memproduksi protein HBsAg100
rekombinan.
2 Menghasilkan klon pembawa gen penyandi HBsAg100 yang telah dikloning
dengan plasmid yang khusus digunakan untuk ekspresi (pGEX-4T-2).
3 Produksi dan isolasi protein HBsAg100-GST rekombinan.
Manfaat Penelitian
Penelitian ini bermanfaat untuk produksi substansi antigenik dalam rangka
memproduksi vaksin dengan DNA rekombinan yang lebih baik dan lebih aman
dibandingkan dengan antigen konvensional yang berasal dari bakteri atau
mikroorganisme lain yang patogen.
Ruang Lingkup
1 Mengisolasi gen penyandi HBsAg100 dari virus Hepatitis B sub tipe adw yang
merupakan sub tipe utama di Indonesia.
2 Memperbanyak HBsAg100 dengan PCR.
3 Membuat plasmid rekombinan melalui ligasi HBsAg100 dengan plasmid
pGEX-4T-2.
4 Melakukan transformasi plasmid rekombinan terhadap E. coli DH5α untuk
kloning, dilanjutkan dengan skrining klon yang membawa plasmid
rekombinan. Sekuensing dilakukan untuk memastikan tidak terdapat mutasi
pada gen target, kemudian dilanjutkan lagi dengan transformasi ke dalam E.
coli BL21 untuk menghasilkan protein HBsAg100-GST rekombinan.
5 Melakukan pengujian antigenisitas protein antigen S rekombinan pada mencit
BALB/c dengan teknik ELISA. Melakukan pengujian imunogenisitas protein
HBsAg100-GST rekombinan melalui respon mencit BALB/c yang diimunisasi
dengan HBsAg100-GST, kemudian melakukan pengambilan serum dan
6
Kerangka Pemikiran
Indonesia merupakan daerah hiperendemik penyakit Hepatitis B. Harga obat
yang digunakan untuk mengatasi penyakit tersebut sangat mahal, sehingga
vaksinasi merupakan metode yang lebih murah dan efektif. Vaksin konvensional
yang digunakan di Indonesia saat ini (Korean Green Cross) merupakan vaksin
yang dihasilkan dari plasma darah orang asing. Virus Hepatitis B merupakan virus
DNA yang memiliki enzim polymerase dengan kecermatan rendah, maka
frekuensi terjadinya mutasi cukup tinggi, sehingga memungkinkan terjadinya
perbedaan sekuen dari gen virus Hepatitis B yang ada di luar negeri dengan virus
Hepatitis B yang terdapat di Indonesia. Penggunaan vaksin galur luar negeri akan
menstimulasi munculnya tanggap kebal (antibodi) spesifik terhadap virus
Hepatitis B yang ada di luar negeri. Sebaliknya, vaksin tersebut kemungkinan
kurang efektif untuk menghasilkan antibodi yang spesifik untuk melawan virus
Hepatitis B yang ada di Indonesia. Oleh karena itu, produksi protein HBsAg100
rekombinan sebagai kandidat vaksin Hepatitis B yang terdapat di Indonesia sangat
mendesak untuk dilakukan sehingga dihasilkan respon antibodi yang mampu
melawan virus tersebut. Berkaitan dengan hal ini, maka sangat perlu dilakukan
penelitian tentang “Produksi Protein Rekombinan HBsAg100-GST sebagai Model
Imunogen untuk Menghasilkan Antibodi pada Mencit”.
Masalah utama untuk menghasilkan antigen permukaan Hepatitis B
menggunakan bakteri E. coli adalah rendahnya tingkat ekspresi. Hal ini
disebabkan karena ekspresi gen penyandi antigen permukaan Hepatitis B
terhambat oleh adanya bagian gen up-stream yang menghasilkan protein bagian
dari antigen yang bersifat hidrophobik (Sheu dan Lo 1995). Selain itu, hasil
penelitian pendahuluan dalam rangkaian penelitian ini menunjukkan bahwa
antigen permukaan Hepatitis B bersifat toksik bagi inang (E. coli). Oleh karena
itu, penelitian yang mengarah pada optimalisasi ekspresi antigen pada E. coli
sangat perlu dilakukan. Pada penelitian ini, optimalisasi ekspresi dilakukan
dengan melakukan kloning dan ekspresi gen yang menghasilkan antigen
hidrophilik yang tetap mempertimbangkan utuhnya bagian-bagian epitop dari
antigen tersebut. Disamping itu, sifat toksik antigen tersebut akan diatasi dengan
mencukupi untuk menghasilkan antigen yang memadai. Pencegahan ekspresi dini
tersebut akan dilakukan dengan pengayaan media melalui penambahan glukosa
untuk mencegah bakteri E. coli mengalami kekurangan nutrisi yang merupakan
penyebab bakteri tersebut mengekspresikan berbagai jenis enzim (termasuk
protein rekombinan) untuk melakukan metabolisme terhadap media.
