Journal of Experimental Marine Biology and Ecology 255 (2000) 215–227
www.elsevier.nl / locate / jembe
Upper thermal tolerances of the beachflea Orchestia
gammarellus (Pallas) (Crustacea: Amphipoda: Talitridae)
associated with hot springs in Iceland
a ,* b
´ David Morritt , Agnar Ingolfsson
a
School of Biological Sciences, Royal Holloway, University of London, Egham, Surrey TW20 0EX, UK
b
´
Institute of Biology, University of Iceland, Grensavegur 12, IS-108 Reykjavik, Iceland Received 5 June 2000; received in revised form 12 August 2000; accepted 18 September 2000
Abstract
The upper thermal tolerance (CTmax) of beachfleas Orchestia gammarellus (Pallas) collected from a number of different locations in Iceland was determined. Differences were recorded between field populations associated with thermal springs and those from non-thermal sites. A number of reciprocal acclimation experiments (where animals from thermal and non-thermal sites were acclimated to the measured ambient temperatures of thermal (17 and 228C) and non-thermal (118C) sites) were performed. Differences between at least one thermal population and a non-thermal population were maintained following this reciprocal acclimation, supporting the hypothesis that population differences were due to non-reversible genetic differences and not local acclimatisation. Animals from one thermal site (Reykjanes) had a mean CTmax537.160.58C when acclimated at 118C and 38.660.38C when acclimated at 228C, whereas animals from a non-thermal site (Hvassahraun) had CTmax values of 35.960.5 and 37.960.38C, respectively. In other cases, differences are best explained by local acclimatisation. Results are discussed in relation to ambient local conditions and the degree of isolation of the different populations.
2000 Elsevier Science B.V. All rights reserved.
Keywords: Amphipoda; Crustacea; Iceland; Talitridae; Thermal springs; Upper thermal tolerance
1. Introduction
The common beachflea Orchestia gammarellus (Crustacea: Amphipoda: Talitridae) (Pallas) is the dominant detritivore in strandline algae on a wide range of rocky, boulder and shingle shores and saltmarshes in NW Europe. Talitrid amphipods play an important
*Corresponding author. Tel.:144-1784-443971; fax:144-1784-470756.
E-mail address: [email protected] (D. Morritt).
role in breakdown of organic debris and subsequent recycling of nutrients in these intertidal habitats, as well as being important food items for a range of invertebrate and vertebrate predators (Wildish, 1988, for review). The ecophysiology of talitrid am-phipods, especially O. gammarellus, has been extensively studied (Spicer et al., 1987; Morritt and Spicer, 1998, for reviews), especially aspects of respiratory physiology, osmo- and ionic regulation and developmental ecophysiology. However, we know relatively little about the thermal tolerances of talitrid amphipods, including O.
gammarellus, with the exception of a recent study by Gaston and Spicer (1998). The
presence in Iceland of populations of O. gammarellus associated with both thermal ´
springs and non-spring sites (Ingolfsson, 1996) offers a unique opportunity to test ideas on fundamental thermal tolerances of populations experiencing different thermal regimes.
Numerous studies have addressed the upper thermal tolerances of closely related groups of species and shown that species from warmer habitats have higher upper thermal tolerance (CTmax) than those from colder habitats. This may be related to latitude (e.g., Moulton et al., 1993, in North American caddisflies), altitude (e.g., Gaston and Chown, 1999, for African dung beetles), microhabitats within a narrow geographical range (e.g., Van der Merwe et al., 1997, for sub-Antarctic weevils), zonation on the shore (e.g., Davenport and MacAlister, 1996, for South Georgian intertidal invertebrates) or even depth distribution in the deep ocean (e.g., Young et al., 1998, for bathyal echinoid larvae). Perhaps more interesting is evidence that different populations within a particular species may have different thermal tolerances depending on their geographic origin, e.g. introduced North American and European populations of the zebra mussel,
Dreissena polymorpha (McMahon, 1996). Indeed, Gaston and Spicer (1998) have
recently demonstrated differences in the upper thermal tolerances of two latitudinally separated populations of the beachflea Orchestia gammarellus within the UK. Further-more, these differences were maintained following acclimation to a range of en-vironmentally realistic temperatures. It is well known that the CTmaxof many arthropod species can be increased by experimental acclimation to elevated temperatures (e.g., Edney, 1964, for isopod species), although this is not always the case (e.g., Quinn et al., 1994). Furthermore, this experimental acclimation can occur relatively quickly, some-times within 24 h (e.g., Colhoun, 1960). A similar process can also occur in nature and here the process is correctly referred to acclimatisation to the prevailing environmental conditions. The main aim of the present paper was to determine the upper thermal tolerances (CTmax) for a number of populations of O. gammarellus from Iceland and, by acclimation at different temperatures, attempt to reverse any differences in tolerance in order to determine whether differences are due to irreversible genetic adaptation or reversible, phenotypic acclimatisation. Experiments were designed with close reference to a similar study (Gaston and Spicer, 1998) on two latitudinally separated UK populations of O. gammarellus in order to allow comparison with those data.
