KULTUR EMBRIO JERUK KEPROK (Citrus nobilis Lour.)
PADA MEDIA MS DENGAN PENAMBAHAN KINETIN
SKRIPSI
UTARI EKA WINDANI
060805024
DEPARTEMEN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
UNIVERSITAS SUMATERA UTARA
KULTUR EMBRIO JERUK KEPROK (Citrus nobilis Lour.) PADA MEDIA MS DENGAN PENAMBAHAN KINETIN
SKRIPSI
Diajukan untuk melengkapi dan memenuhi syarat mencapai gelar Sarjana Sains
UTARI EKA WINDANI 060805024
DEPARTEMEN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM UNIVERSITAS SUMATERA UTARA
PERSETUJUAN
Judul : KULTUR EMBRIO JERUK KEPROK (Citrus nobilis Lour.) PADA MEDIA MS DENGAN PENAMBAHAN KINETIN
Kategori : SKRIPSI
Nama : UTARI EKA WINDANI
Nomor Induk Mahasiswa : 060805024
Program Studi : SARJANA (S1) BIOLOGI
Departemen : BIOLOGI
Fakultas : MATEMATIKA DAN ILMU PENGETAHUAN ALAM
Universitas : SUMATERA UTARA
Diluluskan di Medan, Maret 2011 Komisi Pembimbing :
Pembimbing II Pembimbing I
(Dra.Elimasni, M.Si) (Dra.Isnaini Nurwahyuni, M.Sc) NIP. 1965024 199103 2001 NIP. 19600523 198502 2001
Diketahui/Disetujui oleh
Departemen Biologi FMIPA USU Ketua,
PERNYATAAN
KULTUR EMBRIO JERUK KEPROK (Citrus nobilis Lour.) PADA MEDIA MS DENGAN PENAMBAHAN KINETIN
SKRIPSI
Saya mengakui bahwa skripsi ini adalah hasil kerja saya sendiri, kecuali beberapa kutipan dan ringkasan yang masing-masing disebutkan sumbernya.
Medan, Maret 2011
ABSTRAK
Penelitian tentang “Kultur Embrio Jeruk Keprok (Citrus nobilis Lour.) pada Media MS dengan Penambahan Kinetin” dilakuka n di Laboratorium Kultur Jaringan Tumbuhan Departemen Biologi Fakultas Matematika dan Ilmu Pengetahuan Alam Universitas Sumatera Utara. Penelitian bertujuan mengetahui konsentrasi kinetin terbaik untuk pertumbuhan planlet embrio jeruk keprok (Citrus nobilis Lour.) pada media MS. Penelitian menggunakan Rancangan Acak Lengkap (RAL) non faktorial dengan perlakuan kinetin terdiri dari 5 taraf. Secara umum kinetin berpengaruh terhadap semua variabel pengamatan. Penambahan kinetin berpengaruh nyata terhadap jumlah tunas, berat planlet dan panjang akar. Konsentrasi 1 ppm (K1)
menghasilkan jumlah akar. Konsentrasi 2 ppm (K2) menghasilkan jumlah tunas, berat
planlet, panjang tunas, dan panjang akar. Konsentrasi 3 ppm (K3) menghasilkan
jumlah daun. Pada pengamatan mikroskopik akar menunjukkan bahwa minggu pertama jaringan penyusun akar sudah terbentuk dan tidak ada perbedaan antara semua perlakuan kinetin.
EMBRYO CULTURE OF KEPROK ORANGE (Citrus nobilis Lour.) on MS MEDIA WITH THE ADDITION OF KINETIN
ABSTRACT
Research on "Embryo Culture Keprok Orange (Citrus nobilis Lour.) on MS media with the addition of Kinetin" performed at the Laboratory of Plant Tissue Culture Department of Biology, Faculty of Mathematics and Natural Sciences University of Sumatra Utara. The research aims to identify the best concentration of kinetin to the growth of the embryo plantlets of keprok orange (Citrus nobilis Lour.) on MS medium. Research using Completely Randomized Design (CRD), non-factorial with kinetin treatment consists of 5 levels. In general, kinetin effect on all observed variables. Addition of kinetin significantly affect the number of shoots, weight of plantlets and root length. Concentration of 1 ppm (K1) produces the number of roots.
The concentration of 2 ppm (K2) produces the number of shoots, weight of plantlet,
shoot length and root length. Concentration of 3 ppm (K3) produces the number of
leaves. At the root of microscopic observation showed that the first week of the root constituent tissue has been formed and there was no difference between all treatments kinetin.
PENGHARGAAN
Bismillahirahmannirrahim. Puji dan syukur penulis panjatkan kehadirat Allah SWT.
atas segala nikmat Iman, Islam dan Ikhsan serta karunian-Nya sehingga penulis dapat menyelesaikan skripsi dengan judul ”Kultur Embrio Jeruk Keprok (Citrus nobilis Lour.) Pada Media MS Dengan Penambahan Kinetin”.
Penulis mengucapkan terima kasih yang sebesar-besarnya kepada Ibu Dra. Isnaini Nurwahyuni, M.Sc dan Ibu Dra. Elimasni, M.Si. sebagai dosen pembimbing yang banyak memberi saran, motivasi, waktu dan perhatiannya yang besar dari awal penelitian sampai selesai, hingga penyusunan skripsi ini. Terima kasih juga penulis ucapkan kepada Bapak Riyanto Sinaga, S.Si., M.Si dan Ibu Dra. Emita Sabri, M.Si selaku dosen penguji yang selalu memberikan kritik, saran dan nasihat baik dalam penyempurnaan skripsi ini maupun dalam kehidupan sehari-hari selama di kampus. Terima kasih atas ilmu yang kalian berikan.
Terima kasih untuk Penasehat Akademik penulis Ibu Mayang Sari Yeanny S.Si., M.Si., yang bersedia memberi saran untuk memajukan akademis penulis. Kepada Ibu Dr. Nursahara Pasaribu, M.Sc dan Bapak Kiki Nurtjahja, M.Sc., selaku Ketua dan Sekretaris Departemen Biologi FMIPA USU. Seluruh dosen di Biologi FMIPA USU, khususnya Ibu Dr. Suci Rahayu, M.Sc., dan Ibu Kaniwa Berliani, S.Si., M.Si., yang selalu memberi nasihat dan perhatian kepada penulis. Kak Roslina Ginting dan Bang Enda Raswin selaku staf pegawai Departemen Biologi FMIPA USU.
Ucapan terima kasih yang teramat dalam serta hormat penulis kepada kedua orang tua, Ayahanda tersayang Ahmad Jamaluddin dan Ibunda tercinta Tri Rospitasari, atas kasih sayang, dukungan, nasihat, doa yang tulus dan ikhlas serta perjuangan untuk memberikan semangat pantang menyerah kepada buah hatinya. Adik-adik terkasih, Tia Junna Sonia, Dinda Aulia dan Muhammad Ridho Febrian, atas kasih sayang dan keceriaan kalian yang selalu memberi semangat baru bagi penulis. Nenek tersayang Mariani RS. S.Pd.I dan Kakek K.H. Husein Kasim, yang selalu mendoakan setiap langkah penulis. Terima kasih untuk Ahmad Habibi Siregar., S.Ked., yang selalu memberi inspirasi dan semangat tiada henti, semoga kita dapat meraih cita-cita mulia kita. Seluruh keluarga terdekat penulis, Siswanto Afif, Anie, Siswanti Afif, Sulaiman, Hidayat Afif, Leliana Asli Kesuma, Mulyo Prihatin, Rita Rahmawati, Siti Rohani, Siti Rohima dan Ahmad Hasanuddin yang selalu memberi perhatian yang besar kepada penulis.
merasa lemah dan letih menghadapi dunia kampus dan semoga kita menjadi insan yang sukses dunia akhirat.
Teman-teman seangkatan 2006, Tety, Santi, Rama, Diah, Kasbi, Nikmah, Amie, Yanti, Arico, Leni, Eva, Widya, Jane, Siti, Grisa, Zulfa, Dian, Lena, Afrida, Sulis, Nana, Indah, Zukie, Umri, Sary, Rivo, Hariadi, Rudi, Andri, Helen, Dwi, Christine, Desmina, Hilda, Farida, Dola, Septi, Sutrisno dan Deni. Semoga kekompakan dan perjuangan kita sejak duduk di bangku kuliah hingga memperoleh gelar sarjana membawa kita berada pada tingkat kesuksesan. Untuk Junior 2007, khususnya Misel, Asril, Resti, Dwi, Anggun dan Natalia. Junior 2008, khususnya Intan, Rini, Opy, Pinta, Sirma dan Juju. Junior 2009, khususnya Siska, Adrian, dan Bobby, yang telah mewarnai hari-hari penulis selama di kampus. Untuk senior di Laboratorium Kultur Jaringan, Susilawati, Rico Pranata Tarigan, S.Si., Suria Wulandari Purnama, S.Si., Nikmatul Khoiriah, S.Si. dan Widya Sari Akriyani, S.Si., terima kasih atas ilmu yang kalian berikan. Dan untuk para peneliti di seluruh dunia, semoga terus menciptakan ide dan eksperimen baru.
Penulis mengharapkan skripsi ini berguna bagi peneliti selanjutnya dan dapat digunakan untuk penelitian yang berhubungan dengan jeruk keprok. Sebelum dan sesudahnya penulis mohon maaf atas ketidaksempurnaan skripsi ini dan penulis mengucapkan terima kasih.
Medan, Maret 2011
DAFTAR ISI
1.3 Tujuan Penelitian 4
1.4 Hipotesis 4
1.5 Manfaat Penelitian 4
Bab 2 Tinjauan Pustaka
2.1 Botani Tanaman Jeruk (Citrus nobilis Lour.) 5
2.2 Kultur Jaringan Jeruk 6
2.2.1 Eksplan 7
3.1 Waktu dan Tempat Penelitian 13
3.2 Alat dan Bahan Penelitian 13
3.3 Metoda penelitian 14
3.4 Prosedur Kerja 14
3.5 Variabel Pengamatan 17
Bab 4 Hasil dan Pembahasan
4.1 Tipe Pertumbuhan Kultur 18
4.2 Jumlah Tunas 19
4.3 Berat Planlet (g) 21
4.4 Jumlah Daun (helai) 23
4.5 Panjang Tunas (cm) 24
4.6 Panjang Akar (cm) 26
4.7 Jumlah Akar 29
4.8 Pengamatan Mikroskopik Anatomi Akar 31
4.9 Persentase Kultur yang Hidup (%) 32
4.10 Persentase Kultur Terkontaminasi (%) 32
Bab 5 Kesimpulan dan Saran 5.1 Kesimpulan 34
5.2 Saran 34
DAFTAR TABEL
Halaman
Tabel 4.2 Rata-Rata Jumlah Tunas 19
Tabel 4.3 Rata-Rata Berat Planlet (g) 21
Tabel 4.4 Rata-Rata Jumlah Daun (helai) 23
Tabel 4.5 Rata-Rata Panjang Tunas (cm) 25
Tabel 4.6 Rata-Rata Panjang Akar (cm) 26
DAFTAR LAMPIRAN
Halaman Lampiran A. Komposisi Media MS (Murashige & Skoog) 1962 39
Lampiran B. Layout Penelitian 40
Lampiran C. Data Pengamatan Dan Sidik Ragam Jumlah Tunas 41 Lampiran D. Data Pengamatan Dan Sidik Ragam Berat Planlet (g) 43 Lampiran E. Data Pengamatan Dan Sidik Ragam Jumlah Daun 45 Lampiran F. Data Pengamatan Dan Sidik Ragam Panjang Tunas (cm) 47 Lampiran G. Data Pengamatan Dan Sidik Ragam Panjang Akar (cm) 49 Lampiran H. Data Pengamatan Dan Sidik Ragam Jumlah Akar 51 Lampiran I. Alur Kerja Kultur Embrio Jeruk Keprok (Citrus nobilis Lour.)