Tidak adanya protein disulfide isomerase pada bakteri E. coli menyebabkan
protein rekombinan yang diekspresi menggunakan bakteri ini tidak mampu
mengalami folding secara sempurna. Hal ini berdampak pada rendahnya kelarutan
serta aktifitas dari protein yang dihasilkan. Strategi yang banyak dilakukan untuk
mengatasi masalah tersebut adalah melakukan penggabungan dengan
gluthatione-S-transferase (GST) (Vikis dan Guan 2000; Koschorreck et al. 2005). Oleh karena
itu, penggabungan antigen permukaan Hepatitis B yang akan diproduksi dengan
GST sangat perlu dilakukan. Disamping itu, penggabungan dengan GST juga
akan sangat mempermudah proses pemurnian. Adanya kolum GSTrap yang
tersedia secara komersial merupakan tindak lanjut dihasilkannya plasmid
pGEX-4T-2 yang akhir-akhir ini sangat popular digunakan untuk produksi protein
rekombinan pada bakteri E. coli. Hal ini disebabkan karena plasmid tersebut
merupakan plasmid yang mampu menghasilkan protein rekombinan dalam jumlah
TINJAUAN PUSTAKA
Virus Hepatitis B dan Antigen Permukaan
Menurut Dayal dan Maldonado (1998), Virus Hepatitis B masuk dalam
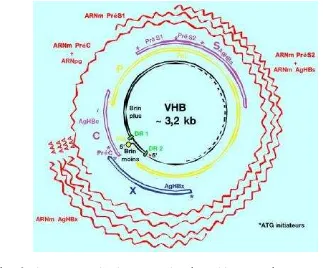
famili virus Hepadna dan mempunyai ukuran genome yang terkecil diantara
semua virus DNA hewan, yaitu dengan ukuran panjang 3,2 kb. Menurut Stannard
(1995), virus Hepatitis B menyebabkan infeksi kronis dan akut pada hati manusia.
Infeksi akut biasanya sampai beberapa bulan saja, sedangkan infeksi kronis bisa
mencapai bertahun-tahun bahkan bisa sampai selama hidupnya. Diameter total
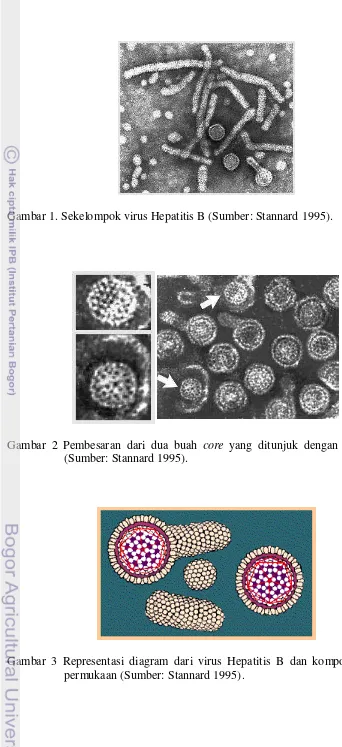
dari virus Hepatitis B adalah 42 nm, sedangkan diameter core atau
nucleocapsidnya adalah 27 nm. Core dilapisi oleh mantel (outer coat) yang
tebalnya sekitar 4 nm. Protein yang terdapat pada mantel disebut surface antigen
atau HBsAg. HBsAg ini kadang-kadang berkembang memanjang membentuk
ekor (tubular) pada salah satu sisi dari partikel virus tersebut. Gambaran virus
Hepatitis B yang lebih jelas, diilustrasikan pada Gambar 1, Gambar 2, Gambar 3
dan Gambar 4.
Virus Hepatitis B (VHB) telah menginfeksi sampai ratusan juta orang di
seluruh dunia dan sekitar 20 juta orang terinfeksi setiap tahun, dan sekitar 78%
penderita berdomisili di Asia (Joshi dan Kumar 2001). Di Indonesia, jumlah
penderita penyakit tersebut mencapai sekitar 15 juta orang. VHB merupakan
penyebab utama sirosis (pengerasan hati) dan kanker hepatoseluler (Human
Hepatocellular Carcinoma) yang merupakan salah satu penyakit terganas
penyebab kematian di seluruh dunia. Jenis kanker ini telah menyebabkan
Gambar 1. Sekelompok virus Hepatitis B (Sumber: Stannard 1995).
Gambar 2 Pembesaran dari dua buah core yang ditunjuk dengan tanda panah (Sumber: Stannard 1995).
11
Gambar 4 Ilustrasi virus Hepatitis B dengan capsid dan internal density yang tampak pada irisan melintang (Sumber: Dryden et al. 2006).
Data dari berbagai penelitian menunjukkan bahwa sebagian besar infeksi
virus Hepatitis B yang bersifat menetap, timbul sebagai akibat infeksi pada waktu
bayi dan anak-anak. Makin muda usia seseorang terkena infeksi virus tersebut,
maka akan lebih besar kemungkinannya untuk menderita infeksi virus Hepatitis B
yang menetap, sehingga lebih besar jumlah resiko untuk menjadi sirosis hati dan
kanker hati primer dikemudian hari (Mulyanto et al. 2002).
Penelitian tentang sebaran geografis virus ini menunjukkan bahwa virus
tersebut tersebar di seluruh dunia. Namun, prevalensi tertinggi ditemukan di Asia
Tenggara dan Sub-sahara Afrika. Mulyanto et al. (1997) membagi zona distribusi
sub tipe virus Hepatitis B di Indonesia berdasarkan perbedaan epitope pada
HBsAg menjadi 4 bagian: (1) zona adw yang merupakan sub tipe utama di
Indonesia meliputi Sumatera, Jawa, Kalimantan bagian Selatan, Bali, Lombok,
Ternate dan Morotai, (2) zona ayw yang meliputi bagian barat dari Nusa Tenggara
dan Maluku, (3) zona adr meliputi Papua, dan (4) campuran antara berbagai sub
Virus Hepatitis B merupakan virus DNA untai ganda dengan panjang
genome mencapai 3.2-3.3 kilo pasangan basa (kpb). Virus yang termasuk famili
hepadnaviridae tersebut memiliki genome yang terbungkus oleh glycoprotein.
Siklus replikasi virus ini dimulai dengan melekatnya protein selubung tersebut
pada hepatosit. Di dalam inti sel hati, sintesis DNA virus disempurnakan dimana
genome virus tersebut diubah menjadi cccDNA (covalently closed circular DNA).
cccDNA inilah yang akan menjadi template untuk sintesis RNA yang kemudian
akan diubah menjadi DNA virus (Lok dan McMahon 2001).