2. Materials and methods
2.1. Upper thermal tolerance in Icelandic populations of Orchestia gammarellus
´
D. Morritt, A. Ingolfsson / J. Exp. Mar. Biol. Ecol. 255 (2000) 215 –227 217
´
where Orchestia gammarellus had previously been recorded by Ingolfsson (1996). At each site, animals were collected with substratum from the natural habitat along with decaying macroalgal debris to provide food. Animals were transported to the Sandgerdi Marine Centre in a sealed plastic container on their natural substratum and maintained in a cooled laboratory (158C) until required. Animals were fed on fucoid algae and chopped carrot ad libitum. During collection of animals, temperature was recorded to the nearest 0.18C with a digital temperature probe (Ama-Digit Precision, Amarell Electronic, Kreuzurstheim, Germany) in the microhabitats from where animals were collected as well as recording sea temperature and thermal spring temperature (where appropriate). Two sites were sampled on the Reykjanes peninsula in southwestern Iceland: a non-thermal site on Puccinellia maritima saltmarsh at Hvassahraun (648019110N,
¨
228099190W) and a saltmarsh site with thermal influence at Reykjanestjorn (638479520N, ¨ 228439120W). In retrospect it may have been inaccurate to describe the Reykjanestjorn population as being exposed to thermal influence: this is discussed later in more detail. Both these sites in southwestern Iceland are situated on Holocene basic and intermediate lava bedrock between 1100 and 11,000 years old. A further three thermal spring sites
¨
were sampled in northwestern Iceland at Hveravik, Steingrimsfjordur (658419470N, ¨
218339550W), Bjarnarstadir, Isafjordur (658499160N, 228299190W) and Reykjanes, ¨
Reykjafjordur (658559250N, 228259530W): no non-thermal sites were sampled in this area as Orchestia gammarellus are only found at sites associated with thermal springs in this part of Iceland. The northwestern sites are situated on upper and middle Miocene bedrock (10–15 million years old). Other characteristics of collecting sites and collection dates are given in Table 1.
Within 24–48 h of collection the wet mass of 150 randomly selected animals was measured 60.1 mg using a microbalance (Scaltec SBC22, Heiligenstadt, Germany) for each population: ovigerous females were debrooded prior to weighing. Debrooding can be performed routinely without damage to the female; indeed, broods can be returned to the marsupium without damage to either female or brood (see Morritt and Spicer, 1996, for details).