Pada Media MSDengan Penambahan Kinetin 53 Lampiran J. Alur Kerja Pembuatan Preparat Anatomi Akar 54 Lampiran K. Data Persentase Kultur Terkontaminasi 55 Lampiran L. Gambar Planlet Pada Tiap Perlakuan Selama 4 Minggu 56 Lampiran M. Gambar Anatomi Mikroskopik Akar Pada Setiap Perlakuan
DAFTAR GAMBAR
Halaman Gambar 3.4.3 Embrio Dari Biji Jeruk Keprok (a) Kulit Biji (b) Biji Utuh
(c) Kotiledon (d) Embrio 15 Gambar 3.4.4 Penanaman Embrio Pada Media MS (a) Media MS Sebelum
Ditanam Embrio (b) Proses Penanaman Embrio (c) Media
Berisi Embrio 16 Gambar 4.1 Planlet Dalam Media MS + Kinetin (a) Daun (b) Batang
(c) Media (d) Akar 18 Gambar 4.2.1 Grafik Kuadratik Jumlah Tunas Dengan Perlakuan Kinetin
Pada Minggu III 19 Gambar 4.2.2 Eksplan Membentuk Tunas Pada Perlakuan K2.3.2 20
Gambar 4.3.1 Respon Kinetin Terhadap Berat Planlet 22 Gambar 4.4.1 Respon Kinetin Terhadap Jumlah Daun 23 Gambar 4.4.2 Eksplan Membentuk Daun Pada Perlakuan K3.4.1 24
Gambar 4.5.1 Respon Kinetin Terhadap Panjang Tunas 25 Gambar 4.7.1 Respon Kinetin Terhadap Jumlah Akar 29 Gambar 4.7.2 Perbandingan Jumlah Akar Pada Perlakuan K0, K1, K2, K3, K4.
(a) (b) (c) (d) (e) Pembentukan Akar 30 Gambar 4.8 Anatomi Akar Pada Kontrol dan Perlakuan K4 Pada Umur
Planlet 4 Minggu (a) Epidermis (b) Hipodermis (c) Korteks
EMBRYO CULTURE OF KEPROK ORANGE (Citrus nobilis Lour.) on MS MEDIA WITH THE ADDITION OF KINETIN
ABSTRACT
Research on "Embryo Culture Keprok Orange (Citrus nobilis Lour.) on MS media with the addition of Kinetin" performed at the Laboratory of Plant Tissue Culture Department of Biology, Faculty of Mathematics and Natural Sciences University of Sumatra Utara. The research aims to identify the best concentration of kinetin to the growth of the embryo plantlets of keprok orange (Citrus nobilis Lour.) on MS medium. Research using Completely Randomized Design (CRD), non-factorial with kinetin treatment consists of 5 levels. In general, kinetin effect on all observed variables. Addition of kinetin significantly affect the number of shoots, weight of plantlets and root length. Concentration of 1 ppm (K1) produces the number of roots.
The concentration of 2 ppm (K2) produces the number of shoots, weight of plantlet,
shoot length and root length. Concentration of 3 ppm (K3) produces the number of
leaves. At the root of microscopic observation showed that the first week of the root constituent tissue has been formed and there was no difference between all treatments kinetin.
BAB 1
PENDAHULUAN
1.1Latar Belakang
Jeruk (Citrus sp.) adalah tanaman tahunan yang berasal dari Asia Tenggara. Sejak ratusan tahun lalu, tanaman ini sudah terdapat di Indonesia, baik sebagai tanaman liar maupun sebagai tanaman pekarangan (Soelarso, 1996). Buah jeruk merupakan salah satu jenis buah yang paling banyak digemari oleh masyarakat. Oleh karena itu tidak mengherankan jika perkembangan tanaman jeruk mengalami perubahan populasi yang cukup tajam. Pada saat ini sebagian petani buah menyadari bahwa komoditas buah jeruk memang dapat meningkatkan taraf hidup masyarakat, terutama jenis komoditas jeruk keprok (Kanisius, 1994). Jeruk keprok (Citrus nobilis Lour.) merupakan salah satu spesies dari sekian banyak spesies jeruk yang sudah dikenal dan dibudidayakan di Indonesia. Mutu dan penampilannya sangat mempengaruhi dan memegang peranan penting dalam perdagangan (Zahara, 2002).
serangan penyakit akar dan batang serta akhir-akhir ini disebabkan oleh suatu penyakit yang dinamakan CVPD (Citrus Vein Phloem Degeneration) (Joesoef, 1993).
Dalam usaha memenuhi kebutuhan jeruk bermutu dan bebas penyakit, teknik perbanyakan tanaman secara in vitro digunakan untuk memperoleh tanaman yang berkualitas, homogen, cepat dan dalam jumlah yang banyak. Kultur jaringan dapat menghasilkan bibit dalam jumlah yang tidak terbatas, dan mewarisi sifat induk (Wiryanta & Raharja, 2003). Sesuai dengan pendapat Suryowinoto (1996), bahwa memperbanyak jeruk keprok dapat dilakukan secara klonal melalui teknik kultur jaringan atau teknik in vitro. Dalam budidaya tanaman dengan menggunakan teknik kultur jaringan, pemberian zat pengatur tumbuh dalam media tanam dan pemilihan eksplan sebagai bahan inokulum awal yang ditanam dalam media perlu diperhatikan karena mempengaruhi pertumbuhan dan perkembangan eksplan tersebut menjadi bibit yang baru.
Menurut Heinz & Mee (1969) dalam (Reinert & Bajaj, 1989), media yang paling baik untuk diferensiasi kalus dan perkembangan planlet adalah media Murashige & Skoog (1962) atau modifikasinya. Media ini kaya akan makroelemen, nitrogen, sukrosa dan vitamin tertentu (Hartman & Kester, 1983). Sementara zat tambahan yang biasa digunakan adalah zat pengatur tumbuh. Zat pengatur tumbuh auksin dan sitokinin dapat diberikan bersama-sama atau auksin saja ataupun sitokinin saja, tergantung dari tujuan (Hendaryono & Wijayani, 1994). Penambahan berbagai macam ekstrak organik pada media kultur sering memberikan respon pertumbuhan yang diinginkan. Bahan organik kompleks tersebut antara lain protein hidrolisa, air kelapa, ekstrak ragi, ekstrak malt, pisang, jus jeruk, jus tomat, dan sebagainya.Tunas pucuk dan satu ruas jeruk manis telah ditanam pada media dasar Murashige & Skoog yang diperkaya dengan 500 mg ekstrak malt (Agisimanto et al., 2006).
akar. Butar-butar (2006) juga melakukan penelitian terhadap kultur biji jeruk keprok (Citrus nobilis Lour. Var. Brastepu) pada media MS diperkaya kinetin (0,0; 0,1; 1,0; 10 mg/l) dan atonik (0,0; 0,5; 1,0; 1,5 mg/l) dari penelitian tersebut diperoleh kombinasi atonik (0,5 ml/l) dan kinetin (1,0 mg/l) pada kultur biji jeruk keprok memberikan pengaruh yang nyata terhadap jumlah tunas. Perlakuan terbaik pada 0,5 ml/l atonik + 1,0 mg/l kinetin yang berpengaruh terhadap berat basah kultur dan jumlah tunas. Selanjutnya Silalahi (2006) juga melakukan penelitian pengaruh konsentrasi media MS dengan kombinasi 2,4-D (1 mg/l) dan BAP (1 mg/l) pada kultur biji jeruk Brastepu (Citrus nobilis Lour. Var. Brastepu), dari penelitian diperoleh pengaruh nyata terhadap tinggi tanaman.
Penelitian lainnya tentang jeruk ini juga pernah dilakukan oleh Devi & Hardiyanto (2007) terhadap kultur nuselus “Japanes Citroen”. Dalam penelitian tersebut digunakan kombinasi antara sitokinin dan ME (Ekstrak Malt). Dari hasil penelitian tersebut diperoleh bahwa kalus akan berkembang baik pada media MS + 3 ppm BA + 500 mg/l ME + 5% sukrosa.
Berdasarkan beberapa penelitian tersebut, maka dalam penelitian ini embrio jeruk keprok dikulturkan dalam media MS dengan penambahan ME dan perlakuan zat pengatur tumbuh kinetin dengan 5 konsentrasi yaitu 0, 1, 2, 3, 4 ppm.
1.2Permasalahan
Beberapa penelitian telah dilakukan untuk kultur jeruk, namun belum menunjukkan hasil yang baik, sehingga masih perlu dilakukan penelitian-penelitian dengan metode yang lebih bervariasi lagi untuk mendapatkan hasil yang lebih maksimal sesuai dengan yang diharapkan. Dalam penelitian ini dilakukan pengkulturan terhadap embrio jeruk dengan konsentrasi zat pengatur tumbuh yang berbeda dalam media MS dengan penambahan kinetin, yang diharapkan dapat menghasilkan kultur yang baik.
1.3Tujuan
Penelitian ini bertujuan untuk mengetahui konsentrasi kinetin terbaik untuk pertumbuhan planlet dari embrio jeruk keprok (Citrus nobilis Lour.) pada media MS (Murashige & Skoog).
1.4Hipotesis
Penambahan kinetin pada berbagai konsentrasi dapat meningkatkan pertumbuhan planlet pada kultur embrio jeruk keprok (Citrus nobilis Lour.).
1.5Manfaat
BAB 2
TINJAUAN PUSTAKA
2.1 Botani Jeruk Keprok (Citrus nobilis Lour.)
Menurut Steenis (2003), tanaman jeruk keprok (Citrus nobilis Lour.) mempunyai sistematika sebagai berikut:
Kingdom : Plantae
Division : Spermatophyta Klass : Angiospermae Subklass : Dicotyledoneae Ordo : Rutales
Family : Rutaceae Genus : Citrus
Spesies : Citrus nobilis Lour.