Selubung virus Hepatitis B (Hepatitis B virus envelope) terdiri dari
membran glikoprotein dimana terdapat 3 bagian protein permukaan yaitu antigen
pre-S1 (119 asam amino), pre-S2 (55 asam aminio) dan S (226 asam amino)
(Jaoude dan Sureau 2005; Barrera et al. 2005). Beberapa ahli menggolongkan
ketiga protein tersebut sebagai protein kecil (small), sedang (middle) dan besar
(large). Antigen pre-S1 memiliki beberapa epitop yang memiliki daya
immunogenik (Hu et al. 2004). Antigen bagian pre-S1 ini dibutuhkan oleh virus
Hepatitis B untuk melakukan infeksi pada korban (Barrera et al. 2005). Menurut
Deng et al (2005), asam-asam amino Pre-S1 pada nomor 21-47 merupakan epitop
yang berfungsi untuk melekatnya virus pada jaringan hati. Oleh karena itu, protein
bagian ini memiliki peranan yang sangat penting untuk siklus virus Hepatitis B.
Antigen pre-S2 diduga mempunyai tingkat imunogenisitas lebih tinggi
dibandingkan HBsAg (Ji et al. 2005) terutama 26 asam amino pada ujung N
(Joung et al. 2004). Antigen pre-S2 mempunyai peranan sangat penting, hal ini
telah dibuktikan secara nyata melalui serangkaian diagnosa (Maruyama et al.
2000). Menurut Hu et al. (2004a), HBsAg telah digunakan secara luas sampai saat
ini sebagai vaksin konvensional, asam-asam amino ke 139-147 pada bagian S
merupakan epitop utama pada protein S (HBsAg). Namun demikian, penghilangan
epitop ini masih tetap dapat memberikan reaksi antigenisitas yang menunjukkan
bahwa masih terdapat epitop lain selain epitop yang terletak pada asam-asam
13
Struktur DNA dan Genome Virus Hepatitis B
Menurut Winarno dan Agustinah (2007), DNA adalah deoxyribo nucleic
acid, yaitu sebuah asam nukleat yang terdiri atas sejumlah nukleotida yang diatur
sedemikian rupa sehingga berbentuk single strand. Biasanya dua buah utas DNA
saling melingkar satu sama lain untuk membentuk sebuah double helix (heliks
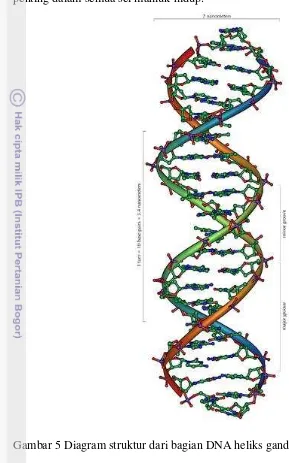
ganda), seperti ditunjukkan oleh Andre (2006) pada Gambar 5. Muladno (2002)
menegaskan bahwa untuk membentuk rangkaian molekul DNA heliks ganda, basa
nitrogen dari setiap nukleotida dalam satu rangkaian akan berpasangan dengan
basa nitrogen dari setiap nukleotida pada rangkaian lainnya melalui ikatan
hidrogen. Pengikatan basa nitrogen dari masing-masing nukleotida tersebut sangat
spesifik. Basa A (Adenine) dari satu nukleotida selalu berikatan dengan basa T
(Thymine) dari nukleotida lainnya, sedangkan basa G (Guanine) selalu
berpasangan dengan basa C (Cytosine). Pasangan A dan T terbentuk dengan dua
ikatan hidrogen, sedangkan pasangan G dan C terbentuk dengan tiga ikatan
hidrogen. Oleh karena itu, pasangan G-C lebih stabil daripada pasangan A-T.
Winarno dan Agustinah (2007) menjelaskan, bahwa gugus basa DNA terdiri
atas empat senyawa berikut: sitosin, adenin, guanin atau timin. Gugus gulanya
adalah deoksiribosa. DNA terdapat di dalam kromosom prokariot dan eukariot
dan di dalam mitokondria eukariot. DNA merupakan materi kebakaan atau
keturunan di hampir semua organisme hidup yang mampu memperbanyak dirinya
sendiri dalam pembelahan inti. Winarno dan Agustinah (2007) menjelaskan
bahwa hipotesa Watson dan Crick merupakan hipotesa berdasarkan X-ray
crystallography yang mengusulkan bahwa DNA merupakan suatu heliks ganda
dari dua uliran rantai fosfat dan gula yang saling bergantian, dengan gula yang
terkait oleh sepasang basa. Selanjutnya Muladno (2002) menjelaskan, bahwa
Watson dan Crick akhirnya memperoleh Nobel dalam bidang biologi modern pada
tahun 1953 setelah menemukan bukti bahwa struktur DNA adalah heliks ganda.
Penemuan ini merupakan tonggak sejarah yang penting terhadap munculnya
teknologi rekayasa genetika yang lahir 20 tahun kemudian, yaitu tahun 1973.
DNA bersama-sama protein (histone) dan molekul ribo nucleic acid (RNA),
susunan yang sangat rumit membentuk kromosom yang merupakan komponen
penting dalam semua sel mahluk hidup.
Gambar 5 Diagram struktur dari bagian DNA heliks ganda (Sumber: Andre 2006).
Melalui suatu proses kimia, kromosom dapat dikeluarkan dari inti sel.