Within 48 h of collection the upper thermal tolerance (CTmax) in air of 30 individuals
Table 1
Site characteristics for populations of Orchestia gammarellus collected in Iceland. Note: the peculiar nature of ¨
the Reykjanestjorn site which is adjacent to a sea water inlet warmed by geothermal energy but the temperature of the microhabitat at which animals were collected was not appreciably warmed
Site Collection Substratum type / Microhabitat Thermal spring Sea water date microhabitat temp. (8C) temp. (8C) temp. (8C) Hvassahraun 6 July 1999 Saltmarsh / under lava stones 11.5–12.5 N /A 11.2
¨
Reykjanestjorn 6 July 1999 Saltmarsh / under lava stones 11.3–12.9 Spring warms 17.3 sea water inlet from beneath Hveravik 10 July 1999 Bedrock-gravel / under stones 13.3–15.0 73.4 6.3 Bjarnarstadir 10 July 1999 Bedrock-gravel / under stones 15.0–25.0 41.8 6.0 Reykjanes 10 July 1999 Bedrock-gravel / under stones 21.962.4 78.0 6.5
from each population, representing as wide a size range as possible, was determined, based on the technique described by Gaston and Spicer (1998). Each individual was weighed and then transferred to an individual small, glass test tube (vol. 4.5 ml) which had a small plug of seawater-soaked tissue paper in the bottom thus maintaining a high relative humidity and reducing the chance of evaporative cooling via desiccation. All sample tubes, held in a rack in a 536 array, had been left to equilibrate in the experimental water bath for at least 2 h prior to experimentation. Animals were allowed to settle for 30 min before experimentation. The temperature of the water bath (Grant Instruments, Cambridge, UK) was gradually increased from a starting point of 118C at a rate of 1.08C per min and all animals were continuously observed under cold light illumination (Intralux 4000-1, Volpi AG, Switzerland). When an individual stopped moving the tube was removed from the water bath and death was confirmed (or otherwise) by the presence of cardiac activity, visible microscopically (Wild M3Z, Heerbrugg, Switzerland) as regular contractions through the dorsal integument. Throughout the experiment the temperature at the paper / air / glass interface inside a tube in the middle of the array was monitored with the digital probe. This was found to be the most accurate measure of the actual temperature experienced by the experimental animals: temperature variation within the array was found to be 0.1–0.28C, whereas the measured water bath temperature at the end of each experiment was 0.8 to 1.08C higher than within the tubes. Where ovigerous females were used in experiments they were first debrooded as it was found that the brood in the female marsupium represented between 10 and 24% of the female body mass, mean 17.464.9, n515 (unpublished observation). This would otherwise lead to an error in calculating body mass–temperature tolerance relationships.
Following determination of the upper thermal tolerances of freshly collected animals, a number of animals were set up at acclimation temperatures in an attempt to reverse any thermal tolerance differences observed between non-thermal and thermal spring populations. Thus approximately 50 animals from the populations from Hvassahraun
¨
´
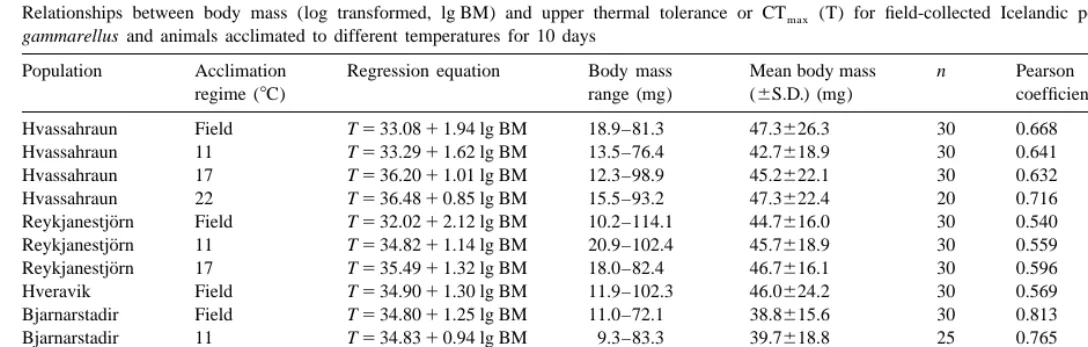
Relationships between body mass (log transformed, lg BM) and upper thermal tolerance or CTmax (T) for field-collected Icelandic populations of Orchestia
gammarellus and animals acclimated to different temperatures for 10 days
Population Acclimation Regression equation Body mass Mean body mass n Pearson Significance
regime (8C) range (mg) (6S.D.) (mg) coefficient
Hvassahraun Field T533.0811.94 lg BM 18.9–81.3 47.3626.3 30 0.668 ,0.001
Hvassahraun 11 T533.2911.62 lg BM 13.5–76.4 42.7618.9 30 0.641 0.001
Hvassahraun 17 T536.2011.01 lg BM 12.3–98.9 45.2622.1 30 0.632 0.001
Hvassahraun 22 T536.4810.85 lg BM 15.5–93.2 47.3622.4 20 0.716 0.001
¨
Reykjanestjorn Field T532.0212.12 lg BM 10.2–114.1 44.7616.0 30 0.540 0.002 ¨
Reykjanestjorn 11 T534.8211.14 lg BM 20.9–102.4 45.7618.9 30 0.559 0.001 ¨
Reykjanestjorn 17 T535.4911.32 lg BM 18.0–82.4 46.7616.1 30 0.596 0.001
Hveravik Field T534.9011.30 lg BM 11.9–102.3 46.0624.2 30 0.569 0.001
Bjarnarstadir Field T534.8011.25 lg BM 11.0–72.1 38.8615.6 30 0.813 ,0.001
Bjarnarstadir 11 T534.8310.94 lg BM 9.3–83.3 39.7618.8 25 0.765 ,0.001
Bjarnarstadir 22 T536.3110.99 lg BM 15.7–90.4 40.1617.4 25 0.811 ,0.001
Reykjanes Field T535.8610.83 lg BM 9.8–116.6 47.6626.8 30 0.575 0.001
Reykjanes 11 T534.3511.70 lg BM 7.2–115.1 51.2625.8 30 0.790 ,0.001
determined as described above. For each experiment, 10 individual control animals were maintained in identical sample tubes at the temperature to which they had been acclimated.