Tanaman jeruk keprok (Citrus nobilis Lour.) diduga berasal dari Asia Tenggara (Purseglove, 1987), kemudian menyebar ke seluruh dunia terutama di daerah subtropik. Pohon jeruk mencapai ketinggian 6-10 m, mempunyai duri yang kuat, ranting yang muda bersudut, biasanya berduri dan bercabang rendah. Jeruk keprok memiliki daun berwarna hijau tua mengkilat pada permukaan atas dan hijau muda pada permukaan bawah tangkai. Buah jeruk tergolong buah sejati, tunggal, dan berdaging (Soelarso, 1996).
Menurut Ball (1997), kulit jeruk ini berdaging tebal dengan permukaan yang kasar dan tidak rata. Biji bersifat poliembrionik dan berwarna sedikit kekuningan sementara embrio berwarna putih. Jeruk keprok ini mengandung sejumlah nutrisi, diantaranya vitamin B1 dan vitamin C. Selain itu jeruk ini juga mengandung glukosa,
pereda berbagai penyakit, misalnya sebagai obat batuk, mengurangi nafsu makan dan menghilangkan rasa mual. Kulit dan biji jeruk mengandung minyak yang dapat digunakan sebagai pengharum rambut, campuran minuman, dan bahan wangi-wangian.
2.2 Kultur Jaringan Jeruk
Penggunaan metode in vitro untuk tanaman jeruk telah dimulai oleh Bove & Morel (1957) dalam Nurwahyuni (2001) dan sejak itu kultur jaringan tanaman jeruk banyak mendapat perhatian. Regenerasi tanaman jeruk secara kultur jaringan telah dilakukan diantaranya dari bagian tunas yang menghasilkan planlet (Bahrany, 2002), bagian kotiledon (Begum et al., 2003) yang menghasilkan kalus.
Teknik kultur jaringan memiliki keuntungan karena dapat menghasilkan bibit klonal yang banyak dalam waktu yang singkat, dapat meningkatkan kualitas tanaman karena mengasilkan tanaman jeruk seragam dan tingkat kesehatan yang lebih baik (Ghorbel et al., 1998, dalam Nurwahyuni, 2003).
2.2.1 Eksplan
Eksplan ini biasanya berupa bagian yang paling kecil dari jaringan atau organ yang diambil dari tanaman yang kemudian akan dikulturkan. Menurut Nusmawarhaeni et
al., (2001), eksplan tanaman yang masih muda menghasilkan tunas maupun akar
adventif lebih cepat bila dibandingkan dengan bagian yang tua. Hal itu karena bagian tersebut memiliki sifat pertumbuhan yang agresif. Jaringan tanaman yang masih muda mempunyai daya regenerasi yang lebih tinggi, sel-selnya masih aktif membelah diri dan relatif mengandung lebih sedikit kontaminan. Sedangkan jaringan tanaman yang sudah tua lebih sedikit beregenerasi dan biasanya mengandung lebih banyak kontaminan (Yusnita, 2003).
Penggunaan eksplan dari jaringan muda lebih sering berhasil karena sel-selnya aktif membelah, dinding sel tipis karena belum terjadi penebalan lignin dan selulosa yang menyebabkan kekakuan pada sel. Gunawan (1995) menyatakan bagian tanaman yang dapat digunakan sebagai eksplan adalah pucuk muda, batang muda, daun muda, kotiledon, hipokotil. Menurut Wattimena (1992) perbedaan dari bagian tanaman yang digunakan akan menghasilkan pola pertumbuhan yang berbeda. Eksplan tanaman yang masih muda menghasilkan tunas maupun akar adventif lebih cepat bila dibandingkan dengan bagian yang tua. Perbedaan dari bagian tanaman yang digunakan akan menghasilkan pola pertumbuhan yang berbeda.
Santoso & Nursandi (2004) menambahkan bahwa langkah pertama untuk menentukan bagian mana dari tanaman yang akan digunakan sebagai eksplan adalah melihat potensi genetik yang ada pada tanaman di lapangan. Untuk itu perlu dilakukan analisis jaringan secara in vivo untuk mengetahui bagian tanaman yang mempunyai kandungan tertinggi senyawa yang diinginkan. Tanaman yang mempunyai kandungan senyawa tertentu dalam jumlah besar akan mampu menghasilkan senyawa yang sama dalam jumlah besar pula apabila tanaman tersebut dikulturkan secara in vitro. Hal ini sesuai dengan teori totipotensi yang dimiliki suatu sel.
Jaringan pada embrio zigotik muda secara alami merupakan jaringan yang embriogenik. Ekplan diberi perlakuan kinetin 1,0 mg/l + sukrosa 10 g/l, kinetin 2,0 mg/l + sukrosa 30 g/l, kinetin 3,0 mg/l + sukrosa 10 g/l menghasilkan kultur dengan persentase hidup yang tinggi yaitu 75%. Konsentrasi kinetin 1,0 mg/l mampu mempercepat saat inisiasi tunas dalam waktu 16,02 (HSI). Konsentrasi sukrosa 30 g/l mampu mempercepat saat inisiasi tunas dalam waktu 16,08 (HSI) (Khurniawati, 2007).
2.2.2 Media Kultur Jaringan
Media tanam dalam kultur jaringan adalah tempat untuk tumbuh eksplan. Media tanam tersebut dapat berupa larutan (cair) atau padat. Media tanam harus berisi semua zat yang diperlukan untuk menjamin pertumbuhan eksplan. Bahan-bahan yang diramu berisi campuran garam mineral sumber unsur makro dan unsur mikro, gula, protein, vitamin, dan hormon tumbuh. Selain itu diperlukan pula bahan tambahan seperti agar, gula dan lain-lain. Dengan demikian keberhasilan kultur jaringan jelas ditentukan oleh media tanam dan macam tanaman. Campuran media yang satu mungkin cocok untuk jenis-jenis tanaman tertentu, tetapi tidak cocok untuk jenis-jenis tanaman yang lainnya (Hendaryono & Wijayani, 1994).
Media MS merupakan media yang dibuat oleh Murashige & Skoog pada tahun 1962, dan sesuai untuk berbagai eksplan spesies tanaman. Dari berbagai komposisi dasar ini kadang-kadang dibuat modifikasi, misalnya hanya menggunakan setengah dari konsentrasi garam-garam makro yang digunakan atau menggunakan komponen garam-garam makro berdasarkan MS yang disesuaikan (Gunawan, 1994). Media yang dikembangkan oleh Murashige & Skoog (MS) untuk kultur jaringan tanaman digunakan secara luas untuk kultivasi kalus pada agar demikian juga kultur suspensi sel dalam media cair. Keistimewaan media ini yaitu kandungan nitrat, kalium dan amoniumnya yang tinggi (Wetter & Constabel, 1991).
merupakan media yang cocok untuk merangsang perkembangan kultur dengan cepat serta mempengaruhi organogenesis dan embriosomatik (Reinert & Bajaj, 1989).
2.2.3 Zat Pengatur Tumbuh
Pada tumbuhan, hormon tumbuh sangat berperan dalam pertumbuhan dan perkembangan tumbuhan (Abidin, 1982). Selain itu zat pengatur tumbuh juga berperan dalam mempercepat terbentuknya kalus serta proses diferensiasi semua fase pertumbuhan dan perkembangan tanaman (Heddy, 1986), juga dapat memberikan arah bagi perkembangan sel tanaman (Pierik, 1987). Faktor yang perlu mendapatkan perhatian dalam penggunaan zat pengatur tumbuh antara lain jenis dan konsentrasi zat pengatur tumbuh dan urutan penggunaannya serta periode masa induksi yang dilakukan dalam teknik kultur jaringan tertentu (Gunawan, 1995). Zat pengatur tumbuh yang biasa digunakan adalah sitokinin dan auksin. Sitokinin yang biasa digunakan 6-Benzil Amino Purin (BAP) dan kinetin, sedangkan auksin yang biasa digunakan adalah IAA, 2,4 D, NAA dan IBA (Maryani & Zamroni, 2005).
Dalam kultur jaringan zat pengatur auksin dan sitokinin sangat berpengaruh (Gunawan, 1995). Auksin adalah zat pengatur tumbuh yang mempengaruhi pertumbuhan kalus. Jenis auksin buatan yang biasa digunakan adalah IBA, 2,4-D dan ANA sedangkan yang alami biasa digunakan IAA (Katuuk, 1989). Asam Naftalen Asetat (ANA) adalah senyawa sintetis yang berhasil dibuat. Senyawa ini tidak mengandung ciri-ciri indol tetapi mempunyai aktifitas biologis seperti IAA, ANA dan 2,4-D merupakan golongan auksin sintetis yang mempunyai sifat lebih stabil dari pada IAA karena tidak mudah terurai oleh enzim-enzim yang dikeluarkan oleh sel atau oleh pemanasan pada proses sterilisasi. Sitokinin alami yang sering digunakan dalam kultur jaringan adalah zeatin, 2-iP, sedangkan untuk sintetik meliputi BAP dan kinetin (Wattimena, 1988).
pengatur tumbuh yang digunakan adalah kinetin. Kinetin (Furfuryl amino purine) merupakan zat pengatur tumbuh eksogen yang termasuk ke dalam golongan sitokinin (Gunawan, 1995). Kinetin adalah N6-furfuril adenin merupakan turunan basa adenin.
Kinetin belum pernah diisolasi dari jaringan-jaringan tanaman, tetapi berasal dari hasil-hasil kromatografi ekstrak tanaman dan kinetin diduga pada tanaman dalam konsentrasi yang rendah (Wattimena, 1998).
Menurut Yusnita, (2003), bahwa kinetin merupakan sitokinin sintetik yang pertama ditemukan oleh Carlos Miller pada ikan kering. Setelah itu ditemukan senyawa sitokinin yang lain dalam endosperma cair jagung, yaitu zeatin. Sitokinin sintetik lainnya adalah BAP (6-benzilaminopurin) dan 2-ip. Fungsi kinetin dapat mendorong pembelahan sel dari sel-sel kalus. Secara umum kinetin juga digunakan untuk menumbuhkan dan menggandakan tunas aksilar atau merangsang tumbuhnya tunas-tunas adventif dengan konsentrasi kinetin yang digunakan berkisar dari 0,1-1,0 mg/l. Gunawan (1995) menambahkan beberapa fungsi sitokinin, antara lain memacu pembelahan sel dalam jaringan meristematik, merangsang diferensiasi sel-sel yang dihasilkan dalam meristem, mendorong pertumbuhan tunas samping dan perluasan daun, menunda penuaan daun serta merangsang pembentukan pucuk dan mampu memecah masa istirahat biji (breaking dormancy).
2.3 Kultur Embrio
Kultur jaringan tanaman merupakan suatu metode atau teknik mengisolasi jaringan, organ, sel, maupun protoplas tanaman, menjadikan eksplan dan menumbuhkannya ke dalam media pertumbuhan yang aseptik sehingga eksplan tersebut dapat tumbuh dan berkembang, berorganogenesis dan dapat beregenerasi menjadi tanaman sempurna (Mattjik, 2005).
panas yang amat panas. Keadaan ini dapat diterapkan pada kultur biji. Kegunaan perbanyakan biji lainnya adalah menghilangkan kontaminasi eksternal seperti jamur, bakteri, dan mikroorganisme pengganggu lainnya yang dimusnahkan pada saat sterilisasi material tanaman (Katuuk, 1989).