Selanjutnya, protein yang berikatan dengan DNA dilisiskan dengan enzim
proteinase, sedangkan RNA yang masih berada di sekitar DNA dikatalisis atau
diurai dengan enzim RNAse. Selanjutnya, DNA yang telah terbebas dari protein
(histone) dan RNA siap direkayasa atau dimanipulasi dalam teknologi rekayasa
genetika (Muladno 2002). Old dan Primrose (1989) menjelaskan, bahwa istilah
manipulasi gen dapat diterapkan pada beberapa macam teknik genetika in-vivo
15
yang tepat untuk istilah manipulasi gen sebagai akibat adanya peraturan
Pemerintah untuk mengendalikannya. Di Inggris, manipulasi gen didefinisikan
sebagai pembentukan kombinasi baru materi yang dapat diturunkan dengan
melakukan penyisipan (insertion) molekul-molekul asam nukleat, yang dihasilkan
dengan cara apapun di luar sel, ke dalam suatu virus, plasmid bakteri atau sistem
pembawa lainnya yang memungkinkan terjadinya penggabungan ke dalam
organisme inang secara tidak alami tetapi selanjutnya mampu melakukan
penggandaan lagi. Definisi resmi ini menekankan penggandaan molekul asam
nukleat asing (asam nukleat ini hampir selalu DNA) di dalam tubuh organisme
inang yang berbeda. Kemampuan untuk melintasi penghalang spesies alami dan
memasukkan gen-gen dari organisme apapun ke dalam suatu organisme inang
yang tidak berhubungan merupakan satu ciri penting manipulasi gen. Ciri penting
kedua berupa kenyataan bahwa relatif sepotong kecil DNA tertentu digandakan
dalam tubuh organisme inang.
Setiap organisme mempunyai sebuah genome yang mengandung semua
informasi biologik yang diperlukan untuk membangun dan memelihara kehidupan
organisme tersebut. Informasi biologik yang terkandung di dalam genome dikode
oleh DNA yang terkandung di dalam genome yang dibagi ke dalam unit-unit
khusus yang disebut gen (Barnum 2005). Menurut Winarno dan Agustinah (2007),
genome dalam arti sederhana berarti satu set lengkap mengandung informasi
genetika yang dimiliki oleh suatu organisme. Selanjutnya dijelaskan oleh
Muladno (2002), bahwa setiap mahluk hidup mempunyai sel yang di dalam inti
selnya terdapat kromosom dengan jumlah berbeda-beda untuk setiap mahluk
hidup. Manusia mempunyai 23 pasang kromosom dalam setiap intinya. Sapi
mempunyai 30 pasang kromosom, lalat buah Drosophila mempunyai empat
pasang kromosom, bakteri E. coli mempunyai satu kromosom. Beberapa peneliti
lain menambahkan, bahwa virus hepatitis B mempunyai 1 kromosom (Dayal dan
Maldonado 1998; Mason et al. 1998; Burda et al. 2001; Muljono dan
Soemohardjo 2003; Anzola 2004; Wagner et al. 2004; Beck dan Nassal 2007;
GenBank 2008; Nurainy et al. 2008).
Menurut Muladno (2002), total kromosom dalam inti sel dinamakan
bisa dikatakan bahwa genom manusia terdiri atas 23 pasang kromosom, genom
sapi terdiri atas 30 pasang kromosom, dan seterusnya. Apabila DNA dari genom
tersebut direntang secara linear, maka ukuran panjang rentangan DNA pada
genom tersebut berbeda-beda pada setiap organisme seperti dijelaskan pada Tabel
1. Rentangan DNA dari genom tersebut disebut genomic DNA.
Berdasarkan data pada Tabel 1, maka dapat dijelaskan bahwa besarnya
ukuran genom tidak mencerminkan besarnya ukuran makhluk hidup, seperti
apabila membandingkan ukuran tubuh manusia dengan ukuran tubuh tikus yang
sangat jauh berbeda, tetapi keduanya mempunyai ukuran genom yang hampir
sama. Demikian pula, ukuran tubuh cacing yang jauh lebih besar dari ukuran
tubuh lalat buah Drosophila, tetapi ukuran genome cacing lebih sedikit daripada
ukuran genome lalat buah Drosophila.
Tabel 1 Ukuran genome dari beberapa makhluk hidup dalam bentuk haploid
Jenis mahluk hidup Ukuran genome Lalat buah Drosophila 140 000 000** Tikus 3 000 000 000** Manusia 3 300 000 000** Sumber: *Nurainy et al. 2008, **Lewin 1990.
Muladno (2002) menyatakan, bahwa virus dan bakteri merupakan makhluk
hidup yang sederhana dan mempunyai ukuran genome yang kecil sehingga
organisasi genomenya juga sederhana. Oleh karena itu, kedua makhluk hidup ini
menjadi “bahan utama” untuk penelitian-penelitian dalam bidang genetika molekuler. Seperti telah diuraikan di atas, bahwa virus Hepatitis B mempunyai
ukuran genome yang terkecil diantara semua virus DNA, yang menurut beberapa
peneliti (Dayal dan Maldonado 1998; Burda et al. 2001; Muljono dan
Soemohardjo 2003; Anzola 2004; Wagner et al. 2004; Beck dan Nassal 2007;
17
3.2 kb. Dayal dan Maldonado (1998) menjelaskan bahwa virus hepatitis B
mempunyai ciri-ciri sebagai berikut:
1 Genome virus hepatitis B yang menurut mereka ditemukan oleh William S.
Robinson dari Stanford University School of Medicine merupakan genome
yang terkecil dengan ukuran panjang 3,2 kb. Diagram organisasi genome
virus Hepatitis B diilustrasikan pada Gambar 7.
2 Informasi genetik dibawa melalui DNA rantai ganda.
3 DNA penyusun genome terdiri atas rantai yang lebih panjang yang disebut
rantai negatif (-) lengkap, dan rantai positif (+) tidak lengkap, terdapat sekitar
15-50% merupakan rantai tunggal.
4 Genome sirkuler dipertahankan dalam bentuk longgar oleh pasangan basa
dari sekuen overlap 240 nukleotida dan Direct Repeats (DR) pendek antara
kedua rantai pada ujung 5’.