All data analyses were performed using SPSS and Unistat statistical packages with reference to Zar (1996).
3. Results
Comparison of population log transformed body masses demonstrated that all populations had significantly different body sizes with the exception of the Hvassahraun and Reykjanes populations (Kruskal–Wallis, H548.5, d.f. 4, P,0.001: Dunns multiple comparisons test). Body masses of animals used in experimental determinations of CTmaxwere log transformed, allowing linear regression of CTmaxagainst log body mass: regression equations and body size ranges are presented in Table 2 for field-collected animals and animals acclimated to different temperatures. There was a significant positive relationship between CTmaxand log body mass for every natural population and experimental acclimation (Pearson correlation coefficient P#0.002 in each case). There was no mortality in each set of control animals run at acclimation temperatures alongside each experimental CTmax determination.
Homogeneity of slopes and intercepts of the lines relating body mass and CTmax for different populations at the same acclimation temperatures were compared using ANCOVA: results are displayed in Table 3. From these analyses it is clear that there are
¨ significant differences in the intercepts between the Hvassahraun and Reykjanestjorn populations and the Hvassahraun and Reykjanes populations at both acclimation temperatures tested. For a given body mass the CTmax was significantly greater in the
¨
118C acclimated Reykjanestjorn population than the Hvassahraun population; the situation is apparently reversed for 178C acclimated animals. For a given body mass the CTmax was significantly higher for Reykjanes animals than for Hvassahraun animals regardless of acclimation temperature. The comparison of the Hvassahraun and
Table 3
Results of ANCOVA comparing regressions of different populations acclimated to the same acclimation temperatures
Comparison Homogeneity Significance Homogeneity Significance
of slope of intercept
Hvassahraun–Reykjanestjorn 118C F50.85, d.f. 1,56 P.0.35 F563.91, d.f. 1,57 P,0.001 Hvassahraun–Bjarnarstadir 118C F54.45, d.f. 1,47 P,0.05
´
D. Morritt, A. Ingolfsson / J. Exp. Mar. Biol. Ecol. 255 (2000) 215 –227 221
Fig. 1. Comparison of the predicted population distribution of upper thermal tolerances (CTmax) for ¨
Bjarnarstadir populations was less conclusive: either slopes were heterogeneous or intercepts were not significantly different.
The regression equations derived for each population were used to predict population CTmaxdistributions with reference to the summer-collected population size data for each population (c.f. Gaston and Spicer, 1998, for summer-collected UK populations). The resulting plots of predicted population upper thermal tolerances represent an illuminating ‘snapshot’ and are shown for those comparisons that were significantly different
¨
(ANCOVA): Fig. 1a–c for the Hvassahraun–Reykjanestjorn comparison and Fig. 2a–c for the Hvassahraun–Reykjanes comparison. There was a significant difference in the predicted population CTmax (Table 4) between all field-collected populations (Kruskal– Wallis H5541.9, d.f. 4, P,0.001. Dunn’s multiple comparisons test: Reykjanes.
¨
Hveravik.Bjarnarstadir.Hvassahraun.Reykjanestjorn). There was a significant differ-ence in the predicted population CTmax between all populations acclimated to 118C (Kruskal–Wallis H5387.1, d.f. 3, P,0.001. Dunn’s multiple comparison test:
¨
Reykjanes.Reykjanestjorn.Bjarnastadir.Hvassahraun). There was a significant dif-ference in the predicted population CTmax between the two populations acclimated to 178C (Mann–Whitney U56408.5, Z5 27.204, P,0.001) with the Hvassahraun site
¨
having a significantly higher population CTmax than the Reykjanestjorn site. For 228C acclimated animals there was also a significant difference between all tested populations
(H5214.3, d.f. 2, P,0.001. Dunn’s multiple comparison test: Reykjanes.