Biji masing-masing spesies tanaman memiliki viabilitas tertentu. Misalnya pada tanaman pisang, tingkat kesuburan dan pembelahan sel pada biji lambat dan membutuhkan waktu yang lama untuk bereproduksi (Crouck et al., 1998). Sementara tanaman anggrek memiliki biji yang sangat lembut. Sehingga perbanyakan dengan biji membutuhkan perlakuan yang khusus. Biji adalah awal permulaan yang tepat dalam kultur jaringan (George & Sherrington, 1984). Setelah sterilisasi permukaan, biji dapat ditumbuhkan dalam media yang kaya akan nutrisi untuk membentuk kalus, atau dikecambahkan dalam media sederhana tanpa zat pengatur tumbuh untuk menghasilkan benih yang bebas dari kontaminan (George & Sherrington, 1984). Menurut Katuuk (1989) bahwa eksplan biji jeruk dapat diambil dari berbagai umur, tetapi sebaiknya dari buah yang muda yaitu 100-200 hari setelah polinasi.
Penelitian tentang kultur embrio sudah pernah dilakukan oleh Kosmiatin & Mariska (2005), embrio yang akan dikulturkan diisolasi dari polong yang disimpan di dalam lemari pendingin. Sebelum embrio diisolasi, polong disterilkan kemudian embrio diisolasi dengan cara membuka kulit biji. Selanjutnya embrio dikulturkan pada media perkecambahan. Embrio umur 1, 2, dan 3 minggu setelah polinasi dikulturkan pada medium dasar Knudson dan Knudson yang dimodifikasi dengan BA 1 mg/l. Sebagai pembanding, embrio dikulturkan pada medium MS yang ditambah dengan IAA 0,01 mg/l dan kinetin 0,1 mg/l (Gosal & Bajaj, 1983).
Kultur embrio belum matang yang diambil dari biji memiliki beberapa aplikasi, yaitu inkompatibilitas antar spesies atau kultivar yang timbul setelah pembentukan embrio akan menyebabkan aborsi embrio. Embrio seperti ini dapat diselamatkan dengan cara mengkulturkan embrio yang belum matang dan menumbuhkannya pada media kultur yang sesuai. Aplikasi lain yaitu untuk menyelamatkan embrio yang sudah matang agar tidak mati akibat serangan hama dan penyakit.
2.4 Planlet yang Baik
Plantlet yang baik adalah yang organnya lengkap, mempunyai pucuk dan akar, warna pucuknya hijau pekat artinya tidak tembus pandang, seperti kaca atau disebut vitrifikasi dan pertumbuhan akar bagus. Menurut Berliana (1998), ciri-ciri bibit yang berkulitas baik yaitu planlet tampak sehat dan tidak berjamur, ukuran planlet seragam, berdaun hijau segar dan tidak ada yang menguning. Selain itu planlet tumbuh normal, tidak kerdil, komposisi daun dan akar seimbang, pseudobulb atau umbi semu mulai tampak dan sebagian kecil telah mengeluarkan tunas baru, serta memiliki jumlah akar serabut 3 – 4 akar dengan panjang 1,5 – 2,5 cm.
BAB 3
BAHAN DAN METODA
3.1 Waktu dan Tempat Penelitian
Penelitian ini dilaksanakan pada bulan Oktober sampai dengan Desember 2010 di Laboratorium Kultur Jaringan Departemen Biologi, Fakultas Matematika dan Ilmu Pengetahuan Alam, Universitas Sumatera Utara, Medan. Pengamatan mikroskopik dilakukan di Laboratorium Ilmu Dasar Universitas Sumatera Utara. Sedangkan bahan tanaman berupa jeruk keprok berasal dari pasar Situmba, desa Gunung Tua Baringin, Kecamatan Sipirok, Kabupaten Tapanuli Selatan, Sumatera Utara.
3.2 Alat dan Bahan Penelitian
Adapun alat-alat yang digunakan dalam penelitian ini adalah autoklaf, entkas, lampu ultraviolet, botol kultur, aluminium foil, pipet serologi, alat diseksi, gelas piala, gelas ukur, neraca analitik, erlenmeyer, cawan petri, kertas saring, shaker, bunsen, pH meter, pipet serologi, pipet tetes, spatula, mikroskop, gelas objek dan penutup gelas.
Bahan penelitian yang digunakan adalah embrio jeruk keprok (Citrus nobilis Lour.) yang sudah masak, komposisi media MS, larutan pemutih (NaOCl 5,25%), fungisida benlate, tween, alkohol 70%, akuades, Ekstrak Malt (ME) 500 mg/l, kinetin, NAA 0,05 mg/l, agar-agar, gula, hara makro, hara mikro, iron, vitamin dan Myo-inositol.
3.3 Metoda Penelitian
K0 : 0 ppm
K1 : 1 ppm
K2 : 2 ppm
K3 : 3 ppm
K4 : 4 ppm
Dengan jumlah ulangan sebanyak enam pada setiap perlakuan, maka jumlah botol percobaan seluruhnya 30 satuan percobaan. Penelitian dilakukan dengan pengamatan sebanyak empat kali dalam waktu empat minggu, maka total botol seluruhnya adalah 120 satuan percobaan.
3.4 Prosedur Kerja 3.4.1 Sterilisasi Alat
Proses sterilisasi ini bertujuan agar seluruh alat yang digunakan terbebas dari kontaminasi. Alat-alat yang digunakan dalam percobaan ini seperti alat diseksi, cawan petri yang diisi kertas saring, botol kultur, erlenmeyer dan pipet volume dicuci dengan deterjen lalu dibilas dengan air mengalir, dikeringkan dan disterilisasi dalam autoklaf pada suhu 121 0C dengan tekanan 15 Psi selama 60 menit. Bersamaan dengan alat dimasukkan juga botol berisi akuades dan aluminium foil.
3.4.2 Pembuatan Media
Larutan MS dibuat dengan memasukkan hara makro, myo-inositol, dan sukrosa ke dalam gelas ukur 1000 ml yang terlebih dahulu telah berisi akuades. Lalu hara mikro, iron, vitamin, sukrosa, 500 mg/l Ekstrak Malt (ME) dan 0,05 mg/l NAA dimasukkan ke dalam gelas ukur yang ditambah akuades hingga 1000 ml, kemudian larutan dibagi menjadi 5 bagian sesuai perlakuan. Selanjutnya kinetin dimasukkan untuk masing-masing perlakuan. Derajat keasaman (pH) larutan diukur setiap perlakuan dengan menggunakan pH meter sebesar 5,8. Untuk mendapatkan pH yang optimal maka ditambahkan NaOH 0,1 N atau HCl 0,1 N.
Pada setiap perlakuan dimasukkan agar yang telah ditimbang sebanyak 3,5 g dan media dipanaskan sampai mendidih. Larutan dituang ke dalam botol kultur steril yang kemudian ditutup dengan aluminium foil dan diikat dengan karet, selanjutnya disterilkan dalam autoklaf pada suhu 1210C dengan tekanan 15 Psi selama 30 menit. Botol kultur ditempatkan di rak-rak kultur untuk menghindari kontaminasi (Reinert & Bajaj, 1989).
3.4.3 Sterilisasi Embrio
Eksplan berupa embrio dari biji jeruk dibersihkan di bawah air mengalir selama 30 menit, dibilas dengan akuades steril. Kulit biji terluar dilepaskan lalu direndam dalam benlate (0,2 g/100 ml) dan ditambahkan 2 tetes tween 20 lalu dishaker selama 120 menit. Biji tanpa kulit terluar tersebut direndam dengan alkohol 70% selama 1 menit, dibilas 3 kali dengan akuades steril, direndam dalam larutan pemutih 5% selama 5 menit dan dibilas 3 kali dengan akuades steril, selanjutnya dengan larutan pemutih 2,5% selama 5 menit, dibilas 3 kali dengan akuades steril. Biji tersebut diletakkan di dalam cawan petri kemudian dikeringkan dengan kertas saring steril. Berikut Gambar 3.4.3 biji yang di dalamnya terdapat embrio sebagai eksplan.
Gambar 3.4.3 Embrio Dari Biji Jeruk Keprok (a) Kulit Biji (b) Biji Utuh (c) Kotiledon (d) Embrio
3.4.4 Penanaman Embrio
Penanaman eksplan dilakukan di dalam entkas yang telah disterilisasi dengan sinar Ultraviolet (UV). Botol-botol berisi media yang sudah disterilkan, dibuka tutupnya dengan menggunakan pinset yang sudah dicelupkan pada alkohol dan telah dibakar. Lampu bunsen disediakan untuk mencegah kontaminasi. Kemudian setelah tutup botol dibuka, bagian sekitar tutup botol dilewatkan di atas api bunsen untuk memperkecil kontaminasi. Eksplan diambil dari dalam cawan petri dengan menggunakan pinset steril, lalu embrio digores pada permukaan untuk memperbesar keluarnya tunas lalu dimasukkan ke dalam botol kultur dengan menggunakan pinset steril. Botol kultur ditutup dengan aluminium foil dan disusun dirak kultur. Berikut Gambar 3.4.4 penanaman embrio pada media MS.
(a) (b) (c)
Gambar 3.4.4 Penanaman Embrio Pada Media MS (a) Media MS Sebelum Ditanam Embrio (b) Proses Penanaman Embrio (c) Media Berisi Embrio
3.4.5 Pemeliharaan Kultur
Eksplan yang telah ditanam di dalam botol kultur diletakkan pada rak pemeliharaan dengan kondisi ruangan yang steril, suhu berkisar 25oC. Intensitas cahaya dengan penyinaran lampu neon 500 lux. Botol-botol yang berisi eksplan tersebut disusun dengan rapi sehingga memudahkan dalam pengamatan. Diupayakan ruangan dalam keadaan steril atau dengan menyemprotkan alkohol 70% setiap hari.
3.4.6 Pengamatan Mikroskopik
perlakuan dan pengamatan setiap minggunya. Bagian akar diiris tipis secara melintang lalu diamati di bawah mikroskop cahaya dengan perbesaran 4x10.
3.5 Variabel Pengamatan
Pengamatan dilakukan empat kali dengan interval waktu satu minggu. Variabel yang diamati dalam penelitian ini adalah :
a. Tipe Pertumbuhan Kultur
h. Pengamatan mikroskopik akar dari planlet dengan objek preparat segar yang diambil dari setiap pengamatan (interval pengambilan planlet 1 minggu)
i
.
Persentase kultur yang hidup (%)Jumlah eksplan yang tumbuh
Persentase kultur yang hidup = X 100 %
Jumlah eksplan seluruh perlakuan
j.