5 Ujung 5’ dari rantai negatif (-) mengikat protein secara kovalen, sedangkan
ujung 5’ dari rantai positif (+) mengikat oligoribonukleotida sebagai RNA
primer.
6 Nukleokapsid berbentuk icosahedral.
7 Nukleokapsid dikelilingi oleh amplop lipid.
8 Diameter Nukleokapsid 27 nm.
9 Melalui gambar mikrograf elektron, virus tampak spherical (bentuk bola)
dengan diameter total 42 nm.
10 Mengandung 4 Overlapping Open Reading Frames (ORF): S, C, P, dan X
yang disandi pada rantai DNA (-).
11 Nomer Triangulasi=3 (T=3).
12 Gen S terdiri atas 3 bagian: yaitu pre-S1, pre-S2, dan S. Tiga macam protein
yang berbeda diproduksi oleh kombinasi dari ke tiga gen tersebut dalam
kombinasi yang berbeda: protein L (large protein), yaitu protein pada amplop
yang dibuat berdasarkan pengkodean dari gen-gen pre-S1, pre-S2, dan S;
protein M (middle-sized proteins/medium surface protein) pada amplop yang
dikode oleh gen pre-S2 dan gen S; dan protein S yang merupakan komponen
Gambar 6 Diagram organisasi genome virus hepatitis B (Sumber: Wagner 2004).
Nurainy et al. (2008) melalui risetnya yang berjudul Genetic Study of
Hepatitis B Virus in Indonesia Reveals a New Subgenotype of Genotype B in East
Nusa Tenggara dengan menggunakan isolat Virus Hepatitis B 2059Java, berhasil
menemukan sekuen genom lengkap Virus Hepatitis B (Lampiran 2). Selanjutnya
dijelaskan, bahwa sekuen genome lengkap tersebut tersusun atas 4 overlapping
open reading frames yang masing-masing mempunyai sekuen sebagai berikut:
1 Gen P (sekuen:2307-3215, 1-1623); yang menyandi pembentukan enzim-enzim
DNA polymerase, reverse trancriptase, dan RNAse.
2 Gen S yang terdiri atas 3 bagian, yaitu: gen pre-S1, gen pre-S2, dan gen S. Tiga
jenis protein amplop VHB yang berbeda diproduksi berdasarkan pengkodean
dari kombinasi tiga gen-gen ini dalam kombinasi yang berbeda. (1) Protein L
(large protein) dikode oleh kombinasi dari gen-gen pre-S1, pre-S2, dan S
(sekuen: 2848-3215, 1-835). (2) Protein M (medium protein) dikode oleh
kombinasi dari gen pre-S2 dan gen S (sekuen: 3205-3215, 1-835). (3) Protein S
(HBsAg) dikode oleh gen S (155-835).
3 Gen X (sekuen: 1374-1838); yang menyandi pembentukan protein X yang
peranannya belum diketahui secara pasti, tetapi diduga berperan dalam aktivasi
19
rekombinan (Barnum 2005). Selanjutnya, Winarno dan Agustinah (2007)
menjelaskan, bahwa teknologi rekayasa genetika merupakan kegiatan
bioteknologi modern dengan teknologi DNA rekombinan (rDNA) untuk
melakukan pemindahan atau transfer suatu sifat tertentu yang dibawa gen, yang
tersusun dalam DNA, dari suatu spesies yang sama atau berbeda untuk
menghasilkan spesies baru yang lebih unggul. Muladno (2002) juga menyatakan,
seiring dengan kemajuan teknologi molekuler, perpindahan gen dapat terjadi antar
organisme yang sama sekali tidak berkerabat dekat, misalnya gen manusia
dipindahkan ke bakteri atau gen manusia dipindahkan ke hewan ternak.
Perpindahan gen tersebut mengakibatkan terbentuknya molekul DNA yang
berasal dari sumber yang berbeda dapat digabungkan menjadi DNA rekombinan.
Teknik menggabungkan molekul DNA tersebut dikenal sebagai Teknik DNA
Rekombinan.
Biasanya DNA rekombinan merupakan gabungan antara DNA vektor yang
merupakan molekul DNA yang dapat mereplikasi diri dan DNA asing yang
biasanya berupa gen dari suatu mahluk hidup. Vektor tersebut berfungsi sebagai
pembawa DNA asing yang berasal dari suatu organisme untuk dipindahkan ke
dalam organisme lain. Gen yang terkandung pada DNA rekombinan di dalam
organisme resipien diharapkan dapat diekspresikan untuk menghasilkan protein
(Muladno 2002). Menurut Glick dan Pasternak (1994), teknologi DNA
rekombinan, juga disebut kloning gen atau kloning molekuler, adalah suatu istilah
yang mencakup sejumlah protokol percobaan yang bertujuan untuk transfer
informasi genetik (DNA) dari suatu organisme ke organisme yang lain. Percobaan
DNA rekombinan biasanya mengikuti prosedur sebagai berikut:
1 DNA target (DNA asing, DNA insert, DNA klon) dari organisme donor
atau DNA vektor (vektor kloning) untuk membentuk suatu bentuk baru yang
disebut molekul DNA rekombinan (rDNA).
2 DNA rekombinan tersebut ditransfer ke dalam sel inang. Proses introduksi
DNA rekombinan ke dalam suatu sel inang atau bakteri disebut transformasi.
3 Selanjutnya sel-sel bakteri tersebut dikultur dan diseleksi atau dimurnikan,
kemudian diisolasi.
4 Sel-sel bakteri inang yang telah dimurnikan dan diisolasi, siap dikultur atau
dikembangbiakkan untuk memproduksi protein spesifik yang dikode oleh DNA
klon yang terkandung dalam DNA rekombinan.
Pembuatan DNA rekombinan memerlukan bantuan dua macam enzim.