Hvassahraun.Bjarnarstadir).
4. Discussion
It is clear that, in field populations, whilst the thermal populations from northwestern Iceland all had a significantly higher CTmax than the non-thermal Hvassahraun
¨
population, the CTmax of the apparently thermal Reykjanestjorn population was lower than the non-thermal population. This is was also born out in later comparisons and suggests that the amphipods at this site are not really influenced by the underground heating of the associated sea water inlet which had a measured temperature of 178C.
¨
Indeed, the Reykjanestjorn population was the only population in which acclimation at 118C increased the value of CTmax, again suggesting that the population does not experience significantly elevated environmental temperatures. If one looks at Table 1
¨
one can see that actual microhabitat temperatures measured at Reykjanestjorn were not appreciably different to those measured at the non-thermal site, suggesting that
¨
describing the Reykjanestjorn population as being under thermal influence may have been erroneous in spite of the proximity of a geothermally warmed inlet.
The findings support the hypothesis that the Reykjanes population of Orchestia
gammarellus, which is undoubtedly constantly exposed to elevated ambient
´
D. Morritt, A. Ingolfsson / J. Exp. Mar. Biol. Ecol. 255 (2000) 215 –227 223
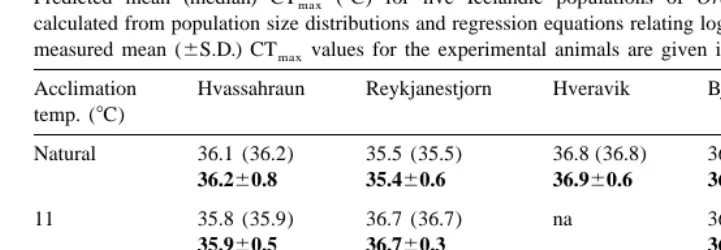
Table 4
Predicted mean (median) CTmax (8C) for five Icelandic populations of Orchestia gammarellus. Values calculated from population size distributions and regression equations relating log body mass to CTmax. Actual measured mean (6S.D.) CTmaxvalues for the experimental animals are given in bold for comparison Acclimation Hvassahraun Reykjanestjorn Hveravik Bjarnarstadir Reykjanes temp. (8C)
Natural 36.1 (36.2) 35.5 (35.5) 36.8 (36.8) 36.6 (36.6) 37.1 (37.2)
36.260.8 35.460.6 36.960.6 36.760.3 37.260.4
11 35.8 (35.9) 36.7 (36.7) na 36.2 (36.2) 36.9 (37.1)
35.960.5 36.760.3 36.360.3 37.160.5
17 37.8 (37.9) 37.6 (37.7) na na na
37.960.4 37.760.3
22 37.8 (37.9) na na 37.8 (37.8) 38.6 (38.6)
37.960.3 37.960.2 38.660.3
non-thermal population) or 228C (measured mean substratum temperature for thermal populations). Whilst acclimation to the higher temperature increased CTmax, acclimation to 118C resulted in a small decrease in CTmax of 0.28C. These results support the hypothesis that the increased CTmax in the population is due to irreversible genetic adaptation rather than local acclimatisation. The situation is not as clear with the Bjarnarstadir thermal population where it would appear that differences in thermal tolerance are reversible by acclimation to temperatures experienced by other O.
gammarellus populations in Iceland. The source of heat at this population is more
localised and it would seem that while the presence of the geothermal spring allows the amphipods to survive in an otherwise hostile environment, the thermal influence may not have resulted in a genetically fixed change in temperature tolerance, only in local acclimatisation.
´
D. Morritt, A. Ingolfsson / J. Exp. Mar. Biol. Ecol. 255 (2000) 215 –227 225
wrack fauna and those described by Moore and Francis (1985) for a Clyde Sea population. Moore and Weeks (1995) have recorded that O. gammarellus from The Azores are capable of surviving temperatures in excess of 408C for short periods of time. It should be noted, however, that these Azorean populations are shifted landward from their typical distribution on most NW European shores and consequently experience a more extreme temperature regime.