Persentase kontaminasi keseluruhan (%)Persentase kultur terkontaminasi dihitung setiap hari sejak awal hingga akhir
Jumlah eksplan yang terkontaminasi
Persentase terkontaminasi = X 100 %
Jumlah eksplan seluruh perlakuan
3.6Analisis Data
Data penelitian menggunakan metode RAL non-faktorial ini dianalisis dengan
Análysis of Variant (ANOVA). Sedangkan untuk menguji beda nyata antara perlakuan
dilakukan dengan Uji Jarak Duncan (UJD) atau disebut dengan Duncan Multiple
BAB 4
HASIL DAN PEMBAHASAN
4.1 Tipe Pertumbuhan Kultur
Hasil penelitian kultur embrio jeruk keprok pada media MS + ME + NAA dengan penambahan berbagai variasi kinetin diperoleh 100% tunas planlet. Hal ini sesuai dengan persentase yang diharapkan yaitu memperoleh 75% hasil kultur hidup dan bebas kontaminasi. Penelitian menggunakan perbandingan konsentrasi auksin lebih kecil dari konsentrasi sitokinin, sehingga menghasilkan planlet. Diketahui bahwa rasio auksin dan sitokinin jika diberikan dengan perbandingan konsentrasi yang sama, maka akan menghasilkan kalus. Namun dengan konsentrasi lebih besar atau lebih kecil akan menghasilkan akar dan tunas.
Pada tunas sudah terbentuk daun dan batang berwarna hijau. Menurut Yusnita (2003), bahwa ciri-ciri tunas yang baik adalah daun berwarna hijau dan hijau tua. Pemeliharaan kultur dengan cahaya optimal mampu merangsang eksplan untuk menghasilkan klorofil yang mendukung untuk pertumbuhan tunas planlet. Setelah 5 hari tunas planlet terbentuk dari sisi embrio yang dilukai. Dengan demikian diketahui bahwa media MS dengan penambahan kinetin 0,1,2,3 dan 4 ppm merupakan media yang baik untuk pertumbuhan tunas planlet jeruk keprok. Berikut Gambar 4.1 planlet dalam media MS + kinetin.
Gambar 4.1 Planlet Dalam Media MS + Kinetin (a) Daun (b) Batang (c) Media (d) Akar
b
d a
4.2 Jumlah Tunas
Berdasarkan pada Lampiran C sidik ragam, kinetin memberikan pengaruh berbeda nyata terhadap jumlah tunas. Perbandingan jumlah tunas dapat dilihat pada Tabel 4.2 di bawah.
(2 minggu), M3 (3 minggu), M4 (4 minggu). Angka-angka pada kolom yang sama bila diikuti dengan
huruf yang tidak sama berbeda nyata pada taraf 5% (huruf kecil), 1% (huruf besar) menurut Uji Jarak Duncan (UJD).
Tabel 4.2 menunjukka n jumlah tunas tertinggi pada pengamatan minggu ke tiga terdapat pada perlakuan K2 yaitu 2.83 yang tidak berbeda nyata terhadap perlakuan K1
dan K3 serta berbeda sangat nyata dengan perlakuan K0 dan K4. Perlakuan kinetin
pada minggu pertama menunjukkan jumlah tunas tertinggi pada perlakuan K4 yaitu
2.50 dan terendah yaitu K1 yaitu 1.16. Pada minggu ke dua jumlah tunas tertinggi
terdapat pada perlakuan K1 dan K3 yaitu 2.16 dan terendah pada K4 yaitu 1.66. Pada
minggu ke tiga jumlah tunas tertinggi terdapat pada perlakuan K2 yaitu 2.83 dan
terendah pada K0 yaitu 1.00. Pada minggu ke empat jumlah tunas tertinggi terdapat
pada perlakuan K2 yaitu 2.83 dan terendah pada K1 yaitu 1.16. Berikut Gambar 4.2.1
grafik kuadratik pada pengamatan minggu ke tiga.
Pada Gambar 4.2.1 diketahui pada minggu ke tiga terjadi peningkatan jumlah tunas dari konsentrasi 0 ppm (K0), 1 ppm (K1), sampai 2 ppm (K2). Namun pada konsentrasi
3 ppm (K3) dan 4 ppm (K4) terjadi penurunan jumlah tunas. Hal ini menunjukkan
bahwa konsentrasi optimum untuk jumlah tunas terdapat pada perlakuan 2 ppm (K2).
Sesuai dengan prinsip kerja hormon tumbuhan yaitu pada konsentrasi rendah mampu menimbulkan suatu respon fisiologis (Salisbury & Ross, 1992).
Kinetin berpengaruh terhadap penggandaan tunas melalui kultur jaringan yang dapat terlihat dari bertambahnya jumlah tunas pada konsentrasi optimum. Kinetin mampu memacu pembentukan tunas pada kultur dengan cara meningkatkan pembelahan dan pembesaran sel. Hal ini sesuai dengan pendapat George & Sherington (1984), bahwa kinetin sangat efektif mendukung secara langsung maupun tidak langsung dalam pembentukan pucuk. Secara umum kinetin juga digunakan untuk menumbuhkan dan menggandakan tunas aksilar atau merangsang tumbuhnya tunas-tunas adventif (Yusnita, 2003). Dengan konsentrasi kinetin yang digunakan berkisar dari 0.1-1.0 mg/l (Gunawan, 1995).
Sitokinin dalam konsentrasi rendah akan dapat memacu perkembangan tunas sedangkan sitokinin konsentrasinya tinggi dalam media kultur akan dapat merangsang pembentukan atau penggandaan tunas (Magoon & Singh, 1995; Goh et al., 1995
dalam Nurwahyuni, 2005). Menurut Kusumo (1984) dalam Kasi (2008), kinetin
berperanan dalam pembelahan sel dan morfogenesis, sedangkan auksin berperanan dalam mengatur pertumbuhan dan pemanjangan sel. Hal ini menunjukkan bahwa sitokinin dan auksin berperanan saling melengkapi dalam menginduksi tunas. Eksplan yang membentuk tunas dapat dilihat pada Gambar 4.2.2 berikut:
Gambar 4.2.2 Eksplan Membentuk Tunas Pada Perlakuan K23.2
Dari Gambar 4.2.2 dapat dilihat bahwa terdapat tunas yang bercabang yang tumbuh dari embrio jeruk. Menurut Hartmann & Kester (2002), bahwa jika dalam media kultur diberikan konsentrasi sitokinin lebih tinggi dibandingkan dengan auksin maka akan merangsang pembentukan dan multiplikasi tunas.
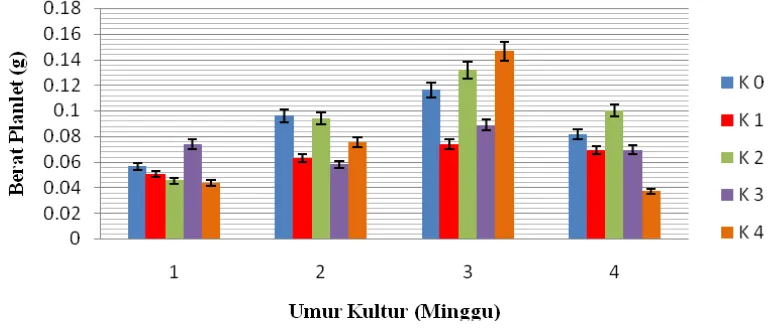
4.3 Berat Planlet (g)
Berdasarkan pada Lampiran D sidik ragam, kinetin memberikan pengaruh berbeda nyata terhadap berat planlet. Perbandingan berat planlet dapat dilihat pada Tabel 4.3 di bawah ini.
Tabel 4.3 Rata-Rata Berat Planlet (g)
Waktu Pertumbuhan
huruf yang tidak sama berbeda nyata pada taraf 5% (huruf kecil), 1% (huruf besar) menurut Uji Jarak Duncan (UJD).
Tabel 4.2 dapat diketahui planlet pada minggu pertama yang memiliki berat tertinggi pada perlakuan K3 yaitu 0,073 yang tidak berbeda nyata terhadap perlakuan K0 dan
berbeda sangat nyata terhadap perlakuan K1, K2 dan K4. Pada minggu pertama berat
planlet tertinggi pada perlakuan K3 yaitu 0.073 dan terendah K4 yaitu 0.043. Pada
minggu ke dua berat planlet tertinggi pada perlakuan K2 yaitu 0.098 dan terendah K3
yaitu 0.058. Pada minggu ke tiga berat planlet tertinggi pada perlakuan K4 yaitu 0.146
dan terendah K1 yaitu 0.073. Pada minggu ke empat berat planlet tertinggi terdapat
pada perlakuan K2 yaitu 0.105 dan terendah K4 yaitu 0.037.
Pada penelitian ini nilai berat planlet tertinggi diperoleh pada perlakuan K3
pengaruh terhadap berat basah kultur. Katuuk (1989), menambahkan bahwa sitokinin yang dikombinasikan dengan auksin dapat memacu pembesaran sel yang diikuti dengan meningkatnya bobot kultur terutama oleh meningkatnya pengambilan air oleh sel tersebut. Respon kinetin terhadap berat planlet untuk setiap minggunya dapat dilihat pada Gambar 4.3.1 di bawah.
Gambar 4.3.1 Respon Kinetin Terhadap Berat Planlet
Gambar 4.3.1 di atas dapat dilihat bahwa setiap minggu penambahan kinetin cenderung menurunkan berat planlet kecuali pada minggu ke tiga. Terdapat perbedaan terhadap konsentrasi kinetin yang menghasilkan berat planlet terbaik, hal ini mungkin disebabkan oleh faktor genetik dari eksplan yang digunakan memiliki perbedaan dalam merespon suatu perlakuan. Hal ini yang menjadi kelemahan dari data destruktif, karena hasil kultur yang diamati setiap minggu berasal dari eksplan berbeda, sehingga pola pertumbuhan tidak selalu akurat. Ketidakseragaman dalam pemilihan eksplan yang digunakan dapat juga menjadi faktor perbedaan hasil berat planlet setiap minggunya. Ekplan berasal dari embrio jeruk yang berbeda-beda kondisi genetiknya, sehingga hasil pertumbuhannya menunjukkan ketidakseragaman. Seperti yang dinyatakan oleh Gunawan (1987) , bahwa setiap sel, jaringan organ dan tanaman yang berbeda memberikan respon yang berbeda terhadap media tumbuh yang sama.
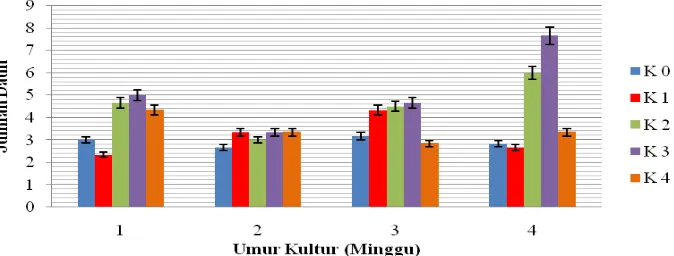
4.4 Jumlah Daun (helai)
Berdasarkan Lampiran E sidik ragam, kinetin memberikan pengaruh berbeda nyata terhadap jumlah daun. Perbandingan jumlah daun dapat dilihat pada Tabel 4.4 di bawah ini.