Pertama, enzim endonuclease (restriction enzyme) berperan sebagai pemotong
molekul DNA. Kedua, enzim ligase berfungsi untuk menggabungkan
molekul-molekul DNA yang telah dipotong oleh enzim restriksi (Muladno 2002; Barnum
2005).
Ilustrasi tahapan pembuatan DNA rekombinan pada Gambar 8 menunjukkan
bahwa enzim endonuclease yang digunakan untuk memotong kedua sumber DNA
adalah BamHI. Enzim restriksi ini memotong kedua molekul DNA tersebut pada
lokasi yang sama dengan membentuk potongan sticky end atau kohesif.
Selanjutnya enzim ligase DNA menggabungkan kedua molekul DNA tersebut
dengan ikatan kovalen menjadi satu molekul DNA rekombinan.
Muladno (2002) menegaskan bahwa pada prinsipnya kloning DNA adalah
proses penggandaan jumlah DNA rekombinan melalui proses perkembangbiakan
sel bakteri (biasanya E. coli). Hal ini dilakukan dengan memasukkan DNA
rekombinan yang dihasilkan dari proses penggabungan tersebut di atas ke dalam
sel E. coli. Selanjutnya sel ini diinkubasi pada suhu optimal sehingga sel dapat
berkembangbiak secara eksponensial. Masuknya molekul DNA rekombinan ke
dalam sel akan mengubah fenotip sel tersebut, sehingga proses pemasukan
molekul DNA ke dalam sel juga disebut transformasi. Sel yang digunakan dalam
proses transformasi ini biasanya disebut dengan sel kompeten. Plasmid
merupakan molekul kecil yang berukuran sekitar lima ribu pasang basa yang
terdapat di dalam sel bakteri Escherichia coli, posisinya terpisah dengan
21
kromosom, kebanyakan berupa rangkaian molekul DNA untai ganda dan biasanya
berbentuk bulat. Selanjutnya Glick dan Pasternak (1994) menerangkan, bahwa ori
adalah sekuen nukleotida yang merupakan tempat diawalinya atau dimulainya
sintesis DNA pada saat replikasi.
Barnum (2005) menjelaskan, ada beberapa cara dalam melakukan kloning
DNA, dan metode yang digunakan bervariasi tergantung pada tipe DNA, tipe sel
inang, dan tujuan akhir dari kloning DNA. Contohnya, tipe vektor yang
digunakan untuk kloning akan tergantung kepada apakah DNA klon akan tetap
berada di dalam vektor atau akan disisipkan ke dalam kromosom sel inang.
Vaksin Hepatitis B
Menurut Dayal dan Maldonado (1998), vaksin hepatitis B yang pertama kali
mendapat lisensi (pada tahun 1981) adalah Heptavax-B dan telah dipasarkan oleh
Merck Sharp dan Dhome. Vaksin tersebut diperoleh dari hasil ekstraksi dan
pemurnian antigen HBsAG dari serum penderita hepatitis B kronis. Selanjutnya
dijelaskan bahwa vaksin rekombinan atau sebagai hasil rekayasa genetika yang
telah berhasil diproduksi secara komersial adalah Recombivax HB Chiron Corp
dan Merck serta Engerix-B oleh SmithKline Biologicals. Vaksin rekombinan
tersebut diproduksi dari hasil kloning gen HBsAg yang terdapat di dalam yeast.
Joung et al. (2004) menyatakan, hampir semua vaksin hepatitis B konvensional
yang sudah mendapat lisensi saat ini adalah vaksin yang dihasilkan dari plasma.
Namun, keberhasilan program imunisasi menyebabkan pasien hepatitis B yang
akan menjadi sumber vaksin tersebut semakin berkurang yang berakibat pada
semakin terbatasnya darah yang dapat digunakan sebagai sumber vaksin. Oleh
sebab itu produksi vaksin hepatitis B dengan menggunakan plasma semakin sulit
dilakukan. Kekhawatiran terhadap adanya kontaminan pada darah terutama oleh
virus berbahaya seperti HIV, menimbulkan kekhawatiran tersendiri untuk
menggunakan vaksin yang bersumber dari plasma tersebut.
Menurut Mulyanto et al. (1997), Indonesia merupakan daerah endemik
sedang sampai tinggi untuk penyakit hepatitis B, sehingga WHO menghimbau
untuk segera melaksanakan usaha pencegahan. Pengobatan terhadap penderita
vaksinasi merupakan tindakan yang lebih tepat. Vaksinasi secara besar-besaran
dinilai efektif untuk mencegah terjadinya penyakit ini. Selanjutnya Mulyanto et
al. (2002) menyatakan, bahwa pada tahun 1987, Pemerintah Indonesia
menjadikan Pulau Lombok sebagai model immunisasi massal Hepatitis B pertama
di dunia. Hal ini disebabkan karena tingkat endemik penyakit tersebut di Pulau
Lombok sangat tinggi. Hasil proyek percontohan tersebut cukup menggembirakan
sehingga pemerintah mulai memperluas program immunisasi ke 4 propinsi yang
lain di tahun 1991 dan kemudian ke 6 propinsi lainnya pada tahun 1992. Saat ini
pemerintah Indonesia telah mengintegrasikan vaksinasi Hepatitis B untuk balita
ke dalam Program Pengembangan Imunisasi (Extended Program of
Immunization).
Berbagai upaya untuk menghasilkan vaksin yang mengandung antigen
rekombinan telah dilakukan, diantaranya dengan memproduksi antigen
menggunakan sel tanaman maupun ragi. Keberhasilan ekspresi antigen tersebut
telah dilaporkan melalui kultur sel tembakau (Kumar et al. 2003), maupun ragi
(Maruyama et al. 2000; Lu et al. 2002; Ddeman & Zyl 2003; Hu et al. 2004).