In all the Icelandic populations examined we found a positive relationship between increasing body size and CTmax. Whilst this agrees with the majority of the results presented by Gaston and Spicer (1998) and other studies on amphipod crustaceans (e.g., Sutcliffe et al., 1981) there are just as many studies which either found no relationship (e.g., Lazo-Wasem, 1984; Agnew and Taylor, 1986) or the reverse relationship, with smaller individuals being more tolerant to elevated temperature (e.g., Marsden, 1985). There is thus no general trend in this group of crustaceans as regards the relationship between body size and upper temperature tolerance. Gaston and Spicer (1998) showed that thermal acclimation affected the CTmax recorded for O. gammarellus with an increase in median upper thermal tolerance of 3–48C (between 5 and 208C). In the present study (depending on which population one considers) there was an increase in CTmax of between 1.6 and 2.08C between acclimation temperatures of 11 and 228C. These latter results are similar to the two comparable studies on amphipods discussed in Gaston and Spicer’s paper, namely Paramelita nigroculus (acclimation temperatures 8.5 and 208C — difference 28C, Buchanan et al., 1988) and Arcitalitrus sylvaticus (acclimation temperatures 10 and 208C — difference 2.68C, Lazo-Wasem, 1984).
The interesting evolutionary question is just how divergent are these populations? How long have the thermal spring populations in northwestern Iceland been isolated? It appears quite likely that they are glacial relict populations having been isolated for
´
2500–4800 years (Ingolfsson 1996), say 5000–10,000 amphipod generations (based on the assumption of two generations per year, see Wildish, 1988). Quite apart from the interesting evolutionary questions these results pose there may also be implications for the reproductive cycle of these thermal populations of O. gammarellus. It is known that temperature is an important cue in initiating breeding in several amphipod species including O. gammarellus (Barnett, 1971; de March, 1977; Morritt and Stevenson, 1993). Thermal spring populations in northwestern Iceland would be expected to have a significantly lengthened breeding season compared with non-spring populations. Growth rates and moulting frequency (also temperature-dependent) would also be expected to be greater (e.g., Charniaux-Cotton, 1957). Preliminary observations certainly support the hypothesis that thermal and non-thermal populations were at different stages in their reproductive cycles at time of sampling (DM, unpublished observation), although there was no apparent difference in body masses of the non-thermal population at Hvas-sahraun and the thermal population at Reykjanes.
Acknowledgements
Research Council (NERC) Advanced Research Fellowship (GT5 /ALS / 94 / 2). DM acknowledges the EC-TMR Large Scale Facility programme for financial support and NERC for funding internal travel costs in Iceland. Gudmundur Vidir Helgasson is thanked for organising many aspects of the visit and for help with equipment. We would also like to thank Linda Wendel, Jon Olafsson, Caroline Nicholson and Emilia Jonsdottir for their help on collecting trips. [SS]
References
Agnew, D.J., Taylor, A.C., 1986. Effects of oxygen tension, temperature, salinity, and humidity on the survival of two intertidal gammarid amphipods. Mar. Ecol. Prog. Ser. 32, 27–33.
Backlund, H.O., 1945. Wrack fauna of Sweden and Finland, ecology and chorology. Opusc. Ent. (Suppl.) 45, 1–236.
Barnett, P.R.O., 1971. Some changes in intertidal sand communities due to thermal pollution. Proc. R. Soc. London 177, 353–364.
Buchanan, J.A., Stewart, B.A., Davies, B.R., 1988. Thermal acclimation and tolerance to lethal high temperature in the mountain stream amphipod Paramelita nigroculus (Barnard). Comp. Biochem. Physiol. 89A, 425–431.
´ ´ ´ ´ `
Charniaux-Cotton, H., 1957. Croissance, regeneration et determinisme endocrinien des characteres sexuels ´
d’Orchestia gammarella (Pallas) Crustace, Amphipode. Ann. Sci. Nat. Zool. Biol. Anim. Ser. 11 (19), 411–560.
Colhoun, E.H., 1960. Acclimatization to cold in insects. Entomol. Exp. Appl. 3, 27–37.
Davenport, J., MacAlister, H., 1996. Environmental conditions and physiological tolerances of intertidal fauna in relation to shore zonation at Husvik, South Georgia. J. Mar. Biol. Assoc. UK 76, 985–1002. Edney, E.B., 1964. Acclimation to temperature in terrestrial isopods. I. Lethal temperatures. Physiol. Zool. 37,
364–377.