Tabel 4.4 Rata-Rata Jumlah Daun (helai)
Waktu Pertumbuhan
Dari Tabel 4.4 dapat diketahui bahwa jumlah daun tidak berbeda nyata untuk semua perlakuan. Pada minggu pertama jumlah daun tertinggi terdapat perlakuan K3 yaitu
5.00 dan terendah pada K1 yaitu 2.33. Pada minggu ke dua jumlah daun tertinggi pada
perlakuan K1, K3, K4 yaitu 3.33 dan terendah pada K0 yaitu 2.66. Pada minggu ke tiga
jumlah daun tertinggi pada perlakuan K3 yaitu 4.66 dan terendah K4 yaitu 2.83. Pada
minggu ke empat jumlah daun tertinggi pada perlakuan K3 yaitu 7.66 dan terendah
pada K1 yaitu 2.66.
Pada konsentrasi kinetin yang tinggi yaitu 4 ppm (K4) pertumbuhan jumlah
daun meningkat pada minggu pertama dan kedua, sedangkan pada minggu ke tiga dan ke empat pertumbuhan jumlah daun menurun. Respon kinetin terhadap jumlah daun untuk setiap minggunya dapat dilihat pada Gambar 4.4.1 di bawah.
Gambar 4.4.1 diketahui bahwa kinetin cenderung memengaruhi jumlah daun seiring dengan peningkatan konsentrasi kinetin. Kinetin yang termasuk dalam sitokinin mampu menghasilkan jumlah helai daun yang tinggi. Menurut Barden et al., (1987) bahwa perkembangan sel pada daun sebagian dikontrol oleh sitokinin. Sitokinin yang ditambahkan pada media kultur jaringan dapat merangsang pembelahan sel dan diferensiasi sel. Daun Menurut Davies (1987) dalam Bintang (2002), mengatakan bahwa sitokinin yang optimum akan lebih memengaruhi pembentukan daun dari pada pembentukan organ tanaman lain. Berikut Gambar 4.4.2 eksplan yang membentuk daun.
Gambar 4.4.2 Eksplan Membentuk Daun Pada Perlakuan K34.1
Penelitian Veronika (2007), apabila dibandingkan pengaruh jumlah daun, dihasilkan atonik 1,5 ml/L + kinetin 0,1 mg/L menunjukka n jumlah daun terendah dan pada perlakuan kinetin 2 mg/L, menghasilkan jumlah daun yang lebih banyak. Hal ini menunjukkan bahwa pemberian kinetin tunggal lebih baik dalam memacu pembentukan daun dari pada pemberian kinetin yang dikombinasikan dengan atonik.
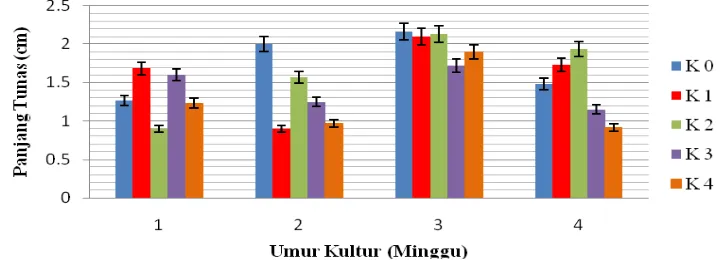
4.5 Panjang Tunas (cm)
Berdasarkan Lampiran F sidik ragam, kinetin memberikan pengaruh yang tidak berbeda nyata terhadap panjang tunas. Perbandingan panjang tunas dapat dilihat pada Tabel 4.5 di bawah ini.
Tabel 4.5 Rata-Rata Panjang Tunas (cm)
Tabel 4.5 dapat diketahui bahwa panjang tunas tidak berbeda nyata untuk semua perlakuan. Panjang tunas tertinggi pada minggu pertama terdapat pada perlakuan K1
yaitu 1.68 dan terendah pada K2 yaitu 0.90. Pada minggu ke dua panjang tunas
tertinggi terdapat pada perlakuan K0 yaitu 2.00 dan terendah pada K1 yaitu 0.90. Pada
minggu ke tiga panjang tunas tertinggi pada perlakuan K0 yaitu 2.16 dan terendah
pada K3 yaitu 1.71. Pada minggu ke empat panjang tunas tertinggi terdapat pada
perlakuan K2 yaitu 1.93 dan terendah yaitu K4 0.90. Respon kinetin terhadap panjang
tunas untuk setiap minggunya dapat dilihat pada Gambar 4.5.1 di bawah.
Gambar 4.5.1 Respon Kinetin Terhadap Panjang Tunas
Dari Gambar 4.5.1 dapat diketahui bahwa pada grafik dapat dilihat pada minggu pertama pertumbuhan panjang tunas terjadi secara fruktuatif. Pada minggu ke dua panjang tunas cenderung mengalami penurunan. Pada minggu ke tiga panjang tunas cenderung konstan. Pada minggu ke empat panjang tunas cenderung membentuk pola kuadratik.
Secara umum, pada minggu ke dua perlakuan K0 menunjukkan panjang tunas
penambahan auksin dan sitokinin eksogen kedalam media kultur tidak selalu dibutuhkan. Salisbury & Ross (1995) menambahkan bahwa batang dan akar yang sedang memanjang tidak membutuhkan sitokinin dalam jumlah tinggi, meskipun tanaman harus membutuhkan sitokinin untuk perpanjangan batang dan akar, kandungan sitokinin endogennya sudah mencukupi. Hal ini diduga bahwa sitokinin eksogen akan menghambat pertumbuhan in vitro yang disebabkan karena konsentrasi sitokinin tersebut dalam tanaman menjadi berlebihan. Hal tersebut sesuai dengan penelitian ini yang memberikan pengaruh panjang tunas terbaik pada perlakuan tanpa penambahan kinetin. Berikut Gambar 4.5.2 perbadingan panjang tunas pada setiap konsentrasi kinetin.
Gambar 4.5.2 Perbandingan Panjang Tunas Pada Perlakuan K0, K1, K2, K3, K4. (a) (b) (c) (d) (e) Pembentukan Tunas
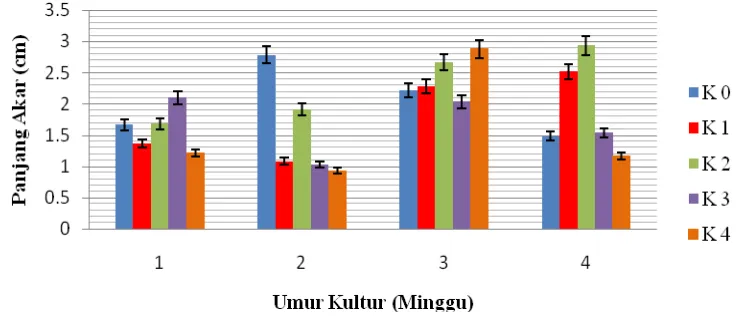
4.6 Panjang Akar (cm)
Berdasarkan Lampiran G sidik ragam, kinetin berpengaruh nyata terhadap panjang akar. Perbandingan panjang akar dapat dilihat pada Tabel 4.6 di bawah ini.
Tabel 4.6 Rata-Rata Panjang Akar (cm)
(2 minggu), M3 (3 minggu), M4 (4 minggu). Angka-angka pada kolom yang sama bila diikuti dengan
huruf yang tidak sama berbeda nyata pada taraf 5% (huruf kecil), 1% (huruf besar) menurut Uji Jarak Duncan (UJD).
a
b c
Tabel 4.6 di atas menunjukka n bahwa dari empat minggu pengamatan ternyata pengaruh kinetin terhadap panjang akar berbeda nyata pada minggu ke dua. Pada minggu ke dua tersebut, panjang akar tertinggi terdapat pada perlakuan K0 yaitu 2.78
yang tidak berbeda nyata pada K2 dan berbeda nyata pada K1, K3 dan K4. Pada minggu
pertama panjang akar tertinggi terdapat pada perlakuan K3 yaitu 2.10 dan terendah K4
yaitu 1.21. Pada minggu ke dua panjang akar tertinggi terdapat pada perlakuan K0
yaitu 2.78 dan terendah K4 yaitu 0.93. Pada minggu ke tiga panjang akar tertinggi
terdapat pada perlakuan K4 yaitu 2.88 dan terendah pada K3 yaitu 2.03. Pada minggu
ke empat panjang akar tertinggi terdapat pada perlakuan K2 yaitu 2.93 dan terendah K4
yaitu 1.16. Respon kinetin terhadap jumlah akar untuk setiap minggunya dapat dilihat pada Gambar 4.6.1 di bawah.
Gambar 4.6.1 Respon Kinetin Terhadap Panjang Akar
Gambar 4.6.1 dapat dilihat bahwa pada minggu ke dua pemberian konsentrasi kinetin yang semakin meningkat akan cenderung menurunkan pertumbuhan panjang akar. Dalam hal ini peningkatan konsentrasi kinetin yang diberikan ke dalam media akan menghambat pertumbuhan akar karena kinetin endogen yang ada dalam eksplan sudah mencukupi untuk pertumbuhan akar.
sitokinin lebih rendah dari auksin, maka ini akan mengakibatkan stimulasi pada pertumbuhan akar.
Menurut Gunawan (1995), bahwa pemberian sitokinin eksogen dengan konsentrasi tinggi ditambah dengan adanya sitokinin endogen yang tinggi di dalam akar akan menghambat pertumbuhan dan pembentukan akar tanaman. Hal ini juga sesuai dengan pendapat Salisbury & Ross (1995) bahwa batang dan akar yang sedang memanjang terkadang tidak memerlukan sitokinin walaupun kedua organ ini mungkin membutuhkan hormon tersebut untuk memanjang, karena kandungan sitokinin dalam jaringan sudah mencukupi. Berikut Gambar 4.6.2 panjang akar pada perlakuan 0,1,2,3 dan 4 ppm.
Gambar 4.6.2 Perbandingan Panjang Akar Pada Perlakuan K0, K1, K2, K3, K4. (a) (b) (c) (d) (e) Pembentukan Akar
Gambar 4.6.2 di atas dapat dilihat bahwa akar berkembang dengan baik sehingga dapat menyerap unsur hara secara optimal dari media MS. Pembentukan akar pada kultur jaringan biasanya diinduksi setelah terbentuk tunas. Namun akar juga dapat langsung terbentuk dari pucuk maupun planlet, jika ke dalam media diberikan sitokinin yang mencukupi. Mattjik (2005) menyatakan bahwa bila eksplan pada kultur pertama langsung membentuk akar, maka sedikit kemungkinannya dapat membentuk tunas.