Meskipun vaksin tersebut memberikan hasil yang sangat memuaskan, sistem
produksi menggunakan hewan maupun tanaman terhambat oleh lamanya waktu
yang diperlukan, terjadi variasi pada produk akhir, terkontaminasi oleh
bahan-bahan kimia pertanian serta kesulitan dalam meningkatkan hasil produk akibat
rumitnya faktor-faktor regulasi. Penggunaan bakteri E. coli menjadi pilihan
terakhir karena waktu yang lebih singkat, harga media lebih murah serta teknologi
pembiakan maupun komponen-komponen yang dibutuhkan untuk optimalisasi
transkripsi maupun translasi telah dikuasai (Ali 2006).
Produksi bagian yang sederhana dari antigen permukaan Hepatitis B dengan
menggunakan bakteri E. coli bisa dilakukan, namun demikian berbeda dengan
hasil ekspresi yang dilakukan dengan induk semang hewan maupun tumbuhan
yang dapat menghasilkan produksi yang tinggi, ekspresi menggunakan E. coli
menghasilkan produk yang sedikit. Hal ini disebabkan oleh sifat antigen
permukaan Hepatitis B yang toksik bagi E. coli. Maeng et al. (2001) melakukan
percobaan ekspresi gen virus Hepatitis B secara parsial yang diikuti dengan
gluthathion-S-23
transferase (GST), dengan demikian ekspresi gen dan kelarutan antigen
permukaan Hepatitis B pada E. coli tersebut bisa ditingkatkan. Ekspresi gen
tersebut dilakukan dibawah kontrol promoter tac. Hasil penelitian tersebut
menunjukkan bahwa terjadi peningkatan tingkat ekspresi antigen pre-S1 yang
digabung dengan GST. Koschorreck et al. (2005) juga melaporkan terjadi
peningkatan solubilitas protein rekombinan yang digabung dengan GST.
Terjadinya peningkatan kelarutan ini sangat penting untuk proses pemurnian
antigen rekombinan dengan menggunakan kolum yang telah tersedia secara
komersial.
Aplikasi Rekayasa Genetik di Bidang Peternakan
Menurut Winarno dan Agustinah (2007), WHO telah meramalkan bahwa
populasi dunia akan berlipat dua pada tahun 2020 sehingga jumlahnya akan lebih
dari 10 milyar. Oleh karena itu, produksi pangan juga harus ditingkatkan.
Kendalanya adalah jumlah sisa lahan dunia yang belum termanfaatkan saat ini
sangat kecil dan terbatas. Teknologi rekayasa genetika atau GMO akan
memainkan peranan yang sangat penting dalam mengatasi kendala tersebut.
Teknologi rekayasa genetika dapat menjadi strategi yang sangat bagus untuk
meningkatkan produksi pangan. Wiryosuhanto dan Sudradjat (1992) menjelaskan,
bahwa teknologi rekayasa genetika merupakan alat yang dapat membantu
pengusaha industri peternakan untuk meningkatkan status kesehatan ternaknya,
meningkatkan feed convertion, dan memperpendek waktu pemeliharaan sampai
mencapai berat badan yang sesuai dengan permintaan pasar. Teknologi rekayasa
genetik memberi harapan untuk diaplikasikan di dalam pembangunan peternakan
di masa datang, yaitu bioteknologi molekuler yang berkaitan dengan keberhasilan
teknologi kloning, ternak transgenik, dan vaksin. Winarno (2004) menyatakan,
bahwa teknologi rekayasa genetika juga mampu membantu masalah yang
dihadapi oleh industri susu dalam mengatasi masalah turunnya suplai renin.
Renin, yang sekarang lebih dikenal dengan nama chymosin, adalah enzim
proteolitik yang digunakan dalam pembuatan keju oleh industri susu. Secara
konvensional, renin diproduksi dari abomasum (lambung ke-4) anak sapi.
lain dengan memproduksi renin mikroba yang diekstraksi dari fungi, namun sifat
biokimianya tidak sama dengan renin dari abomasum anak sapi. Hal ini bisa
diatasi dengan cara mentransfer gen dari anak sapi yang mengkode pembentukan
renin ke dalam bakteri, selanjutnya bakteri tersebut digunakan sebagai mesin
pembentuk renin yang sifat biokimianya sama dengan renin dari abomasum anak
KLONING GEN
SR100
DALAM RANGKA PRODUKSI
PROTEIN REKOMBINAN SEBAGAI MODEL
IMUNOGEN UNTUK MENGHASILKAN
ANTIBODI
Slamet Riyadi1, Rarah R.A. Maheswari2, Mirnawati Sudarwanto3, Fransiska R. Zakaria4, Muhamad Ali5
1
Mahasiswa Program Studi Ilmu Ternak, Sekolah Pascasarjana IPB, 2Fakultas Peternakan IPB,
3
Fakultas Kedokteran Hewan IPB, 4Fakultas Teknologi Pertanian IPB,
5
Ketua Lab. Mikrobiologi & Bioteknologi Fak. Peternakan UNRAM
@
E-mail: [email protected], phone: 085217886729
Abstrak
Sejak satu dekade yang lalu, muncul paradigma baru dalam teknik pembuatan vaksin. Penggunaan mikroorganisme virulen yang dilemahkan ataupun yang dibunuh telah diganti dengan penggunaan vaksin sub unit yang lebih efektif dengan teknologi DNA rekombinan. Melalui penggunaan teknologi tersebut, gen tertentu dari mikroorganisme virulen dapat dikloning, diekspresi dan dievaluasi penggunaannya sebagai vaksin. Pada penelitian ini, telah dikloning bagian gen penyandi protein hidrofilik dari protein S (aa 100-164) dari antigen permukaan virus hepatitis B untuk digunakan sebagai penghasil kandidat vaksin rekombinan. Gen tersebut kemudian diligasi dengan vektor pGEX-4T-2 dan disekuensing. Pensejajaran hasil sequensing tersebut dengan sekuen asli virus hepatitis B menunjukkan kesamaan. Hasil utama dari penelitian ini adalah klon pembawa gen penyandi protein S yang selanjutnya dapat digunakan untuk menghasilkan kandidat vaksin rekombinan hepatitis B.