Gaston, K.J., Chown, S.L., 1999. Elevation and climatic tolerance: a test using dung beetles. Oikos 86, 584–590.
Gaston, K.J., Spicer, J.I., 1998. Do upper thermal tolerances differ in geographically separated populations of the beachflea Orchestia gammarellus (Crustacea: Amphipoda). J. Exp. Mar. Biol. Ecol. 229, 265–276.
´
Ingolfsson, A., 1996. The distribution of intertidal macrofauna on the coasts of Iceland in relation to temperature. Sarsia 81, 29–44.
Lazo-Wasem, E.A., 1984. Physiological and behavioural ecology of the terrestrial amphipod Arcitalitrus
sylvaticus. J. Crust. Biol. 4, 343–355.
de March, B.G.E., 1977. The effects of photoperiod and temperature on the induction and termination of reproductive resting stage in the freshwater amphipod Hyalella azteca (Saussure). Can. J. Zool. 55, 1595–1600.
Marsden, I.D., 1985. Some factors affecting survival and oxygen uptake in a subtropical beach flea. J. Exp. Mar. Biol. Ecol. 88, 213–225.
McMahon, R.F., 1996. The physiological ecology of the zebra mussel, Dreissena polymorpha, in North America and Europe. Am. Zool. 36, 339–363.
Moore, P.G., Francis, C.H., 1985. On the water-relations and osmoregulation of the beach-hopper Orchestia
gammarellus (Pallas) (Crustacea: Amphipoda). J. Exp. Mar. Biol. Ecol. 94, 131–150.
Moore, P.G., Weeks, J.M., 1995. Observations on the natural history of some supralittoral and terrestrial talitrid amphipods from the Azores. Ac¸oreana Suppl. 1995, 79–91.
Morritt, D., Spicer, J.I., 1996. Developmental ecophysiology of the beachflea Orchestia gammarellus (Pallas) (Crustacea: Amphipoda). I. Female control of the embryonic environment. J. Exp. Mar. Biol. Ecol. 207, 191–203.
´
D. Morritt, A. Ingolfsson / J. Exp. Mar. Biol. Ecol. 255 (2000) 215 –227 227 Morritt, D., Stevenson, T.D.I., 1993. Factors influencing breeding initiation in the beachflea Orchestia
gammarellus (Pallas) (Crustacea: Amphipoda). J. Exp. Mar. Biol. Ecol. 165, 191–208.
Moulton, S.R., Beitinger, T.L., Stewart, K.W., Currie, R.J., 1993. Upper temperature tolerance of four species of caddisflies (Insecta, Trichoptera). J. Freshwater Ecol. 8, 193–198.
Quinn, J.M., Steele, G.L., Hickey, C.W., Vickers, M.L., 1994. Upper thermal tolerances of 12 New Zealand stream invertebrate species. N.Z. J. Mar. Freshwater Res. 28, 391–397.
Spicer, J.I., Moore, P.G., Taylor, A.C., 1987. The physiological ecology of land invasion by the Talitridae (Crustacea: Amphipoda). Proc. R. Soc. B 232, 95–124.
Sutcliffe, D.W., Carrick, T.R., Willoughby, L.G., 1981. Effects of diet, body size, age and temperature on growth rates in the amphipod Gammarus pulex. Freshwater Biol. 11, 183–214.
Van der Merwe, M., Chown, S.L., Smith, V.R., 1997. Thermal tolerance limits in six weevil species (Coleoptera, Curculionidae) from sub-Antarctic Marion Island. Polar Biol. 18, 331–336.
Wildish, D.J., 1988. Ecology and natural history of aquatic Talitroidea. Can. J. Zool. 66, 2340–2359. Withers, P.C., 1992. Comparative Animal Physiology. Saunders College Publishing, New York.
Young, C.M., Ekaratne, S.U.K., Cameron, J.L., 1998. Thermal tolerances of embryos and planktotrophic larvae of Archaeopneustes hystrix (A. Agassiz) (Spatangoidea) and Stylocidaris lineata (Mortensen) (Cidaroidea), bathyal echinoids from the Bahamian Slope. J. Exp. Mar. Biol. Ecol. 223, 65–76.