Penelitian Veronika (2007), apabila dibandingkan pengaruh panjang akar, dihasilkanatonik 1,5 ml/L + kinetin 0,1 mg/L memperlihatkan panjang akar terendah bila dibandingkan dengan kinetin 2 mg/L. Hal ini menunjukkan bahwa pemberian
kinetin tunggal lebih baik dalam memacu pembentukan panjang akar dari pada pemberian kinetin yang dikombinasikan dengan atonik.
4.7 Jumlah Akar
Berdasarkan Lampiran H sidik ragam, kinetin tidak berpengaruh nyata terhadap jumlah akar. Perbandingan jumlah akar dapat dilihat pada Tabel 4.7 di bawah.
Tabel 4.7 Rata-Rata Jumlah Akar
Waktu Pertumbuhan Kultur
Perlakuan M 1 M 2 M 3 M 4
K 0 1.00 2.83 1.16 1.33
K 1 2.00 1.00 1.50 2.50
K 2 1.00 2.50 2.16 1.50
K 3 1.00 0.83 1.33 0.66
K 4 1.33 1.50 1.00 0.66
Keterangan: K0 (0 ppm), K1 (1 ppm), K2 (2 ppm), K3 (3 ppm), K4 (4 ppm), M1 (1 minggu), M2 (2
minggu), M3 (3 minggu), M4 (4 minggu)
Tabel 4.7 di atas diketahui bahwa jumlah akar tidak berbeda nyata untuk semua perlakuan. Minggu pertama menunjukkan jumlah akar tertinggi pada perlakuan K1
yaitu 2.00 dan terendah K0, K2 dan K3 yaitu 1.00. Pada minggu ke dua jumlah akar
tertinggi pada perlakuan K0 yaitu 2.83 dan terendah K3 yaitu 0.83. Pada minggu ke
tiga jumlah akar tertinggi terdapat pada perlakuan K2 yaitu 2.16 dan terendah K4 yaitu
1.00. Pada minggu ke empat jumlah akar tertinggi terdapat pada perlakuan 2.50 dan terendah K3 dan K4 yaitu 0.66. Respon kinetin terhadap jumlah akar dengan untuk
setiap minggunya dapat dilihat pada Gambar 4.7.1 di bawah.
Gambar 4.7.1 dapat dilihat bahwa perlakuan kinetin pada minggu pertama cenderung konstan. Pada minggu ke dua cenderung menurun. Pada minggu ke tiga membentuk pola grafik kuadratik. Pada minggu ke empat cenderung menurun terhadap jumlah akar. Pada grafik minggu ke dua menunjukkan bahwa perlakuan K0 menghasilkan
jumlah akar tertinggi dibandingkan dengan perlakuan K1, K2, K3 dan K4. Pemberian
konsentrasi yang semakin meningkat akan menurunkan jumlah akar. Hal ini diduga penambahan sitokinin eksogen tidak diperlukan lagi. Sitokinin dalam hal ini kinetin berfungsi menekan pertumbuhan akar. Adanya hormon endogen pada eksplan mungkin sudah mencukupi untuk memacu pertumbuhan akar.
Menurut Gunawan (1995), bahwa pemberian sitokinin eksogen dengan konsentrasi tinggi ditambah dengan adanya sitokinin endogen yang tinggi didalam akar akan menghambat pembentukan akar. Hal ini sesuai dengan pendapat Salisbury & Ross (1995), bahwa batang dan akar yang sedang memanjang terkadang tidak memerlukan sitokinin walaupun kedua organ ini mungkin membutuhkan hormon tersebut untuk memanjang, karena kandungan sitokinin dalam jaringan sudah mencukupi. Van & Trinh (1990) dalam Marlin (2005), menyatakan bahwa suplai hormon secara endogen sangat mempengaruhi proses morfogenesis pada kultur in
vitro, sejalan dengan laju metabolismenya. Dengan demikian, tanpa suplai auksin dan
sitokinin secara eksogen, akar tanaman akan tetap tumbuh dan memanjang. Menurut Hartmann (2002), bahwa IAA dan sitokinin banyak diproduksi pada biji yang sedang tumbuh. Berikut Gambar 4.7.2 eksplan membentuk akar pada konsentrasi 0,1,2,3 dan 4 ppm.
Gambar 4.7.2 Perbandingan Jumlah Akar Pada Perlakuan K0, K1, K2, K3, K4. a) (b) (c) (d) (e) Pembentukan Akar
a
b c
d
4.8 Pengamatan Mikroskopik Anatomi Akar
Alur kerja pengamatan mikroskopik akar dapat dilihat pada Lampiran J. Dan hasil pengamatan mikroskopik akar dapat dilihat pada Lampiran M. Dari hasil pengamatan mikroskopik akar segar yang diiris secara melintang. Pada akar muda bila dilakukan potongan melintang akan terlihat bagian-bagian dari luar ke dalam yaitu epidermis, korteks, endodermis, xilem, floem dan silinder pusat.
Menurut Tjitrosomo (1983), jaringan akar muda dapat ditemukan tiga daerah utama yaitu silinder pembuluh, korteks dan epidermis. Ketiganya dapat dikenali dengan mikroskop dari irisan melintang bagian akar. Ditengahnya terdapat silinder pembuluh yang dibangun oleh jaringan pembuluh dan parenkim. Bulu akar merupakan modifikasi dari sel epidermis akar, bertugas menyerap air dan garam-garam mineral terlarut, bulu akar memperluas permukaan akar. Sedangkan korteks letaknya langsung di bawah epidermis, sel-selnya tidak tersusun rapat sehingga banyak memiliki ruang antar sel. Endodermis merupakan lapisan pemisah antara korteks dengan silinder pusat. Sel-sel endodermis dapat mengalami penebalan zat gabus pada dindingnya dan membentuk seperti titik-titik, dinamakan titik
caspary. Lalu silinder pusat merupakan bagian terdalam dari akar. Sel-sel xilem yang
berdinding tebal menyalurkan air dan mineral, sedangkan floem yang menyalurkan bahan makanan. Untuk lebih jelas dapat dilihat pada Gambar 4.8 di bawah ini.
Gambar 4.8 Anatomi Akar Pada Kontrol dan K4 Pada Umur Planlet 4 Minggu
(a) Epidermis (b) Hipodermis (c) Korteks (d) Endodermis (e) Xilem (f) Floem
Pengamatan mikroskopik akar pada kontrol dan perlakuan tidak menunjukkan perbedaan anatomi yang signifikan. Akar pada kontrol dan perlakuan memiliki akar normal. Panjang akar dan panjang tunas seimbang. Akar memiliki tekstur yang padat dan tidak banyak mengandung air. Akar yang normal dan tidak banyak mengandung air memudahkan dalam penyayatan tipis pada akar. Sayatan yang tipis tersebut memudahkan dalam mengamati di bawah mikroskop cahaya dan bagian anatomi lebih mudah dibedakan.
4.9 Persentase Kultur yang Hidup (%)
Dari penelitian yang didapat bahwa persentasi kultur yang hidup mencapai 100% dari total keseluruhan 120 botol. Walaupun terjadi kontaminasi sebanyak 10% namun planlet sudah berhasil tumbuh. Hal ini mungkin disebabkan karena kemampuan tanaman tersebut bertahan pada kondisi ruangan kultur ataupun ruang di dalam botol. Menurut Santoso & Nursandi (2004), hampir dapat dipastikan bahwa keberhasilan kegiatan kultur jaringan sangat ditentukan dan tergantung oleh pemilihan media yang digunakan. Teknik kultur jaringan menekan lingkungan yang cocok agar eksplan dapat tumbuh dan berkembang. Wareigh & Philips (1998) dalam Marlin (2005) menambahkan bahwa zat pengatur tumbuh sangat berperan penting untuk memacu proses pertumbuhan in vitro. Oleh sebab itu perlu konsentrasi yang tepat karena zat pengatur tumbuh berbeda-beda untuk setiap jenis tanaman yang digunakan.
4.10 Persentase Kultur Terkontaminasi (%)
yang kurang bersih dan terdapat botol kultur lain yang telah terkontaminasi namun tidak segera dipindahkan.
Jumlah kultur yang terkontaminasi tergolong kecil. Nilai persentase jumlah kultur terkontaminasi yang kecil ini juga menunjukka n bahwa teknik sterilisasi yang dilakukan sudah mencapai tahap sempurna. Sterilisasi yang dilakukan meliputi sterilisasi alat, eksplan, media dan ruangan. Kultur jaringan memerlukan kecermatan tinggi dan keadaan serba steril baik tempat keja, alat-alat maupun bahan yang digunakan serta tangan orang yang bekerja harus selalu steril (Rahardja, 1994).
Pada penelitian ini, kultur terkontaminasi oleh bakteri dan jamur. Pada kultur yang terkontaminasi oleh jamur ditandai oleh munculnya hifa-hifa halus berwarna cokelat dan hitam pada media kultur. Kontaminasi ini diakibatkan oleh rusaknya aluminium foil sebagai penutup botol kultur sehingga kemungkinan terjadi kontak dengan lingkungan luar yang memudahkan jamur untuk masuk dan mengkontaminasi media. Jamur ini akan berkembang dengan cepat karena nutrisi yang dibutuhkannya tersedia pada media. Hal ini tentu saja akan menghambat aktifitas pertumbuhan eksplan sampai akhirnya mematikan sel-sel eksplan.
Ini sesuai dengan pendapat Gunawan (1988) dalam Sitepu (2007), bahwa kontaminasi dapat terjadi setiap saat dalam masa kultur, yang mungkin berasal dari eksplan, mikroba, lingkungan kerja, serta kecerobohan dalam pelaksanaan kultur jaringan. Pada media kultur yang mengandung gula, vitamin dan mineral sangat disenangi jamur dan bakteri untuk tumbuh. Katuuk (1989) dalam Sitepu (2007), menyatakan bahwa bila kontaminasi tidak dihilangkan maka media yang mengandung gula, vitamin dan mineral akan tumbuh jamur dan bakteri secara cepat karena media tersebut disenangi jamur dan bakteri.
BAB 5
KESIMPULAN DAN SARAN
5.1Kesimpulan
a. Tipe pertumbuhan kultur embrio jeruk keprok pada media MS dengan penambahan kinetin adalah planlet, dengan 100% kultur hidup dan 10 % kontaminasi.
b. Penambahan kinetin dalam kultur embrio jeruk keprok pada media MS berpengaruh nyata terhadap variabel jumlah tunas, berat planlet dan panjang akar.
c. Dari pengamatan minggu keempat, konsentrasi kinetin terbaik untuk inisiasi jumlah tunas, berat planlet, panjang tunas dan panjang akar yaitu 2 ppm (K2).
Konsentrasi kinetin terbaik untuk jumlah daun adalah 3 ppm (K3). Dan
konsentrasi kinetin terbaik untuk jumlah akar adalah 4 ppm (K4).
d. Semua variabel memberikan respon yang baik terhadap media MS tanpa penambahan kinetin.
5.2 Saran
a. Peneliti harus menggunakan jeruk keprok yang berkualitas lebih baik, sehingga eksplan berupa embrio jeruk keprok memiliki kemampuan untuk tumbuh optimal.