Kata kunci:DNA rekombinan, vaksin, pGEX-4T-2, Hepatitis B.
Abstract
Since one decade ago, a new paradigm of vaccine design is emerging. Instead of attenuated virulent microorganisms or killed virulent microorganisms, effective subunit vaccines were developed using recombinant DNA technology. By using the technology, selected genes of the virulent microorganisms can be cloned, expressed, and evaluated as vaccine components. In this research, hydrophilic domain of S protein (aa 100-164)-encoding gene of hepatitis B surface antigen was cloned for vaccine candidate production. The gene was ligated with pGEX-4T-2 vector and sequenced. Sequences alignment of the amplified fragment with genome of hepatitis B virus indicated that the sequences were identical. A major result achieved from this research was clones carrying S antigens-encoding gene that could be used further for production of recombinant hepatitis B vaccine candidates.
Pendahuluan
Teknologi kloning merupakan terobosan baru di bidang rekayasa genetika.
Menurut Winarno dan Agustinah (2007), kloning adalah pengembangbiakan suatu
mahluk hidup yang persis sama dengan induknya tanpa melalui pembuahan,
seperti stek pada tanaman, tetapi kloning melalui rekayasa genetika jauh lebih
rumit. Muladno (2002) menjelaskan, bahwa pada prinsipnya kloning DNA adalah
proses penggandaan jumlah DNA rekombinan melalui proses perkembangbiakan
sel bakteri (biasanya E. coli). Proses penggandaan tersebut dilakukan dengan
memasukkan DNA rekombinan ke dalam E.coli, diikuti dengan inkubasi sel E.coli
pada suhu optimal sehingga sel berkembangbiak secara eksponensial. Selanjutnya
dijelaskan pula, bahwa menggandakan jumlah molekul DNA tidak hanya dapat
dilakukan dengan memanfaatkan mekanisme kehidupan mikroorganisma, tetapi
dapat juga dilakukan melalui teknik PCR (Polymerase Chain Reaction).
Perkembangan teknologi molekuler (seperti kloning) yang sangat pesat telah
membuka era baru dalam menghasilkan berbagai jenis vaksin maupun obat yang
dibutuhkan oleh hewan, ternak maupun manusia. Penggunaan teknologi tersebut
telah memudahkan dihasilkannya berbagai sub unit vaksin yang jauh lebih efektif
jika dibandingkan dengan vaksin yang dihasilkan dengan teknologi konvensional
menggunakan mikroorganisme virulen yang dilemahkan ataupun telah dibunuh.
Indonesia merupakan daerah endemis sedang sampai tinggi untuk penyakit
hepatitis B, oleh karena itu pada tahun 1987, WHO menetapkan Pulau Lombok
sebagai model imunisasi masal hepatitis B pertama di dunia. Hasil proyek tersebut
menunjukkan penggunaan vaksin konvensional mampu menurunkan prevalensi
hepatitis B hanya sampai 70% (Mulyanto et al. 2002). Hasil imunisasi tersebut
dinyatakan belum optimal, hal ini antara lain disebabkan vaksin konvensional
yang digunakan (Korean Green Cross) berasal dari plasma darah orang asing
sehingga tidak mampu menstimulasi munculnya tanggap kebal (antibodi) spesifik
yang mampu melawan virus hepatitis B yang terdapat di Indonesia (Joung et al.,
2004).
Berdasarkan permasalahan di atas, pada penelitian ini dilakukan rekayasa
terhadap gen penyandi antigen permukaan hepatitis B untuk menghasilkan antigen
27
penyandi antigen permukaan hepatitis B di atas akan digabung (fusi) dengan gen
penyandi enzim gluthation-S-transferase (GST) untuk meningkatkan ekspresi
maupun solubilitas antigen yang sangat penting untuk aktivitas maupun proses
purifikasi (Sheu dan Lo 1995; Vikis dan Guan 2000; Koschoreck et al. 2005). Gen
penyandi antigen permukaan hepatitis B yang digunakan pada penelitian ini
adalah gen yang diisolasi dari virus hepatitis B sub tipe adw sebagai sub tipe
utama di Indonesia. Hal ini dimaksudkan untuk membuat kandidat vaksin galur
lokal yang mampu memberikan respon antibodi yang spesifik sesuai dengan
genetik virus hepatitis B yang terdapat di Indonesia.
Dihasilkannya kandidat vaksin hepatitis B rekombinan dengan teknologi
rekayasa DNA menggunakan bakteri diharapkan dapat menggantikan metode
produksi vaksin konvensional dari plasma yang memiliki kelemahan seperti
rendahnya imunogenisitas, sumber plasma yang terus berkurang (karena jumlah
penderita penyakit hepatitis B menurun sejalan dengan keberhasilan program
vaksinasi), serta kekhawatiran adanya kontaminasi penyakit lain pada serum
donor. Gen penyandi antigen tersebut diisolasi dari virus hepatitis B yang terdapat
di Indonesia, sehingga antigen ini diharapkan dapat menghasilkan kandidat vaksin
rekombinan hepatitis B yang sesuai dengan genetik virus tersebut di Indonesia.
Bahan dan Metode
Bahan Penelitian
Amplifikasi fragmen S (asam amino nomor 100-164) dari gen penyandi
antigen permukaan virus hepatitis B, digunakan plasmid pGET-HB (disediakan
oleh Prof.Mulyanto, Laboratorum Hepatitis Mataram) yang membawa gen-gen
permukaan virus hepatitis B sebagai cetakan. Amplifikasi tersebut menggunakan