DAFTAR PUSTAKA
Abidin, Z. 1985. Dasar-dasar Pengetahuan tentang Zat Pengatur Tumbuh. Bandung: Angkasa.
Agisimanto, D., Huse, S., Sullistiowati, E. 2006. Induksi Tunas Jeruk Pacitan (Citrus
sinensis L. Osbeck cv. Pacitan) Pada Beberapa Konsentrasi BAP. Balai
Penelitian Tanaman Jeruk dan Buah Subtropika. Malang: Universitas Muhammadiyah Malang.
Bahrany, A. M. 2002. Effect of Phytohormones on In Vitro Shoot Multiplication and
Rooting of Lime Citrus aurantifolia (Chrism) Swing. Scientis Holticulture.
Ball, J. S. 1997. Fruit Growing. New Delhi: Kalyani Publishers. Barden, J. A. 1987. Plant Science. America: McGraw-Hill, Inc.
Barus, W.A 1999. Pengaruh Aplikasi Nitrogen dan Kinetin terhadap Pertumbuhan, Hasil dan Komponen Mutu Tembakau Deli. Tesis. Prgram Pasca Sarjana. Medan: Universitas Sumatera Utara.
Begum, F.. Amin, Azad. 2003. In Vitro Plant Regeneration Cotyledone Derived Callus of Tree Varietas Pummelo (Citrus grandis) L.osb. Biological
sciences 3(8) 752.
Berliana, Y. 1998. Optimasi Konsentrasi Sukrosa dan Adaptasi Awal Planlet Salak (Salacca sumatrana Beccari). Pada Kondisi Kelembapan Rendah. Tesis. Medan: Program Pasca Sarjana USU. Hal: 8.
Bintang, R. 2002. Pengaruh IAA dan Kinetin Pada Kultur Biji Jeruk Brastepu (Citrus
nobolis Lour. Var. Brastepu) Pada Media MS. Skripsi. Medan: Fakultas
Matematika Dan Ilmu Pengetahuan Alam. USU.
Butar-Butar, R. 2006. PertumbuhanKultur Biji Jeruk Brastepu (Citrus nobilis Lour. Var. Brastepu) Pada Media MS Diperkaya Atonik dan Kinetin. Skripsi. Medan. Fakultas Matematika dan Ilmu Pengetahuan Alam. USU.
Chapman, A., Blervacq, A. S., Tissier, J. P., Delbreil, B., Vasseur, J., and Hilbert, J. L. 2000. Cell Wall Differentiation During Early Somatic Embryogenesis in Plants. I. Scaning and Tranmission Elevtron Microscopy Study on Embryos Originating From Direct, Indirect, and Adventitios Pathways. Canadian
Crouck, J. H., Vuylsteke, D., and Ortiz, R. 1998. Perspevtive on The Aplication of Biotechnology to Assist The Genetic Enchasment of Plaintain and Banana (Musa spp.). EJB Electrinoc Journal of Biotechnology 1(1).
Davies, T. J. 1993. Plant Hormones and There Prolea in Plant Growth and Development: Martinus Nijhoff Publisher. pp.15
Departemen Pertanian, 1997. Kultur Jaringan. http//www.deptan.co.id. Diakses pada tanggal 17 Maret 2010.
Devi, N. F. Hardiyanto, Jati, 2007. Penelitian Tanaman Jeruk & Buah Subtropika. Jurnal Holtikultura Edisi Khusus 3 : 229-238.
George, E. T. and Sherrington, P. D. 1984. Plant Propagation by Tissue Culture.
Handbook and Directory of Commercial Laboratories. England: Exegetics
Limited. Eastern Press. Hal: 7.
Gosal, S. S. and Y. P. S. Bajaj. 1983. Interspecific Hybridization Between V. mungo and V. radiata Through Embryo Culture. Euphytica 32: 129-137.
Gunawan, L. W. 1995. Teknik Kultur In Vitro Dalam Holtikultura. Jakarta: Penebar Swadaya. Hal: 6, 41-43, 50-51.
Hartmann, H. T dan Wijayani, A. 1983. Plant Propagation Principles and Practice. 4th Edition. Pretince Mall, Inc. Englewood cliffs: New Jersy.
Hartmann, H. T., Kester, D., E, Davies, F. T., Geneve, R.L. 2002. Plant Propagation
Principles and Practies. 6th Editon. New Delhi: Prentice Hall Of India Private Limited.
Heddy, S. 1986. Hormon Tumbuhan. Jakarta: Penerbit Rajawali. Hal: 3.
Hendaryono, D. P. S. & A. Wijayani. 1994. Kultur Jaringan (Pengenalan dan
Petunjuk Perbanyakan Tanaman Secara Vegetatif Modern). Yogyakarta:
Kanisius.
Joesoef, M. 1993. Penuntun Berkebun Jeruk. Jakarta: Bhratara.
Kanisius, A. A. 1994. Budidaya Tanaman Jeruk. Yogyakarta: Kanisius.
Kasi, P. D dan Sumaryono. 2008. Perkembangan Kalus Embriogenik Sagu (Metroxylon sagu Rottb.) Pada Tiga Sistem Kultur In Vitro. Menara
Perkebunan76(1): 1-10.
Katuuk, J. R. P. 1989. Teknik Kultur Jaringan, Pengenalan dan Petunjuk
Perbanyakan Tanaman Secara Vegetatif Modern. Yogyakarta. Penerbit
Khurniawati, R. 2007. Pengaruh Konsentrasi Kinetin dan Sukrosa Terhadap Pertumbuhan Tunas Tanaman Krisan (Chrisanthemum morifolium Ram.)
Secara In Vitro. dalam Final Project Departement of Agronomi 3(1) Malang:
Universitas Muhammadiyah Malang.
Kosmiatin, M. & Mariska, I. 2005.Kultur Embrio Dan Penggandaan Kromosom Hasil
PersilanganKacang Hijau Dan Kacang Hitam. Jurnal Bioteknologi
Pertanian 10(1) : 24-34.
Marlin. 2005. Regenerasi In Vitro planlet jahe bebas penyakit layu bakteri pada beberapa tahap konsentrasi 6-benzylaminopurin (BAP) dan 1-Napthalene Acetic Acid (NAA). Jurnal Ilmu-Ilmu Pertanian Indonesia 7(1): 8-14.
Mattjik, N. A. 2005. Peran Kultur Jaringan Dalam Perbaikan Tanaman. Bogor: Fakultas Pertanian IPB.
Nurwahyuni, I. 2001. Perbanyakan Tanaman Jeruk Manis (Citrus sinensis) Secara Kultur Jaringan. Medan: Laporan Penelitian. FMIPA. USU.
Nusmawarhaeni, S., Diah, P., Endang, P. P. 2001. Mengenal Buah Unggul Indonesia. Cetakan kesembilan. Jakarta: Penebar Swadaya. Hal: 24.
Pierik, R. L. M. 1987. In Vitro Culture of Higher Plants. Netherland: Martinus Nijhoff Publisher. pp. 50
Pierik, R. L. M. 1987. In vitro Culture of Higher Plants. Martinus Nijhoff Publishers, Dordrecht Boston Lancaster. pp. 344.
Preeven, S., Pawar, V., Ahlawat. Y.S. 2003. Somatic Embryoge nesis and Plant Regeneration In Kinnow Mandarin. Journal Plant Biochemistry &
Biotechnology 12 : 163-165.
Purseglove. 1987. Teknik Sambung Pucuk Menggunakan Tiga Stadia Entres pada
Bibit Tanaman Jeruk Keprok (Citrus nobilis).
Reinert, J and Bajaj, Y. P. S. 1989. Applied and Fundamental Ascepts of Plant Cell
Tissue, and Organ Culture. New Delhi: Narosa Publishing House.
Santoso, U dan F. Nursandi. 2004. Kultur Jaringan Tanaman. Cetakan Kedua. Malang: Universitas Muhammadiyah. Hal: 63, 115, 137-138.
Silalahi, R. R. 2006. Pengaruh Konsentarsi Media MS Dengan Kombinasi 2,4 D dan BAP Pada Kultur Biji Jeruk Brastepu (Citrus nobilis Lour. Var. Brastepu).
Skripsi. Medan: Fakultas Matematika Dan Ilmu Pengetahuan Alam. USU.
Sitepu, B.R. 2007. Inisiasi In Vitro Biji Jeruk Keprok Brastepu (Citrus nobilis Lour. Var. Brastepu) Dengan Kombinasi NH4NO3 Dan Sukrosa Pada Media MS. Skripsi. Medan: Fakultas Matematika dan Ilmu Pengetahuan Alam. USU.
Soelarso, R. B. 1996. Budi Daya Jeruk Bebas Penyakit. Yogyakarta: Kanisius.
Steenis, J. H. 2003. Flora untuk Sekolah di Indonesia. Cetakan Kesembilan. Jakarta: PT. Pradnya Paramita.
Suryowinoto, M. 1996. Pemuliaan Tanaman Secara In Vitro. Yogyakarta: Kanisius. Tjitrosomo, S. S. 1983. Botani Umum 2. Bandung: Angkasa. Hal: 127-129.
Veronika, E. S. 2007. Kultur Biji Jeruk Keprok Brastepu (Citrus nobilis Lour. var. Brastepu). Pada Media MS Dengan Penambahan Glisin dan Zat Pengatur Tumbuh. Skripsi. Medan: Fakultas Matematika dan Ilmu Pengetahuan Alam. USU.
Wattimena, G. A. 1992. Zat Pengatur Tumbuh Tanaman. Bogor: Pusat Antara Universitas IPB. Hal: 18.
Wetter, L. R. dan F. Constabel. 1991. Metode Kultur Jaringan Tanaman (edisi bahasa Indonesia). Bandung: ITB.
Wiryanta, W. dan Rahardja, P. C. 2003. Aneka Cara Memperbanyak Tanaman. Jakarta: Agromedia Pustaka.
Wulandari, S., Syafii, W., Yossilia. 2004. Respon Eksplan DaunTanaman Jeruk Manis (Citrus sinensis L.) Secara In Vitro Akibat Pemberian NAA dan BA. Jurnal
Biogenesis 1(1) : 21-25.
Yusnita, 2003. Kultur Jaringan Cara Memperbanyak Tanaman Secara Efisien. Cetakan Pertama. Jakarta : Agromedia Pustaka. Hal: 8, 46-49, 56.
Lampiran A. Komposisi Media MS (Murashige & Skoog) 1962
Lampiran B. Layout Penelitian
Lampiran C. Data Pengamatan dan Daftar Sidik Ragam Jumlah Tunas
Daftar Sidik Ragam Jumlah Tunas
SK DB JK KT Fh FT 5 % FT 1 %
Daftar Sidik Ragam Jumlah Tunas
Minggu III
Daftar Sidik Ragam Jumlah Tunas
SK DB JK KT Fh FT 5 % FT 1 %
Data Sidik Ragam Jumlah Tunas